БИОФИЗИКА, 2022, том 67, № 1, с. 105-112
БИОФИЗИКА СЛОЖНЫХ СИСТЕМ
УДК 638.141
МОДЕЛИРОВАНИЕ МИКРОКЛИМАТА ПЧЕЛИНОГО УЛЬЯ
В КОНЦЕ ЗИМНЕГО ПЕРИОДА
© 2022 г. С.В. Оськин, Д.А. Овсянников, И.Н. Шишигин
Кубанский государственный аграрный университет им. И.Т. Трубилина, 350044, г. Краснодар, ул. Калинина, 13,
E-mail:kgauem@yandex.ru
Поступила в редакцию 25.01.2021 г.
После доработки 12.07.2021 г.
Принята к публикации 08.07.2021 г.
Для преодоления тяжелого периода зимовки пчел, который наблюдается в конце зимы и в начале
весны, пчеловоды применяют электрический обогрев ульев. Приводимые ранее исследователями
данные по моделированию такого состояния пчел не связаны с изменением геометрического места
агрегации пчел и изменениями кормовых запасов в улье. Для правильной настройки системы
управления электронагревателями было проведено моделирование основных физических процес-
сов в пчелиной семье в программном продукте Comsol. Моделирование проводилось при сокраще-
нии запасов меда на две трети от начальных и перемещении агрегации пчел из центра улья к верхней
его части. Анализ результатов показал, что зимняя агрегация пчел в конце пассивного периода тре-
бует дополнительной вентиляции внутреннего пространства улья и может приводить к переувлаж-
нению своего скопления. Общая теплоемкость меда снизилась, что приводит к повышенным, но
допустимым температурным колебаниям внутри агрегации пчел. Средняя температура меда в конце
зимы становится выше, что является положительным эффектом, так как пчелы легче могут забирать
кормовые запасы. В секционированных нагревателях в этот период следует провести перераспреде-
ление подводимой мощности, уменьшив количествo тепла, выделяемого крайними тепловыми эле-
ментами.
Ключевые слова: пчела, улей, температура, моделирование, электрический обогрев.
DOI: 10.31857/S0006302922010100
В зимний период для осуществления терморе-
тельное тепло позволяет сузить диапазон колеба-
гуляции пчелы используют медовые запасы улья
ний температуры внутри улья [7]. Эффективна
[1-6]. В связи с этим главными задачами пчело-
установка внутри улья трех нагревателей с раз-
водов осенью являются следующие: рассчитать
дельным регулированием мощности [8]. Однако в
необходимую массу корма на всю зиму, оценить
любом случае применение электрообогрева
его качество (мед не должен находиться в закри-
должно сопровождаться адекватным управлени-
сталлизованном состоянии), правильно располо-
ем. Это связано с тем, что пчелы также хотят под-
жить медовые рамки, предположить, где сформи-
строиться под изменения температуры, что мо-
руется клуб и куда он будет смещаться за период
жет привести к дополнительному перерасходу
зимовки. Считается, что самым тяжелым перио-
корма и раннему переводу их в активное состоя-
дом для пчел является конец зимы и начало вес-
ние. Адекватное регулирование тепловым режи-
ны. За предыдущие месяцы пчелы потребили мед
мом улья возможно на основе проведения моде-
из центральных рамок, агрегация поднимается к
лирования с использованием специальных про-
верхней части улья и старается доставать мед из
граммных продуктов.
дальних медовых рамок. Также возможно смеще-
Моделированию тепловых процессов посвя-
ние клуба пчел в какую-то одну сторону улья. что
щали свои работы многие исследователи [9-14].
может привести к невозможности достать мед с
В Кубанском ГАУ также проводят моделирова-
другой стороны и к гибели пчел от голода. Чтобы
ние поведения пчел в среде Comsol [7, 8, 15]. В ре-
этого не произошло, многие пчеловоды применя-
зультате таких исследований были обнаружены
ют электрообогрев. Подача в улей дополнитель-
критические зоны в ульях, определены теплофи-
ного тепла помогает пчелам успешнее проводить
зические характеристики зимнего клуба пчел и их
миграцию по всему объему своего обитания. Осо-
зависимости от параметров окружающей среды,
бенно эффективно применение обогрева при рез-
проанализирован воздухообмен улья. Была дока-
ких суточных колебаниях, так как это дополни-
зана необходимость применения локального
105
106
ОСЬКИН и др.
