БИОФИЗИКА, 2022, том 67, № 2, с. 274-282
БИОФИЗИКА КЛЕТКИ
УДК 573
НАРУШЕНИЕ РЕГУЛЯЦИИ СИСТЕМЫ цАМФ В ОБОНЯТЕЛЬНЫХ
НЕЙРОНАХ В МОДЕЛИ ШИЗОФРЕНИИ, ИНДУЦИРУЕМОЙ
ВВЕДЕНИЕМ (+)-МК-801 КРЫСАМ
© 2022 г. Е.В. Бигдай, А.А. Синегубов
Институт физиологии им. И.П. Павлова РАН, 199034, Санкт-Петербург, наб. Макарова, 6
E-mail: artem_sinegubov@aol.com
Поступила в редакцию 06.12.2021 г.
После доработки 06.12.2021 г.
Принята к публикации 19.01.2022 г.
Одним из ключевых патогенетических звеньев шизофрении является нарушение хода развития
нервной системы. Сходные изменения наблюдаются в рамках модели данной патологии, индуци-
руемой введением антагонистов NMDA-рецепторов на ранних стадиях развития. В работе проде-
монстрировано, что в данной модели шизофрении проявляются функциональные изменения в од-
ном из крупных очагов постнатального нейрогенеза - обонятельном нейроэпителии. Обонятель-
ные сенсорные нейроны модельных животных обладают сниженной чувствительностью к
активатору аденилатциклазы и нарушениями регуляции ее активности со стороны кальмодулин-за-
висимых сигнальных путей.
Ключевые слова: обоняние, шизофрения, нейрогенез, NMDA
DOI: 10.31857/S0006302922020107
Шизофрения - сложное психическое заболе-
висимых транскрипционных факторов в меха-
низме развития алкоголизма [2].
вание, клеточные и молекулярные субстраты ко-
торого до конца не изучены. Современные пред-
Исследования на трансгенных животных уста-
ставления о патофизиологии этого заболевания
новили, что цАМФ-сигнализация важна для обу-
особое внимание уделяют глутаматэргической
чения и памяти, как для ее приобретения, так и
системе, в первую очередь связанной с функцио-
для потери контекстной памяти [3]. Сигнальный
нированием NMDA-рецепторов. Накопленные
путь цАМФ вовлекается в регуляцию нейрогене-
данные позволяют говорить о данной патологии
за, выживаемости нейронов и синаптической
как о болезни развития нервной системы, при ко-
связи, которые участвуют в патогенезе депрессии
торой ведущим звеном патогенеза является ано-
[4].
мальный ход нейрогенеза [1].
Синтез цАМФ изменен при ряде нейропсихи-
Циклический аденозинмонофосфат (цАМФ)
ческих расстройств, включая шизофрению. Так, в
играет очень важную роль в различных типах кле-
тромбоцитах у пациентов с шизофренией обнару-
ток, включая нейроны, поскольку влияет на раз-
жено снижение его продукции [5]. При исследо-
витие клеточного цикла, и при повышении его
вании остроты обоняния у пациентов с шизофре-
внутриклеточного содержания подавляет мито-
нией с помощью одорантов, трансдукция кото-
генную передачу сигналов через фосфорилирова-
рых вовлекает внутриклеточный цАМФ, было
ние протеинкиназой A фактора Raf. Доказана
показано, что внутриклеточные сигнальные пути
роль циклического аденозинмонофосфата в ме-
цАМФ разрегулированы при шизофрении, и раз-
ханизме формирования наркотической зависи-
регулированная цАМФ-сигнализация может
мости, обусловленной аномальной его регуляци-
вносить вклад в патогенез шизофрении [6].
ей в нейронах коры головного мозга, полосатого
При использовании вестерн-блот-анализа и
тела и голубого пятна. Показана роль цАМФ-за- посмертных образцов из хорошо охарактеризо-
ванной коллекции пожилых пациентов с шизо-
Сокращения: цАМФ - циклический аденозинмонофосфат, френией было показано, что белки и фосфопро-
PDE - фосфодиэстераза, ACIII - аденилатциклаза III,
теины, содержащие митоген-активированную
ОСН - обонятельный сенсорный нейрон, ЦНС
-
центральная нервная система, IBMX - 3-изобутил-1-
протеинкиназу и цАМФ-связанные сигнальные
метилксантин, СаМ - кальмодулин.
пути аномально экспрессируются в передней по-
274
НАРУШЕНИЕ РЕГУЛЯЦИИ СИСТЕМЫ цАМФ
275
ясной извилине и дорсолатеральной префрон-
ствии между DISC1 и PDE4 будут влиять на ката-
тальной коре при шизофрении. Ассоциирован-
болизм митохондриального цАМФ с сопутствую-
ный с митоген-активированной протеинкиназой
щим физиологическим и психиатрическим исхо-
путь активирует факторы транскрипции, влияю-
дом. Объединяющая связь между шизофренией и
щие на обучение, память, пролиферацию клеток
биполярным аффективным расстройством, а так-
и апоптоз [7-9].
же между DISC1 и PDE4 может происходить на
когнитивном уровне обучения и памяти и на мо-
Следует отметить, что концентрация цАМФ
лекулярном уровне передачи сигналов цАМФ
изменяется при шизофрении не только в цитозо-
[13]. Не случайно PDE4 является неотъемлемым
ле различных клеток, но и во внеклеточном про-
компонентом механизмов действия различных
странстве. Уровни цАМФ уменьшаются в обоня-
классов антидепрессантов [12].
тельной слизи у этих пациентов, и измерения
обонятельной остроты и циклических нуклеоти-
В регуляции уровня цАМФ в клетках помимо
дов в носовой слизи предлагаются для выявления
PDE участвует второй механизм - активность
патологии у пациентов, которые жалуются на по-
аденилатциклаз. У млекопитающих клонировано
терю обоняния [10].
и охарактеризовано по крайней мере девять близ-
Помимо цАМФ при шизофрении изменяется
ко родственных изоформ аденилатциклаз, каж-
активность протеинкиназы A. Это один из клю-
дая из которых кодируется разными генами на
чевых ферментов цАМФ-сигнального пути, ко-
разных хромосомах.
