БИОФИЗИКА, 2021, том 66, № 4, с. 784-790
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 577.3:612.8
МОДЕЛЬ ГЕНЕРАЦИИ АКТИВНОСТИ
В ЭПИЛЕПТИЧЕСКОМ ФОКУСЕ
© 2021 г. Е.Л. Машеров
Национальный медицинский исследовательский центр нейрохирургии имени академика Н.Н. Бурденко МЗ РФ,
125047, Москва, 4-я Тверская-Ямская ул., 16
E-mail: EMasherow@nsi.ru
Поступила в редакцию 29.07.2020 г.
После доработки 20.08.2020 г.
Принята к публикации 21.08.2020 г.
Предложена модель генерации активности в эпилептическом фокусе, основанная на предположе-
нии о синхронизации колебаний, возникающих в ходе регулирования уровня постоянного потен-
циала, и синхронизации нейронных разрядов с медленными колебаниями. В ходе развертывания
приступа предположены четыре фазы - нарастания медленной активности, высокочастотных
вспышек, синхронизации с появлением острых пиков, угасания приступа. Проведено сопоставле-
нием модели с записью электрокортикограммы во время операции.
Ключевые слова: эпилепсия, ЭЭГ, высокочастотная активность, медленноволновая активность,
регулирование, синхронизация.
DOI: 10.31857/S0006302921040189
новых комплексов. Это позволяет поставить два
Интерес, который вызывает эпилепсия, обу-
вопроса - какова роль этих вспышек в формиро-
словлен как тем, что, с одной стороны, она явля-
вании эпилептического очага и по какой причине
ется третьей по частоте среди органических пора-
они трудно регистрируемы со скальпа, хотя хоро-
жений мозга, так и тем, с другой стороны, что
шо проявляются при регистрации с коры. Ответ
проявляется в виде специфических электрогра-
на эти вопросы позволит прояснить патогенез
фических паттернов, и понимание механизмов их
эпилепсии и будет способствовать разработке ме-
возникновения позволяет прояснить механизмы
тодов выявления эпилептической активности,
функционирования мозга [1, 2]. Разнообразие
включая как аппаратуру и методику регистрации,
форм заболевания выражается и в разнообразии
так и алгоритмы обработки электроэнцефало-
эпилептиформных паттернов на электроэнцефа-
грамм.
лограмме (ЭЭГ), однако для них, как правило, ха-
рактерны высокая амплитуда и острая форма
волн [3]. Среди прочих типичных для паттернов
МОДЕЛЬ СИНХРОНИЗАЦИИ
разрядной активности свойств отмечаются спон-
РАЗНОРОДНЫХ ИСТОЧНИКОВ
танное возникновение и независимость от фоно-
БИОЭЛЕКТРИЧЕСКОЙ АКТИВНОСТИ
вой активности, однотипно повторяющиеся по
КАК МЕХАНИЗМ ФОРМИРОВАНИЯ
форме комплексы волн, тенденция к усилению и
ПРИСТУПА
рекрутированию [1]. Запись непосредственно с
В качестве источника электроэнцефалограм-
поверхности коры позволила регистрировать не
мы в настоящее время в литературе принимается
наблюдаемые при обычной (скальповой) ЭЭГ
суммирование дипольных возбуждающих и тор-
вспышки высокочастотной активности, проявля-
мозящих синаптических потенциалов [2]. Исто-
ющиеся в диапазоне, превышающем не только
рически первое объяснение - образование волн
обычно принимаемую верхнюю границу β-ритма
суммированием потенциалов действия, что поро-
35 Гц, но и границу γ-ритма 70 Гц, и включающие
дило термины «синхронизация» и «десинхрони-
частоты до 250-500 и даже 1000 Гц. Как правило,
зация», обозначавшие появление и исчезновение
появление таких вспышек предшествует появле-
высокоамплитудной ритмической активности -
нию видимого на снимаемой со скальпа ЭЭГ эпи-
было отвергнуто с появлением микроэлектрод-
лептиформного паттерна, в частности, пик-вол-
ной записи. Также было отвергнуто объяснение
электроэнцефалограммы колебаниями уровня
Сокращениe: ЭЭГ - электроэнцефалограмма.
постоянного потенциала [4]. Однако представля-
784
МОДЕЛЬ ГЕНЕРАЦИИ АКТИВНОСТИ
785
ется целесообразным для объяснения феноменов
дипольной и квадрупольной составляющей будут
ЭЭГ во всем их разнообразии вернуть в рассмот-
меняться. Кроме того, на величину регистрируе-
рение и эти механизмы, не отрицая, а дополняя
мого потенциала влияет угол между диполем или
ими представление о возбуждающих и тормозя-
квадруполем и направлением на точку регистра-
щих синаптических потенциалах как источниках
ции. Этим, в частности, можно объяснить то, что
формирования биоэлектрической активности. В
вспышки высокой частоты, хорошо видимые на
работе [5] нами предложена модель формирова-
монополярной записи, не проявляются на бипо-
ния низкочастотной (δ-, θ- и, предположительно
лярной. Потенциал действия, создаваемый нерв-
α-диапазонов) составляющей ЭЭГ как колебаний
ным импульсом, идущим по аксону, можно рас-
уровня постоянного потенциала, возникающих в
сматривать, как квадрупольный, и при увеличении
процессе его регулирования. Для объяснения
расстояния он быстро убывает, его регистрация
эпилептиформных феноменов, протекающих за
возможно лишь в непосредственной близости.
