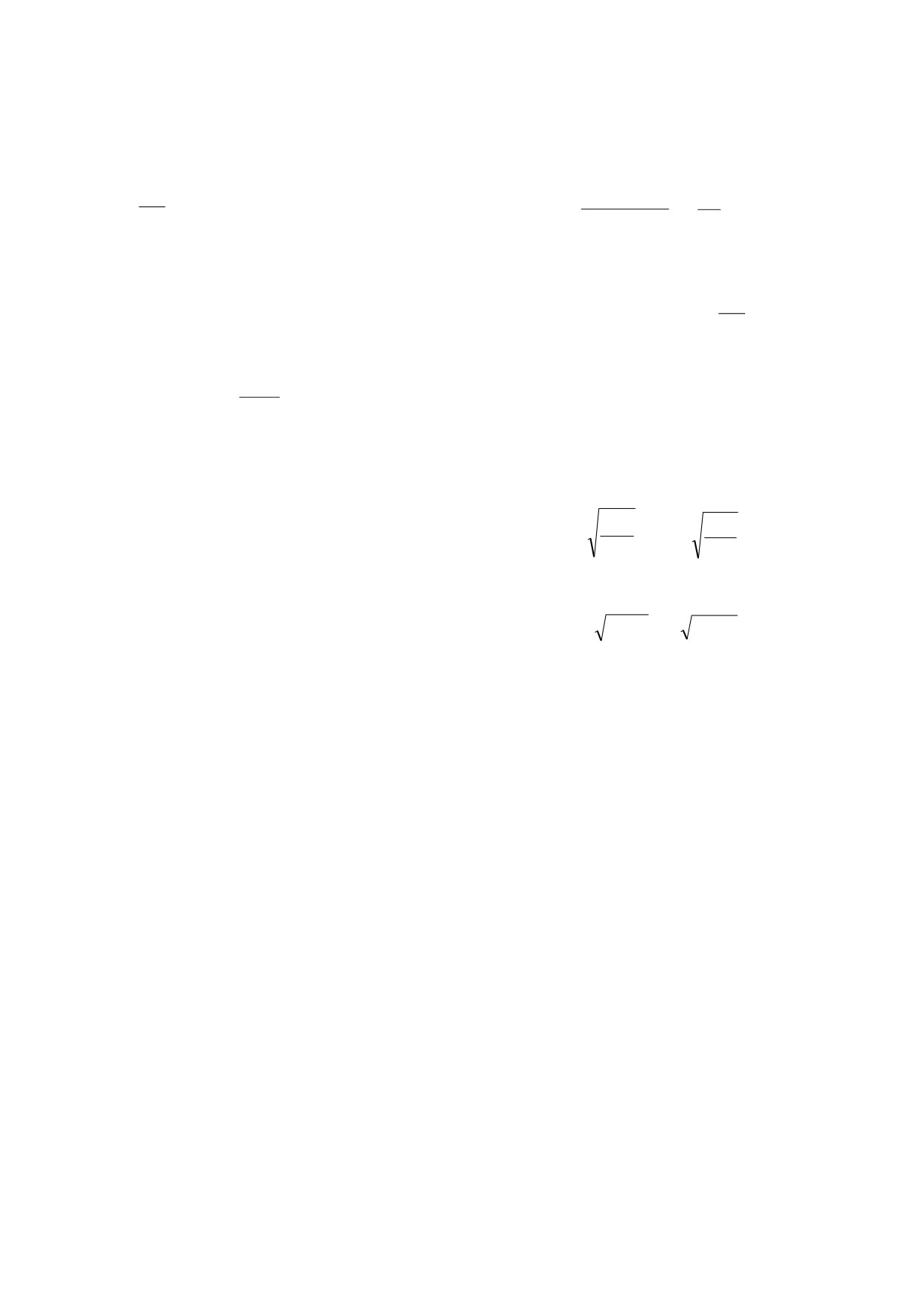БИОФИЗИКА, 2022, том 67, № 5, с. 1030-1040
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 577.3
ЭВОЛЮЦИОННАЯ ОПТИМАЛЬНОСТЬ ПРИЗНАКОВ ОРГАНИЗМА.
ВИДОВАЯ ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
КАК ПРОДУКТ ЭВОЛЮЦИИ
© 2022 г. Л.Л. Овсянников*, #
*Первый Московский государственный медицинский университет имени И.М. Сеченова МЗ РФ
(Сеченовский Университет), ул. Трубецкая, 8/2, Москва, 119991, Россия
#E-mail: tatiana-chap@yandex.ru
Поступила в редакцию 18.05.2022 г.
После доработки 18.05.2022 г.
Принята к публикации 24.06.2022 г.
Согласно дарвиновской теории естественного отбора в ходе конкуренции за ресурс выживает
наиболее приспособленная популяция. Величины признаков особей выжившей популяции будем
называть эволюционно-оптимальными. Строится модель биологического сообщества, состоящего
из конкурирующих популяций и растительного ресурса. Модель включала уравнение динамики
численности популяции, уравнение энергетического баланса половозрелой особи, уравнения
динамики плотности пищевого ресурса. Для определения эволюционно-оптимальных значений
признаков Ханиным и Семевским был предложен критерий, согласно которому эволюционно-
оптимальные значения признаков доставляют максимум коэффициенту размножения, равному
нулю в состоянии равновесия сообщества. Было показано, что существует эквивалентный критерий
минимума получаемой особью энергии. С помощью этих критериев продемонстрирован метод
определения эволюционно-оптимальных значений веса тела и толщины шерстного покрова особи.
Предполагается, что видовая продолжительность жизни, как и другие признаки организма, также
является продуктом эволюции. С помощью критерия минимума энергии, получаемой особью,
определяется видовая продолжительность жизни как функция внешних факторов и других
параметров организма. Полученные результаты были подтверждены численными экспериментами
для конкретных значений параметров.
