БИОФИЗИКА, 2021, том 66, № 6, с. 1144-1163
БИОФИЗИКА КЛЕТКИ
УДК 595.76, 51.76, 532.22
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО
КРИЗИСА БЫСТРО РАСТУЩЕЙ ПОПУЛЯЦИИ
© 2021 г. А.Ю. Пеpеваpюxа
Санкт-Петербургский Федеральный исследовательский центр РАН, 199178, Cанкт-Петеpбуpг, 14-я линия, 39
E-mail: temp_elf@mail.ru
Поступила в редакцию 14.04.2021 г.
После доработки 23.04.2021 г.
Принята к публикации 26.04.2021 г.
Статья посвящена моделированию кризисного и порогового развития популяционного процесса
при образовании новой популяции в конкурентной среде. В процессе расселения резкое включение
факторов биотического противодействия до момента исчерпания ресурсов для дальнейшего роста
численности способно вызвать глубокий популяционный кризис. Многие популяции проходили
через этап «бутылочного горлышка», включая человеческие в момент неолитического краха. Все-
ленцы с высокой репродуктивной активностью нередко агрессивно воздействуют на биосистемы.
Появление эффективной конкуренции может не только вызвать классические циклические флук-
туации, но и привести к полному вымиранию популяции после ряда высоких пиков численности.
Классическими примерами ситуаций индуцированного кризиса являются два альтернативных сце-
нария - опыты Г. Гаузе с интродукцией инфузории-хищника и гибелью двух видов и эксперименты
с внесением бактериофагов в колонии активно делящихся бактерий, но обладающих динамически
адаптирующимся противовирусным механизмом. Нами предложены модификации модели для ак-
туальных сценариев кризисных эффектов в популяционной динамике. В уравнениях с запаздыва-
нием учтен пороговый уровень для воздействия биотической среды на размножение инвазивного
вида и агрегированный характер запаздывающей регуляции с двумя временными факторами. Вы-
числительные сценарии описали как эффект завершения процесса после повторного пика, так и
успешного прохождения популяционного кризиса за счет быстрой адаптации. Глубокие кризисные
явления свойственны динамике локальных популяций при столкновении с новыми для них виру-
сами.
Ключевые слова: сценарии инвазий, моделирование экстремальной динамики популяций, биотическое со-
противление, пороговая регуляция с запаздыванием, исчезновение видов, неолитический крах, биологи-
ческий контроль вредителей, иммунный ответ, CRISPR-Cas9.
DOI: 10.31857/S0006302921060107
В данной работе мы продолжаем совершен-
чем люди на Юге Европы, где доля смертельных
ствование методов математической биологии для
исходов очень высока. Полностью не выяснен
анализа волатильных и стремительных переход-
фактор инфекционного потенциала переболев-
ных процессов. Большинство изложенных в
ших COVID пациентов, но с длительно положи-
предыдущей работе [1] идей о моделировании
тельными тестами ПЦР, взятыми после выздо-
сценариев распространения вирусной инфекции
ровления. В объяснении важных свойств динами-
ки и перспектив современной эпидемической
получили в настоящее время подтверждение в ис-
ситуации успешно участвуют российские специа-
следованиях. Многие неочевидные факторы, вы-
листы, опубликовавшие ряд интересных работ
деленные нами как значимые на момент написа-
[2-4].
ния статьи [1] в начале пандемии в форме гипо-
Целью данной работы является математиче-
тез, были установлены при детальном изучении
ское описание сценариев популяционного кри-
новой болезни или обоснованы эксперименталь-
зиса, который способен спонтанно последовать
но к апрелю 2021 г. Ответы на возникшие в начале
за фазой быстрого роста еще до момента заполне-
пандемии некоторые другие вопросы остаются
ния видом доступной экологической ниши. Ана-
неясными. Пока не получено достоверного объ-
лиз вариабельности явления актуален для прак-
яснения, например, почему жители Дальнего Во-
тических задач - обоснования методов биологи-
стока оказались в среднем устойчивее к вирусу,
ческого контроля опасных вселенцев.
1144
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО КРИЗИСА
1145
Рис. 1. Динамика общей смертности в Англии с января 2020 г. по февраль 2021 г. (данные правительственного отчета,
ДИНАМИЧЕСКИЕ ОСОБЕННОСТИ
те число «COVID superspreaders» оказывается в
СОВРЕМЕННОЙ ПАНДЕМИИ
локальном случае на раннем этапе важнее, чем
среднее по популяции значение R0 базового ре-
В апреле 2020 г. мы отметили принципы ди-
вергенции протекания эпидемического процесса
продуктивного числа передачи вируса. Динамика
COVID-19 и неизбежность его вариабельности,
первых этапов региональных эпидемий в Европе
основываясь на аналогиях. Особенностью совре-
сильно различалась в начале 2020 г. во многом из-
менной пандемии стала роль суперраспространи-
за наличия активных групп таких распространи-
телей в запуске эпидемического процесса y
телей - трудно предсказуемый для прогнозирова-
COVID, что подтверждалось многократно в раз-
ния сценариев эффект.
ных странах. Проблема для моделирования здесь
Вторая фаза роста заболеваемости COIVID зи-
состоит в том, что многие больные не заражают
мой 2021 г. в эпидемическом цикле в Великобри-
совсем никого, но отдельные заражают очень
тании (предположительно виновен «британский
многих. Эффект от действия массовых разносчи-
штамм») стала существенно больше первой по
ков известен давно, описан на примере начала
числу случаев, но отмечено заметное снижение
эпидемии ВИЧ в 1980 г. ВИЧ проник в США еще
статистики смертности по сравнению с весной
в 1950-е гг., но одного гиперактивного распро-
2020 г. (рис. 1). Это значимое явление для прогно-
странителя оказалось достаточно, чтобы в 1979-
за характера эпидемии.
1980 гг. стремительно ускорить динамику эпиде-
мического процесса. Установленные в работе [5]
Затухающая динамика волн эпидемии оказы-
актуальные отличия состоят том, что суперрас-
вается качественно схожей с импульсной «пило-
пространители характерны именно для корона-
образной» вспышкой насекомых вредителей ле-
вируса в большей степени, чем для сезонного
са, для которой в работе [6] нами было предложе-
гриппа, пик вирусной нагрузки и пик симптома-
но сложно настраиваемое уравнение с четырьмя
тики COVID могут чаще не совпадать. В результа-
включениями разных значений запаздывания:
dN
N(t
− τ
=
rN(t
−τ)exp(−bN(t
−τ))
−
q
,τ>τ
1
(1)
dt
A
-
N(t
-τ
1
)
Уравнение (1) генерирует затухающие ряды
вспышки, и в Иране, Нью-Йорке, Великобрита-
пиков, где второй пик в «серии волн» способен
нии, ЮАР это отличие действительно подтверди-
стать больше первого в зависимости от N(0). Ра-
лось. Гораздо реже в регионах локально отмечал-
нее нами было отмечено, что осцилляционный
ся на графиках вместо двух «волн» вариант доста-
характер эпидемии COVID-19 следует отличать от
точно длительного эпидемического
«плато».
сценария повторного запуска Λ-образной
Единичная эпидемическая быстрая Λ-образная
БИОФИЗИКА том 66
№ 6
2021
1146
ПЕPЕВАPЮXА
Рис. 2. Осциллирующая динамика эпидемии COVID в Южной Африке с марта 2020 г. по март 2021 г. (данные New-
York Times).
вспышка, как с гриппом H1N1 в 1918 г. в Фила-
за время между пиками не успеет потерять свою
дельфии, отмечена в
2020 г. в Шри-Ланке.
эффективность и устойчивость накопится в по-
Эволюция COVID оказалась сложнее гриппа «ис-
пуляции, значит, и амплитуда колебаний заболе-
панки».
ваемости в ЮАР стабилизируется. В работе [7]
было подтверждено, что Т-клеточная память и
Хрестоматийный пример двухпиковой осцил-
эффекторная функция CD8-клеток способна ра-
лирующей эпидемии наблюдался в 2020 г. в ЮАР
(рис. 2). Появление нового штамма не поменяло
ботать против множества эпитопов коронавиру-
динамический режим колебаний, поэтому третий
са. Более того, перекрестно-реактивные Т-клетки
найдены в исследовании [7] у людей без подтвер-
пик заболеваемости можно прогнозировать на
жденной инфекции SARS-CoV-2.
уровне, не превосходящем заболеваемость перво-
го пика. Все масштабные пандемии даже измен-
Безусловно, не все появляющиеся новые вари-
чивых ОРВИ вирусов когда-то завершаются. За
ации Spike-белка вируса будут способны вызы-
счет Т-лимфоцитов CD4 и CD8 иммунный ответ
вать скачкообразные режимы всплесков тяжелых
случаев СOVID. У переболевших без иммуносу-
прессии с вероятностью, большей 0.5, их Т-клет-
ки смогут распознавать эпитопы у новых штам-
мов, как B.1.351 из ЮАР. Важно выявлять ситуа-
ции, когда резко меняется качественный ход
эпидемии, и анализировать лучше статистику
случаев госпитализации.
Гораздо опаснее сценарий эпидемии с интер-
валом экстремального роста заболеваемости в
марте 2021 г. мы видим в Бразилии. Обратим вни-
мание на быстроту, с которой фаза затухания с
уменьшением амплитуды колебаний с пиками в
августе и в январе (как и в ЮАР), вдруг резко в
марте сменилась другим режимом - Λ-образной
вспышкой с экспоненциальным ростом. Новый
всплеск, очевидно, превосходит первоначальную
скорость распространения болезни. На рис. 3
представлена динамика волн госпитализаций с
СOVID в южном бразильском штате Риу-Гранди-
ду-Сул. С запаздыванием на две недели в середи-
Рис. 3. Динамика экстремального роста случаев гос-
не марта 2021 г. аналогичный эпидемический
питализаций c COVID-19 в штате Риу-Гранди-ду-Сул
в начале марта 2021 г. (по данным отчета на сайте
всплеск начался после затишья в городах
Чили. Вероятно, бразильский штамм B.1.1.248 в
БИОФИЗИКА том 66
№ 6
2021
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО КРИЗИСА
1147
Рис. 4. Статистика динамики заболеваемости COVID-19 обобщенная по всем штатам Мексики (данные с сайта New-
2021/22 г. будет наиболее проблемным в так назы-
Таким образом, интервал менее двух месяцев
ваемой «эпохе войны вакцин и штаммов» [8]. Си-
между прошедшим пиком и началом новой фазы
туация в Южной Америке предполагает триггер-
роста заболеваемости - признак появления (на-
ное событие, которое смогло резко изменить ха-
копления нескольких) опасных изменений y ви-
рактеристики эпидемического процесса вируса
руса. В Исландии наблюдается типичная динами-
спустя год после его начала, и это явление требует
ка шестимесячных интервалов между пиками: ап-
внимания исследователей. Интересно выяснение
рель, октябрь, апрель. Для Бразилии или Индии
доли повторных заражений среди случаев марта
неактуальна сезонность вспышек ОРВИ, но это
2021.
важный фактор для регионов севера Европы и,
Сценарий изменения динамики эпидемии на
безусловно, для России.
рис. 3 бифуркационный. Более того, здесь мы ви-
дим признаки скачкообразной потери установив-
Обобщенные данные случаев по странам с вы-
шегося режима. Такая трансформация качествен-
сокой численностью населения (США, Индоне-
ной картины процесса на юге Бразилии подразу-
зия, Индия, Бразилия), но неравномерной плот-
мевает резкое увеличение базового r-параметра
ностью мало пригодны для анализа. Графики
вируса, которое произошло из-за мутации, но не
складываются из непохожих групп локальных
в начале марта, а раньше - это процесс инерци-
сценариев. По обобщенным данным заболевае-
онный. Статистика учета госпитализаций реаги-
мости в США прослеживается динамика эпиде-
рует с задержкой. Модель сценария с таким раз-
мии из трех возрастающих волн, но в Нью-Йорке
рушением образовавшегося цикла после бифур-
и Мичигане динамка качественно иная. В Мекси-
кации Андронова-Хопфа N*(t
< t*; rτ) и
ке улучшали методику учета статистики, но при
появлением вместо цикла при значении r ≥ r* тра-
этом график заболеваемости в Мексике каче-
ектории limt→∞N(t; r*τ) = +∞ получена в работе
ственно не похож по динамике на Нью-Йорк,
[9] в расширенной модификации логистического
ЮАР и Бразилию (рис. 4). Эпидемический про-
уравнения с запаздыванием:
цесс развивался в Мексике сравнительно медлен-
нее сопредельных стран при очень высокой плот-
dN
N(t
−τ)
=
rN t)
1−
(ϒ-
N(t
−τ)),ϒ<
K
(2)
ности населения в столице страны.
