БИОФИЗИКА, 2021, том 66, № 6, с. 1123-1132
БИОФИЗИКА КЛЕТКИ
УДК 577.352.5
ГИПОТЕЗА КОНТАМИНАЦИИ МЕМБРАН НЕЙРОНОВ, ЭПИЛЕПСИЯ
И КЕТОГЕННАЯ ДИЕТА
© 2021 г. Ю.Д. Нечипуренко*, Р.С. Гарсия Рейес**, Х.Л. Эрнандес Касерес**
*Институт молекулярной биологии им. В.А. Энгельгардта РАН, 119991, Москва, ул. Вавилова, 32
E-mail: nech99@mail.ru
**Кубинский Центр нейронаук, 11600, Плайя, Гавана, 25-я ул., 15202, Куба
E-mail: jose.caceres@cneuro.edu.cu
Поступила в редакцию 01.07.2021 г.
После доработки 30.08.2021 г.
Принята к публикации 31.08.2021 г.
В свете новых данных рассматривается гипотеза Альтрупа о контаминации мембран нейронов при
эпилепсии. Эта гипотеза связывает пароксизмальную деполяризацию, наблюдаемую при эпилеп-
тической активности, с пейсмекерным потенциалом одиночного нейрона. Физико-химические ха-
рактеристики мембраны, текучесть и контаминации влияют на ее способность проводить импульс
и перезаряжаться. Обсуждаются предложенные ранее способы лечения эпилепсии, основанные на
кетогенной диете и их возможные объяснения в связи с гипотезой Альтрупа. Среди возможных ме-
ханизмов действия кетогенной диеты подчеркивается антиэпилептическое действие кетоновых тел,
роль повышенного синтеза глутатиона и эффект полиненасыщенных жирных кислот и холестери-
на, включенных в состав диеты. Эти три механизма приводят к регуляции текучести и других био-
физических свойств липидного бислоя, а также к деконтаминации мембраны от амфифильных
примесей, в соответствии с гипотезой Альтрупа.
Ключевые слова: эпилептическая активность, пароксизмальная деполяризация, пейсмейкерный потен-
циал, гипотеза Альтрупа, контаминация мембран, латеральное давление, кетогенная диета, глутати-
он, полиненасыщенные жирные кислоты.
DOI: 10.31857/S0006302921060077
есть гигантский возбуждающий постсинаптиче-
Эпилепсией страдают около 50 млн человек во
ский потенциал. В нейрофизиологии принято
всем мире [1], описано более сорока клинических
изучить любое явление в простейшем объекте, где
типов эпилепсии [2]. Несмотря на это разнообра-
это явление может проявляться. Это позволило Э.
зие, общим электрофизиологическим следом для
Канделю изучить основы механизмов памяти в
большинства типов эпилептической активности
нервной системе аплизии [8], а А. Ходжкину и Э.
является пароксизмальная деполяризация (ПД).
Хаксли - основы нервного импульса в гигант-
Во время эпилептической активности некоторые
ском аксоне кальмара [9].
нейроны проявляют большую и длительную де-
поляризацию, это и есть пароксизмальная депо-
Рядом исследователей изучалось возникнове-
ляризация мембранного потенциала (см. работу
ние и распространение эпилептической активно-
[3] и рис. 1).
сти в буккальных ганглиях наземной улитки Helix
Наши знания о нейрофизиологических осно-
pomatia [7, 10-12]. Helix имеет довольно простую
вах эпилепсии далеки от полноты, и есть основа-
нервную систему, что способствует проведению
ния для надежды, что понимание биофизических
биофизических экспериментов [13, 14]. В частно-
основ ПД позволит прояснить патофизиологию
сти, оба буккальных ганглия Helix имеют по четы-
эпилепсии и очертить рациональные способы ее
ре гигантских нейрона с каждой стороны, и из
лечения. В рамках доминирующей идеи о том, что
них только нейрон Б3 способен систематически
эпилепсия является результатом дисбаланса меж-
развивать ПД в присутствии проконвульсивных
ду возбуждением и торможением в нервной ткани
средств, таких как пентилентетразол, этомидат,
[4-6], авторы работы [7] предложили, что ПД и
гептанол (а также при пониженной концентра-
ции магния и т.п.).
Сокращения: ПД - пароксизмальная деполяризация, КД -
кетогенная диета, ПНЖК - полиненасыщенные жирные
Уже давно обсуждается сходство между пове-
кислоты.
дением этой простой клеточной модели и разви-
1123
1124
НЕЧИПУРЕНКО и др.
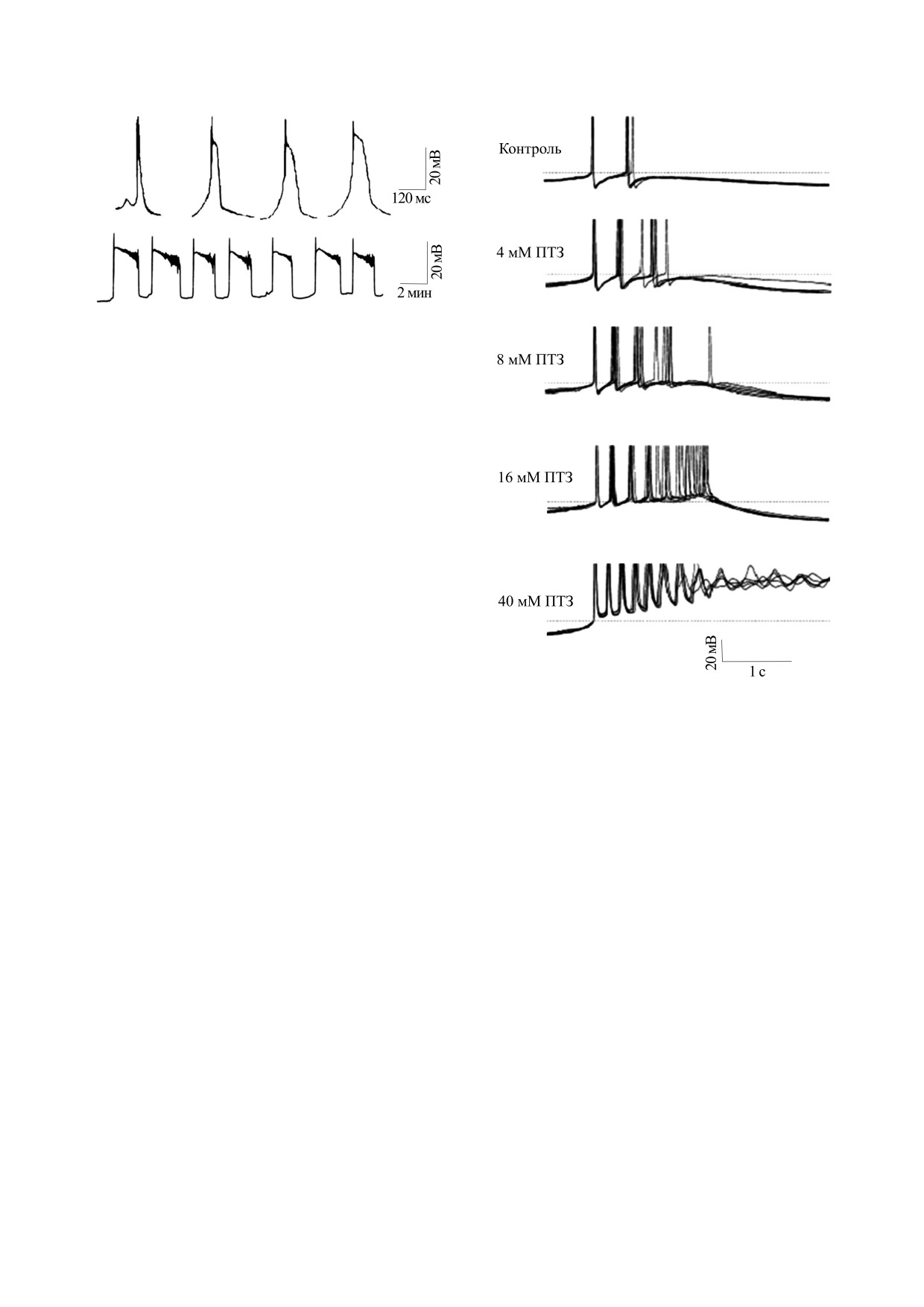
Рис. 1. Сравнение пароксизмальных деполяризаций,
регистрируемых внутриклеточно в нейронах коры
кошки (вверху) и в нейроне Helix Б3 (внизу). Верхняя
часть рисунка - адаптировано из работы [3] (измере-
ния проводились с местным применением пентилен-
тетразолa), нижняя часть рисунка - после примене-
ния 40 мМ пентилентетразола (неопубликованные
данные авторов).
