БИОФИЗИКА, 2022, том 67, № 6, с. 1068-1078
МОЛЕКУЛЯРНАЯ БИОФИЗИКА
УДК 577.3
ОБЗОР МАТЕМАТИЧЕСКИХ МОДЕЛЕЙ, ОПИСЫВАЮЩИХ
МЕХАНИЧЕСКИЕ ДВИЖЕНИЯ В МОЛЕКУЛЕ ДНК
© 2022 г. М.В. Беженар*, **, #, А.А. Елкина*, **, Х.Л. Эрнандес Касерес***, М.Г. Барышев*, **,
А.О. Сулима**, С.С. Джимак*, **, В.А. Исаев**
*ФИЦ «Южный научный центр РАН», 344006, Ростов-на-Дону, пр. Чехова, 41
**Кубанский государственный университет, 350040, Краснодар, ул. Ставропольская, 149
***Кубинский Центр Нейронаук ул. 25-ая, угол к 198-oй, Плайя, Гавана, Куба
#E-mail: mia1610@yandex.ru
Поступила в редакцию 01.07.2022 г.
После доработки 15.08.2022 г.
Принята к публикации 19.08.2022 г.
Рассмотрены различные механические модели ДНК, воспроизводящие ее структуру. Более подроб-
но описана эволюция механической модели молекулы ДНК, предложенной С.В. Ингландером, ко-
торый предположил, что нелинейные эффекты могут играть важную роль в динамике ДНК. Обсуж-
дается механическая модель молекулы ДНК, учитывающая вращательное движение азотистых ос-
нований вокруг сахаро-фосфатной цепи, а также влияние внешнего периодического воздействия на
динамику молекулы. Показано, что при численном решении модели без упрощений при периоди-
ческом внешнем воздействии молекула ДНК начинает колебаться, причем специфика колебаний
молекулы зависит от последовательности нуклеотидов в цепи. Важно, что с помощью данной мате-
матической модели можно рассчитать частоту и амплитуду колебаний отдельного участка ДНК.
Расчеты показывают, что зоны открытых состояний чаще появляются на границах гена и в областях
с преобладанием А-Т-пар оснований.
Ключевые слова: механическая модель молекулы ДНК, энергия связи, водородные связи, стэкинг,
торсионный момент, открытые состояния.
DOI: 10.31857/S0006302922060035, EDN: LJAVZJ
Экспериментальное исследование динамики
верситетов к исследованию динамики ДНК и от-
ДНК и возникновения открытых состояний огра-
крытых состояний, с другой стороны
- о
ничено пространственным разрешением доступ-
необходимости проведения подобных исследова-
ных биофизических инструментов [1]. По этой
ний именно с помощью математического моде-
причине основным методом исследования от-
лирования, позволяющего получить гораздо
крытых состояний ДНК является математиче-
больше информации, отражающей особенности
ское моделирование [2]. Несмотря на большое
динамики ДНК. Отмечается, что механические
количество упрощений, методы математического
деформации ДНК широко распространены и во-
моделирования позволяют с большой точностью
влечены в различные жизненно важные клеточ-
учесть различные аспекты поведения и функцио-
ные процессы. Кроме того, авторы работы [3] от-
нирования молекулы ДНК.
метили, что отсутствие высокопроизводительных
В работе [1] описана попытка эксперимен-
инструментов для измерения механических
тального наблюдения за динамикой открытых со-
свойств ДНК ограничивает изучение ее механи-
стояний в реальном времени. Эксперимент про-
ки. По этой причине особо важным методом ис-
водили на коротких кольцевых молекулах ДНК,
следования молекулярной динамики ДНК явля-
при этом делали большое количество микрофото-
ется математическое моделирование на основе
графий на электронном микроскопе, а затем с по-
механических моделей ДНК.
мощью суперкомпьютера из этих фотографий со-
Механические модели играют важную роль
здавали короткие видео. В результате удалось на-
для теоретического исследования внутренней по-
глядно зафиксировать возможные места
движности молекулы ДНК. В этом случае основа-
возникновения открытых состояний в точках пе-
ния ДНК представляют в виде маятников, а взаи-
региба молекулы ДНК. С одной стороны, это го-
модействия между основаниями описывают с по-
ворит о большом интересе ведущих мировых уни-
мощью замены водородных связей на упругие
1068
ОБЗОР МАТЕМАТИЧЕСКИХ МОДЕЛЕЙ
1069
связи. Такой подход был предложен в работе [4],
последующие работы позволили развить механи-
ческую модель описания динамики ДНК, учесть
разнообразные формы внешнего воздействия,
различные эффекты диссипации и неоднородно-
сти молекулы [5].
Остановимся подробнее на рассмотрении раз-
личных механических моделей ДНК.
Составными частями каждого нуклеотида яв-
ляются сахар, фосфат и основание, которые в
Рис. 1. Механический аналог двойной спирали, пред-
механических моделях представляются в виде
ставляющий собой линейные цепочки маятников (ос-
упругих нитей и маятников (дисков). Рассмотрим
нований), соединенных пружинами (сахаро-фосфат-
взаимодействие между двумя соседними нуклео-
ными стержнями).
