БИОФИЗИКА, 2022, том 67, № 6, с. 1056-1067
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 577.32, 579.61
СПЕЦИФИЧЕСКИЕ И НЕСПЕЦИФИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ
ЛИПОПОЛИСАХАРИДА Yersinia pseudotuberculosis
С МОНОКЛОНАЛЬНЫМИ АНТИТЕЛАМИ, ОХАРАКТЕРИЗОВАННЫЕ
МЕТОДОМ АТОМНО-СИЛОВОЙ МИКРОСКОПИИ
© 2022 г. А.А. Бывалов*, **, #, В.С. Белозёров*, **, Б.А. Ананченко*, И.В. Конышев*, **
*Вятский государственный университет, Московская ул., 36, Киров, 610000, Россия
**Институт физиологии Коми научного центра УрО РАН, Первомайская ул., 50, Сыктывкар, 167982, Россия
#E-mail: byvalov@nextmail.ru
Поступила в редакцию 15.04.2022 г.
После доработки 13.07.2022 г.
Принята к публикации 15.07.2022 г.
Методом атомно-силовой микроскопии проведена оценка силы связи зонда, функционализиро-
ванного липополисахаридом Yersinia pseudotuberculosis, c моноклональными антителами, иммоби-
лизованными на слюде, а также вклада в это взаимодействие неспецифических факторов. Показано
отсутствие выраженных взаимодействий между «чистым» зондом и «чистой» или обработанной тем
или иным способом слюдой. Модификация «чистой» слюды (3-аминопропил)триэтоксисиланом
вызывала повышение силы взаимодействия с кантилевером, покрытым липополисахаридом. Обра-
ботка аминированной слюды глутаровым альдегидом приводила к еще большему возрастанию силы
неспецифического связывания. Последующая иммобилизация моноклональных антител на слюде,
предварительно обработанной (3-аминопропил)триэтоксисиланом и затем глутаровым альдеги-
дом, вызывала резкое ослабление взаимодействия, существенно менее выраженное при использо-
вании антител, комплементарных липополисахариду, по сравнению с гетерологичными антитела-
ми. Сила специфической связи с антителами антигенов, характеризующихся склонностью к агрега-
ции, таких как липополисахариды, по степени выраженности может уступать неспецифическому
компоненту взаимодействия, включающему «дезагрегацию», «развертывание» агломератов биомо-
лекул, иммобилизованных на поверхность зонда и слюды.
Ключевые слова: атомно-силовая микроскопия, сила связи, липополисахарид, моноклональные анти-
тела.
DOI: 10.31857/S0006302922050023, EDN: LIYGNE
взаимодействия. Для большинства же систем из-
Атомно-силовая микроскопия (АСМ) - один
меряются сложные, многокомпонентные взаи-
из наиболее информативных биофизических ме-
модействия, определяемые совокупностью от-
тодов, широко используемый в различных обла-
стях биологии. Сфера применения АСМ включа-
дельных, далеко не всегда идентифицируемых,
ет, в частности, исследование силовых характери-
межмолекулярных связей [6, 7]. При проведении
стик взаимодействия биообъектов на клеточном,
подобных исследований, направленных на опре-
субклеточном и молекулярном уровнях [1-5].
деление силы специфической связи между двумя
Измеряемая сила таких взаимодействий состав-
целевыми биомолекулами, одной из серьезных и
ляет, как правило, от единиц-десятков пиконью-
трудноразрешимых проблем является выявление
тонов до десятков-сотен наноньютонов [6]. Сре-
чисто «специфического» сигнала, свидетельству-
ди используемых модельных систем есть такие,
ющего о разрыве определяемой связи, на фоне
на которых можно с достаточно высокой вероят-
неспецифических сигналов фотодетектора. По-
ностью оценивать единичные межмолекулярные
нятия
«специфическая» и
«неспецифическая»
связь достаточно относительны и иногда они
Сокращения: АСМ
- атомно-силовая микроскопия, трактуются по-разному. Под специфическим вза-
тела, APTES - (3-аминопропил)триэтоксисилан,еГАти– имодействием обычно понимается лишь такое
взаимодействие, которое является предметом
глутаровый альдегид, ФБР - фосфатный буферный рас-
твор.
данного исследования. Специфическое связыва-
1056
СПЕЦИФИЧЕСКИЕ И НЕСПЕЦИФИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ
1057
ние для одного эксперимента может являться не-
связи, трактовка результатов подобных исследо-
специфическим для другого, имеющего иную
ваний во многом зависит от типа взаимодейству-
цель. Так, при оценке силы связи в системе «ан-
ющих биообъектов и условий проведения экспе-
тиген-антитело» для некоторых бактериальных
риментов.
белков (например, иммуноглобулинсвязываю-
В работах, направленных на количественное
щих белков ряда бактерий, в том числе Yersinia
определение сил связи между биообъектами, в
pseudotuberculosis [8]) взаимодействие с Fc-участ-
целях минимизации неспецифических взаимо-
ком антител рассматривается как специфиче-
действий существенным обстоятельством при-
ское, а все остальные - как неспецифические.
знается выбор способа иммобилизации на твер-
Однако в подавляющем большинстве исследова-
дой фазе (зонд или подложка) целевых молекул.
ний иммунохимических взаимодействий прово-
Так, прямой контакт между минеральным суб-
дится оценка силы специфического взаимодей-
стратом и биомолекулами приводит к изменению
ствия антигенной детерминанты с Fab-фрагмен-
их пространственной ориентации и снижению
том комплементарных антител. В настоящей
подвижности, что вызывает ослабление специ-
работе именно так трактуется специфическое
фического взаимодействия [5]. В этой связи для
взаимодействие, а под неспецифическим пони-
определения силы специфического взаимодей-
маются иные сопутствующие (дополнительные)
ствия предпочтительным признается использова-
силы связи.
ние гибких и длинных линкеров, например,
Взаимодействия между биологическими моле-
полиэтиленгликоля, что позволяет более точно
кулами и различными субстратами намного
отделять исследуемое специфическое взаимодей-
сложнее, чем между небиологическими молеку-
ствие от неспецифической адгезии. Линкер обес-
лами или поверхностями. Эта сложность обу-
печивает присоединенной молекуле свободу вра-
словлена рядом факторов: одновременным уча-
щения и тем самым делает сайты связывания бо-
стием множества различных молекул и различ-
лее доступными. Кроме того, он обеспечивает
ных нековалентных сил - ван-дер-ваальсовых,
прочное, ковалентное присоединение биомоле-
электростатических, сольватационных (гидрата-
кул к зонду с относительно низкой плотностью,
ционных, гидрофобных), стерических, энтро-
что позволяет регистрировать единичные межмо-
пийных и «специфических», а также гибкостью
лекулярные взаимодействия, увеличивает рассто-
биологических макромолекул и текучестью мем-
яние между поверхностью и молекулой, что ниве-
бран, если одним из участников взаимодействия
лирует возможные воздействия (например, элек-
является бактериальная клетка [9-11].
