БИОФИЗИКА, 2022, том 67, № 1, с. 73-81
БИОФИЗИКА КЛЕТКИ
УДК 573.22
ИЗМЕНЕНИЕ ПОКАЗАТЕЛЯ ИНТЕГРИРОВАННОЙ ИНФОРМАЦИИ
ДЛЯ АКТИВНОСТИ НЕЙРОНОВ ГИППОКАМПА В ПРОЦЕССЕ
ФОРМИРОВАНИЯ ОПЕРАНТНЫХ НАВЫКОВ У КРЫС
© 2022 г. И.А. Нажёсткин*, О.Е. Сварник*, **
*Московский физико-технический институт (национальный исследовательский университет),
117303, Москва, ул. Керченская, 1а/1
**Институт психологии РАН, 129366, Москва, ул. Ярославская, 13/1
E-mail: olgasvarnik@gmail.com
Поступила в редакцию 29.07.2021 г.
После доработки 16.08.2021 г.
Принята к публикации 30.08.2021 г.
Одним из подходов к оценке сложности систем является теория интегрированной информации, ко-
торая предлагает математические средства для подсчета уровня интеграции системы и была перво-
начально разработана для вычисления уровня сознания. Данный подход был использован для оцен-
ки изменения активности нейронных популяций гиппокампа в процессе приобретения навыков
разной сложности. Установлено, что с увеличением успешности выполнения навыка наблюдается
увеличение показателя интегрированной информации Φ. При этом показатель интегрированной
информации изменяется с периодичностью в трое-четверо суток до достижения животным резуль-
тата. Полученные результаты свидетельствуют в пользу того, что временная дезорганизованность
нейронной активности является необходимым этапом возникающей организации нейронной ак-
тивности, способствующей достижению результата целым организмом.
Ключевые слова: самоорганизация, сложные системы, обучение, теория интегрированной информации,
мозг, нейрон.
DOI: 10.31857/S0006302921060069
В основе способности живых существ к обуче-
теория интегрированной информации, которая
нию или приобретению новой информации ле-
изначально была предложена для оценки уровня
жат перестройки активности нейронных сетей, в
сознания [2-4]. Поскольку предполагается, что
частности, головного мозга. Известно, что актив-
сознание связано с интеграцией разного рода ин-
ность отдельных нейронов может существенно
формации, степень интеграции может отражать
изменяться на различных временных шкалах как
тот уровень, на котором сейчас находится созна-
в реальных сетях мозга, так и in vitro [1]. Такие из-
ние.
менения проявляются и на уровне отдельных
Основная идея теории интегрированной ин-
нейронов, и на уровне популяций клеток, и на
формации заключается в том, чтобы оценить,
уровне когнитивных процессов, например, при
сколько информации способна произвести систе-
колебаниях субъективного восприятия.
ма и насколько она способна к ее интеграции, т.е.
Известно, что в процессе приобретения ново-
к получению целостной картины на основе мно-
го опыта наблюдается ряд закономерностей ней-
жества частных элементов [5]. Оценка системы
ронного уровня. Так, начальные этапы научения
производится, во-первых, на основании числа от-
характеризуются увеличением частоты нейрон-
личающихся друг от друга состояний, в которых
ной активности и вариативности этой активно-
она может находиться (генерируемое системой
сти, оцениваемой по разным параметрам. Однако
количество информации). Чем больше число раз-
головной мозг представляет собой совокупность
личающихся состояний, тем больше информация.
огромного числа сложных самоорганизующихся
Во-вторых, оценка производится на основании
нейронных сетей, и для оценки процессов, позво-
оценки зависимости частей системы друг от друга.
ляющихся живым организмам осуществлять раз-
Система должна иметь способность к интеграции
нообразные адаптации, необходимы подходы,
информации, т.е. ее части должны быть взаимо-
хотя бы до некоторой степени учитывающие эту
связанными, зависимыми друг от друга. Незави-
сложность. Одним из таких подходов может стать
симые компоненты системы практически беспо-
73
74
НАЖЁСТКИН, СВАРНИК
лезны, так как несмотря на то, что они могут зако-
ринта с помощью генетического алгоритма. Ин-
дировать большое количество состояний, не
тегрированную информацию рассчитывали для
получится сделать из них какой-либо вывод. В ка-
параметров робота, изменяемых во время обуче-
честве яркой иллюстрации вышесказанного авто-
ния. С ходом обучения наблюдался рост Φ, кроме
ры теории приводят аналогию со светочувстви-
того, наблюдалась его корреляция с оценкой ка-
тельной матрицей фотоаппарата - состояние каж-
чества выполнения задачи (зависела от расстоя-
дого пикселя матрицы независимо от других, и,
ния до цели в лабиринте и количества успешных
несмотря на то, что матрица способна закодиро-
прохождений).