Ключевые слова: эволюционная оптимальность, равновесное состояние, конкуренция популяций,
адаптивные признаки, продолжительность жизни.
DOI: 10.31857/S0006302922050210, EDN: JRBZVQ
вокупности признаков. В результате на больших
В современной науке принято считать, что все
временных промежутках выживает только одна
признаки организмов являются адаптивными в
популяция, другие элиминируются. Значения
том смысле, что являются результатом естествен-
признаков выжившей популяции будем называть
ного отбора. С появлением компьютеров появи-
эволюционно-оптимальными. В работах [3-11]
лась возможность моделировать естественный
было показано, что эволюционно-оптимальные
отбор [1, 2] и определять величины признаков ор-
значения признаков удовлетворяют некоторым
ганизмов, выживших в ходе отбора. Суть модели
критериям оптимальности, а в работах [12, 13]
естественного отбора, предложенной в данной
приводится обзор критериев оптимальности
работе, состоит в следующем. Строится модель
адаптивных признаков. Мы остановимся на кри-
динамики простейшего биологического сообще-
терии Ханина-Семевского [5, 7, 14], согласно ко-
ства, состоящего из популяции организмов и пи-
торому совокупность значений признаков побе-
щевого ресурса, потребляемого популяцией. Мо-
дившей популяции доставляет максимум коэф-
дель включает уравнение динамики численности
фициенту размножения, равному нулю в
популяции, динамики плотности ресурса и урав-
состоянии равновесия.
нение энергетического баланса особи. На основе
В работах [1, 12, 13] представлен обзор экстре-
модели биологического сообщества строится мо-
мальных принципов, позволяющих определять
дель конкуренции за ресурс популяций, отличаю-
эволюционно-оптимальные значения признаков.
щихся величинами какого-либо признака или со-
Оказалось, что количество энергии, потребля-
Сокращение: ПОЖ - половозрелый отрезок жизни.
емое особью выжившей популяции, минималь-
1030
ЭВОЛЮЦИОННАЯ ОПТИМАЛЬНОСТЬ ПРИЗНАКОВ ОРГАНИЗМА
1031
но. В результате был сформулирован критерий
Назовем выражение
минимума потребляемой энергии [15-18], исполь-
D
D
зуя который, можно определить эволюционно-
p
h
F(f,P,h)=
f
-(
+
(3)
оптимальные значения совокупности признаков.
P
h
В настоящей работе в качестве адаптивного
коэффициентом размножения.
признака рассматривается видовая продолжи-
Энергетический баланс организма особи. Пи-
тельность жизни. В работах [19-21] отмечается,
таясь растительной пищей, особь потребляет
что величина видовой продолжительности жизни
энергию в количестве
является продуктом эволюции. Однако явной
формализации этого понятия нет. Используя
V
G =
,
(4)
критерий минимума потребляемой энергии, по-
N
лучаем эволюционно-оптимальное значение ви-
где V - плотность энергии.
довой продолжительности жизни.
Полученную энергию организм расходует на
Согласно гомеостатической теории старения
следующие виды жизнедеятельности: основной
[20], причиной старения считается накопление
обмен и передвижение (W0), воспроизводство
оксидативных повреждений, приводящее к сни-
(Wf), развитие и функционирование адаптивного
жению с возрастом гомеостатических способно-
признака (Wh), привес (Wb). Таким образом, по-
стей организма. Из-за снижения гомеостатиче-
ской способности доставка кислорода уменьша-
лучаем
ется, так что энергетический ресурс организма
G = Wf + W0 + Wh + Wb.
(5)
падает. Это ведет к тому, что коэффициент усвое-
ния пищи уменьшается с возрастом. Это ведет к
Напишем формулы, отражающие характер за-
дефициту энергии, расходуемой на жизнедея-
висимости этих компонентов от различных пара-
тельность организма. Дефицит пополняется за
метров организма:
счет жировых накоплений. При этом с увеличе-
dP
нием возраста вес тела особи сначала растет, за-
Wf = cf, W0 = aP, Wh = Kh, Wb = b
,
(6)
dt
тем падает до начального значения P0 (веса в мо-
мент наступления половой зрелости). В этот мо-
где с, a, K и b - коэффициенты пропорциональ-
мент наступает смерть организма, поскольку
ности.
жировые ткани организма уже исчерпаны, а даль-
Предполагаем, что процессы в организме
нейшее пополнение энергии должно идти за счет
проходят гораздо быстрее, чем изменяется чис-
тканей жизненно важных органов.
dP
В работе также вводится коэффициент старе-
ленность популяции. В результате
≈ 0, и как
dt
ния, устанавливающий зависимость коэффици-
следствие Wb ≈ 0. Подставив выражения (6) в
ента усвоения пищи от возраста.
формулу (5), получаем
G = c f + aP + Kh.
(6a)
КРИТЕРИИ ЭВОЛЮЦИОННОЙ
Система уравнений (1), (4) и (6а) описывает
ОПТИМАЛЬНОСТИ
динамику биологического сообщества. Положе-
Рассмотрим простейшее биологическое сооб-
ние равновесия определяется следующей систе-
щество, состоящее из популяции плотностью N и
мой:
пищевого ресурса энергетической плотностью V.
Уравнение динамики плотности популяции име-
Dp
Dh
F(f,P,h) = f - (
+
) = 0,
(7a)
ет вид
P
h
dN
G = cf + aP + Kh,
(7б)
=(f -
d)N,
(1)
dt
V
где f - плодовитость, d - смертность. Пусть
G = .
(7в)
смертность зависит от величины адаптивного
N
признака (например, толщины шерстного покро-
Используя модель динамики биологического
ва для северных животных h) и веса тела P. Эту за-
сообщества, составим модель конкуренции попу-
висимость аппроксимируем простейшим выра-
ляций, отличающихся значениями адаптивных
жением
признаков, которыми являются h и P.