короткое время (спайки - 20-50 мс, высокоча-
Этим может объясняться невозможность записи
стотные колебания достигают частоты 500 и более
высокочастотных вспышек со скальпа: удаление
Гц), помимо такого механизма и постсинаптиче-
на толщину мозговых оболочек, черепной кости и
ских потенциалов следует, вероятно, включить и
кожи уже достаточно для ослабления сигнала ниже
имеющие меньшую временную протяженность
порога регистрации.
феномены, в частности потенциалы действия.
Однако разнесенные в пространстве источники
Невозможность зарегистрировать на скальпе
могут синхронизироваться, формируя поле, убы-
потенциал действия, продемонстрированная в са-
вающее медленнее, чем от единичного источника
мом начале развития электроэнцефалографии и
соответствующего вида. Рассмотрим простейший
вызвавшая отказ от объяснения ЭЭГ синхрониза-
случай - круглую площадку радиуса R, в каждой
цией потенциалов действия, обусловлена, как нам
точке которой одинаковые и однообразно, нор-
представляется, разным характером убывания
мально к поверхности площадки, ориентирован-
электрического потенциала при различной кон-
ные квадрупольные источники, и найдем потен-
фигурации зарядов. Мультипольное разложение
циал в точке, расположенной над центром пло-
поля, создаваемого системой зарядов, представля-
щадки на расстоянии d. Создаваемый единичным
ет его, как сумму потенциалов, создаваемого еди-
квадруполем Q в данной точке потенциал равен
ничным зарядом, диполем, квадруполем и мульти-
полями более высокого порядка. Хотя непосред-
ственно сравнивать между собой заряд, дипольный
2
2
3cos
θ-1
3h
1
и квадрупольный потенциалы невозможно, по-
π(r,h)
=
Q
=
Q
−
,
3
5
3
скольку они имеют разные размерности, однако
l
2
2
2
2
2
2
(
r
+
h
)
(
r
+
h
)
для определенной точки регистрации возможно
сопоставлять вклад различных мультиполей в сум-
где l - расстояние от квадруполя до точки реги-
марный потенциал. При этом с ростом расстояния
страции. Тогда суммарный потенциал равен
потенциал, формируемый единичным зарядом,
убывает обратно пропорционально расстоянию,
R
2
диполем - обратно пропорционально квадрату
3h
1
Π(h)
=
Qr
−
dr
расстояния, квадруполем - обратно пропорцио-
5
3
2
2
2
2
0
2
2
нально кубу расстояния. Таким образом, в зависи-
(
r
+
h
)
(
r
+
h
)
мости от расстояния между источником и местом
регистрации, относительные вклады зарядовой,
После замены y = (r2 + h2) и dy = 2rdr получим
2
2
R
+h
5
3
−
-
2
Q
2
Q
h
8
1
2
2
Π(h)
=
3h
y
−
y
dy
=
-9
+
+
3
1
2
2
4
2
2
h
2
2
2
2
h
(
R
+
h
)
(
R
+
h
)
Очевидно, при h << R убывание потенциала
лов действия может сформировать конфигура-
медленное, определяясь вторым слагаемым, об-
цию поля, регистрируемого на достаточно боль-
ратно пропорциональным расстоянию, а по мере
шом расстоянии. Можно предположить, что
роста ускоряется и при h >> R приближается по
отдельные видимые лишь при регистрации с ко-
ры высокочастотные разряды в ходе развития
скорости убывания к квадрупольному.
эпилептической активности синхронизируются
Это позволяет сделать вывод, что суммирова-
во времени, формируя спайк или острую волну,