тием эпилепсии у человека, и сегодня нет сомне-
ний в том, что с точки зрения фундаментальных
механизмов у этих процессов существует ряд об-
щих черт, несмотря на большое филогенетиче-
ское расстояние между человеком и улиткой [10].
Идея о том, что пейсмейкерные потенциалы
являются основой эпилептиформной активности
в нервной системе, обсуждалась в литературе [14].
Возможность генерации ПД в полностью изоли-
рованных нейронах, как у беспозвоночных [10],
так и у позвоночных [11], и тот факт, что ПД со-
провождаются увеличением импеданса нейронов
[7, 12], не согласуется с идеей, что ПД имеют си-
Рис. 2. Постепенное преобразование пейсмейкерного
наптическую природу. С другой стороны, в экс-
потенциала в ПД с увеличением концентрации пен-
периментах на нейроне Б3 было обнаружено, что
тилентетразола (ПТЗ). Изображение составлено на
с увеличением дозы пентилентетразола (как и
основе данных, опубликованных в работе [15].
этомидата) пейсмейкерные потенциалы посте-
пенно трансформируются, их продолжитель-
было проведено математическое моделирование
ность и амплитуда увеличиваются, и можно пред-
и определена область биофизических парамет-
положить, что они превращаются в ПД (см.
ров, обеспечивающая переход от пейсмейкерного
рис. 2).
потенциала к ПД [23]. Было показано, что ПД-
Механизмы генерации пейсмекерных потен-
подобные потенциалы могут появляться как за
циалов еще не выяснены полностью (см., напри-
счет изменения параметров модели, так и посред-
мер, работы [16, 17]). «Минимальная» математи-
ством добавления шума на вход модели (см.
ческая модель для пейсмейкерного потенциала
рис. 3). При уровне шума 0.2 усл. ед. пейсмекер-
нейронов виноградной улитки Helix pomatia была
ные потенциалы удлиняются и проявляют стрем-
предложена в работах Н.И. Кононенко с соавто-
ление к непостоянности, при уровне 0.6 усл. ед.
рами [18-22]. Модель вводит в рассмотрение, во-
возникает гигантская продолжительная деполя-
первых, две стационарные, потенциал-независи-
ризация, напоминающая ПД (рис. 3б,в) [23].
мые проводимости для натрия и калия, которые
образуют линейный ток утечки, во-вторых, на-
триевую стационарную потенциал-зависимую
ГИПОТЕЗА КОНТАМИНАЦИИ МЕМБРАНЫ
S-образную проводимость, которой соответству-
ет отрицательная область сопротивления мем-
У. Альтруп (Ulrich Altrup, 1943-2007), возглав-
бранной вольт-амперной характеристики, в-тре-
лявший Исследовательский институт экспери-
тьих, проводимость, активируемую гиперполяри-
ментальной эпилепсии в Университете Мюнсте-
зацией. С использованием модели Кононенко
ра (Германия), изучал эпилептиформную актив-
БИОФИЗИКА том 66
№ 6
2021
ГИПОТЕЗА КОНТАМИНАЦИИ МЕМБРАН НЕЙРОНОВ
1125
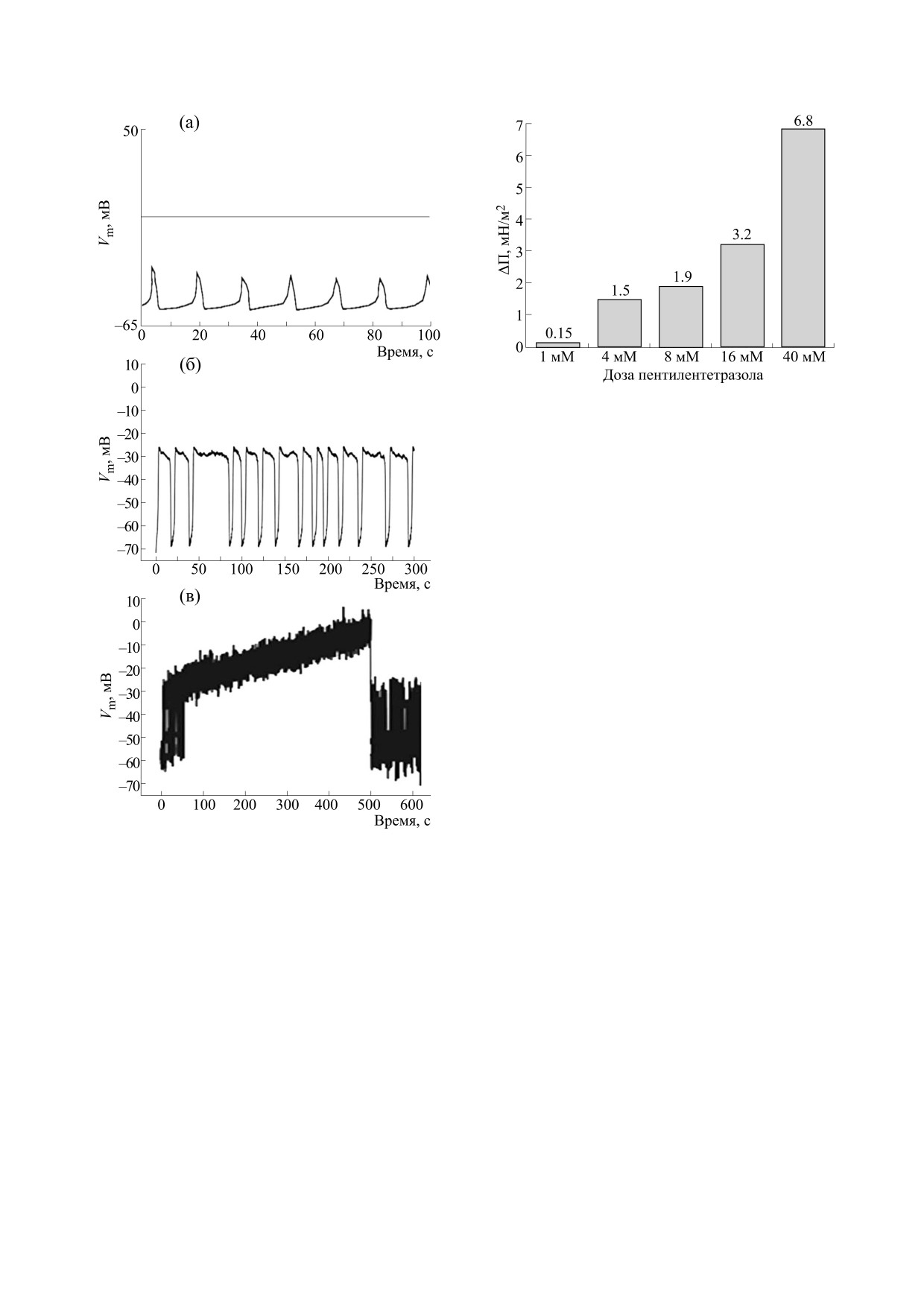
Рис. 4. Средние значения изменения латерального
давления на весах Вильгельми при добавлении раз-
личных доз пентилентетразола. Диаграмма построена
на основе данных, опубликованных в работе [15].