вать огромное количество различных изображе-
Таким образом, теория интегрированной ин-
ний, она неспособна сделать какие-либо выводы,
формации может служить эффективным инстру-
неспособна провести даже простейший анализ
ментом для изучения механизмов обработки ин-
изображения. Только в таком случае - если взаи-
формации в головном мозге в процессе адапта-
мосвязь и взаимозависимость частей системы до-
ции к изменяющимся условиям среды. Однако до
статочно велики - система способна к наилучшей
настоящего времени такого рода исследования не
реакции на внешние события и адаптации к ним,
проводились, что связано прежде всего с техниче-
поскольку способна сделать общий вывод по со-
скими трудностями учета активности больших
стояниям множества своих компонентов. Основ-
популяций нейронов. В настоящей работе прове-
ной метрикой, численно выражающей вышеска-
дена оценка показателя Φ для небольших нейрон-
занное, в данной теории является коэффициент
ных групп гиппокампа экспериментальных жи-
интегрированной информации Φ, показывающий
вотных на разных этапах приобретения простого
разность между количеством информации, произ-
и сложного навыков.
веденным целой системой, и суммой ее невзаимо-
действующих частей. Говоря другими словами,
МАТЕРИАЛЫ И МЕТОДЫ
данный коэффициент показывает, насколько
Вычисление коэффициента интегрированной ин-
уменьшится информация, производимая систе-
формации. По определению, данному авторами,
мой, в случае, если разделить систему на части. В
коэффициент Φ рассчитывается как [10, 11]:
предельном случае, если взаимодействия между
r
частями системы нет, коэффициент Φ равен ну-
Φ
=
H(M
|M
)
−
H
(X
|
X
),
лю, поскольку разделение системы на части ниче-
t,k
t+Δt,k
t
t+Δt
k=1
го не изменит - и вместе, и независимо они будут
генерировать такое же количество информации, т.
где H(A|B) - условная энтропия (энтропия пере-
е. общее число их возможных состояний будет та-
менной A при наличии переменной B), X0, X1 -
ким же, и разность выйдет нулевой. По мнению
векторы состояния системы в моменты времени t
некоторых исследователей, коэффициент Φ пока-
и t + Δt (см. ниже), M0,k, M1,k - подвыборки из си-
зывает, «что система представляет собой изнутри»
стемы в моменты времени t и t + Δt, r - количе-
(what the system is), а не «что система, согласно на-
ство подвыборок. Данное математическое выра-
блюдениям, делает» (what the system does) [6, 7].
жение отражает ранее сказанное во введении -
Кроме изначальной цели в виде выделения со-
математическую разность между количеством
знательных состояний, параметр интегрирован-
информации, производимым разделенной систе-
ной информации успешно применялся для ис-
мой, и производимым всеми ее частями. Количе-
следования роли астроцитов в нейронных сетях
ство информации, генерируемое исходной (не-
in silico [8], а также для оценки поведения других,
разделенной) системой, всегда меньше, так как в
более простых систем. Так, в работе [9] коэффи-
силу ограничений, накладываемых взаимосвязя-
циент Φ рассчитывали для поведения команд лю-
ми компонентов, система способна находиться в
дей и группы редакторов статей в Wikipedia.
меньшем числе состояний. Подвыборки выбира-
Была выявлена корреляция между значением Φ и
ются наиболее независимыми, т.е. таковыми, что
классической метрикой успеха для каждой из си-
разделение системы на них дает минимальное
стем - коллективным интеллектом для групп лю-
увеличение информации по сравнению с целой
дей и качеством статей для редакторов. В работе
системой. Таким образом, выбирается наиболее
[10] был рассчитан коэффициент интегрирован-
репрезентативное разделение, выделяющее наи-
ной информации для виртуальных роботов
более независимые части системы. Более точно
(animats), обучаемых задаче прохождения лаби-
данное разделение определяется как:
r
H(M
|M
)
−
H
(X
|
X
),
t,k
t+Δt,k
t
t+Δt
k=1
MIP
=
argmin
,
(1)
M
(r
−1)minH
(M
)
max
0,k
k
где MIP (minimum information partition) - иско- системы в моменты времени t и t + Δt, r - количе-
мое разбиение, Mt,k и Mt+Δt, k - подвыборки из ство подвыборок, а Hmax(Mt,k) - максимальная
БИОФИЗИКА том 67
№ 1
2022
ИЗМЕНЕНИЕ ПОКАЗАТЕЛЯ ИНТЕГРИРОВАННОЙ ИНФОРМАЦИИ
75
энтропия подвыборки в момент времени t, кото-
стрированных нейронов в данной сессии было
рая равна размеру подвыборки. Нормализация на
меньше 15, все они использовались для анализа и
r -
1
min
H
Mt
в поиске минимума необхо-
никакой отбор не производился [9].