ние пространственно распределенных потенциа- которая уже доступна регистрации и на скальпо-
БИОФИЗИКА том 66
№ 4
2021
786
МАШЕРОВ
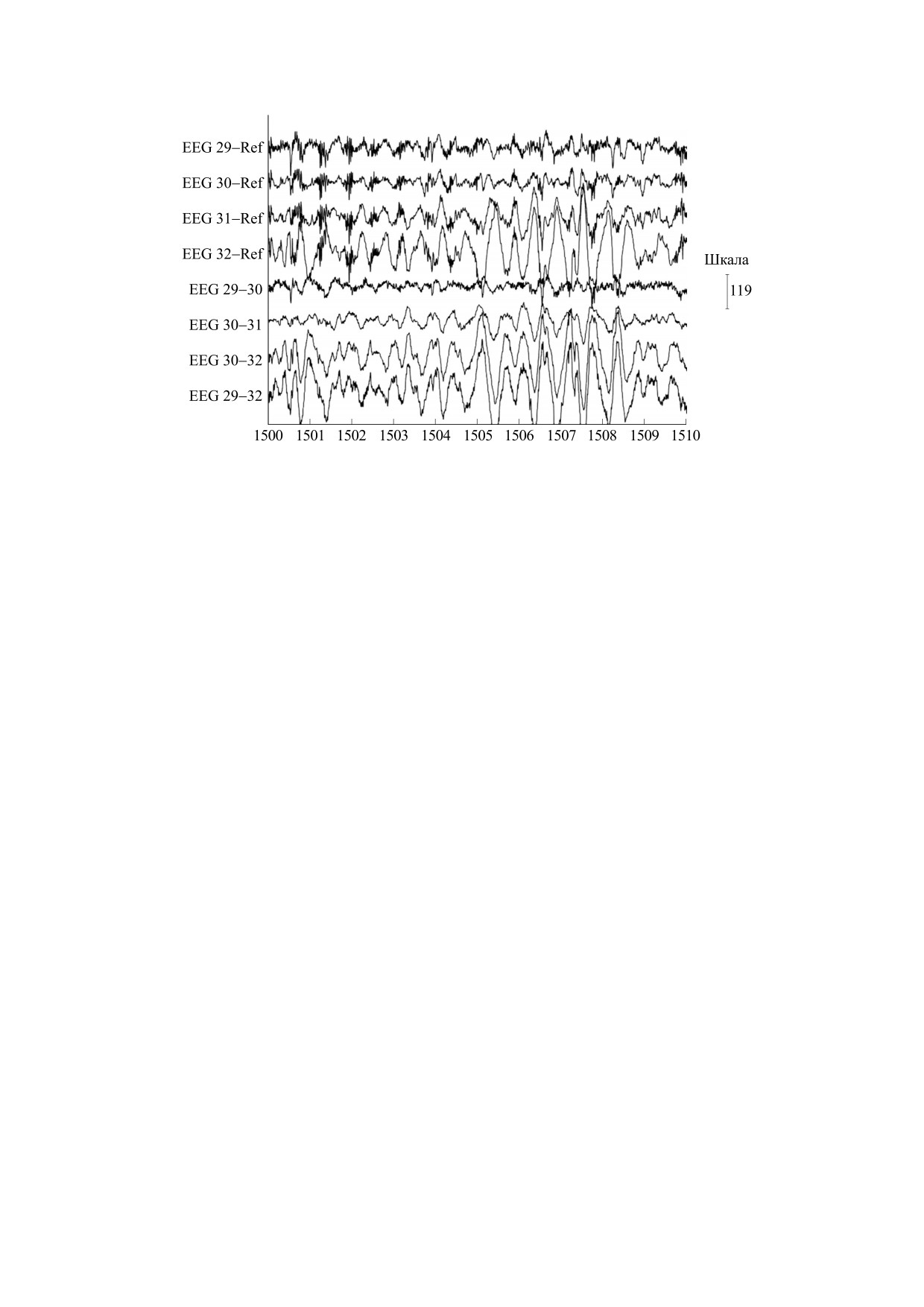
Рис. 1. Фрагмент электрокортикографической записи во время приступа. Высокочастотные вспышки хорошо
заметны на монополярной записи (первые четыре канала), но не на биполярной (последующие четыре канала).
Медленные колебания отчетливо видны в обоих случаях.
вых электродах в ходе обычной ЭЭГ, а не только
формируют достаточно далеко распространяю-
электрокортикографии.
щееся поле, регистрируемое, в частности, как
ЭЭГ. Для данной модели наиболее существенны-
Механизм такой синхронизации может быть
ми представляются два из них - диффузия ионов
выведен из высказанной ранее гипотезы о том,
в нервной ткани и возбуждение нервной клетки
что низкочастотные колебания (от δ- до, предпо-
полем потенциала действия другой. Возможны и
ложительно, части α-диапазона) биопотенциала
иные механизмы синхронизации, которые, дей-
отражают процессы регулирования ионного со-
ствуя на большем расстоянии, могут участвовать
става в ходе поддержания уровня постоянного
в формировании вторичных и зеркальных очагов.
потенциала нейрона [5]. Предполагаемый меха-
низм этого регулятора основан на совместном
В силу диффузии изменение концентрации
действии потенциал-зависимых кальциевых ка-
ионов вблизи нервной клетки - единичного ос-
налов и калий-натриевого насоса, управляемого
циллятора - оказывает влияние на колебания со-
ионами кальция. Отклонение уровня потенциала
седних осцилляторов. Возникает волна, удовле-
меняет поступление ионов кальция, а их накоп-
творяющая волновому уравнению с затуханием
ленная величина регулирует скорость работы ка-
2
∂
u
∂u
лий-натриевого насоса. Такая двухступенчатая
Δu
=α
и пробегающая по ткани от точ-
2
схема может рассматриваться как интегральный
∂t
+β∂t
ки первичного возбуждения. Если простран-
регулятор, который, в отличие от более простого
ственный ее полупериод сравним с размером об-
пропорционального регулятора, является астати-
ласти, с которой регистрируется сигнал, эта вол-
ческим, т. е. способен поддерживать требуемое
значение параметра при различной нагрузке. Это
на отчетливо проявляется на ЭЭГ.
взаимодействие может быть в линейном прибли-
Изменение концентрации ионов меняет воз-
жении описано дифференциальным уравнением
будимость клеток, способствуя спонтанным раз-
второго порядка, решением которого являются
рядам, так что вспышки высокочастотных коле-
затухающие синусоидальные колебания, частота
баний есть пачки таких разрядов, возникающие
которых зависит, с одной стороны, от концентра-
на определенной фазе медленной волны. Разряды
ции ионов кальция, а с другой стороны, посколь-
отдельных нейронов возникают независимо, но с
ку калий-натриевый насос использует АТФ, - от
определенной вероятностью могут при очеред-
концентрации АТФ. Ее дефицит приводит к за-
ной вспышке оказаться настолько близки по вре-
медлению работы насоса, что проявляется в сни-
мени, что одновременное их действие окажется
жении частоты колебаний и росте их амплитуды.