новной механизм эпилептогенеза должен быть
неспецифическим.
К этому следует добавить, что разнообразные
ионные механизмы, связанные с каналами утеч-
ки, калиевыми каналами, большим набором ли-
ганд-зависимых каналов и др., подвержены влия-
нию в одном и том же диапазоне концентраций
пентилентетразола (а также этомидата). Эти на-
блюдения указывают на существование неспеци-
фического механизма эпилептогенеза. Альтруп с
соавторами обнаружил при помощи весов Виль-
гельми, что эпилетогенные вещества пентилен-
тетразол и этомидат вызывают в одном и том же
диапазоне эпилептогенных концентраций посте-
пенное повышение латерального давления ли-
пидных монослоев [7]. На рис. 4 приводятся ре-
зультаты этих экспериментов при добавлении
пентилентетразола.
Совокупность экспериментальных данных
привела Альтрупа к гипотезе, что эпилептоген-
ные амфифильные вещества проникают в липид-
Рис. 3. Постепенное преобразование пейсмейкерного
ный бислой мембраны нервной клетки и что эти
потенциала в ПД при добавлении шума. Расчеты про-
включения увеличивают латеральное давление в
водили при помощи модели Кононенко [20]: (а) -
спонтанные пейсмекерные потенциалы, полученные
бислое, вызывая ПД в тех нейронах, которыe об-
авторами работы [21]; (б) и (в) - результат добавления
ладают способностью генерации пейсмейкерных
белого гауссового шума (0.2 и 0.6 усл. ед. соответ-
потенциалов. Повышенное латеральное давление
ственно).
снижает текучесть мембраны, влияя на подвиж-
ность мембранных белков и вызывая переход от
жидкого конденсированного состояния к твердо-
ность в буккальных ганглиях Heliх почти 30 лет. В
му конденсированному состоянию.
последние годы деятельности он выдвинул гипо-
Белковo-липидные взаимодействия и возбуди-
тезу мембранной контаминации. Альтруп пола-
мость нейронов. Белковo-липидные взаимодей-
гал, что если вещества, резко различающиеся по
ствия и их связь с возбудимостью обсуждались в
механизму действия, такие как гептанол, пенти-
течение нескольких десятилетий в биофизиче-
лентетразол, этомидат (как и низкая концентра-
ских исследованиях [24]. Так, авторы работы [25]
ция магния), способны индуцировать ПД, то ос-
обнаружили, что агенты, изменяющие химико-
БИОФИЗИКА том 66
№ 6
2021
1126
НЕЧИПУРЕНКО и др.
физические свойства мембран (например, про-
мом деле это результат сочетания эффектов как
стые спирты и ацетон), способны изменять срод-
уже открытых каналов, так и закрытых или инак-
ство Na,K-ATФазы к моновалентным катионам.
тивированных. Прямая механическая стимуля-
ция аксона могла бы вызывать открытие натрие-
Наиболее известной моделью генерации и рас-
вых каналов, в ходе которого генерируется энер-
пространения нервного импульса является мо-
гия, необходимая для поддержания движения
дель Ходжкина и Хаксли [9]. Ее биофизическая
механического солитона [32].
правдоподобность и способность объяснить мно-
гие свойства возбуждения нейронов привели к
Если вышеописанные изменения механиче-
тому, что эта модель получила всеобщее призна-
ских свойств двойного липидного бислоя, сопро-
ние в нейронауках и кардиологии. А. Ходжкин и
вождающие потенциал действия, окажутся тесно
Э. Хаксли обнаружили экспериментально, что
связаны с процессом нервной активности, то ста-
электрическая емкость мембраны остается неиз-
нет понятным, почему любое вещество, способ-
менной во время возбуждения. Поэтому роль ли-
ное изменять физико-химические свойства двой-
пидов в этой модели игнорируется и любая моди-
ного липидного слоя, может изменять и возбуди-
фикация ионных каналов рассматривается как
мость нейронов. Такой подход мог бы дать
результат взаимодействий электростатической
правдоподобное объяснение эффекту ингаляци-
природы. В начале 1970-х годов И. Тасаки пред-
онных анестезий, эффективность которых корре-
ложил теорию, которая не вписывается в модель
лирует с растворимостью применяемых веществ в
Ходжкина и Хаксли: он предположил, что рас-
липидах [33], а также известному антиэпилепти-
пространение нервного импульса сопровождает-
ческому эффекту ряда пищевых добавок, таких
ся волной механического возбуждения вдоль ак-
как омега-3-полиненасыщенные жирные кисло-
сона [26]. Эти идеи не нашли экспериментально-
ты [34], витамин Д [35] и др. [36]. Можно также
го подтверждения, в то время как регистрация
ожидать, что изменение механических свойств
воротных токов и токов через единичные ионные
неспецифических мембранных эпилептогенных
каналы наряду с другими наблюдениями свиде-
амфифильных веществ повлияет и на пейсмей-
тельствовали в пользу модели Ходжкина и Хаксли
керные потенциалы, вызывая аномальные изме-
[27].
нения, которые приводят к эпилептогенезу, в со-
ответствии с гипотезой контаминации мембран
В последнее время появились доказательства
[37].