тростатической природы) подложки или
В силу различного толкования терминов «спе-
кантилевера на изучаемый процесс [5, 18, 19].
цифическое» и «неспецифическое» связывание,
При выборе условий осуществления экспери-
недостаточной изученности механизмов взаимо-
ментов такого рода и анализе получаемых резуль-
действия
«нефункционализированный зонд
-
татов следует учитывать влияние ряда факторов
нефункционализированная подложка», влияния
на формирование неспецифической составляю-
на это взаимодействие иммобилизованных био-
щей оцениваемых взаимодействий. Выражен-
молекул и др. доказательных общепринятых кри-
ность неспецифического связывания в значи-
териев идентификации специфических сигналов
тельной мере зависит от химического состава и
на силовых кривых до настоящего времени не
физико-химических особенностей, в частности,
представлено. Одни авторы принимают за неспе-
ионной силы раствора, в котором проводится
цифическое связывание такое, которое на хроно-
эксперимент [20-22]. При оценке результатов
грамме сигнала отведения проявляется неизмен-
подобных исследований следует учитывать, что
ностью угла его наклона до момента пересечения
увеличение времени контакта функционализиро-
кривой базисной линии непосредственно после
ванного зонда с субстратом (dwell time), а также
такового безотносительно к картине последую-
скорости нагружения (loading rate) приводит к
щего сигнала [12, 13]. Многие авторы при оценке
усилению неспецифического взаимодействия
силовых характеристик взаимодействий ориен-
[23]. В целях идентификации специфического
тируются на силу последнего отрыва, который и
сигнала, его обособления от неспецифической
отражает специфическое взаимодействие [14, 15].
компоненты в состав реакционной среды вклю-
В ряде экспериментов при наличии на хроно-
чают компонент, комплементарный одной из це-
грамме сигнала отведения нескольких скачков
левых биомолекул, который препятствует специ-
первый из них часто считают свидетельством раз-
фическому связыванию. Его удаление (отмыва-
рыва неспецифической связи [16]. На гистограм-
нием) из среды восстанавливает специфическое
мах распределения сил отрыва за неспецифиче-
взаимодействие [24]. Для снижения вклада в оце-
ское взаимодействие принимают, как правило,
ниваемое взаимодействие неспецифической со-
низкоамплитудные отрывы [17]. Существуют и
ставляющей предлагается использование в реак-
иные подходы к разграничению специфической
ционной среде детергентов (например, Твин 20),
и неспецифической составляющих оцениваемой
веществ с низкой реакционной способностью,
БИОФИЗИКА том 67
№ 6
2022
1058
БЫВАЛОВ и др.
применяемых, в частности, в составе блокирую-
гали облучению ультрафиолетом в течение
щих буферов в иммунохимических реакциях, та-
15 мин, а слюду скалывали непосредственно пе-
ких как бычий сывороточный альбумин [25]. Од-
ред модификацией. Модификацию аминными
нако в полной мере избавиться от неспецифиче-
группами поверхностей кантилевера и свеже-
ской составляющей регистрируемых сигналов
сколотой слюды в течение полутора часов прово-
взаимодействия практически невозможно. В этой
дили в эксикаторе в атмосфере аргона, рядом
связи важным представляется понимание струк-
с субстратами помещали две емкости, содержа-
туры интегрального сигнала для каждого отдель-
щие
30 мкл
(3-аминопропил)триэтоксисилана
ного эксперимента, осуществляемого при тех или
(APTES) (Sigma-Aldrich, США) и 10 мкл триэтил-
иных заранее заданных условиях.
амина («Reachem», Россия). После инкубации
слюду и зонд помещали в сухожаровой шкаф при
В настоящей работе мы попытались оценить
температуре 60°C на 1 ч для десорбции несвязан-
значимость неспецифической составляющей во
ных ковалентно с минеральными поверхностями
взаимодействии липополисахарида (ЛПС)
молекул APTES.
Y. pseudotuberculosis с моноклональными антите-
лами, комплеменарными и гетерологичными.
Функционализация кантилевера. Функциона-
лизацию поверхности кантилевера препаратом
ЛПС проводили с использованием Acetal-PEG-
МАТЕРИАЛЫ И МЕТОДЫ
NHS [31]. Аминированный зонд помещали на 2 ч
Биомолекулы, иммобилизуемые на поверхность
в 0.5 мл раствора хлороформа, содержащего 1 мг
кантилевера и слюды. Для функционализации по-
полиэтиленгликоля, любезно предоставленного
верхности кантилевера использовался ЛПС, вы-
доктором H. Gruber (Institute for Biophysics, Jo-
деленный из культуры клеток Y. pseudotuberculosis
hannes Kepler University of Linz, Linz, Austria), и
1b методом водно-фенольной экстракции по
30 мкл триэтиламина. После окончания реакции
Вестфалю [26]. Клетки штамма-продуцента вы-
зонд промывали трижды по 5 мин в хлороформе.
ращивали в жидкой питательной среде на основе
Для перевода ацетальной группы полиэтиленгли-
гидролизата казеина при температуре 10°C, кото-
коля в альдегидную зонд помещали в профиль-
рая способствует образованию полноразмерных
трованный через насадку Millipore (размер пор -
О-боковых цепей на молекуле ЛПС.
0.22 мкм) 1%-й раствор лимонной кислоты в де-
Поверхность слюды была покрыта монокло-
ионизированной воде на 10 мин. После актива-
нальными антителами (МКАт) двух типов:
ции ацетальной группы осуществляли промывку
МКАт2, являющимися комплементарными эпи-
зонда в деионизированной воде трижды по 5 мин
топу на О-боковых цепях ЛПС [27], и МКАт7,
и помещали его на 1 ч в раствор, содержащий
100 мкл ЛПС (50-100 мкг/мл) и 20 мМ NaCNBH3
комплементарными белковому эпитопу внешней
мембраны Y. pseudotuberculosis [28]. Перед исполь-
(Sigma-Aldrich, США). Непосредственно перед
зованием в процессе функционализации препа-
этой операцией раствор ЛПС центрифугировали
раты ЛПС и антител центрифугировали при
(13000 g, 15 мин), надосадочную жидкость филь-
13000 g в течение 15 мин и перед добавлением в
тровали через насадку Millipore (размер пор -
реакционную смесь фильтровали через насадку
0.22 мкм). Несвязавшиеся альдегидные группы
Millipore с диаметром пор 0.22 мкм (Merck, Гер-
блокировали добавлением в реакционную смесь
мания).
5 мкл 1 М моноэтаноламина («Reachem», Россия)
с последующей инкубацией в течение 10 мин.
Аминирование поверхностей кантилевера и слю-
Процесс функционализации завершали трех-
ды. При изучении силового взаимодействия ис-
кратным отмыванием зонда деионизированной
пользовали кантилеверы PNP-DB (NanoWorld,
водой трижды по 5 мин и его сушкой с использо-
Швейцария). Кантилеверы данного типа имеют
ванием фильтровальной бумаги.