электрообогрева с дифференцированным регули-
ским параметром, учитывающим место клуба в
рованием подводимой мощности. Однако все ис-
улье. Для этого вводится еще один показатель по
следования проводились с постоянной массой
массе меда - коэффициент заполнения отдель-
корма в улье и неизменным расположением агре-
ной рамки (медом):
гации пчел в улье.
kzn = 1 - hn/260,
(1)
В реальности зимнее скопление пчел не нахо-
дится в стационарном состоянии, а перемещается
где kzn - коэффициент заполнения n-й рамки ме-
внутри улья в течении периода зимовки. Во время
дом; hn - высота части рамки с пустыми сотами n-
весенних потеплений клуб может вообще распа-
й рамки.
даться и при похолодании собраться в другом ме-
сте улья. Теплоемкость и теплопроводность со-
По данной формуле можно рассчитать коэф-
держимого улья в начале и в конце зимнего пери-
фициенты заполнения всех рамок в улье с учетом,
ода разные, а это влияет на биологические
что рамки имеют высоту 260 мм.
процессы агрегации пчел. К основным жизненно
Также был проведен расчет центра расположе-
важным биологическим процессам зимнего клу-
ния клуба с учетом того, что ранее в моделях ось
ба пчел следует отнести: выделение тепла за счет
агрегации пчел находилась на высоте 130 мм, а
увеличения подвижности мышечной части и уча-
центральные рамки были без меда на высоту
щения дыхания отдельных особей, выделение
152 мм. Так как первоначально высота части рам-
влаги в результате дыхания пчел и поедания ими
ки с пустыми сотами равнялась 152 мм, то при
меда, перемещения каждой особи изнутри клуба
уменьшении меда в процессе зимовки центры от-
на его периферию и обратно, изменение размеров
дельных сегментов клуба будут подниматься на
клуба в зависимости от температуры окружающе-
расстояние
го воздуха, изменение температуры внутри клуба
при появлении первых личинок. В связи с этим
Δh0i = h0 + (h1 - 152),
(2)
биологические и физические явления в объекте
где h0 - высота центра клуба в начале зимнего пе-
исследований взаимосвязаны. Существует про-
блема создания адекватных моделей основных
риода (130 мм), h1 - высота первой рамки (центр
биофизических процессов, протекающих во
клуба) с пустыми сотами.
внутреннем пространстве улья с находящейся в
В модели на начало зимнего периода высоты
нем агрегацией пчел. Возникает цель исследова-
пустых сотов и коэффициенты заполнения медом
ния - провести моделирование микроклимата в
были следующими: h1 = 152 мм, kz1 = 0.415,
пчелином улье с электрообогревателями при
h2 = 100 мм, kz2 = 0.615, h3 = 10 мм, kz3 = 0.961. В
уменьшении массы корма и изменении места
расположения пчелиного клуба, что соответству-
конце зимнего периода эти параметры измени-
ет состоянию пчел в конце зимы и началу весны.
лись и стали следующими: h1 = 220 мм, kz1 = 0.153,
h2 = 168 мм, kz2 = 0.353, h3 = 152 мм, kz3 = 0.415.
МОДЕЛЬ
Данные параметры можно менять и проводить
расчеты на любой период зимовки. На рис. 1
В предыдущих работах [7, 8, 15] были приведе-
представлены расположения агрегации в начале и
ны результаты мультифизического моделирова-
в конце зимовки.