тидами из одной цепи, эти связи являются силь-
ными ковалентными связями, и они моделиру-
ются гармоническими потенциалами. Так, на-
тельности же от типа пары, как было показано
пример, в работе [6] исследованы механические
выше, зависит энергия связи между ними.
взаимодействия между соседними нуклеотидами.
В этой же работе было показано, что последова-
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ ЯКУШЕВИЧ
тельность азотистых оснований коренным обра-
зом влияет на процесс структурообразования
Важное предположение впервые в 1980 г. сде-
нуклеиновых кислот, кроме того, отмечено влия-
лали авторы работы [4]: нелинейные эффекты
ние взаимодействия с растворителем на стабиль-
могут играть важную роль в динамике ДНК.
ность последовательности.
Принципиальная возможность образования от-
крытых состояний в ДНК была показана методом
Между нуклеотидами, которые принадлежат к
водородно-тритиевого обмена. Согласно модели,
разным цепям, существуют слабые водородные
ДНК может быть представлена в виде двух линей-
связи. В некоторых работах сделано предположе-
ных цепочек маятников (сахаро-фосфатные ос-
ние, что силы притяжения и отталкивания в дан-
нования), соединенных пружинами (взаимодей-
ной системе почти равны, и соответствующую
ствия между основаниями), как показано на
потенциальную энергию можно описать потен-
рис. 1.
циалом Морзе. Минус такого подхода состоит в
том, что парные потенциалы не зависят от окру-
Полная энергия описанной системы в зависи-
жения, то есть нуклеотиды в такой последова-
мости от углов поворота θn задается следующим
тельности не отличаются друг от друга, в действи-
гамильтонианом:
2
2
dθ
mh
n
S
2
H
=
+
(θ
−
θ
)
+
mgh(1-
cosθ )
,
(1)
n
n−1
n
2
dt
2
где dθn - угол поворота n-го маятника, S - коэф-
Если жесткость S >> mgh, то ширина солитона
фициент крутильной жесткости маятника, m и
равна множеству постоянных решетки. Провора-
h - масса и длина маятников, g - гравитационная
чивание только одного из маятников через угол
постоянная.
2π в пределах одной постоянной решетки потре-
Поэтому дифференциальное уравнение дви-
бовало бы значительной энергии пружины; та-
жения n-го маятника имеет следующий вид:
ким образом, солитон распространяется на тол-
щину L. Для большей толщины затраты потенци-
2
2
d
θ
n
альной энергии слишком велики, что приводит к
mh
=
S(θ
n
−
2θ
+
θ
n-1
)
+
mghsin θ .
n
(2)
2
сохранению солитона неповрежденным.
dt
Решением уравнения (2) является выражение:
Модель, разработанная авторами работы [4],
была улучшена сначала в работах [7, 8], а затем
2nl
S
θ
=
4arctan
exp
,
L
=
2l
,
(3)
n
(
)
уже усовершенствована авторами работы [9], ко-
L
mgh
торые, рассмотрев дискретную модель плоских
где l - расстояние между двумя соседними маят-
ротаторов, показали, что помимо солитонов, ко-
никами. Функция θn ? θ(n) является солитоном
торые описывают повороты оснований, суще-
излома.
ствуют достаточно большие отклонения углов по-
Данное решение интерпретируем как движе-
ворота между соседними парами оснований. Из
ние открытого состояния вдоль цепочки ДНК. этого наблюдения был сделан вывод об обяза-
БИОФИЗИКА том 67
№ 6
2022
1070
БЕЖЕНАР и др.
Согласно данной модели, ДНК представляет со-
бой две цепочки дисков, которые связаны между
собой продольными и поперечными пружинка-
ми. При этом основания ДНК - диски, взаимо-
действие между основаниями - пружинки. Мо-
дель учитывает водородные связи между парами
оснований, а также стэкинг между соседними па-
рами оснований; недостаток данной модели со-
Рис. 2. Схематическое представление модели Л.В. Яку-
стоит в том, что не учитываются колебания осно-
шевич.
ваний в крайних парах (рис. 2).
В своей работе Л.В. Якушевич рассматривала
тельном учете дискретных эффектов при изуче-
фрагмент молекулы ДНК, состоящий из трех пар
нии динамики ДНК.
оснований, и полагала, что происходит только
Биомеханический подход также в своих рабо-
колебание центральной n-й пары оснований, а
тах использовала Л.В. Якушевич. Ею в 1989 г. бы-
пары оснований по краям ((n - 1) и (n + 1)) за-
ла предложена одна из лучших теоретических мо-
фиксированы, причем колебание происходит
делей ДНК, рассматривающая открытое состоя-
только в горизонтальной плоскости. Функция
ние как результат вращения пар оснований [10].
Лагранжа для такой системы имеет вид:
2
2
dφ
dφ
1
1
2
L
single pair
=
I
1
+
I
2
-
K
1-2
[r
1
(r
1
+
r
2
+
a)(1−
cosφ
1
)
+
2
dt
dt
(4)
2
a
+
r
2
(r
1
+
r
2
+
a
)(1 −
cosφ
2
)
-
1r2
1-
cos
(φ
1
+
φ
2
))
+
,
2
где φ1(t) и φ2(t) - углы отклонения 1-го и 2-го ма-
ков, a - расстояние между массами маятников в
положении равновесия (φ1 = φ2).
