БИОФИЗИКА, 2021, том 66, № 6, с. 1133-1136
БИОФИЗИКА КЛЕТКИ
УДК 577.3
ВЛИЯНИЕ ИОНОВ ЦЕЗИЯ НА ИНГИБИТОРНЫЙ ЭФФЕКТ ПУРИНОВ
В МИОНЕВРАЛЬНОМ СИНАПСЕ
© 2021 г. А.Е. Хайруллин*, В.В. Валиуллин, А.Ю. Теплов, А.Р. Шайхутдинова,
Н.М. Каштанова, С.Н. Гришин
Казанский государственный медицинский университет, 420012, Казань, ул. Бутлерова, 49
*E-mail: khajrulli@yandex.ru
Поступила в редакцию 16.06.2021 г.
После доработки 25.06.2021 г.
Принята к публикации 01.07.2021 г.
Для выявления участия K+-каналов в механизме ингибиторного пресинаптического действия
пуриновых соединений в нервно-мышечном синапсе холоднокровных животных изучали эффекты
АТФ и аденозина на параметры токов концевой пластинки нервно-мышечного синапса лягушек
Rana ridibunda, в том числе и на фоне модулирующих синаптическую передачу ионов цезия.
Наблюдалось различие между эффектами ионов цезия на амплитуду токов концевой пластинки при
его получасовом и часовом сроке действия, что отмечалось нами ранее и в присутствии ионов
бария, являющегося блокатором некоторых подтипов калиевых каналов. В случае часового
воздействия цезия ингибиторный эффект аденозина на амплитуду токов концевой пластинки
оставался стабильным, в то время как угнетающее действие на амплитуду постсинаптических
ответов АТФ при этом уменьшалось в ~2.5 раза. Полученные данные могут свидетельствовать о том,
что пресинаптический угнетающий эффект АТФ на секрецию трансмиттера из нервных окончаний
реализуется через Cs+-чувствительные калиевые каналы нервной терминали.
Ключевые слова: мионевральный синапс, ионы цезия, ингибиторный эффект пуринов, блокаторы
калиевых каналов, АТФ.
DOI: 10.31857/S0006302921060089
калиевые каналы и потенциал-зависимые калие-
Широко известно собственное модуляцион-
вые каналы. В реализации пресинаптического
ное действие по принципу отрицательной обрат-
действия пуринов отмечено участие каналов за-
ной связи котрансмиттера АТФ на квантовый со-
став медиатора в синапсах фазных мышц амфи-
держанного выпрямления и А-типа калиевых ка-
налов - представителей потенциалозависимых
бий [1-3]. Выделяясь из синаптических везикул
калиевых каналов [8, 10].
нервного окончания мотонейрона совместно с
основным медиатором ацетилхолином, АТФ
Ранее нами было показано, что 4-аминопири-
угнетает квантовую секрецию медиатора из пре-
дин, блокатор калиевых каналов А-типа, суще-
синаптических окончаний мотонейронов, дей-
ственно снижает ингибиторный эффект АТФ на
ствуя на P2-рецепторы, в то время как конечный
секрецию медиатора [10]. Это может означать,
продукт его распада - аденозин - реализует свое
что калиевые каналы А-типа участвуют в ингиби-
действие через аденозиновые рецепторы [4]. Из-
торном эффекте АТФ на экзоцитоз медиатора в
вестно, что пурины - это ингибиторы синаптиче-
синаптическую щель. Однако, несмотря на то что
ской передачи [5], а такое действие можно оказы-
4-аминопиридин считается специфическим бло-
вать, либо блокируя кальциевые, либо активируя
катором А-типа калиевых каналов [11], во многих
калиевые ионные каналы [6].
случаях обнаруживается его блокирующая спо-
собность и на калиевые каналы задержанного вы-
Многочисленными исследованиями показа-
прямления, что объясняется близкими структур-
но, что калиевые каналы обнаруживают наиболь-
ными, кинетическими и фармакологическими
шее разнообразие подтипов по сравнению с дру-
свойствами этих подтипов ионных каналов [8].
