БИОФИЗИКА, 2022, том 67, № 3, с. 456-466
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 577.323.23+544.341.2.001.24+544.312.001.24
СВЯЗЫВАНИЕ ЛИГАНДОВ С НУКЛЕИНОВЫМИ КИСЛОТАМИ
В РАСТВОРЕ И НА МИКРОЧИПАХ
© 2022 г. Ю.Д. Нечипуренко*, **, А.C. Бучельников**
*Институт молекулярной биологии им. В.А. Энгельгардта РАН, Москва, ул. Вавилова, 32
E-mail: nech99@mail.ru
**Севастопольский государственный университет, 299053, Севастополь, ул. Университетская, 33
Поступила в редакцию 02.04.2022 г.
После доработки 02.04.2022 г.
Принята к публикации 04.04.2022 г.
Многие процессы в живой клетке включают в себя равновесное связывание лигандов (белков,
биологически активных соединений и олигонуклеотидов) с нуклеиновыми кислотами. Модели и
методы теории физической адсорбции позволяют количественно описать такое связывание в
случае, когда лиганд занимает протяженный участок на ДНК и когда существуют кооперативные
взаимодействия между адсорбированными молекулами лиганда. Применение модели Изинга
позволяет построить решение уравнений адсорбции в самом общем виде, для широкого класса
систем. Физико-химические эксперименты в модельных системах in vitro, где присутствуют, с
одной стороны, такие биологические макромолекулы как белки, сложные белковые комплексы,
полимеры РНК и ДНК, а с другой - малые молекулы лигандов, позволили пролить свет на
множество механизмов, лежащих в основании клеточных процессов. Диаграммы футпринтинга
или данные, полученные на микрочипах, позволяют получить детальную информацию о системе,
где происходит связывание лигандов с ДНК. Краткий обзор теоретических исследований в этой
области содержится в данной работе. Описано, как можно использовать расчеты энтропии
адсорбционной системы, чтобы представить происходящие в ней процессы кооперативного
связывания. В приложении дана методика анализа связывания на микрочипах.
Ключевые слова: связывание белков с ДНК, кооперативные взаимодействия, изотермы адсорбции,
модели адсорбции, модель Изинга, микрочип.
DOI: 10.31857/S000630292203005X, EDN: ANDOHM
Развитие экспериментальных методов анализа
ми, которые занимают на полимере соседние
связывания лигандов с нуклеиновыми кислотами
центры связывания. Лиганды при связывании на
после открытия структуры ДНК привело к появ-
макромолекуле могут как способствовать связы-
лению сотен теоретических работ, в которых рас-
ванию, так и препятствовать ему, в первом случае
сматривалось связывание лигандов с линейными
говорят о положительной кооперативности, во
полимерами. Подходы статистической термоди-
втором — об отрицательной кооперативности
намики к данным системам были применены
или антикооперативности взаимодействия.
впервые в работах Д. Брэдли и С. Лифсона [1] и
Описание систем, в которых наблюдаются ко-
С. Латта и Х. Собера [2]. Уравнения адсорбции,
оперативные взаимодействия, было проведено
описывающие связывание лигандов, получили
впервые в работах [5, 6]. Г.В. Гурский, А.С. За-
широкое распространение после работы Д. Кро-
седателев и М.В. Волькенштейн решили ряд тео-
зерса [3] (следует заметить, что с точки зрения
ретических задач, которые позволили описывать
статистической термодинамики задача адсорб-
связывание биологически активных соединений
ции протяженного лиганда на линейном полиме-
с нуклеиновыми кислотами при помощи систем
ре принадлежит классу решеточных моделей, и
алгебраических уравнений и рекуррентных соот-
здесь впервые решение получено Л. Тонксом еще
ношений. Описанию кооперативного связыва-
в 1936 г. [4]).
ния лигандов с ДНК была посвящена также рабо-
Важный этап развития теории адсорбции свя-
та Дж. МакГи и П. фон Хиппеля [7]. В этой работе
зан с учетом кооперативных взаимодействий
были наглядно истолкованы полученные Заседа-
между адсорбированными лигандами. Коопера-
телевым с соавторами уравнения для простейше-
тивность может быть обусловлена, например,
го случая контактных кооперативных взаимодей-
контактными взаимодействиями между лиганда-
ствий между адсорбированными лигандами.
456
СВЯЗЫВАНИЕ ЛИГАНДОВ С НУКЛЕИНОВЫМИ КИСЛОТАМИ
457
В конце 70-х годов ХХ века двумя группами ис-
СВЯЗЫВАНИЕ ПРОТЯЖЕННЫХ ЛИГАНДОВ
следователей был обнаружен аллостерический
НА ПОЛИМЕРЕ
эффект ДНК — возможность распространения
Пусть в растворе при постоянной температуре
вдоль ДНК кооперативных взаимодействий меж-
и постоянном давлении находятся молекулы ли-
ду адсорбированными лигандами. В работах оте-
ганда и молекулы полимера с адсорбированными
чественных исследователей этот эффект называ-
лигандами. С точки зрения физики процесс обра-
ли дальнодействием и описывали при помощи
тимого связывания лигандов на поверхности
потенциала кооперативных взаимодействий [8,
макромолекулы представляет собой явление фи-
9]. Аллостерические взаимодействия лежат в ос-
зической адсорбции и относится к классу реше-
новании механизмов регуляции генов (см., на-
точных моделей: решетка-полимер состоит из ре-
пример, работы [10-13]).
акционных центров (сайтов), которые могут свя-
зывать лиганд [26, 29].
Дальнейшее развитие теории адсорбции в
применении к связыванию лигандов с нуклеино-
Обозначим число лигандов, адсорбированных
выми кислотами позволило учесть ситуации, ко-
на полимере в равновесии, как q, а длину полиме-
гда лиганды образуют различные типы комплекса
ра - N. Предположим, что лиганд протяженный,
или связываются на ДНК в различных ориента-
то есть он при связывании закрывает L звеньев
циях [14, 15]. В русле этого подхода для широкого
полимера, делая их недоступными для связыва-
класса адсорбционных задач применялась одно-
ния других молекул лиганда.
мерная модель Изинга [16]. Заметим, что эта мо-
В случае L << N заполнение полимера лиган-
дель для описания ферромагнетизма была пред-
дом r (которое определяется как q/N) и концен-
ложена в 1920 г. В. Ленцем [17]. Э. Изинг смог
трация свободного лиганда в растворе х связаны
найти решение уравнений, которые возникают в
следующим соотношением [5]:
соответствующей этой модели математической
задаче (поэтому в ряде источников используется
L
r
1-
rL
название «модель Ленца-Изинга») [18]. Исполь-
=
1−
rL + r),
(1)
(
)(
Kx
1−
rL + r
зование математического решения задачи ад-
сорбции для этой модели позволило получить си-
где K — равновесная константа связывания ли-
стему уравнений адсорбции в самом общем виде
ганда с одним связывающим местом на полимере.
[16, 19, 20].
Модель, описывающая связывание протяжен-
Заседателев, Гурский и Волькенштейн полу-
ного лиганда на полимере, рассматривалась и до
чили уравнения, описывающие кооперативное
работ Заседателева с соавторами, соответствую-
связывание протяженного лиганда на полимере,
щие уравнения были получены ранее в работах
[1-3], однако эти уравнения имели другую, пара-
однако они не рассматривали дисперсию адсорб-
ции (так как воспользовались приближением
метрическую форму записи. Уравнение (1) мож-
но назвать центральным уравнением в теории ад-
«бесконечного полимера»). Позже И. Эпштейн
сорбции протяженных лигандов. В англоязычной
показал, что приближение бесконечного полиме-
литературе за ним закреплено имя «уравнение
ра хорошо описывает ситуацию, когда длина
МакГи-фон Хиппеля», так как эти авторы объяс-
полимера уже в десяток раз превышает длину ли-
нили смысл данного уравнения при помощи
ганда [21]. В ряде работ было показано, что дис-
условных вероятностей. Однако первыми в таком
персия адсорбции может быть весьма информа-
виде уравнения записали отечественные исследо-
тивной характеристикой системы [22-25].