Dp
D
h
d
=
+
,
(2)
МОДЕЛЬ КОНКУРЕНЦИИ ПОПУЛЯЦИЙ
P h
где Dp и Dh - коэффициенты, учитывающие вли-
Пусть имеется k популяций. Ni - численность
яние внешних факторов.
популяции с величинами признаков hi и P i.
БИОФИЗИКА том 67
№ 5
2022
1032
ОВСЯННИКОВ
Динамика численности i-й популяции описы-
чин P и h с помощью критерия Ханина-Семев-
вается уравнением
ского.
Составим функцию приспособленности F. Для
dN
i
=(f
-
d
)Ni,
(8)
этого выразим f из формулы (7б) и подставим в
i
i
dt
формулу (7a). Получаем
D
D
p
h
G - aP - Kh Dp
Dh
где di =
+
F(G,P,h) =
- (
+ a
).
(14а)
P
h
P
h
i
i
c
Уравнение энергетического баланса особи i-й
Из совокупности пар выбираем пару Pi,hi.
популяции имеет вид
Приравняем Fi = F(Pi,hi) нулю. Отсюда находим:
G = cfi + aPi + Khi,
(9)
D
D
p
h
Gi = G(Pi,hi) = aPi +Khi + c(
+
).
где G - количество энергии, получаемой особью с
P
h
i
i
пищей. Оно одинаково для особей всех популя-
ций и определяется выражением
Для остальных (k
- 1) пар определяем
F(Gi,Pj,hj), где j ≠ i. В том случае, если F(Gi,Pj,hj) <
V
0, пара Pi,hi будет эволюционно-оптимальной,
G =
(10)
∑Ni
т.е. популяция особи которой имеют эти значе-
ния победит в конкуренции.
Система уравнений (8)-(10) имеет k нетриви-
Найдем глобальный максимум F. Для этого
альных равновесных состояний, в которых чис-
продифференцируем уравнение (14a) по P и h,
ленность только одной j-й популяции отлична от
Приравняв нулю производные, получаем
нуля. Численности остальных (k - 1) популяций
равны нулю.
Dp
c
Dh
c
Nj > 0,
(11a)
P* =
; h* =
(15)
a
K
Fi = fi - dj = 0, если i = j,
Приравняв уравнение (14a) нулю и подставив
Ni = 0,
(11б)
выражения (15), получаем:
−---− -
D cap
D cKh
G* = 2
+ 2
(16)
Fi = fi - di
≠
0, если i ≠ j и i =
1,
к
-1,
Если среди совокупности k популяций со зна-
Gj = G = cfj + aPj + Khj,
(12)
чениями Pi,hi (i = 1,2,…,k) имеется популяция со
значениями (P*,h*), то она и победит в конкурен-
V
Nj = .
(13)
ции. Значения признаков выжившей популяции
G
удовлетворяют критерию Ханина-Семевского,
Равновесие (11) будет асимптотически устой-
поскольку
чивым, если выполняется условие Fi = fi - di < 0,
F(G*,P*,h*) = maxF(G*,Pi,hi)= 0
или
(величины остальных (k - 1) значений отрица-
Fj = max (F1,F2,…,Fk) = 0.
(14)
тельны).
На числовом примере убедимся, что значения
Предложенная модель является математиче-
P* и h* удовлетворяют критерию Ханина-Семев-
ской формализацией дарвиновской теории есте-
ского.
ственного отбора, согласно которой в процессе
конкуренции за ресурс выживает наиболее при-
Пусть a = 100 ккал·кг-1·сут-1, K = 120 ккал·см-1·
способленная популяция. Мерой приспособлен-
·сут-1, c = 91250 ккал/особь, Dp = 0.11 кг·сут-1,
ности является коэффициент размножения Fj,
Dh = 0.033 см·сут-1. В результате по формулам
удовлетворяющий выражению (14).
(15) и (16) получаем оптимальную совокупность
Это выражение (впервые было получено
М.А. Ханиным [14] и Ф.Н. Семевским [7]) будем
P* = 10 кг, h* = 5 см, G* = 3200 ккал·сут-1. Подста-
вив эти значения в выражение (14а), получаем
называть критерием Ханина-Семевского. Ис-
пользуя критерий Ханина-Семевского, можно
F(G*,P*,h*) = 0. При любой другой совокупности
определить величины совокупности признаков
признаков F(G*,P,h) < 0.
особей популяции, победившей в ходе конкурен-
Полученные значения (P* = 10 кг, h* = 5 см)
ции за ресурс. Эти величины будем называть эво-
будут эволюционно-оптимальными в том смыс-
люционно-оптимальными. Проиллюстрируем
ле, что популяция с особями, имеющими данную
определение эволюционно- оптимальных вели-
совокупность значений выиграют конкуренцию у
БИОФИЗИКА том 67
№ 5
2022
ЭВОЛЮЦИОННАЯ ОПТИМАЛЬНОСТЬ ПРИЗНАКОВ ОРГАНИЗМА
1033
Рис. 1. Динамика конкуренции двух популяций, отличающихся значениями признаков.
популяций с другой совокупностью значений.
можно получить, используя критерий минимума
Продемонстрируем это на модели конкуренции.
получаемой энергии. Для этого из уравнения (7a)
выразим f и подставим в формулу (7б). Получаем
МОДЕЛЬ КОНКУРЕНЦИИ
Dp Dh
ДЛЯ ДВУХ ПОПУЛЯЦИЙ
G(P,h) = c(
+
) + aP + Kh.