зонд, заточенный методом термоокисления, за
счет чего на его поверхности формируется тонкий
Функционализация слюды. После окончания
слой оксида кремния [29]. В качестве подложки
аминирования пластинки слюды помещали в 8%-
использовали слюду. Слюда относится к группе
й раствор глутарового альдегида (ГА) на 30 мин.
природных алюмосиликатов со слоистой струк-
Данного времени достаточно для связывания глу-
турой. При скалывании слюды формируется мо-
тарового альдегида со свободными аминными
лекулярно-гладкая грань, содержащая на своей
группами на минеральной поверхности. Далее
поверхности силанольные группы (Si-OH). На-
пластинки отмывали фосфатным буферным рас-
личие на поверхности термоокисленного зонда и
твором (ФБР) трижды по 5 мин в объеме 3 мл, пе-
слюды силанольных групп позволяет проводить
реносили в ФБР, содержащий антитела в концен-
модификацию их поверхности с использованием
трации 50-100 мкг/мл, и инкубировали 1 ч при
органических силанов [30].
комнатной температуре. После функционализа-
Перед проведением модификации для удале-
ции пластинки тщательно отмывали ФБР. По
ния загрязнений с поверхности зондов их подвер-
окончании каждой из стадий функционализации
БИОФИЗИКА том 67
№ 6
2022
СПЕЦИФИЧЕСКИЕ И НЕСПЕЦИФИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ
1059
часть пластинок слюды отбирали и хранили в
кривых проводили в программе Nova_Px_3.4.0.
ФБР, оставшиеся подвергали последующим про-
Выбраковку графиков, имеющих отклонения, а
цедурам функционализации. Таким образом,
также обработку прошедших выбраковку графи-
кроме чистой (ничем не обработанной) слюды в
ков и расчет силовых характеристик проводили в
работе использовали четыре типа пластин, обо-
автоматическом режиме с использованием про-
значенных далее как «APTES» (обработанные
граммного обеспечения, разработанного автора-
только APTES), «ГА» (обработанные последова-
ми. За отрыв программа принимала скачок сигна-
тельно APTES и ГА), «МКАт2» (обработанные
ла фотодетектора, превышающий средний фоно-
последовательно APTES, ГА и МКАт2), «МКАт7»
вый шум не менее чем в четыре раза. Значения
(обработанные последовательно APTES, ГА и
оцениваемых параметров представлены в виде
МКАт7).
средних (Хmean ± I99).
Иммуноферментный анализ. Подтверждение
Общее число силовых кривых, описывающих
факта сенсибилизации слюды моноклональными
взаимодействие чистого (ничем не обработанно-
антителами проводили с использованием твердо-
го) кантилевера с чистой слюдой - 1325; слюдой,
фазного иммуноферментного анализа. Для этого
покрытой МКАт2 - 1325; слюдой, покрытой
в лунки 96-луночного планшета помещали пред-
МКАт7 - 1325; слюдой, покрытой APTES - 325;
варительно аминированные и обработанные пре-
слюдой, покрытой глутаровым альдегидом - 325;
паратами антител стеклянные пластинки площа-
взаимодействие ЛПС-кантилевера с чистой слю-
дой - 1575; взаимодействие со слюдой, покрытой
дью ~20 мм2. Методика функционализации стек-
МКАт2 - 1575; слюдой, покрытой МКАт7 - 1575;
ла была идентична той, которую использовали
слюдой, покрытой APTES - 325; слюдой, покры-
при функционализации слюды. Процедура твер-
той глутаровым альдегидом - 325. Для программ-
дофазного иммуноферментного анализа стан-
ной выбраковки не подлежащих дальнейшему
дартна и описана ранее [32]. Результаты реакции
анализу графиков были предложены четыре кри-
оценивали по показателю ОП492.
терия: 1) горизонтальность базисной линии на
Атомно-силовая микроскопия. Изучение по-
последних 100 точках, по которым определяются
верхности образцов слюды проводили полукон-
положение базисной линии и амплитуда шума;
тактным методом на воздухе в режиме регистра-
2) отсутствие изменений сигнала в положитель-
ции сигнала «высота» с использованием зондов
ную область относительно базисной линии после
NSG30 с радиусом закругления иглы
10 нм
последнего отрыва (линейность всей базисной
(TipsNano, Эстония). Сканирование проводили
линии); 3) линейность силовой кривой отведения
на микроскопе «NtegraPrima» («NT-MDT», Рос-
по первым 40 точкам (плотный контакт между
сия) при частоте развертки 1 Гц, изображения по-
зондом и подложкой), которые используются для
лучены с разрешением 512×512 пикселей. Анализ
последующего пересчета показаний фотодетек-
полученных изображений и расчет показателей
тора в значения силы; 4) отсутствие резких, пре-
шероховатости проводили с помощью программ-
вышающих шум, изменений сигнала в положи-
ного обеспечения Nova_Px v.3.4.0. Эффектив-
тельную область на силовой кривой от момента
ность процедуры модификации и функционали-
отведения кантилевера до пересечения с базис-
зации поверхности слюды оценивали по показа-
ной линией (момент контакта). Силовые кривые,
телям шероховатости: Ra (среднеарифметическая
не соответствующие представленным выше кри-
шероховатость) и Rz (шероховатость поверхности
териям, в определении силовых характеристик не
были использованы.
по выбранным пяти максимальным высотам и
впадинам) [33].
Силовая спектроскопия. Оценку силовых ха-
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
рактеристик межмолекулярного взаимодействия
В настоящей работе мы оценивали методом
проводили с использованием атомно-силового
АСМ силовые характеристики взаимодействия
микроскопа «Ntegra Prima» с измерительным
между зондом, функционализированным препа-
вкладышем AU-208 и зондов PNP-DB
ратом ЛПС и слюдой, покрытой комплементар-
(NanoWorld, Швейцария), имеющих радиус за-
ными к О-антигену липополисахарида антитела-
кругления около 10 нм и жесткость консоли
ми МКАт2. Для оценки влияния на названное
0.06 нН/нм. Все измерения проводили в жидкой
специфическое взаимодействие иных факторов
среде (ФБР)при значении скорости нагружения
измеряли силу связи между этим зондом и «чи-
(loading rate), равной 37 нН/с (скорость подведе-
стой», интактной слюдой, слюдой, аминирован-
ния и отведения образца была равна 0.5 мкм/с),
ной с помощью APTES, а также слюдой, обрабо-
время контакта зонда и образца - 1 с, частота дис-
танной APTES с последующей инкубацией в
кретизации - 500 Гц.
растворе ГА, - две названные операции последо-
Процедура проведения силовой спектроско-
вательно предшествуют обработке слюды антите-
пии изложена в работе [32]. Обработку силовых
лами. Кроме того, дополнительным референс-
БИОФИЗИКА том 67
№ 6
2022
1060
БЫВАЛОВ и др.