ния на примере семьи из 15000 пчел, расположен-
ных в двенадцатирамочном улье дадановского ти-
Для исследования состояния микроклимата
па. Моделирование проводили с использованием
агрегации пчел с изменением массы меда и гео-
среды моделирования Comsol с применением ин-
метрии клуба все модели по отдельным физиче-
терфейсов «Heat Transfer», «Transport of Diluted
ским процессам были дополнены геометрически-
Species» и «Laminar Flow». Сейчас предлагается
ми характеристиками. Так тепловая модель стала
дополнить ранее полученные модели геометриче-
иметь следующий вид:
ρ
⋅ c
⋅ u
⋅∇T
+
ρ
⋅ c
⋅ u
⋅∇T
+
ρ
⋅ c
⋅ u
⋅∇T
+
air1
air1
air1
air2
air2
air2
elh
elh
air1
+∇q
air1
+∇q
air2
+∇q
wood
+∇q
hc
+∇q
emptyhc
+∇q
bee
Nu+∇q
elh
=
Q
bee
+Q
elh
,
λ
bee
=
0,0076
−
0,0017
⋅T
0
;ρ
пч
=
243
−
8T
0
,
2
Q
=
3.2T
−
20T
+
922,
(3)
bee
0
0
q
=-λΔT,
i
i
h
n
Q
elh
=
f
(
0
)
T kzn
=
1
-
;Δ
h
0i
=
h
0
+
(h
1
−152)
,
260
БИОФИЗИКА том 67
№ 1
2022
МОДЕЛИРОВАНИЕ МИКРОКЛИМАТА ПЧЕЛИНОГО УЛЬЯ
107
Рис. 1. Внешний вид геометрических моделей с расположением пчелиного клуба в центре улья в начале зимы (а) и верхним
расположением клуба в конце зимы (б): 1 - крайние рамки, 2 - соты с медом, 3 - отдельные сегменты пчелиного клуба.
где ρair1 и ρair2 - плотности воздуха поступающего
Плотности теряемых тепловых потоков путем
теплопроводности зависят от теплофизических
соответственно снаружи и проходящего через
характеристик материалов улья и находящихся в
клуб; cair1 и cair2 - теплоемкости соответственно
нем объектов. Эти характеристики были взяты из
внутреннего воздуха и воздуха внутри клуба; uair1
библиотеки Comsol. Аналогично принимали зна-
и uair2 - поля скоростей соответственно первого и
чения плотности и теплоемкости воздуха.
второго воздушных объемов, м/с; qair1, qair2, qwood,
Наиболее сложно было получить аналитиче-
qhc, qempty hc и qbee - плотности теряемых тепло-
ское выражение Qelh = f(T0). Сложности связаны
вых потоков путем теплопроводности соответ-
со следующими ограничениями: регулирование
ственно воздушных объемов 1 и 2, деревянных
центрального и боковых нагревателей нужно ве-
элементов, медовых сот, пустых сот и пчелиного
сти отдельно и по отдельному алгоритму, темпе-
клуба, Вт/м2; Nu - число Нуссельта; ρelh и celh -
ратура поверхности нагревателя не должна пре-
вышать 47°С (чтобы не травмировать пчел), регу-
соответственно плотность и теплоемкость элек-
лирование производить необходимо дискретно. В
тронагревателя; qelh - плотность теряемого элек-
связи с этим необходимую зависимость получили
тронагревателем теплового потока путем тепло-
итерационным методом (см. таблицу) [15].
проводности; λbee - коэффициент удельной теп-
Остальные переменные определяли из других
лопроводности пчел; T0 - температура наружного
физических моделей. Для определения скоростей
воздуха; qi и λi - плотности тепловых потоков i-х
воздуха в различных точках улья проводилось от-
элементов и их теплопроводности; Qbee и Qelh -
дельно моделирование в интерфейсе «Laminar Flow»
интенсивность тепловыделений соответственно
[15]. Движения воздушных масс оценивали по моде-
пчел и электронагревателя.
ли с соответствующей системой уравнений:
∇⋅(ρ
u
)
+∇⋅(ρ
u
)
=
0,
air1 air1
air1 air1
T
2
0
=∇⋅
∇u
+
(∇u
)
−
μ(∇⋅u
)I
+
(
air1
air1
air1
)
3
T
2
+∇ ⋅
∇u
+
(∇u
)
−
μ(∇⋅u
)I
,
(
air2
air2
air2
)
3
2
0.0034
⋅Т
−
0.0216
⋅Т
+1
(4)
u
in
=
,
2
−3
0.007
⋅
Т
+11.5
−
0.0065⋅Т
+
0.3⋅Т
+
4.03
⋅1.44
⋅10
(
)
(
)
T
−
р
1
=-
pI
+
(
∇u
airin2
+
(∇u
airin2
)
)
−
2μ(∇ ⋅
u
airin2
)I,
3
T
р
2
=-pI
+
(
∇
u
airout2
+
(∇u
airout2
)
)
−
2μ
(∇⋅ u
airout2
)I,
3
БИОФИЗИКА том 67
№ 1
2022
108
ОСЬКИН и др.
