БИОФИЗИКА, 2021, том 66, № 6, с. 1082-1115
БИОФИЗИКА КЛЕТКИ
УДК 577.3: 535.379
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
БИОЛОГИЧЕСКИХ ОБЪЕКТОВ.
II. МЕТОДЫ, ОСНОВАННЫЕ НА БИОЛОГИЧЕСКОМ
ДЕТЕКТИРОВАНИИ
© 2021 г. И.В. Володяев*,
*, И.И. Концевая**,
Л.В. Белоусов
А.Е. Наумова***, Е.В. Наумова****
*Московский государственный университет имени М.В. Ломоносова, 119991, Москва, Ленинские горы, 1
**Гомельский государственный университет им. Ф. Скорины,
246019, Гомель, ул. Советская, 104, Республика Беларусь
***Санкт-Петербургский государственный университет, 199034, Санкт-Петербург, Университетская наб., 7/9
****Институт физики полупроводников им. А.В. Ржанова СО РАН,
630090, Новосибирск, просп. Академика Лавpентьева, 13
E-mail: naumova@yandex.ru
Поступила в редакцию 07.06.2021 г.
После доработки 20.07.2021 г.
Принята к публикации 27.07.2021 г.
Во второй части обзора методов исследования сверхслабого свечения биологических объектов
представлена методология биологического детектирования. В течение первых 25 лет (1923-1948 гг.)
почти все исследования сверхслабого свечения проводились именно с биологическими детектора-
ми, причем регистрировалась только компонента, относящаяся к среднему ультрафиолетовому
диапазону (в ранних работах - митогенетическое излучение), которую обнаруживали по измене-
нию митотического режима детектора. Эти исследования остаются спорными из-за недостаточной
доказательности методик, но многие полученные результаты, революционные для того времени,
уже подтверждены, а научная проблематика остальных представляет значительный интерес (напри-
мер, фундаментальные результаты, касающиеся деления клеток и канцерогенеза, ранняя онкодиа-
гностика), поэтому анализ их методологии актуален для дальнейшей экспериментальной проверки
этих исследований. В обзоре приведен анализ основных методик с использованием в качестве де-
текторов корней лука, дрожжевых и бактериальных культур, а также проведено сопоставление ме-
тодики лукового детектирования с ближайшим современным аналогом (Allium-тест).
Ключевые слова: сверхслабое свечение биологических объектов, митогенетическое излучение, митогене-
тический эффект, биологические тест-системы, Allium-тест, биофотоника.
DOI: 10.31857/S0006302921060053
получил достаточно убедительного и полного
Этот раздел обзора посвящен методам, отно-
теоретического объяснения либо опровержения,
шение к которым в научном сообществе очень
также он не был ни опровергнут, ни доказан экс-
неоднозначно. Эти мнения, как правило, опира-
периментальными методами. В первые 25 лет по-
ются не на оригинальные статьи, а на их переска-
чти все масштабные исследования ССС проводи-
зы и личные мнения авторов, которые также не
лись именно с помощью методов биологического
читали оригинальных статей, и колеблются от
детектирования, и, cудя по значительному коли-
полного доверия до полного неприятия, не имея
честву верных результатов, опередивших время и
достаточной базы для научной аргументации.
подтвержденных только десятки лет спустя (на-
Митогенетический эффект, на котором было ос-
пример, существование пептидных онкомарке-
новано биологическое детектирование сверхсла-
ров крови), заявленные методики по крайней ме-
бого свечения (ССС), к настоящему времени не
ре в ряде научных школ достаточно эффективно
«работали». Вместе с тем с точки зрения совре-
Сокращения: ССС - сверхслабое свечение, МГЭ - митоге-
менного уровня доказательности эти методы био-
нетический эффект, УФ-ССС - сверхслабое свечение в
ультрафиолетовом диапазоне, МГИ - митогенетическое
логического детектирования имели существен-
излучение.
ные недостатки, а их известные описания недо-
1082
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1083
Рис. 1. Эксперимент А.Г. Гурвича с луком: (а) - по работе [2], (б) - по работе [3].
статочно полны для уверенного воспроизведения
Однако при биологическом детектировании ССС
без дополнительных исследований и анализа пер-
использовался только эффект стимуляции.
воисточников. Мы считаем важным представить
Митогенетическое излучение (МГИ)2 - компо-
подробный аналитический обзор методов, ис-
нента ССС, способная вызвать МГЭ в объекте-
пользованных в оригинальной литературе 1922-
реципиенте.
1948 гг., и привлечь внимание к работам, экспе-
риментальная проверка которых представляет
Реципиент митогенетического излучения3 -
значительный интерес как с точки зрения фунда-
клеточная культура или ткань, «компетентная» к
ментальной, так и прикладной науки. Для обзора
МГЭ, т.е. способная отреагировать проявлением
был использован уникальный архив научных
МГЭ на УФ-ССС.
публикаций, собранный А.Г. Гурвичем и его на-
Индуктор митогенетического излучения - объ-
учной династией.
ект-источник УФ-ССС, способный вызвать МГЭ
в биологическом реципиенте при оптическом
контакте и соблюдении ряда других необходимых
МИТОГЕНЕТИЧЕСКОЕ ИЗЛУЧЕНИЕ
условий.
И МИТОГЕНЕТИЧЕСКИЙ ЭФФЕКТ.
В обширной серии работ 1920-1940-x гг. (см.
ПОНЯТИЕ БИОЛОГИЧЕСКОГО ДЕТЕКТОРА
ниже) МГЭ был продемонстрирован на разнооб-
Определения. Митогенетический эффект
разных парах биологических индукторов и реци-
(МГЭ) - изменение режима клеточного деления
пиентов, были показаны широкая общность яв-
в культуре микроорганизмов, клеток или ткани
ления и отсутствие видоспецифичности эффекта.
под воздействием сверхслабого свечения в уль-
трафиолетовом диапазоне (УФ-ССС)1 другого
В классическом эксперименте А.Г. Гурвича [2]
биологического объекта, химической системы
использовались два луковых корня: один - в ка-
или физического источника.
честве источника ССС (индуктора), второй - в
качестве реципиента (детектора) (см. рис. 1). Вер-
Как правило с увеличением продолжительно-
хушку корня-индуктора направляли на зону деле-
сти воздействия сначала наблюдали стимуляцию,
ния детектора и после окончания такой индукции
затем эффект исчезал и далее наблюдали подав-
делали гистологические срезы зоны деления. Бы-
ление митозов (митогенетическая депрессия).
ло показано, что количество митотических фигур
на препарате со стороны, подвергнутой облуче-
1 Все работы 1920-1940-х гг. уверенно относили компоненту
нию ССС, достоверно выше, чем на противопо-
ССС, обуславливающую эффект (т.е. митогенетическое
излучение), к ультрафиолетовому диапазону. В последние
два десятилетия появились единичные работы, утвержда-
2 В старой литературе использовали также термины «лучи
ющие возможность сходных с митогенетическим эффек-
Гурвича», «митотические лучи».
том дистантных нехимических взаимодействий посред-
3 В «митогенетический» период часто использовали также
ством ССС видимого диапазона (см., например, работу
слово «детектор», но в рамках обзора мы будем так назы-
[1]). На этих эффектах не было построено методик биоло-
вать только реципиенты, использованные в качестве де-
гического детектирования физических параметров ССС, и
текторов излучения в общепринятом техническом смысле,
они не рассматриваются в обзоре.
например в устройствах для спектрального анализа ССС.
БИОФИЗИКА том 66
№ 6
2021
1084
ВОЛОДЯЕВ и др.
ванными» по отношению к контрольному состо-
янию) и «более или менее значительный клеточ-
ный комплекс, притом в достаточно близком со-
седстве клеток друг с другом» ([16], с. 332).
В первые годы после открытия эффекта наи-
более распространенным детектором оставались
корни лука. Однако «луковые» методики оказа-
лись трудоемкими, субъективными и требовали
большой затраты времени. Их сменили дрожже-
вые и бактериальные методики, которые были
проще, допускали количественную оценку эф-
фекта без субъективного и трудоемкого подсчета
под микроскопом определенных клеток, давали
лучшую воспроизводимость результатов, могли
использоваться круглый год. В СССР наиболее
широкое распространение получили дрожжевые
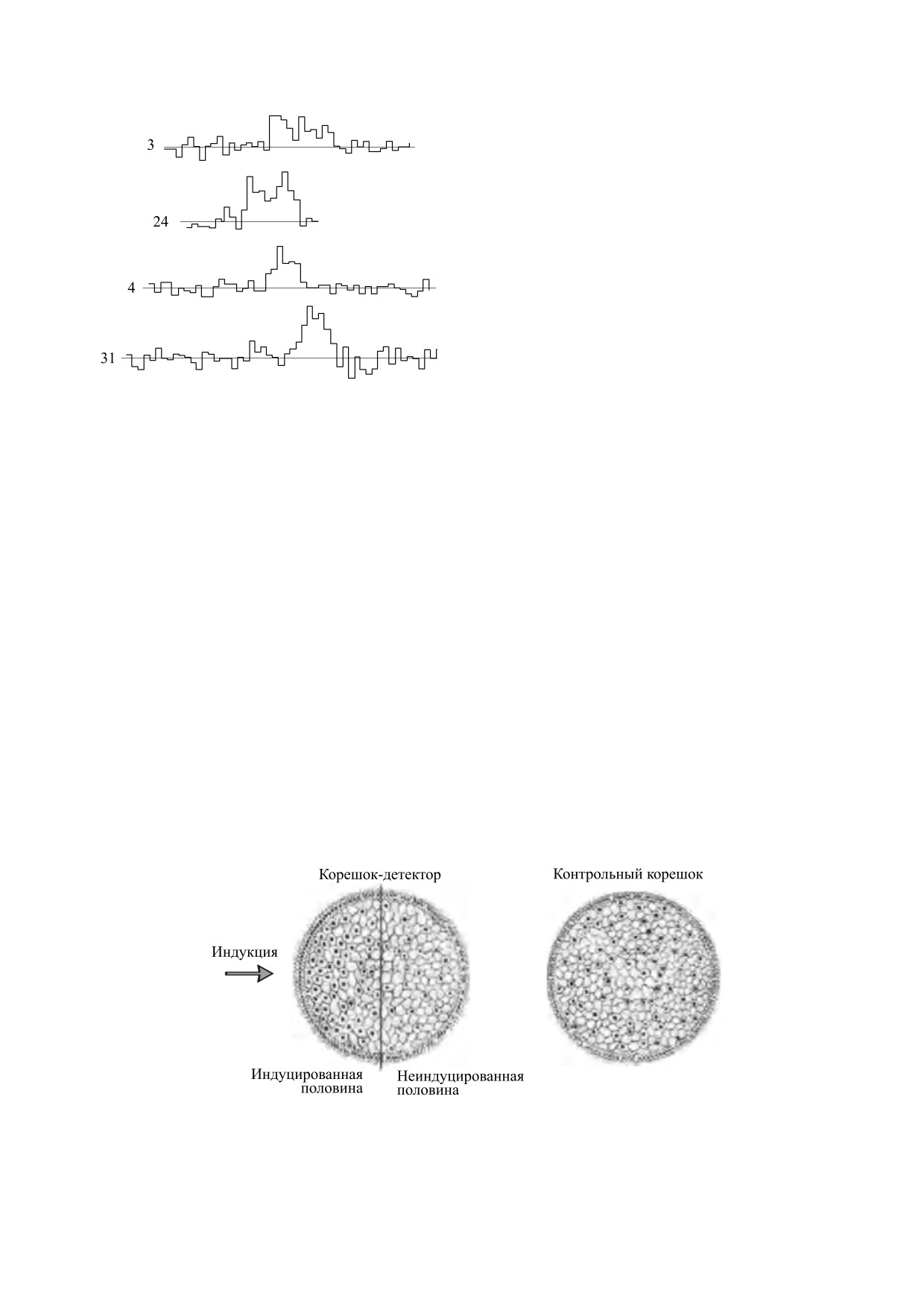
Рис. 2. Разница в количестве митозов между облучен-
детекторы [8, 16, 17], в западных странах - бакте-
ной и необлученной сторонами корня в последова-
риальные [5, 18].
тельных продольных сечениях (по работе [3]).
Несмотря на то что помимо этих реципиентов
МГЭ был продемонстрирован на целом ряде дру-
ложной стороне - именно и только в той области,
гих биообъектов, методики для большинства из
куда был направлен корень-индуктор (рис. 2 и 3).
них не были развиты и отработаны в такой мере,
чтобы уверенно использовать их в качестве детек-
Реципиенты митогенетического излучения. К
торов для фундаментальных или прикладных ис-
реципиентам МГЭ, согласно данным различных
следований ССС. Например, методика с рогови-
работ, относятся:
цей глаза лягушки [16], по данным авторов обес-
- дрожжевые и бактериальные культуры в лаг-
печивавшая очень надежные контроли (различие
периоде [5-7];
в числе митозов двух глаз одной лягушки не более
- «стареющие» дрожжевые культуры [8, 9];
1-3%), была весьма трудоемкой и применялась в
небольшом числе работ [16, 19, 20]. Помимо пере-
- некоторые ткани [10, 11];
численных, был разработан подход, не связанный
- меристема растений [2, 4];
с изменением митотического режима - он был
- сетчатка глаза (тритона, лягушки, крысы)
основан на изменении под действием ССС про-
[12]);
ницаемости биологических мембран (об этом су-
дили по динамике окрашивания раствора пиг-
- развивающиеся эмбрионы (морского ежа
[13, 14], лягушки [4]; дрозофилы [15]).
ментом лепестков цветов) [21], однако и он не по-
лучил распространения.
Общими чертами реципиентов являются
спонтанное (вне всяких воздействий) протекание
В настоящем обзоре подробно рассмотрены
клеточного деления, субоптимальные условия
только луковые (см. раздел «Allium-тест»), дрож-
роста [16] (т.е. «способность быть простимулиро-
жевые и бактериальные (раздел «Дрожжевые и
Рис. 3. Схематическое изображение поперечных срезов луковых меристем: (а) - облученный корень, (б) - необлучен-
ный корень (по работе [4]).
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1085
бактериальные методики детектирования мито-
точное количество почек или чтобы рост дрожже-
генетического излучения») детекторы.
вой или бактериальной культуры проявился в по-
Индукторы митогенетического эффекта. К ин-
мутнении, достаточном для его оценки нефело-
дукторам МГЭ относятся:
метром).
- культуры микроорганизмов [9, 13, 22-24] и
4. Количественная оценка параметра, характе-
клеток [16] в период активного роста;
ризующего эффект (например, для дрожжей это
мог быть подсчет абсолютного количества клеток
- меристема растений [2, 4];
в поле зрения или индекса почкования, измере-
- работающие мышцы [16, 25];
ние объема дрожжевой культуры мицетокрита-
- возбужденные нейроны [26];
ми4 или оценка степени помутнения культуры
- кровь здорового человека [27-29];
нефелометром или колориметром), статистиче-
- злокачественные опухоли (наиболее актив-
ская обработка результатов. В 1920-40-х гг. обыч-
ные из известных индукторов) [4, 20, 25, 30-34];
но эффект количественно характеризовали как
- ткани в процессе резорбции или регенера-
разницу в процентах между средними значения-
ции [35-38];
ми в экспериментальной и контрольной группах
(отмечая среднеквадратичное отклонение у каж-
- ряд химических реакций [39-41].
дой группы).
Также ряду авторов удалось получить МГЭ в
Условия детектирования. Оптический кон-
реципиентах с помощью многократно ослаблен-
такт. Оптический канал между индуктором и ре-
ного УФ-излучения от физических источников
ципиентом должен быть прозрачен для света с λ >
[42-46]. В работе [47] была исследована зависи-
190 нм. «Даже кварц можно использовать, только
мость величины МГЭ, наблюдаемого в дрожже-
если он очень высокой чистоты» [51].
вом детекторе, от длины волны и интенсивности
искусственного УФ-излучения.
Расстояние между индуктором и реципиентом.
Индукторами МГЭ не являются:
Оптимальное расстояние - от 1 до 10 мм; макси-
мальное расстояние, при котором наблюдается
- не растущие или медленно растущие культу-
МГЭ, зависит от индуктора и реципиента, дли-
ры клеток и микроорганизмов [16, 18];
тельности индукции и специальных условий, та-
- внутренние органы [18];
ких как «прерывистая индукция» (см. работу
- кровь пациентов с онкологическими [27-29]
[52]).
и некоторыми другими заболеваниями (анемия,
Длительность индукции. МГЭ имеет нелиней-
сепсис, пневмония, скарлатина) [29, 48], а также
ную дозовую зависимость с отчетливым эффек-
кровь пожилых, утомленных или истощенных
том подавления в области высоких доз [33, 53, 54]
людей [16, 48-50].
(см. рис. 4). Длительность индукции необходимо
специально подбирать для каждого нового набо-
ОБЩАЯ МЕТОДИЧЕСКАЯ СХЕМА
ра условий (по крайней мере в диапазоне 1-
БИОЛОГИЧЕСКОГО ДЕТЕКТИРОВАНИЯ
120 мин).
Примеры оптимальной длительности индук-
Общая последовательность детектирования ми-
ции, упоминаемые в литературе:
тогенетического эффекта. Общая последователь-
ность методики биологического детектирования
1) один-два часа (индуктор и реципиент - кор-
была следующей:
ни лука) [16];
1. Подготовка биологического детектора (как
2) 30 мин (индуктор и реципиент - дрожжевые
экспериментальной группы, так и контрольной),
культуры) [6];
т.е. приведение биологического детектора в над-
3) 15-30 мин (индуктор и реципиент - бакте-
лежащее физиологическое состояние (например,
риальные культуры). Индукция длительностью
проращивание корней лука, культивирование
60 мин давала нулевой эффект; индукция дли-
дрожжей или бактерий до нужной стадии в нуж-
тельностью 120 мин приводила к подавлению ро-
ных условиях и т.д.).
ста культуры [54].
2. Экспозиция (индукция) - приведение де-
Условия освещения. Индукцию надо проводить
текторов экспериментальной группы в оптиче-
на «рассеянном дневном свету» [57]. МГЭ не на-
ский контакт с объектом, ССС которого исследу-
блюдается ни на ярком свету, ни в полной темно-
ется.
те [16].
3. Период развития/проявления эффекта (по-
МГЭ не наблюдается при наличии внешнего
сле вывода из оптического контакта с индуктором
ультрафиолетового излучения уже при относи-
детектор требует некоторого времени, чтобы сти-
тельно малых интенсивностях. Так, МГЭ пропа-
муляция деления проявилась в достаточной сте-
пени для количественной оценки - например,
4 Мицетокрит - сосуд для точного измерения объема жид-
чтобы в дрожжевой культуре образовалось доста-
кой культуры.
БИОФИЗИКА том 66
№ 6
2021
1086
ВОЛОДЯЕВ и др.
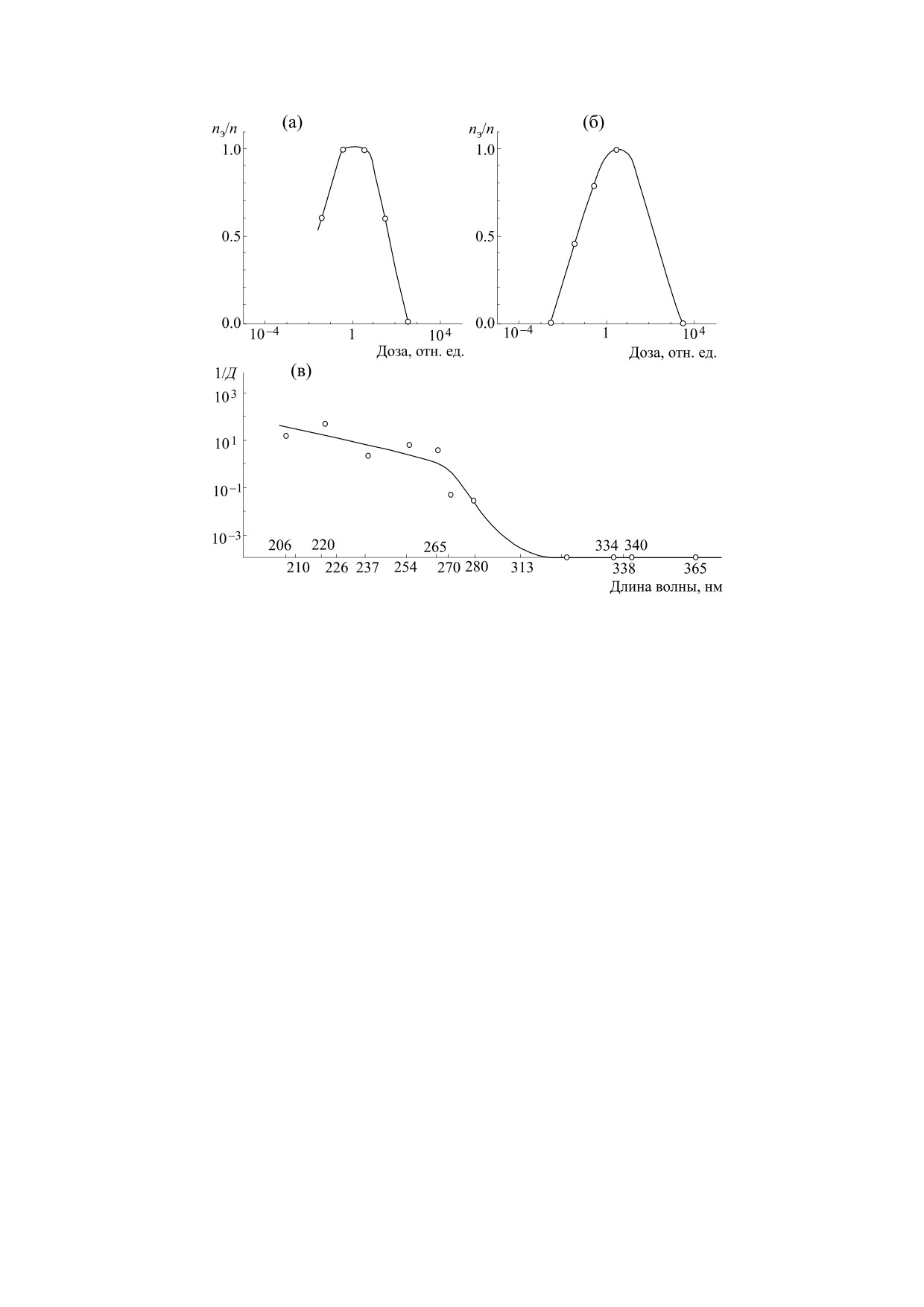
Рис. 4. Дозовые кривые и спектр действия митогенетического излучения: (а) - зависимость вероятности индукции
почкования дрожжевых клеток от дозы при длине волны 206 нм, пэ — число опытов с положительным
митогенетическим эффектом; п — общее число опытов; (б) - то же, но при длине волны 220 нм; (в) - спектр действия,
Д - доза, соответствующая максимальному митогенетическому эффекту. (Из работы [55], построено по данным
работы [56].)