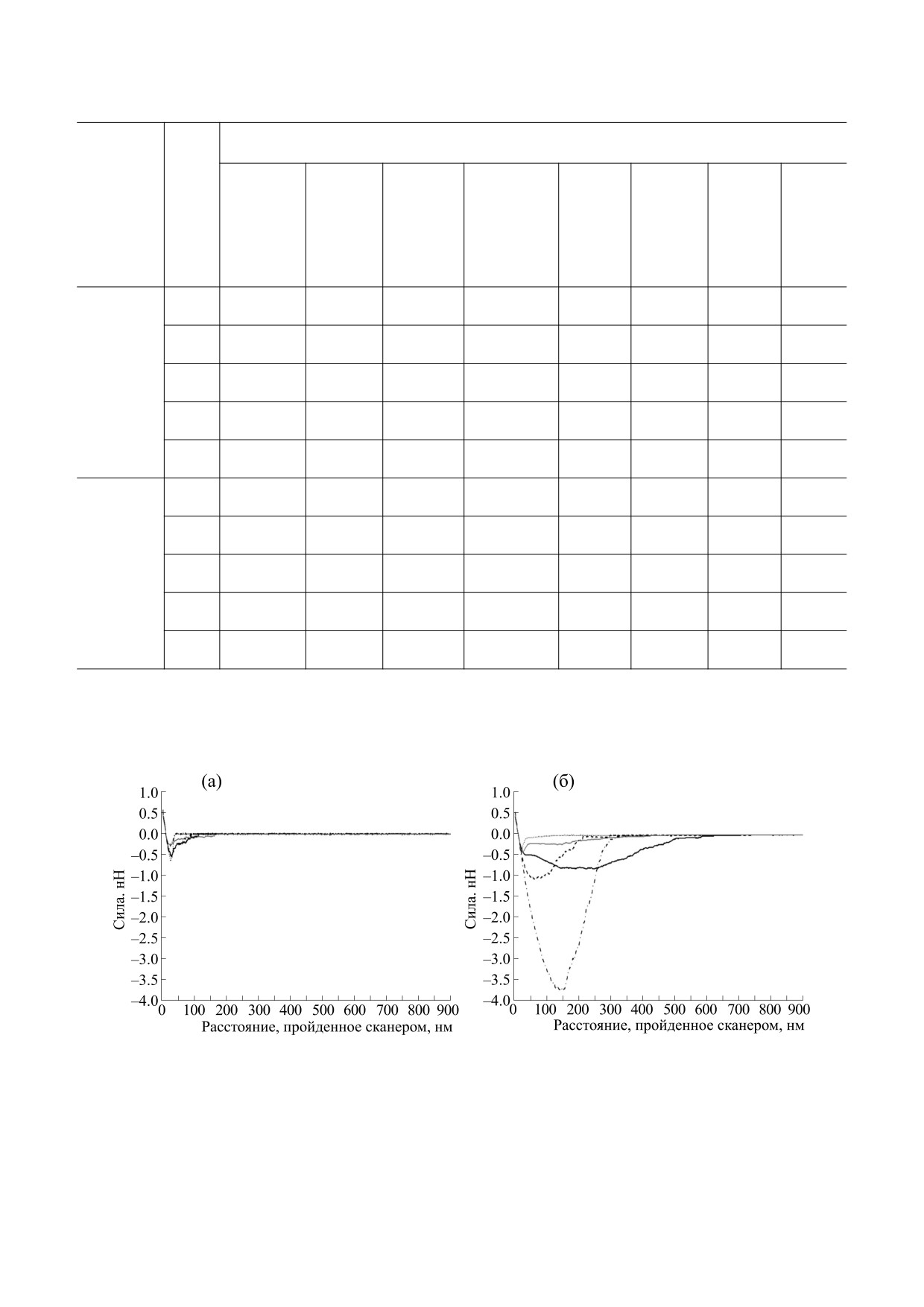
Рис. 1. (а) - Принципиальная схема измерения силовых характеристик взаимодействия методом АСМ. (б) - Пример
силовой кривой взаимодействия «МКАт2 - ЛПС» где 1 - базисная линия, 2 - момент контакта зонда с субстратом, 3 -
максимальная адгезия, 4 - момент разрыва последней связи, 5 - дистанция последнего отрыва, 6 - работа сил адгезии
(площадь).
препаратом служила слюда, функционализиро-
ционализированной антителами МКАт2 и
ванная по той же схеме, но с заменой антител
МКАт7, характеризуется достаточно равномер-
МКАт2 на моноклональные антитела МКАт7, не
ным и плотным распределением отдельных агре-
взаимодействующие с ЛПС. Все пять вариантов
гатов, имеющих приблизительно одинаковые
слюды были использованы и для соответствую-
размеры (рис. 2г,д). Аналогичная картина по-
щих измерений с «интактным», ничем не обрабо-
верхности функционализированной слюды на-
танным зондом этой же серии. Схематическое
блюдалась и при ином методе ковалентной иммо-
изображение процедуры экспериментов пред-
билизации антител [32]. Эффективность сенси-
ставлено на рис. 1а.
билизации поверхности слюды антителами была
подтвержденаи методом твердофазного иммуно-
В предварительных экспериментах методом
ферментного анализа: средние значения ОП492
атомно-силовой микроскопии оценивали струк-
для субстрата, обработанного МКАт2 и МКАт7,
туру поверхности слюды вышеуказанных пяти
составили соответственно 1.981 и 1.551; в контро-
типов. Приведенные в табл. 1 результаты свиде-
ле (слюда, обработанная APTES) это значение со-
тельствуют о том, что каждая последующая опе-
ставило 0.291.
рация обработки слюды, особенно иммобилиза-
ция антител, приводила к повышению ее шерохо-
Для характеристики силы взаимодействия
ватости по показателям Ra и Rz. Иллюстрирует
ЛПС-МКАт определяли несколько показателей,
вышесказанное рис. 2, на котором представлены
измеряемых при отведении пьезостолика от кан-
АСМ-изображения поверхности использованных
тилевера (рис. 1а,б): количество отрывов (1);
в работе субстратов. Поверхность слюды, функ-
среднюю силу (амплитуду) всех отрывов (2); силу
Таблица 1. Шероховатость образцов слюды
Шероховатость
№
Слюда
Ra, нм
Rz, нм
1
Чистая
0.019 ± 0.002
0.089 ± 0.011
2
APTES
0.031 ± 0.004
0.136 ± 0.029
3
ГА
0.041 ± 0.010
0.191 ± 0.038
4
МКАт2
0.330 ± 0.055
1.277 ± 0.332
5
МКАт7
0.277 ± 0.048
1.133 ± 0.260
БИОФИЗИКА том 67
№ 6
2022
СПЕЦИФИЧЕСКИЕ И НЕСПЕЦИФИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ
1061
Рис. 2. АСМ-изображения поверхности свежесколотой слюды (а), слюды, обработанной APTES (б), обработанной
последовательно APTES и ГА (в), обработанной последовательно APTES, ГА, МКАт2 (г) или МКАт7 (д) при размерах
области сканирования 0.5 × 0.5 мкм.