достаточным для возбуждения разряда в соседних
Потенциал единичного осциллятора мал, и его
клетках. Различие между синхронизацией на вто-
регистрация затруднена. Однако имеют место ме-
рой и синхронизацией на третьей стадии состоит
ханизмы синхронизации колебаний, которые
в том, что на второй стадии колебания ионной
БИОФИЗИКА том 66
№ 4
2021
МОДЕЛЬ ГЕНЕРАЦИИ АКТИВНОСТИ
787
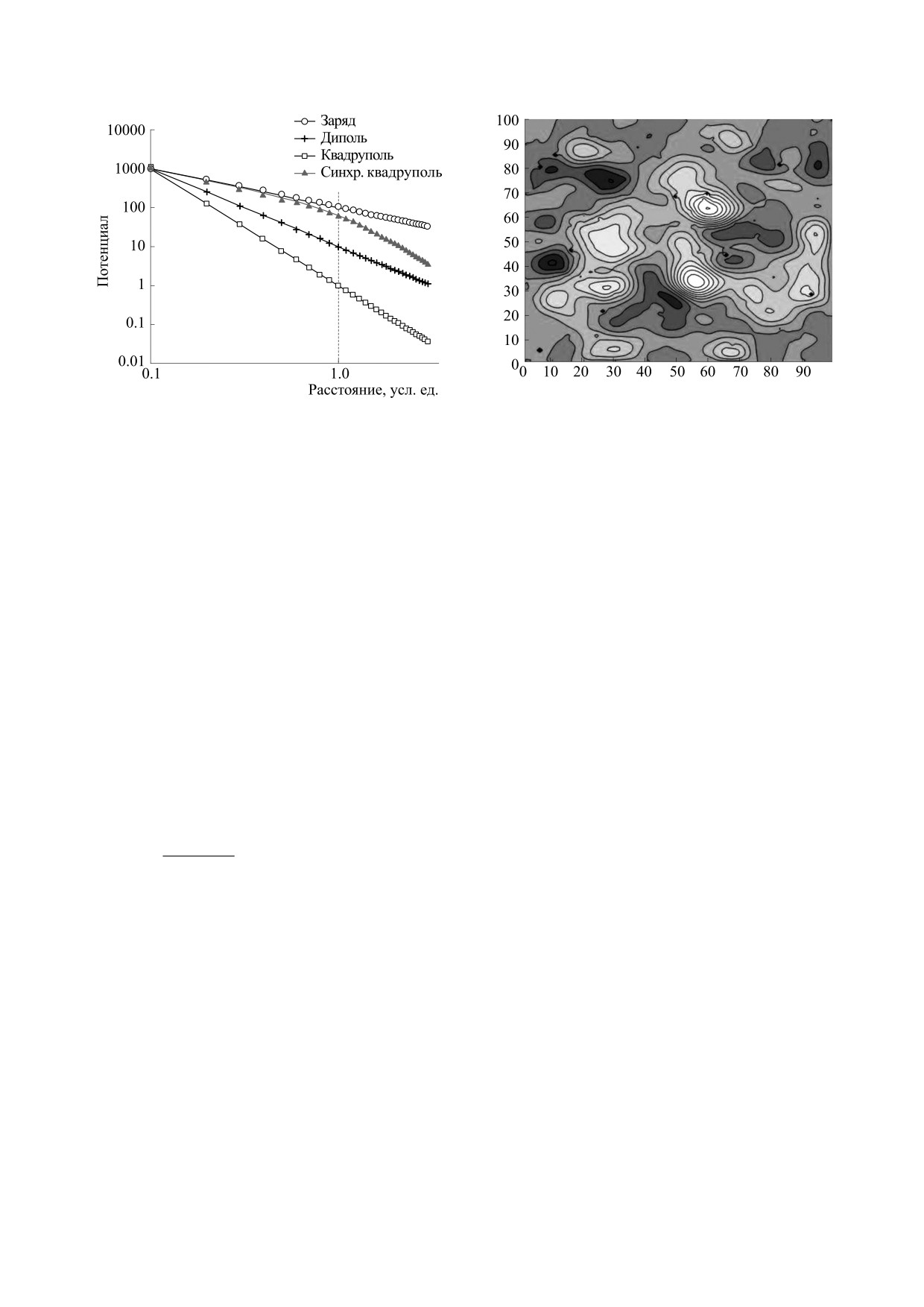
Рис. 2. Убывание создаваемого различными конфигу-
Рис. 3. Численная модель распространения осцилля-
рациями потенциала в зависимости от расстояния
ций при возбуждении случайно выбранных от-
(шкала по осям абсцисс и ординат логарифмическая,
дельных осцилляторов пуассоновским потоком им-
расстояние по оси абсцисс - в усл. ед.).
пульсов.
концентрации меняют вероятность спонтанного
ионного состава и синхронизации фаз медленных
проявления потенциала действия, и повышение
волн. В результате максимальная возбудимость
этой вероятности проявляется в учащении разря-
нейронов имеет место одновременно по всему
дов, что и регистрируется как высокочастотная
участку нервной ткани, в этот момент параметр
активность, огибающая которой синхронизиро-
интенсивности пуассоновского процесса λ при-
вана с низкочастотной активностью. На третьей
нимает значения существенно большие, чем на
же стадии между собой синхронизируются по
второй стадии, P(k) ≈ 1, т. е. разряды становятся
времени отдельные разряды, формируя острый
постоянно повторяющимися.