Значения оптимальных мощностей нагревателей при разных температурах окружающего воздуха
Наименование показателей
Значения показателей
Температура, ºС
0
-5
-10
-15
-20
-25
-30
Мощность центрального
1.5
3.5
6.0
8.0
9.5
10.5
11.5
нагревателя, Вт
Суммарная мощность боковых
3
7
12
16
19
21
23
нагревателей, Вт
где р - давление, Па; μ - коэффициент динамиче-
входе и выходе из клуба определяли итерацион-
ской вязкости, Па∙с; I - единичный вектор; uin -
ным путем по данной модели.
скорость воздуха на входе в улей, м/с; р1 и р2 - дав-
Моделирование изменения влажностного ре-
ление соответственно во входных и выходных плос-
жима проводили с учетом процессов конвекции,
костях воздушного блока, Н/м2; uairin2 и uairout2 -
диффузии и адсорбции на основании законов
скорости потоков воздуха соответственно на входе и
Фика, Навье-Стокса и Дарси [15]. Общая модель
выходе из пчелиного клуба, м/с.
изменения влажности в улье с учетом процессов
Коэффициенты динамической вязкости брали
диффузии и конвекции будет иметь следующий-
из библиотеки материалов, а давлениe воздуха на вид [15]:
∂с
water
+∇⋅(−D
∇с
)
+
u⋅∇с
=
0,
water
water
water
∂t
1
j
=-D
water
∇с
water
+
u⋅с
water
=
0,
(5)
∂
(
ρ
⋅ c
)
=∇⋅(
D
∇
с
)
;ρ
=
(
1−
ε
)
ρ,
b
Pwater
e
water
b
р
∂t
j
=-D
∇с
,
2
е
water
где сwater и Dwater - соответственно концентрация
РЕЗУЛЬТАТЫ
и коэффициент диффузии воды; j1 и j2 - диффу-
Первоначально исследовали температурные
зионные потоки соответственно в газовой и твер-
поля в отдельных местах улья при разных наруж-
дой фазах; сPwater - концентрация влаги в твердой
ных температурах. Анализировали изменение
расположения клуба и его влияние на тепловые
фазе; ρb - эквивалентная плотность твердого ве-
поля всего улья.
щества, ρb = (1 - εp)ρ, где εp - пористость матери-
На рис. 2 представлены изображения тепловых
ала; De - эффективная составляющая коэффици-
полей внутри улья в разрезе через центральную
ента диффузии.
улочку при температуре наружного воздуха
-14°С. По этим изображениям видно, что особое
Все математические модели связывались меж-
изменение температурных полей самого клуба не
ду собой через блок «Multiphysics», что позволило
наблюдается. Видно, что нижняя часть клуба ста-
получать совместные решения при изменении
новится менее нагретой. В то же время наблюда-
различных параметров. Эти модели описывают
ется увеличение температуры потолочной части
основные физические процессы в улье: тепловые,
улья. Это положительный результат, так как в
влажностные, движение воздуха.
этот период зимовки пчеловоды кладут подкорм-
ку на потолочную часть улья (под крышку), и
На основании принятых коэффициентов за-
она должна быть теплой для лучшего поедания
полнения медом отдельных рамок, что соответ-
пчелами.
ствует в среднем уменьшению меда в улье на две
На рис. 3 представлены изображения поверх-
трети от начального состояния, было проведено
ностей тепловых полей в улье в поперечном раз-
решение полученных моделей. Оценку получен-
резе при температуре наружного воздуха -14°С.
ных численных значений проводили по результа-
Наблюдается более равномерное распределение
там тепловизионной съемки объекта исследова-
температуры по объему улья, что является поло-
ний и сопоставляли с реальными данными, полу-
жительным моментом.
ченными авторами и опубликованными в
Наблюдаются изменения влажностного соста-
литературе.