того, что распространение потенциала действия
Кетогенная диета и гипотеза контаминации мем-
сопровождается волной механической деформа-
бран. Около 30% пациентов с эпилепсией рези-
ции клеточной мембраны, которая движется с по-
стентны к лекарственной терапии (это более чем
стоянной скоростью [28]. Разные авторы стара-
15 млн человек в мире). Одним из терапевтиче-
ются построить модель, способную описать все
электрохимические, механические и термодина-
ских вмешательств, способных улучшить состоя-
ние более половины пациентов с тяжелой эпи-
мические явления, связанные с генерацией и
лепсией, является кетогенная диета. Типичная
проведением нервного импульса. Гипотеза о ме-
кетогенная диета (КД) очень богата жирами и
ханическом солитоне кажется нам самой привле-
бедна углеводами (обычно пропорция жиров к
кательной. Однако понадобится целый ряд экс-
периментальных доказательств, а также теорети-
нежирам равна четырем к одному) и применяется
для лечения рефрактерной эпилепсии. Значение
ческих разработок, чтобы идея о нервном
такой диеты в клиническом контексте было отме-
импульсе как о солитоне нашла подтверждение. В
чено еще в 1920-х годах в связи с успешным ее
работах [28-31] было высказано предложение,
применением для лечения эпилепсии.
что открытие ионных каналов, участвующих в ге-
нерации нервного возбуждения, требует механи-
Первая ссылка на то, что пост может оказывать
ческих изменений в двойном липидном слое. В
антиэпилептическое действие, появилась в 1911 г.
пользу этого предположения приводится аргу-
Парижские врачи Ж. Гуелпа и А. Мари использо-
мент, что во время активации ионных каналов
вали пост для лечения двадцати пациентов и со-
происходит деформация мембраны [28, 29]. С
общили об облегчении их симптомов [38]. В нача-
другой стороны, измерения энтальпии во время
ле 1920-х годов Х. Конклин предположил, что
распространения потенциала действия не соот-
эпилепсия проявляется в результате «интоксика-
ветствуют предсказаниям Ходжкина и Хаксли, но
ции мозга веществами, поступающими из кишеч-
поддерживают модель механического импульса
ника». Конклин использовал диету, состоящую
[30, 31].
только из воды, в течение 30 дней для лечения па-
циентов с рефрактерной эпилепсией и достиг
Авторы работы [31] утверждают, что потенци-
успеха - улучшения состояния у более половины
ал действия, смоделированный Ходжкиным и
пациентов [39].
Хаксли, отражает сумму всех потенциальных из-
менений по всей мембране, зарегистрированных
Представление о том, что кетоновые тела яв-
на относительно большой площади, так что на са-
ляются главным эффектором в противоэпилеп-
БИОФИЗИКА том 66
№ 6
2021
ГИПОТЕЗА КОНТАМИНАЦИИ МЕМБРАН НЕЙРОНОВ
1127
тическом механизме поста, привело в 1921 г. Р.
низм начинает использовать жирные кислоты.
Уайлдера к идее предложить диету с высоким со-
Обычно это происходит посредством β-окисле-
держанием жира и низким содержанием углево-
ния, и в результате генерируется ацетил-КоА, ко-
дов для лечения эпилепсии. Он получил обнаде-
торый входит в цикл Кребса для генерации АТФ.
живающие результаты и назвал такую диету кето-
Однако при длительном голодании не весь аце-
генной
[40]. Полезные эффекты этой диеты
тил-КоА может метаболизироваться через цикл
подтвердились к 1930-м годам, но потом она пе-
Кребса, и значительная часть ацетил-КоА преоб-
рестала использоваться врачами, что было связа-
разуется в кетоновые тела: ацетоацетат и β-гид-
но с появлением в 1938 г. фенитоина и позже -
роксибутират, а также ацетон (образующийся пу-
других антиэпилептиков. Однако за последние
тем неферментативного декарбоксилирования
30 лет эпилепсия, невосприимчивая к фармако-
ацетоацетата). При этом кетоновые тела могут
терапии, привлекла внимание научного сообще-
обеспечить до двух третей энергетических по-
ства, и кетогенная диета снова вызвала интерес
требностей мозга.
специалистов. Единственным диетическим мето-
Кетогенная диета, влияя на общий энергети-
дом лечения эпилепсии, имеющим научное обос-
ческий обмен веществ, может создавать смеще-
нование и разработанные принципы практиче-
ния в метаболизме, приводящие к снижению вы-
ского применения, является именно кетогенная
работки лактата и альфакетоглутарата. В резуль-
диета, физиологической основой которой явля-
тате уменьшается концентрации глутамата и
ется создание в организме условий, сходных с те-
снижается риск эксайтотоксичности.
ми, что наблюдаются при длительном посте. Сре-
Лактат способен подавлять липолиз [48]. Уве-
ди вариантов КД встречаются классическая КД,
личенный уровень липолиза в результате умень-
диета, обогащенная триглицеридами средней
шенного уровня лактата при КД вместе с поступ-
длины, модифицированная диета Аткинса и дие-
лением с пищей специфических липидов и жир-
та с низким индексом гликемии [41-45].
ных кислот может оказывать определенные
Заметим, что повышенное содержание кето-
изменения в мембранах, способствуя механиз-
новых тел в организме не обязательно связано с
мам борьбы против контаминации липидного
диабетическим кетоацидозом - несовместимым с
бислоя. Важную роль при этом может играть
жизнью состоянием. Концентрация кетоновых
ограниченная доступность глутамата. Снижение
тел в крови повышается также в результате фи-
уровня глутамата приводит к увеличению абсорб-
зиологического ответа на эпизоды кратковремен-
ции цистеина, и это вызывает повышение уровня
ного или умеренного голодания. Целью КД явля-
глутатиона с соответствующим удалением ненуж-
ется достижение кетоза, избегая кетоацидоз. Так,
ных амфифильных веществ, которые могут за-
в состоянии кетоза, индуцируемого КД, уровни
грязнять мембрану нейронов. Таким образом,
глюкозы, инсулина и pH поддерживаются близ-
при повышенной защите от окислительного по-
кими к нормальным значениям, даже если кон-
вреждения, обеспечиваемой глутатионом, клетки
центрация кетоновых тел в крови почти на два
могут избегать эффекта контаминации мембран
порядка превышает нормальные значения, но все
при эпилептогенезе [49].
же остается в три раза ниже значений, характер-
Многие данные указывают на то, что КД про-
ных для диабетического кетоацидоза (при кото-
являет свое противосудорожное действие посред-
ром уровень глюкозы в крови увеличивается в три
ством синергизма, включающего несколько ме-
раза, а концентрация инсулина падает почти до
ханизмов. Таким образом, противоэпилептиче-
нуля и pH опускается ниже 7.3, что несовместимо
ский эффект КД может найти объяснение вне
с жизнью [46]).
традиционного подхода к действиям на синапти-
При применении КД у 50-60% пациентов, ре-
ческом уровне, и многие из ее эффектов могут
быть проанализированы с точки зрения гипотезы
зистентных к нескольким противосудорожным
о мембранной контаминации. В контексте гипо-
лекарствам, уменьшается по крайней мере в два
тезы Альтрупа мы сосредоточимся далее на следу-
раза частота припадков и примерно 20% пациен-
ющих трех аспектах КД:
тов достигают полного подавления судорог. Кро-
ме того, кетогенная диета способна улучшить са-
1) противосудорожный эффект кетоновых тел;
мочувствие и когнитивные способности [47].