дал в присутствии газовой горелки, в солнечный
2. Следует принимать во внимание отражение,
день при открытых окнах лаборатории и пр., но
поглощение и рассеяние излучения в стекле,
наблюдался при закрытых двойных ставнях в глу-
кварце и других материалах конструкции, по-
бине комнаты, т.е. при фильтрации двумя окон-
скольку они могут значительно снизить эффект
ными стеклами [16].
или дать артефактную стимуляцию в контроле.
Динамика развития эффекта после индукции.
3. Большинство экспериментов выполняли в
МГЭ проявляется не сразу. Так, если регистриро-
аэробных условиях, но без специального насы-
вать изменение плотности культуры, то макси-
щения атмосферы (или среды) каким-либо газом.
мальный эффект наблюдается через один-четыре
Несколько попыток получить МГЭ в анаэробных
часа после окончания индукции [7, 54]; если ре-
условиях не увенчались успехом [16].
гистрировать индекс почкования (на дрожжевых
4. Было отмечено, что временной характер
культурах), то через 30-120 мин [6]. Однако если
приведения реципиента в оптический контакт с
учитывать только почки меньше 1/10 размера ма-
исследуемым индуктором имел существенное
теринской клетки, то МГЭ заметен уже через 5-
значение - более резкая подача ССС к реципиен-
10 мин после окончания индукции [40, 58]. Было
ту, в том числе многократная прерывистая подача
показано, что время развития эффекта суще-
с помощью вращающихся дисков с прорезями,
ственно зависит от продолжительности индукции
была эффективнее, в то время как слишком мед-
[58].
ленное увеличение интенсивности ССС, напри-
Некоторые специальные условия. 1. Суспензи-
мер путем плавного сближения с индуктором до
онные культуры можно индуцировать только в
обычного расстояния наблюдения МГЭ, нивели-
очень тонком слое (<0.5 мм) [18, 54]. «Более тол-
ровало эффект. Например, при плавном прибли-
стые слои суспензии поглощают излучение и пол-
жении к источнику в течение 5-6 мин реципиент
ностью убирают эффект» [18].
не реагировал, тогда как при ускорении сближе-
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1087
ния до 3 мин детектирование было стабильно [16,
шем
эксперименты
по
методикам,
33].
предложенным М. Пол, не проводились, а данная
методическая работа была проигнорирована в по-
следующих критических обзорах - возможно, от-
ЛУКОВЫЕ МЕТОДИКИ ДЕТЕКТИРОВАНИЯ
части из-за отсутствия достаточного статистиче-
МИТОГЕНЕТИЧЕСКОГО ИЗЛУЧЕНИЯ
ского материала, отчасти из-за того, что на смену
«Основной опыт» Гурвича. А.Г. Гурвич и его со-
луковым уже пришли более простые дрожжевые и
трудники с 1922 по 1928 гг. провели ряд серий экс-
бактериальные детекторы [81].
периментов с корешками лука обыкновенного
Ложноположительные результаты. Результаты
(Allium сера) в качестве детектора в разных моди-
работ [73, 82] с корнями Allium cepa и Vicia faba,
фикациях, в том числе классический вариант, ко-
подтверждающие МГЭ, А.Г. Гурвич расценивал
гда излучателем выступали тоже корни лука [59,
как неубедительные с методической точки зре-
60].
Опыты проводили в лабораториях
ния, в частности он подверг критике использова-
А.Г. Гурвича в Симферополе, Москве и Ленин-
ние в качестве детекторов двудольных растений и
граде [61] с разными сортами лука при разных
особенно корней бобовых вместо лука [62, 78, 83].
температурных, световых и временных условиях.
Общее количество таких «луковых» эксперимен-
Критика результатов Гурвича. Среди исследо-
тов по разным данным было свыше 150 [60] или
ваний, которые не подтвердили результаты
около 300 [59, 62], причем А.Г. Гурвич упоминал
А.Г. Гурвича, наиболее значимыми являются ра-
только два отрицательных результата, получен-
боты Б. Россманна
[84]5. В ответ на них
ных в его лаборатории, которые объяснял посто-
А.Г. Гурвич опубликовал более подробные про-
ронними причинами [3, 63]. Т. Рейтер и Д. Габор
цедуры и множество статистических материалов,
выполнили более 200 опытов с биологическим
подтверждающих его результаты, а также отме-
детектированием УФ-ССС (из них 125 опытов с
тил, что автор не повторял его эксперименталь-
луковыми детекторами), представили подробные
ные условия и неудачно выбрал излучающий био-
методики и протоколы экспериментов [4], в част-
объект (корень гороха) [86-89].
ности, они использовали луковые детекторы в
Сильная критика относительно существова-
экспериментах с отражением, преломлением и
ния митогенетических лучей и выводов Гурвича
спектральным разложением МГИ (см. раздел
по
«луковому» опыту была высказана
«Эксперименты, подтверждающие, что МГЭ обу-
Б.П. Токиным [90], который, тем не менее, ссы-
словлен электромагнитным излучением биообъ-
лается в своем резюме не на положительные ре-
ектов в УФ-диапазоне»).
зультаты нескольких сотен опытов своих сотруд-
С помощью «луковой» методики было проде-
ников, а на критические работы [91-95]. Ключе-
монстрировано МГИ различных биообъектов,
вой аргумент Б.П. Токина о возможности
причем значительное внимание уделялось выс-
альтернативного объяснения МГЭ не излучени-
шим растениям - были исследованы корни лука,
ем, а стимуляцией митозов парами эфирного
гороха, подсолнечника, гиацинта, донце лука,
масла, испускаемыми донцем луковицы, несо-
первые листья подсолнечника, семядоли подсол-
стоятелен, поскольку не охватывает эксперимен-
нечника, клубни и лептомы картофеля, кашицы
ты с химической изоляцией, эксперименты с ис-
из желтой репы, донца лука, листьев Sedum и др.
точниками, не имеющими подобного химическо-
[2, 8, 16, 45, 64-74]; было также продемонстриро-
го фактора; отсутствуют также доказательства
вано МГИ бактерий Bacterium tumefaciens [75, 76],
влияния на митозы такой малой разницы в кон-
дрожжевых культур [66], злокачественных опухо-
центрации паров, которая возникает между облу-
лей [30].
чаемой и тыльной сторонами корня.
В публикациях были изложены методики по-
становки эксперимента [2, 4, 67, 77-79], описаны
В известной критической работе [91]6 авторы
(с предоставлением фотографий) разные устрой-
на основании своих экспериментов сделали вы-
ства, приспособления и материалы [3, 78, 80].
вод, что корень лука как детектор МГИ не может
Позже была опубликована выдающаяся по мето-
быть использован из-за значительной асиммет-
дической проработке статья М. Пол по луковым
рии в количестве делящихся клеток между поло-
детекторам [71], в которой были проанализирова-
винками корня (до 50% в их экспериментах), и
ны опыты других исследователей, учтена вся кри-
оценили результаты школы А.Г. Гурвича как не-
тика и предложены новые подходы для исключе-
корректные из-за отсутствия при каждой новой
ния возможных артефактов, в частности, разра-
ботан метод по унификации отдельных фаз
5 См. также современное издание [85].
6 Следует отметить, что авторы работы [91] признают воз-
митоза. Методика была опробована всего на не-
можность стимуляции митозов слабым внешним УФ, т.е.
скольких экспериментах, результаты которых,
принципиальную возможность биообъектов детектировать
несмотря на методические отличия, согласовыва-
УФ-излучение, но отрицают само существование УФ-из-
лись с данными школы А.Г. Гурвича. В дальней-
лучения у биообъектов.
БИОФИЗИКА том 66
№ 6
2021
1088
ВОЛОДЯЕВ и др.
постановке опыта отдельного отрицательного
A. cepa считается эталонным растительным тест-
контроля.
объектом для анализа мутагенности, митотоксич-
ности и токсичности различных факторов [103,
М.Н. Моисеева многократно повторила ос-
119].
новной «луковый» опыт, придерживаясь, по ее
Широкое распространение тесты с A. cepa по-
словам, в мельчайших подробностях методики
А.Г. Гурвича [92-96]. Ее замечания были в основ-
лучили из-за того, что они являются более чув-
ном направлены на присутствие внешних факто-
ствительными и простыми по сравнению с биоте-
ров (механических воздействий, света, тепла, га-
стами животных9, несмотря на большие различия
зообразных химических веществ и т.п.), влияние
в организации и метаболизме между раститель-
которых якобы совершенно игнорировалось
ными и животными клетками, многими авторами
школой А.Г. Гурвича. При этом, судя по доступ-
сделан вывод о возможности экстраполировать
ным материалам, автор включала в критический
полученные результаты A. cepa на животные орга-
анализ только те работы А.Г. Гурвича, которые
низмы, включая человека [100, 102, 116, 118-142].
подтверждали ее мнение. В частности, в основ-
Кроме этого, лук обыкновенный толерантен к
ных работах А.Г. Гурвича неоднократно было
разным условиям проращивания (рН от 3.5 до
подчеркнуто сверхосторожное обращение с кор-
11.0) и доступен практически круглогодично [102,
нями [17, 83], применение специального оборудо-
143, 144]. В работе [100] суммированы оригиналь-
вания и материалов, использование авторских
ный и модифицированный варианты Allium-те-
механических конструкций для предотвращения
ста, в работах [129, 145-147] перечислены основ-
незапланированных отклонений в положении
ные значимые подходы постановки опыта.
биообъектов.
Вопросы методики и стандартизации Allium-
Allium-тест. Растительная тест-система Alli-
теста поднимались в единичных публикациях,
um cepa L. Allium-тест был разработан в 1938 г.
например [148]. Проанализировав более 200 ра-
[97-99] и стандартизован в 1980-х годах [100-
бот по Allium-тесту, мы выяснили, что мнения
102]. Общий подход в работе с луком обыкновен-
исследователей в отношении постановки теста и
ным остается аналогичным предложенному
условий инкубации материала сильно различают-
А.Г. Гурвичем, хотя методика значительно про-
ся. Ниже мы сравниваем общие этапы гурвичев-
ще. Экспериментальная группа луковиц с проро-
ской луковой методики детекции УФ-ССС и Alli-
щенными корнями7 подвергается воздействию
um-теста для выяснения наиболее значимых
исследуемых факторов/условий, затем для оцен-
условий проведения опытов, которые требуют
ки их влияния определяются морфологические и
стандартизации и, возможно, дополнительных
микроскопические параметры корней (в частно-
исследований.
сти, одним из факторов, к которым чувствителен
Реализация отдельных этапов Allium-теста.
Allium-тест, является УФ-излучение). Этот тест
1. Сорт и размер луковицы. В отличие от методик
является наиболее близким современным анало-
лукового детектирования МГИ, исследователи
гом лукового детектора среди биологических те-
Allium-теста не акцентируют внимание на сезон-
стов, прошедших стандартизацию и активно ис-
ных эффектах в постановке опыта и не всегда
пользующихся на практике в настоящее время8.
упоминают сорт лука, с которым проводят тести-
Наряду с Allium cepa L. используются и другие
рование [101, 118, 128, 133, 148-168]. В тех случаях,
тест-объекты (среди растений - наиболее часто
когда упоминают название сорта, это, как прави-
Pisum sativum L. (горох), Vicia faba L. (бобы), Hor-
ло, лук «Штуттгартен Ризен» [144, 146, 169-175].
deum vulgare L. (ячмень), Zea mays L. (кукуруза))
Размер луковиц в работах большинства иссле-
[118]. Тем не менее в современных исследованиях
дователей небольшой: диаметр 1.5-2.0 см, вес не
более 3-4 г. В единичных работах применяли лу-
7 Корни предварительно проращивают в так называемом
ковицы больших размеров: весом до 15-30 г [165],
оригинальном варианте Аллиум-теста, который наиболее
удобен при тестировании физических факторов. В моди-
40-60 г [167].
фицированном варианте теста луковицы помещают непо-
2. Среда для проращивания. Проращивание как
средственно в исследуемый раствор без предварительного
проращивания корешков, модифицированный вариант
правило выполняли на воде неопределенного со-
чаще используется при тестировании химических веществ
става: «пригодной для питья» [129, 147], водопро-
и здесь не рассматривается.
водной или фильтрованной водопроводной [129,
8 О широком применении лука обыкновенного свидетель-
130, 146, 154, 176]. Редко использовали дистилли-
ствует ряд примеров тестирования химических и физиче-
ских агентов: антибиотики
[103,
104], антиоксиданты
рованную воду [132, 141, 147, 158, 172-174, 177-
[105], гербициды [106], грибковые экстракты [107], инсек-
тициды [108], лекарственные растения [109], наночастицы
9 Это подтверждено в совместных международных исследо-
[110], промышленные отходы [111], пестициды [112], ра-
ваниях в рамках Программы ООН по окружающей среде,
диоактивность [113], синтетические растительные гормо-
Всемирной организации здравоохранения, Международ-
ны [114], тяжелые металлы [115], ядовитые растения [116,
ной программы о химической безопасности, Агентства по
117].
охране окружающей среды США [118, 119].
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1089
186] и крайне редко - специальные питательные
Предварительные эксперименты в работе [71]
среды либо деионизированную воду [148].
подтверждают предположение о влиянии сроков
постановки опыта, но результаты показали хоро-
В основных протоколах Allium-теста указана
шее прорастание луковиц с марта по ноябрь. Так-
обязательная смена воды либо тестируемой жид-
же влияние времени года на постановку индукци-
кости. Эксперименты, описанные в работе [187],
онного опыта отмечали и другие исследователи
свидетельствуют, что цикл деления клеток силь-
[18, 193, 195].
но зависит от парциального давления кислорода
как фактора окружающей среды, и авторы счи-
В современном исследовании с использовани-
тают, что митотические стадии, на которые наи-
ем Allium-теста на сорте «Штуттгартен Ризен»
более негативно влияет низкое давление кисло-
[143] была подтверждена сезонность чувствитель-
рода - это метафаза и анафаза.
ности лука [143]. В ряде работ предложен способ
синхронизации и активизации деления клеток
В работе [188] проанализированы клеточные
[177, 196, 197].
реакции на повреждения, вызванные оксидатив-
ным стрессом.
Размер луковиц. Выбор объекта исследования
выполняется на двух этапах: 1) при отборе луко-
3. Температура. Проращивание обычно вы-
виц для проращивания придаточных корней;
полняли при комнатной температуре, которая
2) при отборе используемых в опыте корней
колебалась, в зависимости от времени года и кли-
[70, 78].
матических условий региона, от 20 до 30°С [118,
А.Г. Гурвич отдавал предпочтение большим
129, 144, 153-155, 159, 160, 162, 172, 177, 179, 185,
(многолетним) луковицам диаметром 5 см и бо-
186, 189]. В единичных работах температура под-
лее, поскольку именно они дают «сильные, мито-
держивалась на одном уровне (22 или 25°С) с ис-
тически активные корни» [78]. В отличие от это-
пользованием фитотронов или иных инкубаторов
го, в работах по Allium-тесту предпочтение в ос-
[153-155, 172, 179, 180].
новном отдается более мелким луковицам (см.
4. Освещенность. По отношению к световому
параграф «Реализация отдельных этапов Allium-
режиму при постановке теста мнение исследова-
теста»).
телей также различалось. Большинство тестов
Среда для проращивания лука. Для проращива-
выполняли при естественном освещении (при
ния луковиц во всех известных нам работах по
условии защиты от прямых солнечных лучей),
МГЭ использовали водопроводную отстоянную
очень редко устанавливали 12-часовой световой
воду. Отмечалось, что воду следует менять каж-
цикл [180]. В некоторых работах особо подчерк-
дый день [70, 78], рекомендовано было погружать
нуто, что луковицы должны оставаться в темноте
в воду только подошву лука на глубину примерно
100% времени [147, 155, 163, 190, 191].
1 см. В работах по Allium-тесту этот вопрос также
5. Время фиксации. Корни лука проращивают
мало стандартизован (см. параграф «Реализация
до достижения ими длины 1-2 см [129, 192].
отдельных этапов Allium-теста).
В единичных работах указано время фикса-
Температура. Практически все эксперименты
ции: с 8:00 до 9:00 [143, 177] [149] и в 11:00
по МГЭ проводились при нестандартизируемой
[109,164].
комнатной температуре. При этом А.Г. Гурвич
Анализ этапов индукционного «классического»
подчеркивал, что предпочтительна «умеренная
лукового опыта. Проанализируем этапы индукци-
комнатная температура» (не выше 15°С) [78], а в
онного «классического» лукового опыта в работах
работе [70] автором была указана средняя ком-
по МГЭ.
натная температура от 18°С в зимний сезон до
26°С летом.
Cорт лука и сезонность. Ни исследователями,
ни критиками практически не упомянуты ис-
В своем аналитическом обзоре применительно
пользуемые в опыте сорта лука. Мимолетно дан-
к дрожжевым культурам O. Ран писал: «К сожале-
ный вопрос был затронут в работе [70] и чуть по-
нию, не было заявлено, что 12°C считается нор-
дробнее - в работе [193]. Только в двух работах,
мальной комнатной температурой в Москве и Ле-
использующих биологическое детектирование,
нинграде, и исследователи, следуя [методиче-
была отмечена значимость сорта используемого
ским] указаниям
[авторов] буквально и при
американских комнатных температурах, несо-
материала - картофеля [74] и лука10 [66].
мненно, получили бы совершенно другую физио-
Также А.Г. и Л.Д. Гурвичи подчеркивают, что
логическую стадию» [198]. По-видимому, это так-
для того, чтобы объективно обнаружить индук-
же справедливо и для луковых экспериментов.
цию, необходимо выполнять эксперимент с зим-
В работе [71] М. Пол подчеркивала необходи-
ним и ранневесенним материалом [2, 63, 78, 194].
мость стабилизации внешних условий с начала
образования корня до конца индукционного экс-
10Были исследованы три коммерческих сорта, распростра-
ненных на рынках Нью-Йорка - испанский, техасский
перимента, чтобы исключить по возможности
красный и техасский белый.
факторы, которые могли повлиять или размыть
БИОФИЗИКА том 66
№ 6
2021
1090
ВОЛОДЯЕВ и др.
небольшой длины [78], но полагал, что получение
эффекта при этом затруднено из-за существенно-
го влияния собственного МГИ-детектора [67].
Для экспериментов выбирали «крепкий, иде-
ально прямой» корень, «идеально сформирован-
ный и с наиболее симметричным кончиком» [17,
78, 203]. При этом луковицу и остальные корни
обычно обрезали, так что экспериментальный
корень контактировал только с частью луковицы
или небольшим кусочком донца.
Автор работы [204], напротив, оставлял луко-
вицу целой, но также удалял оставшиеся корни; в
работах [4, 205] в качестве детектора был исполь-
зован отрезанный корень лука.
В работах по Allium-тесту как правило исполь-
зовались более короткие корни (1-2 см) - см. па-
раграф «Реализация отдельных этапов Allium-те-
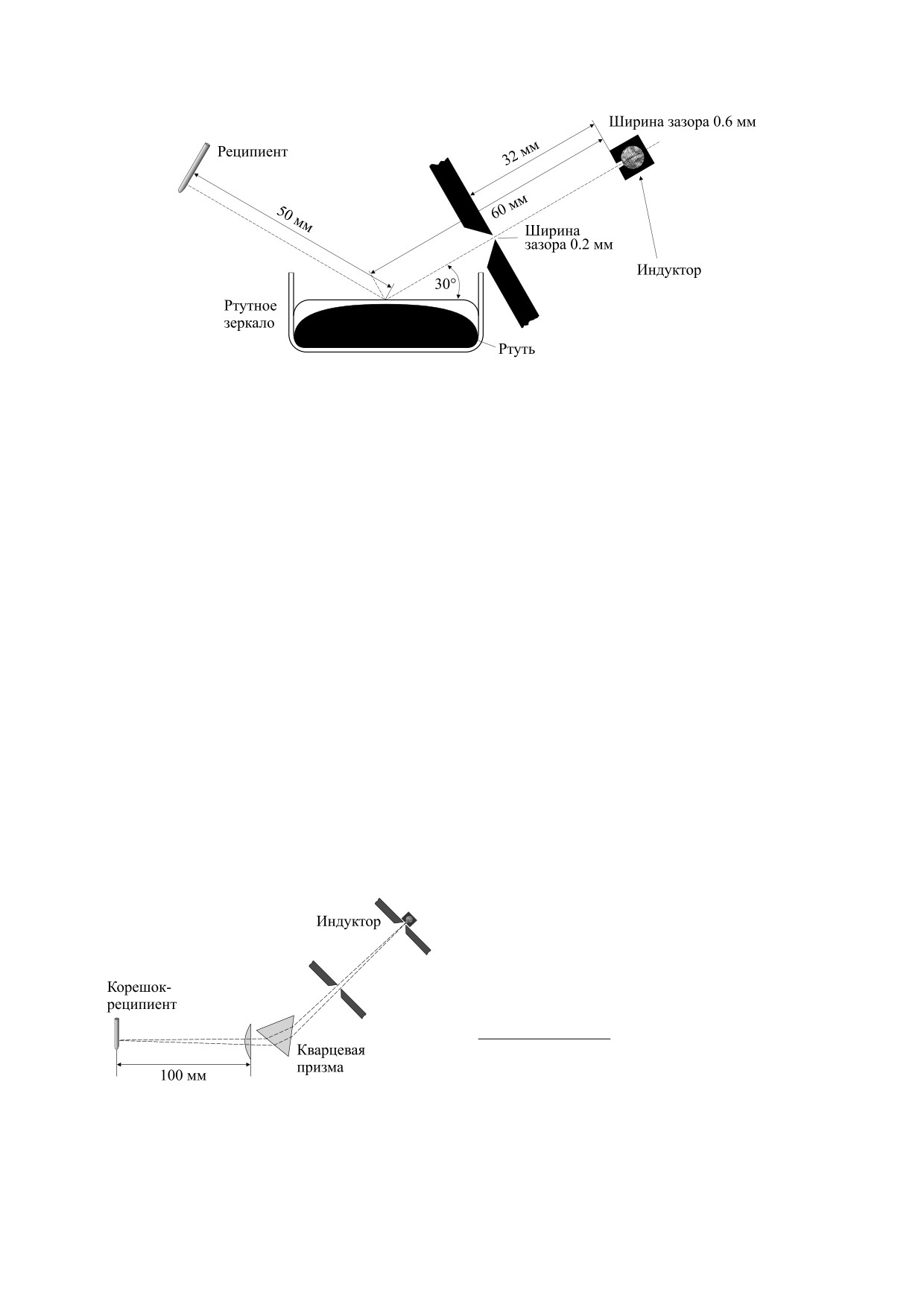
Рис. 5. Установка по получению МГЭ на корнях лука
ста».