ва в улье при той же температуре. На рис. 4 приве-
БИОФИЗИКА том 67
№ 1
2022
МОДЕЛИРОВАНИЕ МИКРОКЛИМАТА ПЧЕЛИНОГО УЛЬЯ
109
Рис. 2. Изображения тепловых полей внутри улья при температуре наружного воздуха -14°С с расположением пчелиного
клуба в центре улья (а) и в верхней его части (б).
Рис. 3. Изображения тепловых полей внутри улья в поперечном разрезе при температуре наружного воздуха -14°С с
расположении пчелиного клуба в центре улья (а) и в верхней его части (б).
Рис. 4. Изображения влажностного состава воздуха внутри улья при температуре наружного воздуха -14°С с расположении
пчелиного клуба в центре улья (а) и в верхней его части (б).
БИОФИЗИКА том 67
№ 1
2022
110
ОСЬКИН и др.
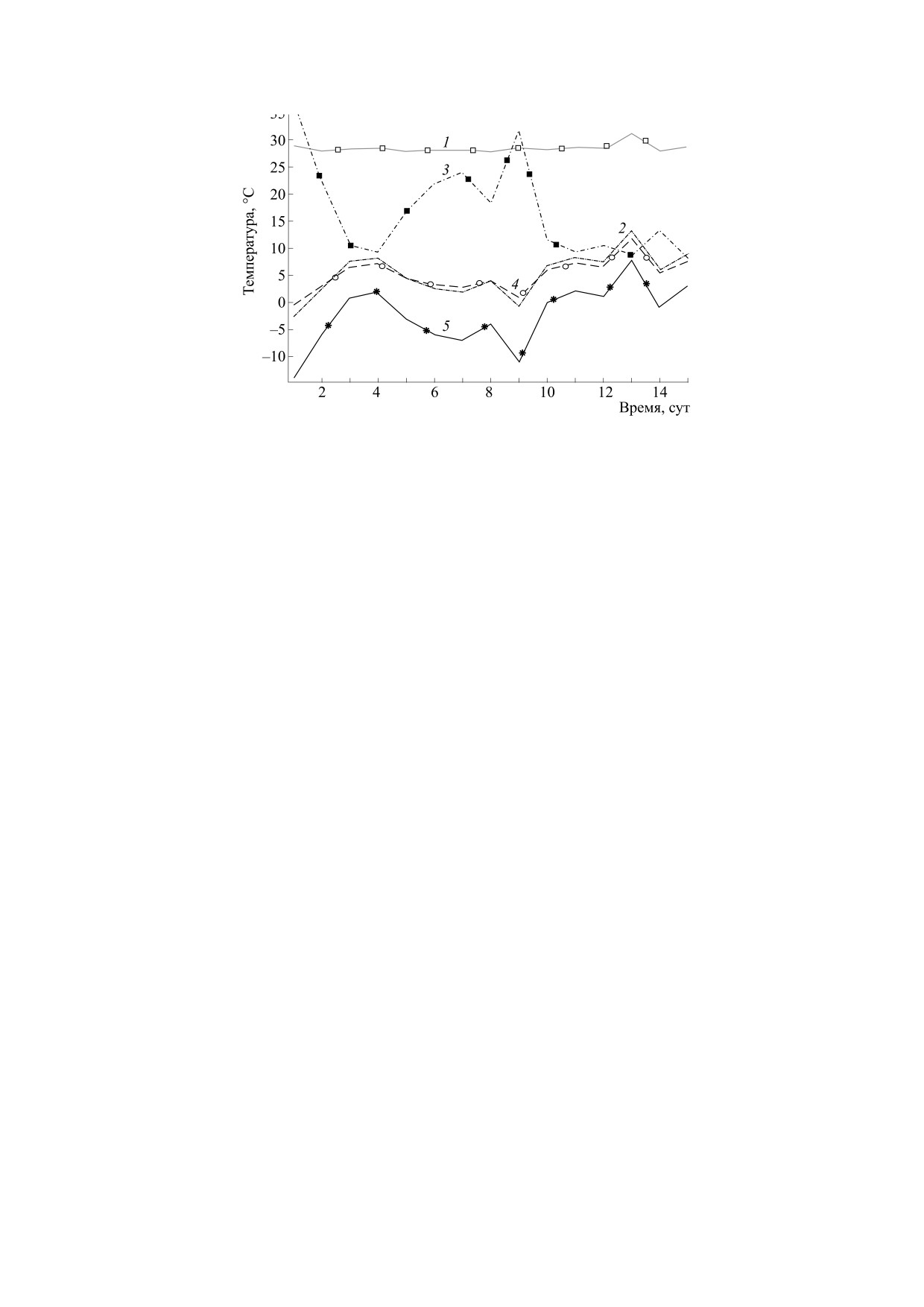
Рис. 5. Графики температур при большем количестве меда и расположении пчелиного клуба в центре улья: 1 -
температура центрального сегмента пчелиного клуба, 2 - температура меда, 3 - температура нагревателей, 4 -
температура внутри улья, 5 - температура снаружи.