пик. Более формально это можно описать моде-
Таким образом серия разрядов соседних ней-
лью, в которой разряды i-го нейрона образуют
ронов, возникающая на определенной фазе про-
пуассоновский процесс с параметром λ, завися-
бегающей по ним медленной волны, перерожда-
щей от концентрации ионов ci в окрестности ней-
ется в синхронный разряд, который уже может
рона, как λi = λ(сi). Суммарный поток также будет
быть зарегистрирован на отдалении от места ге-
пуассоновским процессом с λ = λi. Если для воз-
нерации, проявляясь в виде острого пика на ЭЭГ.
буждения разряда необходимо возникновение на
В зависимости от степени одновременности раз-
промежутке времени Δt не менее k спонтанных
рядов это может быть спайк (длительность менее
разрядов, то вероятность такого события выража-
70 мс), острая волна (более 70 мс) или множе-
ется через неполную гамма-функцию, как
ственные спайки. Возникающее вследствие изме-
нения концентрации ионов и последующего про-
Γ(k
+1
,λ)
P k)
=
1−
цесса ее регулирования медленное колебание, в
k!
свою очередь, создает условия для формирования
нового острого пика. Замыкается цепь обратной
На второй стадии, когда медленные волны пе-
связи и возникает длительная серия однообраз-
ремещаются по участку нервной ткани и макси-
мальная возбудимость клеток не возникает одно-
ных паттернов, например «пик-волна». Прекра-
временно, суммарная интенсивность потока λ не
щаются они либо по истощению клеток, либо по
поступлению внешних импульсов, способных
столь велика и P(k) существенно меньше едини-
сбить синхронизацию.
цы. Поэтому отдельные разряды возникают неза-
висимо, проявляясь в виде пачек высокочастот-
Развитие эпилептической активности, таким
ных колебаний. Однако P(k) и не столь мала, что-
образом, представляется четырехстадийным. На
бы в какой-то момент одновременное импульс от
первой стадии появляется медленная активность,
k и более нейронов не имел бы места. Это событие
на второй медленные волны пространственно
приводит к генерации потенциала действия и
синхронизируются, и на определенной фазе вол-
других нейронов, формируя высокоамплитудный
ны возникают спонтанные разряды нейронов,
импульс, уже доступный регистрации со скальпа.
проявляющиеся на электрокортикограмме, как
Эта одновременность генерации потенциала дей-
вспышки высокочастотной активности, на тре-
ствия приводит к одновременности колебаний
тьей стадии эти вспышки синхронизируются во
БИОФИЗИКА том 66
№ 4
2021
788
МАШЕРОВ
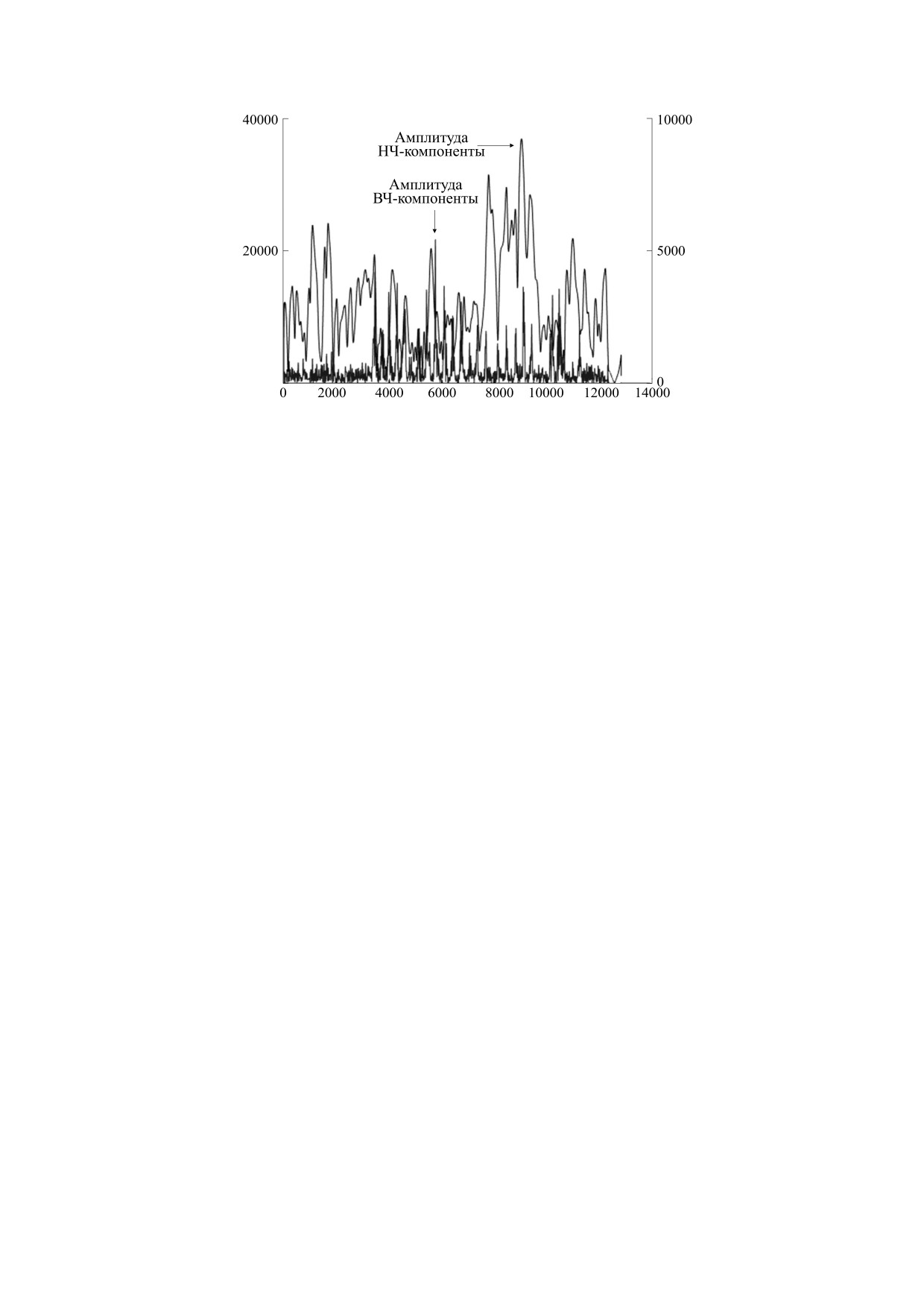
Рис. 4. Мгновенные мощности составляющих сигнала в мкВ2. По оси абсцисс время от начала записи в отсчетах
(частота записи 512 Гц).