(
)
dt
K
Сравнение сценариев подтвердило, что меры
Разрушение цикла в недиссипативном реше-
«локдауна» не способны остановить уже начав-
нии уравнения (2) описывает момент трансфор-
шуюся фазу стремительного роста заболеваемо-
мации процесса из-за резкого увеличения r-пара-
метра, но не варианты его завершения. Сценарий
сти, если губительный для экономики «локдаун»
с бифуркацией фактически непредсказуем и от-
вводят только по факту риска исчерпания боль-
ражает независимое случайное событие. Эпиде-
ничных мест, - пример попытки управления про-
мия гриппа Н1N1 «swine flu» в 2009 г. проходила в
цессом на основе данных, которые отражают его
этих регионах c двумя пиками в июле и в ноябре.
динамику с отставанием.
БИОФИЗИКА том 66
№ 6
2021
1148
ПЕPЕВАPЮXА
ВАРИАТИВНЫЕ ОСОБЕННОСТИ
в локальной популяции. Однако при распростра-
СЦЕНАРИЕВ ИММУННОЙ АКТИВАЦИИ
нении вируса по всем континентам закон боль-
ших чисел будет играть против нас. Вирус начал
Помимо ограничительных мер «локдаунов»,
меняться и путем делеций - потерь участков в его
социальных норм и традиций общения на дина-
большом геноме. Достаточно предсказуемо мута-
мику региональных эпидемий оказывают влия-
ции затрагивают Spike-белок, но отмечено, что у
ние отдельные варианты иммунного ответа, кото-
данного сложного трансмебранного белка есть и
рые у локальных групп иногда действительно
консервативные участки [14]. Автор статьи решил
имеют особенности. Подтвердилось [10], что на-
вакцинироваться, понимая, что отлична от нуля
боры перекрестно-реактивных В-клеток, образо-
вероятность ситуации, когда январе 2022 гг. будет
ванные в организме в результате предыдущих
циркулировать иной штамм, и выработанные по-
контактов с другим эндемическим бета-корона-
сле вакцины IgG антитела могут оказаться недо-
вирусом, встречаются у некоторых пациентов с
статочно аффинными к новому варианту RBD-
сильными симптомами. Такие клетки памяти
домена шипа-мутанта. Однако чем больше людей
(MBC) представляют небольшую, но стабильную
и как можно быстрее в мире станут устойчивыми
часть среди встречающихся исследователям ре-
к вирусу, тем меньше вероятность опасных собы-
пертуаров клеточных рецепторов лимфоцитов, и
тий - появления у вируса новых значимых мута-
данные клетки «не способствуют нейтрализую-
ций. Большинство действий при иммунном отве-
щей реакции антител» согласно работе [10]. Акти-
те в организме не полностью детерминировано и
вация B-клеток иммунной памяти не помогала
вероятность успеха все-таки превосходит. По-
справиться с новым вирусом. В различных регио-
пуляционный «стадный» иммунитет после массо-
нах циркулировал отличающийся состав ранее
вой вакцинации не должен пропадать стреми-
известных четырех коронавирусов. По нашей ги-
тельно при появлении новых штаммов, но
потезе из-за явления «слепого пятна» или «перво-
уменьшаться линейно, по аналогичным вероят-
родного антигенного греха» [11] в иммунном от-
ностным причинам, которые сопутствуют про-
вете это повлияло весной 2020 г. на явные локаль-
цессу презентации антигена. У большой группы
ные различия процентного количества тяжелых
вакцинированных иммунодоминатные пептиды
случаев COVID и доли смертельных исходов.
и B- и Т-клеток будут различаться, образуя слож-
Свойства эффекта таковы, что при повторных
ное распределение. Иммунитет «весь и сразу» эф-
волнах эпидемии в 2021 г. этот эффект перестанет
фективности не потеряет. Многие авторы обос-
быть значим, но позднее может вновь активиро-
новали важность изучения клеточного иммунно-
ваться из-за накопления мутаций в РНК вируса.
го ответа от Т-киллеров с рецептором CD8 на
Возникает «антителозависимое усиление инфек-
спектр антигенов - участков консервативных
ции», когда комплекс «вирус-антитело» непро-
структурных вирусных белков и нуклеопротеи-
чен и вирус избегает уничтожения в эндосомах
нов, но такой ответ формируется с непостоянным
лейкоцитов и заражает их [12]. ADE в 2020 г. мог
запаздыванием.
быть локальным и эпизодическим, но в перспек-
тиве эффект ADE способен стать массовым и
Особая роль NK-лимфоцитов натуральных
циклическим региональным эффектом для ранее
киллеров в иммунном ответе действительно ока-
переболевших COVID. Клетки иммунной памяти
залось высока, что следовало согласно гипотезе
MBC, ранее реагировавшие на неактуальный те-
задержки в сценарии активации Т-клеток. У па-
перь эпитоп, через сезон заболеваемости ОРВИ
циентов с серьезными воспалительными ослож-
становятся уже помехой - с субоптимальными
нениями было отмечено снижение эффекторных
антителами при столкновении с немного изме-
функций NK-клеток [15]. Теперь, сопоставив
нившими антигеном. Примером служит ситуация
между собой новые результаты нескольких иссле-
с неудачей массовой вакцинации от вируса Денге
дований, мы можем обосновать причину важно-
из-за его четырех разных серотипов, после вакци-
сти этой группы клеток. Каждое звено иммуните-
нации некоторые привитые переболели лихорад-
та нацелено решать свой круг задач, но допуска-
кой Денге серьезнее, чем в среднем в контроль-
ется дублирование их функций. NK-клетки
ной группе [13]. Эффект ADE для заразившихся
обеспечивают антителозависимую цитотоксич-
вирусом Денге явление вероятностное, но 10%
ность, а виновниками осложнений оказались
неудачных случаев после прививки уже серьезная
альвеолярные макрофаги. Антитела IgG
проблема.
против S-белка SARS-CoV-2, не содержащие в
Фактор накопления генетических отличий и
своей Fc-области сахара фукозы («афукозилиро-
появление новых штаммов SARS-CoV-x был оче-
ванные» иммуноглобулины), из-за их повышен-
виден сразу после объявления ВОЗ о пандемии. У
ной связывающей способности с Fc-рецептором
коронавирусов есть механизм, отвечающий за
y макрофагов запускают высвобождение этими
точность репликации их РНК, такой вирус может
клетками уже излишних сигнальных молекул -
оставаться стабильным долгие годы, циркулируя
провоспалительных цитокинов [16]. Из-за поло-
БИОФИЗИКА том 66
№ 6
2021
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО КРИЗИСА
1149
жительной обратной связи это усиливает систем-
процессов при нарушении выравненности сло-
ное воспаление.
жившегося биологического разнообразия. Мы
Наиболее проблемным остается подтвержде-
рассмотрим моделирование нескольких альтер-
нативных сценариев развития инвазионных про-
ние нашей модельной гипотезы об эффекте зави-
цессов y активных вселенцев, но которые сопро-
симости от дозы в сценарии развития инфекци-
онного процесса, когда начальный уровень зара-
вождаются стремительным усилением конку-
жения N(0) сильно определяет быстроту и
рентного противоборства. От борьбы иммунной
качество клеточного иммунного ответа. В вычис-
системы и вируса сценарии при инвазиях отлича-
ются большими возможностями совместной
лительной модели сценария мы можем использо-
адаптации видов, когда вселенец может победить
вать только количественную величину - дозу, но
не относительную оценку ее вирулентности. Пря-
в борьбе за экологическую нишу, стать домини-
мые опыты на людях с высокими дозами невоз-
рующим видом. У вируса нет устойчивой «ниши»,
можны по понятным причинам. Ряд результатов
но только уровень минимального неразрушаю-
щего среду воздействия. Длительное состояние
позволяют более уверенно говорить о факторе
при высокой вирусной нагрузке для организма
порогового уровня той начальной дозы зараже-
ния, которая может вызывать тяжелое течение
хозяина невозможно, что отражено в нашем мо-
заболевания COVID из-за активации каскада
дельном сценарии инфекционного процесса для
воспалительных цитокинов, включая IL-1β, за-
гепатита C из [1]. Безусловно, заражение всех до-
ступных клеток печени означает летальный вари-
пускающих как местное, так и системное воспа-
ант исхода болезни.
ление. В работе [17] описаны эксперименты с
повторным заражением вирусом макак-резусов,
Представим, что инвазивный вид с высокой пло-
отмечено, что антитела могут защищать от зара-
довитостью «r-параметром» активно распространя-
жения SARS-CoV-2 дозозависимым образом, но
ется в новом ареале, на первом этапе это возможно с
только если ранее полученные IgG будут выше
динамикой, близкой к экспоненциальному закону
порогового титра, необходимого для защиты. Не-
роста N(t) = N(0)ert - qN(t), q < r < 1. В некоторый
обходимы экспериментальные работы c оценкой
момент времени t = T вид начинает встречать актив-
качества всего спектра иммунного ответа на раз-
ное противоборство со стороны автохтонного (или
ные дозы коронавируса, подобные опытам, опи-
специально интродуцированного) биотического
санным в работе [18] для разных доз гриппа
окружения. Ситуации, когда вид-вселенец, интен-
H3N2. Разработчики классической вакцины
сивно размножаясь, разрушает свою среду обитания
VLA2001 с инактивированным коронавирусом
полностью, как завезенные на остров Беринга олени
подтвердили выраженную дозозависимую им-
или люди на острове Пасхи, остаются редкими при-
мунную реакцию на их препарат. В контексте за-
мерами. Триггерный фактор выступает как меха-
висимости развития осложненного течения
низм регуляции численности с запаздывающей ре-
COVID от исходной дозы введенного вируса, по
акцией. Известны несколько вариантов для разви-
нашему мнению, недооценены методы усиления
тия подобных событий и интересных реальных
вакцинами иммунного комплекса слизистых
примеров.
оболочек - первого защитного барьера и важного
Человеческие популяции за приделами Афри-
для группы людей с ослабленным Т-клеточным
ки вполне можно считать инвазивными, им при-
иммунитетом и, как следствие, длительным тече-
ходилось отвоевывать свою «нишу». Как устано-
нием COVID.
вили археологи, после проникновения в Европу и
Множество отдельных, иногда разрозненных
активного расселения в северных регионах, посе-
и кажущихся далеких друг от друга результатов
ления Homo sapiens испытали резкий кризис, из-
исследований постепенно складываются общую
вестный как неолитический крах (Neolithic crash)
мозаику - вариантов действия иммунного ответа,
около 6 тыс. лет до н.э. [19]. Затяжной кризис
которые интересно классифицировать и с мате-
продолжался полтысячелетия (рис. 5) [19]. Исто-
матической теоретизацией.
рики дискутируют о причинах и последствиях на-
стигшего наших предков длительного демогра-
БИОЛОГИЧЕСКИЕ ИНВАЗИИ
фического спада.
И АКТИВАЦИЯ ПРОТИВОДЕЙСТВИЯ
После кризиса в неолите образовалось дли-
СРЕДЫ
тельное равновесие, сменившиеся новым трен-
дом повышения численности населения Европы
Рассмотрим математически не менее интерес-
только около 4 тыс. лет до н.э. По масштабам ис-
ную другую проблему экодинамики переходных
торической шкалы пример продолжительного, но
процессов, которой ранее уделяли мало внима-
не самого глубокого кризиса.
ния. Спонтанные вселения стали регулярными и
даже неизбежными явлениями для экологии био-
Подобный тип резких изменений численности
систем. Практически важны и интересны для ма-
в экодинамике называется «boom-and-bust», что
тематического описания экстремальные формы
можно перевести как «взлет и крушение». Мате-
БИОФИЗИКА том 66
№ 6
2021
1150
ПЕPЕВАPЮXА
множения поддерживает популяцию при исто-
щении кормовой базы за счет каннибализма [21],
что как раз позволяет ему вытеснять конкурен-
тов, проходя минимумы численности. Для мно-
гих видов существует минимальный уровень чис-
ленности L, хотя для других видов его нет.