гими ионными каналами [7-9]; так, различают
кальций-зависимые калиевые каналы, калиевые
Для выявления конкретного подтипа калие-
каналы внутреннего выпрямления, двупоровые вого канала, предположительно участвующего в
пресинаптическом ингибиторном действии
Сокращениe: ТКП - токи концевой пластинки.
АТФ, мы провели эксперименты на нервно-мы-
1133
1134
ХАЙРУЛЛИН и др.
шечном препарате озерной лягушки в цезиевой
среде, учитывая, что ионы цезия блокируют кана-
лы задержанного выпрямления, но не калиевые
каналы А-типа [12].
МЕТОДЫ
Эксперименты проводили на изолированном
нервно-мышечном препарате
«седалищный
нерв - портняжная мышца» лягушек Rana ridib-
unda [6]. В течение эксперимента нервно-мышеч-
ный препарат непрерывно перфузировали рас-
твором Рингера для холоднокровных животных
следующего состава (в мМ/л): NaCl - 115.0, KCl -
2.5, CaCl2 - 1.8, NaHCO3 - 11.0, pH раствора под-
держивали на уровне 7.3.
Для непрямого раздражения препарата нерв
располагали на серебряных электродах в герме-
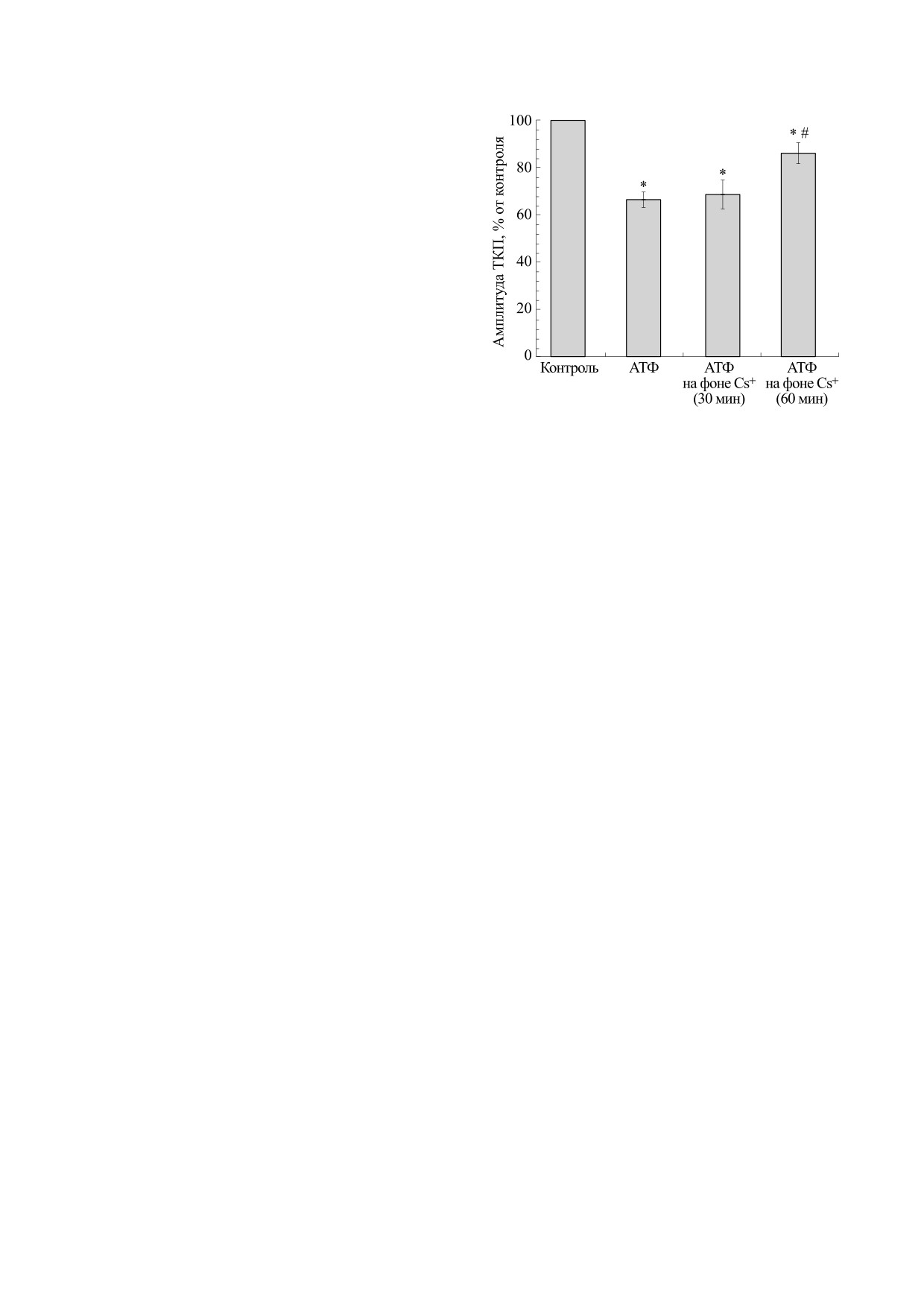
Эффекты АТФ в концентрации
100 мкМ на
тичной увлажненной камере, изолированной от
амплитуду ТКП в контроле и на фоне получасового и
ванночки, в которую помещали мышцу. Для ре-
часового действия ионов цезия; * - р < 0.05 от
гистрации токов концевой пластинки (ТКП)
контроля; # - р < 0.05 от базового действия АТФ.
нерв раздражали прямоугольными импульсами
сверхпороговой амплитуды длительностью 0.2-
0.4 мс с частотой 0.03 Гц [10, 13].
концентрации было аналогичным. При этом, как
и при действии аденозина, в случае добавления
Вызванную квантовую секрецию оценивали
АТФ падение амплитуды ТКП было обратимым,
по параметрам вызванных токов концевой пла-
что было обнаружено при отмывке образцов фи-
стинки [6]. Для отведения ТКП потенциальный и
зиологическим раствором Рингера.
токовый микроэлектроды фокусировали и вводи-
Цезий в концентрации 2 мМ к 30-й минуте