ятников, I1 и I2 - моменты инерции этих маятни-
ков, K1-2 — жесткость горизонтальной пружины,
Уравнения, соответствующие данной функ-
соединяющей маятники, r1 и r2 — длины маятни-
ции, имеют следующий вид:
2
dφ
(-)
(+)
d
φ
1
1
I
=-
U
sin φ
+
U
sin
(φ
+
φ
)
−
S
+
S
φ
−β
,
1
1
1
1−2
1
2
(
1
1
)
1
1
dt
dt
(5)
2
dφ
2
(-)
(+)
d
φ
2
I
2
=-
U
2
sin φ
2
+
U
1−2
sin
(φ
1
+
φ
2
)
−
(
S
2
+
S
2
)
φ
2
−β
2
,
dt
dt
где β1 и β2 - коэффициенты диссипации.
При развитии данной модели Л.В. Якушевич вве-
ла дополнительные члены в уравнения и получила [5]:
2
d
φ
1,n
t)
'
I
−
K
[
φ
t)
−
2φ
t)
+
φ
(t)]
−
k
R
(
R
+
R
)
sinφ
−
1,n
2
1,n
n+1,1
n,1
n-1,1
1
-2,n
1,n
1,
n
2,n
n,1
dt
d
φ
n,1
t)
−
k
1
−2,n
R
1,
n
R
2,n
sin(φ
n,1
−
φ
n
,2
)
=-β
1,n
+
F
0
cos(
Ω
t),
dt
(6)
2
d
φ
2,n
t)
'
I
2,n
−
K
2,n
[
φ
n+
1,2
t)
−
2φ
n,2
t)
+
φ
n-1,2
(t)]
−
k
1
-2,n
R
2,
n
(R
1,n
+
R
2,n
)
sin φ
n,2
−
2
dt
d
φ
t)
n,2
-
k
1
-
2,n
R
1,
n
R
2,n
sin(φ
n,2
−
φ
n
,1
)
= -β
2,n
+
F
0
cos(
Ω
t)
,
dt
где φi,n(t) - угловое отклонение n-го азотистого
K’i,n - константа, характеризующая крутильную
основания i-й цепочки; Ii,n - момент инерции n-
жесткость сахаро-фосфатной цепочки; k1-2,n -
го азотистого основания i-й цепочки; Ri,n - рас-
константа, характеризующая взаимодействие
стояние от центра масс n-го азотистого основа-
между основаниями внутри пар; F0 и Ω - соот-
ния i-й цепочки до сахаро-фосфатной цепочки;
ветственно амплитуда и частота внешнего перио-
БИОФИЗИКА том 67
№ 6
2022
ОБЗОР МАТЕМАТИЧЕСКИХ МОДЕЛЕЙ
1071
дического поля; n - номер основания в последо-
усреднены коэффициенты I1,n, I2,n, K’1,n, K’2,n,
вательностях S и Sk.
R1,n, R2,n и k1-2,n методом концентраций. Были
При решении этих уравнений был введен ряд
получены следующие уравнения:
упрощений и усреднений. В частности, были
2
d
φ
1,n
t)
'
I
1
−
K
1
[
φ
n+1,1
t)
−
2φ
n,1
t)
+
φ
n-1,1
(t)]−
k
1-2
R
1
(
R
1
+
R
2
)
sin φ
n,1
−
2
dt
d
φ
t)
n,1
−
k
1−2
R
1
R
2
sin(φ
n,1
−
φ
n,2
)
=-
β
1
+
F
0
cos
(Ωt)
,
dt
(7)
2
d
φ
1,
n
t)
'
I
2
−
K
2
[
φ
n
+
1,2
t)
−
2φ
n
,2
t)
+
φ
n-
1,2
(t)]−
k
1-2
R
12
(
R
1
+
R
2
)
sin φ
n,2
−
2
dt
d
φ
t)
n,2
-
k
R
R
sin(φ
−
φ
)
=-
β
+
F
cos(Ωt).
1-2
1
2
n,2
n,1
2
0
dt
Как показано в нашей работе [11], усреднение
ную связь между n-м диском на одной цепи и
коэффициентов уравнений приводит к уменьше-
(n + h)-м диском на другой, где 2h - длина витка
нию в 100 раз величины амплитуды угловых коле-
спирали в единицах основания [12, 13].
баний азотистых оснований.