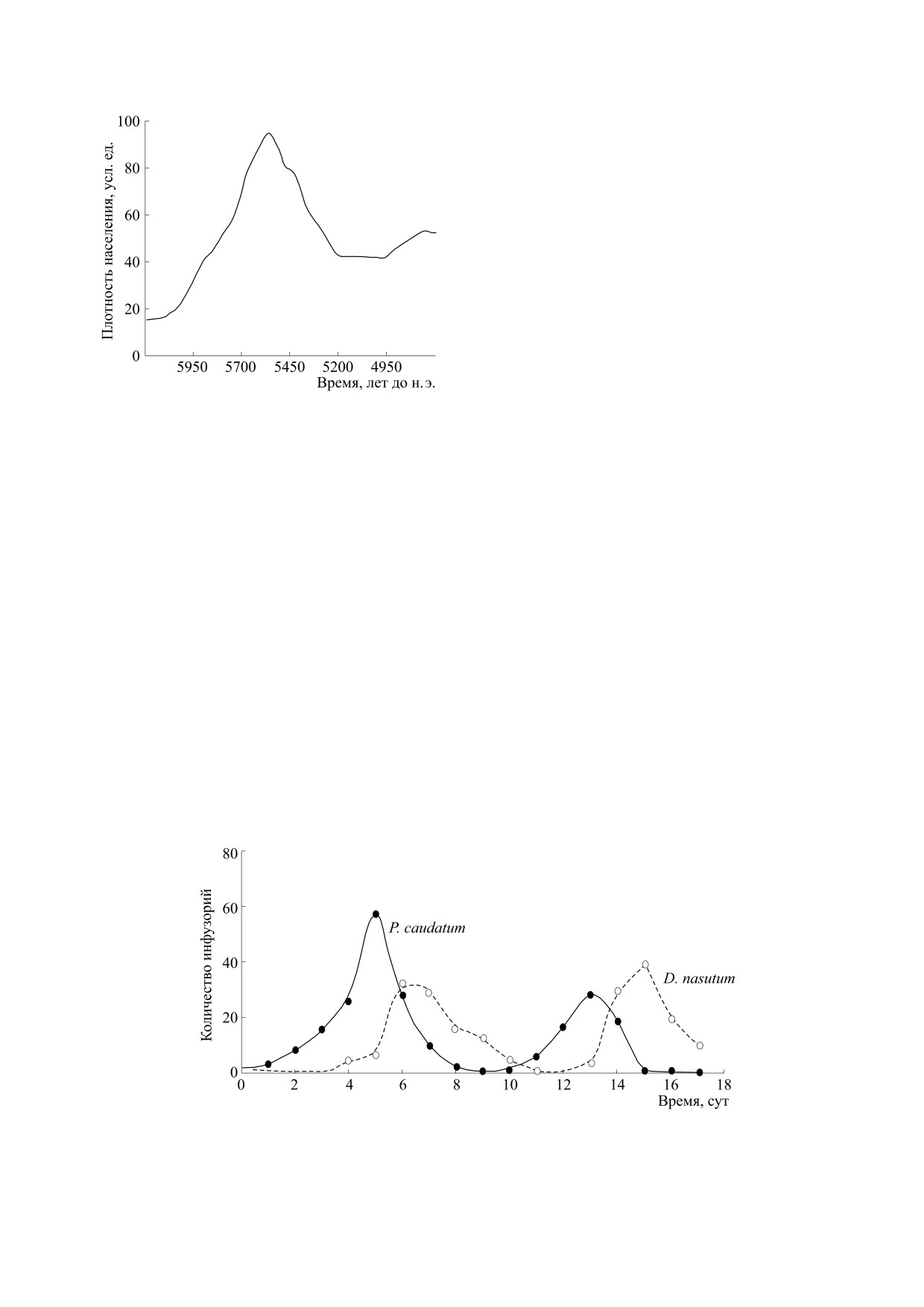
На примере наших неолитических предков
мы видим благополучный сценарий успешного
преодоления затяжного минимума N(t) → L,
L > δ ∈ и низкой эффективности воспроизвод-
ства. Существует много видов, не преодолевших
внезапный кризис, в том числе среди инвазивных
популяций после начала активного освоения но-
вого ареала. Обзор случаев краха популяций при-
веден в работе [22]. Многие из них имели высо-
кий репродуктивный потенциал, как вселенный в
Ставропольском крае американский жук-листоед
Рис. 5. Динамика плотности населения Севера Евро-
Zygoramma suturalis [23].
пы при неолитическом крахе (по работе [19]).
ЭКСПЕРИМЕНТАЛЬНЫЕ НАБЛЮДЕНИЯ
матический сценарий в работе [20] c однократной
ДВУХ ЯВЛЕНИЙ КРИЗИСА
краткой вспышкой - только его частный случай.
Изменения с пиками и кризисами достаточно ха-
В экспериментах получено наглядное подтвер-
рактерны и важны, так как сопровождаются эф-
ждение различных вариантов кризисной динами-
фектом «бутылочного горла». Этот эффект связан
ки. Классические серии экспериментов Г. Гаузе
с генетическими изменениями, так как после
1934 г. c двумя видами инфузорий можно интер-
прохождения «горлышка бутылки» продолжаю-
претировать и как внезапное включение запазды-
щаяся развиваться популяция оказывается по-
вающего сопротивления, но и как лабораторную
томками малой группы. Число вариантов y ал-
модель поведения агрессивного вселенца в новой
лельных генов может резко снижаться, и подоб-
среде. В пробирку с логистически (N(t) → K) рас-
ные явления кризисов могут рассматриваться как
тущей колонией Paramecium caudatum добавля-
факторы эволюции и причины внутривидовой
лись (t = tD) естественные враги, агрессивные
дифференциации. Генетики по мтДНК опреде-
хищники Didinium nasutum. Взаимодействие ин-
лили ряд видов, переживших в прошлом этап ма-
фузорий в опытах без убежищ заканчивалось уни-
лой группы.
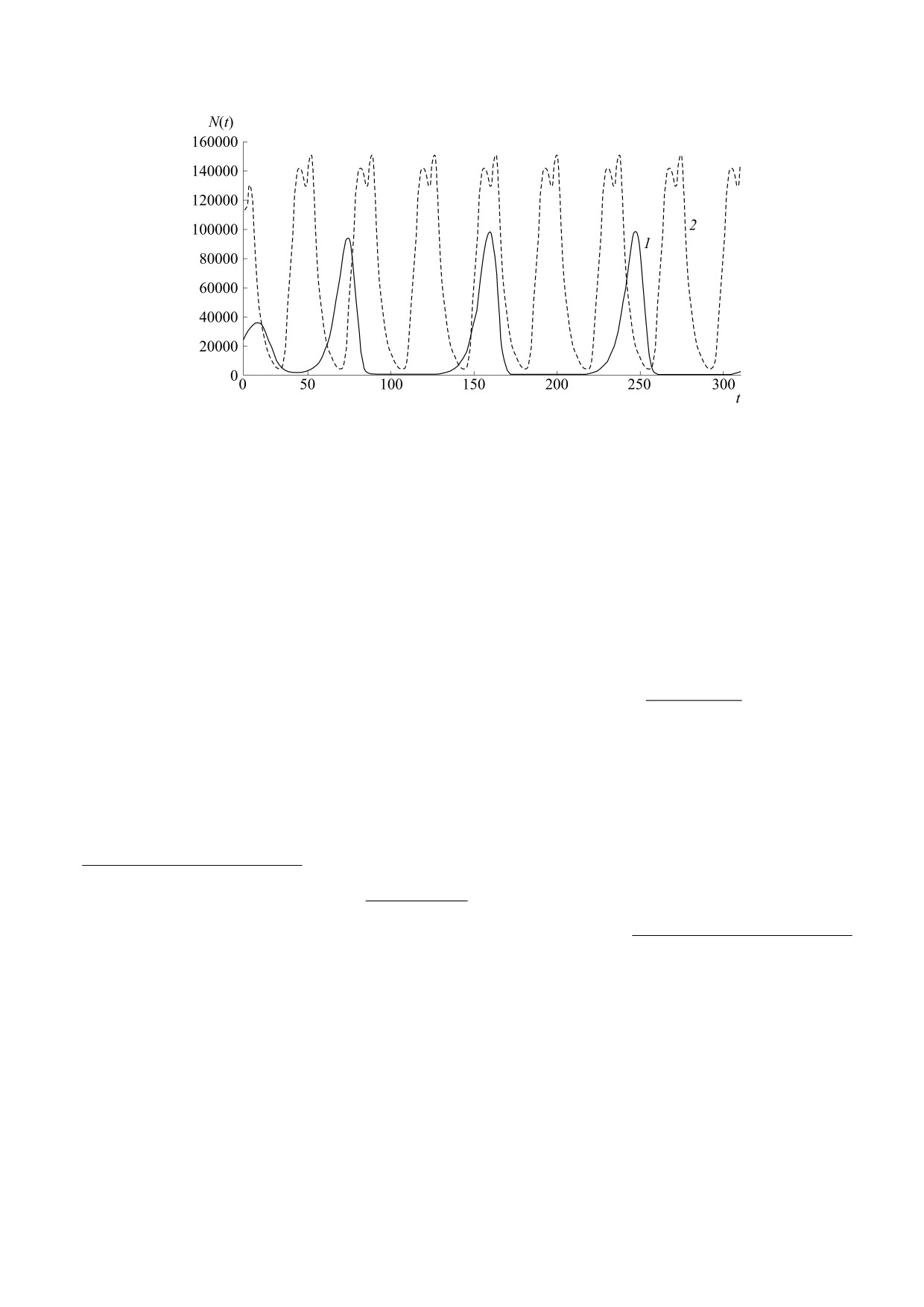
чтожением инфузории-жертвы, и следом после
Для других вселенцев резкие колебания чис-
второго максимума численности погибал лишен-
ленности, наоборот, нормальны и не мешают
ный пищи хищник (рис. 6) [24]. Второй максимум
приспособленности. Так, вредоносный вселенец
у Didinium оказывался выше первого и совпадал с
гребневик Mnemiopsis leidyi после массового раз-
минимумом у жертвы.
Рис. 6. Динамика двух видов инфузорий с гибелью после второго пика в опытах Гаузе [24].
БИОФИЗИКА том 66
№ 6
2021
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО КРИЗИСА
1151
Опыты описаны в некоторых книгах как экс-
развитие в каждом отдельном случае, и мы вы-
периментальное опровержение модели В. Воль-
нуждены снова иметь дело с вероятностными из-
терра и теории об устойчивой цикличности видов
менениями». Так Гаузе предвидел не изученное
в двухуровневом трофическом взаимодействии,
тогда явление, что математически называется
но их трактовка в работах самого Гаузе несколько
стохастическим размыванием сепаратрисы. Не-
отличается. Гаузе писал о неполной обусловлен-
которое возмущение начальных условий при
ности лабораторных опытов в работе [24]: «когда
N(0) ± ε [25] необязательно приведет к иному
имеется большое число особей Paramecium и Di-
асимптотическому положению траектории при
dinium, оказывается возможным по известным
существовании альтернативных аттракторов.
уравнениям предсказать течение процесса борь-
Границу областей притяжения аттракторов вме-
бы за существование. Однако в критические мо-
сто L-порога составит область рассеянного мно-
менты смены одного цикла следующим, когда
жества точек. Данное свойство наблюдается в на-
число особей невелико, различные «случайные
шей модели (3), где H - предпороговая величина
моменты» приобретают большое значение. В ре-
вирусной нагрузки для организма при условии
зультате оказывается невозможным определить
N(0) < H < K для данной инфекции:
dN
K
=
rN t)ln
(H -N(t
−
h)),H <K,h
>τ
(3)
dt
N(t
-τ)
Эффект размывания сепаратрисы не противо-
активность. Отмеченные y Гаузе «случайные мо-
речит опытам и наблюдениям, ведь повышение
менты» связаны с чувствительностью его лабора-
начальной дозы на n + 1 вирион не обязательно
торной системы относительно возмущения ее на-
приводит к летальному исходу. Известно из опы-
чального состояния в моменты t = 0 и в t = tD. Ма-
тов, приведенных в работе [18], что существуют
тематически интересны эксперименты, где
шансы выживания y мышей и при огромной дозе
свободная динамика жертвы не логистическая, а
гриппа. Уравнение (3) - наглядный, но не един-
циклическая. Такие эксперименты можно прове-
ственный пример стохастических свойств y де-
сти, добавив к инфузориям их паразита Holospora
терминированной модели, но вычислительные
undulata.
эксперименты с уравнением (3) не могут описать
Рассматриваемый нами сценарий кризиса в
спонтанное разрушение колебаний y возникшего
активной фазе инвазии часто связан с началом
популяционного цикла.