(из работы [4]).
Экспериментальная установка. Особое внима-
ние авторы уделяли экспериментальной установ-
результаты эксперимента. При этом были учтены
ке (см. рис. 5).
работы о тормозящем влиянии света на деление
А.Г. Гурвич подчеркивал, что максимальная
[199-201] и о влиянии температуры на продолжи-
иммобилизация корня детектора абсолютно не-
тельность митоза в корнях лука [202].
обходима для успеха эксперимента [78]. Для этого
автор помещал корень в стеклянный капилляр, а
Освещенность. Согласно данным работ
центрирование корня в капилляре проводил с по-
А.Г. Гурвича [59, 78], при апробировании разно-
мощью микроскопа [2].
образных условий нормального освещения и за-
темнения было доказано отсутствие влияния
У других авторов, за исключением работы [82],
естественного и искусственного освещения на
расположение двух корней во время эксперимен-
скорость роста корней, регулярность формы кор-
та всегда было таким же. Аналогичные установки
ня, обилие митозов и радиальную симметрию ми-
использованы в работах [4, 92, 70] и др. При этом,
тозов внутри корня. В то же время автор ссылает-
по мнению М.Н. Моисеевой [96], подобные кон-
ся на доказательства негативного влияния темно-
струкции могут приводить к артефактам за счет
вых условий культивирования корней лука в
неравномерного прикосновения корня к капил-
работах [4, 73, 82, 85]. Ряд авторов особо подчер-
ляру и механостимулированных митозов в этой
кивает необходимость полной защиты от возмож-
части корня11.
ного внешнего ультрафиолетового излучения
Во избежание подобных эффектов автор рабо-
(см. главу «Условия детектирования» раздела
ты [82] проращивал часть корней свободно в воде
«Общая методическая схема биологического де-
или во влажном воздухе (используя цинковый
тектирования»).
ящик с небольшим окошком). Позже M. Пол
Следует подчеркнуть, что по отношению к
[71], придерживаясь методики работы [82], ис-
световому режиму при постановке Allium-теста
пользовала только свободно растущие корни, без
мнение исследователей также различалось - от
размещения их в капиллярах.
использования естественного освещения до стро-
Расстояние от индуктора до детектора. Рас-
гого соблюдения 12-часовой периодичности или
стояние от индуктора до детектора в опытах Гур-
полной темноты (см. параграф «Реализация от-
вича составляло от 1.5-2.0 до 12 мм [2, 67], в
дельных этапов Allium-теста»).
успешных опытах других исследователей - до
В работе [71] специально отмечалась необхо-
38 мм (максимальное испытанное расстояние)
димость стабилизации режима освещенности для
[206]. Не только воздух, но и вода, тонкая кварце-
корректной работы с луковым детектором.
вая пластина и даже стекло в очень тонких пла-
стинках (толщиной не более 50 мкм), размещен-
Выбор корней для опыта. А.Г. Гурвич рекомен-
ное между индуктором и детектором, не пред-
довал проращивать луковицы до появления кор-
ставляют собой препятствия для МГИ; более
ней длиной не менее 6-7 см и не более 18 см [67];
позднее верхний предел был снижен до 10-12 см
11 Следует отметить, что неизбежные многократные прикос-
[17]. В то же время отдельные исследователи ра-
новения корня к внутренней поверхности трубки скорее
ботали с корнями длиной от 1 до 4 см [70, 71, 82].
могут увеличить разброс результатов, чем вносить систе-
А.Г. Гурвич также допускал применение корней
матическую ошибку.
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1091
толстый слой стекла задерживал МГИ и нивели-
В работах [4, 71, 84, 204], напротив, предпри-
ровал МГЭ [4, 65].
нимались попытки выявить регулярные колеба-
ния в делениях клеток меристемы корней лука,
Локализация эффекта вдоль корня. Согласно
вслед за работами [199, 200, 202, 211]. Тем не ме-
данным А.Г. Гурвича [207], эффект индукции ло-
нее из-за неоднозначности полученных результа-
кализован латерально, но распространяется про-
тов авторы не смогли подтвердить эти наблю-
дольно на расстояние до 250 мкм [208] (см.
дения.
рис. 2). По результатам авторов работы [4] в про-
дольном направлении эффект индукции ограни-
Проблема изучения ритмичности клеточных
чен примерно 150 мкм. В работах [73,82] наблю-
делений у растений представляет повышенный
дался более диффузный эффект в латеральном
интерес [200, 211-220], поскольку до сих пор не
направлении (возможно, из-за недостаточно
решен вопрос ни о видоспецифичности данного
жесткой фиксации корней во время индукции).
признака, ни об определяющих его механизмах
Следует отметить, что при длительных индук-
[213, 217] (см. обобщение результатов исследова-
циях начинал играть роль собственный рост кор-
ний ритмов клеточных делений меристемы в ра-
ня. Так, за три часа индукции корень мог вырасти
ботах [213, 217]). Несмотря на многочисленные
на 1.5-2.0 мм [67].
данные по суточной ритмике митозов в меристе-
мах побегов и корней, результаты разных авторов
Продолжительность индукции. Продолжитель-
часто не согласуются и даже противоречат друг
ность индукции, как уже рассмотрено в главе
другу. В том числе различны и данные по Allium
«Условия детектирования» раздела «Общая мето-
cepa [145, 169, 187, 202, 211, 214, 216, 218, 219, 221-
дическая схема биологического детектирования»,
233]. Величина митотического индекса стабили-
необходимо подбирать для каждого нового набо-
зируется в корнях лука при длине 2-4 см и оста-
ра условий проведения эксперимента. В научной
ется постоянной в процессе дальнейшего роста
школе А.Г. Гурвича продолжительность индук-
[234].
ции составляла как правило 30-40 мин и более
[78], вплоть до одного-двух часов [16]. Согласно
С нашей точки зрения и исследователям фено-
данным работ [209] и [82], МГЭ постоянен при
мена МГЭ, и специалистам, работающим с Alli-
воздействии в течение 0.5-3.0 ч и 2-6 ч соответ-
um-тестом, следовало тщательнее исследовать су-
ственно; согласно работе [53], эффект становится
точную периодичность митозов и учитывать эти
резко отрицательным при непрерывной индук-
данные в экспериментах по МГЭ.
ции меристемы лука в течение 12-26 ч несколько
Фиксация и окрашивание. Согласно методике
раз обновляемыми культурами дрожжей («мито-
школы А.Г. Гурвича, обработку корешков выпол-
генетическое истощение» или «депрессия»). По
няли фиксатором Боуэна на протяжении трех-
данным работы [70], после первых двух часов ин-
пяти часов [78]. Однако в соответствии с данными
дукции наблюдается существенное увеличение
последних лет фиксаторы Боуэна, Кларка, Кар-
числа делящихся клеток меристемы; в дальней-
нуа, а также формалин приводят к продольной,
шем (через 4-5 ч) наступает явная депрессия, т.е.
поперечной и объемной деформации (сжатию)
падение числа митозов на индуцированной сто-
[235]. Также отмечено мацерирующее действие
роне.
фиксатора Боуэна на апексы корня. Эти данные
Продолжительность развития эффекта. После
объясняют проблемы, с которыми столкнулась
окончания индукции корни отставляются друг от
группа А.Г. Гурвича [78] при подсчете делящихся
друга - требуется некоторое время на развитие
клеток.
эффекта [2, 18, 210]. Это время обычно составляет
от 30 мин до 2 ч [6]. Согласно данным работы [4],
Согласно рекомендациям, данным в работе
[63], для подсчета митозов подходят только об-
эффект может быть обнаружен через час после
окончания индукции (возможно, что и раньше) и
разцы, резко окрашенные железным гематокси-
наблюдается по крайней мере еще пять часов по-
лином и столь же резко дифференцированные, в
сле индукции. В то же время M. Пол в большин-
идеале толщиной 10 мкм: «ядра покоя» (согласно
стве случаев фиксацию корешков проводила сра-
сегодняшней терминологии и пониманию - ядра
в интерфазе) должны быть обесцвечены насколь-
зу после окончания индукции [71].
ко возможно, чтобы можно было точно иденти-
Время фиксации корней. А.Г. Гурвич неодно-
фицировать ранние стадии профазы, когда нити
кратно упоминал неоднородность корней и их
хроматина четко окрашиваются.
разную митотическую активность на момент
фиксации [20, 59, 78, 79], при этом попытки вы-
Техника подсчета митозов. Технике подсчета
яснения суточной периодичности митозов в кон-
митозов в школе А.Г. Гурвича уделялось особое
трольных образцах, по-видимому, не предприни-
внимание [78]. В основу были положены следую-
мались. В работах [78, 79] упоминается время
щие основные правила: 1) каждый срез всегда
фиксации образцов (10:00), однако без объясне-
считается несколько раз, и из них берут среднее
ния причин.
[2, 83, 89]; 2) каждый счет, насколько это возмож-
БИОФИЗИКА том 66
№ 6
2021
1092
ВОЛОДЯЕВ и др.
но, пересчитывается другим сотрудником без
захватывал 3-4 клеточных пласта периблемы
знания результатов первого.
[83].
Приведем основные приемы в методике мик-
M. Пол [71] также проводила подсчет во всех
роскопирования,
используемые
школой
тканях конуса нарастания корня, включая калип-
А.Г. Гурвича и другими исследователями.
троген и дерматоген, за исключением клеток кор-
1. Гистологические срезы. Изучали в основном
невого чехлика.
продольные срезы толщиной 10 мкм, параллель-
В негативных работах методика подсчета отли-
ные направлению индукции, что позволяло четко
чалась: в работе [82] не учитывались митозы в пле-
дифференцировать митозы [3, 78, 236]. Метод по-
роме, а автор работ [84, 204] при подсчетах игнори-
перечных срезов был использован в работе [4]
ровал дерматоген и дополнительно калиптроген и
(рис. 3). В опытах M. Пол и поперечные, и про-
корневой чехлик.
дольные срезы показали наличие МГЭ, но автор
По вопросу о распределении митозов по цито-
отметила преимущество продольных срезов (про-
гистологическим зонам корня в литературе суще-
дольная ось ядра и митозов обычно ориентирова-
ствуют противоречивые сведения. Наибольшая
на вдоль оси корня, поэтому на продольных сре-
доля делящихся клеток отмечена в дерматогене и
зах гораздо легче различать отдельные стадии,
периферических слоях периблемы в работах [183,
чем на поперечных).
227, 240, 241], другие данные показывают наи-
Если была идентифицирована
«клеточная
большую митотическую активность плеромы
слизь» центральных столбцов, затрагивающих
[171, 242, 243] или утверждается отсутствие суще-
оставшуюся часть плеромы, а тем более перибле-
ственной разницы в распределении митозов по
му и дерматоген, то счет не проводили [78].
зонам меристемы [244].
При подсчетах делящихся клеток предвари-
3. Фазы клеточного цикла. Важным моментом
тельно определяли среднюю плоскость [3] (в ра-
методологии, как подчеркивалось в работе [70],
боте [204] были отмечены проблемы с ее опреде-
является разграничение фаз митоза, подлежащих
лением). Митозы в каждой секции подсчитыва-
подсчету. По этому вопросу мнения исследовате-
ются слева и справа от средней линии каждой
лей также различались [71]. Согласно рекоменда-
секции. По данным микроскопических измере-
циям А.Г. Гурвича [89], необходимо подсчиты-
ний на продольных срезах, длина верхушечной
вать митозы от самой ранней профазы до поздней
меристемы составляет 800-1200 мкм, длина кор-
телофазы в двух половинах каждой секции [63].
невого чехлика - 200-300 мкм [71, 222, 237].
При этом разделительная линия фиксируется пу-
Было подчеркнуто, что эффект получается
тем осмотра при малом увеличении. В работах
всегда на пяти-семи медиальных (центральных)
[84, 204] автор критиковал учет профазы, кото-
[238] и парамедиальных срезах из десяти, сделан-
рую трудно отличить от ядер в состоянии покоя,
ных подряд, т.е. на протяжении приблизительно
считая включение этой фазы основным источни-
60-80 мкм вдоль корня, если индукция происхо-
ком ошибки при подсчетах12. Тем не менее он
дит на расстоянии от 2 до 38 мм [2, 3, 8, 17, 83,
упоминал, что в ряде опытов при подсчете профаз
206]. При соприкосновении корней возникала
он также получил доминирование клеток с мито-
более широкая полоса индукции (около 180 мкм)
зами на индуцированной стороне корня. Соглас-
[67]. В большинстве поздних опытов сотрудники
но представлениям А.Г. Гурвича [78, 83, 89], эф-
Гурвича ограничивались подсчетом числа мито-
фект индукции, напротив, проявлялся в основ-
зов в 10-15 медиальных срезах (по 10 мкм) [2, 83,
ном в ранней профазе, не учитываемой в работах
194].
[84, 204], и именно эти методические расхожде-
Примеры исследований с негативными ре-
ния были ответственны за отрицательные резуль-
зультатами: в работах [93, 94, 96, 239] авторы под-
таты. Кроме того, по данным А.Г. Гурвича [89]
считывали именно периферические зоны одина-
40% от общего избытка митозов находится в дер-
ковой ширины по обе стороны от средней плос-
матогене, который в указанных работах игнори-
кости; в работе [82] подсчитывались все секции
рован.
корня; автор работы [204] также исследовал ко-
О большей продолжительности профазы по
рень целиком, а затем сравнивал с цифрами, под-
сравнению с другими стадиями митоза свиде-
считанными на центральных срезах.
тельствуют работы [221, 233, 245, 246]. Установле-
2. Ткани. А.Г. Гурвич рекомендовал проводить
но, что суточный митотический цикл Allium cepa
подсчет делящихся клеток от самого кончика
корня до зоны растяжения, учет при подсчете
12 При этом автор работы [204], высказывая сомнение в соб-
всех слоев конуса нарастания из-за их плохой
ственном правильном определении клеток на стадии про-
фазы, критиковал А.Г. Гурвича за небрежность и отсут-
дифференциации: дерматогена и периблемы,
ствие четких критериев подсчета. После этой критики
плеромы, корневого чехлика, клеток калиптроге-
А.Г. Гурвич опубликовал дополнительные подробности
на [78]. При этом основной эффект индукции об-
процедуры и множество статистических материалов, под-
наруживался на дерматогене и субдерматогене и
тверждающих его результаты [88, 89, 203].
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1093
включает 54.14 ± 1.07% интерфазы, 39.07 ± 0.99%
существующие протоколы о луковом корне в ка-
профазы, 2.39 ± 0.26% метафазы, 1.56 ± 0.26%
честве детектора и максимально объективно ин-
анафазы и 2.77 ± 0.38% телофазы [246]. Таким об-
терпретировать полученные данные. Аналогич-
разом, утверждаемое А.Г. Гурвичем влияние
ное мнение было высказано авторами работы [4].
МГИ на раннюю профазу митоза действительно
При этом исключаются только протоколы, в ко-
должно регистрироваться в первую очередь при
торых экспериментальные ошибки (или неточно-
подсчете ядер в профазе и может нивелироваться
сти в подсчетах) не только предполагались, часто
при их исключении из подсчета.
безответственно, но и фактически доказывались.
Такое статистическое исследование было пред-
Методика, описанная в работе [4], существен-
принято в работе [247]. Ее автор графически отоб-
но отличается - авторы считали «зрелые ядра» в
разил
все
результаты,
опубликованные
дополнение к митозам в обычном смысле, в итоге
А.Г. Гурвичем
и его сотрудниками (около
их подсчеты нельзя сопоставить с результатами
200 опытов) (рис. 6).
других специалистов [70, 71].
Л.В. Белоусов [248] специально подчеркивает,
Контроль. Исследования распределения мито-
что новаторским в исследованиях А.Г. Гурвича
зов в интактных корнях выполняли А.Г. Гурвич
было то, что он использовал статистическую об-
[78, 83], М.Н. Моисеева [93, 94, 96] и еще ряд ав-
работку картин распределения клеточных деле-
торов. А.Г. Гурвич заявлял, что общее количество
ний в различных растительных и животных тка-
митозов в двух половинах корня отличается мак-
нях.
симум на 5%, М.Н. Моисеева обнаружила почти
полную симметрию. В работе [239] были отмече-
Надо отметить, что исследователи Allium-те-
ны колебания не более 4%. Напротив, подсчет в
ста также подчеркивали внутриклоновую вариа-
работе [204] показал разницу в ±10%, а в работах
бельность митотического индекса Allium cepa
[73, 82] были обнаружены колебания до ±15%.
как в контрольном, так и в экспериментальном
Значительные различия обнаружил и автор рабо-
материале, и высказывали предположение о
ты [205]. Однако следует подчеркнуть, что методы
внутривидовой вариабельности признака [119,
подсчета у всех авторов были разные.
148, 249]. При проведении Allium-теста многие
исследователи следуют рекомендациям работ
М.Н. Моисеева [93, 94, 96] и авторы работы
[102, 129], используя для микроскопического ана-
[91] оценивают как некорректные результаты
лиза отбор проб из пяти луковиц по пять корней
экспериментов и школы Гурвича, и ряда других
на луковицу; другие следуют «статистическому
исследователей (например, представленные в ра-
стандарту» работы [249]: три луковицы и три кор-
боте [66]) из-за отсутствия отдельного отрица-
ня на луковицу.
тельного контроля при каждой новой постановке
Возможности «модернизации» методики луково-
опыта. В то же время А.Г. Гурвич отмечает следу-
го детектора в целях повышения ее убедительности.
ющее: «Специальные контроли со свободно рас-
На основании данных по разнородности корней
тущими корешками нами не предпринимались,
отчасти потому, что на основании долголетней
лука в работе [18] был поднят вопрос о корректно-
предшествовавшей работы с ними мне достаточ-
сти их использования в качестве детекторов; в ка-
честве более надежных детекторов предлагались
но была известна их симметрия», «…за нашу про-
дрожжевые и бактериальные культуры (см.
должительную работу у нас получился богатый
ниже).
материал опытов с «нулевым эффектом», могу-
щим служить наилучшим контролем первона-
Аналогичный вопрос ставили и исследователи
чальной симметрии корешков» [83]. Делается
Allium-теста. Тем не менее было отмечено [148],
примечание, что в первых сериях (около 40 опы-
что это не компрометирует именно лук в качестве
тов) выполняли подсчет очень большого количе-
тест-системы, поскольку описанная вариация ти-
ства срезов каждого корешка и убедились в бес-
пична для любого биологического материала.
полезности этой длительной процедуры [83].
Тщательный анализ тестового материала, соот-
ветствующая экспериментальная процедура и
Статистика. Разнородность корней в смысле
статистический анализ полученных результатов
количества митозов, даже в пределах одной луко-
позволяют использовать тест с низким риском
вицы, отмечена в работах А.Г. Гурвича [20, 67],
ошибок.
O. Рана [18] и др. (в то же время использование
«идеально симметричных корней» позволяло по-
Лук успешно использовался в качестве детек-
лучать очень близкие значения в пределах одного
тора для регистрации МГИ тканевых культур рас-
корня - см. выше). Тем не менее, несмотря на ма-
тительного или животного происхождения [89],
лое применение статистических методов в биоло-
резорбированных и регенерирующих тканей [38,
гии того времени, авторы признавали необходи-
89], культур микроорганизмов [66, 75, 76], злока-
мость статистического анализа результатов. По
чественных опухолей [30]. Следует подчеркнуть,
мнению А.Г. Гурвича [89], единственно правиль-
что индукторами не могут выступать не растущие
ное и подходящее решение - это обобщить все
или медленно растущие культуры [16, 203, 250]
БИОФИЗИКА том 66
№ 6
2021
1094
ВОЛОДЯЕВ и др.
Рис. 6. Обобщенные результаты А.Г. Гурвича и соавторов с корнем лука в качестве детектора. По оси абсцисс - общее
количество митозов; по оси ординат - эффект увеличения количества митозов относительно контроля. Кружки ука-
зывают на положительную индукцию, черные точки - на отсутствие митогенетического эффекта. Линией указаны
пределы погрешности. (Построено по данным работы [247].)