торый обеспечивает многочисленные клеточные
Отдельные изоформы аденилатциклаз предло-
реакции, зависящие от фосфорилирования бел-
жены в качестве инструментов для исследования
ков. Следовательно, дисфункции в различных
специфических механизмов, вовлекающихся в
компонентах передачи сигналов цАМФ могут
реакции клеток различного типа на их стимуля-
быть вовлечены в патогенез шизофрении [11].
цию. Например, аденилатциклаза I предложена
Как известно, концентрация внутриклеточно-
как детектор для исследования механизмов сти-
го цАМФ регулируется посредством таких кле-
муляции с участием αs субъединицы G-белка и
точных механизмов, как активация аденилатцик-
кальций/кальмодулина, а аденилатциклаза II -
лазы, гидролизующей АТФ с образованием
для исследования механизмов стимуляции, в ко-
цАМФ, и фосфодиэстеразы (PDE), понижающей
торые вовлекаются αs субъединица и βγ-димер G-
концентрацию цАМФ в цитозоле. Обнаружено
белка или PKC; аденилатциклаза V или аденилат-
одиннадцать изоформ этого фермента. Так, PDE4
циклаза VI были предложены в качестве детекто-
специфически гидролизует цАМФ и широко экс-
ров для ингибирования, в механизме которого
прессируется во многих типах клеток и тканях,
принимает участие кальций (или NO для адени-
включая нервные клетки.
латциклазаы VI) [15].
В 1970-х годах была разработана концепция
В настоящее время внимание к себе стала при-
«циклической нуклеотидной системы» как мощ-
влекать аденилатциклаза III (ACIII). Впервые ее
ного инструмента, расширяющего возможности
обнаружили в обонятельном нейроэпителии, где
для исследования лекарств. Эта система помимо
она локализуется в обонятельных жгутиках сен-
цАМФ включает аденилатциклазы и фосфодиэс-
сорных нейронов и является важным компонен-
теразы. При этом большое внимание было сосре-
том механизма обонятельной трансдукции. Од-
доточено на цАМФ-PDE и цГМФ-PDE, которые
нако помимо основного компонента в механизме
рассматривались как мишени для разработки но-
обонятельной трансдукции ACIII играет незаме-
вых лекарств, в частности, для улучшения когни-
нимую роль в постнатальном созревании обоня-
тивных функций. Были разработаны специфиче-
тельных сенсорных нейронов (ОСН) и поддержа-
ские ингибиторы PDE4 для многих заболевaний,
нии ультраструктуры обонятельных жгутиков у
среди которых болезнь Альцгеймера, когнитив-
мышей в течение всего взрослого периода [16].
ные нарушения, болезнь Паркинсона, депрессия
и шизофрения [12]. По мнению авторов, профи-
Вместе с тем ACIII экспрессируется и в других
лактика и лечение этих заболеваний требует по-
типах клеток, локализуясь в так называемых пер-
нимания изменений механизмов внутриклеточ-
вичных ресничках, и играет там важную роль.
ной передачи сигналов, связанных с нарушением
Так, длина этих ресничек строго контролируется,
регуляции изофермента PDE, а также других ком-
поскольку укорочение их вызывает такие тяжкие
понентов сигнальной системы цАМФ.
последствия, как полицистоз почек или гидроцe-
Показано, что PDE вовлекается в процесс на-
фалия. Регуляторную роль в нормальном цилио-
рушения регуляции внутриклеточной сигналь-
генезе выполняет ACIII-цАМФ-сигнальный путь
ной системы цАМФ при шизофрении. В этом
[17]. Он обнаруживается в первичных ресничках в
сигнальном пути участвует DISC1 - белок, коди-
фибробластах, синовиоцитах, астроцитах, а так-
руемый одноименным геном. Предполагается,
же в плазматической мембране первичных ресни-
что функциональные изменения во взаимодей-
чек почти всех нейронов в мозге взрослых мышей
БИОФИЗИКА том 67
№ 2
2022
276
БИГДАЙ, СИНЕГУБОВ
и может использоваться в качестве их маркера
прессии GluN2B-субъединицы NMDA-рецепто-
[18].
ров в гиппокампе [5]. Таким образом, нарушения
в ЦНС, связанные с функционированием дофа-
Известно, что от астроцитов в субвентрику-
минергической и глутаматэргической системами
лярной зоне первичная ресничка вытягивается в
являются результатом отсутствия или потери
полость желудочка, заполненную спинномозго-
вой жидкостью, и служит механизмом, с помо-
ACIII в обонятельном эпителии. ACIII в основ-
щью которого эти ненейронные клетки опреде-
ном обонятельном эпителии играет жизненно
важную роль в патофизиологии депрессии и тре-
ляют химический состав окружающей их среды.
вожного состояния.
Тот факт, что некоторые астроциты экспрессиру-
ют ACIII-положительные реснички, указывает на
ACIII в первичных ресничках нейронов регу-
то, что в них цАМФ участвует в качестве вторич-
лирует морфологию дендритов, подтверждая тот
ного мессенджера для воздействия на клеточные
факт, что потеря ACIII ведет к атрофии нейронов.
процессы. Эти данные обеспечиваются дополни-
Общегеномное ассоциативное исследование у
тельным механизмом, который может влиять на
людей недавно выявило причастность аденилил-
роль астроцитов в регуляции синаптической пе-
циклазы III к большому депрессивному рас-
редачи в нервной системе [18].