эпизоотии. Известна другая лабораторная систе-
ма, где взаимная адаптация происходит достаточ-
В первой фазе для t ∈ [0, tK] в безбифуркацион-
но быстро. В колонию кишечной палочки, расту-
ном сценарии наблюдаются затухающие осцил-
щей по логистическому закону ∀N(0), N(t) → K с
ляции и limt→tK N(t) = K, но равновесие K стано-
небольшим «овершутом» N(t) > K, добавляли ви-
вится неустойчивым. В момент t → tK, N′(t) → 0 и
рус-бактериофаг и наблюдали необычную дина-
N(t) → K знак правой части выражения (3) будет
мику [26]. Бактериофаг активно подавлял бакте-
зависеть от значения Eh = (H - N(t - h)), выпол-
рии, но потом внезапно выживаемость у остав-
няющего роль возмущающего воздействия в за-
шихся микроорганизмов резко увеличивалась.
висимости от состояния траектории в момент tK -
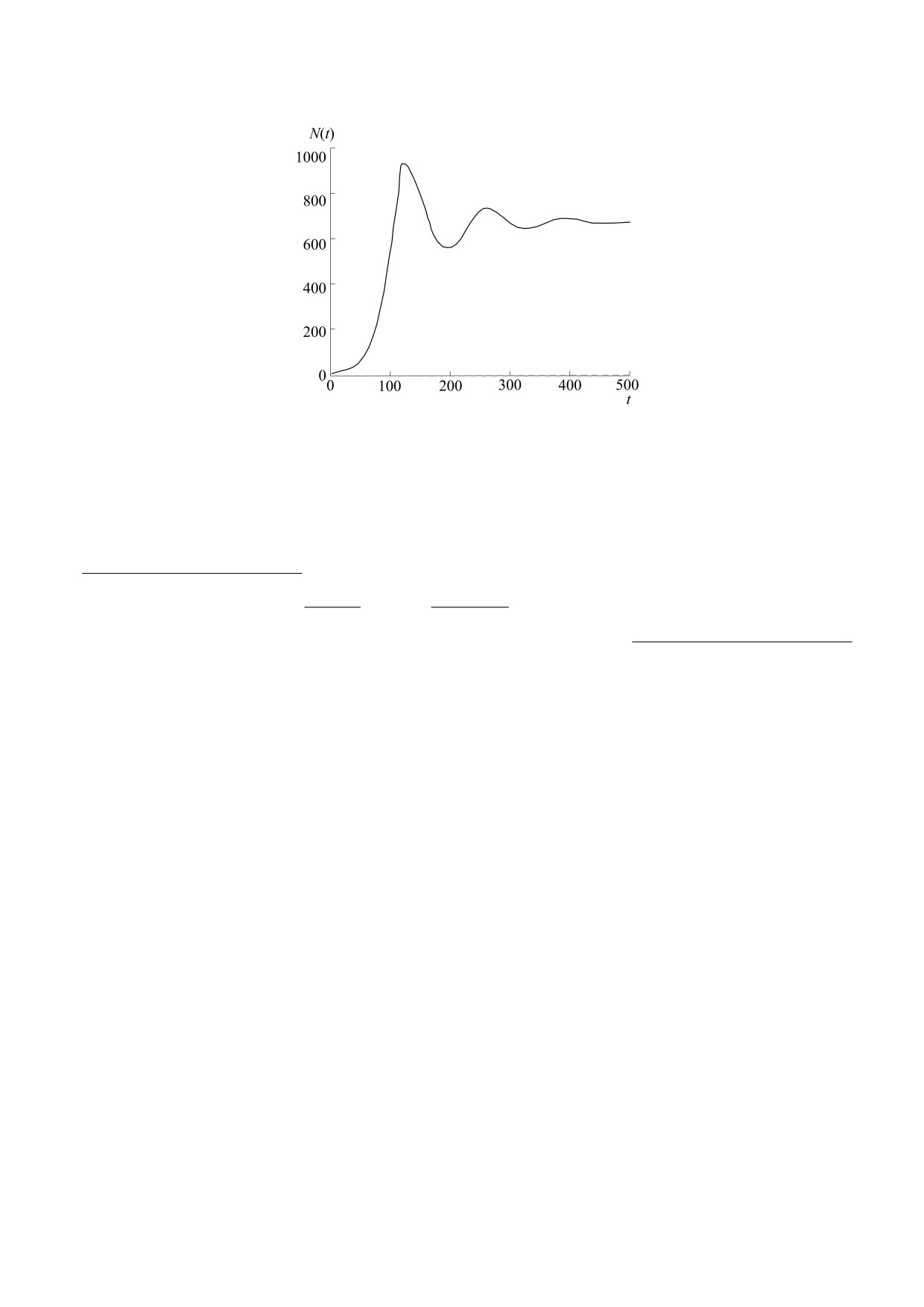
Система с вирусом проходила минимум и стаби-
лизировалась, как показано на рис. 7. Искус-
h, которое может быть положительным или отри-
цательным, что определяется при задании N(0). В
ственно созданное противоборство E. coli/бакте-
вычислительных сценариях получено два вариан-
риофаг служит лабораторной постановкой для
та: limt→∞ N(t) = +∞ при Eh > 0 и limt→∞ N(t) = H
вполне реальной ситуации при инвазии. Напри-
мер, активность разрушительного вредителя все-
при Eh < 0. Немонотонный характер приближе-
ленца непарного шелкопряда Lymantria dispar в
ния траектории N(t) → K к промежуточному рав-
лесах Северной Америки начал резко снижать па-
новесию K определяет дихотомию сценариев.
тогенный для бабочек грибок-интродуцент Ento-
При малом r-параметре формула (3) покажет про-
mophaga maimaiga, но для эффективной адапта-
сто затухающие осцилляции, в нелинейных моде-
ции грибка к новой бабочке потребовались годы
лях параметры неравноправны, не все из них ме-
[27].
няют фазовый портрет.
Для исследователей прохождения кризиса из-
Сценарий с вселением хищной инфузории не
за адаптаций E. coli был неутешительный резуль-
подразумевает совместной адаптации видов.
тат. В 1930-е гг. вирусы планировали использо-
Главным противником хищника-вселенца в та-
вать в клинической практике как антимикробные
ком сценарии стали собственные пищевые по-
средства [28]. Только много десятилетий спустя
требности и некомпенсируемая репродуктивная
смогли понять истинную причину происходящих
БИОФИЗИКА том 66
№ 6
2021
1152
ПЕPЕВАPЮXА
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ
ЗАПАЗДЫВАЮЩЕГО ПРОТИВОДЕЙСТВИЯ
Предложим модели, которые будут способны
феноменологически описывать экстремальные си-
туации после первой фазы активной инвазии. Ин-
вазии делают поведение биосистем менее предска-
зуемым [32]. В водоемах безобидные донные все-
ленцы на самом деле могут запускать цепочку
изменений в потоке переноса органических ве-
ществ [33]. В работе [6] подробно изложена исто-
рия появления идеи моделирования с учетом эф-
фекта проявления запаздывания в биологических
системах и «эредитарности» по [34], - зависимости
функционала регуляции процесса от ряда предше-
Рис. 7. Динамика колонии бактерии Escherichia coli с
ствующих состояний системы [35]. Методик вы-
адаптивной антивирусной системой CRISPR-Cas
числительного описания временного последей-
после заражения вирусом-фагом: 1 - численность
ствия известно четыре. Наиболее обосновано и ча-
бактерий, 2 - вирионов [26].
сто применяемыми являются дифференциальные
уравнения с отклоняющимся аргументом. Метод
В. Вольтерра с интегро-дифференциальными си-
в пробирке с вирусом и кишечной палочкой про-
стемами и дискретные функциональные итерации
цессов. В 2020 г. была присуждена Нобелевская
[36], включающие рекурсионное звено вычисле-
премия за исследование механизма действия
ний xn+1 = Ψ(xn, xn-1) - Q(xn-i), применяются ре-
CRISPR-Cas9, но в области геномного редакти-
же. В работе [37] предложено непрерывно-собы-
рования, а не бактериологии. Короткие палин-
тийное представление модельного времени для ги-
дромные повторы, регулярно расположенные
бридных моделей c учетом влияния стадийности
группами CRISPR в геноме многих бактерий,
онтогенеза и задержки развития при малой скоро-
представляли вирусные «сигнатуры» для внутри-
сти роста особей на их выживаемость.
клеточного антивируса, действующего по прин-
Современный обзор результатов в области раз-
ципу молекулярных ножниц, средствами для
работки модификаций непрерывных эредитар-
опознавания и разрезания чужой ДНК эндо-
ных моделей математической биологии проведен
нуклеазой Cas. Как знает любой пользователь
в работе [38]. Важно понимать несколько особен-
персонального компьютера, сигнатуры для анти-
ностей применения уравнений с запаздыванием
вирусов нужно обновлять регулярно. Горизон-
при сущностной биологической интерпретации
тальный перенос генов среди бактерий [29] поз-
результатов вычислений, часто в форме релакса-
воляет быстро преодолеть острый кризис и со-
ционных колебаний [39]. При выборе методики
здать новое равновесие. Сейчас возможности
моделирования необходимо представить заранее,
что, усложняя модель, мы сужаем диапазон зна-
геномного редактирования кажутся нам безгра-
чений в пространстве ее параметров, при котором
ничными - от создания устойчивых сортов кар-
поведение траектории будет для нас интересно и
тофеля до решения проблем онкологии [30], но
биологически обосновано. Популяционные мо-
когда-то CRISPR у E. coli сократил перспективы
дели с большим числом бифуркационных пара-
бактериофагов в лечении.
метров часто демонстрируют нелинейные явле-
ния, которые оказываются за рамками сущност-
Таким образом, мы рассмотрели два динами-
ной экологической интерпретации. Подобные
чески схожих случая прохождения кризиса «бу-
эффекты с метаморфозами аттракторов, возник-
тылочного горлышка», вызванного различными
новением «окон периодичности» и удвоением пе-
биологическими причинами, но имеющими об-
риода цикла связаны с переходом к хаотическому
щий принцип запаздывающей регуляции и адап-
поведению траектории, что трудно предвидеть.
тации. Сценарии y Гаузе не предполагали эволю-
Уравнения с запаздыванием появились в мате-
ционной адаптации, только создание простран-
матической биологии для описания флуктуаций
ственной неоднородности. Однако для многих
численности - возникающих циклических режи-
вселений насекомых и рыб адаптация - важней-
мов различной формы [40]. Как известно [41], по-
ший фактор [31]. По описанному сценарию с
пуляционные циклы образуются у изолирован-
кризисом развиваются отдельные нежелательные
ных популяций с большим репродуктивным по-
для нас процессы, потому интересно получить и
тенциалом даже при постоянстве лабораторных
сравнить математические модели обоих вариан-
условий. На амплитуду колебаний в лаборатор-
тов динамики.
ных опытах может оказывать влияние скорость
БИОФИЗИКА том 66
№ 6
2021
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО КРИЗИСА
1153
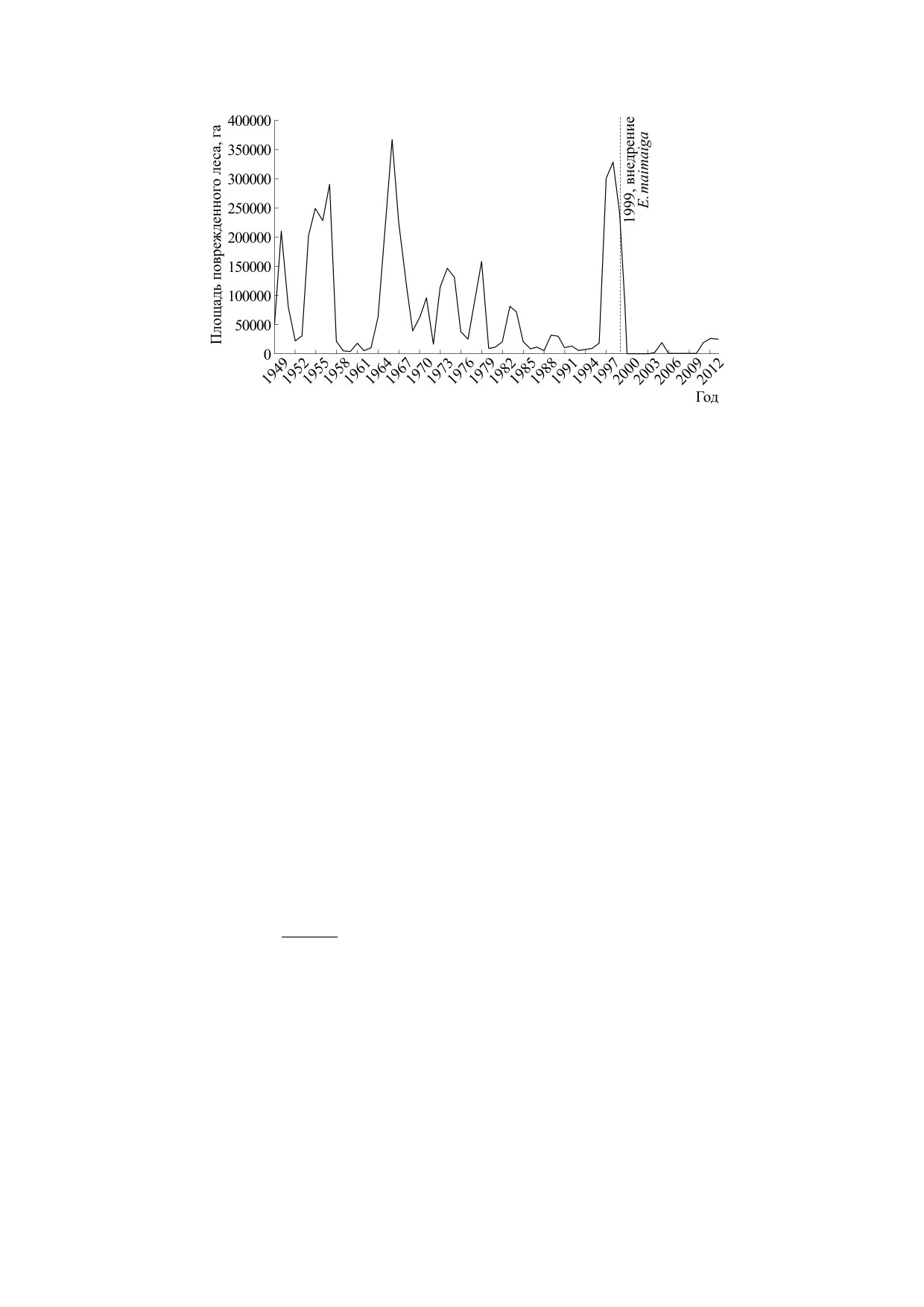
Рис. 8. Динамика серии затухающих вспышек непарного шелкопряда Lymantria dispar в Болгарии по величине площа-
ди пораженного леса с 1949 по 2013 г. [52].