(см. главу «Условия детектирования» раздела
- использовать только крупные луковицы;
«Общая методическая схема биологического де-
- отбирать корни с длиной не менее 6 см;
тектирования»), в то время как детектор на пери-
- использовать методику подсчета митозов,
од индукции должен быть в неоптимальных усло-
предложенную А.Г. Гурвичем.
виях роста и обладать пониженной митотической
Из перечисленного значимым «белым пят-
активностью. Таким образом, в «луковом методе»
ном» является знание суточной периодичности
должны использоваться детектор и индуктор с
митозов конкретных сортов лука. Более того, для
противоположными характеристиками в отно-
шении клеточного цикла. Данный критерий не
циклов Allium cepa существует значимое взаимо-
действие между митотическими стадиями и пери-
был учтен ни одним из исследователей, посколь-
одами фиксации материала. Данная зависимость
ку в период активных экспериментов с использо-
свидетельствует о том, что необходимо отрабаты-
ванием лука обыкновенного (1920-е годы) пред-
вать не только время фиксации материала для ци-
ставления о клеточном и митотическом цикле
были минимальными, противоречивыми, часто
тологического исследования, но и время поста-
новки луковиц на проращивание.
неверными. Именно современные знания о ми-
тотическом цикле [251] помогают объяснить от-
дельные подходы в «луковом» методе, например
ДРОЖЖЕВЫЕ И БАКТЕРИАЛЬНЫЕ
«развитие эффекта», а также свидетельствуют о
МЕТОДИКИ ДЕТЕКТИРОВАНИЯ
том, что ошибочно применять одновременно
МИТОГЕНЕТИЧЕСКОГО ИЗЛУЧЕНИЯ
корни лука одного сорта в качестве индуктора и
Впервые МГЭ на дрожжевых культурах был
детектора13.
показан в работе [8], на бактериальных - в работе
Мы полагаем, что можно сформулировать сле-
[5]. Результаты этих авторов были подтверждены,
дующие рекомендации для «лукового метода»:
например, в работах [6, 23, 252] и подверглись
- работать с конкретным сортом лука, предва-
критике в работах [253, 254]. Среди наиболее по-
рительно выяснив максимумы и минимумы су-
дробных обзоров МГЭ на культурах микроорга-
точной митотической активности;
низмов следует отметить работы [16, 18, 20, 40];
- проращивать в естественных световых усло-
наиболее значимыми критическими обзорами
виях;
являются работы [51, 81, 255]. В основном в СССР
применяли дрожжевые детекторы, на Западе -
13 Сведения о суточных максимумах митотической активно-
бактериальные. В целом наиболее широкое рас-
сти недостаточны и противоречивы. Вполне вероятно, что
пространение получили дрожжевые детекторы.
даже при использовании в качестве индуктора и детектора
разных сортов нельзя подобрать время суток, оптимальное
Особенности методик детектирования сверхсла-
для высокой эффективности и индуктора, и детектора.
бого свечения с помощью дрожжевых и бактериаль-
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1095
ных культур. Фаза роста. МГЭ был показан на
Характеризация митогенетического эффекта в
дрожжевых и бактериальных культурах в лаг-пе-
дрожжевых и бактериальных культурах. Для ха-
риоде [6, 7, 54], или в периоде пост-диауксии (не-
рактеризации МГЭ в дрожжевом детекторе, ин-
известном в то время) [8]. В первом случае для по-
кубируемом на плотной агаризованной среде,
лучения реципиентной культуры необходимо ис-
пользовались методом подсчета абсолютного ко-
пользовать инокулят в фазе пост-диауксии
личества клеток в поле зрения и индекса почкова-
(дрожжевые культуры в возрасте 1-6 суток, и луч-
ния, т.е. количества почек, отнесенного к общему
ше агаровые, чем суспензионные [18], или бакте-
количеству клеток (размер учитываемых почек
риальные двух-четырехсуточные культуры [5, 7,
также варьировал в разных работах [16]).
54]). На более молодых (в период активного ро-
Для оценки роста суспензионной культуры ис-
ста) или более старых (G0) культурах МГЭ не на-
пользовали, кроме подсчета клеток, и физиче-
блюдается [6, 18].
ские методы. Например, прирост объема культу-
ры оценивали с помощью мицетокритов, сосудов
Период чувствительности. Любая культура-ре-
для точного измерения объемов (так называемый
ципиент имеет период чувствительности («окно
объемный метод Брайнеса) [49], плотность кле-
рецептивности») к МГЭ, зависящий от возраста
культуры, взятой для засева. Чем ближе к стацио-
ток культуры характеризовали оптическим мето-
дом - с помощью нефелометров [252] или коло-
нарной фазе используемый инокулят, тем позже
риметров [256].
«открывается» окно рецептивности у полученной
из него культуры микроорганизма. Так, дрожже-
Методики использования дрожжевых детекто-
вые культуры в лаг-периоде, полученные из одно-
ров наиболее подробно изложены в работах [9, 16]
суточной культуры, чувствительны к индукции в
и в диссертациях по онкодиагностике 1940-х го-
период до одного часа после засева; культуры, по-
дов [28, 257, 258]. Бактериальные методики опи-
лученные из шестисуточного инокулята - в пери-
саны в работах [5, 7, 259]. В методической статье
од 2.0-2.5 ч после его внесения [6]. На культурах,
по бактериальному детектированию [259] отмеча-
полученных из десятисуточной исходной суспен-
ется, что по сравнению с дрожжами бактериаль-
зии, МГЭ не наблюдается [6]. Бактериальные
ные детекторы дают более выраженный эффект,
культуры, инокулированные двух-четырехсуточ-
но менее стабильный.
ным посевным материалом, чувствительны сразу
Негативные работы и контркритика. Ниже при-
после посева на питательную среду [54]. В другом
ведены методические особенности основных
варианте индуцировали исходный двух-четырех-
критических работ (в скобках курсивом даны на-
суточный инокулят, после чего высевали его на
ши комментарии; ссылки и методику «позитив-
питательную среду [7].
ных работ» см. выше).
Плотность культуры. МГЭ не наблюдается на
Ричардс, Тейлор, 1932 [254]. Основные отличия
«слишком густых» [18] или «слишком разбавлен-
условий эксперимента от рекомендованных в
ных» культурах [16].
«позитивных работах»:
1. Использованы суспензионные культуры
Рекомендованная плотность культуры состав-
S. cerevisiae, без указания фазы роста (исходя из
ляет:
условий, описанных авторами, культура реципи-
- 20000 кл/мл (стафилококк, суспензионная
ента находилась в фазе экспоненциального ро-
культура) [5];
ста). (МГЭ наблюдается только в лаг-период или в
период пост-диауксии. Авторы процитировали эту
- 50-5000 кл/мл (E. coli, суспензионная куль-
рекомендацию, но предпочли отклониться от нее.)
тура) [7];
2. Использовали культуральную среду с опти-
- отдельные клетки на плотной агаризованной
мальным составом и температуру инкубации
среде, не образующие групп (S. cerevisiae, агаро-
28°C. (МГЭ наблюдается в субоптимальных усло-
вая культура): «Клетки должны быть достаточно
виях, включая «обедненный» состав питательной
далеко друг от друга, чтобы не влиять друг на дру-
среды и пониженную температуру культивирова-
га» ([18], c. 68).
ния.)
Условия роста культуры. МГЭ не наблюдается
3. Культуру реципиента помещали в пробирки
на «стандартных» культуральных средах. Наилуч-
объемом 1.5-2.0 мл и фиксировали их в большом
ший эффект достигается при использовании
контейнере с культурой индуктора. (МГЭ в сус-
обедненной среды («стандартный бульон, разбав-
пензионных культурах наблюдается только в очень
ленный в 10 раз» [7]), однако некоторые результа-
тонких слоях (<0.5 мм).)
ты получены, наоборот, на среде с избыточным
4. Индукция продолжалась от 2 до 24 ч. (Дрож-
количеством субстрата («18-баллинговое сусло»
жевые культуры демонстрируют достоверный
[40]).
МГЭ при длительности индукции 15-60 мин. Более
Эффект лучше проявляется при пониженной
длительная индукция приводит к подавлению эф-
температуре [16].
фекта.)
БИОФИЗИКА том 66
№ 6
2021
1096
ВОЛОДЯЕВ и др.
5. Плотность культуры измеряли непосред-
чатках, делали строго восемь пересевов культуры
ственно после индукции. (Индукция приводит к
перед началом эксперимента (конкретное число
изменению плотности культуры через один-четыре
пересевов никак не объясняется), использовали
часа после ее окончания.)
заземление для кварцевых пластинок и даже для
самих себя. К сожалению, огромная работа, про-
Так как в работе сделан ряд критических нару-
шений в методике, результаты авторов, по наше-
деланная авторами, также не имеет отношения к
му мнению, не могут использоваться для провер-
феномену МГЭ.
ки МГЭ.
Кройхен, Бейтман,
1932
[260], Накайдзуми,
Голлендер, 1937 [253]. Основные отличия усло-
Шрайбер, 1931 [261], Вестенберг, 1935 [262]. В этих
вий эксперимента от рекомендованных в «пози-
критических работах также нарушен целый ряд
тивных работах»:
рекомендованных условий. Так, в работе [261] ис-
пользовали культуры дрожжей в фазе экспонен-
1. В качестве инокулята (для реципиента) ис-
циального роста (9-12 ч при 25°C); в работе [260]
пользовали агаровые культуры в возрасте 15-19 ч
использовали культуру реципиента в концентра-
(позже авторы стали использовать 39-часовые
ции на два-три порядка выше, чем рекомендова-
культуры). (Культуры E. coli приобретают чув-
но в «позитивных работах». К сожалению, нам не
ствительность к индукции после 48 ч роста (при
удалось найти оригинал работы [262], однако в
37C) и, в любом случае, после окончания периода
монографии [59] разобрана методика этой работы
активного роста. Здесь кривые роста не приведены,
с указанием критических нарушений.
а возраст культуры выбран на основании посторон-
них соображений.)
Попытки повторить МГЭ в «новое время». Бы-
2. Посевную культуру смывали с агаризован-
ли попытки воспроизведения МГЭ в дрожжевых
культурах, индуцированного дрожжевой культу-
ной среды солевым раствором (NaCl, KCl, CaCl2 и
рой [263] и слабым искусственным УФ-излучени-
вода). Полученную суспензию клеток помещали
ем [264] после того, как авторы данных работ по-
в чашки диаметром 2 см и высотой 1-2 см и инду-
лучили в 1970-1980-е гг. достоверные результаты
цировали от 5 с до 12 мин при 37°C и постоянном
по физической регистрации ССС в УФ-диапа-
перемешивании. (МГЭ в суспензионных культурах
зоне.
наблюдается только в очень тонких слоях
(<0.5 мм); авторы работ [6,54] специально отмеча-
К сожалению, данные авторы не имели ин-
ли, что МГЭ невозможно обнаружить в толстом
формации об условиях, необходимых для получе-
слое суспензии детектора.
ния эффекта, и сделали ряд критических ошибок
в методике. Ниже приведены использованные
МГЭ на бактериальных культурах наблюдается
ими условия (в скобках курсивом даны наши за-
при длительности индукции 15-30 мин. В любом
мечания):
случае, для любых новых параметров эксперимента
необходимо подбирать длительность индукции в
1. В качестве индуктора использовали дипло-
диапазоне по крайней мере от 1 мин до 2 ч. В «пози-
идный лабораторный штамм S. cerevisiae (фаза
тивных работах» этот параметр всегда тщатель-
экспоненциального роста).
но отрабатывали.
2. Культура реципиента была: (1) в стационар-
Использованный солевой раствор - крайне спе-
ной фазе (G0 - 10 суток при 28°C и насыщении
цифическая среда для индукции. Нет ни одной рабо-
кислородом) или (2) в лаг-периоде - сразу после
ты с использованием подобных сред.
высева инокулята на свежую питательную среду.
Постоянное перемешивание суспензии с реципи-
(МГЭ не наблюдается на стационарных культурах
ентом - также крайне специфическое условие, до
или культурах, высеянных из них. Работать можно
того ни разу не проверявшееся.)
только с культурами в области пост-диауксии.)
3. Культуру реципиента использовали в кон-
3. Использовали в качестве реципиента сус-
центрации (1.5-2.0)⋅105 кл/мл. (В работе [54]
пензионные культуры при 28°C, продуваемые
кислородом. (МГЭ наблюдается в субоптимальных
использовали концентрацию 2104 кл/мл; в работе
условиях, включая пониженную температуру. Про-
[7] - 50-5000 кл/мл. При больших концентрациях
дувание кислородом никогда не использовалось в
эффект не наблюдался.)
«позитивных работах». Ранее подобные условия ис-
Таким образом, авторы использовали совер-
пользовали только в работе [253], в которой МГЭ
шенно новые условия эксперимента без проверки
не был зафиксирован.)
и оптимизации. Некоторые параметры однознач-
4. Индукцию проводили с реципиентом в про-
но противоречили рекомендациям «позитивных»
бирках объемом 10 мл. (МГЭ на суспензионных
работ (возраст посевной культуры, толщина слоя
культурах наблюдается только в очень тонких сло-
реципиента при индукции, плотность культуры
ях (<0.5 мм).)
реципиента). В то же время авторы очень тща-
тельно подошли к стандартизации второстепен-
5. Индукция начиналась сразу после (или не-
ных условий: работали только в резиновых пер-
посредственно перед) высева инокулята на све-
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1097
жую питательную среду. (Культура-реципиент
чувствительна к индукции только в определенный
период («окно рецептивности»), который зависит
от физиологического состояния инокулята. Авторы
никак не проверяли этот параметр.)
6. Единственное проверенное время индукции
было 30 мин. (Длительность индукции необходимо
оптимизировать по крайней мере в диапазоне 1-
120 мин для любых новых условий.)
Таким образом, авторы работы [263] использо-
вали совершенно новые условия, без оптимиза-
ции ключевых параметров эксперимента. В неко-
торых случаях использованные условия прямо
противоречили рекомендациям «ранних авторов»
(возраст культуры-реципиента, «окно рецептив-
Рис. 7. Эксперимент, подтверждающий прямолиней-
ное распространение и нехимическую природу фак-
ности», метод индукции).
тора, обусловливающего МГЭ. Индуктор - кусочек
головки лука, размещен в герметично закрытой квар-
Попытки воспроизвести МГЭ предприняли
цевой пробирке, реципиент - корень лука. (Из рабо-
еще в ряде лабораторий (см., например, работы
ты [4], с небольшими изменениями.)
[265, 266]). Авторам удалось получить хорошие
результаты, но не удалось воспроизвести эффект
в повторных сериях экспериментов [266]. Судя по
ОСНОВНЫЕ СХЕМЫ ИССЛЕДОВАНИЯ
описанной ими методике, авторы не были знако-
СВЕРХСЛАБОГО СВЕЧЕНИЯ
мы с рекомендациями «ранних работ» и оставили
В УЛЬТРАФИОЛЕТОВОМ ДИАПАЗОНЕ
без внимания ряд важных параметров экспери-
С БИОЛОГИЧЕСКИМИ ДЕТЕКТОРАМИ
мента.
Эксперименты, подтверждающие, что митогене-
К подобным работам, недостаточно полно
тический эффект обусловлен электромагнитным
учитывающим рекомендации ранних авторов,
излучением биообъектов в ультрафиолетовом диа-
можно отнести и нашу собственную (И.В. и Л.Б.)
пазоне. Первые эксперименты после открытия
попытку повторить МГЭ на дрожжевых культурах
МГЭ были направлены на исследование обуслав-
в 2010-2013 гг. Изучив проблематику по книгам и
ливающего его фактора. Было установлено, что
обзорным статьям (И.В.), мы попытались вос-
эффект имеет не химическую природу, размеще-
произвести условия эксперимента, однако полу-
ние индуктора или детектора в герметичной квар-
чили маловоспроизводимые результаты
[267,
цевой колбе не влияло на эффект (см. рис. 7).
268]. После оптимизации условий нам удалось
получить достоверную воспроизводимую стиму-
Практически одновременно Д. Габор с
ляцию культуры-реципиента, которая в итоге
Т. Рейтером и А.Г. Гурвич показали, что фактор,
оказалась вызванной не ССС, а углекислым га-
обусловливающий митогенетический эффект,
зом, продуцируемым культурой-индуктором и
проявляет свойства электромагнитного излуче-
стимулирующим рост культуры-реципиента
ния - распространяется прямолинейно (рис. 7),
[269]. Позднее более тщательный анализ ориги-
отражается (рис. 8), преломляется в кварцевой
нальных работ по МГЭ позволил понять сделан-
призме (рис. 9) (в более поздних работах были по-
ные методические ошибки [270]:
лучены детальные спектры МГИ - см. ниже).
- сверхобедненная среда для реципиента, на
Размещение между индуктором и детектором
которой не мог проявиться МГЭ, а только стиму-
пластин из материалов с различными окнами
ляция химическими факторами;
прозрачности (слюда, стекло, желатин, кварц, де-
рево и т.п.) [4, 16, 271] позволило отнести это из-
- слишком высокая плотность культуры-ре-
лучение к УФ-диапазону. Кварцевая пластинка
ципиента;
не влияла на эффект, со стеклянной же пластин-
кой он исчезал. Также подтвердили УФ-диапазон
- неотработанная фаза роста посевной культуры;
излучения первые довольно простые экспери-
менты с призмами (см. рис. 9). Впоследствии этот
- неоптимизированная длительность индукции.
вывод был косвенно подтвержден тем, что мито-
Таким образом, негативный результат нашей
генетический эффект наблюдался под действием
собственной работы [267-269] также не имеет от-
многократно ослабленного УФ-излучения физи-
ношения к проверке МГЭ и не должен тракто-
ческих источников (искровых разрядов, ртутных
ваться в рамках данного вопроса.
ламп) [42-47].
БИОФИЗИКА том 66
№ 6
2021
1098
ВОЛОДЯЕВ и др.
Рис. 8. Эксперимент с отражением УФ-ССС от поверхности ртути. (Схема из работы [4], с незначительными
изменениями.)
Метод спектрального митогенетического анали-
ром требовала значительных затрат времени на
за. Схема спектрального митогенетического ана-
последовательную экспозицию детекторов. Даль-
лиза. Первые работы по исследованию физиче-
нейшее усовершенствование повысило разреше-
ских свойств МГИ, проведенные с помощью
ние спектров до 5 Å [274].
призм и зеркал, получили дальнейшее развитие
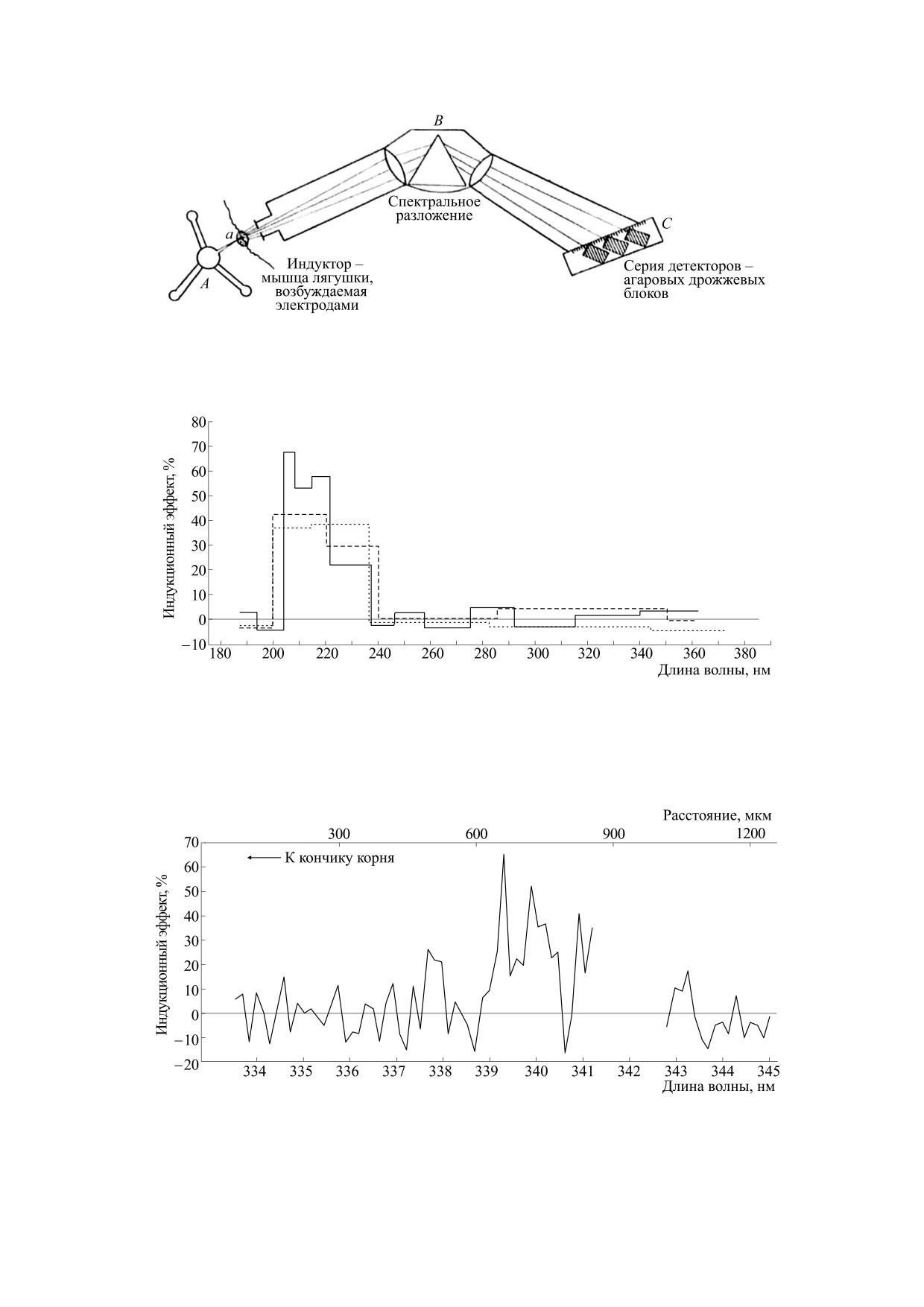
С помощью спектральных установок с биоло-
с применением монохроматоров. В работах [44,
гическими детекторами была обнаружена и ис-
272] были представлены установки для получе-
следована УФ-хемилюминесценция целого ряда
ния детальных спектров МГИ (рис. 10-12).