стройству в результате потери синапсов и соот-
ACIII локализуется также в ресничках клеток
ветственно изменения нейрональной активности
сосудистого сплетения в мозге. Сосудистые спле-
в лимбической и корковой областях. Более того,
тения - это структуры, обнаруженные в боковых,
уровень экспрессии ACIII в крови считается био-
третьем и четвертом желудочках головного мозга;
маркером депрессии у людей. Исследование на
их основные функции - производство и гомео-
мышах продемонстрировало, что потеря ACIII у
стазис спинномозговой жидкости. Некоторые
них приводит к снижению нейрональной актив-
клетки сосудистого сплетения имеют небольшие
ности, изменению режима сна и поведению, по-
пучки подвижных ресничек, другие же клетки -
добному депрессии, предоставляя убедительные
единственную первичную ресничку. Разрушение
доказательства, подтверждающие, что ACIII яв-
этих ресничек связано с повышенным содержа-
ляется геном-кандидатом [20]. Это исследование
нием хлора в спинномозговой жидкости в сочета-
также предполагает новую стратегию борьбы с де-
нии с гидроцефалией. Интересно, что повышен-
прессией, направленную на внутриклеточную
ный уровень хлоридов в спинномозговой жидко-
сигнальную систему цАМФ, в частности, в рес-
сти сочетается с повышенной внутриклеточной
ничках нейронов.
концентрацией цАМФ в эпителии сосудистого
Значительное снижение экспрессии ACIII об-
сплетения. Таким образом, нарушение регуляции
наружено в обонятельном сенсорном эпителии у
ACIII-опосредованной передачи сигналов цАМФ
больных шизофренией [21].
в эпителиальных клетках может быть причиной
Признание проблем, связанных с нарушением
возникновения гидроцефалии [19].
регуляции системы цАМФ при шизофрении, по-
Активность аденилатциклазы в первичных
служило поводом для разработки кандидатных
ресничках нейронов центральной нервной систе-
молекулярных подходов в определении причин,
мы (ЦНС) может иметь решающее значение для
лежащих в основе патологии развития при шизо-
некоторых форм обучения и памяти. ACIII явля-
френии
[22]. Существует множество доказа-
ется ключевым медиатором передачи сигналов
тельств того, что, по крайней мере, когнитивные
цАМФ в первичных ресничках мозга. Поскольку
и негативные симптомы шизофрении являются
она экспрессируется в первичных ресничках ней-
результатом дисфункции глутаматергической си-
ронов гиппокампа, исследовали ACIII-/- мышей
стемы. Литературные данные свидетельствуют о
на наличие нескольких форм обучения и памяти
дефиците функции NMDA-рецепторов при ши-
показали, что ACIII-/- у мышей наблюдается на-
зофрении, что является одной из основных при-
рушение обучаемости и памяти на временное
чин этого заболевания [23].
диссоциативное пассивное избегание. Так как
NMDA-рецепторы на нейронах гиппокампа и
ACIII экспрессируется исключительно в первич-
церебрального кортикального слоя участвуют в
ных ресничках, это означает, что сигналы цАМФ,
процессах обучения и памяти, вовлекая в этот
которые генерируются внутри первичных ресни-
процесс цАМФ-зависимую PDE4 [4]. Вместе с
чек, способствуют некоторым формам обучения
тем показано, что активация NMDA-рецепторов
и памяти, включая потерю контекстуальной обу-
вызывает повышение концентрации цАМФ в об-
словленности страха [4].
ласти СА1 гиппокампа посредством активации
У мышей с генетическим нокаутом аденилил-
аденилатциклаз, что свидетельствует о том, что
циклазы ACIII в обонятельном эпителии умень-
метаболизм циклического АМФ играет важную
шается продукция цАМФ, что приводит к депрес-
роль в индукции зависимой от NMDA-рецепто-
сии и вызывает тревогу. Эти состояния связаны с
ров долговременной потенциации в области CA1
подавлением дофаминергической системы и экс-
гиппокампа [22]. Поэтому ослабление функции
БИОФИЗИКА том 67
№ 2
2022
НАРУШЕНИЕ РЕГУЛЯЦИИ СИСТЕМЫ цАМФ
277
NMDA-рецепторов при шизофрении может ини-
получить представление о связанных с шизофре-
циировать нарушение регуляции системы цАМФ
нией изменениях в развитии нервной системы,
в нейронах ЦНС через снижение активности
реакции на стресс и путях регуляции экспрессии
ACIII и служить одним из молекулярных меха-
генов [30].
низмов, лежащих в основе этого заболевания [23].
Обонятельные клетки в первичной культуре
Следует отметить, что важную информацию о
являются функционально зрелыми, экспрессиру-
функционировании этого фермента в нейронах
ющими рецепторные и внутриклеточные меха-
ЦНС в норме и при дисфункции NMDA-рецеп-
низмы не только для обонятельной трансдукции
торов может дать изучение активности ACIII в
(в частности, ACIII), но и для нейромедиаторных
ОСН, поскольку появляется все больше свиде-
путей, участвующих в психоневрологических
тельств того, что обонятельная дисфункция явля-
расстройствах, таких как биполярная депрессия
ется эндофенотипом шизофрении, таким об-
или шизофрения [24].
разом, биологические основы обонятельной
Животных, предварительно анестезированные
дисфункции могут дать ключ к патофизиологиче-
с применением эфира, подвергали цервикальной
ским механизмам, влияющим на нейроны в раз-
дислокации. Препарирование обонятельных за-
личных областях мозга [27]. Учитывая многие из-
витков I-IV проводили на льду при постоянном
вестные клеточные и молекулярные сходства в
орошении аэрированным буферным раствором
нейрогенезе, дифференцировке и созревании
следующего состава: 150 мМ NaCl, 20 мМ HEPES,
между обонятельным эпителием и центральной
5 мМ D-глюкозы, 5 мМ KCl, pH 7.4. Изолирован-
нервной системы, исследование регуляции внут-
ные фрагменты обонятельного эпителия с подле-
риклеточной цАМФ сигнализации в ОСН полез-
жащей соединительной тканью инкубировали в
но для понимания аномального развития нерв-
буферном растворе с добавлением 1% фракции V
ной системы при шизофрении [28].
бычьего альбумина и 1 мг/мл коллагеназы типа IV
Однако, как видно из выше изложенного, фер-
в течениe 15 мин при 37°C. Раствор декантирова-
ментативная активность ACIII определялась либо
ли и ткань дополнительно дважды промывали бу-
посредством биохимического анализа в изолиро-
ферным раствором без добавок.