восполнения питательных веществ [42], как в из-
циклы y мелких грызунов в северных широтах
вестных опытах Николсона с мухами Lucilia
представляют собой череду увеличивающихся го-
cuprina [43].
довых значений численности поколений с резким
максимумом в конце периода. После максимума
Наиболее известны в популяционном модели-
популяция снова приходит к минимальному
ровании три уравнения с запаздыванием, способ-
L-уровню [49].
ные генерировать колебания, в том числе сложной
формы [44], - модели Хатчинсона, Николсона и
Пилообразные вспышки y насекомых-вреди-
Гополсами, имеющие ряд модификаций [45]. По-
телей представляют собой отдельные серии спо-
явлению устойчивого цикла N*(t) вокруг теряющей
радических Λ-образных пиков с тенденцией
уменьшения амплитуды, где максимальным ча-
устойчивость стационарной точки отвечает наибо-
сто оказывается второй пик [50]. Нами показано,
лее распространенная бифуркация Андронова-
что сценарий, известный в динамике насекомых-
Хопфа, отражающая мягкую потерю устойчивого
вредителей как «sawtooth-like oscillation» - пило-
режима [46]. Для моделирования установившихся
гармонических колебаний у изолированной попу-
образная вспышка численности с разновеликими
затухающими пиками, отличается от иных вари-
ляции без внешних факторов воздействия, или яв-
антов развития вспышек [51]. Для нарушения
ления «овершута» - краткого превышения при ин-
равновесия и для запуска очередного переходно-
вазии быстро растущей популяцией К-емкости ни-
го режима колебаний сценарий требует внешнего
ши ∃tm < tc, ∀N(0) < K : N(tm) > K, N(tc) < K,
воздействия. На рис. 8 представлена динамика се-
используются модификации модели Хатчинсона.
рии затухающих вспышек непарного шелкопряда
Предложенное в работе [47] Райтом уравнение (4)
Lymantria dispar в лесах Болгарии. Очередная се-
на основе идей Г. Хатчинсона в свою очередь явля-
рия в 2000 г. резко завершилась на десятилетие
ется расширением модели Ферхюлста с запаздыва-
из-за активности энтомапатогенного грибка
нием τ при регуляции размножения:
Entomophaga maimaiga, нового паразита для гусе-
ниц [52]. Со временем при совместной эволюции
dN
N(t
−τ)
=
rN t)
1−
(4)
некоторые насекомые, как, например, уже при-
(
)
dt
K
способившиеся термиты, приобретают способ-
Режим затухающих осцилляций limt→∞ N*(t) = K
ность чувствовать и избегать опасные грибки.
можно получить в (4) при 1 < rτ < π/2, N(0) < K, но
Сценарий кратких вспышек, характерных для
мы не сможем получить в (4) сценарий остановки
динамики псиллид на юге Австралии, представ-
колебаний со снижением до минимальной ста-
ляет особое явление в биокибернетике с другим
бильной группы особей limt→∞ N*(t) = L, даже с
механизмом регуляции - пороговым развитием
независимой убылью. При rτ = π/2 + ε точка-фо-
фаз процесса [53]. Порог численности для начала
кус потеряет устойчивость и реализуется бифур-
вспышки y Psyllidae достижим из состояния сто-
кация рождения цикла [48]. Гармонические коле-
хастических колебаний. Наличие порога связано
бания с равномерным затуханием редко встреча-
с активностью ос-сверхпаразитов. Вспышка y
ются в экологической реальности. Известные
псиллид завершается переходом к долгому апери-
БИОФИЗИКА том 66
№ 6
2021
1154
ПЕPЕВАPЮXА
Рис. 9. Сравнение динамики колебательных режимов (4) и (7) при одинаковых τ: 1 - релаксационный цикл в модели
Хатчинсона (4), 2 - флуктуации в уравнении Николсона (7).
одическому режиму. Пороговые эффекты важны
Таким образом, при увеличении значения rτ > 3π
для промысловых популяций как причина кол-
модель Хатчинсона теряет возможности популя-
лапса запасов [54].
ционной интерпретации колебаний, так как в
Известной проблемой модели (4) при практи-
длительном малочисленном состоянии велика
ческом применении уравнения становится свой-
роль любых случайных возмущений даже для по-
ство, когда при увеличении значения rτ образую-
пуляции без действия эффекта Олли.
щийся цикл негармонической формы с увеличе-
Для преодоления недостатков циклического
нием амплитуды получает минимумы в
решения уравнения (4) была предложена модель
окрестности нуля: limt→∞ min N*(t,rτ) = 0 + ε.
Гополсэми [55], где действующая емкость эколо-
Продолжительные глубокие «околонулевые» ми-
гической ниши уже не статична:
нимумы не соответствуют наблюдениям о попу-
dN
K -N(t
−τ)
ляционной динамике видов, для которых отмече-
=
rN t)
(5)
ны эруптивные фазы при стремительном разви-
dt
K + jN(t
−τ)
тии вспышек численности. Масштабы вспышек
Модель (5) мы модифицировали, неравномер-
численности вредителей отражают размеры пора-
но увеличив нелинейности в числителе и в знаме-
женных ими лесных площадей. Отсутствие по-
нателе, чтобы следующий пик колебаний начи-
гибшего леса в графиках отчетов лесного хозяй-
нался от положения min N*(t,r) выше стабильного
ства означает, что локальная популяция находит-
ся в состоянии со стабильной разреженной
L-порога численности, а Ψ(Nk(t - h)) нормирует
плотностью, но совсем не близка к вымиранию.
скорость затухания колебаний:
2
dN
С -N
(t
−τ)
k
=
rN t)
−Ψ[N
(t
−
h)],С <K
(6)
3
dt
K
+γN
(t
-τ)
Для разрушающих среду видов вредителей ем-
dN = rN
(t
−τ)exp(
−bN
(t
−τ))
−δN t),
(7)
кость ниши здесь не остается статичной. Альтер-
dt
нативными модификациями для моделей (4) и (5)
можно считать уравнения без явной фиксации
где δ - показатель независимой смертности. Мо-
уровней, как достижимого оптимального значе-
дель (7) допускает апериодический режим флук-
ния численности С или использования предела
туаций при увеличении τ (рис. 9). Эксперименты,
емкости экологической ниши K. Уравнение blow-
описанные в работе [56], подразумевали конку-
f lies equation было предложено для описания дан-
ренцию между личинками и имаго. Модель blow-
ных (подсчета личинок и имаго) экспериментов c
f lies equation только феноменологически описы-
лабораторной популяцией мух при большой ам-
вает релаксационные колебания общей числен-
плитуде колебаний [56]:
ности, используя свойства функции Рикера.
БИОФИЗИКА том 66
№ 6
2021
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО КРИЗИСА
1155
Обобщение моделей (4) и (7) в модель (8) для
Онтогенетическая задержка развития поло-
возрастной структуры не получило широкого
возрелого организма - только один из временных
применения:
факторов и значимый далеко не для всех ситуа-
ций. Важнее те показатели задержки, что регули-
−τ
2
∂N
1
−bN(t−τ)
=
r
1−
dv s)N(t
+
s)
N t)e
(8)
руют репродуктивную активность. Величину τ в
(
)
∂t
K -τ
1
модели разрушения осцилляций (2) мы полагаем
Вместо экспоненциальной функции регуля-
результатом взаимодействия вида и конкретной
биотической среды, формирующегося в данном
ции Рикера f(x) = axe-bx при модификации моде-
ареале. Величину действующего запаздывания
ли
(7) можно использовать неунимодальную
популяционных моделей могут составлять онто-
функцию f(x) для формализации внутривидовой
генетические свойства, интервалы времени эта-
конкуренции без использования фиксированных
пов развития, скорость восстановления необхо-
значений численности порогов конкуренции или
димых для жизни вида ресурсов, время миграции
насыщения ниши. Унимодальные варианты f(x)
и др. При инвазиях значимо время выработки от-
рассмотрены в работе [57]. Отдельно можно рас-
вета со стороны паразитов или патогенов.
смотреть модификации с использованием идеи
Для борьбы иммунитета и вируса в организме за-
модели Базыкина о влиянии критически низкой
паздывание - индивидуально зависимая характе-
численности L в виде (N - L), но c введением за-
ристика всех реакций активации, среднее значе-
паздывания, как в модели Хатчинсона:
ние может варьироваться в локальных субпопуля-
dN
N(t
−τ)
циях.
=
rN t)
1−
(N t)
−
L)v.
(9)
(
)
dt
K
В работе [59] показано, что репродуктивную
Наблюдаемая нами цикличность численности
составляющую популяционной модели rN(t)
вида в реальности может быть связана с суще-
адекватнее представлять без запаздывания, но
ствованием репродуктивно изолированных суб-
для пространственной компоненты величина τ
популяций. Такие группы промысловая стати-
будет значима. Уравнения с запаздыванием
стика не имеет возможности различать в отчетах,
N(t-τ) широко применяются не только для опи-
как отмечено для волжской севрюги [58]. Общие
сания колебательной динамики популяций [60]
данные статистики о популяциях со сложной
или при моделировании иммунных реакций (в
структурой вводят в заблуждение.
современном развитии работ Г.И. Марчука [61],)
Для моделирования кризисных эффектов при
но и для анализа внутриклеточных процессов, на-
быстрой инвазии необходимо расширить приме-
пример, для синтеза белков, репликации нуклеи-
няемые методы описания динамики с запаздыва-
новых кислот и др. В работе [62] при моделирова-
нием не только для внутривидовой регуляции, но
нии альтернативного сплайсинга РНК параметр
и оказываемого противодействия, которое в свою
запаздывания аналогично трактовался как агре-
очередь может изменяться пороговым образом,
гированная величина из двух основных составля-
например, из-за разной начальной дозы зараже-
ющих, суммарного времени сплайсинга пре-
ния гриппом [15].
мРНК в мРНК и транспорта мРНК из клеточного
ядра в цитоплазму клетки. При построении моде-
лей жестко контролируемых процессов важно со-
ПРЕДСТАВЛЕНИЕ ЗАПАЗДЫВАНИЯ
хранять поведение траектории предсказуемым в
В РЕГУЛИРУЮЩЕМ ФУНКЦИОНАЛЕ
вычислительных экспериментах, так как реше-
ние может стать чувствительным по отношению к
В развиваемой нами методике моделирования
малому возмущению начальных условий N(0) ± ε
параметр запаздывания для динамики размножа-
и выбору функции-предыстории φ0 на интервале
ющихся в конкурентной среде популяций нужно
трактовать как агрегированную характеристику,
времени φ0 (ζ),-τ ≤ ζ < 0.