(22)
P h
Имеются две популяции численностями
N1 и N2. Модель конкуренции имеет вид:
Обозначим Gi = G(Pi,hi). Каждой паре значе-
ний (Pi,hi) соответствует величина Gi. Согласно
dN
D
D
1
p
h
критерию минимума потребляемой энергии, в
=
(f
-(
+
))N
,
(17)
1
1
dt
P h
конкуренции побеждает популяция с такими зна-
1
1
чениями признаков, которые доставляют мини-
мум потребляемой энергии G.
dN2
Dp Dh
G* = G(P*,h*) = minG(Pi,hi).
= (f2 - (
+
))N2,
(18)
dt
P
h
2
2
Алгоритм определения эволюционно-опти-
мальных значений пар (Pi,hi) следующий: 1) для
V
G =
,
(19)
каждой пары значений (Pi,hi) вычислить по фор-
N
+
N
1
2
муле (22) величину Gi; 2) выбрать пару, которой
G = cf1 + aP1 + Kh1,
(20)
соответствует минимальное значение G.
Найдем значения признаков, доставляющих
G = cf2 + aP2 + Kh2.
(21)
глобальный минимум функции G. Продифферен-
Проведем численные расчеты для конкретных
цируем уравнение (22) по P и h. Приравняв нулю
значений параметров. Используем следующие
производные, получаем
начальные условия: N1(0)
= N2(0)
=
= 0.001 особь·м-2, V = 8 ккал·м-2·сут-1; значения
Dp
c
Dh
c
P* =
, h* =
(23)
признаков: P1= P* = 10 кг, h1 = h* = 5 см, P2 = 8 кг,
K
a
h2 = 6 см.
Соответствующее значение функции будет
Результаты расчетов приведены на рис.
1.
равно
Видно, что выживает первая, наиболее при-
способленная популяция, то есть та, которая
G* = G(P*,h*) = 2
D cap
+ 2
D cKh
(24)
удовлетворяет критерию Ханина-Семевского
(F(3200,10,5) = 0, а F(3200,8,6) = -0.00084).
Нетрудно убедиться, что G* = G(P*,h*) =
= minG(P,h).
КРИТЕРИЙ МИНИМУМА ПОЛУЧЕННОЙ
Как видно, эволюционно-оптимальные значе-
ЭНЕРГИИ
ния, полученные из критериев Ханина-Семев-
В работах [15-18] показано, что эволюционно-
ского (формула (15)) и энергетического (формула
оптимальные значения совокупности признаков
(23)), совпадают.
БИОФИЗИКА том 67
№ 5
2022
1034
ОВСЯННИКОВ
Убедимся в том, что значения признаков побе-
[20], едва ли не единственной причиной старения
дившей популяции удовлетворяют критерию ми-
считается накопление оксидативных поврежде-
нимума потребляемой энергии. Для этого
ний [22, 23], приводящее к снижению гомеоста-
сравним G1 = G(P1,h1) и G2 = G(P2,h2). По форму-
тических способностей организма с возрастом.
Из-за снижения гомеостатической способности
ле (4.1) получаем G1
=3200 ккал·сут-1; G2 =
доставка кислорода уменьшается, так что энерге-
= 3276,5 ккал·сут-1. Как видим, количество по-
тический ресурс организма падает. Поскольку
требляемой энергии у победившей популяции
расход энергии измеряется в кислородном эк-
меньше.
виваленте, а количество кислорода в клетках ор-
Хотя оба представленных критерия эквива-
ганизма падает с возрастом, то коэффициент
лентны, энергетический критерий более простой
усвоения пищевой энергии также уменьшается с
в понимании и использовании. В дальнейшем
возрастом. Эту зависимость будем аппроксими-
при определении эволюционно-оптимальных
ровать выражением
значений признаков организмов будем использо-
φ = 1 - mτ,
(26)
вать энергетический критерий.
где τ - возраст взрослой особи (τ ∈ [0, TМ - t0]),
Эволюционно-оптимальное значение видовой
t
продолжительности жизни. Естественно предпо-
0 - возраст наступления половой зрелости; TМ -
ложить, что видовая продолжительность жизни
полная продолжительность жизни; m - коэффи-
является таким же продуктом эволюции, как и
циент старения. Предполагаем, что старение
структурно-функциональные признаки орга-
начинается с возраста наступления половой зре-
низма.
лости.
Применим вышеизложенную теорию для
Подставив выражение (26) в формулу (25), по-
определения эволюционно-оптимального значе-
лучаем количество энергии, содержащееся в по-
ния видовой продолжительности жизни.
лученной пище:
Введем обозначения: TМ - промежуток от
V
Gp = qφ,
(27)
рождения до смерти. Его назовем полной продол-
N
жительностью жизни; t0 - возраст наступления
Для особи возраста TМ выражение (27) запи-
половой зрелости; T = TМ - t0 - половозрелый
шем в виде
отрезок жизни.
Gp(T) = G(1 - mT),
(27a)
ЭНЕРГЕТИЧЕСКИЙ БАЛАНС ОРГАНИЗМА
где T = TМ - t0 - половозрелый отрезок жизни
В ареале произрастает растительность, кото-
V
особи, G = q.
рая служит пищей для популяции животных. В
N
результате особи популяции получают энергию,
Расход энергии на воспроизводство. Очевидно,
которую расходуют на различные виды жизнеде-
что энергозатраты на воспроизводство пропор-
ятельности: основной обмен, передвижение, ре-
циональны плодовитости f и определяются выра-
продукцию, привес. Рассмотрим все компонен-
жением
ты, входящие в уравнение энергетического ба-
ланса.
Wf = сf,
(28)
Энергия, получаемая особью. Пусть V - плот-
где с - коэффициент пропорциональности, соот-
ность растительности, производимой в экосисте-
ветствующий энергии, затрачиваемой родителем
ме в единицу времени. N - плотность численно-
на выкармливание одного потомка.
сти популяции; q - удельная калорийность пищи.
Затраты энергии на основной обмен и передви-
Количество энергии, потребляемое одной осо-
жение. Эти затраты определим известным выра-
бью, составляет
жением [24]:
V
G = q.