времени, образуя регистрируемые в том числе и
участка приведенной записи это время составляет
на скальповой ЭЭГ острые волны, четвертая ста-
96 мс, а для третьего - 69 мс.
дия - угасание эпилептической активности.
Возникает гипотеза, что это отражает четыре
стадии развития эпилептического приступа. На
АНАЛИЗ ДАННЫХ
первой стадии, предшествующей приступу, в си-
Для проверки этого предположения подверг-
лу различных причин - нарушений гемодинами-
ки, снижения оксигенации, механического сдав-
нем частотной фильтрации отрезок записи элек-
трокортикограммы, выделив низкочастотную
ления опухолью или гематомой, метаболических
(1-5 Гц) и высокочастотную (40-100 Гц) компо-
отклонений или иных факторов - нарушается
ненты. Фильтрованный сигнал был подвергнут
функционирование регулятора, поддерживаю-
преобразованию Гильберта и получены оценки
щего уровень постоянного потенциала. Величина
мгновенных амплитуд компонентов сигнала
отклонения растет, как и время возвращения к
(представляет интерес также и оценка мгновен-
нормальному значению регулируемого парамет-
ных частот, но для нее требуется более узкополос-
ра, что проявляется, как высокоамплитудные ко-
ная фильтрация или переход к методу Гильберта-
лебания низкой частоты. В силу изменения кон-
Хуанга [6]).
центрации ионов меняется и возбудимость ней-
ронов, приводя к возникновению спонтанной
Видна картина изменений во времени обоих
импульсной активности. Это составляет вторую
видов активности и их соотношения. На данном
фрагменте можно выделить четыре участка - от
стадию, на которой эти импульсы, возникающие
начала фрагмента до приблизительно 7.5 с (отсчет
на определенной фазе медленноволновой актив-
ности, проявляются в виде вспышек высокой ча-
3800), от 7.5 с до 17.2 с (отсчет 7800), от 17.2 с до
23 с (отсчет 10800) и следующий за 23 секундами.
стоты. В свою очередь такие вспышки вызывают
Для первого характерна малая амплитуда высоко-
колебания концентрации ионов и этим запуска-
частотной активности и нерегулярная низкоча-
ют медленные волны, синхронизируясь с ними и
стотная, во втором появляются регулярные
между собой. Возникает контур положительной
вспышки высокочастотной активности, синхро-
обратной связи, в котором синхронизированные
низирующиеся с низкочастотной, в третьем ам-
разряды вызывают медленное колебание, а оно,
плитуда низкочастотной активности растет, а вы-
меняя возбудимость ткани, способствует на опре-
сокочастотной несколько снижается, в четвертом
деленной фазе колебания появлению синхрони-
угасает как низкочастотная, так и высокочастот-
зированного разряда. Степень временной син-