появляющуюся при наложении нескольких био-
системных факторов и частных сопутствующих
Исходя из изложенных принципов o роли ре-
процессов. Временное последействие в составе
гуляции с последействием, модификации наших
регуляторных функций разрабатываемых уравне-
уравнений будут включать внутреннюю регуля-
ний относится не к характеристикам отдельного
цию репродуктивной активности с τ (возможно, с
биологического вида, а к системному взаимодей-
использованием пороговых состояний численно-
ствию с участием окружения вида, часто не явно-
сти вида) и отдельно функционал запаздывающе-
му, а опосредованному. В модели протекания ви-
го внешнего противодействия среды. Используем
русной инфекции при разной силе иммунного от-
логарифмическую функцию регуляции ln(K/N) в
вета (3) мы выделили отдельно репликационный
модели как базовую для дальнейших специаль-
и адаптационный временные показатели для яв-
ных модификаций при стационарной емкости
лений запаздывания при активации разных ком-
ниши, какая существовала в опытах с инфузория-
понентов иммунитета.
ми и бактериофагами:
БИОФИЗИКА том 66
№ 6
2021
1156
ПЕPЕВАPЮXА
максимумов у решений уравнения Хатчинсона
(4) и нашей базовой модели (10).
При стремительной инвазии изначально ма-
лая группа вида-вселенца за некоторое, но огра-
ниченное короткое время после вселения
может быстро достичь огромной численности
(N(tK) >> K), на порядок превосходящей итоговое
равновесие со средой. Стремительный рост сме-
няется аналогичным резким сокращением попу-
ляции и осцилляциями вокруг равновесия, и та-
кой слабо колеблющийся диапазон численности
не оказывает давления на среду. Динамика кон-
центрации бактериофага в экспериментах после
внесения вируса укладывается в полученный сце-
нарий, но антагонисты бактерии демонстрируют
качественно иной путь.
К вспышке по сценарию (10) способна исход-
ная группа с N(0) значительно меньше K. Извест-
Рис. 10. Сравнение динамики моделей при одинако-
вых K = 15000, τ = 48, N(0) = 15: 1 - единичная
ны другие опытные примеры остановки вспыш-
вспышка в нашем уравнении (10) при r = 0.0175; 2 -
ки. Так, после начала искусственного выращива-
релаксационный цикл в модели (4) при r = 0.07.
ния деликатесного гребешка Chlamys farreri в
морских садках удалось быстро увеличить его
продукцию (рис. 11). Потом вирусная инфекция в
dN
K
=
r
ln
N t).
(10)
1997 г. в шесть раз сократила производство гре-
dt
N(t
-τ)
бешка, и никакими мерами не получилось вос-
становить производство до объемов 1996 г. [63].
В модели (10) можно получить затухающие и
Неоднократно попытки борьбы с вредителями
циклические колебания, но экологически более
путем создания стабильных популяций выпуска-
интересным представляется другой вариант пове-
емых паразитических наездников заканчивались
дения решения уравнения. Данный сценарий ос-
неудачей. Сокращение численности отмечалось
недавно, например, даже для инвазионных мол-
цилляционного достижения балансового равно-
люсков из семейства Dreissenidae, заполонивших
весия в решении уравнения (10) отличается от
водоемы в Северной Америке и ставших в США
сценариев известных моделей и возможен, когда
большой экономической проблемой. Инвазив-
N(0) мало, но τ достаточно велико (рис. 10). Пара-
ные виды моллюсков Dreissena polymorpha и D. ros-
метры в сравнительном вычислительном экспе-
triformis bugensis начали так конкурировать между
рименте подобраны для совпадения значений
собой [64], что их совокупная плотность биомас-
Рис. 11. Продукция в тыс. тонн искусственной популяции морского гребешка Chlamys farreri с кризисом в 1997 г. из-
за эпизоотии, вызванной вирусом герпеса устриц [63].
БИОФИЗИКА том 66
№ 6
2021
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО КРИЗИСА
1157
сы будет меньше, чем когда нишу захватывает
один из видов.
Модель описывает единичный всплеск чис-
ленности инвазивного вида. Повторные пики
численности [65] после первичной вспышки ха-
рактерны для насекомых, повреждающих лес.
Вредители переходят к пилообразным колебани-
ям, о которых писали выше. Сценарий развития с
Λ-образных кратких вспышек y тлей и псиллид
требует других метаморфоз аттракторов [66]. Ди-
намика модели (10) не описывает случай перете-
кания острой вирусной инфекции в хроническую
болезнь, так как в таком сценарии высокая вирус-
ная нагрузка фиксируется продолжительное вре-
мя около неустойчивого равновесия - предела
развития.
МОДЕЛЬ «СЦЕНАРИЯ ГАУЗЕ»
Рис. 12. Сценарий гибели популяции после второго
С РАЗРУШЕНИЕМ КОЛЕБАНИЙ
максимума колебаний: 1 - решение модели (11) при
Давление со стороны среды может линейно за-
r = 0.0175, δ = = 0.007, N(0) = 103, τ = τ1 = 48,
висеть от предшествующего состояния вселенца.
K = 15000; 2 - поведение уравнения (4) с аналогичной
модификацией c δ при r = 0.05.
Учтем теперь в модификации с фактором проти-
воборства модели (10) отложенную реакцию био-
тической среды, но пожертвуем удобным свой-
произошел их внезапный коллапс, а вытеснен-
ством ∀t, N(0) > 0, N(t) ≥ 0:
ные им автохтонные виды муравьев восстановили
ареал [67].
dN
K
=
r
ln
N t)
−δN(t
−τ
),
(11)
1
dt
N(t
-τ)
где δ - показатель фактора убыли, обусловленно-
МОДЕЛИРОВАНИЕ ПРОХОЖДЕНИЯ
го действием паразитов или вирусных инфекций.
КРИЗИСА «БУТЫЛОЧНОГО ГОРЛЫШКА»
Поведение модели (11) в вычислительном экспе-
Сценарий гибели инвазивных популяций, ак-
рименте показывает прекращение существова-
тивно истощающих необходимые им ресурсы и
ния популяции только после повторной вспыш-
столкнувшихся с линейно заданным противодей-
ки. Второй максимум решения превосходит пер-
ствием с запаздыванием, является актуальным
вый пик активности (рис. 12), но вторая точка
для ситуации мер искусственного подавления не-
минимума находится вне допустимых значений,
желательных вселенцев. В современной практике
и в расчетах модели при N(t > τ) < -ε вычисли-
все чаще одни виды-интродуценты (например,
тельный эксперимент завершается алгоритмом
паразитические перепончатокрылые) использу-
инструментальной среды с фиксацией гибели по-
ются для борьбы с другими вселенцами в качестве
пуляции.
средств биологического контроля [68], но полно-
Модель (11) феноменологически воспроизво-
го подавления удается добиться редко. Интересен
дит динамику эксперимента Гаузе с инфузория-
опыт Южной Кореи по длительной систематиче-
ми, но это лабораторный и искусственный сцена-
ской борьбе с карантинным вредителем амери-
рий in vitro, где гибель хищника-вселенца опреде-
канской белой бабочкой Hyphantria cunea [69]. В
лена нехваткой быстро истраченных пищевых
моделируемом сценарии вырабатываемое средой
ресурсов. Момент t0 времени исчезновения
(или интродуцентом) противодействие может
N(t0) = 0 для (11) зависит от N(0), при начальных
определяться состоянием инвазивной популяции
условиях N(0) = 100 максимум будет единствен-
на пике численности и в момент времени, когда
ным. Сценарий с исчезновением наблюдается в
численность популяции по независимым причи-
решении (11) для больших значений r-параметра,
нам прошла свой максимум.
при уменьшении r можно увидеть регулярные ко-
Внешнее давление может быть организовано
лебания minN*(t) > ε. Наиболее известный неже-
сложнее, чем пропорциональное изъятие -δN(t-τ).
лательный инвазивный вид, для которого неод-
Реакция выпускаемых паразитических ос опреде-
нократно описано спонтанное исчезновение по-
лена кучностью их жертв, а патогенные вирусы
сле вспышки
- аргентинский муравей
начинают быстрее распространяться в плотных
Linepithema humile. В Новой Зеландии агрессив-
скоплениях. Часто активно размножающегося
ный вселенец создал крупные колонии, но потом
вселенца настигает эпизоотия при повышении
БИОФИЗИКА том 66
№ 6
2021
1158
ПЕPЕВАPЮXА
Рис. 13. Сценарий кризиса быстрорастущей популяции в модели (12) при r = 0.0717, K = 15000, τ = 48, δ = 10, J = 103,
q = 0.0042, m = 2, N(0) = 10.
численности задолго до уровня, когда опасный
модификацию модели для наиболее интересной
вселенец сможет сам разрушительно воздейство-
ситуации, когда запаздывающее противодей-
вать на среду, при приближении к некоторому
ствие быстро растущей популяции задается нели-
порогу численности J < K. Рассмотрим третью
нейно и пороговым образом:
m
dN
K
N
(t
−τ)
=
r
ln
N t)
−δ
-
qN t),N(0)
<
J
K,
(12)
2
dt
N(t
-τ)
(J
-
N t))
где J - пороговый уровень для усиления агрессии
J, показатель m определяет глубину кризиса. При
биотического окружения, m - показатель силы
q = 0 сохраняется колебательная динамика. Мо-
реакции среды на плотность вселенца. Усложнен-
дель применима для сценария инвазии малочис-
ный c точки зрения биологической кибернетики
ленных групп, при приближении начальных
механизм регуляции неожиданно ведет к более
условий к порогу J кризис становится критиче-
пластичному сценарию эволюции. Увеличение
ским V-образным, при N(0) > J решение уравне-
численности от исходной малой группы N(0) < J в
ния (12) продемонстрирует одну из форм логи-
первой стадии процесса идет по обычному логи-
стического роста. Значительная величина τ в мо-
стическому закону, но вместо N(t) → K оказыва-
дели не приводит здесь к устойчивому циклу.
ется в переходном режиме колебаний гораздо
Величина порога J по своей сути в нашей модели
раньше, чем в моделях (4) или (7). В новой модели
зеркальна величине L в модели (9) как нижней
(12) вместо плавного замедления роста после точ-
пороговой численности особей из модели Базы-
ки перегиба или остановки в стадии с превышаю-
кина, предложенной им для описания действия
щим «овершутом» N(tK) > K начинается этап рез-
эффекта Олли. Причины возникновения порого-
кого кризиса при t > 2τ (рис. 13). Обобщенно мо-
вых состояний для популяций и переходных ба-
дель с постоянными факторами запаздывания τ,h
рьеров в экодинамике рассмотрены в [70]. При-
представляется балансом функции регуляции эф-
мер возникновения порога сверху, который со-
фективности воспроизводства Φ и функции про-
здается из-за активности сверхпаразита,
тиводействия биотической среды Ψ, где важней-
приведен в работе [71] о механизмах регуляции
шим является соотношения показателей m и d:
размножения эвкалиптовых псиллид. Порог, ко-
торый преодолевает популяция в дискретной мо-
dN
d
m
=
rN t)Φ[N
(t
−τ)]−Ψ[N
(t
−
h)].
дели с бифуркационным завершением вспышки,
dt
возник не у самих вредителей, а у их естественных
При приближении к порогу N(t) → J убыль
врагов из-за атак сверхпаразитов [72]. В работе
резко превосходит рождаемость, и популяция
[73] отмечено, что удельная эффективность пара-
проходит состояние «бутылочного горлышка». В
зитизма специально выпускаемых наездников се-
результате прохождения кризиса траектория ре-
мейства Trichogrammatidae возрастает с увеличе-
шения стремится к равновесию на уровне меньше
нием плотности яиц бабочек.
БИОФИЗИКА том 66
№ 6
2021
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО КРИЗИСА
1159
В результате образующаяся инвазивная попу-
цированных растений [74]. Бактерии и миксоми-
ляция с большим репродуктивным потенциалом
цеты ведут химическую войну, в результате
может продемонстрировать опасную вспышку
которой был открыт пенициллин, но со временем
численности, только если выполнится условие
первые антибиотики потеряли эффективность.
N(t) > J, но при этом оказываемое сильное сопро-
Инвазионные виды часто нарушают баланс в
тивление не приводит к критическому минимуму
трофической цепи [75], запуская эффект домино
численности. Популяция успешно проходит «бу-
и вызывая резкие колебания y других видов.
тылочное горлышко», так как давление среды на
Предложенные уравнения пригодны для исполь-
популяцию, зависящее от предшествующего ее
зования в составе систем для моделирования не-
состояния, ослабляется, если текущая числен-
посредственного трофического взаимодействия.