(25)
W0 = aPγ,
(29)
N
где P - масса тела. В дальнейшем упрощенно по-
Но не вся потребленная с пищей энергия усва-
лагаем γ = 1.
ивается организмом особи. Количество усвоен-
ной особью энергии определяется выражением
Энергозатраты на привес. Данные затраты бу-
дем определять выражением
Gp = Gφ,
(25a)
dP
где φ - коэффициент усвоения пищевой энергии.
Wb = b
,
(30)
dτ
Влияние старения на коэффициент усвоения.
Согласно гомеостатической теории старения
где b - удельная калорийность тела особи.
БИОФИЗИКА том 67
№ 5
2022
ЭВОЛЮЦИОННАЯ ОПТИМАЛЬНОСТЬ ПРИЗНАКОВ ОРГАНИЗМА
1035
В результате уравнение энергетического ба-
ность зависит от следующих параметров:
D
-
ланса особи возраста τ примет вид
учитывают постоянные факторы среды,
β
- учи-
Gp = Wf + W0 + Wb
(31)
тывают физиологическое состояние организма,
или
- адаптивные признаки. Тогда
a
dP
G(1 - mτ) = сf + aP + b
(31a)
d = d(D,
β,
a
).
(35)
dτ
В равновесии уравнение (33) принимает
Масса тела особи возраста TМ, как будет пока-
вид
зано, равна P0, т.е. массе особи возраста t0. Пере-
пишем уравнение энергетического баланса (31a)
dn
для особи возраста TМ:
= - d(τ)n
(36)
dτ
dP(T
)
с начальным условием
G(1 - mT) = сf + aP0 + b
(32)
dτ
T
M
n(0) =
(τ)
f
(τ)
dτ
(37)
∫n
ДИНАМИКА БИОЛОГИЧЕСКОГО
t0
СООБЩЕСТВА С УЧЕТОМ ВОЗРАСТНОЙ
Решение уравнения (36) имеет вид
СТРУКТУРЫ ПОПУЛЯЦИИ
ttt
Предлагается аналитическая модель сообще-
n(τ) = n0exp(-
(
)
),
(38)
ства, учитывающая возрастную структуру попу-
∫d
0
ляции.
Пусть n(τ,t) - плотность численности популя-
где n0 = n(0) - численность новорожденных. Под-
ции, т.е. численность особей возраста τ в момент
ставив выражение (38) в уравнение (37), получаем
времени t. Число особей в интервале возраста от t1
T
M
ttt
до t2 определяется выражением
n0 = n0
(τ
)exp(-
(
)
)дτ.
(39)
∫f
∫d
τ
2
t0
0
N(t) =
n(
τ
,
t
)d
τ
(33а)
∫
Сократив обе части на n0 ≠ 0, получаем
τ
1
T
M
ttt
Уравнение динамики плотности численности
(τ
)eхp(-
(
)
)дτ - 1 = 0.
(40)
популяции представим известным уравнением
∫f
∫d
t0
0
Лотки [25].
Уравнение (6.8) устанавливает связь между
дn
дn
+
= -d(τ,t)n,
(33)
смертностью d и плодовитостью f в равновесном
дt
дτ
состоянии сообщества.
где d(τ,t) - смертность.
Динамика популяции, состоящей из двух воз-
Граничное условие
растных групп - ювенильной и половозрелой. Юве-
нильные особи. Возраст: от момента рождения до
T
M
возраста наступления половой зрелости
n(0,t) =
(τ)n
(τ
,
t
)d
τ
(34)
∫f
t
0(τ∈[0,t0]). Упрощенно полагаем, что ювениль-
t0
ные особи потребляют энергию только с молоком
определяет количество новорожденных в момент
матери.
времени t. Здесь f(τ) - плодовитость (количество
Плодовитость ювенильных особей равна
потомков, произведенных особью возраста τ),
нулю.
t0 - возраст наступления половой зрелости, TМ -
Смертность ювенилов dU обусловлена внеш-
полная продолжительность жизни особи.
ними факторами и зависит от возраста.
Смертность зависит от ряда причин: 1) внеш-
Половозрелые особи. Возраст: τ ∈ [t0,TМ], где
ние факторы определяются климатом, наличием
TМ - продолжительность жизни особи (от рожде-
хищников и паразитов; 2) внутренние факторы
ния до смерти).
обусловлены физиологическим состоянием орга-
низма; 3) приспосабливаясь к условиям внешней
Плодовитость: упрощенно будем полагать, что
среды, организм развивает адаптивные признаки
плодовитость f постоянна во всех возрастах.
a
, величина этих признаков также оказывает
Смертность: упрощенно полагаем, что смерт-
влияние на смертность. Таким образом, смерт-
ность половозрелых особей dМ обусловлена
БИОФИЗИКА том 67
№ 5
2022
1036
ОВСЯННИКОВ
внешними факторами и не зависит от возраста
обусловливает высокую смертность и как след-
(dМ = const).
ствие низкую продолжительность жизни. И на-
оборот, высокая продолжительность жизни обу-
В результате численность новорожденных
словливает низкую плодовитость.
определяем формулой
В природе наблюдаются виды c практически
T
M
нулевой взрослой смертностью. При этом они
n0 = f
n(
τ
)d
τ
,
(41)
∫
имеют высокую продолжительность жизни. На-
t0
пример, калифорнийский кондор живет доволь-
где n(τ) - плотность численности половозрелых
но долго (примерно 60 лет) и при этом имеет
особей. Она определяется выражением
очень низкую плодовитость (один потомок за два
n(τ) = n(t0)exp(-dМτ).
(42)
года).
Если предположить, что смертность соответ-
Плотность численности половозрелых особей
в возрасте t0 - n(t0) - определяется выражением
1
ствует величине d =
, то динамику плотности
t0
wT
n(t0)= n0
exp(-d τ dτ )
,
(43)
численности взрослых особей популяции можно
∫
представить выражением
0
где du - ювенильная смертность.
dN
1
= (f -
)N.