ванных и суспендированных мембранных препа-
Суспензию диссоциированных клеток готови-
ратах обонятельных жгутиков [29], либо при
ли путем тритурирования с использованием по-
оценке уровня ее экспрессии в обонятельном
лированных в пламени горелки стеклянных пи-
эпителии у пациентов с шизофренией, либо по
петок в буферном растворе (145 мМ NaCl, 20 мМ
концентрации цАМФ в тромбоцитах и обоня-
HEPES, 5 мМ D-глюкозы, 5 мМ KCl, 2 мМ CaCl2,
тельной слизи у этих пациентов. Нам не удалось
1 мМ MgCl2, pH 7.4) с добавлением 1% (w/v)
обнаружить данные, полученные при прямой
фракции V бычьего альбумина для обеспечения
стимуляции ACIII в норме и при патологии.
более равномерного окрашивания препарата, по-
Поэтому в данной работе мы попытались отве-
скольку липофильные флуоресцентные красите-
тить на вопрос, изменяется ли активность самой
ли склонны к образованию эмульсии в водных
ACIII без влияния на нее сопряженного с ней G-
растворах. Полученную суспензию смешивали с
белка при шизофрении.
1 мМ стоковым раствором флуоресцентного кра-
Для этого мы исследовали прямую стимуля-
сителя Фура-2-ацетоксиметилового эфира (Fura-
цию ACIII, минуя ее рецептор-сопряженную ак-
2 AM) в безводном диметилсульфоксиде до ко-
тивацию, в изолированных обонятельных сен-
нечной концентрации 5 мкМ. Суспензию поме-
сорных нейронах ОСН крыс линии Вистар в кон-
щали на предварительно обработанные поли-l-
троле и в модели шизофрении, индуцируемой
лизином стекла и инкубировали в течение 1 ч при
введением (+)-МК-801.
37°C. Остатки неосажденной суспензии одно-
кратно отмывали от стекол буферным раствором.
Активность ACIII в ОСН в ответ на стимуля-
МАТЕРИАЛЫ И МЕТОДЫ
цию оценивали посредством прижизненной флу-
В качестве объекта исследования использова-
оресцентной микроскопии по изменению интен-
ли изолированные обонятельные сенсорные ней-
сивности свечения Fura-2 AM, характеризующей
роны крыс линии Вистар, полученных из ЦКП
изменение содержания ионов кальция в цитозоле
«Биоколлекция» Института физиологии РАН.
ОСН. Для этого препарат помещали в открытую
Как известно, ОСН представляют модель клеток,
камеру объемом 500 мкл на предметный столик
полученных из обонятельного эпителия, в каче-
микроскопа МИКМЕД-2 (ЛОМО, Россия), осна-
стве модельной системы для изучения молеку-
щенного камерой, дихроичным зеркалом и свето-
лярных механизмов, участвующих в шизофрении
фильтрами для выделения полосы возбуждения
и других нервно-психических расстройствах. Не-
люминесценции 380 нм и эмиссии 510 нм. Реги-
давние исследования патофизиологии шизофре-
страцию сигнала проводили с помощью програм-
нии с использованием этой модели позволили
мы VirtualDub 2.1.
БИОФИЗИКА том 67
№ 2
2022
278
БИГДАЙ, СИНЕГУБОВ
Для прямой стимуляции ACIII применяли
ской обработки данных применяли статистиче-
форсколин. Его высокие концентрации обнару-
ский пакет STATISTICA Base. Статистическую
живаются исключительно в нейронах, что позво-
значимость полученных данных оценивали с по-
ляет отделить в ходе анализа нейрональную фрак-
мощью дисперсионного анализа.
цию от всех прочих клеток в смешанной первич-
ной культуре из обонятельного нейроэпителия.
Помимо форсколина применение ацетоксимети-
РЕЗУЛЬТАТЫ
ловых эфиров дополнительно позволяет выявить
Визуальные наблюдения за крысами, которым
нейрональные клетки в смеси других типов кле-
вводили (+)-МК-801, выявили характерные для
ток, так как он относительно селективно метабо-
шизофрении негативные симптомы. В частности,
лизируется в них. Чтобы получить реакцию на
животные были социально изолированы. В отли-
стимуляцию только ACIII, исключив активность
чие от контрольной группы крысы, получавшие
PDE, применяли смесь из 15 мкМ форсколина,
препарат, проводили значительное количество
специфического активатора аденилатциклаз, и
времени на удалении от матери и при принуди-
100 мкМ 3-изобутил-1-метилксантина (IBMX) -
тельном отделении от общей группы не возвра-
эффективного блокатора Ca2+/кальмодулин-за-
щались назад. Таким образом, эти результаты да-
висимой PDE в обонятельных нейронах [31]. Та-
ют основание для заключения о том, что измене-
ким образом, наблюдаемые изменения концен-
ния во внутриклеточной сигнализации, которые
трации внутриклеточного кальция будут обуслов-
мы ожидаем получить в ОСН от животных в мо-
лены накоплением цАМФ и последующей
дели шизофрении в нашей работе, позволят гово-
активацией циклонуклеотид-зависимых Са2+-
рить о клеточных и молекулярных механизмах,
каналов.
которые лежат по крайней мере в основе негатив-
ной симптоматики.
Роль механизмов регуляции аденилатциклаз-
ной системы оценивалась с помощью LY294002 и
Активность ACIII оценивали по изменению
кальмидазолия хлорида - ингибиторов соответ-
интенсивности флуоресценции Fura-2 в ответ на
ственно фосфоинозитид-3-киназы и кальмоду-
стимуляцию. Уменьшение сигнала свидетель-
лина. Применяемые в опыте концентрации под-
ствовало о нарастании концентрации внутрикле-
бирали эмпирически.
точного кальция. Результаты наших исследова-
ний выявили, что под действием смеси форско-
Эксперименты проводили на модели шизо-
лина (15 мкМ) и 3- IBMX (100 мкМ) во всех
френии, индуцируемой введением (+)-MK-801.