ность становится значительно меньше порога.
В уравнение (12) вместо q можно ввести явное
Модель с усложненной регуляцией противо-
влияние вида-антагониста P:
действия среды оказывается примером адаптаци-
онной динамики при прохождении глубокого
m
кризиса, подобного ситуации колонии бактерий с
dN
K
P
(t
−ξ)
=
r
ln
N t)
−δ
(13)
2
противовирусным механизмом CRISPR после
dt
N(t
-τ)
(J
−
N t))
интродукции вируса-бактериофага. Полученный
в новой модели сценарий актуален не только для
Для описания эффекта убежища (рефугиума) с
коэволюции паразита/хозяина. Насекомые вре-
возможностью исчезновения реликтовой мало-
дители аналогично приспосабливаются к выво-
численной популяции можно использовать сте-
димым устойчивым сортам генетически модифи-
пень 1/3 для действия нижнего L-порога:
dN
K
3
=
r
ln
N t)
N(t
−κ)
−
L
−Ψ[P(t
−ξ)], 0
< κ<τ
(14)
dt
N(t
-τ)
Динамику конкурирующего при взаимодей-
гибридные модели с событийным преставлением
ствии вида P(t) интересно описывать с колебани-
времени [79], так как необходимо формализовать
ями по другому механизму регуляции, что мы об-
логику управления.
судим в следующей статье. Предложенные моде-
ли
(12) и
(14) объясняют многочисленные
ЗАКЛЮЧЕНИЕ
примеры неудачных целенаправленных интро-
дукций.
В предложенных четырех моделях (10), (11),
(12) и (14) для специфических случаев популяци-
Дальнейшее усложнение поведения траекто-
онных процессов мы рассмотрели варианты рез-
рии решения, например, с введением в балансо-
ких изменений численности, которые некоторы-
вое уравнение для Φ[Nd(t - τ)] и Φ[Nd(t - τ)]
ми авторами относились к типу «эруптивных»
функций Бивертона-Холта или Рикера, как было
экологических процессов или, по терминологии
выполнено в работе [76], не представляется био-
современных работ, динамике формы «Boom and
логически обоснованным. В работе [77] показа-
bust» [80]. Для каждой из моделей мы указали
но, в каких случаях при вычислительном анализе
примеры экологических ситуаций, наиболее
лучше избегать излишней нелинейности в моде-
соответствующие полученному поведению реше-
лях биосистем, как хаотизации по сценарию Фей-
ния уравнения. Согласно классификации в рабо-
генбаума с бесконечным каскадом бифуркаций
те [81], процессы изменений в биологических
периода p = 2i, i → ∞ цикла ϕ(x*)n = ϕ(x*)n+p y ите-
сообществах различаются по формам как «гради-
раций xn+1 = ϕ(xn). В итерационных моделях из-
ентные» или «эруптивные». Градиентные случаи
быточных эффектов может быть несколько, на-
легко объяснимы, например, сукцессия. Ситуа-
пример, разрушение странных аттракторов, обра-
ция изменилась в пользу одного из видов незави-
зование
фрактальных границ областей
симо от него, что можно отразить увеличением K,
притяжения, появление окон периодичности с
и последующим монотонным, хоть и быстрым,
переходами «хаос ↔ цикл», где циклы подчиня-
ростом численности. Эруптивная и немонотон-
ются порядку теоремы А.Н. Шарковского [78], но
ная динамика на самом деле многообразна, не
не экологическим принципам. Для каждого мате-
ограничивается цикличностью фаз роста и сни-
матического эффекта, возникающего в предло-
жения у видов-конкурентов и представляет от-
женной модели, необходима сущностная биоло-
дельную проблему для методики моделирования
гическая интерпретация. Для ситуации при
биосистем.
управляемом антропогенном воздействии или
В работе [82] отмечено 16 ситуаций, когда на-
модели сценария кризиса, вызванного эксплуата-
блюдалось стремительное распространение инва-
цией биоресурсов, целесообразно разрабатывать
зионного вида, но затем следовал аналогичный
БИОФИЗИКА том 66
№ 6
2021
1160
ПЕPЕВАPЮXА
стремительный коллапс. Явление коллапса авто-
порог) без непосредственного взаимодействия
рами трактуется как снижение численности на
встречается в нашей жизни. Из-за вирусной ин-
90% от оптимума, и такое снижение длится не ме-
фекции может начинаться бактериальная пнев-
нее трех лет. Мы рассмотрели варианты в моделях
мония, если популяция альвеолярных макрофа-
(10) и (12), когда фазы стремительного увеличе-
гов истощается [85]. Многоуровневые схемы ре-
ния перемежаются с глубокими кризисами,
гуляции для контроля размножения видов с
вплоть до риска внезапного исчезновения в моде-
высоким репродуктивным потенциалом интерес-
ли (14). Удалось продемонстрировать на приме-
ны для биологической кибернетики, но сложны
рах графиков из опытов Гаузе и экспериментов с
для непосредственного математического описа-
бактериофагами, что можно эффективно исполь-
ния их звеньев. Скорость размножения видов и
зовать уравнения с запаздыванием не только для
длина их жизненного цикла может оказаться в
классической задачи моделирования эндогенных
разных и даже в несравнимых временных мас-
долговременных и устойчивых осцилляций чис-
штабах. Упрощение до замкнутой системы «хищ-
ленности [83] или затухающей серии пилообраз-
ник-жертва» приводит к результату N = 0 из экс-
ных вспышек бабочек. Включение в модели с
периментов Гаузе. Процесс эволюционной адап-
N(t-τ) порогового регулирующего функционала
тации создает иногда неожиданные барьеры,
Ψ[N(t-h)] актуально и для описания свойств экс-
которые препятствуют тотальному доминирова-
тремальных форм инвазивной динамики и для
нию даже эффективного хищника или паразита
сценариев с быстрой адаптацией. Анализировать
[86]. Например, субвирусные паразиты вирофаги
запаздывание при регуляции актуально для про-
используют для своего размножения крупные ви-
цессов различной природы [84], имеющих не-
русы, заражающие амеб, и так невольно повыша-
сколько выраженных отдельных стадий развития
ют выживаемость самих амеб [87]. Бактерии рода
и условий переходов между стадиями.
Bdellovibrio паразитируют в других бактериях и
могут в перспективе использоваться в клиниче-
Линейное и зависимое от предшествующего
ской практике [88]. Самая сложная известная нам
состояния внешнее воздействие Ψ = -δN(t-τ) в
модели (11) способно привести к гибели популя-
система обнаружена у плесневых грибков в тро-
пиках [89], где борются десятки конкурирующих
ции. В модификации (12) в момент быстрого ро-
видов и химически подавляют друг друга. Острая
ста моделируемая популяция попадает в глубо-
конкуренция за ограниченный ресурс становится
кий кризис раньше, чем достигает теоретически
фактором активного видообразования, появле-
возможного уровня экологической ниши. Одна-
ко регуляция в этом сценарии сложнее, и это до-
ния множества видов-эндемиков из одного се-
мейства не только у микроорганизмов, как, на-
полнение позволяет успешно миновать стадию
пример, в случае 35 видов рыб семейства Gobiidae
бутылочного горлышка, что демонстрирует в
в Каспийском море [90]. При изучении эволюци-
опытах колония E. сoli. Разработанные нами мо-
онной борьбы в микромире предстоит еще много
дели описывают сценарии смены динамических
режимов без бифуркаций, такие переходы в сце-
интересных для практики открытий, так, по-ви-
димому
[91], археи получили независимо от
нариях не требуют изменений значений парамет-
CRISPR-Cas бактерий отдельный антивирусный
ров по ходу модельного эксперимента. При моде-
механизм.
лировании процессов важно понимать, какие из
наблюдаемых изменений допустимо представ-
При разработке систем уравнений согласно
лять бифуркациями аттракторов - возникнове-
нашему принципу целесообразно разделять типы
нием топологически неэквивалентных фазовых
эффектов запаздывания. Мы выделили репро-
портретов у траектории при возмущении внут-
дуктивное запаздывание, регуляционное запаз-
реннего параметра. Параметры в нелинейных мо-
дывание как фактор отложенной реакции среды и
делях неравноправны по своему влиянию на то-
адаптационное запаздывание. Популяции (или
пологию фазовых портретов. Сдвиг вектора пара-
организму) на выработку приспособительных ре-
метров уравнения - это самый простой способ,
акций необходимо некоторое время, и такая ве-
но не всегда биологически обоснованный. Внут-
личина задержки может изменяться в процессе
ренние характеристики биосистем меняются эво-
инвазии. В интересных случаях развития инвазии
люционно согласованно. В большинстве случаев
происходит динамическая корректировка оказы-
скачкообразное изменение характеристик выгля-
ваемого воздействия со стороны биотической
дит нереалистично (исключая быстрые мутации у
среды. Воздействию подвергается не просто чис-
вирусов и прокариотов), потому мы в серии работ
ленность популяции, как было в случае коллапса
развиваем иные математические средства.
запасов трески Северной Атлантики из-за незна-
Рассмотренная в модели (12) сложная порого-
чительной, но системной ошибки при определе-
вая регуляция, обоснованная тем, что вид-регу-
нии допустимого промыслового изъятия поло-
лятор сам имеет ограничивающий фактор, не яв-
возрелых особей рыб
[92]. Трансформируется
ляется уникальным явлением. Ситуация, когда
функция регуляции эффективности воспроиз-
один вид разрушает порог (или, наоборот, создает
водства, и показатель m в модели (12) может зави-
БИОФИЗИКА том 66
№ 6
2021
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО КРИЗИСА
1161
сеть от t. Существуют процессы, где все три фак-
7. M. Shrotri, M. van Schalkwyk, N. Post, and D. Eddy,
тора запаздывания могут конкурировать - раз-
PLoS One 16 (1), e0245532 (2021).
личные средства иммунного ответа имеют разное
8. S. Fung, K. Yuen, Z. Ye, et al., Emerging Microbes &
время активации и ингибирования, как и циклы
Infections 9 (1), 558 (2020).
создания копий вируса. Дисрегуляция иммунно-
9. A. Y. Perevaryukha, Izvestiya Vysshikh Uchebnykh Za-
го ответа, в том числе и особенно его гиперакти-
vedeniy. Prikladnaya Nelineynaya Dinamika 25 (2), 51
вация, только усиливают патогенез инфекции
(2017).
COVID и ВИЧ [93]. Циклические решения при
10. M. Sakharkar, G. Rappazzo, and W. Wieland-Alter,
моделировании борьбы вирус/иммунитет неакту-
Science Immunol. 6 (56), eabg6916 (2021).
альны.
11. F. Tavasolian, M. Rashidi, and G. R. Hatam, Front.
Отметим, что логико-алгоритмические мето-
Immunol. 11 601886 (2021).
ды способны учесть еще больше факторов собы-
тийных изменений при моделировании противо-
12. Ю. Д. Нечипуренко, А. А. Анашкина и О. В. Мат-
борства пространственно неоднородных популя-
веева, Биофизика 65 (4), 824 (2020).
ций
[94]. При распространении инвазивных
13. R. Shukla, V. Ramasamy, R. Shanmugam, et al., Front.
видов в новом ареале часто возникают фронты
Cell. Infection Microbiol. 10, 572681 (2020).
повышенной плотности, что наблюдается и в
14. N. Suryadevara, S. Shrihari, P. Gilchuk, and L. A. Van
обычных микробных сообществах [95]. В алго-
Blargan, Cell 184, 1 (2021).
ритмы клеточных автоматов можно включать
15. C. van Eeden, L .Khan, and M. S. Osman, Int. J. Mol.
факторы запаздывания [96], но по специальным
Sci. 21 (17), 6351 (2020).
правилам [97]. В настоящее время перспективно
развивать программные реализации интеллекту-
16. M. Larsen, L. de Graaf, and E. Sonneveld, Science 371
альных клеточных автоматов с вариабельной
(6532), eabc8378 (2021).
плотностью решеток клеток и со стохастической
17. K. McMahan, J. Yu, N. Mercado, et al., Nature 590,
составляющей алгоритма, которые могут приме-
630 (2021).
няться для прогнозирования пространственно
18. I. Marois, A. Cloutier, E. Garneau, and V. Richter, J.
неоднородных процессов в биосистемах.
Leukocyte Biol. 92 (1), 107 (2012).
19. 19. S. Colledge, J. Conolly, and E. Crema, Quaternary
ФИНАНСИРОВАНИЕ РАБОТЫ
Res. 93 (3), 1 (2019).