2) связь глутатиона с противоэпилептически-
Доказательства эффективности КД ставят серьез-
ми эффектами КД;
ные проблемы перед теоретиками-эпилептолога-
3) значение полиненасыщенных жирных кис-
ми. Далее мы обсудим возможные механизмы
лот (ПНЖК) и холестерина в действии КД.
КД, рассматривая их возможную связь с гипоте-
Противосудорожное действие кетоновых тел.
зой Альтрупа о мембранной контаминации.
Довольно давно была предложена гипотеза о том,
Метаболизм в условиях кетогенной диеты. При
что кетоновые тела могут действовать непосред-
длительном голодании или при сахарном диабе-
ственно как фармакологические агенты. Х. Кейт
те, когда клетка не имеет доступа к глюкозе, орга-
в 1933 г. сообщил, что приступы, вызванные туй-
БИОФИЗИКА том 66
№ 6
2021
1128
НЕЧИПУРЕНКО и др.
оном у кроликов, были купированы предвари-
рода в изолированных митохондриях. Это указы-
тельной обработкой ацетоацетатом [50]. Позже
вает на эффект КД посредством активации
это было подтверждено на модели мышей, чув-
антиоксидантных механизмов жирными кисло-
ствительным к аудиогенным припадкам [51]. Так-
тами и кетонами. Также было показано, что аце-
же есть раннее сообщение о противосудорожном
тон способен увеличивать текучесть липидной
действии ацетона [52]. Авторы работы [53] сооб-
мембраны [62], действуя в противоположном на-
щили о дозозависимом подавлении судорожных
правлении, чем про-судорожные вещества, и это
приступов внутрибрюшинным введением ацето-
объясняет его антиэпилептический эффект в
на, в четырех отдельных моделях припадков и
контексте гипотезы Альтрупа. В целом следует
эпилепсии на животных. В поддержку идеи о том,
подытожить, что среди множества возможных
что эффект ацетона является одним из основных
механизмов противосудорожного действия кето-
механизмов действия КД, можно привести тот
новых тел, их влияние на вязкость мембраны и
факт, что пациенты с эпилепсией, которые поло-
соответствующая стабилизация липидного бис-
жительно реагировали на КД, демонстрировали
лоя может носить фундаментальное значение.
высокие уровни ацетона в головном мозге (выше
Глутатион и механизм действия кетогенной дие-
1 мМ) [54]. Этот результат привел к тому, что ав-
ты. Снижение уровня глутамата при сниженном
торы работы [53] выдвинули идею о том, что аце-
потреблении глюкозы приводит к увеличению
тон является причинным фактором противосудо-
абсорбции цистеина, и это вызывает повышение
рожных эффектов КД.
уровня глутатиона с соответствующим удалением
Что касается возможных молекулярных мише-
ненужных амфифильных веществ, которые могут
ней для кетоновых тел, то было обнаружено, что
загрязнять мембрану нейронов. Аналогично, из-
VGLUT2 (переносчик глутамата в синаптические
быток глутамата в синапсах, высвобождающийся
пузырьки) ингибируется ацетоацетатом [55] в
при судорогах, препятствует абсорбции цистеи-
концентрациях, которые ожидаются при КД.
на, необходимого компонента глутатиона. Глута-
Этот эффект может противодействовать эксайто-
тион-трипептидный тиол обнаружен практиче-
токсичности, связанной с глутаматом. В то же
ски во всех клетках, он участвует в окислительно-
время кетоновые тела, по-видимому, не изменя-
восстановительных реакциях, и защищает клетку
ют синаптическую передачу на уровне гиппокам-
от активных форм кислорода [24-26]. Глутатион
па, что находится в полном соответствии с выше-
путем прямой конъюгации выводит токсины и
сказанным и наблюдениями Альтрупа [7, 15]. Так,
многие чужеродные и токсические соединения. В
авторы работы [56], используя методы клеточной
частности, за счет устранения свободных радика-
электрофизиологии, обнаружили, что миллимо-
лов глутатионом стабилизируется структура мем-
лярные концентрации бета-гидроксибутирата и
браны [33].
ацетоацетата не влияют на: 1) возбуждающие
Роль глутатиона при эпилепсии хорошо из-
постсинаптические потенциалы и популяцион-
вестна. При помощи модели пилокарпиновых су-
ные разряды в пирамидных нейронах CA1 после
дорог на крысах обнаружено, что концентрации
стимуляции коллатералов Шаффера; 2) спонтан-
глутатиона при судорогах уменьшились на 54 и
ную эпилептиформную активность в модели при-
58% в стриатуме и лобной коре соответственно
ступа гиппокампально-энторинальной коры; 3)
[63]. В модельных экспериментах на животных
ионные токи, вызываемые глутаматом, каинатом
судороги связаны с производством большого ко-
и гамма-аминомасляной кислотой в нейронах
личества активных форм кислорода, которые
гиппокампа.
приводят к окислительному повреждению белков
В частности, был идентифицирован рецептор
и перекисному окислению липидов и могут вли-
β-гидроксибутирата GPR109a (HCAR2), который
ять на активность антиоксидантных ферментов
активируется у мышей с эпилепсией mutUNG1
супероксиддисмутазы и глутатионпероксидазы.
при КД, что предполагает избирательное влияние
Следовательно, уровни глутатиона в головном
КД на механизмы рецепторов кетоновых тел
мозге снижаются. Так, в работе [64] было обнару-
[57, 58].
жено значительное снижение примерно на 35%
среднего отношения глутатион/вода даже в обла-
Другие исследования показывают, что кетоно-
стях мозга, удаленных от эпилептогенного очага
вые тела также могут действовать как нейропро-
[65]. При этом ожидается, что глутатион способ-
текторные средства [59, 60]. Так, авторы работы
ствует снижению уровня контаминации мембра-
[61] показали, что бета-гидроксибутират и ацето-
ны, приводящий к развитию эпилепсии [66].
ацетат (в миллимолярных концентрациях)
предотвращают гипервозбудимость, вызванную
Полиненасыщенные жирные кислоты и кетоген-
перекисью водорода, в острых неокортикальных
ная диета. Кетогенная диета усиливает мобилиза-
срезах головного мозга и диссоциированных ней-
цию ПНЖК из жировой ткани в печень и мозг
ронах, и что кетоновые тела способны значитель-
[57, 67]. Одна из разновидностей этой диеты -
но снижать образование активных форм кисло-
КД, обогащенная среднецепочечными триглице-
БИОФИЗИКА том 66
№ 6
2021
ГИПОТЕЗА КОНТАМИНАЦИИ МЕМБРАН НЕЙРОНОВ
1129
ридами, и является основным методом лечения
ной контаминации может объяснить этот меха-
не поддающейся лекарственному лечению эпи-
низм, в этом и состоит основной тезис нашей ра-
лепсии. Было обнаружено, что на пентилентет-
боты.
разольной модели судорог в срезах гиппокампа
Причины контаминации мембран нервных
крыс выявляется специфичная для точки ветвле-
клеток при эпилептогенезе могут быть самыми
ния активность, которая сравнима с антиэпилеп-
разными: некроз мозговой ткани (скажем, в ре-
тическим эффектом вальпроевой кислоты. Это
зультате травмы), выделение токсинов в случае
показывает, что жирные кислоты, присутствую-
эпилепсии, вызванной нейроинфекцией, или вы-
щие в КД, могут прямым образом оказывать про-
деление анфифильных веществ при инфламации
тивосудорожное действие [68].