Введение в гамильтониан члена, который от-
Л.В. Якушевич получила аналитическое реше-
вечает за «геликоидальное» взаимодействие, с од-
ние данной модели при усреднении коэффици-
ной стороны учитывает спиральную структуру, а
ентов уравнений. Было получено аналитическое
с другой - способно качественнее описать дина-
решение в виде кинка:
мику молекулы ДНК. В дальнейшем модель Яку-
шевич претерпевала несколько изменений, в
1,Ω
φ t)=
A
1,Ω
cos(Ωt
+
φ
01
),
(8)
частности, стало учитываться влияние внешней
f
среды за счет появлений коэффициентов дисси-
01
где
A
1,Ω
=
- амплитуда,
2
пации. В работе [14] Л.В. Якушевич с соавторами
2
2
2
(
ω
01
-
Ω
)
+
(
δ
1
Ω
)
показали, что в рассматриваемой асимметричной
модели двойной цепи ДНК могут существовать
2δ
Ω
три типа топологических солитонов, имитирую-
1
φ
=
arctan-
- фаза.
01
2
щих локализованные состояния с открытыми па-
ω2-Ω
(
01
)
рами оснований. Солитоны могут перемещаться
Однако данное решение не учитывает спи-
вдоль макромолекулы с постоянной скоростью,
ральный характер структуры ДНК и взаимодей-
меньшей скорости звука, а также они устойчивы
ствие угловых смещений азотистых оснований.
по отношению к тепловым колебаниям.
Кроме того, решение типа кинка может быть по-
Результаты численных расчетов показали, что
лучено только в случае однородной синтетиче-
данная модель солитонов может объяснять струк-
ской ДНК.
туру молекулы ДНК. В работе [11] было показано,
На основе модели Л.В. Якушевич было пред-
что необходимо учитывать колебания двух нитей
ложено учесть спиральную структуру ДНК, до-
ДНК, это позволяет описать динамику открытых
бавляя в уравнение член, учитывающий торсион-
состояний молекулы ДНК:
2
i
i
d
ϕ
1
t)
i
i−1
i
i+1
I
=
K
ϕ
t)
−2ϕ
t)
+ϕ
t)-
1
2
1
1
1
1
dt
(9)
i
i
i
i
i
i
i
i
i
i
i
−
k
R
R
+
R
sinϕ
−
k
R
R
sin(
ϕ
−ϕ
)
+
F
(
t
)
,
i
=
2,
n
−
1,
12
1
(
1
2
)
1
12
1
2
1
2
1
2
1
d
ϕ
1
1 t)
1
2
1
1
1
1
1
1
1
1
1
1
I
=
K
ϕ
t)
+ϕ
t)-
k
R
(
R
+
R
)
sinϕ1 -
k
R
R
sin(ϕ1 -ϕ
)
+
F t),
1
2
1
1
1
12
1
1
2
1
12
1
2
1
2
1
(10)
dt
2
n
n
d
ϕ
1
t)
n
n−
1
n
I
=
K
ϕ
t)
−ϕ
t)
-
1
2
1
1
1
dt
(11)
n
n
n
n
n
n
n
n
n
−k
R
R
+
R
sin
ϕn -
k
R
R
sin(ϕn -ϕ
)
+
F t)
,
12
1
(
1
2
)
1
12
1
2
1
2
1
БИОФИЗИКА том 67
№ 6
2022
1072
БЕЖЕНАР и др.
2
i
i
d
ϕ
2
t)
i
i-1
i
i+1
I
=
K
ϕ
t)
−2ϕ
t)
+ϕ
t)-
2
2
2
2
2
2
dt
(12)
i
i
i
i
i
i
i
i
i
i
i
−k
12
R
1
(
R
1
+
R
2
)
sinϕ
2
−
k
12
R
1
R
2
sin(ϕ
1
−ϕ
2
)
+
F
2
(t),
i
=
2,n
−
1,
2
1
1
d
ϕ
2
t)
1
2
1
1
1
1
1
1
1
1
1
1
1
1
I
=
K
ϕ
t)
+ϕ
t)-
k
R
R
+
R
sinϕ
−
k
R
R
sin(ϕ
−ϕ
)
+
F t),
(13)
2
2
2
2
2
12
2
(
1
2
)
2
12
1
2
2
1
2
dt
2
n
d
ϕ
2
n t)
n
n−1
n
n
n
n
n
n
n
n
n
n
I
=
K
ϕ
t)
+ϕ
t)-
k
R
R
+
R
sin
ϕn -
k
R
R
sin(ϕn -ϕ
)
+
F t),
(14)
2
2
2
2
2
12
2
(
1
2
)
2
12
1
2
2
1
2
dt
i
оснований уменьшается, что приводит к их ста-
i
i
ϕ
d t)j
где
F t)
= -β
+
F
cos(Ωt
)
билизации; в свою очередь, уменьшение вязкости
j
j
0
dt
среды приводит к обратному эффекту - увеличе-
Как оговаривалось выше, данный член отвечает
нию скорости угловых отклонений азотистых ос-
за эффекты диссипации и периодическое воздей-
нований и увеличению угловых деформаций ни-
ствие, причем F0 = 0.526⋅10-22 Дж, ω = 0.4⋅1012 c-1.
тей ДНК. Таким образом, уменьшение вязкости
среды вызывает нестабильность молекулы ДНК,
В работе [11] было показано, что ряд упроще-
которая со временем только увеличится [18].
ний (усреднение характеристик цепочки ДНК)
При воздействии торсионного момента на мо-
приводит к снижению достоверности решения.