Работа выполнена при финансовой поддержке
20. А. Ю. Переварюха, Биофизика 61 (2), 395 (2016).
Российского фонда фундаментальных исследова-
21. J. Javidpour, C. Molinero, and E. Ramírez-Romero,
ний (грант № 17-07-00125) и за счет бюджетной
Commun. Biol. 3 212 (2020).
темы № 0073-2019-0003 Санкт-Петербургского
22. D. Simberloff and L. Gibbons, Biol. Invasions 6 (2),
Федерального исследовательского центра РАН.
161 (2004).
23. С. V. Pushkin, Rus. J. Biol. Invasions 1, 42 (2008).
КОНФЛИКТ ИНТЕРЕСОВ
24. G. F. Gause, The Struggle for Existence (Williams &
Автор заявляет об отсутствии конфликта инте-
Wilkins, Baltimore, 1934).
ресов.
25. Г. М. Заславский, Стохастичность динамических
систем (Наука, М., 1984).
26. R. E. Lenski, Adv. Microb. Ecol. 10, 1 (1988).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
27. T. Andreadis and R. Weseloh, Proc. Natl. Acad. Sci.
Настоящая работа не содержит описания ис-
USA 87 (7), 2461 (1990).
следований с использованием людей и животных
в качестве объектов.
28. C. Diez-Villasenor, C. Almendros, and J. Garcia-Mar-
tinez, Microbiology 156 1351 (2010).
29. К. С. Пугач, A. В. Лопатина и К. В. Северинов,
СПИСОК ЛИТЕРАТУРЫ
Молекуляр. биология 46 (2), 195 (2012).
1. А. Ю. Переварюха, Биофизика 66 (2), 384 (2021).
30. Н. А. Жигалова, С. В. Женило, А. В. Артемов и
2. А. Ф. Ванин, А. В. Пекшев, А. Б. Вагапов и др.,
Е. Б. Прохорчук, Молекуляр. биология 51 (5), 836
Биофизика 66 (1), 183 (2021).
(2017).
3. Б. И. Кузник, В. Х. Хавинсон и В. С. Смирнов,
31. В. Г. Ильичев, Биофизика 57 (2), 356 (2012).
Успехи геронтологии 33 (6), 1032 (2020).
32. А. Б. Медвинский, Биофизика 51 (6), 1033 (2006).
4. А. Ф. Ванин, Биофизика 65 (4), 818 (2020).
33. А. Е. Бобыpев, В. А. Буpменcкий, Е. А. Кpикcунов
5. A. Goyal, D. B. Reeves, and E. Cardozo-Ojeda, eLife
и др., Биофизика 55 (6), 1138 (2010).
10, e63537 (2021).
34. G. E. Hutchinson, Ann. N. Y. Acad. Sci. 50, 221
6. А. Ю. Переварюха, Биофизика 65 (1), 138 (2020).
(1948).
БИОФИЗИКА том 66
№ 6
2021
1162
ПЕPЕВАPЮXА
35. V. V. Mikhailov and Yu. S. Reshetnikov, Informatsion-
61. Г. И. Марчук, А. А. Романюха и Г. А. Бочаров, в сб.
no-Upravliaiushchie Sistemy, No 4, 31 (2018).
Математические вопросы кибернетики (Наука, М.,
1989), вып. 2, сс. 5-70.
36. А. И. Михайлов, А. Е. Бобырев, Т. И. Булгакова и
А. Д. Шереметьев, Журн. общ. биологии 80 (6), 418
62. В. В. Когай, Т. М. Хлебодарова, С. И. Фадеев и
(2019).
В. А. Лихошвай, Вычисл. технологии 20 (1), 38
(2015).
37. А. Ю. Переварюха, Журн. общ. биологии 81 (3), 174
(2020).
63. G. Ximing and S. E. Ford, Philos. Trans. Roy. Soc. B:
Biol. Sci. 371 (1689), 206 (2016).
38. S. Ruan, in Delay Differential Equations and Applica-
tions (Springer, Berlin, 2006), pp. 477-517.
64. A. Y. Karatayev, L. E. Burlakova, and D. K. Padilla,
Hydrobiologia 746, 97 (2015).
39. A. Y. Perevaryukha, Cybernetics and Systems Analysis
52 (4), 623 (2016).
65. A. N. Frolov, Entomol. Rev. 95 (2), 147 (2015).
40. R. M. May, Stability and Complexity of Ecosystems
66. A. Y. Perevaryukha, Entomol. Rev. 95 (3), 397 (2015).
(Princeton University Press, 1974).
67. N. Hanley and M. Roberts, People and Nature 1, 124
41. J. Van Dyken, J. Theor. Biol. 460 (1), 115 (2019).
(2019).
42. W. S. Gurney, S. P. Blythe, and R. M. Nisbet, Nature
68. M. Cooling, S. Hartley, D. A. Sim, P. J. Lester, Biol.
287, 17 (1980).
Lett. 8 (2), 430 (2012).
43. A. J. Nicholson, Aust. J. Zool. 2 (1), 9 (1954).
69. W. I. Choi and Y.-S. Park, Insect Sci. 19 (5), 535
(2012).
44. А. Ю. Колесов и Н. Х. Розов, Тр. мат. института
им. Стеклова 277 101 (2012).
70. T. Andersen, J. Carstensen, E. Hernández-Garcia, and
C. M. Duarte, Trends Ecol. Evol. 24 (1), 49 (2009).
45. А. Ю. Колесов и Н. Х. Розов, Мат. сб. 202 (6), 51
(2011).
71. L. R. Clark, Austral. J. Zool. 12 (2), 150 (1964).
46. G. Oster and J. Guckenheimer, in The Hopf Bifurcation,
72. A. Y. Perevaryukha, J. Comput. Systems Sci. Int. 60
Еd. by J. Marsden and M. McCracken (Springer-Ver-
(1), 87 (2021).
lag, Berlin, 1976), pp. 327-345.
73. M. J. Grieshop, P. W. Flinn, and J. R. Nechols, J. In-
47. E. M. Wright, J. für die Reine und Angewandte Math-
sect Science 10 (1), 99 (2010).
ematik 194 (1), 66 (1955).
74. А. В. Pуcаков, А. Б. Медвинcкий, Б. Л. Ли и
48. S. A. Kashchenko, Automat. Control Comput. Sci. 47
М. М. Гоник, Биофизика 54 (4), 733 (2009).
(3), 470 (2013).
75. Б. В. Адамович, Р. З. Ковалевская, Н. П. Радчикова
49. C. J. Krebs and J. H. Myers, Adv. Ecol. Res. 8 (2), 267
и др., Биофизика 60 (4), 769 (2015).
(1974).
76. H. Wan and H. P. Zhu, Math. Biosci. Engineer. 11,
50. T. Royama, W. E. Mackinnon, and E. G. Kettela, Eco-
1395 (2014).
logy 86 (5), 1212 (2005).
77. Т. Ю. Борисова и И. В. Соловьева, Мат. машины и
системы, № 1, 71 (2017).
51. A. Y. Perevaryukha, Cybernetics and Systems Analysis
55 (1), 141 (2019).
78. О. А. Иванов, Вестн. СПбГУ. Математика. Меха-
ника. Астрономия 6 (3), 422 (2019).
52. M. Zubrik, A. Hajek, and D. Pilarska, J. Appl. Ento-
mol. 140 (8), 565 (2016).
79. V. V. Mikhailov and A. V. Spesivtsev, Studies Comput.
53. L. R. Clark, Austral. J. Zool. 12 (3), 362 (1964).
Intelligence 868, 449 (2020).
80. D. L. Strayer, M. D'Antonio, M. S. Fowler, and
54. T. N. Solov’eva, Information and Control Systems 4,
J. Geist, Ecol. Lett. 20, 1337 (2017).
60 (2016).
81. A. A. Berryman, in Encyclopedia of Entomology
55. K. Gopalsamy, Stability and Oscillations in Delay Dif-
(Springer, Dordrecht, 2008).
ferential Equations of Population Dynamics (Springer,
Netherlands, 1992).
82. K. Aagaard and J. L. Lockwood, Biol. Invasions 18,
1667 (2016).
56. С. Д. Глызин, Моделирование и анализ информа-
ционных систем 3, 365 (2017).
83. А. Е. Бобыpев, В. А. Бурменский, Е. А. Криксунов
и др., Биофизика 58 (2), 334 (2013).
57. T. L. Kaplan and T. A. Yorke, J. Math. Analysis and
Applications 48 (2), 317 (1974).
84. И. В. Арженовский и А. В. Дахин, Регионология 28
(3), 470 (2020).
58. V. A. Dubrovskaya and I. V. Trofimova, J. Belarus.
State University. Biology 3, 76 (2017).
85. A. Smith and A. P. Smith, Sci. Rep. 6, 38703 (2016).
59. G. Bocharov and F. Rihan, J. Comput. Appl. Math.
86. Ю. С. Букин, Вавиловский журн. генетики и се-
125 (2), 183 (2000).
лекции 18 (2), 320 (2014).
60. K. P. Hadeler and G. Bocharov, Canad. Appl. Math.
87. B. La Scola, C. Desnues, and I. Pagnier, Nature 455,
Quarterly 11 (2), 159 (2003).
100 (2008).
БИОФИЗИКА том 66
№ 6
2021
МОДЕЛИРОВАНИЕ СЦЕНАРИЕВ ГЛУБОКОГО ПОПУЛЯЦИОННОГО КРИЗИСА
1163
88. D. Raghunathan, P. M. Radford, and C. Gell, Sci. Rep.
93. Г. Р. Хасанова, И. Г. Мустафин и В. А. Анохин,
9 4293 (2019).
Эпидемиология и инфекционные болезни, № 3, 47
(2012).
89. P. T. H. Nguyen, O. V. Pettersson, and P. Olsson, Eur.
J. Plant Pathol. 127, 73 (2010).
94. А. В. Яцишин, А. Ю. Переварюха и В. В. Михай-
лов, Мат. машины и системы, № 2, 90 (2019).
90. В. В. Кузик, Т. Ю. Борисова и П. П. Гераскин,
Журн. эволюц. биохимии и физиологии 56 (7), 623
95. М. А. Цыганов и Г. Р. Иваницкий, Биофизика 51
(2020).
(6), 1008 (2006).
91. M. Pohlschroder and S. V. Albers, FEMS Microbiol.
96. А. Ю. Переварюха, Динамические системы 7 (2),
Rev. 42 (6), 719 (2018).
157 (2017).
92. W. E. Schrank and N. Roy, Marine Resource Econom-
97. A. V. Nikitina, V. M. Kozlov, and A. A. Filina, Comput.
ics 28 397 (2013).
Math. Information Technol. 1 (1), 35 (2019).
Simulation of Scenarios of the Deep Population Crisis
of the Rapidly Growing Population
A.Yu. Perevaryukha
St. Petersburg Federal Research Center, Russian Academy of Sciences, 14-ya Linia 39, St. Petersburg, 199178 Russia
The article is devoted to the modeling of threshold development of the population process during the forma-
tion of a new population in a competitive environment. Invaders with high reproductive activity often aggres-
sively affect autochthonous biosystems. In the process of settling invaders in the area, a sharp turn on of biotic
countermeasures until the exhaustion of resources for further growth in numbers can cause a deep population
crisis for a new species. Many populations went through the bottleneck stage, including humans at the time
of the Neolithic crash in Europe. Invaders with high reproductive potential in many cases have deleterious
effects on biosystems. The emergence of effective competition can not only cause classical cyclical fluctua-
tions, but also lead to the complete extinction of the population after a series of high peaks in its abundance.
Two alternative scenarios are classical samples of situations of an induced population crisis. First, G. Gause
conducted experiments with an introduction of predatory ciliates and two ciliates species were driven to ex-
tinction. Second, a series of experiments were carried out with bacteriophages that were introduced into col-
onies of actively dividing bacteria with a dynamically adapting antiviral mechanism. We have proposed mod-
ifications of the model for scenarios which are enough to constitute a crisis in population dynamics. In equa-
tions with deviating arguments in the time variable, a threshold level for activating the impact of biotic
conditions on the reproduction of invasive species and an aggregated nature of the lagging regulation with two
time factors are taken into account. The computational scenarios described both the effect of the completion
of the process after a repeated outbreak, and the successful passage of the population crisis due to rapid ad-
aptation. Deep crisis phenomena are characteristic of local population dynamics when organisms interact
with viruses that are new to them.
Keywords: invasion scenarios, modeling of extreme population dynamics, biotic resistance, lagged threshold regu-
lation, species extinction, Neolithic crash, biological pest control, immune response, CRISPR-Cas9
БИОФИЗИКА том 66
№ 6
2021