биохимических и химических реакций. На рис. 13
Индуктор размещали перед входной щелью
представлены митогенетические спектры различ-
монохроматора, а на месте выходной щели уста-
ных ферментативных реакций - расщепления
навливали линейку независимых биологических
креатинфосфата, глюкозы и др. [20].
реципиентов (как правило, агаровых блоков с
Сенсибилизированная флуоресценция. В. Фран-
дрожжевой культурой) - детекторов МГИ. При
кенбургер, развивая свою гипотезу о свободнора-
этом МГЭ наблюдался не во всех детекторах, а
дикальном механизме ультрафиолетовой хеми-
только в определенных, и заранее известное по-
люминесценции, предположил, что энергия, об-
ложение спектральных линий на этой линейке
разовавшаяся при рекомбинации радикалов, мо-
позволяло получить
«биологический спектр
жет передаваться веществу, не принимающему
МГИ».
участия в реакции, и высвечиваться с характер-
Существенно повысить разрешение за счет
ным для него спектром14 [275]. Спектральный
дальнейшего уменьшения ширины агаровых бло-
анализ подтвердил его гипотезу, это явление бы-
ков было невозможно, и из спектра излучения
ло названо сенсибилизированной флуоресценци-
стали выделять полосы шириной по 10 Å с помо-
ей и исследовано в различных модельных систе-
щью монохроматора [273]. Схема с монохромато-
мах. Например, добавление в водный раствор об-
лученного глицина15 солей NaH2PO4 или NaCl
приводит к появлению характерных спектраль-
ных полос ионов этих веществ [277, 278] (см.
рис. 14). Исследования сенсибилизированной
флуоресценции в различных модельных системах
были использованы для выяснения механизмов
УФ-ССС биологических объектов. Например,
анализ данных МГИ-спектров различных систем
привел авторов к заключению, что так называе-
14 Сама возможность передачи энергии возбужденного со-
стояния атома/молекулы другому атому/молекуле с после-
дующим высвечиванием была известна ранее.
15 Предварительно облученный глицин широко использо-
Рис. 9. Эксперимент с преломлением УФ-ССС в
вался в экспериментах «митогенетического периода» как
кварцевой призме. (Схема из работы [4], с незначи-
система с наиболее простым спектром - излучение наблю-
тельными изменениями.)
далось только в полосе 229-230 нм [276].
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1099
Рис. 10. «Митогенетический» спектральный анализ. Схема эксперимента по работе [44], с незначительными измене-
ниями.
Рис. 11. Три спектра МГИ от мышцы лягушки в состоянии тетануса, полученные с использованием биологических
детекторов - агаровых дрожжевых культур (по работе [44], с незначительными изменениями). Наблюдается
митогенетический эффект (изменение индекса почкования относительно контрольной дрожжевой культуры).
Различие в дискретности спектров связано с разной шириной блоков детектора.
Рис. 12. Спектр МГИ саркомы крысы, полученный с использованием биологических детекторов - корешков лука (по
работе [4], с незначительными изменениями).
БИОФИЗИКА том 66
№ 6
2021
1100
ВОЛОДЯЕВ и др.
Рис. 13. Спектры некоторых типичных источников ССС ферментативных реакций (из работы [20]): 1 - расщепление
креатинфосфата, 2 - расщепление глюкозы, 3 - расщепление нуклеиновой кислоты и лецитина, 4 - расщепление
пептидов, 5 - расщепление мальтозы, 6 - расщепление сахарозы, 7 - расщепление мочевины, 8 - расщепление ли-
пидов.
мый гликолитический спектр (см. спектр 2 на
Концепция сенсибилизированной флуорес-
рис. 13), т.е. спектр, который наблюдался при
ценции была подкреплена сравнением МГЭ-
ферментативном раcщеплении глюкозы и других
спектра бензольной воды с предварительно облу-
гликолитических процессах и был характерен для
ченным глицином [280] и обычных спектров флу-
ССС многих биологических объектов (крови, ра-
оресценции паров бензола [281] (см. рис. 15 и таб-
ковых опухолей и др.), обусловлен возбуждением
лицу).
целых молекул глюкозы [279], а спектр расщепле-
Специфика митогенетических спектров (отли-
ния нуклеиновой кислоты фосфатазой (см.
чия митогенетических спектров от спектров, по-
спектр 3 на рис. 13), наблюдаемый в ССС печени,
лученных стандартными методами). Спектраль-
нерва и др., связан не с целой молекулой нуклеи-
ный анализ УФ-излучения с биологическими де-
новой кислоты, а с фосфатным ионом (см.
текторами был ограничен областью очень низких
рис. 14).
интенсивностей и относительно узким спек-
тральным диапазоном чувствительности биоло-
гических детекторов. Исследуемый диапазон
Рис. 15. Сравнение МГЭ-спектров сенсибилизиро-
Рис. 14. Сенсибилизированная флуоресценция в вод-
ванной флуоресценции бензола и обычного спектра
ных растворах облученного глицина. Спектры облу-
флуоресценции паров бензола: сверху - спектр флуо-
ченного глицина, глицина с NaH2PO4, глицина с Na-
ресценции паров бензола; снизу - МГЭ-спектр флу-
Cl (использованы дрожжевые детекторы). Характер-
оресценции водного раствора бензола с добавлением
ные спектральные полосы глицина и ионов PO4
предварительно облученного и высушенного глици-
обозначены буквами G и P соответственно. (Данные
на (около 0.08% бензола и 1% глицина, по результа-
взяты из работы [278], см. также детали эксперимента
там 90 экспериментов). Приведены спектральные
в работе [277].)
данные из работы [280].
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1101
МГЭ-спектр сенсибилизированной флуоресценции бензола в сравнении с данными П. Прингсгейма по
флуоресценции паров бензола
Флуоресценция паров
Полосы, выделенные
Средний митогенетический
Число
бензола (П. Прингсгейм
монохроматором (Е.А. Гордон
эффект, % (дрожжевой
экспериментов
[282])
[280])
детектор)
2280-2300
0
5
2300-2360
0
3
2420-2460
41
6
2460-2480
12
3
2541
2500-2560
35
7
2560-2580
16
3
2602
2580-2620
25
4
2620-2645
8
4
2645-2660
3
2
2667
2660-2685
45
6
2685-2720
4
2
2739
2730-2760
42
8
2760-2780
38
7
2780-2810
5
2
2815
2810-2840
30
2
2840-2860
1
2
2860-2880
3
2
2895
2880-2900
26
4
2920-2960
0
2
–2980
2960-3000
45
5
распространялся от 190 нм, что соответствует гра-
пытки оценить относительную интенсивность,
нице прозрачности кварцевой оптики, до длин-
снимая спектры с различной продолжительно-
новолнового порога чувствительности биологи-
стью экспозиции [59]. Предполагалось, что излу-
ческих детекторов (примерно 290-325 нм) [16, 18,
чение имеет большую интенсивность на тех поло-
26, 44, 283-285]. Самая длинноволновая полоса
сах, на которых эффект проявлялся при мини-
ССС биологических источников, зарегистриро-
мальной экспозиции. Основные недостатки
ванная с помощью спектрального анализа с
МГИ-спектрального анализа были связаны с ис-
МГЭ-детекторами, - 287-291 нм (источником
пользованием биологического принципа детек-
была текущая протоплазма в листьях Elodea
тирования. Измерения одного спектра с разреше-
[285]), но обычно митогенетические спектры об-
нием 1 нм требовали проведения сотен экспери-
наруживали в пределах 190-250 нм.
ментов и около трех-четырех месяцев работы.
Такие спектры существенно отличались от
Для грубой оценки наличия спектральных осо-
традиционных спектров, получаемых с помощью
бенностей, характерных для наиболее распро-
физических приборов, тем, что показывали толь-
страненных спектров, обнаруживаемых при фер-
ко наличие или отсутствие излучения в пределах
ментативных реакциях гликолиза, расщепления
дискретных полос спектра, ничего не говоря о его
пептидных связей, окисления и действия фосфа-
интенсивности. Следует отметить, что были по-
тазы, т.е. четырех наиболее распространенных у
БИОФИЗИКА том 66
№ 6
2021
1102
ВОЛОДЯЕВ и др.
биологических объектов типов спектров, исполь-
лучены по митогенетическим спектрам излуче-
зовали упрощенную методику: непосредственно
ния нервов [26, 284, 289-295].
перед детекторами располагали шаблоны с про-
Обзоры применений спектрального МГИ-
резями; прорези соответствовали характерным
анализа в биологии представлены в работах [16,
полосам четырех основных спектров, каждая из
18, 20, 33, 40].
которых отсутствовала в остальных трех спектрах.
Несмотря на все эти недостатки, биологиче-
ские детекторы оставались непревзойденными по
ПРИКЛАДНОЕ ПРИМЕНЕНИЕ МЕТОДОВ
чувствительности к слабому УФ-излучению по
БИОЛОГИЧЕСКОГО ДЕТЕКТИРОВАНИЯ
сравнению с физическими детекторами того вре-
Митогенетическое излучение крови в норме и
мени (см часть III настоящего обзора), поэтому
при различных заболеваниях. Митогенетическое
именно схемы с биологическими детекторами ис-
излучение крови исследовалось довольно широко
пользовались для спектрального анализа в 1930-
из-за того, что кровь является доступным актив-
1940-х гг.
ным индуктором митогенетического эффекта и
Применения и значение спектрального анализа
представляет интерес с точки зрения медицин-
митогенетического излучения. Спектральный ана-
ской диагностики. Было показано, что хорошо
лиз был применен Г.М. Франком вначале с един-
излучает кровь у здоровых людей и целого ряда
ственной целью - доказать, что митогенетиче-
млекопитающих, птиц и амфибий, а также гемо-
ские лучи действительно относятся к средней
лимфа крабов и моллюсков. «Healthy blood never
ультрафиолетовой области [44], но очень скоро
fails. If a failure occurs, it is time to test either the yeast
спектральный МГЭ-анализ стал инструментом
or the apparatus» - писал Г. Гезениус [296]. Было
для неинвазивных бесконтактных исследований
показано, что излучение крови млекопитающих
биохимии живых организмов. Как писал
главным образом обусловлено гликолитически-
А.Г. Гурвич: «… в противоположность любым
ми процессами, крови амфибий - окислительны-
биохимическим методам, анализ совершается не
ми процессами [16, 297]. Подробный обзор ран-
после, а во время функционирования» [16]. Чув-
них работ по митогенетическому излучению кро-
ствительность метода была чрезвычайно высока
ви был представлен во втором томе справочника
для того времени, например, при расщеплении
по общей гематологии в 1934 г. [298] (цитируется
всего 10-7-10-8 г глюкозы наблюдался отчетли-
по работе [18]).
вый гликолитический спектр; в системе «желу-
Было обнаружено, что при раковых заболева-
дочный сок-белок» по спектру белок обнаружи-
ниях различных типов излучение крови исчезает
вался при концентрации 10-6 г.
на начальных стадиях малигнизации, задолго до
Результаты митогенетического спектрального
появления первых морфологических признаков
анализа хорошо согласовывались с известной ин-
злокачественного роста, за счет появления в кро-
формацией, например внутренние некротиче-
ви высокоспецифичного вещества, которое ту-
ские области раковых опухолей демонстрировали
шит УФ-хемилюминесценцию [297]. Это веще-
протеолитический спектр, в то время как в пери-
ство было названо «раковым тушителем». Было
ферийных областях с активным метаболизмом
показано, что при онкологических заболеваниях
излучение имело гликолитический и нуклеотиче-
кроме прекращения излучения крови наблюдает-
ский характер (т.е. совпадающий со спектром
ся также и прекращение излучения мочи [29]. Са-
расщепления нуклеиновой кислоты фосфатазой)
ми злокачественные опухоли, напротив, неодно-
[286]. Спектральный анализ позволял получить и
кратно были отмечены как наиболее активные
новую информацию. Например, ранее исследо-
источники митогенетического излучения ([16,
ватели столкнулись с явлением, которое не могли
30], см также работы [4, 20, 25, 30-34]).
объяснить - МГИ крови и глаза кролика полно-
Физико-химические свойства тушителя были
стью прекращалось при голодании и возобновля-
исследованы достаточно подробно, хотя его хи-
лось примерно через неделю. Спектральный ана-
мический состав не был идентифицирован [20,
лиз показал, что во время голодания излучение с
297]. Тушитель содержится в свободной от белков
гликолитическим спектром исчезает, а примерно
фракции крови, имеет отрицательный электриче-
через неделю появляется излучение со спектром,
ский заряд, в растворе довольно стоек, но термо-
характерным для процессов протеолиза [33]. По-
лабилен, диффундирует через коллодийную пе-
сле затухания гликолитической компоненты из-
регородку только при электрофорезе, выдержи-
лучения она быстро восстанавливалась при инъ-
вает высушивание при комнатной температуре,
екциях глюкозы [20]).
адсорбируется на каолин и может быть отмыт от
В середине 1930-х гг. в СССР спектральный
него щелочным раствором [20, 297]. Была разра-
МГИ-анализ стал одной из основных областей
ботана методика его выделения из крови. В моно-
исследований, связанных с МГИ [17, 20, 274, 276,
графии [297], посвященной тушителю, по анали-
287, 288]. Наиболее обширные данные были по-
зу результатов исследований было сделано за-
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1103
ключение, что раковый тушитель представляет
появляется в крови еще на обратимых стадиях
собой «пептид с преобладанием энольной формы
канцерогенеза [28, 297, 304]. При воздействии от-
пептидной связи», имеющий нитевидную форму
носительно малой дозой канцерогенов опухоли
и относительно небольшую длину (около 3 нм).
образовывались только у части мышей. Через 32-
Именно в работах по раковому тушителю впер-
53 суток после введения канцерогена у всех этих
вые утверждалось, что пептид крови может яв-
мышей в крови появлялся тушитель, опухоли ди-
ляться онкомаркером. Не исключено, что най-
агностировались к 191-222 суткам. У мышей, в
денное вещество представляло собой не один
крови которых тушитель не появлялся, опухоли
пептид, а целую группу различных пептидов,
не образовывались [28, 304]. Клинические иссле-
обладающих тушащими свойствами. Активная
дования пациентов с различной локализацией
специфичная часть пептидов должна была иметь
опухолей показали высокую чувствительность и
довольно малые размеры, поскольку после
специфичность (согласно данным авторов >95%)
«осторожного гидролиза» в соляной кислоте по-
онкодиагностики, основанной на раковом туши-
лученное вещество диффундировало через кол-
теле, и ее ценность для определения эффективно-
лодийную пленку без потери тушащих свойств
сти проведенной терапии [20, 28, 257, 297, 304].
[297].
Некоторые другие применения митогенетиче-
Обширные экспериментальные исследования
ского детектирования. Биологические МГЭ-де-
показали, что митогенетическое излучение крови
текторы использовались для исследований не
прекращается или ослабевает при ряде физиоло-
только биообъектов. С их помощью были прове-
гических состояний: физическом утомлении [49],
дены широкие спектральные исследования ССС
умственной нагрузке [50], голодании [16], в по-
различных биохимических систем, моделирую-
жилом возрасте [49], а также при различных забо-
щих ферментативные процессы в живом организ-
леваниях (пернициозная анемия, лейкемия,
ме [273, 276, 305], а также неорганических хими-
пневмония, сепсис [33, 298, 299], хронический
ческих реакций [39, 306]. Была показана чувстви-
тонзиллит [300], диабет, цирроз печени и др. [16,
тельность луковых
[4] и дрожжевых
[307]
297]). Было обнаружено, что во всех случаях, не
детекторов к слабому искусственному УФ. Дрож-
связанных с онкологическими заболеваниями,
жевой детектор использовали для определения
кровь перестает излучать из-за появления ве-
наличия радиоактивного осадка на коже после
ществ, не обладающих свойствами тушителя, но
радиоактивных ванн (оценка объемным методом
поглощающих УФ-излучение. Они были названы
Брайнеса) [308].
гасителями [20, 297]. Гасители представляют со-
бой низкомолекулярные продукты патологиче-
ОБСУЖДЕНИЕ БИОЛОГИЧЕСКИХ
ского метаболизма, например, кетоновые тела
МЕТОДОВ ДЕТЕКТИРОВАНИЯ
при диабете. Было показано, что, в отличие от га-
сителей, раковый тушитель не поглощает УФ-из-
МИТОГЕНЕТИЧЕСКОГО ИЗЛУЧЕНИЯ
лучение и не влияет на прозрачность тонких сло-
Особенности биологических методов детектиро-
ев крови существенным образом. Принципиаль-
вания сверхслабого свечения: достоинства и не-
ные отличия свойств тушителя и гасителей
достатки. Использование в качестве детекторов
позволили разработать методику, которая с помо-
биологических объектов имело целый ряд недо-
щью дрожжевых детекторов митогенетического
статков. Во-первых, оно было чрезвычайно трудо-
излучения позволяла идентифицировать наличие
емким и длительным. Например, одна регистрация
тушителя в крови [28, 257, 258].
УФ-излучения с помощью лукового детектора тре-
Тушитель имеется только в крови людей и жи-
бовала тщательной подготовки гистологических
вотных со злокачественными опухолями и состо-
срезов кончиков корней, включая этапы фикса-
яниями, которые относят к предраковым, появ-
ции, окрашивания, парафинирования, подсчета
ляется на ранних стадиях заболевания. Излуче-
под микроскопом числа митозов в различных зо-
ние крови у крыс исчезало на девятые-
нах меристемы при разных увеличениях микро-
двенадцатые сутки после прививки опухоли здо-
скопа, общее поле составляло несколько тысяч
ровому животному, тогда как прощупываться
клеток, причем подсчет под микроскопом деля-
опухоль начинала через 28-32 суток после при-
щихся клеток был не только долгим, но и субъек-
вивки [301]. В экспериментах на мышах раковый
тивным. Так называемая агаровая дрожжевая ме-
тушитель появлялся в крови на первые-вторые
тодика требовала подсчета под микроскопом поч-
сутки после имплантации карциномы и исчезал
кующихся клеток на суммарном поле, содержащем
через трое-восемь суток после ее радикального
более 10000 клеток; только на подсчет уходило
хирургического удаления; в случае стойкого со-
около трех-четырех часов, не считая времени на
хранения тушителя в крови неизменно наблюда-
приготовление детектора, индуктора, препаратов
лись рецидивы [302, 303]. В экспериментах с регу-
для микроскопии, а также длительности самого де-
лярным смазыванием кожи мышей канцероген-
тектирования [309]. Для получения одного спектра
ными веществами было показано, что тушитель
обычно проводили несколько сотен эксперимен-
БИОФИЗИКА том 66
№ 6
2021
1104
ВОЛОДЯЕВ и др.
тов по дрожжевому детектированию, что занимало
довольно субъективный выбор («еще не почка,
три-четыре месяца. Так, например, в работе [305]
почка, уже не почка») и не только требовал ма-
спектр излучения в системе «мочевина-уреаза»
стерства, но и не мог гарантировать от системати-
был построен на основании результатов 300 экспе-
ческих ошибок (в поздних работах подчеркива-
риментов.
лась необходимость слепого метода проведения
подобных экспериментов).
Во-вторых, работа с биологическими детекто-
рами требовала очень тщательного соблюдения
Любой биологический детектор использовал-
методики. Некоторые авторы не смогли повто-
ся однократно, не мог быть предварительно про-
рить известные эксперименты по митогенетиче-
тестирован или прокалиброван. Преодолеть про-
скому эффекту именно из-за нарушения методи-
блемы вариабельности биологических детекторов
ки работы с биологическими детекторами.
можно было только многократным повторением
Подробно их ошибки разобраны в обзорной ра-
экспериментов и хорошей постановкой методики
боте [52] и выше в настоящем обзоре. В-третьих,
с предельно точным воспроизведением значимых
вариабельность как самих биологических детекто-
условий. Например, вывод о наличии или отсут-
ров, так и многочисленных внешних факторов,
ствии излучения в одной полосе спектра при из-
влияющих на их чувствительность, делала такой
мерениях дрожжевым детектором делали после
метод регистрации слабого УФ-излучения доволь-
серии не менее десяти экспериментов с согласо-
но нестабильным. Биологические системы слож-
ванными результатами16.
нее технических устройств, им свойственна зна-
чительная естественная вариабельность. Биоло-
МГЭ-детекторы были чувствительны только в
гическая реакция системы не может быть точно
узком диапазоне длин волн, примерно от 190 до
количественно предсказана, как бы подробно ни
290-320 нм (см., например, рис. 4в), из-за погло-
щения в кварцевых элементах возможности де-
были описаны ее текущие физиологические па-
тектирования излучения с длинами волн менее
раметры, внешние условия и история развития.
Например, на современном уровне знаний невоз-
190 нм не могли быть исследованы. Биологиче-
можно предсказать со 100%-й вероятностью даже
ское детектирование не позволяло количествен-
всхожесть семян, хотя это не противоречит био-
но характеризовать интенсивность излучения, но
применялось для грубой сравнительной оценки
логическому закону их прорастания. Биологиче-
интенсивностей биообъектов по минимальной
ские законы имеют вероятностный характер, и
митогенетический эффект не является исключе-
продолжительности экспозиции, при которой
нием из этого правила.
наблюдался МГЭ (при прочих равных условиях).