ватели (как и уравнения, описывающие коопера-
При связывании на матрицах нуклеиновых
тивные взаимодействия [5]). Позже было показа-
кислот может быть задействовано одновременно
но, что уравнение (1) может быть использовано
несколько систем реакционных центров: в живой
для построения асимптотического метода анали-
клетке с однотяжевыми и двутяжевыми участка-
за изотерм адсорбции и для классификации моде-
лей адсорбции [26, 30].
ми молекул ДНК и РНК связывается большое
число различных соединений. В монографии [26]
Интерпретировать уравнения адсорбции в
рассматривается как конкуренция лигандов за
рамках вероятностного подхода можно следую-
одни и те же связывающие места, так и координа-
щим образом (см., например, работу [26]). Рас-
ция при связывании лигандов на одной или двух
смотрим полимер с адсорбированными лиганда-
системах реакционных центров. Эта работа в не-
ми как результат проведения схемы испытаний
котором смысле содержит итог развития теории
Бернулли с двумя исходами. Введем в рассмотре-
адсорбции в применении к описанию связывания
ние вероятность встретить в последовательности
лигандов с нуклеиновыми кислотами с 70-х годов
из свободных звеньев полимера и адсорбирован-
прошлого века, в нее вошли как оригинальные
ных лигандов свободное звено (не закрытое ли-
работы, так и ряд обзоров [27, 28].
гандом):
БИОФИЗИКА том 67
№ 3
2022
458
НЕЧИПУРЕНКО, БУЧЕЛЬНИКОВ
1−
rL
Наиболее полную информацию об избира-
P
=
(2)
тельном связывании лигандов на разных местах
1-
rL
+
r
ДНК дают диаграммы футпринтинга, на которых
Тогда уравнение (1) можно записать в следую-
представлены данные о том, как связанный ли-
щем виде:
ганд закрывает отдельные межнуклеотидные свя-
L
зи в ДНК от расщепления разного рода агентами.
1
P
=
(3)
Ранее в работах [33, 34] было показано, как мож-
Kx
1−
P
но рассчитывать такие диаграммы и сравнивать
Параметр P изменяется от 0 до 1, и если зада-
результаты с экспериментальными данными в ра-
вать значение этого параметра, то можно из урав-
ботах (см. также монографию [26], где данному
нений (2) и (3) легко найти значения x и r и по-
анализу посвящена отдельная глава).
строить изотерму адсорбции.
Вероятностная интерпретация уравнения (1)
ПРИМЕНЕНИЕ ТЕОРИИ АДСОРБЦИИ
заключается в том, что правая часть уравнения
ДЛЯ ОПИСАНИЯ БИОЛОГИЧЕСКИХ
соответствует относительному числу свободных
СИСТЕМ
связывающих мест на полимере с адсорбирован-
ными лигандами (одно место составляют L рас-
Математические модели позволяют количе-
положенных подряд не закрытых лигандом зве-
ственно описать процессы, происходящие в жи-
ньев полимера). Величина r дает относительное
вой клетке и в модельных системах при связыва-
число адсорбированных лигандов. В таком случае
нии лигандов с ДНК. Кооперативные взаимодей-
уравнение (1) можно рассматривать как уравне-
ствия между лигандами были обнаружены и
ние химического равновесия для лиганда
описаны в работах [9, 10, 26, 35]. Позже были по-
длины L.
лучены данные, подтверждающие участие таких
В реальной ситуации полимер имеет не беско-
взаимодействий в процессах генетической регу-
нечную длину, благодаря этому в системе наблю-
ляции, и Д. Крозерс уделил этому особое внима-
дается дисперсия: разные полимеры в растворе
ние [36] (см. также специальный номер журнала
несут разное число адсорбированных лигандов.
«Biopolymers», посвященный его памяти [37]).
Ранее было показано, как построить схему испы-
Методы статистической термодинамики при-
таний Бернулли и оценить сходимость величины
менялись для анализа широкого класса систем:
q/N к значению r. Эту величину можно интерпре-
рассматривалось связывание белков и антибио-
тировать как вероятность события, что опреде-
тиков с ДНК [38], проводился анализ расположе-
ленное звено полимера занято лигандом [26].
ния нуклеосом на сателлитной ДНК [39], было
Заметим, что уравнения (1)-(3) являются до-
описано связывание матричной РНК с рибосо-
вольно простыми алгебраическими уравнения-
мальными РНК в составе рибосомы [40]. Также
ми, и при усложнении моделей адсорбции - учете
эти методы были использованы для анализа ко-
образования разных типов комплекса и коопера-
оперативных взаимодействий лигандов с рибосо-
тивных взаимодействий — вместо уравнений (1)-
мой [41, 42], для описания шума экспрессии генов
(3) появляется более сложная система уравнений
[25, 43], для анализа вхождения малых РНК в
(см., например, работы [19, 26, 31, 32]).
комплекс РНК-индуцированного сайленсинга
Отдельно следует упомянуть об избиратель-
генов RISC [44] и анализа данных по связыванию
ном связывании лигандов с ДНК, когда разным
олигонуклеотидов на микрочипах [29, 45-49].
связывающим местам соответствуют разные
Множество работ в области применения тео-
энергии связывания. В этом случае принято гово-
рии адсорбции к описанию связывания лигандов
рить о связывании на гетерополимере. Для опи-
с нуклеиновыми кислотами контрастирует с от-
сания такого связывания вместо одной констан-
сутствием обобщающих монографий в этой обла-
ты химического равновесия K, описывающей
сти. Хотелось бы видеть современные труды, где
связывание лиганда с единичным связывающим
были бы описаны разные аспекты такого связы-
местом на ДНК, вводят в рассмотрение набор
вания, подобно тому, как описано связывание
констант {Ki} и набор соответствующих измене-
лигандов с белками в монографии Дж. Ваймана и
ний свободной энергии Гиббса {ΔGi}, которые
С. Гилла
[29]. В одной из глав монографии
связаны соотношением Ki = exp(-ΔGi/RT). Ана-
Г. Стормо рассматриваются термодинамические
аспекты связывания белков с ДНК [50], однако в
лиз связывания и расчет уравнений адсорбции в
ней нет систематического изложения методов
этом случае удобно проводить при помощи по-
анализа такого связывания.
строения большой статистической суммы ад-
сорбционной системы или с использованием ме-
Заметим, что относительно недавно вышел
тода рекуррентных соотношений [6]. Пример
ряд работ, в которых содержится описание си-
применения модели адсорбции лигандов на гете-
стем, где связывание лигандов с матрицами нук-
рополимере приведен в «Приложении».
леиновых кислот происходит по новым схемам
БИОФИЗИКА том 67
№ 3
2022
СВЯЗЫВАНИЕ ЛИГАНДОВ С НУКЛЕИНОВЫМИ КИСЛОТАМИ
459
(см., например, работы [51, 52]). Развиты новые
новых методов и технологий в молекулярной
модели адсорбции, получены уравнения связыва-
биологии появилась надежда решить и фунда-
ния. В монографии [26] такие модели впервые из-
ментальные задачи: разобраться в том, как рабо-
ложены системно: с единой точки зрения впер-
тают факторы транскрипции, малые РНК и ле-
вые описана совокупность методов и подходов
карственные соединения, которые регулируют
статистической термодинамики, позволяющих
процессы экспрессии генов. Теоретические и
анализировать системы, в которых изучается свя-
экспериментальные методы, разработанные за
зывание лигандов с нуклеиновыми кислотами in
последние годы, дают возможность вплотную по-
vitro. На ряде примеров показано, как такие под-
дойти к решению этих задач. Заметим, что кон-
ходы работают, какие параметры связывания они
цептуально молекулярная биология уже созрела
позволяют оценить.