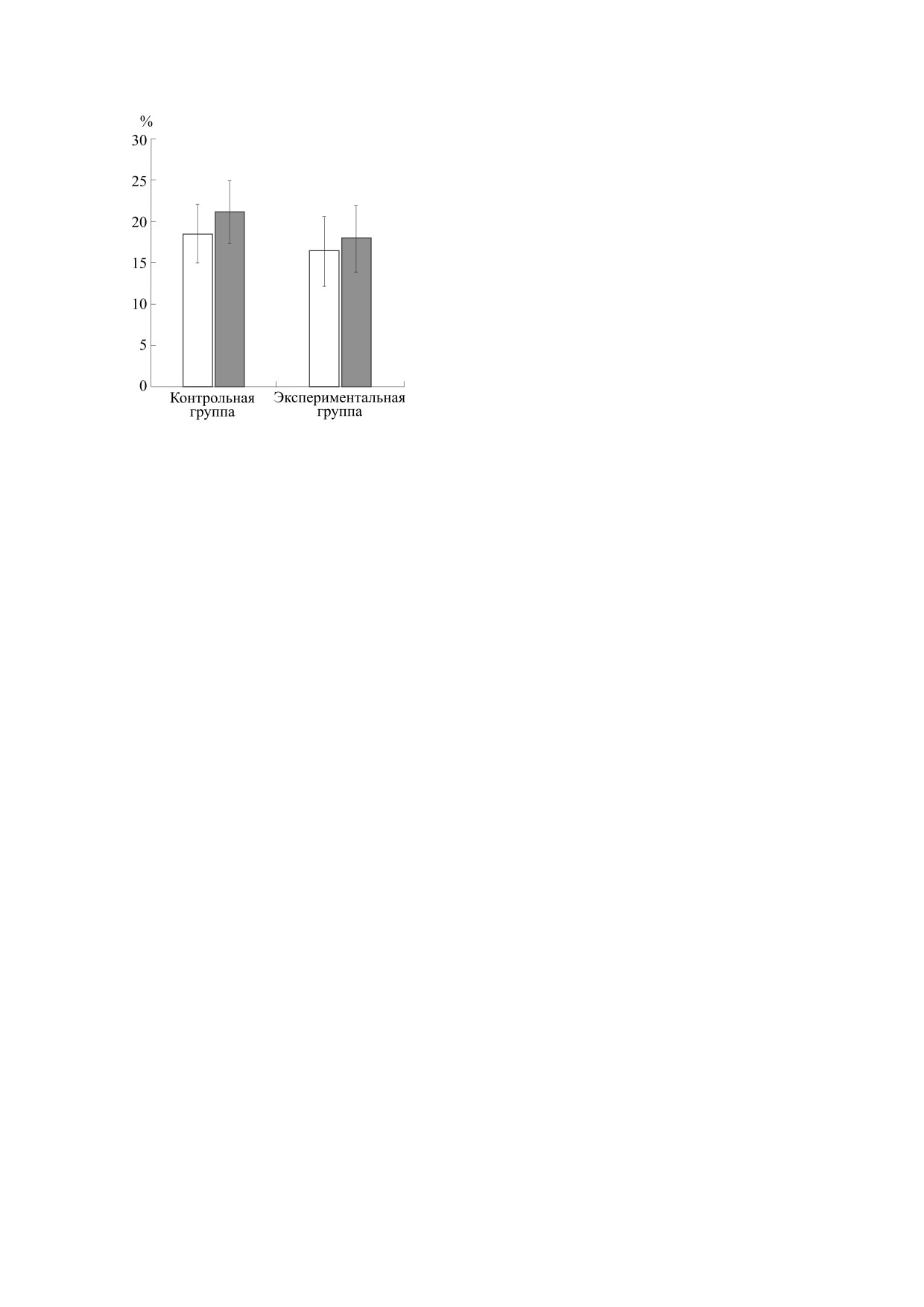
случаях наблюдалось уменьшение флуоресцен-
(+)-MK-801 представляет собой антагонист
ции в клетках контрольной группы на 18.5 ± 3.6%
NMDA-рецептора, который является основой
(n = 48; p = 0.05), а у экспериментальных живот-
для глутаматной модели шизофрении, а индуци-
ных - на 16.4 ± 4.1% (n = 54; p = 0.05). Следова-
руемая им социальная изоляция крыс является
тельно, при активации ACIII совместно с ингиби-
признанной моделью негативных симптомов ши-
рованием PDE концентрация цитозольного каль-
зофрении [32].
ция в ОСН увеличивалась как в контрольной, так
Протокол введения препарата основывался на
и в экспериментальной группе животных. Как
литературных данных, в которых была продемон-
видно, в экспериментальной группе реакция на
стрирована способность (+)-MK-801 индуциро-
стимуляцию ACIII форсколином, увеличиваю-
вать персистирующие изменения в поведении и
щим ее активность, статистически значимо была
развитии нервной системы при системном введе-
меньше, чем в контроле. Вероятно, это обуслов-
нии в ранний постнатальный период. Инъекции
лено ослаблением активации фермента в нейро-
производились с частотой один раз в сутки под-
нах в группе животных с измененной функцией
кожно в дозировке 0.5 мг на килограмм в течение
NMDA-рецепторов.
первой недели жизни. В качестве контрольной
группы выступали животные из того же помета,
Известно, что кальмодулин (СаМ) модулирует
получавшие 0.9% раствор NaCl равного объема
активность ряда компонентов пути обонятельной
[33].
трансдукци. Показано, что обонятельные жгути-
ки содержат кальмодулин. Поэтому мы предпо-
Обработку полученных изображений и кван-
ложили, что изменения в системе внутриклеточ-
тификация уровней флуоресценции проводили с
ной сигнализации могут частично объяснить раз-
помощью программы FIJI (NIH, США). Для это-
личия в реакциях на стимуляцию ACIII в обеих
го каждую отдельную клетку отмечали как ROI
группах.
(region of interest). Для каждого отдельного ROI
измеряли средний уровень интенсивности сигна-
Для этого выполняли преингибирование каль-
ла и выражали его в единицах, представляющих
модулина в течение 15 мин с помощью кальмида-
собой отношение интенсивности в данный мо-
золия хлорида (100 мкM), с последующей стиму-
мент времени к усредненной интенсивности сиг-
ляцией смесью форсколина (15 мкM) и IBMX
нала в отсутствие стимуляции. Для статистиче-
(100 мкM). Оказалось, что, как и в предыдущей
БИОФИЗИКА том 67
№ 2
2022
НАРУШЕНИЕ РЕГУЛЯЦИИ СИСТЕМЫ цАМФ
279
серии, интенсивность флуоресценции под дей-
Результаты анализа полученных данных пока-
ствием стимулирующей ACIII смеси падала, сви-
зали, что при прямой стимуляции ACIII форско-
детельствуя о накоплении Са2+ в цитозоле ОСН.