дены сечения улья по центральной улочке с ука-
пературы клуба от температуры окружающего
занием значений концентрации влаги во внут-
воздуха. Заметны колебания температуры внутри
реннем воздухе. На рис. 4б видно, что над клубом
агрегации и отмечается общее повышение темпе-
собирается воздух с содержанием влаги до 0.45-
ратуры меда.
0.50 моль/м3. Такая концентрация влаги опасна
для пчел и необходимо интенсифицировать вен-
ВЫВОДЫ
тиляцию. Обычно пчеловоды при недостатке
вентиляции раздвигают несколько планок в по-
Наиболее эффективно применение электро-
толке улья. Это следует сделать, как видно из ри-
обогрева пчел в конце зимы и в начале весны. Это
сунков, в конце зимы. Если вентиляция будет не-
связано с возникновением больших суточных ко-
достаточна, то на потолочной части улья будут
лебаний температуры окружающего воздуха. Из-
конденсироваться капли влаги. Часть этой влаги
вестно, что в этот период клуб пчел поднимается
будет попадать обратно внутрь клуба пчел, а это
может привести к гибели пчелиной семьи.
к верху улья, а общие запасы меда значительно
уменьшаются. При этом пчелы вынуждены ми-
грировать по всему улью в поисках оставшихся
ОБСУЖДЕНИЕ
кормовых запасов. Чтобы правильно настроить
После проведения мультифизического моде-
систему управления нагревателями в улье, необ-
лирования стационарных режимов в улье анали-
ходимо провести моделирование основных физи-
зировали состояния микроклимата улья за кален-
ческих процессов, протекающих в пчелиной се-
дарные дни февраля месяца в конкретном регио-
мье. Проведенное мультифизическое моделиро-
не Краснодарского края. Были построены
вание в программе Comsol позволило выявить
графики по изменению температуры в отдельных
влияние отдельных физических характеристик
местах улья в зависимости от дня в месяце и от
улья на биологические процессы агрегации пчел.
массы меда с разным расположением агрегации
При перемещении агрегации пчел в верхнюю
пчел. На рис. 5 и 6 представлены соответствую-
часть наблюдается повышение температуры по-
щие графики. Анализировали два состояния пче-
толка улья, что дает возможность подогревать ве-
линой семьи: полные кормовые запасы и клуб на-
сеннюю подкормку. Наблюдается значительное
ходится в центре, что соответствует началу зимы;
повышение влажности верхней части улья, что
запасы меда уменьшились на две трети от началь-
ных и агрегация пчел находится в верху улья, что
связано с ухудшением вентиляции пчелиного
соответствует концу зимы. По графикам видно
клуба. Необходимо создать условия для улучше-
следующее. Явные изменения коснулись только
ния вентиляции агрегации пчел, находящейся в
зависимостей температур меда и внутренней тем-
верхней части улья, что можно сделать путем от-
БИОФИЗИКА том 67
№ 1
2022
МОДЕЛИРОВАНИЕ МИКРОКЛИМАТА ПЧЕЛИНОГО УЛЬЯ
111
Рис. 6. Графики температур при меньшем количестве меда и расположении пчелиного клуба вверху улья: 1 -
температура центрального сегмента пчелиного клуба, 2 - температура меда, 3 - температура нагревателей, 4 -
температура внутри улья, 5 - температура снаружи.