для количественного подхода к описанию регуля-
ции генов (см., например, работу [68]) и, как мы
Методы анализа данных в области теории ад-
отмечали во «Введении», аллостерические взаи-
сорбции дают в руки исследователя разнообраз-
модействия между лигандами могут играть важ-
ный теоретический «инструментарий», позволя-
нейшую роль в такой регуляции [69].
ющий анализировать как реальные биологиче-
ские системы, так и модельные системы, в
Для строгого термодинамического описания
которых изучается связывание лигандов с нукле-
кооперативного связывания необходимо пони-
иновыми кислотами. Примером такого инстру-
мание того, как изменяются функции состояния
ментария является модель Изинга, которая хоро-
адсорбционной системы, среди которых ключе-
шо известна в самых разных приложениях стати-
вую роль, по нашему мнению, играет статистиче-
стической физики. Любопытно, что развитие
ская, или конфигурационная, энтропия. Рас-
теории связывания лигандов с двумерными плат-
смотрим далее, как описанный формализм мож-
формами микрочипов закономерно привело к
но применить для расчетов энтропии на примере
применению двумерной модели Э. Изинга [29,
простейшей модельной системы, в которой суще-
45-49].
ствуют кооперативные взаимодействия между ад-
сорбированными лигандами.
Детальный анализ связывания биологически
активных соединений с нуклеиновыми кислота-
ми содержится в работах М.П. Евстигнеева с со-
СТАТИСТИЧЕСКАЯ ЭНТРОПИЯ
авторами [53-55], отдельно следует упомянуть
ПРИ ОПИСАНИИ КООПЕРАТИВНОГО
оригинальную концепцию интерцепторно-про-
СВЯЗЫВАНИЯ
текторного действия ДНК-связывающих лиган-
дов [53], развиваемую этими авторами.
Ниже мы постараемся изучить поведение ста-
тистической энтропии на простом примере ад-
Обширный класс фундаментальных и при-
сорбции малых молекул на линейном полимере и
кладных задач молекулярной биофизики можно
показать, что расчеты энтропии позволяют про-
решать при помощи теории адсорбции. Любо-
лить свет на происходящие в системе процессы.
пытны новые подходы, которые позволяют свя-
Заметим, что данный обзор концептуально свя-
зать адсорбцию с конденсацией ДНК [56-60] и с
зан с работой [70], вышедшей пять лет назад. В
плавлением ДНК (см., например, работу [61]),
настоящей публикации мы расширили и обнови-
исследовать противовирусную активность лиган-
ли список литературы, а также включили расчеты
дов (см., например, работу [62]), связывание ли-
конфигурационной энтропии [71].
гандов с квадруплексами [63-65]. Особый инте-
рес представляют работы, в которых изучается
В дополнение к величинам K и x, введенным
кооперативное связывание белков с ДНК (см.,
выше при рассмотрении модели адсорбции ли-
например, работу [66]).
гандов на гомополимере, обозначим через ω па-
раметр кооперативности. Значения ω < 1 соот-
Статистическая термодинамика, описываю-
ветствуют отрицательной кооперативности, или
щая связывания лигандов с ДНК и РНК, нахо-
антикооперативности; при ω = 1 адсорбция неко-
дится на переднем крае науки, в области развития
оперативна, параметр ω > 1 соответствует поло-
новых технологий постгеномной эры. Модели ад-
жительной кооперативности [72].
сорбции оказываются необходимыми и востребо-
ванными в анализе экспериментов по футприн-
Все термодинамические параметры системы, в
тингу [33, 34], в объяснении экспериментов по
том числе статистическую энтропию, можно рас-
считать, зная большую статистическую сумму си-
гибридизации геномных ДНК и РНК на микро-
чипах [67], в описании регуляции экспрессии ге-
стемы ΞN. Существует несколько методов нахож-
нов при помощи малых РНК (см., например, ра-
дения большой статистической суммы системы, в
боту [44]), при конструировании новых лекар-
которой происходит адсорбция лигандов на по-
ственных соединений и создании биосенсоров
лимерной решетке. Среди них матричный [18] и
[47, 51] - т. е. во многих областях молекулярной
комбинаторный [73] методы являются наиболее
биологии и биотехнологии. В связи с развитием
известными.
БИОФИЗИКА том 67
№ 3
2022
460
НЕЧИПУРЕНКО, БУЧЕЛЬНИКОВ
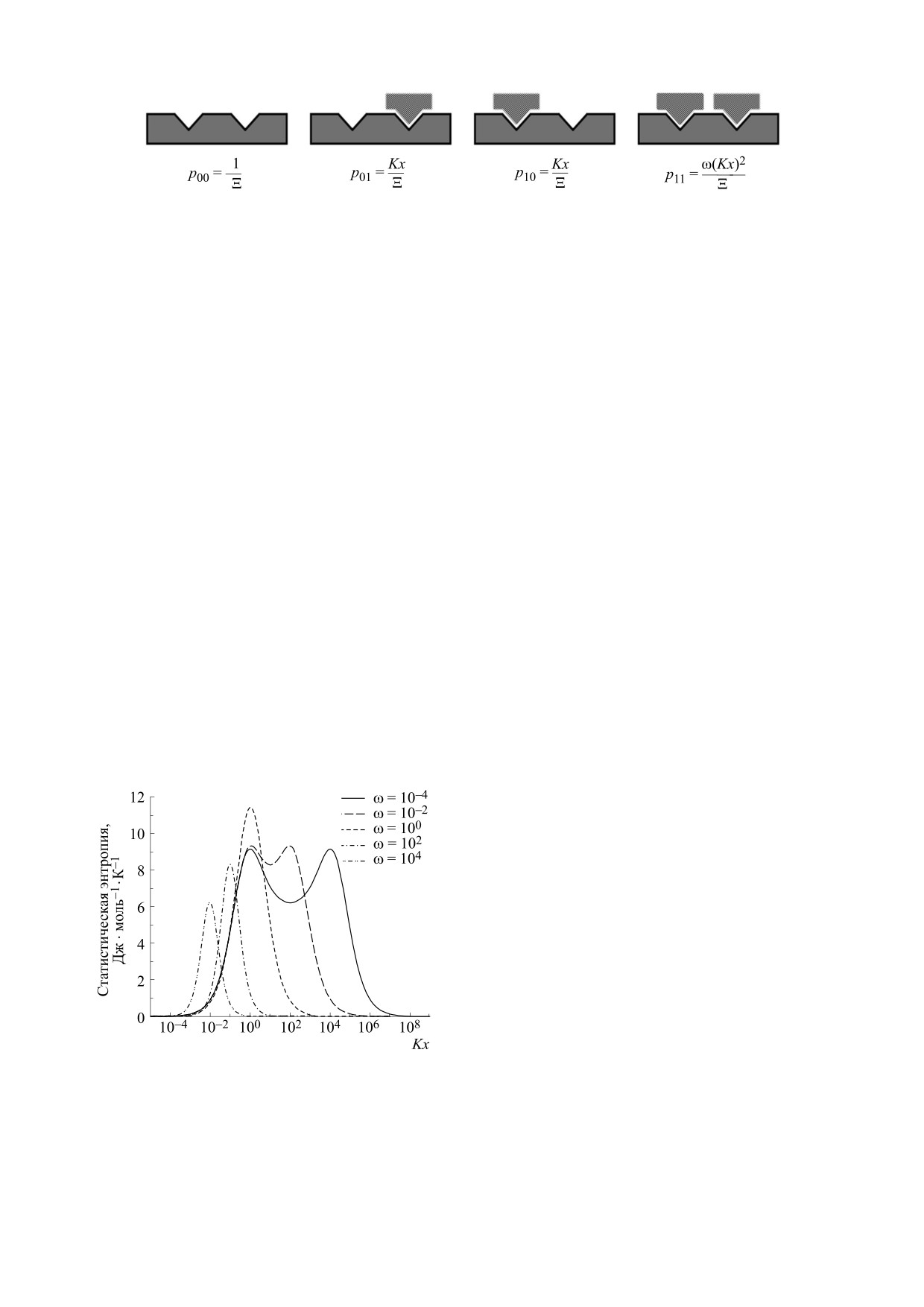
Рис. 1. Показаны разные состояния простейшей решетки и соответствующие им вероятности.