или эксайтотоксичности.
Другим вероятным механизмом противоэпи-
В связи с возможными биофизическими меха-
лептического эффекта кетогенной диеты может
низмами противосудорожного эффекта кетоген-
быть воздействие холестерина. В 1939 г. авторы
ной диеты можно отметить три главных момента.
работы [69] задавались вопросом, действует ли
1. Роль кетоновых тел в стабилизации бислоя
механизм кетогенной диеты через повышение
может быть связана с их влиянием на такие био-
уровня холестерина. Изучалась кокаин-индуци-
рованная модель эпилепсии у крыс, было обнару-
физические свойства бислоя, как текучесть, а
жено, что внутрибрюшные инъекции холестери-
также с их действием в качестве антиоксидантов.
на приводят к увеличению порога судорог. Было
2. Увеличение активности глутатиона способ-
показано, что кетогенная диета вызывает суще-
ствует деконтаминации мембран и препятствует
ственное и монотонное увеличение уровня холе-
действию окислительных агентов.
стерина на протяжении одного года ее примене-
3. ПНЖК, поступающие с КД, играют важную
ния [70].
роль в стабилизации поврежденных мембран в
В настоящее время известно, что холестерин
эпилептической мозговой ткани.
регулирует текучесть мембран, и его противоэпи-
Не исключается вклад других эффектов в дей-
лептический эффект может быть обусловлен
ствие КД: диета может менять сольватацию липи-
этим. Холестерин, содержащийся в плазматиче-
дов, белков и углеводов, а также ряд межмолеку-
ских мембранах эукариот, модулирует текучесть
лярных взаимодействий на уровне мембраны, ко-
мембран, делая мембрану менее жидкой при бо-
торые могут быть нарушены при эпилепсии и
лее высоких температурах (например, температу-
восстановлены в результате применения КД.
ре тела 37°C) за счет ограничения подвижности
фосфолипидов и более жидкой при более низких
Заметим, что возможна контаминация глиаль-
температурах за счет предотвращения плотной
ных клеток при эпилептогенезе - в этих клетках
упаковки фосфолипидов [71].
не развиваются ПД, но контаминация их мем-
бран может привести к последствиям, усугубляю-
Заметим, что гептанол вызывает уменьшение
щим эпилептический процесс. В частности, к за-
мембранной текучести холестериновых доменов
грязнению мембран нейронов продуктами мета-
в липидном бислое, и этим эффектом можно объ-
болизма или к синхронизации эпилептического
яснить эпилептогенный аффект гептанола [72]. В
очага [15].
этом плане также достойно внимания, что пропо-
фол, который оказывает антиэпилептическое
В целом белковo-липидные взаимодействия
действие [73], способен увеличить текучесть мем-
играют ключевую роль в переходе от пейсмекер-
бран [74]. Таким образом, в отличие от других
ного потенциала к ПД, где принимает участие це-
преобладающих теорий по эпилептогенезу, гипо-
лый набор белков-ионных каналов, ионных на-
теза контаминации мембраны может дать рацио-
сосов, рецепторов и т. д. Так, по мнению Альтру-
нальное объяснение противоэпилептическому
па:
«эпилептическая активность может быть
эффекту кетогенной диеты и другим эксперимен-
результатом контаминации мембран нейронов
тально подтвержденным фактам [68].
анфифильными молекулами...». Включение этих
веществ вызывает рост латерального давления в
мембране, что может приводить к увеличению
ОБСУЖДЕНИЕ
электрического сопротивления мембаны, инги-
бированию потенциало-зависимых калиевых ка-
Антиэпилептическое действие КД для многих
налов, ингибированию разных типов лигандо-за-
специалистов является до сих пор загадкой. Кли-
висимых каналов; в конечном итоге все это
нические и экспериментальные свидетельства
приводит к преобразованию пейсмекерных по-
показывают, что кетогенная диета эффективна
тенциалов в ПД [7].
даже при самых тяжелых формах эпилепсии. Тем
не менее общепринятые теории о механизмах
На рис. 5 представлена схема, объясняющая
эпилептогенеза не способны дать этому факту
противосудорожный эффект КД в соответствии с
объяснение. Мы считаем, что гипотеза мембран-
гипотезой Альтрупа.
БИОФИЗИКА том 66
№ 6
2021
1130
НЕЧИПУРЕНКО и др.
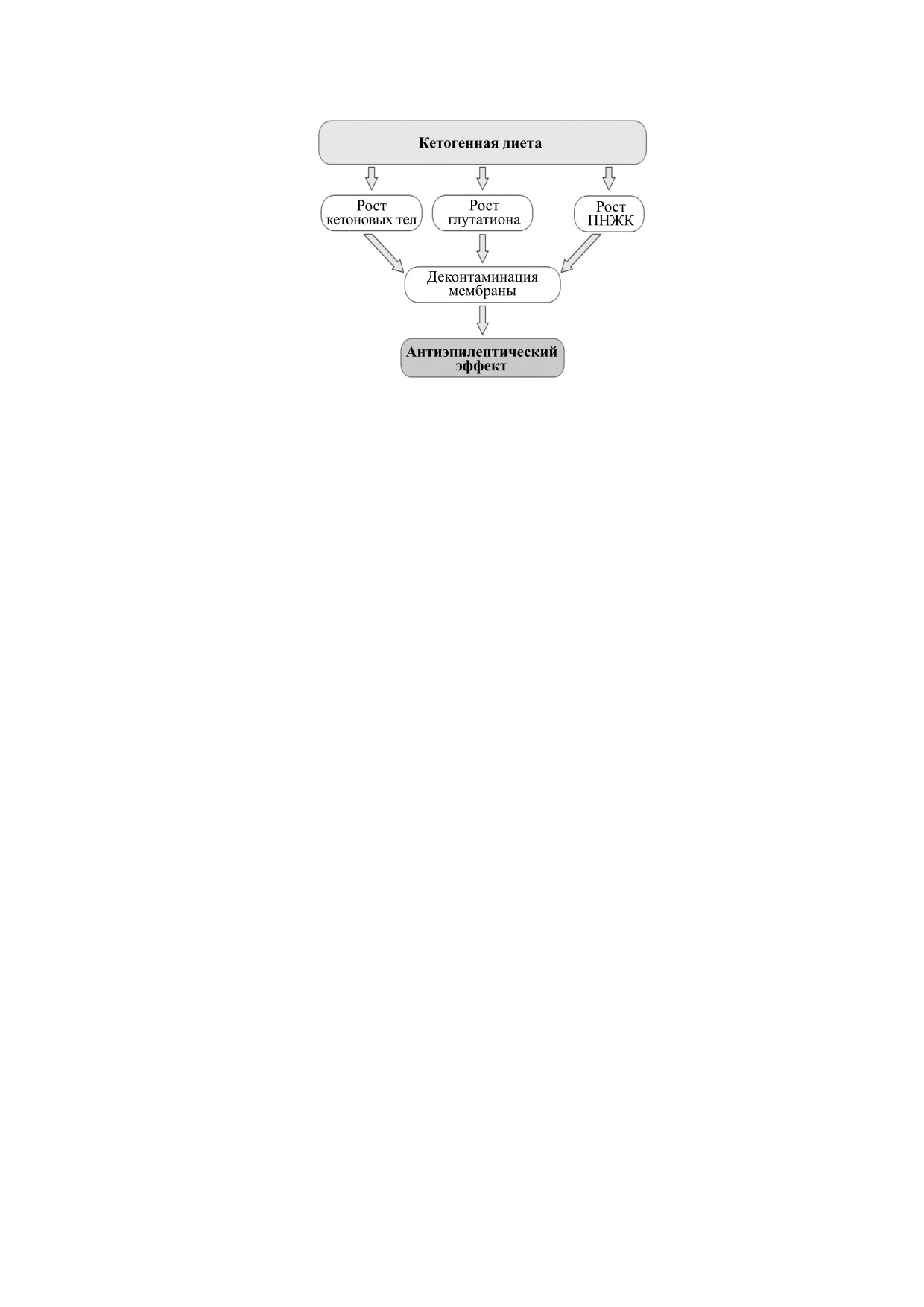
Рис. 5. Схематическое представление механизма действия кетогенной диеты в соответствии с гипотезой контамина-
ции мембраны; ПНЖК - полиненасыщенные жирные кислоты.