(
)
max
(
,k
)
k
дима для исключения влияния размеров подвы-
Для вычисления Φ системы необходимо ука-
борок на получающуюся условную энтропию
зать, что является узлами, а также определить со-
[11]: например, в случае, если система будет раз-
стояния узлов в каждый момент времени. В слу-
бита на две части - большую и малую - то у тако-
чае нейронов головного мозга, узлами являются
го разбиения уменьшение энтропии почти всегда
нейроны, а состояния узлов определяются следу-
будет меньше, чем для равных частей. В итоге бу-
ющим образом. Все время разбивается на отрезки
дет найдено такое разбиение, которое выделит в
(бины) равного размера. В случае если в бине i-й
системе части, связь между которыми наиболь-
нейрон произвел спайк, то считается, что в мо-
шим образом влияет на интеграцию информации
мент времени i узел (нейрон) имел состояние 1, а
в ней.
если спайка не было, то считается, что узел имел
Основная проблема данного метода заключа-
состояние 0.
ется в том, что разбиение на подвыборки может
Размер бина выбирается исходя из компро-
быть найдено только полным перебором. Коли-
мисса, чтобы не допустить потери тонких паттер-
чество разбиений на подвыборки всех возможных
нов нейронной активности и в то же время не уве-
размеров для сети из N узлов выражается числом
личить размер данных, передаваемых алгоритму,
Белла BN. Это число растет быстрее экспоненты;
до такой степени, что вычисление займет недопу-
так, для N = 5 BN = 52, для N = 10 BN = 115975, а
стимо большое время. Исходя из визуального
для N = 20 BN = 51724158235372 [12]. Из-за ограни-
анализа спайковых временных данных, был вы-
бран размер бина 0.06 с.
ченных вычислительных ресурсов компьютеров в
данной работе для анализа использовали только
Еще одной проблемой является то, что энтро-
15 нейронов. Для проведения вычислений отби-
пии, входящие в выражение (1), требуют нор-
рали 15 нейронов с наибольшей вариативностью
мального распределения входных данных [13].
активности (с наибольшей дисперсией распреде-
Для тех данных, чье распределение неизвестно
ления Бернулли), поскольку в случае слабо меня-
или отличается от нормального, существует дру-
ющих свою активность нейронов алгоритм рабо-
гой алгоритм, так называемое авторегрессионное
тает (см. ниже). В случае если количество зареги-
Φ (autoregressive Φ, ΦAR):
det(Σ(X))
2
det(Σ(M
))
1
1
k
min ln
-
ln
X
M
M
k=1
k
2
det Σ(E
))
2
det Σ(E
))
Φ
AR
=
,
L(M)
где (X) - ковариационная матрица для пере-
единицу меньшее число нейронов и повторяли
менной X; EX - остатки (ошибки предсказания)
вычисление.
линейной регрессии, предсказывающей значение
После вычисления полученное значение ΦAR
переменной X в момент времени 1 на основании
нормировали на количество нейронов, фактиче-
значения X в момент времени 0; Mk, k = 1, 2 - две
ски участвовавших в расчете. Это было сделано
подвыборки системы (разбиение системы на две
для обеспечения возможности сравнения резуль-
M
k
части),
L
M
=
1lnmin
2πe
det
Σ
M
-
татов, полученных на различном числе нейронов.
(
)
{
(
)
(
(
k
))
}
k
2
норми-ровочный коэффициент. Подвыборки
Одним из параметров алгоритма является вре-
выбираются так, чтобы значение ΦAR по-
менной промежуток (сдвиг) Δt для вычисления
лучилось минимально возможным [13].
взаимной информации. Коэффициент Φ из опре-
деления (1) показывает разность в информации,
В некоторых случаях алгоритм не мог вернуть
генерируемой системой за некоторое количество
правильное значение. Известно [9], что такое по-
шагов (последовательных состояний). Данное ко-
ведение может происходить, когда узлы сети ме-
личество зависит от конкретной системы и опре-
няют свою активность слишком редко. В данном
случае это означает, что нейрон был почти всегда
деляется тем, через сколько шагов в системе про-
неактивен или, наоборот, имел слишком боль-
исходят значимые изменения. На практике проме-
шую частоту спайков и активировался почти в
жуток определяется перебором
- выбирается
каждом бине. В таком случае заново отбирали на
таковым, чтобы при нем было наибольшим [9].