Рассмотрим простейшую систему с полиме-
S = -R(p00lnp00 + p01lnp01 + p10lnp10 + p11lnp11)S,
ром, имеющим только два центра связывания
или
(N = 2). Большая статистическая сумма такой си-
стемы может быть записана следующим образом:
Kx
S R
(2(1+ωKx)
lnKx +ωKx
lnω)
−
lnΞ
(5)
Ξ = 1 + 2Kx + ω(Kx)2.
Ξ
Формула (5) позволяет представить статисти-
Определим статистическую, или конфигура-
ческую энтропию как функцию двух независи-
ционную энтропию исследуемой системы как ме-
мых переменных - S(Kx, ω). Зависимость энтро-
ру неопределенности, возникающую при адсорб-
пии от величины Kx при разных значениях ω по-
ции лигандов на решетке. Действительно, если
казана на рис. 2 .
известно, что на одной решетке связан один ли-
ганд, существует неопределенность: какой имен-
Мы видим, что зависимость энтропии от кон-
но из двух реакционных центров занят лигандом,
центрации лиганда в растворе и от заполнения
матрицы лигандом оказывается нетривиальной:
а какой свободен? Из статистической термодина-
мики известно, что такая энтропия S в расчете на
при значениях параметра кооперативности ω < 1
моль решеточного полимера равна:
график имеет два максимума.
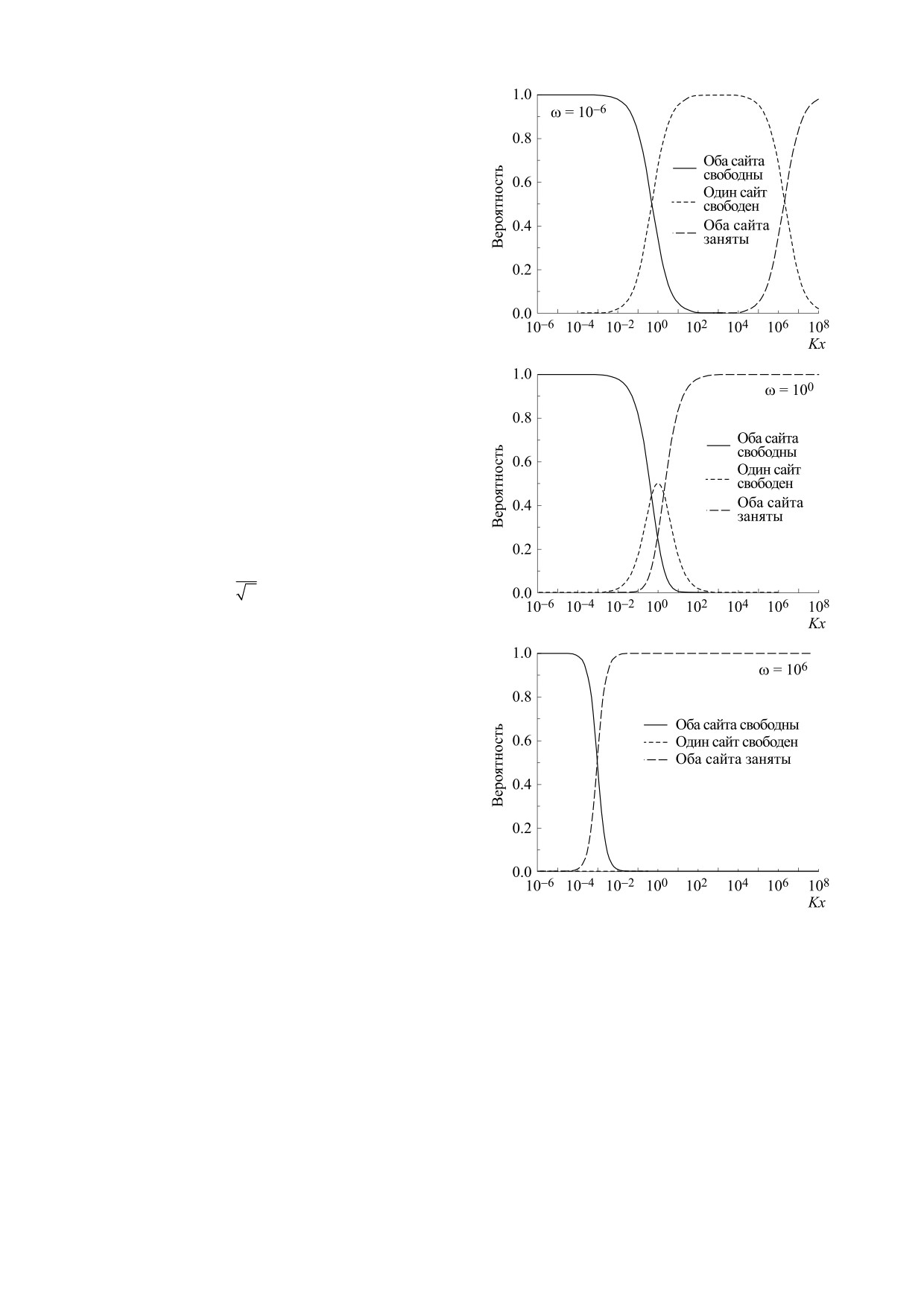
На рис. 3 показаны зависимости плотности ве-
1
1
роятности состояний решетки от Kx. При малом
S =−R
p
ln
p
,
(4)
ij
ij
параметре кооперативности (ω << 1) три состоя-
i=0
j=0
ния последовательно сменяют друг друга. По ме-
где R - универсальная газовая постоянная, pij -
ре увеличения концентрации лиганда начинает
вероятность пребывания системы в одном из че-
снижаться вероятность того, что решетка полно-
тырех возможных состояний. Всего в рассматри-
стью пуста, и увеличиваться вероятность того, что
ваемой системе возможны четыре состояния,
хотя бы один центр связывания решетки занят. С
укажем для каждого из них свою вероятность (см.
дальнейшим ростом концентрации лиганда уве-
pис. 1).
личивается вероятность полностью занятой ре-
Найдем энтропию системы в явном виде на ос-
шетки. В пределе, при ω → 0, можно ожидать, что
новании формулы (4):
последнее событие наступит при бесконечно
большой концентрации лиганда, что невозмож-
но, и это вполне отвечает нашим представлени-
ям. Действительно, при бесконечно антикоопе-
ративном процессе существует запрет на суще-
ствование двух соседних занятых сайтов - модель
«исключенного соседа» [74]. Эта модель может
служить переходом к решеточной модели «газа
твердых стержней», в которой один лиганд при
связывании занимает сразу несколько реакцион-
ных центров [2-4].
При увеличении параметра кооперативности
вероятность полного заполнения решетки лиган-
дами наступает уже при меньших концентрациях.
При этом состояние решетки с одним занятым
сайтом становится все менее вероятным. При
дальнейшем увеличении параметра кооператив-
ности вероятность встретить в растворе наполо-
вину заполненную решетку падает практически
до нуля. В пределе, при ω → ∞ состояние системы
Рис. 2. Зависимость статистической энтропии от произ-
ведения Kx при различных значениях параметра коопе-
определяется двумя возможными конфигурация-
ративности ω.
ми: полностью пустая либо полностью заполнен-
БИОФИЗИКА том 67
№ 3
2022
СВЯЗЫВАНИЕ ЛИГАНДОВ С НУКЛЕИНОВЫМИ КИСЛОТАМИ
461
ная решетка, причем смена конфигураций, т. е.
кооперативный переход, происходит при беско-
нечно малой концентрации лиганда.