Предполагалось, что более интенсивное УФ-
Во всех лабораториях, где проводили тысячи
ССС биообъекта способно стимулировать мито-
экспериментов с дрожжевыми и бактерийными
зы в детекторе при меньших экспозициях. Срав-
детекторами, иногда наблюдали необъяснимые
нивать между собой можно было только ССС
перебои - детекторы переставали реагировать на
биологических объектов, а также модельных хи-
излучение [18, 54], причем, по свидетельству
мических систем, поскольку чувствительность к
А.Г. Гурвича, это наблюдали одновременно в те-
ослабленному излучению физических источни-
чение нескольких дней все сотрудники, ведущие
ков была гораздо ниже [47].
независимые эксперименты. Потом также необъ-
Единственным, но принципиальным преиму-
яснимо способность к детектированию восста-
навливалась. Какой фактор (или факторы) обу-
ществом биологических детекторов, обусловив-
славливал перебои в работе биологических детек-
шим их широкое применение в «митогенетиче-
торов, осталось неясным. Гурвич предполагал
ский» период исследований, была их чрезвычай-
изменения радиофона, тем более что радиосвязь в
но высокая чувствительность к ССС. Длительное
время после обнаружения эффекта биологиче-
те годы активно развивалась. Были и другие пред-
положения, например влияние погодных условий
ские детекторы оставались единственным воз-
и др. [18].
можным средством регистрации МГИ, появив-
шиеся затем модификации счетчиков Гейгера-
Продолжительность экспонирования в каж-
Мюллера имели меньшую чувствительность и не
дой серии экспериментов с любым биологиче-
могли быть использованы, например, для спек-
ским детектором приходилось подбирать индиви-
трального анализа и многих других исследо-
дуально. При увеличении дозы митогенетическо-
ваний.
го излучения стимуляция сменяется его
угнетением (см. рис. 4а,б). Это чрезвычайно за-
16 Следует отметить, что дрожжевой детектор был наиболее
трудняло применение биологических детекторов,
методически проработанным из всех биологических МГЭ-
детекторов. А.Г. Гурвич указывал, что он наилучший по
так как в каждой серии экспериментов приходи-
воспроизводимости [17], приведенное количество повто-
лось подбирать время экспозиции. Сами эффек-
ров относится к этапу его массового применения для спек-
ты были, как правило, невелики (20-30% превы-
трального анализа, после практически предельного разви-
шения над фоном), а подсчет почек подразумевал
тия дрожжевой методики.
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1105
Следует отметить, что развитие биологических
нены на крайне низком уровне. Как писал
методик детектирования было очень сложной за-
А. Голлендер, «особенно жаль, что в этой теме
дачей, требующей масштабных многофакторных
стали участвовать авторы, увидевшие в ней лишь
трудоемких экспериментов, в первую очередь из-
возможность привлечь к себе внимание» [51]. Це-
за естественных трудностей стандартизации био-
лый ряд явлений, исходно относимых авторами к
логических объектов, выступающих в качестве
разряду «дистантных нехимических взаимодей-
детекторов, отсутствия знаний о механизмах де-
ствий» в период 1990-2010-х гг., либо оказались
тектирования, недостаточной информации о том,
артефактами, либо получили чисто «химическое»
какие внешние факторы необходимо стандарти-
объяснение.
зовать.
Следует отметить, что результаты эксперимен-
О современном значении биологического детек-
тов митогенетического периода исследований, не
тирования митогенетического эффекта. Биологи-
имея достаточной теоретической базы для их объ-
яснения на уровне знаний того времени, зачастую
ческое детектирование занимает важное значе-
ние в истории исследований ССС, поскольку са-
приводили к неверным гипотезам, это отмечал и
мо излучение было открыто и исследовалось в
сам А.Г. Гурвич. Кроме того, он подчеркивал, что
первые десятилетия в основном именно с помо-
мультидисциплинарный характер предмета неод-
щью биологических детекторов. В настоящее вре-
нократно подталкивал исследователей к попыткам
работать и выдвигать гипотезы в научных областях,
мя биологические МГЭ-детекторы представляют
интерес только для исследования самого эффек-
в которых они не были специалистами [17].
та, т.е. как реципиенты в схеме наблюдения МГЭ
Как писал O. Ран [198]: «Российские ученые,
(например, для исследования механизма детекти-
которые первыми использовали дрожжи и бакте-
рования, выяснения вопроса о биологической
рии в качестве детекторов митогенетических лу-
роли эффекта). Из-за низкой воспроизводимости
чей, не были бактериологами; это очень очевидно
методик и трудоемкости они не востребованы как
из их инструкций по подготовке культуральной
детекторы в установках для исследования физи-
среды и их описания эксперимента. Ничего не
ческих свойств ССС (например, для спектраль-
сказано о физиологическом состоянии клетки,
ного анализа) или в прикладных задачах, связан-
хотя вскоре они заметили, что клетки не реагиро-
ных с детектированием малых количеств веще-
вали в течение всего периода своего роста. Барон
ства по хемилюминесценции (например,
(1930) заново открыл лаг-фазу роста дрожжей, с
методика онкодиагностики по наличию в крови
которой он не был знаком. Отсюда его инструк-
тушителя хемилюминесценции) как это было в
ция, что дрожжи следует выращивать в течение
1920-1940-х гг. В ближайшем времени исследо-
шести-десяти часов при комнатной температуре,
вания ССС будут проводиться, вероятно, исклю-
чтобы они быстро отреагировали. К сожалению,
чительно с помощью современных технических
не было заявлено, что 12°C считается нормальной
средств (ФЭУ, ЭОП, ПЗС-матриц и т.д.) (см
комнатной температурой в Москве и Ленинграде,
часть III настоящего обзора). Вместе с тем в отда-
и исследователи, следуя этим указаниям букваль-
ленной перспективе при выяснении механизма
но и при американских комнатных температурах,
биологического детектирования ССС, причин
несомненно, достигли бы совершенно иной фи-
более высокой чувствительности [47] биологиче-
зиологической стадии».
ских детекторов к ССС биообъектов по сравне-
Вместе с тем неверная теоретическая интер-
нию с излучением физических источников, раз-
претация не умаляет значимости накопленного в
работке хорошо воспроизводимых методик био-
тот период обширного экспериментального мате-
логическое
детектирование
на
основе
риала.
использования МГЭ или других биологических
Влиятельные критические работы. Существует
эффектов ССС, возможно, найдет свою нишу.
ряд работ с серьезной критикой феномена МГЭ.
Неоднозначное состояние вопроса о принципе
Большинство их них детально разобрано выше в
биологического детектирования митогенетического
сравнении с основными «позитивными работа-
эффекта. Главная проблема в области биологиче-
ми», отмечены принципиальные расхождения в
ского детектирования ССС - крайне неопреде-
методике. По словам O. Рана: «Некоторые авторы
ленное состояние вопроса о самом эффекте, на
считают, что их отрицательные результаты опро-
котором оно основано, т.е. МГЭ. Этот эффект не
вергают положительные результаты других иссле-
только не общепризнан, но часто ассоциируется с
дователей. Это - ошибка. Если два автора полу-
«лженаукой» [310]. Отметим несколько причин
чают разные результаты, это не доказывает, что
такого положения.
один из них прав, а другой ошибается. Это дока-
зывает только, что они поставили разные экспери-
Дискредитация: ложноположительные резуль-
менты» [198].
таты. К сожалению, в литературе встречается
достаточно много работ, якобы подтверждающих
Кроме того, существует ряд критических обзо-
МГЭ или ссылающихся на него, которые выпол-
ров, явно предвзятых в пользу всех (даже плохо
БИОФИЗИКА том 66
№ 6
2021
1106
ВОЛОДЯЕВ и др.
сделанных) «негативных» и против любых «пози-
мировой войны. Во время войны продолжаются
тивных» работ. Так, обзор [81] посвящен в основ-
отдельные попытки апробации и использования
ном поиску несоответствий между «позитивны-
МГЭ для медицинской диагностики, многие из
ми» работами разных авторов (часть из которых
которых остались неопубликованными; другие
действительно не отвечает требованиям к науч-
вошли в диссертации [28, 258, 315, 316], но, по
ной статье, другая же часть сделана на очень вы-
очевидным причинам, фундаментальные иссле-
соком уровне) и делает акцент на дискредитиру-
дования почти прекратились. После 1945 г. ис-
ющие работы по МГЭ, «пропуская» все серьезные
следования МГЭ в СССР возобновились только в
публикации, такие как [6, 23, 54, 252]. Крайне
нескольких лабораториях А.Г. Гурвича и его кол-
жесткая критика результатов по вторичному из-
лег. После сессии ВАСХНИЛ в 1948 г., на которой
лучению, полученных с помощью биологических
митогенетическое направление было подвергну-
детекторов, была основана на их несоответствии
то крайне жесткой критике вместе с генетикой,
закону Стокса: «Это неизвестный до сих пор тип
А.Г. Гурвич окончательно потерял возможность
флуоресценции... Он предполагает присутствие в
вести экспериментальную работу. Впоследствии,
дрожжевой культуре пивного сусла демона, спо-
с подтверждением существования ССС биообъ-
собного делать непрозрачную суспензию про-
ектов с помощью ФЭУ и некоторой реабилитаци-
зрачной для ультрафиолетового света. Подобно
ей феномена, исследования А.Г. Гурвича отчасти
демону Максвелла, демон Гурвича - это просто
продолжились в лабораториях его дочери
изобретение для уклонения - на бумаге - от тер-
А.А. Гурвич и С.В. Конева, однако существенно-
модинамического закона с той важной разницей,
го развития исследования компоненты УФ-ССС
что нас серьезно просят поверить в его существо-
уже не получили. В Европе и США после войны
вание» [81]. Следует отметить, что возможность
исследования МГЭ не возобновила ни одна из ра-
антистоксовых процессов была показана
нее работавших лабораторий. Ситуация усугуби-
Л. Ландау через 12 лет, а экспериментально они
лась тем, что основные работы по МГЭ, опубли-
были продемонстрированы физическими мето-
кованные на русском языке, оказались совер-
дами только 25 лет спустя. Другой пример пред-
шенно недоступны для западных авторов, в
взятой необъективной критики - влиятельная
частности это практически все работы по онкоди-
анонимная публикация в журнале «Nature» [311],
агностике, наиболее интересные с точки зрения
полностью основанная на данных работы [253]
прикладной науки.
(детально проанализированных выше) и исполь-
Проблемы с воспроизводимостью митогенети-
зующая не вполне научную риторику при упоми-
ческого эффекта. Наиболее критический фактор
нании «позитивных» работ.
для «признания» МГЭ - это сложности с его вос-
T. Квикенден с соавторами [263, 264] также
произведением. «Если он такой универсальный,
весьма критичны в отношении «ранних работ».
почему его так сложно поймать?» Это - не только
При этом во всех публикациях авторы ссылаются
стандартный вопрос, но и частый аргумент про-
на три работы [253, 312, 313], в которых видят
тив МГЭ. Хотя, конечно, никакие рассуждения
окончательное опровержение феномена МГЭ,
не могут быть аргументами в экспериментальной
хотя в работах [312, 313] авторы пытались обнару-
науке, этот вопрос действительно приводит в за-
жить с помощью модифицированных газоразряд-
мешательство…
ных счетчиков излучения от объектов, которые,
С нашей точки зрения одна из основных при-
как ранее уже было показано биологическими де-
чин трудностей воспроизведения МГЭ связана с
текторами, не являлись источниками УФ-ССС,
тем, что биологический детектор должен про-
более того, следует отметить низкую чувствитель-
явить чувствительность к внешнему ССС на фоне
ность и существенные конструктивные недостат-
значительного внутреннего фона. Любой объект,
ки используемых ими модификаций газоразряд-
используемый в экспериментах по МГЭ, состоит
ных счетчиков, приводящие к повышенным шу-
из большого числа клеток с характерным рассто-
мам (см. критику этих модификаций в работе
янием между ними ~10-6-10-4 м, расстояние же
[314] и части III настоящего обзора). Критика
до внешнего индуктора МГЭ в любом случае со-
биологической части известной негативной экс-
периментальной работы [253] была представлена
ставляет не менее чем 10-3-10-2 м (при этом, как
выше, критику ее физической части см. в части
известно, средняя интенсивность излучения от
III настоящего обзора.
точечного источника падает пропорционально
квадрату расстояния от него). Также интенсив-
Методические детали экспериментов по МГЭ
ность излучения снижается за счет отражения от
и критика ряда ключевых авторов были проана-
имеющихся границ сред с разным показателем
лизированы также в работе [52].
преломления («кварц-воздух», «кварц-среда» и
Исторические и геополитические факторы.
пр.), кроме того имеется некоторое поглощение и
Большая часть исследований МГЭ относится к
рассеяние в средах, разделяющих индуктор и де-
1920-1930-м гг. и обрывается с началом Второй
тектор (например, отмечалось, что для химиче-
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1107
ской изоляции детектора и индуктора требуется
ственно компрометирует результаты, получен-
кварц высокой чистоты, с кварцем более низкого
ные с их помощью, и затрудняет проверку. Толь-
качества эффект пропадал).
ко на уровне воспроизводимой методики имеет
Таким образом, для достаточно высокого со-
смысл применение биологического объекта в ка-
честве детектора в обычном техническом пони-
отношения сигнал/шум необходимо следующее:
мании этого термина, до этого можно вести речь
1. Культура индуктора должна быть достаточно
только об исследовании самого эффекта.
«сильной», чтобы произвести МГЭ на таком
большом «нетипичном» расстоянии.
Более вероятным представляется, что методи-
2. Клетки реципиента должны быть чувстви-
ки биологического детектирования, разработан-
тельны к внешней индукции.
ные в ряде научных коллективов при экспери-
ментах на большом материале, обеспечивали вос-
3. «Собственный» МГЭ в культуре реципиента
производимость, достаточную для получения
должен быть по возможности подавлен, но так,
верных результатов, и были вполне пригодным
чтобы культура сохранила жизнеспособность и
инструментом исследований. Когда методики
чувствительность к внешним индукторам.
успешно перенимали, например, в пределах шко-
4. Интенсивность сигнала не должна быть по-
лы Гурвича и сотрудничавших с ней научных и
давлена ниже некоторого критического уровня
медицинских организаций, то, вероятно, часто
при преодолении расстояния между индуктором
воспроизводили при этом и некоторые условия
и детектором.
эксперимента, которым не придавалось значения
Следуя этим требованиям, можно легко по-
в самих описаниях методик.
нять большинство условий, необходимых для
воспроизведения МГЭ (см. выше):
Более того, в разных независимых лаборатори-
ях отработанные методики детектирования могли
1. Тщательный отбор индукторов и еще более
быть различны, что и могло быть причиной рас-
тщательная подготовка реципиента, включая
хождения количественных результатов при на-
подбор фазы роста культуры-реципиента и по-
блюдении близких эффектов. Например, луко-
севного материала.
вые методики, которые использовали Д. Габор и
2. Создание специальных условий для культу-
Т. Рейтер
[272], существенно отличались от
ры реципиента
- пониженной температуры,
методик школы А.Г. Гурвича [78], в первую
обедненной среды, низкой плотности культуры и
очередь тем, что Д. Габор и Т. Рейтер работали с
т.д. - так, чтобы культура реципиента оказалась
отрезанными корнями. Это значительно меняло
ослабленной, но при этом жизнеспособной.
режим митогенетического излучения внутри кор-
3. Минимизация потерь излучения между ин-
ня, в частности отрезанный корень не являлся
дуктором и детектором за счет уменьшения рас-
индуктором.
стояния между ними, снижения отражения и рас-
Прошлое и будущее биологического детектиро-
сеяния.
вания. Итак, как мы постарались продемонстри-
4. Специальные предосторожности, такие как
ровать во второй части нашего обзора, «ранние
слабый фоновый видимый свет, экранирование
работы» по МГИ - это не несколько отдельных
внешнего ультрафиолета, очень тонкий слой
сомнительных публикаций (как полагали, напри-
культуры-реципиента и т.д.
мер, в работе [317]), а около тысячи работ, в том
Кажется гораздо более удивительным, что
числе в наиболее высокорейтинговых журналах;
МГЭ вообще удалось обнаружить, чем то, что он
часть этих работ была выполнена очень автори-
оказался таким «капризным».
тетными учеными (проф. O. Ран, США; нобелев-
Недостаточное для воспроизведения описание
ский лауреат Д. Габор, Германия; проф.
методик17 делает их неубедительными и соответ-
Л.К. Вольф, Нидерланды; проф. А.Г. Гурвич,
Б.С. Песоченский, академики Ю. А. Харитон и
17 Из методик того периода наиболее подробно было описа-
Г.М. Франк и др., СССР). Исследования митоге-
но определение в крови тушителя хемилюминесценции в
нетического излучения широко обсуждались на
целях онкодиагностики, имелись обширные результаты, в
международном научном уровне, ключевые экс-
том числе более 1300 протоколов в ведущих клинических
лабораториях, которые показали высокую эффективность
перименты подвергались неоднократной провер-
предложенной онкодиагностики. В этой методике исполь-
ке в научных коллективах разных стран, публика-
зовали наиболее распространенный тогда биологический
ции имели массовый характер и были подкрепле-
детектор - дрожжевой, эффект тушения тестировали при
ны значительным количеством подробных
послесвечении раствора глицина (это один из наиболее ча-
сто используемых контрольных источников МГИ). Вместе
протоколов испытаний. Негативные работы со-
с тем Н.И. Линтварева (Центральный онкологический ин-
ставили менее 3% от общего количества экспери-
ститут), согласно ее отчету в архиве А.Г. Гурвича, ни разу
ментальных работ, выполненных по тематике
не только не смогла обнаружить тушения, но даже зареги-
МГИ, например, это работы [91, 204, 253, 254].
стрировать свечение контрольного источника МГИ при
многократном тщательном повторении экспериментов по
Все они были выполнены с серьезными отклоне-
описанной методике.
ниями от известных методик, которые тщательно
БИОФИЗИКА том 66
№ 6
2021
1108
ВОЛОДЯЕВ и др.
обсуждались в ответных публикациях [16, 18, 59,
текторы представляют собой интерес только для
198, 301, 318]. Более подробно методические
исследования самого эффекта, т.е. как реципиен-
ошибки этих и других негативных работ проана-
ты в схеме наблюдения МГЭ (например, для ис-
лизированы в обзоре [52] (cм. также историю и
следования механизма детектирования, выясне-
результаты исследований митогенетического из-
ния вопроса о биологической роли эффекта). Из-
лучения в обзорах [52, 248, 319-321]). Все аргу-
за низкой воспроизводимости методик и их тру-
менты критиков, утверждавших невозможность
доемкости они не востребованы как детекторы в
заявленных феноменов с теоретической точки
установках для исследования физических свойств
зрения, давно устарели, а экспериментальные ис-
ССС (например, для спектрального анализа) или
следования ССС «митогенетического» периода
в прикладных задачах, связанных с детектирова-
не были опровергнуты.
нием малых количеств вещества по хемилюми-
несценции (например, методика онкодиагности-
В то же время комплекс работ по исследовани-
ки по наличию в крови тушителя хемилюминес-
ям ССС митогенетического периода нельзя счи-
ценции), как это было в
1920-1940-х гг. В
тать и доказанным. Биологические методики, с
ближайшем времени исследования ССС будут
помощью которых были получены эти результа-
проводиться, вероятно, исключительно с помо-
ты, не представляются достаточно полными в том
щью современных технических средств (ФЭУ,
виде, в котором они описаны в старых работах.
ЭОП, ПЗС-матриц и т.д.). Вместе с тем в отдален-
Описание методик, недостаточное для их уверен-
ной перспективе - при выяснении механизма
ного воспроизведения, компрометирует выпол-
биологического детектирования ССС, причин
ненные исследования и затрудняет их проверку
более высокой чувствительности [47] биологиче-
(см. выше).
ских детекторов к ССС биообъектов по сравне-
Авторы настоящего обзора полагают, что ис-
нию с излучением физических источников, раз-
следования ССС «митогенетического» периода -
работке хорошо воспроизводимых методик, -
не фальсификация и не иллюзия, основанная на
биологическое детектирование на основе исполь-
случайной или систематической ошибке, но
зования МГЭ или других биологических эффек-
сложно наблюдаемые эффекты, мало понятные и
тов ССС, возможно, найдет свою нишу.
трудно воспроизводимые в работах того времени.
На основании анализа методик и результатов
того периода мы можем предположить, что цело-
ЗАКЛЮЧЕНИЕ
му ряду научных коллективов, которые вели мас-
Современное состояние проблемы УФ-ССС
штабные исследования ССС в 1923—1948 гг., дей-
можно назвать парадоксальным. С одной сторо-
ствительно удалось подобрать условия, в которых
ны, само существование МГЭ-эффекта не дока-
достаточно стабильно и воспроизводимо работа-
зано и не опровергнуто. С другой стороны, целый
ли МГЭ-детекторы. Перенести эти отработанные
ряд концепций, происходящих из работ по МГЭ с
методики в другие лабораторные условия удава-
использованием биологических детекторов, об-
лось не всегда18; это заставляет предполагать, что
щепризнан и относится к «фактам из учебников».
в методиках были описаны не все значимые усло-
Кратко перечислим их:
вия (возможно, режим освещения, радиофон и
- Существование сверхслабого свечения биоло-
другие условия). Например, почти все спектраль-
гических объектов.
ные исследования проводили в школе Гурвича,
либо исследователями, которые перенимали ме-
- Ультрафиолетовая компонента сверхслабого
тодику непосредственно у представителей этой
свечения [322, 323], по спектру и времени появле-
школы (вероятно, повторяя некоторые важные
ния полностью совпадает с МГИ.
детали, которым не придавалось значения и кото-
- Свободнорадикальный механизм генерации
рые не оговаривались в статьях).
сверхслабого свечения. Впервые (в 1930-е годы!)
Биологическое детектирование занимает важ-
предложен в работах по МГЭ, как вывод из дан-
ное значение в истории исследований ССС, по-
ных «спектрального анализа МГЭ» (см. работу
скольку само излучение было открыто и первые
[52]).