БИОФИЗИКА том 67
№ 1
2022
76
НАЖЁСТКИН, СВАРНИК
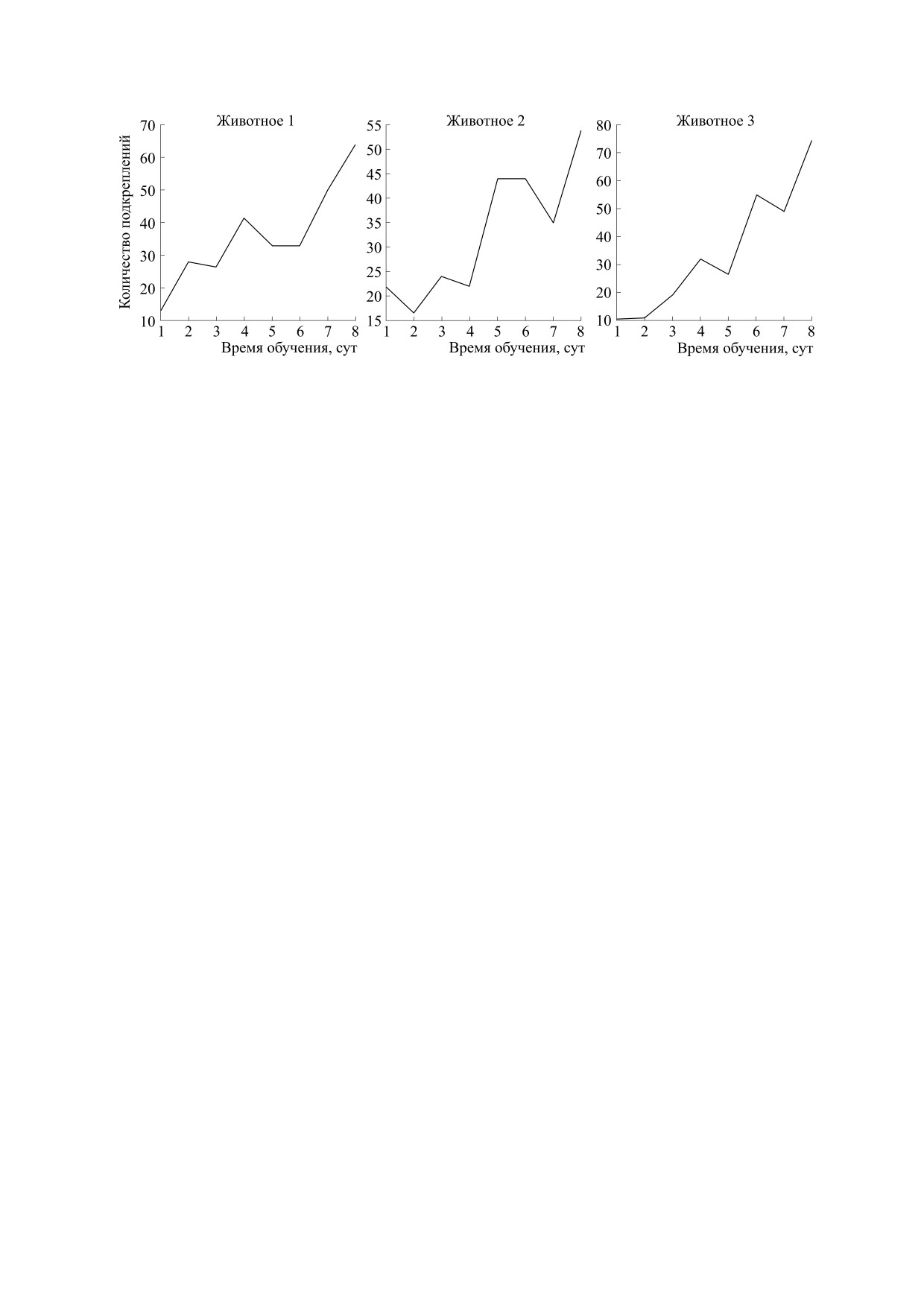
Рис. 1. Кривые обучения для трех животных. Ось абсцисс - время обучения в сутках; ось ординат - количество под-
креплений (выдач пищевого вознаграждения).
С целью установления зависимости между Φ и
вознаграждения следовало перейти из конца од-
параметрами обучения для каждого животного
ного рукава в конец другого по определенным
были рассчитаны следующие метрики:
правилам [15].
- мгновенная скорость обучения - производ-
Обучение проводили в течение восьми дней. В
ная кривой обучения (рассчитывалась численно,
течение каждого дня проводили одну или две обу-
с использованием разностной схемы);
чающих сессии по 15-20 мин. В течение всего
времени обучения проводили непрерывную реги-
- общая скорость обучения - разность между
страцию нейронной активности животного и ви-
количеством вознаграждений в последний и пер-
вый день, деленная на число дней;
деозапись его поведения. В каждой сессии
было зарегистрировано от 15 до 50 нейронов (в
- общая скорость изменения интегрирован-
среднем - 31 нейрон).
ной информации - разность между значениями
Φ в последний и первый день, деленная на число
Значение интегрированной информации Φ
дней.
вычислялось для каждой сессии обучения, а затем
усреднялось по дню. Также рассчитывалось зна-
Для вычислений был использован открытый
чение Φ для воvсьми равных по продолжительно-
код в среде MATLAB (MathWorks Inc., США) [9].