Из сопоставления рис. 2 и 3 следует важный
вывод: энтропия системы максимальна в месте
пересечения плотностей вероятности, т.е. когда
соответствующие события равновероятны. Рав-
ная вероятность наступления событий означает
отсутствие преимущественного состояния ре-
шетки и приводит к неопределенности, количе-
ственной мерой которой является статистическая
энтропия. При отрицательной или положитель-
ной кооперативности процессы связывания ли-
гандов с решеткой зависят друг от друга, в то вре-
мя как при некооперативном связывании эти
процессы протекают независимо. По этой причи-
не наибольшее значение энтропии в максимуме
(см. рис. 2 и 3) наблюдается именно для некоопе-
ративного процесса (ω = 1).
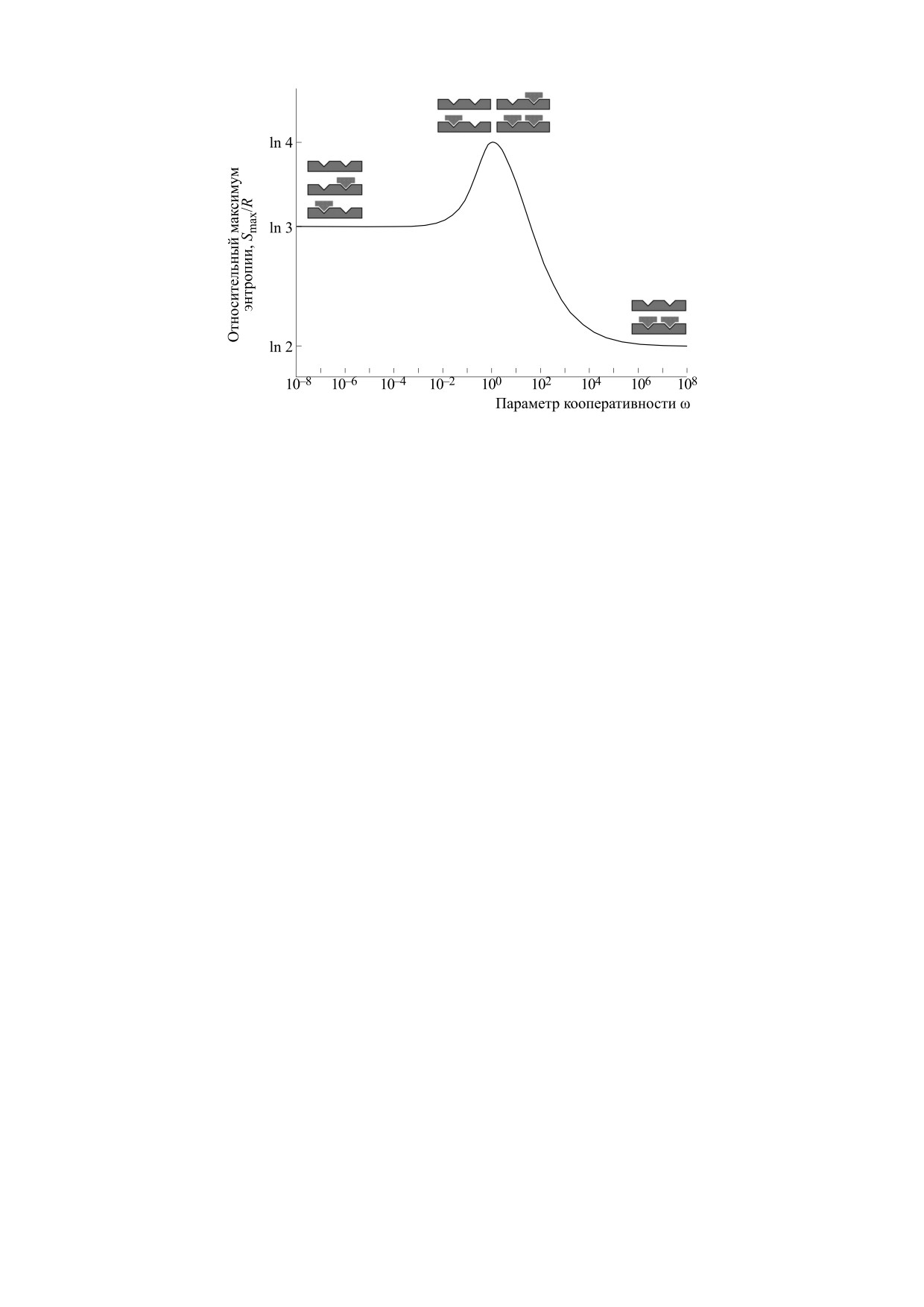
Интересно рассмотреть зависимость макси-
мальных значений энтропии Smax, т.е. значений
энтропии в пике каждого графика на рис. 2, от па-
раметра кооперативности ω. Можно показать,
что
S
1,ω
при
ω≤
1,
Kx
=
1
(
)
ω
S
(Kx,ω)
max
=
1
S
,ω
при
ω≥
1
(
)
ω
График зависимости максимальных значений
энтропии с точностью до универсальной газовой
постоянной от параметра кооперативности изоб-
ражен на рис. 4.
Из рисунка видно, что максимальная стати-
стическая энтропия соответствует системе с не-
кооперативными взаимодействиями (ω = 1), в
которой равновероятны все четыре состояния
системы. По мере уменьшения параметра коопе-
ративности (ω << 1) состояние с полностью за-
полненной решеткой становится все менее веро-
ятным, а при увеличении параметра кооператив-
ности (ω >> 1) все менее вероятными становятся
состояния с наполовину заполненной решеткой.
Как видно из рис. 4, максимальная энтропия
системы с одним молем решеточных полимеров с
точностью до постоянной Авогадро находится в
полном согласии с уравнением Больцмана:
S = RlnΩ = NAklnΩ,
Рис. 3. Зависимости плотностей вероятности состояний
где Ω — термодинамическая вероятность, т. е.
решетки от концентрации лиганда.
число микроскопических состояний системы,
которым можно реализовать данное макроскопи-
ческое состояние, в нашем случае - число кон-
вым и вторым центрами может происходить с
фигураций адсорбционной системы при данном
разными константами. Пример такого случая мы
значении параметра кооперативности.
рассмотрели в работах [41, 42].
Заметим, что в реальных системах, т.е. при
При анализе экспериментальных данных ис-
связывании лигандов с рецепторами, субстратов
следователю необходимо решать так называемую
с рибосомами и др., связывание лигандов с пер-
обратную задачу: восстановить параметры моде-
БИОФИЗИКА том 67
№ 3
2022
462
НЕЧИПУРЕНКО, БУЧЕЛЬНИКОВ
Рис. 4. Зависимость максимума энтропии от параметра кооперативности.
ли на основании кривых связывания. В этом слу-
молекулярных биологов и биофизиков к теории
чае возникает неопределенность, и параметр ко-
адсорбции: по сути, она помогает решать множе-
оперативности можно определить только при
ство актуальных задач, о чем мы писали выше. В
определенных допущениях. Существует также
отечественной и мировой литературе почти не су-
известная проблема, связанная с тем, что резуль-
ществует крупных обзорных работ, в которых с
таты расчета кривых в модели, в которой суще-
единой точки зрения рассматривались бы сотни
ствуют антикооперативные взаимодействия меж-
примеров успешного применения теории адсорб-
ду лигандами, совпадают с результатами расчетов
ции к описанию взаимодействия лигандов с нук-
в случае, если есть гетерогенное связывание, но
леиновыми кислотами. Наша работа продолжает
нет кооперативности. В этом случае на помощь
серию обзоров и глав в монографиях, в которых
исследователю могут прийти эксперименты по
мы ставим перед собой цель заполнить лакуны в
кинетике связывания [75].
описании применения теории адсорбции к свя-
зыванию лигандов с нуклеиновыми кислотами
Ниже, в «Приложении», мы продемонстриру-
[26, 28, 70].