(амплитуду) последнего отрыва (3); дистанцию,
Анализ индивидуальных и усредненных кри-
на которой произошел последний отрыв (4);
вых подведения для всех десяти пар сравнивае-
работу, затрачиваемую на разрыв всех связей и
мых объектов показал отсутствие значимых сви-
соответствующую площади между кривой отведе-
детельств их притяжения или отталкивания до
ния и базисной линией
(5); максимальную
момента касания при использованных условиях
адгезию, определяемую как наибольшее рас-
эксперимента (данные не представлены). Взаи-
стояние от точки на кривой отведения до базис-
модействия регистрировали только после сопри-
ной линии (6); длину ассоциата (агломерата)
косновения и односекундного контакта зонда и
ЛПС-МКАт, под которой в данном эксперименте
субстрата.
понимается расстояние от поверхности слюды
до острия кантилевера в момент разрыва послед-
Как видно из рис. 3а, все неспецифические
ней связи, определяемое как сумма длины ассо-
взаимодействия материала интактного (чистого)
циата ЛПС (LЛПС) и длины ассоциата антител
зонда с разнокачественными вариантами слюды,
включая обработанные антителами двух специ-
(LМКАт) (7).
фичностей, являются маловыраженными отно-
Обобщенные результаты оценки силовых ха-
сительно функционализированного зонда. Из
рактеристик взаимодействий представлены в
числа пяти типов слюды, по-видимому, несколь-
табл. 2 в виде средних значений указанных выше
ко большее взаимодействие с интактным канти-
показателей для всех десяти пар объектов.
левером отмечено для слюды «APTES» (рис. 3а
На рис. 3 и 4 представлены также усредненные
и 4). Эти данные качественно подтверждают ре-
хронограммы сигнала фотодетектора (в коорди-
зультаты авторов работы [11], зарегистрировав-
натах «сила - дистанция, пройденная сканером»)
ших притяжение аминированного с помощью
для всех пяти типов слюды при отведении ин-
APDES зонда (нитрид кремния) к поверхности
тактного и функционализированного ЛПС кан-
стекла; не обработанный APDES зонд указанного
тилеверов. При усреднении хронограмм за нуле-
свойства не проявлял. Авторы объясняют данный
вую точку принимали точку пересечения кривой
эффект адгезии действием электростатических
отведения с базисной линией.
(положительно заряженные аминогруппы и от-
БИОФИЗИКА том 67
№ 6
2022
1062
БЫВАЛОВ и др.
Таблица 2. Влияние метода обработки слюды на силовые характеристики ее взаимодействия с кантилеверами
«чистым» и функционализированным ЛПС Y. pseudotuberculosis
Показатели, Хmean ± I99
Коли-
Дистан-
чество
Кантилевер
Слюда
Сила
Работа сил
Средняя
Максималь-
Длина
ция
Коли-
отведе-
последнего
адгезии,
сила всех
ная адгезия,
ассоциата,
послед-
чество
ний с
отрыва, нН
Дж × 10-15
отрывов, нН
нН
нм
него
отрывов
отрыва-
отрыва, нм
ми, %
Чистая
1.078 ± 0.132
0.12 ± 0.02
1.078 ± 0.132
1.148 ± 0.106
0.7 ± 0.4
18.6 ± 2.2
1.00 ± 0.00
65.5
APTES
1.051 ± 0.260
0.20 ± 0.08
1.042 ± 0.208
1.097 ± 0.248
13.2 ± 9.8
30.7 ± 10.7
1.30 ± 0.21
100.0
Чистый
ГА
0.926 ± 0.506
0.16 ± 0.19
1.063 ± 0.447
0.963 ± 0.501
7.5 ± 6.2
22.9 ± 12.3
1.42 ± 0.67
80.0
МКАт2
0.686 ± 0.184
0.16 ± 0.09
0.778 ± 0.168
0.852 ± 0.207
20.3 ± 6.8
31.7 ± 7.9
1.46 ± 0.30
51.3
МКАт7
0.585 ± 0.187
0.11 ± 0.05
0.602 ± 0.150
0.655 ± 0.174
18.6 ± 5.2
28.3 ± 6.7
1.28 ± 0.21
43.3
Чистая
0.538 ± 0.035
0.10 ± 0.02
0.547 ± 0.034
0.627 ± 0.033
12.2 ± 3.9
21.1 ± 4.0
1.10 ± 0.03
73.5
APTES
1.876 ± 0.302
1.28 ± 0.32
2.117 ± 0.232
2.365 ± 0.322
103.7 ± 34.5
134.9 ± 33.5
1.82 ± 0.20
93.6
Функцио-
нализиро-
ГА
5.461 ± 0.375
6.05 ± 0.61
5.199 ± 0.220
6.032 ± 0.379
77.0 ± 5.2
167.8 ± 8.9
2.76 ± 0.19
99.3
ванный ЛПС
МКАт2
2.267 ± 0.211
2.66 ± 0.45
2.650 ± 0.161
2.383 ± 0.215
85.3 ± 6.6
123.0 ± 9.6
1.96 ± 0.12
78.0
МКАт7
1.169 ± 0.134
0.59 ± 0.16
1.447 ± 0.124
1.251 ± 0.129
33.4 ± 4.0
52.8 ± 5.6
1.53 ± 0.08
72.5
Рис. 3. Усредненные хронограммы сигнала отведения слюды от чистого кантилевера (а) и кантилевера, покрытого ЛПС (б).
Хронограммы взаимодействия кантилеверов выполнены в виде точек (чистая слюда), пунктирной линии (слюда
«APTES»), штрих-пунктирной линии (слюда «ГА»), сплошной черной линии (слюда «МКАт2») и сплошной серой линии
(слюда «МКАт7»).
БИОФИЗИКА том 67
№ 6
2022
СПЕЦИФИЧЕСКИЕ И НЕСПЕЦИФИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ
1063
Рис. 4. Усредненные хронограммы сигнала отведения чистой слюды (а) и слюды, обработанной препаратами APTES (б),
глутарового альдегида (в), МКАт7 (г) и МКАт2 (д) от чистого кантилевера (точки) и кантилевера,
функционализированного ЛПС (сплошная линия).
рицательно заряженная поверхность стекла) и
же обработанной МКАт2 и МКАт7, выход кривых
гидрофобных взаимодействий. Последние опре-
на базисную линию происходит значительно
позднее, через 100 и более нм (рис. 3а).
деляются заменой силанольных групп на поверх-
ности зонда на аминогруппы [11]. Можно отме-
Иная картина наблюдается при использова-
тить, что усредненная кривая для интактной слю-
нии кантилевера, функционализированного
ды и слюды «ГА» выходит на базисную линию
ЛПС (рис. 3б и 4). Если наличие/отсутствие мо-
через ~40 нм после начала отведения пьезостоли-
лекул ЛПС на поверхности зонда мало отражает-
ка; для слюды, модифицированной APTES, а так-
ся на его адгезивности к интактной слюде
БИОФИЗИКА том 67
№ 6
2022
1064
БЫВАЛОВ и др.