лекулу ДНК возникают вращательные движения
Вышеописанный метод намного ближе к истин-
азотистых оснований, что приводит к возникно-
ной биомеханике молекулы ДНК.
вению открытых состояний, которые чаще всего
На основе модели Л.В. Якушевич появилось
образуются на краях молекулы ДНК и в областях
множество других моделей, особенностью кото-
с преобладанием азотистых пар оснований A-T
рых является использование торсионного момен-
[19]. Вероятность появления открытых состояний
та для описания открытых состояний. Такие мо-
при ситуации, когда торсионный момент прикла-
дели в последствии получили название Y-модели.
дывается к определенной части гена, зависит от
В работах [15, 16] было показано влияние изо-
содержания в этой области пар A-T, от размера
топного состояния среды: при изменении изо-
этой области и времени воздействия торсионного
момента на этот участок гена.
топного обмена 2H/1H наблюдалось увеличение
энергии, которое необходимо для разрыва водо-
Необходимо отметить, что данная модель поз-
родной связи, в связи с чем возрастало количе-
воляет учесть стэкинг оснований за счет коэффи-
ство открытых состояний. Представленное в ра-
циента, характеризующего крутильную жест-
боте [17] исследование показало, что наличие
кость сахаро-фосфатной цепочки. При этом, ес-
дейтерия в цепочке нуклеотидов может вызывать
ли знать значения стэкинга для всех комбинаций
в зависимости от величины энергии разрыва во-
пар оснований в цепи, он может быть учтен в аб-
дородной связи как увеличение, так и уменьше-
солютных значениях. Как отмечено в работе [20],
ние вероятности возникновения открытых состо-
при расчете величины стэкинга необходимо учи-
яний. Участие атомов дейтерия в образовании во-
тывать ряд параметров, в том числе, влияние вод-
дородных связей в молекуле ДНК может вызвать
ного окружения.
изменение во времени передачи генетической
информации, что говорит о том, что даже незна-
МОДЕЛЬ ПЕЙЯРДА-БИШОПА
чительное изменение изотопного состояния сре-
И ПЕЙЯРДА-БИШОПА-ДОКСУА
ды способно повлиять на изменение процессов
метаболизма в живых системах.
Модели, представленные выше, являются уг-
ловыми, такие модели предсказывают движение
В работе [17] было показано влияние частот в
солитонов по цепочке ДНК. Хорошо известным
диапазоне от 1011 с-1 до 108 с-1 внешнего перио-
примером трансляционных моделей является мо-
дического воздействия на динамику молекулы
дель Пейярда-Бишопа (PB) [21, 22], а также две
ДНК. Под воздействием внешней периодической
ее расширенные версии. Они представляют собой
силы молекула ДНК может совершать колеба-
геликоидальные модели Пейярда-Бишопа и мо-
тельные движения с частотой отличной от часто-
дель Пейярда-Бишопа-Доксуа. Основным тре-
ты внешнего воздействия.
бованием к трансляционному подходу является