ем применение методов статистической термоди-
намики к анализу связывания на платформах
Теория адсорбции применялась к биологиче-
микрочипов молекул матричных РНК в ситуа-
ским системам на протяжении всего ХХ века и
ции, когда необходимо провести различие между
накопила уже достаточно опыта, чтобы высту-
высокоспецифичным связыванием (сигналом) и
пить в этой области в роли научной дисциплины.
низкоспецифичным (шумом). Заметим, что дан-
Эта дисциплина выработала свой язык, который
ное «Приложение» может иметь большое практи-
позволил сформулировать взгляд на такую фун-
ческое значение для анализа данных по связыва-
даментальную проблему биологии, как регуляция
нию мишеней на биочипах. Кроме известных за-
генетической экспрессии, в рамках представле-
рубежных работ в этой области и работ, которые
ний статистической физики (см., например, ра-
мы используем для развития методов анализа
боты [25, 36, 37, 43]). С другой стороны, примене-
данных [67, 76-78], следует упомянуть примеры
ние теории адсорбции к анализу эксперименталь-
успешного применения биочипов в Институте
ных данных позволяет связать наблюдаемые
молекулярной биологии им. В.А. Энгельгардта
величины с модельными свойствами молекуляр-
РАН (см., например, работы [79-83]).
но-биологических систем: определять энергии
взаимодействия лигандов с нуклеиновыми кис-
лотами, энергии взаимодействия между адсорби-
ЗАКЛЮЧЕНИЕ
рованными лигандами и т.п. Описание реальных
Применение теории адсорбции для описания
систем при помощи теории адсорбции дает воз-
биологических систем обусловлено участием спе-
можность как предсказывать их поведение при
циалистов в области статистической физики в ре-
изменении внешних условий, так и уточнять чер-
шении задач, связанных с биополимерами. В на-
ты моделей связывания лигандов с ДНК. Без при-
стоящее время необходимо привлечь внимание
менения теории адсорбции невозможно сейчас
БИОФИЗИКА том 67
№ 3
2022
СВЯЗЫВАНИЕ ЛИГАНДОВ С НУКЛЕИНОВЫМИ КИСЛОТАМИ
463
представить эффективное решение и таких при-
нями, которые не содержали мест специфичного
кладных задач, как анализ связывания биологи-
связывания, в нашей системе будет присутство-
чески активных соединений на микрочипах и со-
вать мишень со специфичным местом i = s.
здание биосенсоров.
Молекула зонда может пребывать в N + 1 со-
стоянии: либо она будет свободна, либо связана с
одной мишенью из набора N типов (на который
ПРИЛОЖЕНИЕ
мы разбили реальную мишень). Методы стати-
Сравнение кросс-гибридизации со специфиче-
стической термодинамики позволяют оценить
ской гибридизацией. Рассмотрим эксперимен-
вероятность того, что зонд будет связан с i-й ми-
тальную систему, в которой происходит гибри-
шенью. Связывание зонда с i-й мишенью харак-
дизация
- связывание молекул-мишеней из
теризуется равновесной константой Ki.
раствора с олигонуклеотидами-зондами, распо-
Воспользуемся формализмом статистических
ложенными на платформе микрочипа [67] (см.
сумм. Представим большую статистическую сум-
также работу [76]). Заметим, что под терминами
му системы в виде:
микрочип и биочип мы понимаем здесь одно и то
же: матрицы ячеек на плоской поверхности, в
Ξ
=
1+K
x,
N
i
каждой из которых иммобилизованы молекуляр-
i
ные зонды определенного типа.
где x - концентрация свободных молекул мише-
ней в растворе, статистический вес 1 соответству-
Для оценки специфичности гибридизации
ет ситуации, когда мишень не связана с зондом.
здесь мы будем использовать две базы данных,
Тогда вероятность обнаружить зонд связанным с
полученных в двух эквивалентных эксперимен-
i-й мишенью равна
тальных системах, представляющих микрочипы с
одними и теми же пробами [67]. В первой системе
d
lnΞ
N
K
i
x
к микрочипу добавляются мишени, которые име-
P
i
=
=
d
lnK
i
1
K
i
x
ют специфические места связывания для проб, во
+
i
второй - те же самые мишени, только без специ-
Если рассматривается связывание со специ-
фических мест связывания (что можно считать
контролем). Соответственно, в первой базе пред-
фичной матрицей i = s, то
ставлены данные для проб, каждая из которых
K
x
s
может находиться в одном из трех состояний:
Ps
=
1
K
x
быть свободной (не связанной с мишенью) (I),
+
i
i
быть связанной специфично (II) и связанной не-
специфично (III). Во второй базе представлены
Здесь в знаменателе в общей сумме есть член,
данные для проб, для которых нет специфичных
соответствующий j = s, то есть Ks x.
мишеней - и для них возможны только состоя-
Вероятность кросс-гибридизации равна:
ния (I) и (III).
K
x
Рассмотрим связывание одной молекулы зон-
j
j≠s
да с мишенью для первой экспериментальной си-
P
=
(A1)
cross
1
K
x
стемы. Будем считать, что это связывание прохо-
+
i
i
дит в равновесных условиях. Для того, чтобы оце-
нить вероятность связывания зонда на мишени
Если мы сигналом будем называть вероят-
по специфическому механизму, с образованием
ность связывания зонда на специфическом ме-
совершенного дуплекcа, по сравнению с вероят-
сте, а шумом — вероятность кросс-гибридизации,
ностями связывания на всех неспецифических
то оценку величины отношения «сигнал-шум»
местах (приводящего к кросс-гибридизации),
дает отношение соответствующих вероятностей:
применим следующий прием.
P
s
K
s
x
R
=
Предположим, что каждая длинная молекула
SN
P
K
x
cross
j
=
мишени связывается только с одним зондом. В
j≠s
этом случае мы можем представить, что молекула
Для интерпретации экспериментальных дан-
зонда равновесно связывается с набором моле-
ных, полученных в двух описанных выше систе-
кул-мишеней, каждая из которых имеет свой но-
мах, то есть на двух микрочипах, применим наш
мер i =1, …, N. Таким образом, мы заменяем ре-
подход.
альную молекулу мишени на набор коротких мо-
лекул, каждая из которых соответствует одному
Вероятность кросс-гибридизации Pcross мы
месту связывания с молекулой зонда. Подобную
можем оценить на основании данных из экспери-
схему связывания рассматривали ранее авторы
мента во второй системе, где зонды связываются
работы [77]. Однако в отличие от этой работы, где
с мишенями, в которых нет специфического ме-
оценивалось связывание только зондов с мише-
ста. Если предположить, что в этой системе экс-
БИОФИЗИКА том 67
№ 3
2022
464
НЕЧИПУРЕНКО, БУЧЕЛЬНИКОВ
перимент проводится при аналогичных условиях,
факторов может позволить уточнить оценки от-
то весь сигнал гибридизации в этом эксперимен-
ношения «сигнал-шум» в рассматриваемой си-
те будет равен сигналу кросс-гибридизации:
стеме.
Заметим, что оценка специфического и неспе-
K
x′
j
цифического связывания была проведена в рабо-
j≠s
P′
=
,
те [78], авторы которой развивали модель, сход-
cross
1
K x′
+
i
ную с нашей. Однако в этой работе сумму кон-
i≠s
стант, соответствующих неспецифическому
где x´ - концентрация свободных мишеней во
связыванию, заменяли на одну константу с усред-
второй системе. Если предположить, что концен-
ненной энергией.
трации мишеней в этих системах мало отличают-
Мы описали здесь модель, в которой молекула
ся, т. е. что x´ ~ x, то, используя соотношение
зонда может находиться в одном из трех состоя-
(А1), можно показать, что отношение сигнала
ний: быть свободной (I), связываться со специ-
кросс-гибридизации в первой системе к сигналу
фическими мишенями (II), либо с неспецифиче-
кросс-гибридизации во второй системе равно:
скими (III), и эта модель применялась для анали-
за данных в работе [67].