лином повышается концентрация ионов Са2+ в
Однако на фоне ингибирования СаМ амплитуда
ОСН обеих групп. Как известно, активированная
сигнала в обеих группах была выше, чем при со-
ACIII осуществляет гидролиз АТФ, продуцируя
хранении его активности. Так, в контрольной
цАМФ. В условиях ингибирования PDE концен-
группе она достигала
21.14
± 3.7% (n = 61;
трация цАМФ в цитозоле увеличивается еще
p = 0.05), а в экспериментальной - 17.9 ± 4.1%
больше. Рост содержания цитозольного Са2+ при
(n = 45; p = 0.05).
этом достигается за счет его входа через цикло-
Следовательно, меньшая реакция обонятель-
нуклеотид-зависимые (CNG) Са2+-каналы, экс-
ных сенсорных нейронов на смесь раздражителей
прессируемые в обонятельных жгутиках разных
в первой серии экспериментов обусловлена, ве-
животных, включая крыс [28].
роятно, ингибирующим действием СаМ, которое
Однако, как следует из наших результатов, ре-
распространяется и на клетки животных, инъе-
акция ОСН на смесь форсколина и IBMX была
цированных (+)-МК-801.
значимо меньше в обонятельных сенсорных ней-
Другим ферментом, опосредующим ингиби-
ронах от крыс, инъецированных антагонистом
рование пути обонятельной трансдукции, являет-
NMDA-рецепторов. В этих клетках меньше акку-
ся фосфоинозитид-3-киназа. Мы предположили,
мулировалось кальция под действием смеси раз-
что она может модулировать активность адени-
дражителей в результате, вероятно, инактивации
латциклазы III в ОСН. Для проверки нашего
Са2+-тока. Можно предположить, что подавле-
предположения мы перед стимулированием в те-
ние любой из ступеней каскада передачи сигнала
чение
15 мин предварительно ингибировали
может способствовать этому процессу. Мы в на-
клетки в LY294002 (100 мкM). Реакцию на стимул
ших опытах стимулировали напрямую ACIII, ми-
регистрировали от 43 клеток контрольных и 49
нуя мембранные рецепторы.
клеток экспериментальных животных. Результа-
Следовательно, более слабую флуоресценцию,
ты анализа показали, что на фоне ингибирования
наблюдаемую нами в ОСН, стимулируемых сме-
фосфоинозитид-3-киназы интенсивность флуо-
сью форсколина и IBMX у опытных животных,
ресценции под действием стимуляции смесью
можно объяснить ослабленной активацией ACIII
раздражителей статистически значимо не изме-
при шизофрении, инициируемой введением
нялась в обеих группах по сравнению с той, кото-
МК-801. Наше предположение согласуется с дан-
рая регистрировалась в отсутствие ингибитора.
ными, полученными на ОСН у пациентов с шизо-
Таким образом, фосфоинозитид-3-киназа не
френией. В них падала активность сопряженного
влияла на активность сигнального пути цАМФ в
с ACIII G-белка и значительно уменьшалась экс-
обонятельных сенсорных нейронах как в кон-
прессия ACIII, приводя к небольшой продукции
трольных, так в опытных образцах.
цАМФ [28].
Вероятно, аномальная активность ACIII в на-
ОБСУЖДЕНИЕ
шей работе является результатом подавленной
функции NMDA-рецепторов, которые активиру-
Введение антагонистов NMDA-рецепторов
ет ACIII в нейронах ЦНС и обонятельных сенсор-
грызунам является одной из наиболее продуктив-
ных нейронах у крыс, получавших инъекции
ных моделей шизофрении, используемой на до-
(+)-МК-801. Такое предположение основывается
клиническом этапе разработки препаратов. Ее ха-
на том, что NMDA-рецепторы экспрессируются
рактерной чертой является высокое сходство
не только в нейронах ЦНС, но и в ОСН [21]. А
нейробиологических изменений с таковыми у
при шизофрении, как известно, функция
больных и наличие в спектре эффектов так назы-
NMDA-рецепторов ослаблена, что является од-
ваемых «негативных симптомов», составляющих
ной из основных причин этого заболевания [36].
облигатные диагностические критерии для шизо-
Поэтому разрегуляция внутриклеточной сиг-
френии. Наиболее полно они проявляются в слу-
нальной системы цАМФ в нейрональной ткани,
чае хронического и субхронического введения и
наблюдаемая при шизофрении, вероятно, объяс-
не угасают в течение длительного времени после
няется недостаточной активацией ACIII посред-
прекращения действия препарата [1]. Первая не-
ством NMDA-рецепторов, вызывая уменьшение
деля жизни крыс является критическим перио-
потока Са2+ в цитозоль через CNG-каналы и сни-
дом в развитии обонятельного эпителия и синап-
жение возбудимости ОСН, которую наблюдают у
тических связей с нейронами обонятельной луко-
пациентов с шизофренией.
вицы. Исходя из предположения, что (+)-MK-
Однако следует отметить, что в общий кальци-
801 может нарушать процесс развития обонятель-
евый ток вносят вклад ионы, входящие через
ной системы, препарат вводили именно в данный
период.
NMDA-рецепторы, проницаемые для Са2+. Вы-
БИОФИЗИКА том 67
№ 2
2022
280
БИГДАЙ, СИНЕГУБОВ
ляцию. Однако результаты выявили противопо-
ложную реакцию на смесь форсколина с IBMX
как в контрольной, так и в опытной группе жи-
вотных. Анализ наших результатов выявил досто-
верное увеличение реакции на стимул в обеих
группах по сравнению с той, которая наблюда-
лась при сохранении активности СаМ (см. рису-
нок).