Обозначим через w выживаемость новорож-
dt
wT
денных:
В состоянии равновесия справедливо выраже-
t0
ние (50). Подставив это выражение в формулу
w =
exp(-
d τ dτ )
(44)
∫
(31a), получаем дифференциальное уравнение
0
динамики массы половозрелой особи в равновес-
Подставив выражение (44) в выражение (43),
ном состоянии биологического сообщества:
получаем
c
dP
n(t0) = n0w.
(45)
G(1 - mτ) =
+ aP + b
(50a)
wT
dτ
Подставим выражения (42) и (45) в формулу (41).
с начальным условием P(0) = P0.
Получаем
T
M
Это уравнение имеет аналитическое решение,
которое приводим ниже. Обозначим K= (G -
n0 = f n0w
exp(
τ
)
τ
(46)
∫
-d
с
Gm a
t0
-
)/b; r =
; g = .
Сократив обе части формулы (46) на n0 ≠ 0 и
wT
b
b
проинтегрировав, получаем
Получаем:
exp(-d
t )- exp(−d
T
)
M
0
M
M
r
fw
= 1.
(47)
P(τ) = (K +
- rτ)/g +
d
M
g
В том случае, если внешняя смертность
r
равна нулю (dМ = 0), особи гибнут только от
+ (P0 - (K +
)/g)exp(-gτ).
(51)
g
старости, дожив до предельного возраста TМ.
Продемонстрируем решение на конкретном
Поскольку
примере.
exp(-d
t )- exp(−
d
T
)
Пусть значения параметров будут равны:
M
0
M
M
lim
= TМ -t0,
d
M
→0
с = 585 ккал, b = 600 ккал·кг-1, a = 100 ккал·кг-1·
d
M
получаем
·сут-1, P0 = 1,4 кг, T = 25 сут, w = 1, G =
fw(TМ - t0) = 1.
(48)
= 204 ккал·сут-1, m = 0.01 сут-1. На рис. 2 изобра-
Так как
жен график зависимости массы особи от возраста
τ для данных значений параметров.
T = TМ - t0,
(49)
Как видно из графика, сначала масса возраста-
то подставив выражение (49) в уравнение (48),
ет, достигая максимального значения в возрасте
получаем
7 суток. Это объясняется тем, что избыток массы
1
f =
(50))
образуется за счет образовавшихся жировых тка-
wT
ней организма. Затем масса снижается, посколь-
Формула (50) связывает плодовитость с дли-
ку полезная энергия, получаемая особью, умень-
тельностью половозрелого периода жизни особи
шается, и недостаток ее покрывается не только
и соответствует понятию сопряженности. Со-
энергией, получаемой с пищей, но и за счет обра-
гласно этому понятию, высокая плодовитость
зовавшихся жировых тканей. Наконец, наступает
БИОФИЗИКА том 67
№ 5
2022
ЭВОЛЮЦИОННАЯ ОПТИМАЛЬНОСТЬ ПРИЗНАКОВ ОРГАНИЗМА
1037
Рис. 2. Зависимость веса тела взрослой особи от ее
Рис. 3. Зависимость потребляемой энергии от продол-
возраста.
жительности жизни особей популяции.
возраст ТМ. В этом возрасте масса тела становит-
МОДЕЛЬ ДИНАМИКИ
ся равным начальной массе P0. Запас жира исчер-
БИОЛОГИЧЕСКОГО СООБЩЕСТВА
пан, и особь погибает, поскольку необходимая
Уравнения динамики экосистемы представим
энергия будет пополняться за счет тканей жиз-
в виде
ненно важных органов.
dN
1
Оптимальная величина продолжительности
= (f -
)N,
(53a)
dt
wT
зрелого периода жизни Т, при которой потребля-
емая энергия минимальна, является, согласно
G = Vq/N,
(53б)
энергетическому критерию, также эволюционно-
оптимальной величиной.
dP(T
)
f = (G(1 - mT) - aP0 - b
)/с,
(53в)
dτ
Уравнение (51) для особи возраста TМ примет
вид
где N - плотность численности взрослых особей;
Vq =Vq (V - плотность пищевого ресурса, q -
r
удельная калорийность ресурса).
P0 = (K +
- rT)/g +
g
В итоге получаем следующий алгоритм расчета
биологического сообщества.
r
+ (P0 - (K +
)/g)exp(-gT).
(51a)
Шаг 1. Задаем начальные значения N0 и f0.
g
Решая численно дифференциальное уравнение
(53a), получаем численность популяции на следу-
Откуда получаем количество энергии, потреб-
ющем (первом) временном шаге N1.
ляемое каждой особью популяций в конце жизни
Шаг 2. Из уравнения (53б) находим G1.
(оно одинаково для взрослых особей любого воз-
раста).
Шаг 3. По формуле (53в) находим значение
плодовитости f1 на следующем временном слое.
aP +c/(wT)
Переходим на шаг 1 и т.д.
0
G(T) =
(52)
Равновесные значения переменных сообще-
1
+
m
/
g-mT
/(1
−
exp(−
gT
))
ства имеют вид
На рис. 3 изображен график зависимости по-
_
требляемой особью энергии от половозрелого пе-
f
=1/(wT),
(54а)
риода жизни T.
aP +c/(wT)
0
Из рис. 3 видно, что при T* = 18, G* = min(G) =
G(T) =
,
(54б)
1
+
m
/
g-mT
/(1
−
exp(−
gT
))
= 198.1. Это значит, что имеется оптимальная
продолжительность жизни, при которой энергия,
−
_
N
= Vq/G
(54в)
потребляемая особью минимальна.