Численное решения механической математи-
описание комплементарных Н-связей с помо-
ческой модели молекулы ДНК для гена интерфе-
щью нелинейного потенциала. Нелинейность
рона альфа 17 и фрагмента гена дрозофилы пока-
данного потенциала доказывается работами по
зало, что при увеличении вязкости среды ско-
инфракрасной спектроскопии ДНК, такие низ-
рость изменения угловых колебаний азотистых
кочастотные колебательные моды водородных
БИОФИЗИКА том 67
№ 6
2022
ОБЗОР МАТЕМАТИЧЕСКИХ МОДЕЛЕЙ
1073
связей оснований были названы
«дыханием»
ДНК. Модель Пейярда-Бишопа предполагает,
что молекула ДНК состоит из двух цепочек поли-
нуклеотидов и представляет собой две цепочки
дисков, которые связаны между собой продоль-
ными и поперечными пружинами. продольные
взаимодействия представляют собой сильные ко-
валентные связи, моделируемые гармонически-
ми потенциалами. Поперечные взаимодействия
между нуклеотидами, принадлежащими к раз-
ным цепям, являются слабыми водородными вза-
имодействиями, требующими ангармонического
потенциала. Примером, используемым для моде-
ли Пейярда-Бишопа, является потенциал Морзе:
2
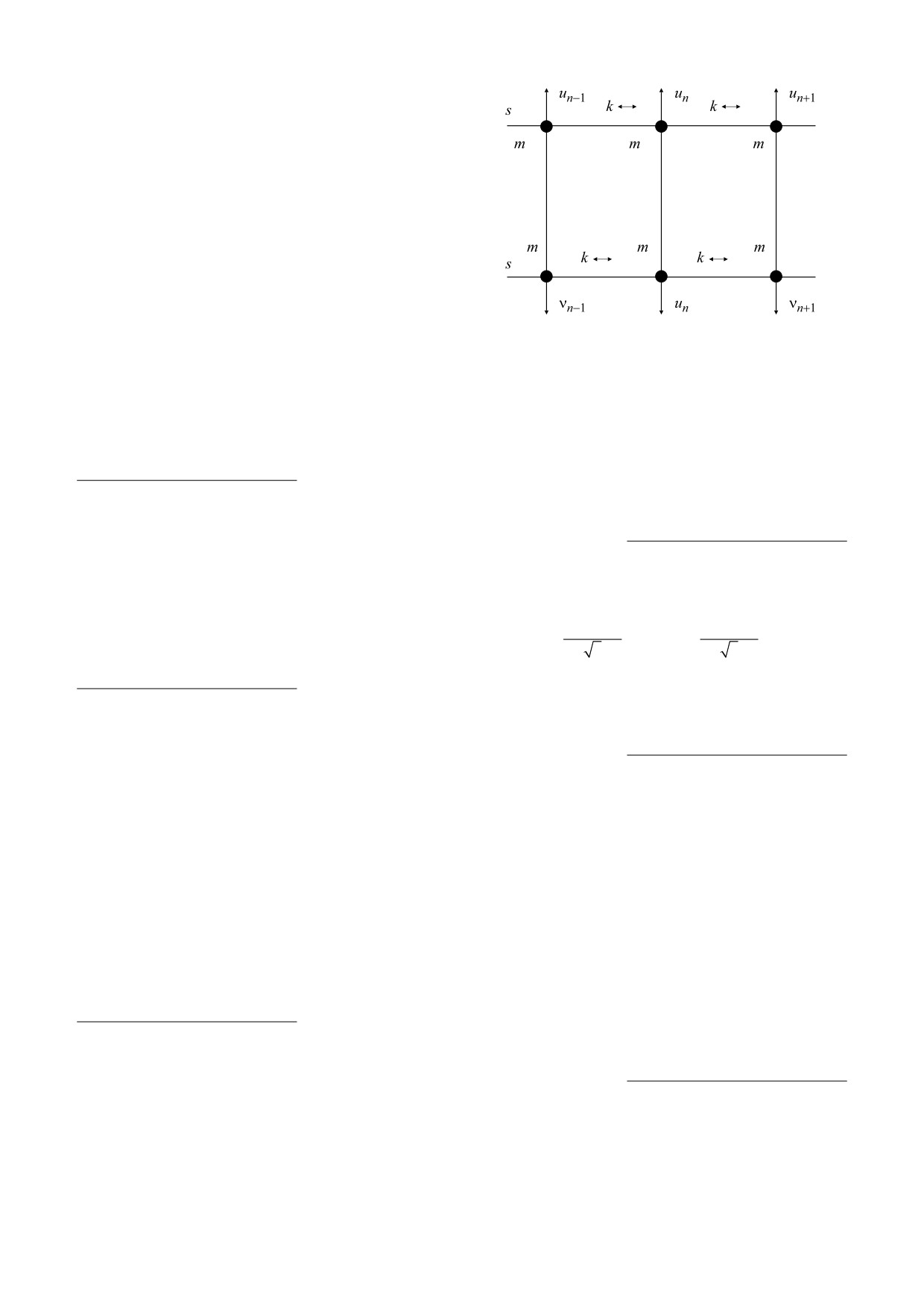
Рис. 3. Сегмент молекулы ДНК, показывающий как
- (un−v
n
)
V
(u
−
v
)
=
D
e
−1
,
(15)
M n
n
ковалентные взаимодействия, определяемые парамет-
ром k, так и водородные связи, моделируемые потенци-
где параметры D и a - соответственно глубина и
алом Морзе.
обратная ширина потенциальной ямы Морзе.
Координаты un и vn представляют собой попереч-
ные смещения нуклеотидов в положении n от их
родной связи. Гамильтониан такой системы ра-
положений равновесия вдоль направления водо-
вен
m
2
2
k
2
2
H
=
u
+
v
+
(u
−
u
)
+
(v
−
v
)
+V
(u
−
v
),
(16)
{
(
n
n
)
n
n−1
n
n-1
}
M n
n
2
2
где m = 300 а.е.м. = 5/1∙10-25 кг - масса нуклеоти-
ной системы и описывает слабые водородные
связи. Для удобства были введены новые коорди-
да, k - константа гармонического взаимодей-
наты, представляющие синфазные и внефазовые
ствия, в то время как u и v представляют соот-
n
n
поперечные движения:
ветствующие скорости. Первые два члена пред-
(u
n
+
v
n
)
(u
n
-
v
n
)
ставляют собой соответственно кинетическую и
x
n
=
,
y
n
=
,
(17)
потенциальные энергии продольной пружины.
2
2
Потенциал Морзе отвечает за нелинейность дан- которые преобразует гамильтониан (16) в (18):
m
2
2
k
2
2
H
=
x
+
y
+
x
−
x
+
y
−
y
+V
y
(18)
(
n
n
)
(
n
n−1
)
(
n
n-1
)
(M n
)
{
}
2
2
Согласно уравнениям (17) и рис. 3 можем за-
яснить денатурацию ДНК, а это означает, что ве-
ключить, что xn(t) и yn(t) описывают синфазные и
личина, которая может отражать степень денату-
внефазовые колебания пары нуклеотидов в поло-
рации молекул ДНК в растворе, представляет со-
жении n соответственно. Другими словами, xn(t)
бой среднее растяжение водородных связей [19].