десятилетия подавляющая часть исследований
- Повышение интенсивности сверхслабого све-
его физических свойств проводилась именно с
чения в ответ на внешнее повреждающее воздей-
использованием биологического детектирова-
ствие [52, 322]. Впервые обнаружено в работах по
ния. В настоящее время биологические МГЭ-де-
МГЭ, в которых названо «деградационным излу-
чением» [16].
18 Следует отметить, что нередко положительные результаты
получали исследователи, начавшие работы с предвзятым
- Временная структура сверхслабого свечения.
негативным отношением к существованию эффекта (на-
По современным данным спонтанное ССС био-
пример, Р. Одюбер) и, наоборот, негативные результаты
логических образцов состоит из отдельных вспы-
получали исследователи, изначально настроенные пози-
тивно (А. Голлендер).
шек длительностью не более 10-3 с [324, 325].
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1109
Этот же вывод сделан в 1930-е гг. в работах по
7. A. J. Ferguson and O. Rahn, Archiv für Mikrobiologie
«фракционированию МГЭ» [16] (см. работу [52]).
4 (1-4), 574 (1933).
8. M. A. Baron, Wilhelm Roux’ Archiv fur Entwicklungs-
К простым совпадениям трудно отнести также
mechanik der Organismen 108 (4), 617 (1926).
и корреляцию результатов онкодиагностики пер-
вичных больных с опухолевыми процессами не-
9. M. A. Baron, Planta/Achiv für wissenschaftliche
установленной этиологии и окончательных кли-
Botanik 10 (1), 28 (1930).
нических диагнозов, полученных значительно
10. H. Guillery, Virchows Archiv 279, 311 (1929).
позже: специфичность и чувствительность мето-
11. Г. К. Хрущев, Архив биол. наук, Сер. Б 35 (1), 317
дики, построенной на использовании митогене-
(1934).
тического эффекта, составляла >95% [28, 257, 297,
12. L. Gurwitsch and A. Anikin, Wilhelm Roux' Arch. En-
304].
twickl. Mech. Org. 113 (4), 731 (1928).
Экспериментальные исследования митогене-
13. J. Magrou and M. Magrou, Rev. Path. Veg. Ent. Agr.
тического излучения с помощью биологических
14, 244 (1927).
детекторов в 1923-1948 гг. позволили сделать ре-
14. M. J. Magrou, M. M. Magrou, and M. P. Reiss,
волюционные, даже парадоксальные для того
Comptes Rendus de l'Academie des Sciences 189, 779
времени научные выводы, которые были под-
(1929).
тверждены спустя десятилетия другими методами
15. L. K. Wolff and G. Ras, Nature 133 (3361), 499 (1934).
(например, существование пептидных онкомар-
16. А. Г. Гурвич and Л. Д. Гурвич, Митогенетическое
керов крови, антистоксовых процессов, фоторе-
излучение (Изд-во ВИЭМ, Ленинград, 1934).
активации и другие). Вместе с тем методики, с по-
17. А. Г. Гурвич и Л. Д. Гурвич, Успехи соврем. биоло-
мощью которых были получены эти верные ре-
гии 16 (3), 305 (1943).
зультаты, в том виде, в котором они описаны в
18. O. Rahn, Invisible radiations of organisms (Gebrüder
старых работах, не представляются достаточно
Bornträger, Berlin, 1936).
убедительными с точки зрения современных тре-
19. Ю. H. Пономарева, Бюл. эксперим. биологии и
бований.
медицины 2 (1), 16 (1936).
Мы считаем тщательную и объективную про-
20. А. Г. Гурвич и Л. Д. Гурвич, Митогенетическое из-
верку МГЭ, с вниманием ко всем накопленным
лучение: Физико-химические основы и приложения в
данным и методическим деталям, важной задачей
биологии и медицине (Медгиз, М, 1945).
как для фундаментальной, так и (в перспективе)
21. A. Potozky, Protoplasma 25 (1), 49 (1936).
для прикладной науки и медицины. Однозначное
22. W. W. Siebert, Zeitsch. Klin. Med. (109), 360 (1928).
доказательство феномена МГИ сыграло бы зна-
23. L. Acs, Centr. Bakt. I Abt. Orig. 120 (1/2), 116 (1931).
чимую роль для всей биологии, опровержение за-
крыло бы важную страницу в истории науки.
24. L. Acs, Zentralblatt für Bakteriologie, Parasitenkunde
und Infektionskrankheiten 126, 125 (1932).
25. W. W. Siebert, Biochem. Z. 202, 115 (1928).
КОНФЛИКТ ИНТЕРЕСОВ
26. А. А. Гурвич, Архив биол. наук, Сер. Б 35 (1), 127
Авторы заявляют об отсутствии конфликта
(1934).
интересов.
27. L. D. Gurwitsch and S. Salkind, Biochemische
Zeitschrift 211 (1-3), 362 (1929).
28. Б. С. Песоченский, Дисс. … д-ра мед. наук (Ленин-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
градский онкологический институт, Ленинград,
Настоящая работа не содержит описания ис-
1942).
следований с использованием людей и животных
29. W. W. Siebert, Biochemische Zeitschrift 226 (4/6), 253
в качестве объектов.
(1930).
30. A. G. Gurwitsch and L. D. Gurwitsch, Biochemische
Zeitschrift 196 (4-6), 257 (1928).
СПИСОК ЛИТЕРАТУРЫ
31. M. Kisliak-Statkewitsch, Zeitschrift für Krebsfor-
1. D. Fels, Plos One 4 (4), e5086 (2009).
schung 29 (1), 214 (1929).
2. A. G. Gurwitsch, Archiv für mikroskopische Anatomie
32. L. Gurwitsch and A. Gurwitsch, Zeitschrift für
und Entwicklungsmechanik 100 (1-2), 11 (1923).
Krebsforschung 29 (1-2), 220 (1929).
3. A. G. Gurwitsch, Das Problem der Zellteilung physiolo-
33. A. G. Gurwitsch and L. D. Gurwitsch, Die mitoge-
gisch betrachtet (Julius Springer, Berlin, 1926).
netische Strahlung. (Springer-Verlag, Berlin, Heidel-
4. T. Reiter and D. Gabor, Zellteilung und Strahlung.
berg, Berlin, 1932).
(Springer-Verlag, Berlin, 1928).
34. H. P. Ypsilanti and R. Paltauf, Zeitschrift für Krebsfor-
5. L. K. Wolff and G. Ras, Ztrbl. f. Bakt. I. 123, 257
schung 32 (1-2), 372 (1930).
(1931).
35. L. J. Blacher and N. W. Bromley, Wilhelm Roux’ Ar-
6. J. B. Tuthill and O. Rahn, Archiv für Mikrobiologie 4
chiv fur Entwicklungsmechanik der Organismen 122
(1-4), 565 (1933).
(1), 79 (1930).
БИОФИЗИКА том 66
№ 6
2021
1110
ВОЛОДЯЕВ и др.
36. L. J. Blacher and N. W. Bromley, Wilhelm Roux' Ar-
63. A. Gurwitsch and L. Gurwitsch, Archiv für mikroskop-
chiv fur Entwicklungsmechanik der Organismen 123
ische Anatomie und Entwicklungsmechanik 104 (1),
(2), 240 (1931).
109 (1925).
37. L. J. Blacher and O. G. Holzmann, Wilhelm Roux’ Ar-
64. С. Я. Залкинд, Природа, № 3, 11 (1934).
chiv fur Entwicklungsmechanik der Organismen 123
65. Л. Л. Васильев, Вестник знания (19-20),
1185
(2), 230 (1931).
(1925).
38. L. J. Blacher, A. I. Irichimowitsch, L. D. Liosner, et al.,
66. D. N. Borodin, Plant Physiol. 5 (1), 119 (1930).
Roux’ Archiv 127, 339 (1932).
67. A. G. Gurwitsch and N. Gurwitsch, Archiv für
39. A. Potozky, Biochemische Zeitschrift 249, 282 (1932).
mikroskopische Anatomie und Entwicklungsmechanik
40. А. А. Гурвич, Проблема митогенетического излуче-
103 (1), 68 (1924).
ния как аспект молекулярной биологии (Медицина,
68. G. Frank and S. Salkind, Wilhelm Roux' Archiv fur En-
Л., 1968).
twicklungsmechanik der Organismen 108 (4),
596
41. O. Rahn, in Cold Spring Harbor Symposia on Quantita-
(1926).
tive Biology (Berlin, 1934), Vol. II, pp. 226-240.
69. A. Hollaender and E. Schoeffel, Quart. Rev. Biol. 6 (2),
42. G. M. Frank and A. G. Gurwitsch, Wilhelm Roux’
215 (1931).
Arch. Entwickl. Mech. Org. 109 (3), 451 (1927).
70. W. Loos, Jahrb. Wiss. Bot. 72 (4), 611 (1930).
43. A. G. Gurwitsch and G. M. Frank, Comptes Rendus
71. M. Paul, Wilhelm Roux' Archiv fur Entwicklungs-
des Séances de l’Académie des Sciences 184,
903
mechanik der Organismen 128 (1), 108 (1933).
(1927).
72. Rawin, Archiv für Mikroskopische Anatomie und En-
44. G. Frank, Biologisches Zentralblatt 49, 129 (1929).
twicklungsmechanik 101, 53 (1924).
45. T. Reiter and D. Gabor, Arch. Exp. Zellforsch 11, 21
73. N. Wagner, Biol. Zentralbl. 47, 670 (1927).
(1931).
74. M. Kisliak-Statkewitsch, Wilhelm Roux' Archiv fur
46. R. Ruyssen, Acta Brev. Neerl. (3), 141 (1933).
Entwicklungsmechanik der Organismen 109 (2), 283
47. J. Chariton, G. Frank, and N. Kannegiesser, Naturwis-
(1927).
senschaften 18 (19), 411 (1930).
75. J. Magrou, Архив биол. наук, Сер. Б 35 (1), 297
48. G. Protti, Radiobiologia 1 (4), 49 (1930).
(1934).
49. С. Н. Брайнес, Архив биол. наук, Сер. Б 35 (1), 325
76. J. Magrou and M. M. Magrou, Bulletin D’Histologie
(1934).
Appliquée 4, 253 (1927).
50. В. В. Ефимов and С. П. Летунов, Архив биол. наук,
77. Л. Я. Бляхер, в кн. Общая биология (пособие для мед-
Сер. Б 35 (1), 157 (1934).
институтов и университетов), под ред. А. Ф. Шел-
51. A. Hollaender, in Biological effects of radiation, Ed. by
ла (Гос. мед. изд-во, М.-Л., 1933) , ч. 1, сс. 66-72.
B. Duggar (McGraw-Hill Book Company, Inc., NY,
78. A. G. Gurwitsch, in Handbuch der biologischen Arbeits-
London, 1936), Vol. 2, pp. 919-958.
methoden, Ed. by E. Abderhalden (Urban &
52. I. V. Volodyaev and L. V. Beloussov, Front. Physiol. 6
Schwarzenberg, Berlin, Wien, 1929), Vol. V, pp. 1401-
(00241), 1 (2015).
1470.
53. H. Sussmanowitsch, Wilhelm Roux' Archiv fur En-
79. A. G. Gurwitsch and L. D. Gurwitsch, Wilhelm Roux’
twicklungsmechanik der Organismen
113
(4),
753
Archiv fur Entwicklungsmechanik der Organismen 113
(1928).
(4), 740 (1928).
54. L. K. Wolff and G. Ras, Zentralblatt für Bakteriologie,
80. A. Gurwitsch and L. Gurwitsch, Wilhelm Roux' Archiv
Parasitenkunde und Infektionskrankheiten 128, 314
fur Entwicklungsmechanik der Organismen 109 (3),
(1933).
362 (1927).
55. Ю. А. Владимиров, Сверхслабые свечения при биохи-
81. J. B. Bateman, Biol. Rev. 10 (1), 42 (1935).
мических реакциях (Наука, М., 1966).
82. N. Wagner, Planta 5 (1), 70 (1928).
56. С. Родионов и Г. М. Франк, Вопросы свето-биоло-
гии и измерения света.(Гос. техн.-теорет. изд-во,
83. А. Г. Гурвич, Журн. Русского ботанич. об-ва 13, 179
М.-Л., 1934).
(1928).
57. A. Potozky, Biologisches Zentralblatt 50, 712 (1930).
84. B. Rossmann, Wilhelm Roux' Archiv fur Entwicklung-
smechanik der Organismen 114 (4), 583 (1929).
58. А. Г. Гурвич и В. Ф. Еремеев, в кн. Сборник работ
по митогенезу и теории биологического поля, под
85. B. Rossmann, Untersuchungen über die Theorie der mi-
ред. А. Г. Гурвича (Изд-во АМН СССР, М., 1947),
togenetischen Strahlen (Springer Berlin Heidelberg,
сс. 5-19.
2013).
59. А. Г. Гурвич и Л. Д. Гурвич, Введение в учение о ми-
86. A. Gurwitsch, Рус.-нем. мед. журн. 6, 345 (1927).
тогенезе (Изд-во Акад. мед. наук СССР, М., 1948).
87. A. A. Gurwitsch, The problem of mitogenetic radiation as
60. A. G. Gurwitsch, Wilhelm Roux' Archiv fur Entwick-
an aspect of molecular biology (Meditsina, Leningrad,
lungsmechanik der Organismen 116 (1), 20 (1929).
1968).
61. A. Gurwitsch, Protoplasma 26, 23 (1936).
88. A. G. Gurwitsch, Wilhelm Roux' Archiv fur Entwick-
62. С. Я. Залкинд и Г. М. Франк, Митогенетические
lungsmechanik der Organismen 113 (2), 406 (1928).
лучи и деление клеток (Госиздат, М.-Л., 1930).
89. A. G. Gurwitsch, Protoplasma 6 (1), 449 (1929).
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1111
90. 90. Б. П. Токин, Митогенетические лучи (Гос. мед.
дикация и биотестирование (издательский центр
изд-во, М., 1933).
«Академия», М., 2007).
91. G. W. Taylor and E. N. Harvey, Biol. Bull. 61 (3), 280
121. Ш. А. Окада, Радиоционная биохимия клетки
(1931).
(Мир, М., 1974).
92. М. Н. Моисеева, Автореф. дисс. … д-ра биол. наук
122. И. М. Прохорова, М. И. Ковалева, А. Н. Фомиче-
(Киев, 1960).
ва и О. В. Бабаназарова, Пространственная и вре-
93. M. Moissejewa, Ukr. Bot. Rev. (5), 41 (1929).
менная динамика мутагенной активности воды оз.
94. M. Moissejewa, Biochemische Zeitschrift
241,
1
Неро (Наука, М., 2008).
(1931).
123. D. Antosiewicz, Acta Soc. Bot. Pol. 56, 543 (1987).
95. M. Moissejewa, Biochemische Zeitschrift 251,
133
124. G. L. Cabrera and D. M. G. Rodriguez, Fundamental
(1932).
and Molecular Mechanisms of Mutagenesis 426 (2),
96. M. Moissejewa, Biochemische Zeitschrift 243 (1-3),
211 (1999).
67 (1931).
125. M. L. Camparoto, R. Teixeira, and P. Vicentini, Gen.
97. A. Levan, Heredias (24), 9 (1938).
Mol. Biol. 25, 85 (2002).
126. M. P. Chrysouli, C. N. Banti, N. Kourkoumelis, et al.,
98. A. Levan, Nature (156), 751 (1945).
J. Inorgan. Biochem. 179, 107 (2018).
99. A. Levan, Heredias (35), 325 (1949).
127. M. P. Chrysouli, C. N. Banti, I. Milionis, et al., Mater
100.G. Fiskesjö, Hereditas 102, 99 (1985).
Sci. Eng. C Mater. Biol. (93), 902 (2018).
101. G. Fiskesjö, Environmental Toxicology and Water
128. S. M. C. M. A. Ferreira, H. L. C. dos Santos,
Quality 8 (3), 291 (1993).
V. G. Jennifer, et al., Braz. J. Pharm. Sci. 56, 1 (2020).
102. G. Fiskesjö, in Plants for environmental studies (CRC
129. G. Fiskesjö, in Methods in Molecular Biology. Vol. 43.
Press LLC, New York, 1997), pp. 308-333.
In vitro toxicity testing protocols, Ed. by S. O. Hare
103. И. И. Концевая, С. В. Жадько и О. Г. Алексеенко,
and C. K. Atterwell (Humana Press Inc., Totowa,
Разнообразие растительного мира, № 1(4), 81
USA, 1995), pp. 119-127.
(2020).
130. W. F. Grant, Mutat. Res. 99, 273 (1982).
104. I. Milionis, C. N. Banti, I. Sainis, et al., J. Biol. Inor-
131. W. F. Grant and K. D. Zura, in Mutagenicity: new hori-
gan. Chem. 23 (5), 705 (2018).
zons in genetic toxicology, Ed. by J. A. Heddle (Acad.
105. O. A. Asita, S. Moramang, T. Rants’o, et al., Caryolo-
Press, New York, 1982), pp. 407-433.
gia 70 (2), 151 (2017).
132. D. Kumar, A. Rajeshwari, P. S. Jadon, et al., J. Envi-
106. E. Kowalska-Wochna, A. Tylicki, and A. Ościłowicz,
ron. Sci. 38, 150 (2015).
Cell. Mol. Biol. Lett. 7 (2002).
133. L. P. Kumar and N. Paneerselwam, Facta Universita-
107. G. L. R. Dávila, A. J. J. Méndez, and W. M. Arango,
tis. Ser.: Medicine and Biology 15 (1), 20 (2008).
Caryologia 71 (2), 166 (2018).
134. T. H. Ma, Mutat. Res. 99, 257 (1982).
108. R. B. Souza, C. P. De Souza, O. C. Bueno et al., Che-
135. T. H. Ma, Mutat. Res. 426, 103 (1999).
mosphere (168), 1093 (2017).
109. S. Pallavi, M. Singh, H. S. Rathore, et al., Ethnobo-
136. M. H. Nielsen and J. Rank, Hereditas 121 (3), 249
tanical Leaflets 12, 927 (2008).
(1994).
137. J. Rank and M. Nielsen, Environmental Mutagenesis
110. T. A. Yekeen, M. A. Azeez, A. Lateef, et al., Caryolo-
and Related Subjects 312, 17 (1994).
gia 70 (4), 366 (2017).
111. J. Dutta, A. Ahmad, and J. Singh, Caryologia 71 (2),
138. H. S. Rathore, Ethnobotanical Leaflets 10, 92 (2006).
139 (2018).
139. K. Sabti, Cytobios 58, 71 (1989).
112. F. Fatma, S. Verma, A. Kamal et al., Chemosphere
140. I. Sainis, C. N. Banti, A. M. Owczarzak, et al., J. In-
(195), 864 (2018).
organ. Biochem. 160, 114 (2016).
113. K. Sax, Getetics (26), 418 (1941).
141. J. Ślusarczyk, M. Dudek, M. Wierzbicka, et al., Cary-
114. A. Ya. Kocaman and E. Kılıç, Caryologia 70 (3), 229
ologia 67 (3), 250 (2014).
(2017).
142. M. K. Stathopoulou, C. N. Banti, N. Kourkoumelis,
115. T. A. Staykova, E. N. Ivanova, and I. G. Velcheva, J.
et al., J. Inorgan. Biochem. 181, 41 (2018).
Cell Mol. Biol. 4, 41 (2005).
143. И. И. Концевая и Т. А. Толкачева, Веснік Віцеб-
116. D. S. Pesnya, Caryologia 66 (3), 275 (2013).
скага Дзяржаўнага Ўніверсітэта,
№ 6(72), 57
(2012).
117. I. Khan, C. Kant, A. Sanwaria, et al., Heart Views 11
(3), 115 (2010).
144. J. Rank, Ekologija 1, 38 (2003).
118. I.-D. Adler, J. Cole, N. Danford, et al., Guide to short-
145. C. Balog, Cytologia 1982 (47), 689 (1982).
term tests for detecting mutagenic and carcinogenic
146. M. Levine, Cancer Res. 5, 107 (1945).
chemicals (World Health Organization, Geneva, 1985).
147. S. Tedesco and H. Laughinghouse, Bioindicator of
119. E. Bonciu, P. Firbas, C. S. Fontanetti et al., Caryolo-
genotoxicity: the Allium cepa test (InTech, Croatia,
gia 71 (3), 191 (2018).
2012).
120. О. П. Мелехова, Е. И. Егорова, Т. И. Евсеева и др.,
148. M. Wierzbicka and D. Antosiewicz, Acta Soc. Bot.
Биологический контроль окружающей среды: биоин-
Pol. 57 (2), 201 (1988).
БИОФИЗИКА том 66
№ 6
2021
1112
ВОЛОДЯЕВ и др.
149. Т. И. Евсеева, Т. А. Майстренко, С. А. Гераськин
179. C. N. Banti and S. K. Hadjikakou, BioProtoc. 9 (14),
и др., Цитология и генетика, № 5, 73 (2005).
e3311 (2019).
150. M. K. Alotaibi and I. O. Barnawi, Pak. J. Bot. 50 (6),
180. M. Ghosh, M. Bandyopadhyay, and A. Mukherjee,
2245 (2018).
Chemosphere 81, 1253 (2010).
151. Ö. S. Aslantürk and A. Celik Tulay, Pakistan J. Biol.
181. R. Liman, U. G. Gökçe, D. Akyıl, et al., Revista Bra-
Sci. 8 (6), 482 (2005).
sileira de Farmacognosia 22 (3), 541 (2012).
152. Ö. S. Aslantürk, Caryologia 72 (2), 29 (2019).
182. T. G. Mashanda, A dissertation submitted in partial
153. G. A. Bich, M. C. Vedoya, and M. G. Medvedeff, Rev.
fulfilment of the requirements for the bachelor of sci-
Cienc. Tecnol. 14 (18), 39 (2012).
ence honours degree in biological sciences (Bindura
154. A. de la Campo, M. Bracho, L. Marcano, et al., Bio-
University of Science Education, Zimbabwe, 2017).
cell 29 (2), 169 (2005).