ли в область концевой пластинки мышечного во-
своего действия вызывал повышение амплитуды
локна. Накопление и усреднение треков ТКП
ТКП на 44.3 ± 9.2% (n = 8, р < 0.05) по сравнению
производили с помощью персонального компью-
с контролем, и этот эффект затем сменялся почти
тера с периодом опроса 5-20 мкс на точку. Расчет
полным восстановлением исходной амплитуды
параметров ТКП проводили при помощи ориги-
постсинаптических ответов. Так, к 60-й минуте
нального программного обеспечения [10]. Все ве-
действия цезия амплитуда ТКП составила 110.1 ±
щества вводили через систему перфузии со ско-
8.3% (n = 8, р > 0.05) от контроля.
ростью 2 мл/мин. При оценке аддитивного дей-
ствия АТФ и аденозин подавали через 30 и 60 мин
На 30-й минуте после внесения цезия эффек-
после введения ионов цезия.
ты пуринов не были модифицированы. Так, аде-
нозин в этих условиях снижал амплитуду ТКП до
72.0 ± 3.6% (n = 7, р <0.05) и, как это показано на
РЕЗУЛЬТАТЫ
рисунке, АТФ полностью воспроизводил свой
базовый эффект по снижению амплитуды ТКП.
При фиксации потенциала на уровне -40 мВ
средняя амплитуда многоквантовых токов конце-
Через час после действия ионов цезия добавле-
вой пластинки составила 138 ± 25 нА (n = 25). Че-
ние 100 мкМ АТФ приводило лишь к небольшому
рез 15 мин добавления в инкубационную среду
угнетению амплитуды ТКП, почти в 2.5 раза
аденозина в концентрации 100 мкМ наблюдали
меньшему по отношению к базовому эффекту
снижение амплитуды ТКП до 70.5 ± 2.9% (n = 11,
этого пурина. В отличие от АТФ, ингибирующий
р < 0.05) от исходных значений. Эффект аденози-
синаптическую секрецию эффект аденозина и
на на амплитуду токов был обратим, и после от-
после часового выдерживания препарата в Cs+
мывания раствором Рингера амплитуда сигнала
сохранялся в полной мере: 69.8 ± 5.1% (n = 8,
восстанавливалась до исходных величин (n = 25;
р < 0.05) от контроля.