сти периодов, на которые делилось время каждой
Весь анализ данных и поведения был выполнен с
сессии.
помощью программ в среде MATLAB. Статисти-
ческий анализ проводили в программе GraphPad
Обучение животных (простой навык). Для дан-
Prism (GraphPad Software Inc., США).
ного вида обучения также был использован набор
открытых данных [17, 18]. Кратко: крысы (n = 4)
Обучение животных (оперантное многодневное
выполняли различные задачи в линейном или
обучение). Для анализа изменений параметра ин-
круговом лабиринте. Каждое животное выполня-
тегрированной информации в ходе многодневно-
ло одну или несколько задач. Перед каждой сес-
го формирования оперантного навыка были ис-
сией в лабиринте и после нее проводили сессию
пользованы открытые данные [14-16]. Наиболее
отдыха в домашней клетке (~4 ч). Микроэлектро-
просты в проведении и практически отработаны
ды были также имплантированы в область CA1
за много лет исследований эксперименты с про-
гиппокампа (AP - 4.0 мм, ML - 2.4 мм). Время
странственным гиппокамп-зависимым обучени-
каждой сессии (45 мин) в лабиринте делили на
ем в лабиринте. Поэтому в настоящей работе бы-
восемь равных периодов, после чего значение Φ
ли использованы открытые данные именно с та-
вычисляли для каждого периода.
ким типом обучения. Крысы линии Long-Evans
(n = 3) были обучены навыку ориентирования в
W-образном лабиринте с целью получения жид-
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
кой пищевой награды. Сначала крысы были при-
Многодневное последовательное обучение слож-
учены к поилке с наградой), после чего проводи-
ному оперантному навыку. Все животные успешно
лась операция по имплантации массива из 21 не-
приобрели требуемый навык за семь дней (рис. 1).
зависимо управляемых микроэлектрода в область
Наибольшая средняя скорость обучения наблю-
CA1 правого гиппокампа (координаты: -3.6 мм
далась у третьего животного, наименьшая - у вто-
AP, 2.2 мм ML), и после реабилитации (5-7 суток)
рого. То же самое можно сказать и о максимально
начиналось обучение. Животным для получения
достигнутом количестве подкреплений.
БИОФИЗИКА том 67
№ 1
2022
ИЗМЕНЕНИЕ ПОКАЗАТЕЛЯ ИНТЕГРИРОВАННОЙ ИНФОРМАЦИИ
77
Рис. 2. (а) - Зависимости коэффициента интегрированной информации Φ от времени обучения при различных сме-
щениях по времени Δt. Ось абсцисс - время обучения в сутках; ось ординат - величина коэффициента Φ. Различные
кривые соответствуют различным значениям Δt. Видно, что наиболее яркая зависимость наблюдается при Δt = 14 би-
нов (2.8 с) у первого животного, Δt = 13 бинов (2.6 с) у второго и Δt = 14 бинов (2.8 с) у третьего. (б) - Зависимости
амплитуды кривой показателя Φ в наиболее характерной точке от смещения по времени Δt. Пунктирной линией по-
казано максимальное значение.
Перед выполнением анализа поведения коэф-
Результаты зависимости Φ от дня обучения для
фициента Φ был выполнен его расчет для различ-
каждого животного при оптимальном Δt пред-
ных значений смещения по времени (Δt). Далее
ставлены на рис. 3. Как видно, интегрированная
определяли, где значения Φ больше и где его из-
информация для всех животных с ходом обучения
изменялась периодически, с периодом в трое-
менения по дням сильнее выражены (рис. 2). Бы-
четверо суток.
ло выяснено, что оптимальным является интер-
вал Δt = 14 бинов (2.8 с) для первого животного,
С целью проведения более подробного анали-
Δt = 13 бинов (2.6 с) для второго и Δt = 14 бинов
за каждая сессия была разбита на восемь перио-
для третьего. Дальнейший анализ проводили
дов, одинаковых по времени. В каждом из перио-
только для Φ, найденных при оптимальном зна-
дов было рассчитано значение Φ при оптималь-
чении Δt.
ном временном сдвиге Δt, а также количество
БИОФИЗИКА том 67
№ 1
2022
78
НАЖЁСТКИН, СВАРНИК
Рис. 3. Зависимости коэффициента интегрированной информации Φ от времени обучения при оптимальном значе-
нии Δt.
вознаграждений. В каждом отдельно взятом пе-
ном обучении сложному навыку наблюдались
риоде характерных закономерностей обнаружено
колебания параметра Φ с периодом трое-четверо
не было. Между количеством вознаграждений и
суток, что согласуется с ранее полученными ре-
значением Φ в каждом периоде была обнаружена
зультатами, свидетельствующими о присутствии
статистически значимая взаимосвязь (рис. 4).
регрессии в успешности во время обучения [23].