183. M. T. Remacle-Dath, Bull. Soc. Royale Sci. Liege 35
155. M. L. Ferrero and C. de la Torre, J. Cell Set. 80, 171
(3-4), 315 (1966).
(1986).
184. M. C. Silva, A. F. Matos, H. L. C. d. Santos, et al.,
156. C. K. Grisolia, A. B. B. de Oliveira, H. Bonfim, et al.,
Braz. J. Pharm. Sci. 56, e18302 (2020).
Gen. Mol. Biol. 28 (2), 334 (2005).
185. T. Sifa, Caryologia 62 (3), 171 (2014).
157. M. F. Knoll, A. C. F. da Silva, T. S. do Canto-Dorow,
186. B. Tutunoglu, O. Aksoy, R. Ozbek, et al., Biologia
et al., Gen. Mol. Biol. 29 (3), 539 (2006).
plantarum 63 (1), 365 (2019).
158. M. Konuk, R. Liman, and İ. H. Ciğerci, Pak. J. Bot.
187. J. F. Lopez-Saez, F. Gonzalez-Bernaldez, A. Gonza-
39 (1), 73 (2007).
lez-Fernandez, et al., Protoplasma 67, 213 (1969).
159. D. Liu and I. Kottke, J. Biosci. 29 (3), 329 (2004).
188. A. M. Guarino, A. Troiano, E. Pizzo, et al., Genes (9),
160. A. Mesi and D. Kopliku, Procedia Technology 8, 19
513 (2018).
(2013).
189. L. P. P. Kumar, N. , Facta universitatis. Ser.: Medicine
161. Y. Mustafa and E. S. Arikan, Caryologia 61 (1), 45
and Biology 14 (2), 60 (2007).
(2014).
190. G. Devnarayan, S. Marmat, H. Rathore, et al., Int.
162. C. A. Ozel, M. Ozkul, F. Unal, et al., Fresenius Envi-
Res. J. Biol. Sci. 5 (8), 53 (2016).
ron. Bull. 27 (12), 8133 (2018).
191. S. A. Ozlem and A. C. Tulay, Pakistan Journal of Bio-
163. A. Özkara, Caryologia 72 (2), 21 (2019).
logical Sciences 8 (6), 482 (2005).
164. H. S. Rathore, S. Bi, S. Anjali, et al., Ethnobotanical
192. В. А. Пухальский, А. А. Соловьев, Е. Д. Бадаева и
Leaflets 10, 272 (2006).
др., Практикум по цитологии и цитогенетике рас-
165. P. K. Renjana, S. Anjana, and J. E. Thoppil, Int. J.
тений (КолосС, М., 2013).
Pharm. Pharm. Sci. 5, 311 (2013).
193. М. Мойсєєва, Вісник Київського Ботанічного Са-
166. C. A. Rosculete, E. Bonciu, E. Rosculete, et al., Int. J.
ду (14), 1 (1932).
Environ. Res. Public Health 16 (1), 75 (2019).
194. A. G. Gurwitsch, Bulletin d’histologie appliquée a la
167. R. Sehgal, S. Roy, and V. L. Kumar, Biocell 30 (1), 9
physiologie et a la pathologie et de technique mi-
(2006).
croscopique 1 (11), 486 (1924).
168. M. I. Soliman, J. Biol. Sci. 1 (11), 1021 (2001).
195. W. W. Siebert, Biochemische Zeitschrift 202, 123
(1928).
169. С. А. Беждугова, Э. М. Гидова, М. М. Биттуева
и др., Международный журнал прикладных и
196. И. Н. Гудков и Д. М. Гродзинский, в сб. Механиз-
фундаментальных исследований, № 3, 96 (2018).
мы радиоустойчивости растений (Наук. думка,
Киев, 1976), сс. 110-137.
170. А. Ю. Буданцев, Фундаментальные исследова-
ния, № 6, 1393 (2014).
197. А. И. Довгалюк, Т. Б. Калиняк и Я. Б. Блюм, Ци-
тология и генетика 35 (1), 3 (2001).
171. П. В. Лебедев, М. Ф. Мельникова и И. А. Уткина,
Цитология 8 (5), 584 (1971).
198. O. Rahn, J. Bacteriol. 28 (2), 153 (1934).
172. О. А. Момот, Автореф. дисс. … канд. биол. наук
199. G. Karsten, Z. Bot. (10), 1 (1918).
(Калуга, 2007).
200. Y. Karsten, Zeit. Bot. 7 (1), 1 (1915).
173. Д. С. Песня, А. В. Романовский и И. М. Прохоро-
201. M. Stalfeit, Studien über die Periodizität der Zellteilung
ва, Ярославский педагогич. вестн., № 3, 80 (2010).
und sich daran anschließende Erscheinungen. Kungl. Sv.
174. Л. Н. Ульяненко, Е. В. Рева и Б. И. Сынзыныс,
Vetenskapakad (Almqvist und Wiksell, Stockholm,
С.-х. биология 52 (1), 183 (2017).
1921).
175. L. Pucciarini, F. Ianni, V. Petesse, et al., Molecules 24
202. H. H. Laughlin, (Publ. Cam. Inst. Wash., 1919), Vol.
(3) (2019).
265, pp. 1-48.
176. G. Fiskesjo, Vatten 37, 232 (1981).
203. А. Г. Гурвич, Архив биол. наук 46 (3), 3 (1937).
177. Т. Евсеева, Е. Белых и Т. Майстренко, Вестн. Ин-
204. B. Rossmann, Wilhelm Roux' Archiv fur Entwick-
ститута биологии Коми НЦ УрО РАН, № 9, 2
lungsmechanik der Organismen 113 (2), 346 (1928).
(2005).
205. F. Wassermann, Bd. 11, 1931, Arch. Experim. Zellfor-
178. Т. И. Евсеева, С. А. Гераськин и Е. С. Храмова,
schung 11 (1931).
Радиоэкология. Охрана окружающей среды, № 3,
206. A. G. Gurwitsch, Botanical Gazette 80 (2),
224
143 (2001).
(1925).
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1113
207. A. G. Gurwitsch, Archiv für mikroskopische Anato-
237. W. A. Jensen и Z. Y. Kavaljian, Am. J. Bot. 45 (5), 365
mie und Entwicklungsmechanik
103
(3-4),
490
(1957).
(1924).
238. А. Г. Гурвич и Л. Д. Гурвич, (1926).
208. L. D. Gurwitsch, Pflüger's Archiv für die gesamte
239. W. Schwarz, Biol. Zentralbl. 48, 302 (1928).
Physiologie des Menschen und der Tiere 197 (1/2)
240. З. П. Ростовцева, Верхушечная меристема (Изд-во
(1922).
МГУ, М., 1969).
209. P. G. Rusinoff, Archiv für mikroskopische Anatomie
241. R. Hagemann, Kulturpflanze (4), 46 (1956).
und Entwicklungsmechanik 104 (1), 121 (1925).
242. И. А. Уткина, Автореф. дис
канд. биол. наук
210. A. G. Gurwitsch, in Atti del I Congresso Interanzionale
(Ин-т экологии растений и животных УНЦ АН
di electro-radio-biologia, Ed. by L. Capelli (Bologna
СССР, 1972).
Venezia, 1934), Vol. II.
243. И. А. Уткина and И. М. Синягина, в сб. Материа-
211. W. E. Kellicott, Bull. Torrey Bot. Club. (31), 529
лы по экологии и физиологии растений уральской
(1904).
флоры (УрГУ, Свердловск, 1976), сс. 42-50.
212. В. Г. Гриф, Цитология 1 (2), 229 (1959).
244. R. М. Savelkoul, Am. J. Bot. 44 (4), 311 (1957).
213. В. Г. Гриф, Цитология 36 (11), 1069 (1994).
245. C. E. Stephens, Cytologia 40, 607 (1975).
246. C. E. Stephens, Cytologia 49, 679 (1984).
214. С. Н. Мовсесян, Научн. тр. Арм. отд. Всес. ботан.
об-ва (5), 129 (1970).
247. J. Schwemmle, Biologisches Zentralblatt (49), 421
(1929).
215. E. Edgar, Fluctuation of mitotic index in the shoot apex
248. Л. В. Белоусов, В. Л. Воейков и Ф.-А. Попп, При-
of Lonicera nitida (University of Canterbury, Canter-
рода, № 3, 64 (1997).
bury, N.Z., 1961).
249. A. Barbеrio, in Water Treatment, Edited by Walid
216. R. C. Friesner, Am. J. Bot. 7 (9), 380 (1920).
Elshorbagy and Rezaul Kabir Chowdhury
(2013),
217. V. G. Grif, V. B. Ivanov, and E. M. Machs, Цитоло-
pp. 317-334.
гия 44 (10), 936 (2002).
250. A. G. Gurwitsch, Radiobiologia 1 (1), 15 (1932).
218. A. Lance, Ann. Sci. natur. Bot. et bid. veg. 11 (13), 301
251. Л. В. Высоцкая, Вавиловский журн. генетики и
(1952).
селекции 18 (1), 81 (2014).
219. H. Rоtta, Planta (37), 399 (1949).
252. G. Frank, Biologisches Zentralblatt 52 (1), 1 (1932).
220. М. Tourte, Rev. Cyt. et Biol, veg. (29), 199 (1966).
253. A. Hollaender and W. D. Claus, An experimental study
221. Г. Г. Батикян and Е. Г. Симонян, Биол. журн. Ар-
of the problem of mitogenetic radiation. (National re-
мении 23 (11), 96 (1970).
search council of the National academy of sciences,
222. А. Ю. Буданцев, Изв. РАН. Сер. биол., № 3, 293
Washington, 1937).
(2009).
254. O. W. Richards and G. W. Taylor, Biol. Bull. 63 (1),
223. R. C. Bishop and R. M. Klein, Genome 13 (3), 597
113 (1932).
(2011).
255. A. Hollaender and W. D. Claus, J. Optic. Soc. Amer.
224. A. C. Chandler, Biol. Bull. (112), 305 (1957).
25, 270 (1935).
225. C. E. Droogleever-Fortuyn, Day and night period in
256. В. Ф. Сарафанов, Архив биол. наук. Сер. Б 35 (1),
nuclear divisions (1926).
309 (1934).
226. S. M. Erickson and W. S. Hillman, Genime 18 (1), 189
257. Е. Е. Авчина, Дисс. … канд. мед. наук (Институт
(2011).
онкологии, Ленинград, 1950).
227. W. A. Jensen and Z. Y. Kavaljian, Am. J. Bot. 45 (5),
258. О. Е. Нудольская, Дисс. … д-ра мед. наук (2-й
365 (1958).
Московский государственный медицинский ин-
ститут, Москва, 1945).
228. A. Lewis, Bot. Gaz. (32), 423 (1901).
259. В. И. Иоффе and Е. С. Биллиг, Архив биол. наук
229. J. F. Lopez-Saez and E. Fernandez-Gomez, Experi-
40 (1), 5 (1935).
entia (21), 591 (1965).
260. K. H. Kreuchen and J. B. Bateman, Protoplasma 22
230. M. Jangala, M. Santoshi, M. Suman, et al., Int. J.
(1), 243 (1934).
Toxicol. Appl. Pharmacol. 2 (2), 18 (2012).
261. M. Nakaidzumi and H. Schreiber, Biochemische
231. A. M. A. Matias and I. K. C. Fontanilla, Science Dili-
Zeitschrift (237), 358 (1931).
man 23 (1), 43 (2011).
262. J. Westenberg, Die "slide-cell" Methode von Wolff und
232. Y. M¸kinen, Ann. Bot. Vanamo 34 (6), 1 (1963).
Ras zum Nachweis von Gurwitsch-Strahlen (N.V.
233. M. D. Solomon and J. A. Trent, Trans. Kansas Acad.
Noord-Hollandsche Uitgeversmaatschappij, Amster-
Sci. 44, 202 (1903).
dam, 1935).
234. A. Gonzalez-Fernandez, J. Lopez-Saez, and
263. T. I. Quickenden and S. S. Q. Hee, Speculat. Sci.
G. Gimenez-Martin, Exp. Cell Res. (43), 255 (1966).
Technol. 4 (5), 453 (1981).
235. А. Ю. Буданцев и А. Ю. Демьянов, Цитология 59
264. T. I. Quickenden and R. N. Tilbury, Radiat. Res. 102,
(6), 447 (2017).
254 (1985).
236. F. Wassermann, Klinische Wochenschrift 9 (10), 433
265. F. Grasso, F. Musumeci, A. Triglia, et al., Photochem.
(1930).
Photobiol. 54 (1), 147 (1991).
БИОФИЗИКА том 66
№ 6
2021
1114
ВОЛОДЯЕВ и др.
266. M. Wainwright, K. Killham, C. Russell, et al., Micro-
295. A. A. Gurwitsch, Annales de Physiologie 14 (2), 182
biol.-UK 143 (1), 1 (1997).
(1938).
267. И. В. Володяев, Р. Н. Ивановский, А. С. Богачук
296. H. Gesenius, Radiobiologia 1 (2), 33 (1932).
и др., в сб. Матер. V Междунар. конгр. «Слабые и
297. А. Г. Гурвич, Л. Д. Гурвич, С. Я. Залкинд и др.,
сверхслабые поля и излучения в биологии и медицине»
Учение о раковом тушителе: Теория и клиника
(Изд-во АМН СССР, Москва, 1947).
(Санкт-Петербург, 2009), с. 128.
298. W. Siebert, in Handbuch der allgemeinen Hämatologie,
268. I. V. Volodyaev, R. N. Ivanovsky, A. S. Bogachuk,
Ed. by H. Hirschfeld and A. Hittmair (Urban &
et al., in Abstr. Res. Symp. “Progress In Electromagnet-
Schwarzenberg, Berlin, Wien, 1934), Vol. 2, p. 1339.
ics” (Moscow, Russia, 2009), p. 1025.
299. H. Gesenius, Biochemische Zeitschrift 226 (4-6), 257
269. I. V. Volodyaev, E. N. Krasilnikova, and R. N. Iva-
(1930).
novsky, PloS One 8 (4), e62808 (2013).
300. M. Heinemann, Klinische Wochenschrift 11 (33),
270. И. В. Володяев и Е. В. Наумова, в сб. Матер. VIII
1375 (1932).
Междунар. конгр. «Слабые и сверхслабые поля и из-
301. С. Я. Залкинд, Природа 1, 53 (1938).
лучения в биологии и медицине» (Санкт-Петербург,
302. Я. С. Кленицкий, Архив биол. наук, Сер. Б 35 (1),
2018), сс. 21-22.
213 (1934).
271. С. Я. Залкинд, Природа, № 2, 111 (1932).
303. 303. М. Б. Новиков, Архив биол. наук 51 (3), 56
272. T. Reiter and D. Gabor, Strahlenther Onkol. (28), 125
(1938).
(1928).
304. Б. С. Песоченский, в Сборник работ по митогенезу
273. J. Ponomarewa, Biochemische Zeitschrift 239, 424
и теории биологического поля, под ред. А. Г. Гурви-
(1931).
ча (Изд-во АМН СССР, Москва, 1947), сс. 102-
114.
274. G. Decker, Protoplasma 25 (1), 515 (1936).
275. W. Frankenburger, Strahlenther Onkol. 47 (2), 233
305. E. G. Prokofiewa, Nature 134 (3389), 574 (1934).
(1933).
306. А. Е. Браунштейн и А. П. Потоцкая, Архив биол.
276. A. Gurwitsch and L. Gurwitsch, L'analyse mi-
наук, Сер. Б 35 (1), 73 (1934).
togénétique spectrale (Hermann & Cie, Paris, 1934).
307. Н. Н. Каннегисер, Ю. А. Харитон и Г. М. Франк,
277. А. Гурвич, Л. Д. Гурвич и А. А. Слюсарев, Архив
в кн. Биофизика живой клетки. Избранные труды.
биол. наук 55 (2), 104 (1939).
(Наука, М., 1982), сс. 161-167.
278. A. G. Gurwitsch and L. D. Gurwitsch, Nature 143
308. К. А. Голшмид and В. К. Модестов, в Сборник ра-
(3633), 1022 (1939).
бот Пермского отделения Уральского филиала Цен-
279. A. G. Gurwitsch and L. D. Gurwitsch, Die mitoge-
трального института курортологии (Пермь,
netische Strahlung (VEB Gustav Fischer Verlag, Jena,
1934), сс. 43-50.
1959).
309. Е. Т. Кулин, Здравоохранение Белоруссии (12), 17
280. H. A. Gordon, Acta Physicochim. USSR 13 (3), 393
(1957).
(1940).
310. I. Langmuir and R. N. Hall, Phys Today 42 (1989).
281. P. Pringsheim and A. L. Reimann, Zeitschrift für
311. Anonim, Nature (December, 11), 1007 (1937).
Physik 29 (1), 115 (1924).
312. J. Gray and C. Ouellet, Proc. Roy. Soc. Lond. Ser. B
282. P. Pringsheim, Phosphoreszenz und Fluoreszenz, 3rd ed.
114 (786), 1 (1933).
(Springer, Berlin, 1928).
313. E. Lorentz, Phys. Rev. (44), 329 (1929).
283. N. Kannegiesser, Biochemische Zeitschrift 236, 415
314. Г. Барт, Архив биол. наук 46 (1), 153 (1937).
(1931).
315. А. Л. Юделес, Дис. … д-ра мед. наук (Свердловск,
284. Л. Д. Гурвич, Архив биол. наук, Сер. Б 35 (1), 141
1944).
(1934).
316. В. П. Нагорянская, Дисс. … канд. мед. наук (Цен-
285. В. Ф. Еремеев, Бюл. эксперим. биологии 45 (6), 95
тральный Онкологический Институт НКЗ
(1958).
РСФСР, Москва, 1945).
286. С. Я. Залкинд, Природа, № 6, 53 (1938).
317. W. S. Metcalf and T. I. Quickenden, Nature 216
287. А. Г. Гурвич и Л. Д. Гурвич, Бюл. эксперим. био-
(5111), 169 (1967).
логии и медицины 4 (6), 474 (1937).
318. С. Я. Залкинд, Архив биологических наук 57 (2-
288. А. Г. Гурвич и Л. Д. Гурвич, Бюл. ВИЭМ (6-7), 29
3), 124 (1940).
(1935).
319. A. A. Gurwitsch, Experientia 44 (7), 545 (1988).
289. A. A. Gurwitsch, Ann. de Physiol. et. Physicochem.
320. V. L. Voeikov and L. V. Beloussov, in Biophotonics and
Biol. 10 (5), 1153 (1934).
Coherent Systems in Biology (Springer US, Boston,
290. G. S. Kalendaroff, Pflüger's Archiv für die gesamte
MA, 2007), pp. 1-16.
Physiologie des Menschen und der Tiere 231, 238
321. E. V. Naumova, A. E. Naumova, D. A. Isaev, et al., J.
(1933).
Biomed. Photonics Engineer. 4 (4), 040201 (2018).
291. И. В. Цоглина, Архив биол. наук, Сер. Б 35 (1), 341
(1934).
322. R. N. Tilbury and T. I. Quickenden, J. Bioluminesc.
Chemiluminesc. 7 (4), 245 (1992).
292. Г. С. Календаров, Архив биол. наук 32 (1), 26
(1934).
323. Н. А. Троицкий, С. В. Конев и М. А. Катибников,
Биофизика 6 (2), 238 (1961).
293. Г. С. Календаров, Архив биол. наук, Сер. Б 35 (1),
183 (1934).
324. L. V. Beloussov, Eur. J. Biophys. 1 (1), 6 (2013).
294. A. A. Gurwitsch, Annales de Physiologie et de Physi-
325. M. Kobayashi and H. Inaba, Appl. Optics 39 (1), 183
cochimie biologique 10 (5), 1166 (1934).
(2000).
БИОФИЗИКА том 66
№ 6
2021
МЕТОДЫ ИССЛЕДОВАНИЯ СВЕРХСЛАБОГО СВЕЧЕНИЯ
1115
Methods of Studying Ultraweak Photon Emission from Biological Objects.
II. Methods Based on Biological Detection
I.V. Volodyaev*,
*, I.I. Kontsevaya**,
L.V. Beloussov
A.E. Naumova***, and E.V. Naumova****
*Lomonosov Moscow State University, Leninskie Gory 1, Moscow, 119991 Russia
**Francysk Skaryna Gomel State University, ul. Sovetskaya 104, Gomel, 246019 Republic of Belarus
***St. Petersburg State University, Universitetskaya nab. 7/9, Saint Petersburg, 199034 Russia
****Rzhanov Institute of Semiconductor Physics, Siberian Branch of the Russian Academy of Sciences,
prosp. Akademika Lavrentieva 13, Novosibirsk, 630090 Russia
The second part of the review, which is devoted to the methods of studying ultraweak photon emission, pres-
ents a methodology of biological detection. During the first 25 years of research (1923-1948), almost all ex-
periments in this area were based on biological detection and only the component belonging to the middle
ultraviolet region of the spectrum (also called mitogenetic radiation at that time) was studied, because it was
observed due to the influence of ultraweak ultraviolet radiation on the mitotic regime of the biological detec-
tor. These experiments remain open to debate till present due to the insufficient validation of methods. Many
of the results obtained from these investigations have been very impressive for that time period and are now
verified, the research questions the other results relate to are of considerable interest (for instance, fundamen-
tal results concerning cell division and carcinogenesis, early cancer diagnostics). Thus, the analysis of the
methodology employed is an emerging issue for the subsequent experimental verification. The details of the
methods with the most common biological detectors (onion roots, yeast and bacterial cultures) are analysed
and the onion detection method is compared to the closest analogue (the Allium-test).
Keywords: ultraweak photon emission from biological objects, mitogenetic radiation, mitogenetic effect, biological
test systems, Allium- test, biophotonics
БИОФИЗИКА том 66
№ 6
2021