БИОФИЗИКА том 67
№ 5
2022
1038
ОВСЯННИКОВ
Рис. 4. Динамика численности популяции.
Продемонстрируем расчет динамики экоси-
Количество энергии, получаемое одной взрос-
стемы на конкретном примере
лой особью:
Для этого решим систему уравнений (53) для
V
q
конкретных значений параметров.
G =
(356)
Пусть N0 = 0.04 особь·м-2 и Vq = 11.2 ккал·м-2·
∑Ni
Плодовитость особи i-й популяции получаем
·сут-1. Для T = 25 сут мы получили энергию при-
из выражений (54):
веса Wb(25) = -11.76 ккал·сут-1 и основного обме-
m
mT
i
на W0(25) = W0(0) = 140 ккал·сут-1.
fi = (G(1 +
-
) - W0)/c,
(57)
g
1−exp(
-
gT
i
)
На рис. 4 изображена динамика численности
популяции. Как видим, численность стремится к
где W0 - энергозатраты на основной обмен и пе-
--
редвижение особи в возрасте Ti. Они определяют-
величине
N
= 0.055 особь·м-2. Как показали
−
ся формулой
расчеты, f
→ f
= 0.04 сут-1, G
=
→G
W0 = aP0.
(58)
= 202.7 ккал·сут-1.
Рассмотрим три популяции со следующими зна-
Из графика на рис. 4 видно, что плотность чис-
чениями продолжительности жизни: T1 = 12 сут,
ленности стремится к равновесному состоянию,
T2 = T* = 18 сут, T3 =25 сут. Этим величинам соот-
полученному из решения системы (53).
ветствуют следующие равновесные значения потреб-
ляемой энергии: G(T1) = 204.9 ккал·сут-1, G(T2) =
МОДЕЛЬ КОНКУРЕНЦИИ ПОПУЛЯЦИЙ
= 198.2 ккал·сут-1; G(T3) = 202.7 ккал·сут-1 и приве-
С РАЗНОЙ ПРОДОЛЖИТЕЛЬНОСТЬЮ
ЖИЗНИ
сов: Wb1 = -8.4 ккал· ·сут-1, Wb2 = -10.0 ккал·сут-1;
Используя модель динамики биологического
Wb3 = -11.4 ккал· ·сут-1.
сообщества, построим модель конкуренции по-
Как видим, минимальное значение потребляе-
пуляций с различной продолжительностью
мой энергии имеет популяция с продолжитель-
половозрелого отрезка жизни (ПОЖ). Покажем,
ностью жизни T2 = 18 сут.
что в соответствии с энергетическим критерием
Для заданных значений продолжительности
эволюционной оптимальности в конкуренции
жизни и представленных значениях параметров
побеждает популяция с оптимальной продолжи-
решаем систему уравнений (55)-(57). Выберем
тельностью жизни (доставляющей минимум по-
следующие начальные условия: N1(0) = N2(0) =
требляемой энергии). Напишем уравнения био-
логического сообщества, содержащего популя-
= N3(0) = 0.013 особь·м-2.
ции с различными значениями ПОЖ.
На рис. 5 представлено решение этой системы.
Динамика численностей популяций:
Из графика видим, что в соответствии с энергети-
dNi
ческим критерием эволюционной оптимально-
1
= (fi -
)Ni.
(55)
сти в конкуренции побеждает популяция с опти-
dt
T
i
мальным значением адаптивного признака (в
БИОФИЗИКА том 67
№ 5
2022
ЭВОЛЮЦИОННАЯ ОПТИМАЛЬНОСТЬ ПРИЗНАКОВ ОРГАНИЗМА
1039
Рис. 5. Динамика численности конкурирующих популяций с разными значениями ПОЖ.
данном случае продолжительности жизни). При
с
wW0
этом получаемая особью энергия минимальна.
T* =
(
1+
- 1),
(62)
wW
cm
Из графика видно, что выживает популяция
0
особи которой имеют оптимальную продолжи-
где W0 = aP0.
тельность жизни Т* = 18 сут. Численности других
Эволюционно-оптимальная величина ПОЖ,
популяций стремятся к нулю.
полученная по упрощенной формуле Т*
=
= 16.7 сут. Достоинством формулы (62) по сравне-
УПРОЩЕННАЯ ФОРМУЛА
нию с алгоритмом определения оптимума Т чис-
ЭВОЛЮЦИОННО-ОПТИМАЛЬНОГО
ленным методом в том, что выявлена явная зави-
ЗНАЧЕНИЯ ПОЛОВОЗРЕЛОГО
симость видового ПОЖ от параметров организма.
ОТРЕЗКА ЖИЗНИ
Напишем уравнение энергетического баланса
ВЫВОДЫ
взрослой особи возраста Т в равновесном состоя-
1. Показано, что величины признаков орга-
нии экосистемы:
низма являются продуктами эволюции и при
G(1 - mT) = Wf(T) + W0(T)+ Wb(T),
(59)
этом удовлетворяют критерию эволюционной
с
dP(T
)
оптимальности Ханина-Семевского.
где Wf(T) =
, W0(T) = aP0, Wb(T) = b
,
2. Определено существование эквивалентного
wT
dt
критерия, критерия минимума потребляемой
или
энергии, позволяющего более простым способом
с
dP(T
)
определять эволюционно-оптимальные значения
G(1 - mT) =
+ W0(T) + b
(60)
адаптивных признаков организма.
wT
dt
Как видим, расчеты для возраста T показыва-
3. В уравнении энергетического баланса орга-
ют, что энергозатраты на привес много меньше
низма учитывался тот факт, что коэффициент
энергозатрат на основной обмен и передвижение
усвоения пищевой энергии уменьшается с воз-
растом пропорционально коэффициенту старе-
(Wb(T) << W0(T)). В результате полагаем Wb(T)
≈
0.