1+
K
x
j
P
cross
j≠s
=
(A2)
P
′
1
K
x
БЛАГОДАРНОСТИ
cross
+
i
i
Авторы считают своим приятным долгом по-
Перейдем к экспериментальным оценкам. В
благодарить А.С. Заседателева, Н.Г. Есипову и
первой системе зонд может находиться в трех со-
В.В. Аристова за полезные дискуссии, А. Анаш-
стояниях: он может быть не связан с мишенью
кину и О. Матвееву - за ценные замечания и об-
(обозначим вероятность этого состояния как P0),
суждение.
быть связанным со специфичным местом (соот-
ветствующая вероятность равна Ps) и быть свя-
ФИНАНСИРОВАНИЕ РАБОТЫ
занным неспецифично (соответствующая веро-
Работа выполнена при частичной финансовой
ятность равна Pcross):
поддержке Программы фундаментальных науч-
P
+P
+P
= 1.
ных исследований в Российской Федерации на
0
s
cross
долгосрочный период
(2021-2030 годы) (тема
Сигнал от общей гибридизации в первой си-
121052600299-1).
стеме пропорционален величине Ps + Pcross. Вы-
читая из этого сигнала
P
′
, мы получаем неко-
cross
КОНФЛИКТ ИНТЕРЕСОВ
торую оценку для Ps, которая в соответствии с со-
Авторы заявляют об отсутствии конфликта
отношением (А2) оказывается тем точнее, чем
интересов.
меньше отношение сигнала к шуму RSN. По сути,
тогда будет сделано следующее приближение: мы
считаем полный набор состояний в первой систе-
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
ме для эксперимента с микрочипом, когда в ми-
Настоящая статья не содержит каких-либо ис-
шени есть специфические места. Потом сигнал
следований с участием людей или животных в ка-
кросс-гибридизации на этом чипе (его оценивает
честве объектов исследований.
формула (А1)) мы заменяем на сигнал кросс-ги-
бридизации, полученный в эксперименте, где нет
специфичного места [67]. Уточнить оценки пара-
СПИСОК ЛИТЕРАТУРЫ
метров мы сможем, если учтем детали экспери-
1. D. F. Bradley and S. Lifson, in Molecular Associations
мента. Следует оценить отличие x´ и x в этих си-
in Biology, Ed. by B. Pullman (Acad. Press, London,
стемах, которое обусловлено двумя факторами:
New York, 1968), pp. 261-270.
во-первых, зонды, специфично связанные с ми-
2. S. A. Latt and H. A. Sober, Biochemistry 6 (10), 3293
шенями, выключают из реакции протяженные
(1967).
участки ДНК, которые прилегают к специфиче-
3. D. M. Crothers, Biopolymers 6 (4), 575 (1968).
скому месту связывания и тем самым уменьшают
4. L. Tonks, Phys. Rev. 50 (10), 955 (1936).
эффективное значение x, во-вторых, в системе,
5. А. С. Заседателев, Г. В. Гурский и М. В. Волькен-
где нет участков ДНК со специфичными местами
штейн, Молекуляр. биология 5 (20), 245 (1971).
связывания, вообще уменьшается общая концен-
6. Г. В. Гурский, А. С. Заседателев и М. В. Волькен-
трация мишеней в растворе, которая равна сумме
штейн, Молекуляр. биология 6, 479 (1972).
концентраций мишеней, связанных на зондах и
7. J. D. McGhee and P. H. von Hippel, J. Mol. Biol. 86
свободных мишеней в растворе. Учет двух этих
(2), 469 (1974).
БИОФИЗИКА том 67
№ 3
2022
СВЯЗЫВАНИЕ ЛИГАНДОВ С НУКЛЕИНОВЫМИ КИСЛОТАМИ
465
8.
A. S. Krylov, S. L. Grokhovsky, A. S. Zasedatelev
41.
S. B. Bourd, M. K. Kukhanova, B. P. Gottikh, et al.,
et al., Nucl. Acids Res. 6 (1), 289 (1979).
Eur. J. Biochem. 135 (3), 465 (1983).
9.
M. Hogan, N. Dattagupta, and D. M. Crothers, Na-
42.
М. К. Куханова, С. Б. Бурд, Л. С. Викторова и др.,
ture 278, 521 (1979).
Молекуляр. биология 18 (3), 691 (1984).
10.
X. Qu, J. O. Trent, I. Fokt et al., Proc. Natl. Acad. Sci.
43.
М. В. Головкин, Ю. Д. Нечипуренко и Г. В. Гур-
USA 97, 12032 (2000).
ский, Биофизика 54 (4), 581 (2009).
11.
D. M. Chenoweth and P. B. Dervan, Proc. Natl. Acad.
44.
O. Matveeva, Yu. Nechipurenko, L. Rossi, et al.,
Sci. USA 106, 13175 (2009).
Nucl. Acids Res. 35 (8), e63 (2007).
12.
A. Balaceanu, A. Pérez, P. D. Dans et al. Nucl. Acids
45.
М. В. Головкин, О. В. Матвеева и Ю. Д. Нечипу-
Res. 46 (15), 7554 (2018).
ренко, Биофизика 54 (5), 820 (2009).
13.
J. Dale, C. P. Howe, H. Toncrova, et al., Phys. Chem.
46.
М. В. Ходыков, А. А. Анашкина, М. В. Головкин
Chem. Phys. 23 (28), 15352 (2021).
и др., Биофизика 56 (6), 1053 (2011).
14.
Г. В. Гурский и А. С. Заседателев, Биофизика 23,
47.
Я. В. Стирманов, М. В. Ходыков, О. В. Матвеева
932 (1978).
и др., Биофизика 58 (6), 981 (2013).
15.
G. V. Gursky and A. S. Zasedatelev, Sov. Sci. Rev. D,
48.
Ya. V. Stirmanov, O. V. Matveeva, and Yu. D. Nechi-
Physicochem. Biol. 5, 53 (1984).
purenko, J. Biomol. Struct. Dyn. 37 (12), 3103 (2019).
16.
Ю. Д. Нечипуренко, А. С. Заседателев и Г. В. Гур-
49.
Yu. D. Nechipurenko, in Advances in Medicine and Bi-
ский, Биофизика 24, 351 (1979).
ology, Ed. by L. V. Berhardt (Nova Science Publishers,
17.
W. Lenz, Z. Phys. 21, 613 (1920).
Hauppauge, New York, 2019), vol. 136, pp. 193-206.
18.
E. Ising, Z. Phys. 31, 253 (1925).
50.
G. D. Stormo, Introduction to protein-DNA interac-
19.
Ю. Д. Нечипуренко и др., Молекуляр. биология
tions: Structure, thermodynamics, and bioinformatics
18, 332 (1984).
(Cold Spring Harbor Laboratory Press, N. Y., 2013).
20.
Yu. D. Nechipurenko and G. V. Gursky, Biophys.
51.
Yu. M. Yevdokimov, S. G. Skuridin, Yu. D. Nechipu-
Chem. 24 (3), 195 (1986).
renko, et al., Int. J. Biol. Macromol. 36, 103 (2005).
21.
I. R. Epstein, Biophys. Chem. 8 (4), 327 (1978).
52.
V. B. Teif, N. Kepper, K. Yserentant, et al., J. Phys.:
22.
Б. Йованович и Ю. Д. Нечипуренко, Молекуляр.
Condens. Matter 27, 064110 (2015).
биология 24, 478 (1990).
53.
M. P. Evstigneev, A. S. Buchelnikov, and V. P. Ev-
23.
V. Arakelyan, Yu. Babayan, and G. Potikyan, J. Bio-
stigneev, Prog. Biophys. Mol. Biol. 149, 131 (2019).
mol. Struct. Dyn. 18, 231 (2000).
54.
С. Ф. Барановский, Д. Н. Чернышев, А. С. Бу-
24.
Ю. Д. Нечипуренко, А. М. Вольф и Ю. М. Евдо-
чельников и др., Биофизика 56 (2), 235 (2011).