Увеличение концентрации Са2+ в цитозоле
может обусловливаться не только активацией
ACIII, но и ингибированием PDE. Как указано
выше, в своих экспериментах стимуляцию ACIII
форсколином мы осуществляли вместе с IBMX -
ингибитором Са/СаМ-зависимой PDE, особенно
эффективным в ОСН [30]. Ca2+ может активиро-
вать Ca2+/СаМ-зависимую PDE, тем самым сни-
жая локальную концентрацию сAMP. Однако на
Са2+-зависимое ингибирование этой PDE не
Изменение флуоресценции обонятельных нейронов
влияют ингибиторы кальмодулина [39]. Следова-
под действием стимула в контрольной и эксперимен-
тельно, снижая активность ACIII, мы сохраняем
тальной группах: светлые столбики - IBMX + форско-
влияние IBMX на PDE. Казалось бы, на фоне ин-
лин, темные столбики - IBMX + форсколин + кальми-
дазолия хлорид.
гибирования Са2+/СаМ-зависимой PDE в отсут-
ствие синтеза цАМФ посредством ACIII трудно
ожидать высокой продукции цАМФ, активирую-
ше было сказано, что под действием (+)-МК-801
щей CNG-зависимые каналы и, вследствие этого,
уменьшается вероятность их открытых состояний
более высокий флуоресцентный сигнал на стиму-
и, как следствие, их проницаемость для Са2+.
ляцию ОСН. Но полученные результаты расхо-
Возможно, и этот фактор обусловливает более
дятся с нашими ожиданиями.
слабый флуоресцентный сигнал ОСН в ответ на
В ОСН кальмодулин активирует также 3',5'-
стимуляцию, а значит, меньшую концентрацию
циклонуклеотид-зависимые фосфодиэстеразы
внутриклеточного Са2+ в ОСН крыс, инъециро-
[31]. Возможно, ингибирование кальмодулина в
ванных (+)-МК-801. И в этом случае уменьшение
наших опытах снижает активность цАМФ-зави-
входящего тока кальция в нейроны будет вносить
симой PDE, вызывая повышение содержания
вклад в изменение возбудимости ОСН.
цАМФ в цитозоле. Показано, что из всех изо-
Полученные нами данные позволяют предпо-
форм цАМФ гидролизуют только PDE4, PDE7 и
ложить, что подавление функции NMDA-рецеп-
PDE8, причем PDE4, которая вовлекается в регу-
торов при шизофрении вызывает подавление ак-
ляцию цАМФ при шизофрении, распространена
тивности нейронов ЦНС по двум механизмам,
в большинстве клеток, включая обонятельные
приводящим к падению концентрации цитозоль-
нейроны [32]. Вместе с Са/СаМ-зависимой PDE
ного кальция: с одной стороны - через разрегуля-
они повышают концентрацию кальция выше ис-
цию системы цАМФ, а с другой - благодаря
ходной, что мы наблюдаем в наших опытах.
ослаблению собственной функции NMDA-ре-
NMDA-рецепторы являются каналами, чьи
цепторов.
физиологические свойства также регулируются
Известно, что в регуляции активности ACIII,
внутриклеточным Са2+, способным ингибиро-
не зависимой от сопряженного с ней G-белка, в
вать активность NMDA-рецепторного канала. В
обонятельных рецепторных нейронах и нейронах
Са2+-зависимой регуляции NMDA-рецептора
ЦНС участвует кальмодулин, который активиру-
также участвует СаМ, связывание с которым вы-
ет аденилатциклазу III [29]. В Cа2+/СаМ-зависи-
зывает четырехкратное снижение вероятности
мую активацию ACIII вовлекаются также
открытия канала. Этот Са2+/СаМ-зависимый ме-
NMDA-рецепторы.
ханизм играет важную роль в защите от эксайто-
Чтобы проверить роль СаМ в регуляции кле-
токсической смерти клеток [33].
точного ответа на стимуляцию форсколином и
Са2+/СаМ ингибирует NMDA-рецепторные
IBMX, ОСН предварительно ингибировали каль-
каналы, вызывая снижение концентрации цито-
мидозолием. Мы ожидали, что ингибирование
СаМ, снижая активность Са/СаМ-зависимой
зольного Са2+. При ингибировании СаМ его бло-
ACIII, будет уменьшать реакцию ОСН на стиму-
кирующее действие на NMDA-рецепторы пре-
БИОФИЗИКА том 67
№ 2
2022
НАРУШЕНИЕ РЕГУЛЯЦИИ СИСТЕМЫ цАМФ
281
кращается, вызывая ток Са2+ через этот канал и
СПИСОК ЛИТЕРАТУРЫ
повышение цитозольного Са2+ в ответ на актива-
1.
E.-J. Choi, Zh. Xia, and D. R. Storm, Biochemistry
цию ACIII, что мы и видим в наших опытах. Но
31, 6492 (1992).
такой механизм работает при нормальном функ-
2.
J. Bradley, D. Reuter, and S. Frings, Science 294, 2176
ционировании NMDA-рецепторов. В группе жи-
(2001).
вотных, инъецированных (+)-МК-801, этот ре-
3.
J. Bradley, W. Bönigk, K.-W. Yau, and S. Frings, Nat.
цептор заблокирован. Следовательно, можно бы-
Neurosci. 7 (7), 705 (2004).
ло ожидать уменьшения реакции на стимуляцию
4.
J. Hanoune and N. Defer, Annu. Rev. Pharmacol.
ОСН. Однако мы зарегистрировали ее увеличе-
Toxicol. 41, 145 (2001).
ние при ингибировании СаМ. Вероятно, в нерв-
5.
Z. Wang, T. Phan, and D. R. Storm, J. Neurosci. 31
ных клетках, принадлежащих группе крыс с мо-
(15), 5557 (2011).
делью шизофрении, Са/СаМ-зависимая PDE
вместе с цАМФ-зависимой PDE при их ингиби-
6.
X. Liu, Y. Zhou, S. Li, et al., J. Affective Disorders
ровании повышают концентрацию кальция выше
268, 28 (2020).
исходной.
7.
B. I. Turetsky and P. J. Moberg, Am J Psychiatry 166
(2), 226 (2009).