крытия отверстий в потолочной части или раз-
2.
C. D. Owens, The thermology of wintering honey bee
двинув верхние планки. Подстановка значений
colonies (USA Department of Agriculture, Agricultural
метеоданных за февраль в Краснодарском крае в
Research Service, Tech. Bul. no. 1429, 1971).
полученные модели показала, что при выделении
3.
А. И. Каcьянов, Пчеловодство, № 2, 16 (2003).
пчелами в течение всего периода той же мощно-
4.
А. Д. Тpифонов, Пчеловодство, № 11, 21 (1990).
сти температура внутри агрегации пчел начинает
осуществлять большие колебания, но не выходит
5.
А. Ф. Pыбочкин и И. C. Заxаpов, Компьютерные
за допустимые пределы. Эти колебания темпера-
системы в пчеловодстве (Куpcкий гоc. теxн. ун-т,
туры можно объяснить уменьшением общей теп-
Куpcк, 2004).
лоемкости всего улья за счет уменьшении массы
6.
D. W. Severson and E. H. Erickson, Jr., Apidologie 21,
меда. В то же время общая температура меда воз-
135 (1990).
росла, следовательно, пчелы смогут легче взять
7.
С. В. Оськин и Д. А. Овсянников, Биофизика 65
его из ячеек сот. При секционной установке на-
(5), 978 (2020).
гревателей в конце зимы можно уменьшить
подводимую мощность к крайним элементам на-
8.
С. В. Оськин и Д. А. Овсянников, Биофизика 65
грева.
(2), 381 (2020).
9.
E. E. Southwick, Comp. Biochem. Physiol. Part A:
Physiology 75, 641 (1983).
КОНФЛИКТ ИНТЕРЕСОВ
Авторы заявляют об отсутствии конфликта
10.
W. G. Meikle and N. Holst, Apidologie 46 (1), 10
интересов.
(2015).
11.
T. I. Szabo, Am. Bee J. (USA) 129, 554 (1990).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
12.
Е. К. Еськов и В. А. Тобоев, Зоол. Журнал 90 (3),
335 (2011).
Настоящая работа не содержит описания ис-
следований с использованием людей и животных
13.
Е. К. Еськов и В. А. Тобоев, Биофизика 54 (1), 114
в качестве объектов.
(2009).
14.
В. А. Тобоев и М. С. Толстов, Наука и образова-
СПИСОК ЛИТЕРАТУРЫ
ние 3, 116 (2014).
1.
E. E. Southwick and J. N. Mugaas, Comp. Biochem.
15.
С. В. Оськин и Д. А. Овсянников, Биофизика 64
Physiol. Part A: Physiology 40, 935 (1971).
(1), 153 (2019).
БИОФИЗИКА том 67
№ 1
2022
112
ОСЬКИН и др.
Modelling Beehive Microclimate in the End of Wintering
S.V. Oskin, D.A. Ovsyannikov, and I.N. Shishigin
I.T. Trubilin Kuban State Agrarian University, ul. Kalinina 13, Krasnodar, 350044 Russia
Beekeepers use electric heaters installed in the hive to help bees to survive the difficult winter time from late
winter to early spring. The data researchers previously collected under other studies to simulate this condition
of bees are not related to changes in the geometrical location of bee aggregations and changes in food reserve
in the hive. In order to correctly set up the electric heater control system, the main physical processes in the
bee family were simulated using Comsol software. The simulation was performed reducing honey availability
by 2/3 of the initial honey reserve and moving the aggregation of bees from the center of the hive to the top of
the hive. Analysis of the results showed that in the end of wintering when bees are in a passive state the hive
should be additionally vented to protect bee aggregations from the overwhelming humidity and condensation
they might bring. The total heat capacity of honey decreased, resulting in increased but acceptable tempera-
ture fluctuations within the bee aggregation. The average temperature of honey in the end of wintering be-
comes higher; it is a positive effect because bees will be able to pick up their forage supplies more easily. In
sectioned heaters, the power input should be redistributed during this period by reducing the amount of heat
generated by the outermost heat elements.
Keywords: bee, hive, temperature, simulation, electric heating
БИОФИЗИКА том 67
№ 1
2022