кимов, Биофизика 48 (5), 802 (2003).
55.
M. P. Evstigneev and A. V. Shestopalova, in Applica-
25.
Ю. Д. Нечипуренко, А. М. Вольф и Г. В. Гурский,
tion of Computational Techniques in Pharmacy and
Биофизика 48 (6), 986 (2003).
Medicine. Challenges and Advances in Computational
26.
Ю. Д. Нечипуренко, Анализ связывания биологиче-
Chemistry and Physics, Ed. by L. Gorb, V. Kuz’min,
ски активных соединений с нуклеиновыми кислота-
and E. Muratov (Springer, Dordrecht, 2014), vol. 17,
ми (Ин-т компл. исслед., М.-Ижевск, 2015).
pp. 21-57.
27.
Ю. Д. Нечипуренко, Биофизика 27, 391 (1982).
56.
D. Yu. Lando and V. B. Teif, J. Biomol. Struct. Dyn.
28.
Ю. Д. Нечипуренко и Г.В. Гурский, Биофизика 48
17, 903 (2000).
(5), 773 (2003).
57.
В. Б. Тейф и Д. Ю. Ландо, Молекуляр. биология
29.
J. Wyman and S. J. Gill, Binding and Linkage: Func-
35, 117 (2001).
tional Chemistry of Biological Macromolecules (Univer-
58.
Ю. Д. Нечипуренко, А. М. Вольф, В. И. Салянов
sity Science Books, Mill Valley CA, 1990).
и др., Журн. эксперим. и теорет. физики 125 (1),
30.
Ю. Д. Нечипуренко, Молекуляр. биология 18,
103 (2004).
1066 (1984).
59.
Е. А. Воробьев, Ю. Д. Нечипуренко, В. И. Саля-
31.
A. R. Wolfe and T. Meehan, J. Mol. Biol. 223 (4),
нов и др., Биофизика 52 (4), 636 (2007).
1063 (1992).
60.
V. B. Teif and K. Bohinc, Prog. Biophys. Mol. Biol.
32.
D. Yu. Lando and Yu. D. Nechipurenko, J. Biomol.
105, 208 (2011).
Struct. Dyn. 26, 187 (2008).
61.
E. Bereznyak, N. Gladkovskaya, E. Dukhopelnykov,
33.
В. Ф. Рябоконь, Ю. Д. Нечипуренко и Г. В. Гур-
et al., AIMS Biophysics 2, 423 (2015).
ский, Докл. РАН 398, 832 (2004).
62.
А. Н. Суровая, Н. П. Бажулина, С. Ю. Лепехина и
34.
Yu. D. Nechipurenko, B. Jovanovic, V. F. Riabokon,
др., Биофизика 61 (2), 270 (2016).
et al., Ann. N. Y. Acad. Sci. 15, 660 (2005).
63.
A. G. Kudrev, Talanta 116, 541 (2013).
35.
Ю. Д. Нечипуренко, А. С. Крылов, А. С. Заседате-
лев и др., Молекуляр. биология 18, 332 (1984).
64.
Н. С. Ильинский, А. М. Варижук, А. Д. Вениами-
нов и др., Молекуляр. биология 48 (6), 891 (2014).
36.
D. M. Crothers, Science 339 (6121), 766 (2013).
37.
J. B. Chaires and S. D. Levene, Biopolymers 103, 469
65.
A. Beniaminov, G. Chashchina, A. Shchyolkina,
(2015).
et al., Biochimie 191, 33 (2021).
38.
Ю. Д. Нечипуренко и Г. В. Гурский, ДАН СССР
66.
M. Melikishvili and M. G. Fried, Biopolymers 103
281, 213 (1985).
(9), 509 (2015).
39.
Ю. Д. Нечипуренко и М. В. Волькенштейн, ДАН
67.
O. Matveeva, Yu. Nechipurenko, N. Nazipova, et al.,
СССР 286, 216 (1986).
Bioinformatics 32 (17), i552 (2016).
40.
Ю. Д. Нечипуренко, Н. В. Попов, М. А. Исаев
68.
E. Segal and J. Widom, Nat. Rev. Genet. 10 (7), 443
и др., Биофизика 40 (6), 1208 (1995).
(2009).
БИОФИЗИКА том 67
№ 3
2022
466
НЕЧИПУРЕНКО, БУЧЕЛЬНИКОВ
69.
S. Kim, E. Broströmer, D. Xing, et al., Science 339
76. J. Hooyberghs, P. Van Hummelen, and E. Carlon,
(6121), 816 (2013).
Nucl. Acids Res. 37, 53 (2009).
70.
Ю. Д. Нечипуренко, в кн. Физико-химические ме-
77. C. Furusawa, N. Ono, S. Suzuki, et al., Bioinformatics
ханизмы и регуляция процессов трансформации
25 (1), 36 (2009).
энергии в биологических структурах, под ред. Г. Ю.
78. L. Zhang, M. F. Miles, and K. D. Aldape, Nat. Bio-
Ризниченко и А. Б. Рубина (Ин-т компл. исслед.,
technol. 21, 818 (2003).
М.-Ижевск, 2017), сс. 102-125.
79. Д. А. Грядунов, Б. Л. Шаскольский, Т. В. Насед-
71.
V. V. Aristov, A. S. Buchelnikov, and Yu. D. Nechipu-
кина и др., Acta Naturae 10 (4), 4 (2018).
renko, Entropy 24 (2), 172 (2022).
80. С. А. Лапа, Р. А. Мифтахов, Е. С. Клочихина и др.,
72.
S. H. Ha and J. E. Ferrell, Science 352 (6288), 990
Молекуляр. биология 55 (6), 944 (2021).
(2016).
81. Д. О. Фесенко, И. С. Абрамов, В. Е. Шершов и
73.
G. Scatchard, Ann. N. Y. Acad. Sci. 51 (4), 660
др., Молекуляр. биология 52 (6), 997 (2018).
(1949).
82. Д. О. Фесенко, Т. О. Гусейнов, С. А. Лапа и др.,
74.
M. S. Rocha, Biopolymers 93 (1), 1 (2010).
Молекуляр. биология 52 (3), 533 (2018).
75.
F. Sevlever, J. P. Di Bella, and A. C. Ventura, PLoS
83. D. O. Fesenko, A. V. Chudinov, S. A. Surzhikov, et al.,
Comput. Biol. 16 (6), e1007929 (2020).
Genet. Test. Mol. Biomarkers 20 (4), 208 (2016).
Ligand Binding to Nucleic Acids in Solution and on Microchips
Yu.D. Nechipurenko*, ** and A.S. Buchelnikov**
*Engelhardt Institute of Molecular Biology, Russian Academy of Sciences, ul. Vavilova 32, Moscow, 119991 Russia
**Sevastopol State University, Universitetskaya ul. 33, Sevastopol, 299053 Russia
The equilibrium binding of ligands (proteins, biologically active compounds, and oligonucleotides) to nucleic
acids underlies many processes in the living cell. Model study and physical adsorption theory methods can be
used to give quantitative description of this binding in cases when the ligand occupies an extended region on
DNA and when cooperative interactions occur between adsorbed ligand molecules. With the Ising model it
is possible to find an analytical solution to the equations of adsorption in the most general form for a wide
range of models. Physico-chemical experiments in in vitro model systems containing biological macromole-
cules such as proteins, the complex web of protein interactions, RNA and DNA polymers, on the one hand,
and small molecule ligands on the other hand, shed light on many mechanisms underlying cellular processes.
Footprinting diagrams or microchip data sheet provide the detailed information on the system where ligands
bind to DNA. This article presents a brief overview of theoretical research in this area. How to use entropy
formula for the adsorption system is shown to learn more about the cooperative binding processes occurring
in this system. An approach for the analysis of binding on microchips is described in the Appendix.
Keywords: DNA-protein binding, cooperative interactions, binding curve, adsorption model, Ising model, micro-
chip
БИОФИЗИКА том 67
№ 3
2022