ЗАКЛЮЧЕНИЕ
ФИНАНСИРОВАНИЕ РАБОТЫ
Работа выполнена при поддержке Кубинского
В совокупности наши собственные данные на-
научного фонда (FONCI) и Проекта нефармако-
ряду с литературными представляют новые фак-
логической антиэпилептической терапии, коор-
ты, которые поддерживают и уточняют гипотезу
динируемого Кубинским Центром нейронаук, а
контаминации мембраны, предложенную У. Аль-
также при финансовой поддержке Программы
трупом. Кетогенная диета может благоприятно
фундаментальных исследований государствен-
изменить свойства двойного липидного слоя ней-
ных академий наук России на 2013-2020 годы (те-
рональной мембраны при помощи различных ме-
мы № 01201363818 и № 01201363820).
ханизмов:
- Прямого действия ацетона и других кетоно-
КОНФЛИКТ ИНТЕРЕСОВ
вых тел (как противоэпилептического средства).
Авторы заявляют об отсутствии конфликта
- Усиления синтеза холестерина, стабилизи-
интересов.
рующего текучесть мембраны и обладающего
противоэпилептическим действием.
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
- Снижения эксайтотоксичности, связанной с
глутаматом.
Настоящая работа не содержит описания ис-
следований с использованием людей и животных
- Снижения уровня лактата, необходимого
в качестве объектов.
для включения липолиза.
- Повышения уровня глутатиона, который
СПИСОК ЛИТЕРАТУРЫ
уменьшает уровень активных форм кислорода и
захватывает эпилептогенные амфифильные ве-
1. K. M. Fiest, K. M. Sauro, S. Wiebe, et al., Neurology
щества.
88, 296, (2017).
2. J. H. Bautista and F. Luders, Epileptic Disord 2 (1), 65
- Стабилизирующего действия ПНЖК.
(2000).
3. J. Walden, H. Straub, and E. J. Speckmann, Acta Neu-
rol. Scand. (Suppl.) 140, 41 (1992).
БЛАГОДАРНОСТИ
4. S. Engelborghs, R. D’Hooge, and P. P. De Deyn, Acta
Авторы считают своим приятным долгом по-
Neurol. Belg. 100, 201 (2000).
благодарить О. Остроумову, Ю. Ермакова, Д. Се-
5. H. F. Bradford, Prog. Neurobiol. 47, 477, (1995).
мёнова и Л. Яковенко за полезные обсуждения,
И. Лавриненко - за техническую помощь. Мы
6. A. Bragin, C. L Wilson, and J. Engel Jr, Epilepsia 41,
также глубоко благодарны рецензенту за внима-
144 (2000).
тельное прочтение нашей работы, положитель-
7. U. Altrup, M. Häder, J. L. Hernández Cáceres, et al.,
ный отзыв и интересные вопросы.
Brain Res. 1122, 65 (2006).
БИОФИЗИКА том 66
№ 6
2021
ГИПОТЕЗА КОНТАМИНАЦИИ МЕМБРАН НЕЙРОНОВ
1131
8. E. Kandel, Behavioral Biology of Aplysia (New York,
34. E. K. Lund, L. J. Harvey, S. Ladha, et al., Ann. Nutr.
1979).
Metab. 43, 290 (1999).
9. A. L. Hodgkin and A. F. Huxley, J. Physiol. 117, 500
35. A. Hollo, Z. Clemens, A. Kamondi, et al., Epilepsy Be-
(1952).
hav. 24, 131 (2012).
10. M. Wiemann, W. Wittkowski, U. Altrup, et al., Cell
36. K. Ji-Eun and C. Kyung-Ok, Nutrients 11, 1309 (2019).
Tissue Res. 286, 43 (1996).
37. A. Sukhotin, N. Fokina, and T. Ruokolainen, J. Exp.
11. E.-J. Speckmann and H. Caspers, Epilepsia 14, 397
Biol. 220, 1423 (2017).
(1973).
38. G. Guelpa and A. Marie, Rev. Ther. Medico-Chirurgi-
12. M. Segal, J. Neurophysiol. 65, 761 (1991)
cale 78, 8 (1911).
39. J. W. Wheless, Epilepsia 49 (8), 3 (2008).
13. Е. С. Никитин и П. М. Балабан, Журн. высш. нерв.
деятельности им. И.П. Павлова 61 (6), 750 (2011).
40. R. M. Wilder, Mayo Clinic Proc. 2, 307 (1921).
14. Т. П. Норекян, Е. С. Никитин, Н. И. Браваренко
41. Y. Zhang, J. Xu, K. Zhang, et al., Curr. Neuropharma-
и др., Журн. высш. нерв. деятельности им.
col. 16, 66 (2018).
И.П. Павлова 51(6), 717 (2001).
42. D. Boison, Curr. Opin. Neurol. 30, 187 (2017).
15. U. Altrup, M. Hader, and U. Storz, Brain Res. 975, 73
43. A. L. Hartman, X. Zheng, E. Bergbower, et al., Epilep-
(2003).
sia 51, 1395 (2010).
16. R. H. Kramer and R. S. Zucker. J. Physiol. 362 (1), 107
44. D. Y. Kim and J. M. Rho, Curr. Opin. Clin. Nutr. Me-
(1985).
tab. Care 11, 113 (2008).
17. T. Budde, L. Caputi, T. Kanyshkova, et al., J. Neurosci.
45. A. L. Hartman, M. Gasior, E. P. Vining et al., Pediatr.
25, 9871 (2005).
Neurol, 36, 281 (2007)
18. N. I. Kononenko, Comp. Biochem. Physiol. 107A, 323
46. A. Paoli, G. Bosco, E. M. Camporesi, and D. Mangar,
(1994).
Front. Psychol. 6, 27 (2015).