группами [36]. Полисахаридная часть ЛПС содер-
жит большое число гидроксильных групп, при
этом даже в случае образования слабых водород-
ных связей в системе «альдегидная группа - гид-
роксильная группа» интегральная сила взаимо-
действия множества отдельных частных связей
может достигать больших величин. Общеизвест-
но также, что альдегидные группы легко вступают
в реакцию с аминогруппами с образованием ко-
валентной связи. Можно предположить, что зна-
чительная сила связи обусловлена в том числе
взаимодействием между свободными аминогруп-
пами молекулы ЛПС и альдегидными группами
подложки, поскольку известно, что содержание
общего белка в препаратах ЛПС, выделенных да-
же жесткими методами, может достигать 1.5-
2.5%. Использованный нами в работе препарат
ЛПС включал в качестве прочно связанной при-
Рис. 5. Средняя сила разрыва последней связи. Довери-
меси 1.3% белка.
тельные интервалы указаны для p = 0.01.
Заключительная стадия функционализации
моноклональными антителами слюды, предвари-
(рис. 4а), то в остальных четырех случаях функ-
тельно обработанной последовательно APTES и
ционализированный ЛПС кантилевер вызывает
затем ГА, при использовании ЛПС-кантилевера
существенно более выраженное взаимодействие с
приводила к резкому падению силы взаимодей-
субстратами по сравнению с интактным кантиле-
ствия, выраженному в намного большей степени
вером. Последнее объясняется в том числе тем,
для слюды «МКАт7» по сравнению с «МКАт2»
что выделенные из бактериальной массы препа-
(табл. 2, рис. 3б). Последнее обстоятельство ука-
раты ЛПС склонны к агрегации во времени и, в
зывает на то, что «пришитые» антитела, как
зависимости от его концентрации и состава рас-
МКАт2, так и МКАт7, экранируют на поверхно-
твора, могут находиться в виде высокомолекуляр-
сти слюды реакционно-активные группы ГА,
ных структур (ассоциатов, агломератов) различ-
способные взаимодействовать с молекулами
ных формы и размера [34]. При взаимодействии
ЛПС. Вместе с тем существенная разница в адге-
функционализированного ЛПС кантилевера и
зивности функционализированного ЛПС канти-
аминированной слюды регистрируются выра-
левера к слюде, покрытой комплементарными
женные неспецифические силовые взаимодей-
этому антигену и гетерологичными антителами,
ствия. Эти данные согласуются с результатами
свидетельствует о весомом вкладе в регистрируе-
работы [35], в которой при помощи установки для
мое взаимодействие «МКАт2-ЛПС» специфиче-
изучения поверхностных сил авторы показали,
ской составляющей.
что сила взаимодействия в системе «ЛПС - ами-
Наиболее часто используемыми и показатель-
нированная APTES слюда» значительно превы-
ными силовыми характеристиками изучаемых
шает взаимодействие в системе «ЛПС - немоди-
взаимодействий в разнообразных системах «ли-
фицированная слюда». Вероятно, между ами-
ганд - рецептор» являются сила последнего раз-
ногруппами модифицированной слюды и
рыва связей и площадь между кривой отведения и
молекулами ЛПС происходит образование боль-
базисной линией (работа сил адгезии). Послед-
шого числа связей, имеющих кислотно-основ-
ний отрыв в такого рода исследованиях, как пред-
ную природу, обусловленную присутствием на
полагается [14], отражает разрыв специфической
молекулах ЛПС свободных фосфатных, карбок-
связи непосредственно между молекулами, им-
сильных и гидроксильных групп.
мобилизованными на поверхность зонда и суб-
страта. Площадь между кривыми подведения и
Силы взаимодействий между функционализи-
отведения соответствует работе, которую необхо-
рованным ЛПС зондом и аминированной слюдой
димо приложить для разрыва всех связей, как
резко возрастают после последующей ее обработ-
специфических, так и неспецифических [37].
ки глутаровым альдегидом. Природа взаимодей-
ствий в системе «ЛПС - слюда «ГА»» не совсем
Рис. 5 и 6 иллюстрируют наличие специфиче-
ясна. Так, считается, что карбогидраты слабо вза-
ской и неспецифической составляющих взаимо-
имодействуют с альдегидами. Вместе с тем, альде-
действий при использовании кантилеверов двух
гиды могут в порядке убывания силы взаимодей-
типов, функционализированного ЛПС и интакт-
ствия реагировать с ε-, α-, вторичными амино-
ного, для двух наиболее демонстративных крите-
группами и гидроксильными функциональными
риев силы разрываемых связей - силе последнего
БИОФИЗИКА том 67
№ 6
2022
СПЕЦИФИЧЕСКИЕ И НЕСПЕЦИФИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ
1065
отрыва (рис. 5) и работе сил адгезии (рис. 6). При-
менительно к интактному кантилеверу по указан-
ным показателям значимость процедуры той или
иной обработки слюды существенно ниже по
сравнению с ЛПС-кантилевером. Обращает на
себя внимание аналогичный характер результа-
тов сравнительной оценки амплитуды последне-
го отрыва и работы, затрачиваемой на разрыв всех
связей. В частности, более высокие значения обо-
их показателей для слюды, покрытой МКАт2, по
сравнению с МКАт7, при использовании функ-
ционализированного ЛПС кантилевера подтвер-
ждают наличие специфического компонента свя-
зи (рис. 5 и 6). Об этом можно судить при сравне-
нии названных систем не только по двум
вышеуказанным показателям, но и таким, как ди-
станция последнего отрыва, длина ассоциата,
среднее количество отрывов, доля отведений,
имеющих хотя бы один отрыв (табл. 2). По-види-
мому, относительно более высокие значения ис-
пользованных показателей для пары «МКАт2 -
Рис. 6. Площадь между кривыми подведения и отведе-
ЛПС» объясняются наличием в препарате ЛПС,
ния (работа сил адгезии). Доверительные интервалы
иммобилизованном на зонде, достаточно круп-
указаны для p = 0.01.