р > 0.05).
Как показано на рисунке, через 15 мин добав-
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
ления в инкубационную среду АТФ в концентра-
ции 100 мкМ наблюдалось снижение амплитуды
В нашем исследовании мы оценивали моду-
ТКП до двух третей от контрольного значения.
лирующее квантовый выброс медиатора дей-
Таким образом, снижение амплитуды ТКП при
ствие АТФ и аденозина на фоне ионов цезия,
действии АТФ и аденозина при одной и той же
блокатора калиевых ионных каналов [12]. Из-
БИОФИЗИКА том 66
№ 6
2021
ВЛИЯНИЕ ИОНОВ ЦЕЗИЯ НА ИНГИБИТОРНЫЙ ЭФФЕКТ
1135
вестно, что помимо достаточно сложных орга-
дован, в том числе, активацией калиевых каналов
нических соединений, таких как ибериотоксин,
задержанного выпрямления.
харибдотоксин и апамин, некоторые подтипы
И, наоборот, отсутствие изменения в ингиби-
калиевых каналов блокируются одно- и двухва-
торном действии аденозина в цезиевой среде го-
лентными катионами - Cs+ и Ва2+ [8, 12]. Ко-
ворит о том, что уже известное почти полное
устранение ингибиторного эффекта этого пурина
нечно, эти катионы не такие высокоспецифич-
под действием 4-аминопиридина [10] связано
ные блокаторы калиевых каналов, как упомяну-
именно с блокадой А-типа калиевых каналов.
тые ранее яды скорпионов и пчел, но, тем не
менее, их использование вносит свой вклад в ис-
Исходя из полученных данных, мы полагаем,
что пресинаптический угнетающий эффект АТФ
следование разнообразных подтипов калиевых
на секрецию трансмиттера из нервных оконча-
ионных каналов [8, 12], а возможные механизмы
их участия в ряде дисфункций мионеврального
ний реализуется, в том числе, через Cs+-чувстви-
перехода не изучены.
тельные калиевые каналы нервной терминали.
При исследовании действия Cs+ на амплитуду
КОНФЛИКТ ИНТЕРЕСОВ
ТКП на различных сроках инкубации обнаруже-
Авторы заявляют об отсутствии конфликта
но, что к 30-й минуте он вызывал повышение ам-
интересов.
плитуды ТКП почти в полтора раза по сравнению
с контролем, а к 60-й минуте амплитуда ТКП вос-
станавливалась до исходной. Подобное различие
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
эффектов кратковременного и долговременного
Проведенные эксперименты полностью соот-
присутствия Cs+ может также получить объясне-
ветствуют действующим национальным и между-
ние медленно развивающимся внутриклеточным
народным нормам в области этики.
действием этих ионов. Аналогичный двухфазный
характер имеет и эффект ионов бария, блокатора
СПИСОК ЛИТЕРАТУРЫ
некоторых подтипов калиевых каналов [7]. Так, в
1. S. N. Grishin and A. U. Ziganshin, Biochemistry
присутствии 2.5 мМ Ва2+ мы наблюдали сначала
(Moscow) Suppl. Ser. A: Membrane and Cell Biology 9
увеличение, а потом снижение амплитуды ТКП
(1), 13 (2015).
2. R. A. Cunha and J. A. Ribeiro, Life Sci. 68 (2), 119
[10]. Для функционирования К+-каналов имеет
(2000).
значение и время взаимодействия с агентами.
3. K. J. Todd and R. Robitaille, Pflugers Arch. 452 (5),
608 (2006).
Ранее были опубликованы данные о частич-
4. G. Burnstock, T. R. Arnett, and I. R. Orriss, Purinergic
ном устранении 4-аминопиридином (блокатором
Signal. 9 (4), 541 (2013).