Температура плавления, заданная численной об-
представляет колебание центра масс нуклеоти-
работкой, значительно выше, чем температура,
дов, в то время как yn(t) - их растяжение.
соответствующая непрерывному приближению.
Фактически модель Пейярда-Бишопа являет-
Это, безусловно, указывает на большую роль дис-
ся частным случаем геликоидальной модели Пей-
кретности в динамике ДНК [25].
ярда-Бишопа, которая будет объяснена ниже.
Рассмотрим усовершенствованные модели
Теперь достаточно указать, что функция xn(t)
Пейярда-Бишопа
- геликоидальные модели
представляет собой линейную волну, в то время
Пейярда-Бишопа и модель Пейярда-Бишопа-
как yn(t) является нелинейной. Важно знать, что
Доксуа. В модели Пейярда-Бишопа-Доксуа гар-
yn(t) зависит не от температуры, а от ее среднего
моническая потенциальная энергия была замене-
значения ?y? [23-25]. Данная модель может объ-
на ангармонической следующим образом:
k
2
k
-α(y
+
n n
y
−1
)
2
(y
−
y
)
1+ρe
(
y
−
y
)
,
(19)
n
n−1
n
n-1
2
2
где ρ и α - константы [26].
БИОФИЗИКА том 67
№ 6
2022
1074
БЕЖЕНАР и др.
Это выражение можно рассматривать как гар-
шопа) и α ≠ 0 (модель Пейярда-Бишопа-Док-
моническое взаимодействие с переменной кон-
суа) - и продемонстрировали преимущество по-
стантой связи [27].
следнего. Отметим, что функция
〈y〉 также
В рамках данной модели показано, что среднее
зависит от остальных параметров, существующих
растяжение водородных связей 〈y〉 зависит от
в модели, таких как D и a, описывающих потен-
температуры. Это демонстрирует преимущество
циал Морзе. Некоторые комбинации дают очень
модели Пейярда-Бишопа-Доксуа по сравнению
резкое увеличение даже для гармонического слу-
с теми моделями, которые предполагают гармо-
чая, а также слишком низкую температуру плав-
ления [28, 29].
ническую аппроксимацию. Функция 〈y〉 сначала
медленно увеличивается, а потом резко возраста-
Хронологически первое и наиболее важное
ет, данный участок графика представляет собой
усовершенствование модели Пейярда-Бишо-
денатурацию. В случае модели Пейярда-Бишо-
па было сделано в работе [30], его мы и называ-
па-Доксуа денатурация протекает довольно рез-
ем геликоидальной моделью Пейярда-Бишо-
ко и происходит при более низких температурах.
па. В уравнение (12) был введен новый термин,
Чтобы быть более точным, авторы сравнили два
описывающий геликоидальные взаимодей-
случая в пределах потенциала, заданного уравне-
ствия, и гамильтониан перепишем следующим
нием (19), - площадь α = 0 (модель Пейярда-Би-
образом:
m
2
2
k
2
2
H
={
u
+
v
+
(u
−
u
)
+
(v
−
v
)
+
(
n
n
)
n
n-1
n
n-1
2
2
(20)
2
K
2
2
−a(u
−
n n
v
)
+
(u
−
v
)
+
(u
−
v
)
+
D
e
−1
},
n
n+h
n
n-h
2
где K - гармоническая постоянная геликоидаль-
Полный математический анализ, включаю-
ной пружины. Как оговаривалось выше, геликои-
щий множество математических деталей, мож-
дальность означает, что после поворота на угол π
но найти в работе [32], очень кратко рассмот-
нуклеотид, принадлежащий одной цепи в пози-
рим основные выводы. Уравнения (17) и (20)
ции n, будет близок как к (n + h)-му, так и (n - h)-
приводят к двум несвязанным динамическим
му нуклеотидам другой цепи [31].
уравнениям движения - линейному и нели-
нейному. Ограничимся нелинейным, который
представляет собой выражение:
-a
2y
n
-a
2y
n
my
n
=
k(y
n+1
+
y
n-1
−
2
y
n
)
−
K
(y
n+h
+
y
n-h
−
2y
n
)
+
2 2aD(e
−1)e
(21)
Конечно, это дискретное дифференциальное
уравнение в частных производных, и наша цель -
y
n
n
= εΦ ( 1)
ε
(22)
получить его решение, т.е. функцию yn(t). Пред-
полагаем, что колебания нуклеотидов достаточно
Для решения уравнения (22) используем полу-
велики, чтобы быть ангармоническими, но все же
достаточно малы для преобразования, чтобы
дискретное приближение, что означает волновые
быть действительными:
решения вида:
iθ
n
i
2θ
n
2
Φ
t)
=
F
(ξ)e
+
ε
F
(ξ)
+
F
(ξ)e
+
сс
+
Ο
ε
(23)
n
1
0
2
(
)
и
действительной. Это модулированная волна, где
F1 является непрерывной функцией, представля-
ξ = (εnl t), θ =
nql
-ωt,
(24)
n
ющей огибающую, в то время как компонент не-
где l = 0.34 нм - расс тояние между двумя сосед-
ними нуклеотидами в одной цепи, ω - оптиче-
сущей eiθn является дискретным. Математиче-
ская частота линейного приближения; q = 2π/λ -
ской основой для уравнения (23) является метод
волновое число, сс представляет собой комплекс-
множественного масштаба или метод разложения
но сопряженные члены, а функция F0 является
по производным [33, 34].