ная активность. При этом длительность высоко-
хронизации разрядов настолько высока, что при
частотных вспышек на третьем участке короче,
записи они сливаются в пик или острую волну, а
чем на втором. Если в качестве оценки длитель-
за счет описанной выше пространственной син-
ности вспышки принять время, за которое ам-
хронизации формируемое ими электрическое по-
плитуда нарастает до максимального значения от
ле может быть зарегистрировано не только в не-
величины в половину максимальной и затем убы-
посредственной близости, но и на удалении от
вает до половины максимальной, то для второго
источника. Это третья стадия, уже доступная ре-
БИОФИЗИКА том 66
№ 4
2021
МОДЕЛЬ ГЕНЕРАЦИИ АКТИВНОСТИ
789
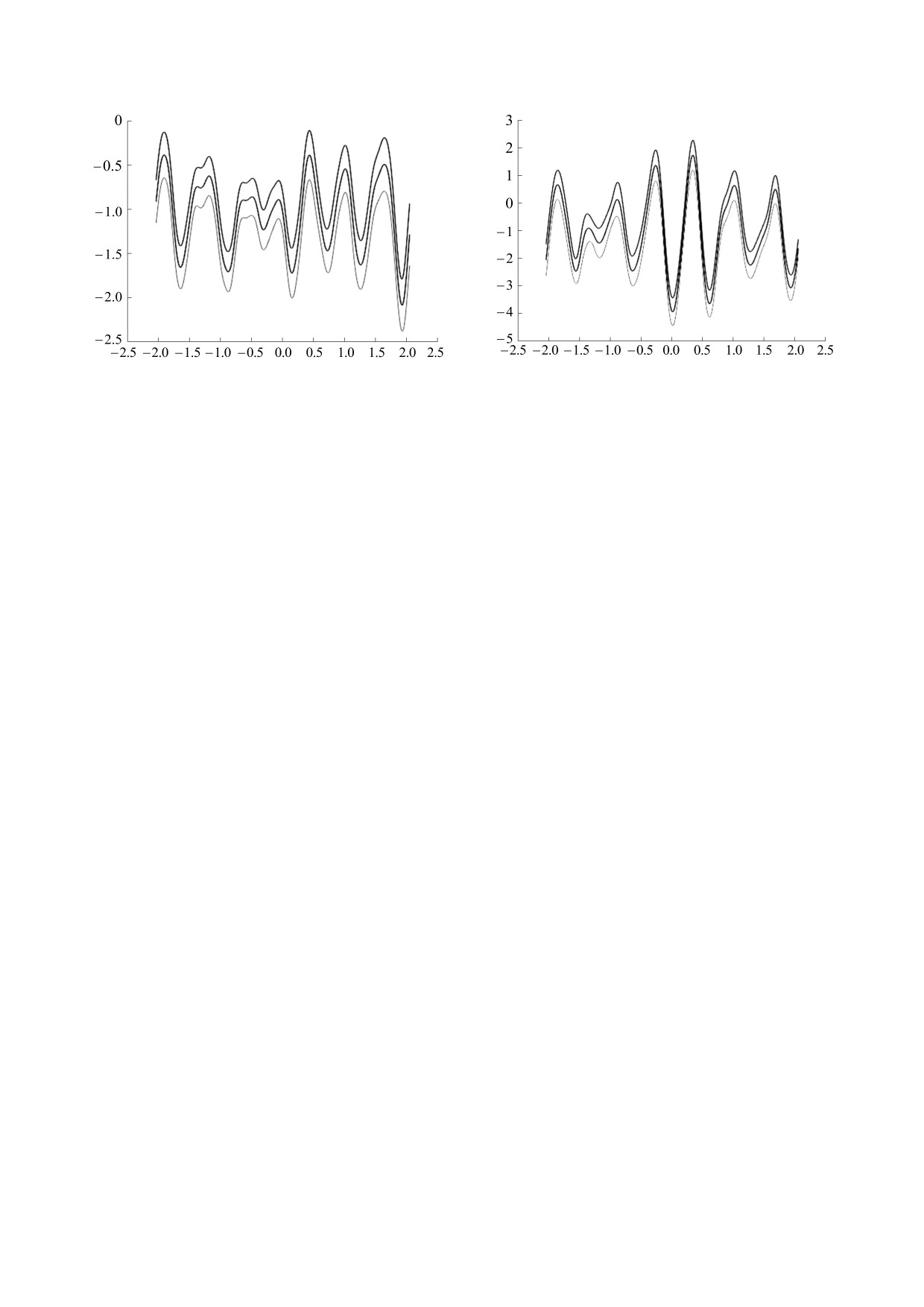
Рис. 5. Усреднение для второй стадии развития эпи-
Рис. 6. Усреднение для третьей стадии развития эпи-
лептической активности. Средние значения (цен-
лептической активности. Средние значения (цен-
тральная линия) и средние ± стандартное отклонение
тральная линия) и средние ± стандартное отклонение
(верхняя и нижняя линии) в мкВ, по оси абсцисс вре-
(верхняя и нижняя линии) в мкВ, по оси абсцисс вре-
мя в мс (нулевое значение соответствует максимуму
мя в мс (нулевое значение соответствует максимуму
мощности высокочастотной активности).
мощности высокочастотной активности).
гистрации электродами на скальпе при клиниче-
ми взаимодействия нейронов - через изменение
ской ЭЭГ, проявляется как «пик-волновый ком-
концентрации ионов в межнейронном простран-
плекс» или иные эпилептиформные феномены.
стве и через непосредственное электрическое
На четвертой стадии активность угасает, по при-
взаимодействие. Более же сложные межнейрон-
чине ли истощения или в силу появления им-
ные связи вообще не вовлекается в рассмотрение.
пульсов от иной структуры мозга, сбивающих
Но то, что при такой степени упрощения получа-
синхронизацию.
ют объяснение некоторые наблюдаемые явления,
позволяет надеяться, что при введении в рассмот-
Для проверки предположения о наличии син-
рение более сложных форм взаимодействия, уче-
хронизации такого рода было произведено усред-
те структуры нервной ткани и топологии нейрон-
нение медленноволновой активности, причем в
ной сети, включении в модель электрических и
качестве триггера усреднения были выбраны пи-
биохимических явлений не только в нейронах, но
ки на высокочастотной компоненте, превышаю-
и в глиальных структурах, данная модель может
щие пороговое значение. Усреднение проведено
быть доведена до полноценно объясняющей и
для отрезков как до, так и после пика, отдельно
практически применимой.