калиевых каналов А-типа и задержанного вы-
5. A. U. Ziganshin, C. H. V. Hoyle, A. E. Khairullin, and
прямления) пресинаптического ингибиторного
S. N. Grishin, Int. J. Mol. Sci. 21 (17), 6423 (2020).
действия АТФ [10]. Подобное наблюдалось и в
6. С. Н. Гришин и А. У. Зиганшин, Биол. мембраны
30 (4), 243 (2013).
обнаруженном нами эффекте АТФ на фоне ионов
7. N. A. Castle, D. G. Haylett, and D.H. Jenkinson,
цезия, которые из двух упомянутых подтипов ка-
Trends Neurosci. 12 (2), 59 (1989).
налов блокируют калиевые каналы задержанного
8. A. Meir, S. Ginsburg, A. Butkevitch, et al., Physiol.
выпрямления. Все это в совокупности может
Rev. 79 (3), 1019 (1999).
означать, что калиевые каналы задержанного вы-
9. J. G. Nicholls, A. R. Martin, B. G. Wallace, and
прямления участвуют в реализации эффектов
P. A. Fuchs, Sinauer Associates, Inc. 672 (2001).
АТФ. При этом ранее всегда отмечалось [14, 15],
10. S. Grishin, A. Shakirzyanova, A. Giniatullin, et al.,
Eur. J. Neurosci. 21, 1271 (2005).
что 4-аминопиридин, помимо прочих эффектов,
11. G. Russo, S. Masetto, and I. Prigioni, Neuroreport 6
резко усиливает приток ионов Са2+ в двигатель-
(3), 425 (1995).
ное нервное окончание, и частичная отмена ин-
12. S. Zhang, Am. J. Physiol. Heart Circ. Physiol. 290 (3),
гибиторного действия АТФ на фоне этого агента
1038 (2006).
13. E. V. Ponomareva, E. V. Kachaeva, E. G. Altaeva, et al.,
могла быть связанна с известным ослаблением
Biophysics 53 (6), 615 (2008).
ингибиторного эффекта пурина на фоне повы-
14. S. N. Grishin, Biochemistry (Moscow) Suppl. Ser. A:
шенной концентрации кальция [10, 14]. Получен-
Membrane and Cell Biology 10 (2), 99 (2016).
ные нами данные свидетельствуют, что ингиби-
15. B. Katz and R. Miledi, Proc. Roy. Soc. Lond. B. Biol
торный эффект АТФ на амплитуду ТКП опосре-
Sci. 205 (1160), 369 (1979).
БИОФИЗИКА том 66
№ 6
2021
1136
ХАЙРУЛЛИН и др.
The Influence of Cesium Ions on the Inhibitory Effect
of Purines in Neuromuscular Synapse
A.E. Khairullin, V.V. Valiullin, A.Yu. Teplov, A.R. Shaihutdinova,
N.M. Kashtanova, and S.N. Grishin
Kazan State Medical University, ul. Butlerova 49, Kazan, 420012 Russia
To reveal the participation of K+-channels in the mechanism of inhibitory presynaptic action of purine com-
pounds in the neuromuscular synapse of cold-blooded animals, we studied the effects of ATP and adenosine
on the parameters of the currents of the end plate of the neuromuscular synapse in frogs Rana ridibunda, in-
cluding against the background of modulating synaptic transmission of cesium ions. A difference was ob-
served between the effects of cesium ions on the amplitude of the end-plate currents during its half-hour and
hourly duration of action, which we noted earlier in the presence of barium ions, which is a blocker of some
subtypes of potassium channels. In the case of an hourly exposure to cesium, the inhibitory effect of adenos-
ine on the amplitude of end-plate currents remained stable, while the inhibitory effect on the amplitude of
postsynaptic ATP responses decreased by ~2.5 times. The data obtained may indicate that the presynaptic in-
hibitory effect of ATP on the secretion of the transmitter from the nerve endings is realized through the Cs+-
sensitive potassium channels of the nerve terminal.
Keywords: myoneural synapse, cesium ions, inhibitory effect of purines, potassium channel blockers, ATP
БИОФИЗИКА том 66
№ 6
2021