ния, который обусловлен накоплением оксида-
Перепишем уравнение (60) в виде
тивных повреждений клеток с возрастом.
с
4. Определена динамика веса тела взрослой
G(1 - mT)
≈
+ W0.
(61)
особи от возраста, согласно которой, вес вначале
wT
растет до определенной величины, затем падает
Отсюда количество полученной энергии будет
до первоначального значения. В этот момент
равно
происходит смерть организма.
с
5. Из системы уравнений равновесного состо-
G
≈
(
+ W0)/(1 - mT).
(61а)
wT
яния сообщества определяется зависимость по-
Далее находим оптимальное Т*, доставляющее
требляемой энергии от продолжительности жизни.
минимум G.
6. Используя критерий минимума потребляе-
Продифференцировав уравнение (61а) по Т и
мой энергии, получаем эволюционно-оптималь-
приравняв производную нулю, получаем
ную величину видового ПОЖ.
БИОФИЗИКА том 67
№ 5
2022
1040
ОВСЯННИКОВ
7. Справедливость предложенной теории под-
8. А. Т. Терехин и Е. В. Будилова, Журн. общ. биоло-
тверждалась моделью конкуренции за ресурс по-
гии, 62 (1), 286 (2001).
пуляций с различными значениями видового
9. E. L. Charnov, Lafe history invariants. Some exploration
of sinnetry in evolutionary ecology (Univ. Press, Oxford,
ПОЖ. В процессе конкуренции выживала попу-
1993).
ляция с ПОЖ, доставляющим минимум потреб-
10. J. Kozlovski and J. Weiner, Amer. Nat., 149 (1), 423
ляемой энергии.
(1997).
11. R. E. Ulanowicz, Ecol. Model., 220 (16), 1886 (2009).
КОНФЛИКТ ИНТЕРЕСОВ
12. П. В. Фурсова, Л. П. Левич и И. Л. Алексеев, Успе-
Автор заявляет об отсутствии конфликта инте-
хи соврем. биологии, 123 (2), 115(2003).
ресов.
13. И. В. Волвенко, Биофизика, 57 (3), 476 (2012).
14. М. А Ханин, Н. Л. Дорфман, И. Б. Бухаров и
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
В. Г. Левадный, Экстремальные принципы в биоло-
гии и физиологии (Наука, М., 1978).
Настоящая работа не содержит описания ис-
15. Л. Л. Овсянников и В. П. Пасеков, Журн. общ.
следований с использованием людей и животных
биологии, 51 (5),709(1990).
в качестве объектов.
16. Л. Л. Овсянников и М. И. Шпитонков, Биофизи-
ка, 65 (1), 1171(2020).
СПИСОК ЛИТЕРАТУРЫ
17. Л. Л. Овсянников и Ю. М. Свирежев, Журн. общ.
биологии, 14 (5), 621(1983).
1. А. П. Левич, Искусство и метод в моделировании си-
стем (Москва-Ижевск, 2012).
18. Л. Л. Овсянников, Тр. Института системного ана-
лиза РАН, 69 (2), 78(2019).
2. А. В. Марков и Е. Б. Наймарк, Количественные за-
кономерности макроэволюции (Геос, М., 1998).
19. Л. А. Гаврилов и Н. С. Гаврилова, Биология продол-
жительности жизни (Наука, М., 1991).
3. Н. В. Волосова, Математические структуры и мо-
делирование, 2 (38), 27 (2016).
20. В. Н. Новосельцев, Ж. А. Новосельцева и
А. И. Яшин, Успехи геронтологии, 12, 149 (2003).
4. Г. Ф. Гаузе, The struggle for existence (Hafner, New
York, 1934).
atpsynthase.info/Gero/ZhBS - 2013.pdf
5. И. Ф.Образцов и М. А. Ханин, Оптимальные био-
механические системы (Медицина, М., 1989).
22. В. К. Кольтовер, Успехи соврем. биологии, 96, 85
(1983)
6. Р. Розен, Принцип оптимальности в биологии (Мир,
М., 1969).
23. D. Harman, Exp. Gerontol., 33, 95(1998)
7. Ф. Н. Семевский и С. М. Семенов, Математиче-
24. M. Kleiber, Higardia, 6, 315 (1932).
ское моделирование экологических процессов (Гидро-
25. A. J. Lotka, Elements of Mathematical Biology (Williams
метеоиздат, Л., 1982).
and Wilkins, Baltimore, 1925).
The Evolutionary Optimality of the Body's Features.
Species Life Span as a Product of Evolution
L.L. Ovsyannikov*
*Sechenov First Moscow State Medical University of the Ministry of Health of the Russian Federation (Sechenov University),
Trubetskaya ul. 8/2, Moscow, 119991 Russia
According to Darwin's theory of natural selection, the fittest population can survive in competition for re-
sources. The values of the traits of the fittest are referred to as evolutionarily optimal trait values. A model of
a biological community that consists of populations that compete to get ahold of plant resources is being built.
The model included the equation of population dynamics, the equation of the energy balance of a mature in-
dividual, the equations of the dynamics of the density of a food resource. To determine the evolutionarily op-
timal values of traits, Khanin and Semevsky proposed a criterion according to which the evolutionarily opti-
mal values of traits deliver the maximum to the reproduction coefficient equal to zero in the equilibrium state
of the community. It was shown that there is an equivalent criterion for the minimum energy received by an
individual. Using these criteria, a method for determining the evolutionarily optimal values of body weight
and the thickness of the coat of species was demonstrated. It is assumed that the species life span, like other
traits of the organism, is also a product of evolution. Using the criterion of the minimum energy received by
an individual, the specific life span is determined as a function of external factors and other parameters of the
organism. The results obtained were confirmed by numerical experiments for specific parameter values.
Keywords: evolutionary optimality, equilibrium state, competition between populations, adaptive traits, life span
БИОФИЗИКА том 67
№ 5
2022