ных ассоциатов (агломератов) субъединиц анти-
гена, отрыв которых от субстрата требует преодо-
количество отрывов» для субстратов без МКАт2,
ления не только сил специфического связыва-
ния, но и опосредуемых ими неспецифических
в первую очередь, для пары «МКАт7 - ЛПС»
сил дезагрегации ассоциата, очевидно, превыша-
(рис. 3, табл. 2). В этой связи можно ли утвер-
ющих таковые для пары «МКАт7 - ЛПС», в кото-
ждать, что каждый последний отрыв в системе
рой нет специфического взаимодействия «анти-
«МКАт2 - ЛПС» содержит специфическую ком-
ген - антитело». Вместе с тем достаточно высокие
поненту, то есть отражает ли он в том числе и раз-
абсолютные значения показателей взаимодей-
рыв связи между Fab-фрагментом МКАт2 и О-ан-
ствия в паре «МКАт7 - ЛПС» могут свидетель-
тигеном липополисахарида? По-видимому,
ствовать о том, что степень выраженности сум-
нельзя, поскольку, как можно судить по пред-
марной неспецифической составляющей во взаи-
ставленным в табл. 2 результатам, силы неспеци-
модействии «МКАт2 - ЛПС» по крайней мере
фического связывания достаточно велики и мо-
сопоставима с таковой у его специфического
гут целиком опосредовать последний отрыв. Да-
компонента, если не выше. Так, по литературным
же если отрыв (в том числе и последний)
данным сила единичной связи между молекулами
включает специфическую в указанном выше
антигена и антитела в подобных системах зави-
смысле составляющую, прикладываемая сила
сит, в частности, от скорости отведения и, как
разрывает не только такую связь, но и сопутству-
правило, не превышает 250-280 пН [38]. В наших
ющую неспецифическую, определяемую в том
экспериментах средняя сила последнего отрыва в
числе и дезагрегацией биомолекул. Поэтому при
паре «МКАт2 - ЛПС» превысила 2.2 нН (табл. 2),
количественной оценке взаимодействия «анти-
что свидетельствует о разрыве сильного неспеци-
ген - антитело» ориентироваться на абсолютные
фического взаимодействия, которое может пре-
вышать специфичекое взаимодействие, и, кроме
значения силы последнего отрыва не следует, во
того, единовременном разрыве множественной
всяком случае, как и в наших экспериментах, для
специфической связи «антиген - антитело», по-
биомолекул, имеющих склонность к агломера-
скольку область контакта острия кантилевера и
ции. Точность таких измерений относительна и в
субстрата значительно превышает размеры одной
силу других причин - шероховатости соприкаса-
белковой молекулы [39, 40].
ющихся поверхностей, адсорбции на них примес-
ных компонентов, способа ковалентного связы-
Следует подчеркнуть, что последний (и часто
вания биомолекул к твердой фазе, определяюще-
единственный) отрыв в оцениваемой связи
го доступность реакционноспособных сайтов для
«МКАт2 - ЛПС» содержит неспецифическую со-
ставляющую. На это указывают относительно
иммуноспецифического взаимодействия, и др.
большие значения показателя «сила последнего
[11, 41]. Кроме того, существуют разные подходы
отрыва» и, косвенно, показателей «дистанция по-
к идентификации отрыва как отрыва, выбраковке
следнего отрыва», «длина ассоциата», «среднее
отдельных хронограмм, иные различия в проце-
БИОФИЗИКА том 67
№ 6
2022
1066
БЫВАЛОВ и др.
дуре измерений, статистической обработке и
со слюдой, предварительно обработанной APTES
трактовке полученных результатов.
и далее ГА, за счет последующего блокирования
препаратами антител ее реакционно-активных
Литературные данные, а также вышеприве-
групп, способных к взаимодействию с ЛПС. Учи-
денные результаты экспериментов свидетель-
тывая вышеизложенное, количественное выявле-
ствуют о том, что применительно к изучению
ние из интегрального сигнала фотодетектора
методом АСМ силы связи между двумя биообъек-
компонента, который бы соответствовал чисто
тами неспецифические связи можно квалифици-
специфической связи «молекула антигена - мо-
ровать следующим образом.
лекула антител», представляется сложной про-
1. Взаимодействие минеральных материалов
блемой вследствие неотъемлемого и не всегда
зонда и подложки (субстрата) между собой.
строго идентифицируемого влияния на разрыва-
2. Взаимодействие каждого из них с биообъек-
емую связь неспецифических факторов. Их при-
том (биомолекулами), иммобилизованным на
рода, количество и степень выраженности опре-
другом.
деляются выбранными условиями проведения
3. К числу неспецифических взаимодействий
эксперимента. В подобного рода исследованиях
следует отнести, очевидно, также и события, ко-
целесообразным и продуктивным представляется
торые вызывают изменения сигнала, отражаю-
использование нескольких, желательно ради-
щие дезагрегацию биомолекул; исследуемые
кально различающихся и взаимодополняющих
биомолекулы могут иметь различные химический
методических подходов, совокупные результаты
состав, физико-химические свойства, простран-
применения которых должны давать более объек-
ственную организацию или представлять собой
тивную, взвешенную оценку подобных взаимо-
агломераты, как, например, препараты бактери-
действий.
альных липополисахаридов [32, 42]. Дезагрегация
такого агломерата может иметь место и в отсут-
ИСТОЧНИКИ ФИНАНСИРОВАНИЯ
ствие специфической связи, но наличие послед-
ней требует повышения суммарной работы, за-
Исследование выполнено при финансовой
трачиваемой на разрыв всех связей.
поддержке Российского фонда фундаментальных
4. Вполне вероятно также, что существенный
исследований в рамках научного проекта № 20-
вклад в силу формируемой связи вносит и компо-
34-90013, а также гранта Президента Российской
нента, определяемая неспецифическим, единич-
Федерации для государственной поддержки мо-
ным или множественным, взаимодействием со-
лодых российских ученых-кандидатов наук
прикасающихся участков биомолекул, не имею-
(№ МК-3383.2021.1.4).
щих прямого отношения к определяемой связи
«антигенная детерминанта - Fab-фрагмент анти-
КОНФЛИКТ ИНТЕРЕСОВ
тел». Указанная компонента, очевидно, также
может влиять на количество работы, требуемой
Авторы заявляют об отсутствии конфликта
для дезагрегации агломератов макромолекул.
интересов.
ВЫВОДЫ
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
Совокупность представленных эксперимен-
Настоящая статья не содержит каких-либо ис-
тальных данных свидетельствует о том, что неспе-
следований с участием людей или животных в ка-
цифические связи, регистрируемые в проведен-
честве объектов исследований.
ных экспериментах, включают взаимодействия
не только между минеральными носителями, но
СПИСОК ЛИТЕРАТУРЫ
и, в гораздо большей степени, взаимодействия,
определяемые иммобилизованными на них био-
1. T. Nandi and S.R.K. Ainavarapu, Emerg. Top. Life Sci.
молекулами. По-видимому, последние регистри-
5 (1), 103 (2021).
руются в результате дезагрегации агломератов
2. M.Y. Amarouch, J. El Hilaly, and D. Mazouzi, Scan-
ЛПС (и в меньшей степени - агломератов анти-
ning 2018, 7801274 (2018).
тел) при отведении сенсибилизированного ЛПС
3. S. Liu and Y. Wang, Scanning 32(2), 61 (2010).
кантилевера от субстрата, покрытого антителами;
4. Z. Bednarikova, Z. Gazova, F. Valle, and E. Bystreno-
указанная неспецифическая составляющая взаи-
va, J Microsc. 280 (3), 241 (2020).
модействия ЛПС кантилевера с субстратом, обра-
5. C. Lo Giudice, A.C. Dumitru, and D. Alsteens, Anal
ботанным комплементарными антителами
Bioanal Chem 411 (25), 6549 (2019).
МКАт2, оказалась существенно выше по сравне-
6. L.S. Dorobantu and M.R. Gray, Scanning 32 (2), 74
нию с гетерологичными антителами МКАт7. При
(2010).