Таким образом, можно предположить, что при
шизофрении поддерживается более низкая кон-
8.
Kyosseva S. Int Rev Neurobiol 59, 201 (2004).
центрация Са2+ в нервных клетках, чем у здоро-
9.
A. J. Funk, R. E. McCullumsmith, V. Haroutunian,
вых людей, в результате более низкой активности
and J. H. Meador-Woodruff, Neuropsychopharma-
ACIII, регулируемой NMDA-рецепторами, и сов-
cology 37, 896 (2012).
местной активности фосфодиэстераз, снижаю-
10.
N. Ma, T. Abel, and P. J. Hernandez, Learn. Mem. 16,
щих концентрацию цАМФ, продуцируемой по-
367 (2009).
средством ACIII, в результате чего падает Са2+-
11.
D. Tardito, G. B. Tura, L. Bocchio, et al., Neuropsy-
ток, снижая нейрональную активность.
chopharmacology 23 (2), 217 (2000).
Результаты, полученные в данной работе, дают
12.
T. Keravis and C. Lugnier Brit. J. Pharmacol. 165,
основания полагать, что в модели шизофрении,
1288 (2012).
индуцируемой введением (+)-МК-801, ингиби-
13.
J. K. Millar, B. S. Pickard, S. Mackie, et al., Science
рующего NMDA-рецепторы в нейронах ЦНС, а
310, 1187 (2005).
также в ОСН, снижается активность внутрикле-
14.
A. R. Kuzel, M. Lodhi, and M. Rahim, Cureus 9 (11),
точной сигнальной системы трансдукции, сопря-
e1878 (2007).
женной с активностью аденилатциклаз. Суммар-
15.
Z. Zhang, D. Yang, et al., Front. Cell. Neurosci. 11 (1)
ное снижение содержания кальция в цитозоле
(2017).
нейрональных клеток, обусловленное как инги-
16.
Y. Ou, Y. Ruan, M. Cheng, et al., Exp. Res. 15, 2802
бированием NMDA-рецепторов, так и ослабле-
(2009).
нием активности аденилатциклаз будет вызывать
снижение нейрональной активности, подобную
17.
M. C. Antal, K. Beanardais, B. Samama, et al., PLoS
той, которую наблюдали у мышей с потерей ACIII
One 12 (1), e0170756 (2017). DOI: 10.1371/jour-
[34]. Это, в свою очередь, снизит эффективность
nal.pone.0170756
синаптической передачи, синаптической пла-
18.
G. A. Bishop, N. F. Berbar, J. Lewis, et al., J. Comp.
стичности, образование синапсов, в которых глу-
Neurol. 505, 562 (2007).
таматные NMDA-рецепторы/каналы играют
19.
X. Chen, J. Luo, Y. Leng, et al., Biol. Psychiatry 80
важную роль [35].
(11), 836 (2006).
20.
K. E. Borgmann-Winter, H.-Y. Wang, R. Ray, et al.,
Schizophrenia Bull. 42 (2) 377 (2016).
КОНФЛИКТ ИНТЕРЕСОВ
21.
C. S. Weickert and D.R. Weinberger, Schizophrenia
Автор заявляет об отсутствии конфликта инте-
Bull. 24 (2), 303 (1998).
ресов.
22.
L. S. Pilowsky, R. A. Bressan, J. M. Stone, et al., Mol.
Psychiatry 11, 118 (2006).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
23.
D. Chetkovich and J. D. Sweatt, J. Neurochem. 61,
1933 (1999).
Эксперименты с применением животных про-
24.
K. E. Borgmann-Winter, N. E. Rawson, H.-Y. Wang,
ведены в соответствии с локальными этическими
et al., Neuroscience 158 (2), 642 (2009).
стандартами, принятыми в ИФ РАН и согласо-
ванными с Европейской конвенцией о защите
25.
S. E. Arnold, L. Y. Han, P. J. Moberg, et al., Arch.
позвоночных животных, используемых для экс-
Gen. Psychiatry 58, 829 (2001).
периментов или в иных научных целях (СEД
26.
R. R. H. Anholt and A. M. Rivers, Biochemistry, 29
№123).
(17), 4049 (1990).
БИОФИЗИКА том 67
№ 2
2022
282
БИГДАЙ, СИНЕГУБОВ
27. J. C. Neill, S. Barnes, S. Cook, et al., Pharmacology &
31. M. Conti, D. Mika, and W. J. Richter, Gen. Physiol.
Therapeutics 128, 419 (2010).
143 (1), 29 (2003).
28. S. C. Shirley, C. J. Robinson, K. Dickinson, et al.,
32. M. D. Ehlers, S. Zhang, J.P . Bernhardt, R. L. Hugan-
Biochem. J. 240, 605 (1986).
ir, Cell 84, 745 (1996).
29. R. H. Kramer and S. A. Siegelbaum, Neuron 9, 897
33. J. Lavoie, A. Sawa, K. Ishizuka, Curr. Opin. Psychia-
(1992).
try 30 (3), 176 (2007).
30. C. B. Klee, T. H. Crouch, and P. G. Richman, Ann.
Rev. Biochem. 49, 489 (1980).
34. I. B. Levitan, Neuron 22, 645 (1999).
Dysregulation of the cAMP System in Olfactory Neurons in a Model of Schizophrenia
Induced by Administration of (+)-MK-801 to Rats
E.V. Bigdai and A.A. Sinegubov
Pavlov Institute of Physiology, Russian Academy of Sciences, nab. Makarova 6, St. Petersburg, 199034 Russia
One of the key features of schizophrenia are neurodevelopmental abnormalities. Those can be reproduced in
rodent model of schizophrenia by perinatal exposure to NMDA-receptor antagonists. Here we show that in
this model (+)-MK-801 induces functional abnormalities in well-known neurogenic niche - olfactory neu-
roepithelium. Olfactory sensory neurons of model animals had lower activity of adenylyl cyclase associated
with differences in calmodulin-dependent regulation of canonical olfactory signaling.
Keywords: sense of smell, schizophrenia, neurogenesis, NMDA
БИОФИЗИКА том 67
№ 2
2022