Также была установлена статистически значимая
Таким образом, оказалось, что изменения по-
корреляция параметра интегрированной инфор-
казателя интегрированной информации соответ-
мации с успешностью реализации формирую-
ствуют изменению поведения животных с тече-
щейся программы поведения.
нием приобретения сложного навыка. Такие
периодические изменения успешности выполне-
Был определен временной промежуток, в тече-
ния навыков, когда после ряда успешных проб
ние которого в нейронных сетях происходит наи-
отмечается спад успешности, отмечаются в лите-
более значимая обработка и интеграция инфор-
ратуре постоянно (см., например, работу [19]).
мации. Было выявлено, что он приблизительно
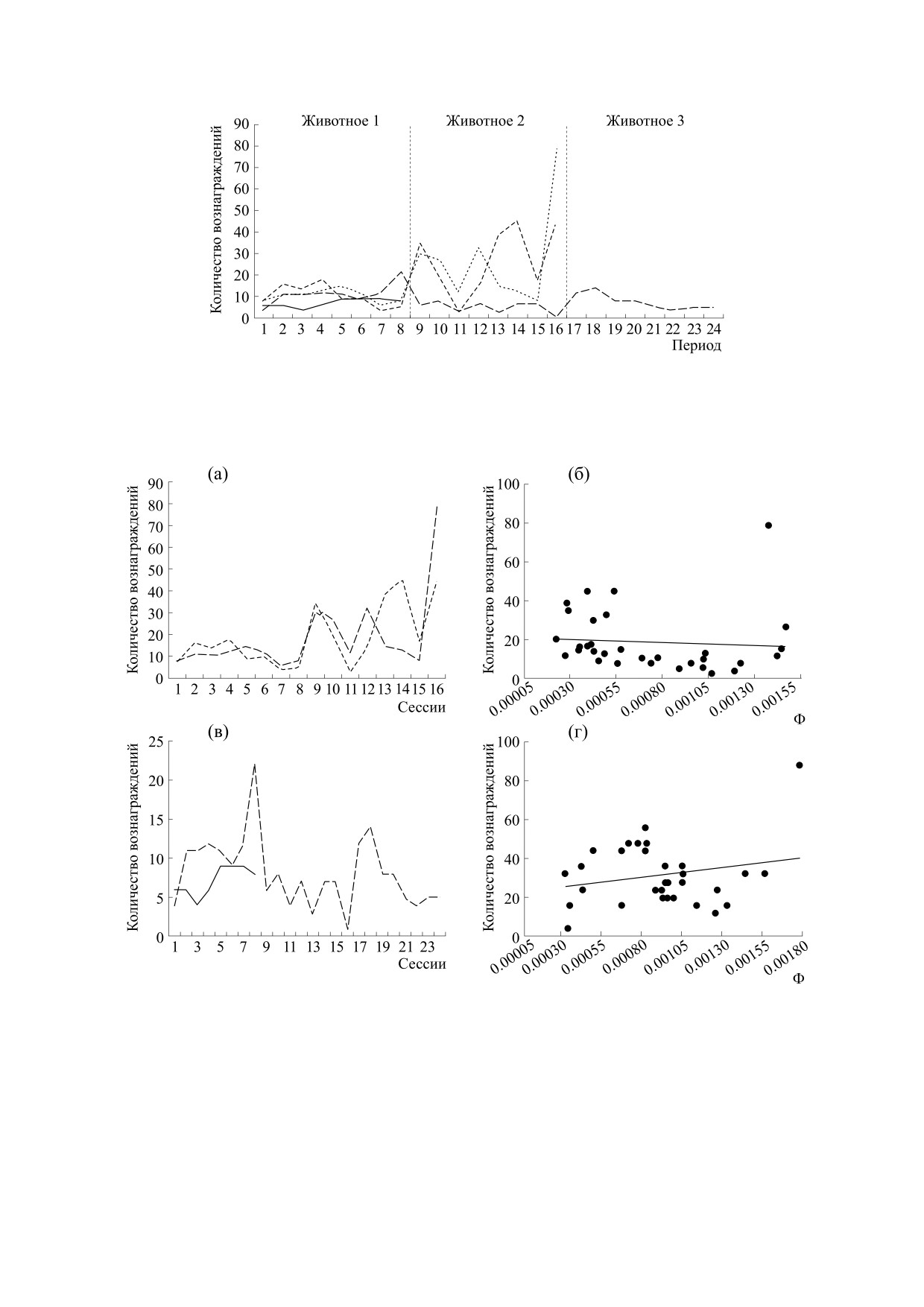
Обучение простому навыку. Двое из четырех
одинаков у различных животных и составляет
животных демонстрировали прогресс с ходом
2.6-2.8 с. Это время по порядку сравнимо со вре-
обучения (рис. 5).
менем одного вознаграждаемого поведенческого
Для животных, имевших прогресс в обучении,
акта.