19. N. I. Kononenko, in: Proc. IMACS Symp. on Mathe-
47. Е. А. Куринная и М. А. Барабанова, Журн.
matical Modelling (Vienna, 1994), pp. 315-318.
неврологии и психиатрии 10, 67 (2005).
20. 20. A. O. Komendantov and N. I. Kononenko, Systems
48. S. Sun, H. Li, J. Chen, et al., Physiology 32, 453 (2017).
Analysis Modelling Simulation 18-19, 725 (1995).
49. C. F. Pereira and C. R. de Oliveira, Neurosci. Res. 37,
21. N. M. Berezetskaya, V. N. Kharkyanen, and N. I. Ko-
227 (2000).
nonenko, J. Theor. Biol. 183 (2), 207 (1996).
50. H. M. Keith, Arch. Neurol. Psych. 29, 148 (1933).
22. N. I. Kononenko. Comp. Biochem. Physiol. 106A, 135
51. K. J. Bough and J. M. Rho, Epilepsia 48, 43 (2007).
(1993).
52. H. M. Keith, Am. J. Dis. Children 41 (3) 532 (1931).
23. R. C. García Reyes, B. Sc. Thesis (Faculty of Mathe-
53. S. S. Likhodii, I. Serbanescu, M. A. Cortez, et al., Ann.
matics, Havana University, 2020).
Neurol. 54 (2), 219 (2003).
24. С. В. Стовбун и Л. В. Яковенко, Вестн. МГУ.
54. K. J. Seymour, S. Bluml, J. Sutherling, et al., Magnetic
Физика 6, 101 (2014).
Resonance Materials in Physics, Biology and Medicine
25. М. Н. Резаева, М. Хеншель, Х. Л. Эрнандес и др.,
8 (1), 33 (1999).
Биофизика 25 (1), 41 (1980).
55. N. Juge, J. A. Gray, H. Omote, et al., Neuron 68 (1), 99
26. И. Тасаки, Нервное возбуждение. Макромолекуляр-
(2010).
ный подход (М., Мир, 1971).
56. L. L. Thio, M. Wong, and K. A. Yamada, Neurology 54
27. W. A. Catterall, A. L.Goldin, and S.G. Waxman, Phar-
(2), 325 (2000).
macol. Rev. 57, 397 (2005).
57. M. M. Hasan-Olive, K. H. Lauritzen, M. Ali, et al.,
28. I. Tasaki and K. Iwasa, Japan. J. Physiol. 32 (1), 69
Neurochem. Res. 44 (1), 22 (2019).
(1982).
58. M. A. Rogawski, W. Löscher, and J. M. Rho, Cold
29. В. Г. Артюхов и М. А. Наквасина, Биологические
Spring Harbor Perspectives in Medicine 6 (5), a022780
мембраны: структурная организация, функции,
(2016).
модификация физико-химическими агентами: Учеб.
59. R. L. Veech, B. Chance, Y. Kashiwaya, et al., IUBMB
пособие (Изд-во ВГУ, Воронеж, 2000).
Life 51 (4), 241 (2001).
30. J. R. Godfrey, M. P. Diaz, M. Pincus, et al., Psycho-
60. R. L. Veech, Prostaglandins, Leukotrienes and Essen-
neuroendocrinology 91, 169 (2018).
tial Fatty Acids 70 (3), 309 (2004).
31. B. C. Abbott, A. V. Hill, and J. V. Howarth, Proc. Roy.
61. A. Mosek, H. Natour, M. Y. Neufeld, et al., Seizure 18
Soc. Lond. Ser. B - Biological Sciences 148 (931), 149
(1), 30 (2009).
(1958).
62. G. Dyrda, E. Boniewska-Bernacka, D. Man, et al.
32. J. F. Howe, J. D. Loeser, and W. H. Calvin, Pain 3 (1),
Mol. Biol. Reports 46 (3), 3225 (2019).
25 (1977).
63. H. V. Júnior, M. M. D. F. Fonteles, and R. M. de Fre-
33. N. P. Franks and W. R. Lieb, Nature 333, 662 (1988).
itas, Oxidative Med. Cell. Longevity 2 (3), 130 (2009).
БИОФИЗИКА том 66
№ 6
2021
1132
НЕЧИПУРЕНКО и др.
64. S. G. Mueller, A. H. Trabesinger, P. Boesiger, et al.,
70. A. Mosek, H. Natour, M. Y. Neufeld, et al., PLoS One
Neurology 57 (8), 1422 (2001).
5 (6), e11162-1 (2010).
65. J. B. Schulz, J. Lindenau, J. Seyfried, et al., Eur. J. Bio-
71. M. H. Martinez-Seara, T. Rog, M. Karttunen, et al.,
chem. 267 (16), 4904 (2000).
Epilepsia 45 (Suppl. 7), 199 (2004).
66. C. A. Shaw, in Glutathione in the nervous system, Ed. by
C. A. Shaw (Taylor & Francis, Lond., 1998).
72. E. L. Bastiaanse, H. J. Jongsma, A. van der Laarse,
67. T. Harayama and T. Shimizu, J. Lipid Res. 61 (8), 1150
et al., J. Membrane Biol. 136 (2), 135 (1993)
(2020.)
73. M. T. Baker, Anesthesia & Analgesia 112 (2), 340
68. P. Chang, K. Augustin, K. Boddum, et al., Brain 139
(2011).
(2), 431, (2016).
69. R. B. Aird and C. Gurchot, Arch. Neurol. Psychiatry
74. H. Tsuchiya, Clin. Exp. Pharmacol. Physiol. 28 (4),
42 (3), 491 (1939).
292 (2001).
Hypothesis on Pollution of Neuronal Membranes, Epilepsy and Ketogenic Diet
Yu.D. Nechipurenko*, R.C. Garcia Reyes**, and J.L. Hernandez Caceres**
*Engelhardt Institute of Molecular Biology, Russian Academy of Sciences, ul. Vavilova 32, Moscow, 119991 Russia
**Cuban Neuroscience Center, 15202 Avenida 25, Playa, La Habana, 11600 Cuba
In the light of new data, Altrup's hypothesis on the contamination of neuronal membranes in epilepsy is deal
with. This hypothesis links the paroxysmal depolarization observed during epileptic activity with the pace-
maker potential of a single neuron. The physicochemical characteristics of the membrane, fluidity and con-
tamination affect its ability to conduct an impulseand recharge. Previously proposed methods of epilepsy
treatment based on ketogenic diet and their possible explanations are discussed in connection with the Al-
trup’s hypothesis. Among the possible action mechanisms of the ketogenic diet, the antiepileptic effect of ke-
tone bodies,the role of increased glutathione synthesis and the effect of dietary polyunsaturated fatty acids
and cholesterol are emphasized. These three mechanisms lead to the regulation of the fluidity and other bio-
physical properties of the lipid bilayer, as well as to the de-contamination of the membrane from amphiphilic
impurities, in accordance with Altrup’s hypothesis.
Keywords: epileptic activity, paroxysmal depolarization shifts, pacemaker potential, hypothesis proposed by Altrup,
membrane pollution, lateral pressure, ketogenic diet, glutathione, polyunsaturated fatty acids
БИОФИЗИКА том 66
№ 6
2021