этом следует подчеркнуть резкое снижение не-
7. C. Formosa-Dague, M. Castelain, H. Martin-Yken,
специфического связывания ЛПС-кантилевера
et al., Microorganisms 6 (2), 39 (2018).
БИОФИЗИКА том 67
№ 6
2022
СПЕЦИФИЧЕСКИЕ И НЕСПЕЦИФИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ
1067
8.
E.V. Sidorin and T.F. Solov'eva. Biochemistry (Mosc).
26. O. Westphal and K. Jann, Methods in Carbohydrate
76 (3), 295 (2011).
Chemistry 5, 83 (1965).
9.
J. Israelachvili, Q Rev Biophys. 38 (4), 331 (2005).
27. A.A. Byvalov, L.G. Dudina, S.G. Litvinets, et al., Prikl.
10.
A. Elbourne, J. Chapman, A. Gelmi, at al., J Colloid
Biokhim. Mikrobiol. 50 (2), 203 (2014).
Interface Sci. 546, 192 (2019).
28. A.A. Byvalov, L.G. Dudina, A.V. Chernyad'ev, et al.,
Mol. Gen. Mikrobiol. Virusol. 33 (2), 32 (2015).
11.
T.S. Tsapikouni and Y.F. Missirlis, Colloids and Sur-
faces B: Biointerfaces 75 (1), 252 (2010).
29. H. He, J. Zhang, J. Yang, and F. Yang, Microsyst.
Technol. 23, 1799 (2017).
12.
O.H. Willemsen, M.M. Snel, K.O. van der Werfs, et al.,
Biophys. J. 75, 2220 (1998).
30. S. Kim, H.K. Christenson, and J.E. Curry, Langmuir
18, 2125 (2002).
13.
L. Chih-Kung, W. Yu-Ming, H. Long-Sun, and
L. Shiming, Micron 38 (5), 446 (2007).
31. A. Ebner, L. Wildling, and H.J. Gruber, Methods Mol.
Biol. 1886, 117 (2019).
14.
M. Utjesanovic, T.R. Matin, K.P. Sigdel, et al., Sci
32. B. Ananchenko, V. Belozerov, A. Byvalov, et al., Int. J.
Rep. 9 (1), 451 (2019).
Biol. Macromol. 156, 841 (2020).
15.
15. C.T. Lim, E.H. Zhou, A. Li, et al., Mater. Sci. Eng.
33. ISO 4287:2000, Geometrical product specification
26 (8), 1278 (2006).
(GPS). Surface texture. Profile method. Terms, defini-
16.
M. Targosz, P. Czuba, R. Biedron, et al., ACTA PHYS-
tions and surface texture parameters, International Or-
ICA POLONICA A 109 (3), 421 (2005).
ganization of Standardization.
17.
E. Brzozowska, A. Leśniewski, S. Sęk, et al., Sci. Rep.
34. N.C. Santos, A.C. Silva, M.A. Castanho, et al., Chem-
8 (1), 10935 (2018).
biochem. 4 (1), 96 (2003).
18.
И.В. Сафенкова, А.В. Жердев и Б.Б. Дзантиев,
35. Q. Lu, J. Wang, A. Faghihnejad, et al., Soft Matter 7
Успехи биологической химии 52, 281 (2012).
(19), 9366 (2011).
19.
A. Beaussart and S. El-Kirat-Chatel, Cell Surf. 5,
36. I. Migneault, C. Dartiguenave, M.J. Bertrand, and
100031 (2019).
K.C. Waldron, Biotechniques 37 (5), 790 (2004).
20.
Y. Wang, J. Wang, J. Sun, et al., J Nanosci Nanotech-
37. R. Popeski-Dimovski, Carbohydr Polym. 123, 146
nol 19 (12), 7584 (2019).
(2015).
21.
S. Ramezanian, H.X. Ta, B. Muhunthan, and N. Abu-
38. H. Gao, X.X. Zhang, and W.B. Chang, Front Biosci.
Lail, Biointerphases 13 (4), 041005 (2018).
10, 1539 (2005).
22.
M.M. Elmahdy, A. Drechsler, C. Gutsche, et al., Lang-
39. Y. Wang, J. Wang, S. Huang, et al., Int J Biol Macro-
muir 25 (22), 12894 (2009).
mol. 134, 28 (2019).
23.
E. Celik and V.T. Moy, J Mol Recognit. 25 (1), 53
40. C. Wang, J. Wang, and L. Deng, Nanoscale Res Lett. 6
(2012).
(1), 579 (2011).
24.
J.R. King, C.M. Bowers, and E.J. Toone, Langmuir 31
41. H.J. Butta, B. Cappella, and M. Kappl, Surface Sci-
(11), 3431 (2015).
ence Reports 59, 1 (2005).
25.
J. Wakayama, H. Sekiguchi, S. Akanuma, et al., Anal
42. W. Kaca, J. Gleńska-Olender, I. Konieczna, et al.,
Biochem. 380 (1), 51 (2008).
Methods Mol Biol. 2021, 273 (2019).
Specific and Non-Specific Interactions of Yersinia pseudotuberculosis
Lipopolysaccharide with Monoclonal Antibodies Assessed by Atomic Force Microscopy
A.A. Byvalov*, **, V.S. Belozerov*, **, B.A. Ananchenko*, and I.V. Konyshev*, **
*Vyatka State University, Moskovskaya ul. 36, Kirov, 610000 Russia
**Institute of Physiology, Кomi Science Centre of the Ural Branch of the Russian Academy of Sciences,
Pervomaiskaya ul. 50, Syktyvkar, 167982 Russia
Atomic force microscopy (AFM) was employed to quantify the interaction force between a probe coated with
Yersinia pseudotuberculosis lipopolysaccharide (LPS) (a LPS-functionalized probe) and monoclonal anti-
bodies immobilized on mica; contribution of nonspecific factors to this interaction was evaluated. No evi-
dence was observed for the presence of discernable interactions between the "pure" probe and "pure" mica or
mica subjected to any treatments. Surface modification of "pure" mica with (3-aminopropyl)triethoxysilane
led to an increase in the forces of interaction between an LPS-covered AFM cantilever tip and mica. Treat-
ment of surface-aminated mica with glutaric aldehyde caused further increase in nonspecific binding force.
Subsequent immobilization of monoclonal antibodies on mica pretreated with (3-aminopropyl)triethoxysi-
lane and then with glutaric aldehyde induced a sharp decrease in the interaction, decrease was significantly
less pronounced in the presence of specific anti-lipopolysaccharide antibodies than decrease in the presence
of heterologous antibodies. The force of specific binding to antibodies of antigens that have a tendency to
aggregate, such as lipopolysaccharides, can be lower than the force of nonspecific interaction, including "dis-
aggregation", "unfolding" of agglomerates of biomolecules immobilized on the probe and mica surface.
Keywords: atomic force microscopy, binding force, lipopolysaccharide, monoclonal antibodies
БИОФИЗИКА том 67
№ 6
2022