для второй (рис. 5) и третьей (рис. 6) стадий ис-
следуемого фрагмента электрокортикограммы,
Аргументом в пользу предлагаемой модели мо-
показаны средние значения и интервал в одно
жет служить связь нарушений электролитного ба-
стандартное отклонение.
ланса, прежде всего гипокальциемии, а также
Можно видеть, что разряд (соответствующий
гипонатриемии и гипомагниемии, с развитием
нулевому отсчету по оси Х) проявляется при вы-
эпилепсии [7]. Нарушение работы контура регу-
раженной негативной полярности низкочастот-
лирования вследствие недостатка кальция может
ной компоненты и способствует, в свою очередь,
приводить к появлению медленной активности,
развитию колебания низкой частоты, формируя
которая, в свою очередь, создает условия для по-
контур обратной связи, порождающий повторя-
явления высокочастотных вспышек, затем фор-
ющуюся активность.
мирующих острые волны.
Для проверки предложенной модели может
быть использовано сопоставление данных элек-
ОБСУЖДЕНИЕ
трокортикографии, скальповой электроэнцефа-
Предлагаемая модель, в силу ее крайне упро-
лографии, магнитоэнцефалографии и других ме-
щенного характера, может рассматриваться лишь
тодов. Это позволит проверить предположение о
как первое приближение к полноценно объясня-
том, что различие частотного состава сигнала, ре-
ющей явление во всей его полноте модели. Тем не
гистрируемого данными методиками, может объ-
менее она позволяет объяснить некоторые фено-
ясняться различием конфигурации источника
мены эпилептиформной активности, как и ЭЭГ
потенциала, а также рассмотреть формы синхро-
вообще. Основным ее недостатком следует при-
низации между собой сигналов, имеющих раз-
знать ограничение лишь элементарными форма-
личную частоту.
БИОФИЗИКА том 66
№ 4
2021
790
МАШЕРОВ
Неясным остается вопрос о механизме перехо-
КОНФЛИКТ ИНТЕРЕСОВ
да к четвертой фазе, прекращения приступа. Если
Автор заявляет об отсутствии конфликта инте-
синхронизация играет основную роль в его воз-
ресов.
никновении, то подача стимулов, ее нарушаю-
щих, может оказаться способом пресечения эпи-
припадка. Возможно, вошедший в клиническую
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
практику метод стимуляции блуждающего нерва
Настоящая работа не содержит описания ис-
[8] использует именно этот принцип для прекра-
следований с использованием людей и животных
щения приступа. В таком случае открывается воз-
можность новых методов лечения эпилепсии, с
в качестве объектов.
использованием вживляемых электродов или
магнитостимуляции области очага. Однако они
СПИСОК ЛИТЕРАТУРЫ
будут требовать выявления вспышек высокоча-
стотной активности и соответствующего измене-
1. В. А. Карлов, Эпилепсия у детей и взрослых женщин
ния методики регистрации сигнала, а также вы-
и мужчин (Медицина, М., 2010).
работки критериев автоматического распознава-
2. Clinical Electroencephalography, Ed. by O. Mecarelli
ния патологической активности в отличие от
(New York, Springer, 2019).
физиологической.
3. Нейрофизиологические исследования в клинике, под
ред. Е. М. Трошиной (НМИЦ нейрохирургии име-
БЛАГОДАРНОСТИ
ни академика Н. Н. Бурденко, М., 2019).
Автор выражает благодарность за помощь и
4. М. Н. Жадин, Биофизические механизмы формиро-
ценное обсуждение Н.А. Архиповой, Л.Б. Ивано-
вания электроэнцефалограммы (Наука, М., 1984).
ву, А.Б. Козловой, М.А. Куликову, В.И. Лукьяно-
5. Е. Л. Машеров, Биофизика 64 (3), 572 (2019).
ву, Л.Б. Окниной, К.Ю. Саркисовой, М.Ю. Челя-
6. N. E. Huang and S. S. P. Shen, Hilbert-Huang Trans-
пиной, Е.В. Шаровой.
form and its Applications (World Scientific, London,
2005).
ФИНАНСИРОВАНИЕ РАБОТЫ
7. M. A. Samuels and A. H. Ropper, Samuel’s Manual of
Neurologic Therapeutics (LWW, Philadelphia, 2017).
Работа выполнена при финансовой поддержке
Российского фонда фундаментальных исследова-
8. C.A. Edwards, A. Kouzani, K. L. Lee, and E. K. Ross,
ний (грант № 18-013-00967а).
Mayo Clinic Proc. 92 (9), 1427 (2017).
A Model of Activity Generation in the Epileptic Focus
E.L. Masherov
N.N. Burdenko National Scientific and Practical Center for Neurosurgery, Ministry of Healthcare of the Russian Federation,
4-ya Tverskaya-Yamskaya ul. 16, Moscow, 125047 Russia
A model of activity generation in the epileptic focus was proposed originating from assumption about synchro-
nization between oscillations which arise when the resting potential level is regulated, and synchronization be-
tween neuronal discharges with slow oscillations. At the onset of a seizure, four phases were identified - an in-
crease of slow wave activity, high-frequency spike activity, synchronization in the presence of sharp peaks, and
extinction of a seizure. This model was compared to electrocorticography data recorded during surgery.
Keywords: epilepsy, EEG, high-frequency activity, slow-wave activity, regulation, synchronization
БИОФИЗИКА том 66
№ 4
2021