наблюдали статистически значимую отрицатель-
Таким образом, результаты настоящего иссле-
ную корреляцию коэффициента Φ и количества
дования показывают, что параметр интегриро-
подкреплений в сессии (рис. 6а,б). Для живот-
ванной информации Φ может стать новым марке-
ных, которые не демонстрировали прогресс, ста-
ром, который дает возможность количественно
тистически значимая взаимосвязь не наблюда-
оценить изменения функциональной связанно-
лась (рис. 6в,г).
сти нейронов в сложной системе при обучении.
Вместе с тем проведенное исследование позволя-
ЗАКЛЮЧЕНИЕ
ет наметить подходы к вопросу о том, что именно
оценивает показатель интегрированной инфор-
Для анализа закономерностей изменения ней-
мации на уровне когнитивных процессов. По-
ронной активности гиппокампа при формирова-
скольку первоначально теория интегрированной
нии сложных и простых навыков у крыс была
информации создавалась для оценки уровня со-
применена теория интегрированной информа-
знания, настоящее исследование ставит вопрос о
ции [5]. Известно, что нейроны демонстрируют
том, можно ли говорить о колебаниях уровня со-
специфическую связь с поведением (см., напри-
знания при приобретении нового навыка.
мер, работы [20-22]), однако для понимания
закономерностей осуществления успешного
В будущем возможно проведение более глубо-
приобретения нового опыта требуются подходы,
ких исследований с использованием других набо-
позволяющие провести глобальную оценку со-
ров данных, полученных в разнообразных усло-
стояния нейронных групп. При последователь-
виях. Возможна и работа с данными, полученны-
БИОФИЗИКА том 67
№ 1
2022
ИЗМЕНЕНИЕ ПОКАЗАТЕЛЯ ИНТЕГРИРОВАННОЙ ИНФОРМАЦИИ
79
Рис. 4. (а) - Коэффициенты интегрированной информации Φ во всех периодах за весь ход обучения для трех живот-
ных. Нижняя ось абсцисс - номер периода (в хронологическом порядке), верхняя ось абсцисс - время обучения в сут-
ках; ось ординат - значение Φ в данном периоде. Вертикальные линии разделяют сутки обучения. (б) - Количество
подкреплений в каждом периоде для трех животных. Ось абсцисс - номер периода (в хронологическом порядке), ось
ординат - значение Φ в данном периоде. (в) - Взаимосвязь между величиной Φ и количеством подкреплений в каж-
дом из периодов. Ось абсцисс - значение Φ; ось ординат - количество вознаграждений в данном периоде. Животное
1: коэффициент корреляции Спирмена r = 0.4697, 95%-й доверительный интервал [0.2993, 0.6110], p = 0.0001 (критерий
Стьюдента). Критерий Стьюдента используется для оценки значимости корреляции. Животное 2: r = 0.2791, 95%-й
доверительный интервал [0.09142, 0.4477], p = 0.0041. Животное 3: r = 0.3196, 95%-й доверительный интервал [0.1296,
0.4869], p = 0.0009.
ми с помощью электроэнцефалографии и
исследований в рамках научного проекта № 20-
подобных методов, как, например, в работе [24].
34-90023.
ФИНАНСИРОВАНИЕ РАБОТЫ
КОНФЛИКТ ИНТЕРЕСОВ
Исследование выполнено при финансовой
Авторы заявляют об отсутствии конфликта
поддержке Российского фонда фундаментальных интересов.
БИОФИЗИКА том 67
№ 1
2022
80
НАЖЁСТКИН, СВАРНИК
Рис. 5. Прогресс обучения навыку по периодам. Ось абсцисс - периоды 1/8 сессии (в хронологическом порядке), ось
ординат - количество вознаграждений. Вертикальные линии разделяют сессии.
Рис. 6. (а) - Кривые обучения (количество вознаграждений за день) для животных, продемонстрировавших прогресс
в обучении. Различные кривые относятся к различным животным. Оформление кривых соответствует оформлению
на рис. 5. (б) - Коэффициенты интегрированной информации Φ и количество подкреплений в экспериментальных
сессиях для животных, продемонстрировавших прогресс в обучении. Ось абсцисс - значения Φ, ось ординат -
количество подкреплений (успешных поведенческих актов); p = 0.0155, r = -0.4243. (в) - Кривые обучения
(количество вознаграждений за день) для животных, не продемонстрировавших прогресс в обучении. (г) -
Коэффициент интегрированной информации Φ и количество подкреплений в экспериментальных сессиях для
животных, не продемонстрировавших прогресс в обучении. Ось абсцисс - значения Φ, ось ординат - количество
подкреплений (успешных поведенческих актов); p = 0.6225, r = -0.09046.
БИОФИЗИКА том 67
№ 1
2022
ИЗМЕНЕНИЕ ПОКАЗАТЕЛЯ ИНТЕГРИРОВАННОЙ ИНФОРМАЦИИ
81
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
13. A. B. Barrett and A. K. Seth, PLoS Comput. Biol. 7 (1),
e1001052 (2011).
Все применимые международные, националь-
ные и институциональные принципы ухода и ис-
14. Sh. P. Jadhav and L. M. Frank, Simultaneous extracel-
пользования животных при выполнении работы
lular recordings from hippocampal area CA1 and medial
были соблюдены.
prefrontal cortex from rats performing a W-track alterna-
tion task (2020). DOI: 10.6080/K02N50G9
15. Sh. P. Jadhav, G. Rothschild, D. K. Roumis, and
СПИСОК ЛИТЕРАТУРЫ
L. M. Frank, Neuron 90 (1), 113 (2016).
1. S. Reinartz, Adv. Neurobiol. 22, 331 (2019).
16. W. Tang, J. D. Shin, L. M. Frank, and Sh. P. Jadhav, J.
2. A. G. Casali, O. Gosseries, M. Rosanova, et al., Sci.
Neurosci. 37 (49), 11789 (2017).
Transl. Med. 5 (198), 198ra105 (2013).
17. A. D. Grosmark, J. Long, and G. Buzsáki, Recordings
3. M. T. Alkire, A. G. Hudetz, and G. Tononi, Science
from hippocampal area CA1, PRE, during and POST
322 (5903), 876 (2008).
novel spatial learning (hc-11 data set) (2016). DOI:
10.6080/K0862DC5
4. J. R. King, J. D. Sitt, F. Faugeras, et al., Curr. Biol. 23
(19), 1914 (2013).
18. Zh. Chen, A. D. Grosmark, H. Penagos, and
M. A. Wilson, Sci. Reports 6 (1), 1 (2016).
5. G. Tononi, Arch. Ital. Biol. 150 (2/3), 56 (2012).
6. L. Albantakis and G. Tononi, Entropy 17 (8), 5472
19. M. H. Erdelyi, Am. Psychol. 65 (7), 623 (2010).
(2015).
20. M. P. Karlsson and L. M. Frank, J. Neurosci. 28 (52),
7. T. Niizato, K. Sakamoto, Yoh-ichi Mototake, et al.,
14271 (2008).
PloS One 15 (2), e0229573 (2020).
21. M. A. Wilson and B. L. MsNaughton, Science 261
8. O. Kanakov, S. Gordleeva, A. Ermolaeva, et al., Phys.
(5124), 1055 (1993).
Rev. E 99 (1), 012418 (2019).
22. R. Q. Quiroga, L. Reddy, G. Kreiman, et al., Nature
9. D. Engel and T. W. Malone, PLoS One 13 (10),
435 (7045), 1102 (2005).
e0205335 (2018).
23. Ю. И. Александров, О. Е. Сварник, И. И. Знаменская
10. J. A. Edlund, N. Chaumont, A. Hintze, et al., PLoS
и др., Вопросы психологии, № 4, 87 (2017).
Comput. Biol. 7 (10), e1002236 (2011).
24. K. Fujii, H. Kanazawa, and Y. Kuniyoshi, in Abstr.
11. D. Balduzzi and G. Tononi, PLoS Comput. Biol. 4 (6),
Book of the Joint IEEE 9th Int. Conf. on Development and
e1000091 (2008).
Learning and Epigenetic Robotics (ICDL-EpiRob)
12. G. C. Rota, Am. Math. Monthly 71 (5), 498 (1964).
(IEEE, 2019), pp. 137-142.
Variations in the value of Integrated Information Theory Estimated from Hippocampal
Neural Activity during Acquisition of Operant Behavior in Rats
I.A. Nazhestkin* and O.E. Svarnik*, **
*Moscow Institute of Physics and Technology, Kerchenskaya ul. 1a/1, Moscow, 117303 Russia
**Institute of Psychology, Russian Academy of Sciences, Yaroslavskaya ul. 13/1, Moscow, 129366 Russia
One of the approaches to quantifying the complexity of systems is the integrated information theory, as a
mathematically complicated measure of how much "integrated information" a system possesses and has been
initially proposed to calculate how much consciousness a neural system has. In this study, we used this ap-
proach to assess whether there are any differences in hippocampal neural activity during skill acquisition of
various degrees of difficulty. It was found that the value of integrated information index Φ increased with the
task progress. Moreover, Φ-value changes with an interval of three-four days until the animal is able to per-
form the skill. The results obtained attest to the fact that the temporary abnormal neural activity (the tempo-
rary disorganization of the system’s state) is an essential stage in reorganization of neural activity contributing
to the achievement of a goal for the whole body.
Keywords: self-organization, complex systems, learning, integrated information theory, brain, neuron
БИОФИЗИКА том 67
№ 1
2022