БИОФИЗИКА том 67
№ 6
2022
ОБЗОР МАТЕМАТИЧЕСКИХ МОДЕЛЕЙ
1075
Приходим к дисперсионному соотношению:
2
iF
+
PF
+Q
F
F
=
0,
(26)
1τ
1SS
1
1
2
4
2
2
ql
2
qhl
где τ и S - новые временные и пространственные
ω
=
a
D
+
k
sin
+
Κ cos
,
(25)
( )
(
)
координаты. Здесь P и Q - коэффициент диспер-
m
2
2
сии и коэффициент нелинейности соответствен-
выражение для групповой скорости VS = dω/dq и
но. Для PQ > 0 решение уравнения (26) равно:
так далее. Особое значение имеет тот факт, что
функции F0 и F2 могут быть выражены через F1, в
S
-
u
τ
iu
(S
-
u
τ)
e
e
c
то время как последняя является решением хоро-
F
(S,τ)
=
A
sec
exp
,
(27)
1
0
L
2P
шо известного разрешимого нелинейного урав-
e
нения Шредингера:
что приводит к окончательному решению [34]:
nl
−
V
t
nl
-V
t
μ
e
e
y t)
=
2Asec
{cos(Θnl
−
Ωt)
+
A
sec
[
+δcos(2(Θnl
-
Ωt))]}.
(28)
n
L
L
2
Выражение для A ≡ εA0, L ≡ Le/ε, волновое чис-
ПРИМЕНЕНИЕ КВАНТОВО-
ло Θ огибающей скорости Ve и частота Ω приведе-
МЕХАНИЧЕСКИХ И МЕТОДОВ
САМОНАПРЯЖЕННЫХ КОНСТРУКЦИЙ
ны в [30].
ПРИ ИССЛЕДОВАНИИ СТРУКТУРЫ ДНК
Чтобы построить график функции yn(t), значе-
Квантово-механические методы часто приме-
ния всех параметров должны быть известны или,
няются для описания сложных биологических
по крайней мере, оценены. Их две группы: мате-
объектов. Эта необходимость может возникнуть
матические (ε, ue, uc) и внутренние параметры (k,
при объяснении поведения клеток и тканей, воз-
никающих в результате коллективных взаимо-
K, a, D, q). Было высказано предположение [34],
действий внутри сложных молекулярных сетей, а
что длина волны охватывает целое число перио-
также в вопросе переноса энергии и заряда в мак-
дов l:
ромолекулах. В отличие от моделей, представлен-
ных выше, данные процессы протекают на моле-
2π
q
=
,
λ
=
Nl, N integer.
(29)
кулярном уровне, поэтому на динамику молеку-
λ
лы ДНК могут влиять квантово-механические
эффекты. Данный подход был предложен в рабо-
Следовательно, можно принять N в качестве
те [35], автор которой описал солитонный меха-
внутреннего параметра вместо q. Обращаем вни-
низм переноса биоэнергии в макромолекуле. В
мание на то, что зависимость от ε может быть
работе [36] показано, что возникающее нелиней-
устранена [30]. Это можно было ожидать, по-
ное взаимодействие, сопровождающее распро-
скольку это только рабочий параметр, который
странение вибрационных квантов вдоль мак-
мы используем для различения больших и малых
ромолекулы, вызывается вибрационными воз-
терминов в расширении ряда (23).
буждениями (экситонами) и акустическим
фононами. Это приводит к формированию соли-
Проблема с математическими параметрами
тона, под которым подразумевают перемещаю-
практически решена с использованием идеи ко-
щуюся вдоль молекулярной цепи локальную об-
герентного режима [31, 34], предполагая, что ско-
ласть продольных сжатий водородных связей в
рости огибающей и несущей волны равны:
молекуле белка. В работе [37] описан солитон
кручения, который представляет собой динами-
чески устойчивую локальную область деформа-
Ve
=Ω.
(30)
Θ
ции торсионных углов макромолекулы. Рассмат-
ривалась цепь молекул, находящихся на опреде-
Это означает, что функция yn(t) одинакова в
ленном расстоянии, а в качестве потенциала -
любой позиции n. Другими словами, волна сохра-
торможение звена цепи, которое возникает при
няет свою форму во времени, что указывает на
перекрывании орбиталей при вращении вокруг
возможную стабильность [34]. Обращаем внима-
валентной связи. Одним из недостатков подоб-
ние на то, что уравнение (28) является однофаз-
ных моделей является то, что солитон обладает
ной функцией, если выполняется уравнение (30).
коротким временем жизни (~ 10-12-10-13 с) при
БИОФИЗИКА том 67
№ 6
2022
1076
БЕЖЕНАР и др.
температурах, близких к биологическим, что го-
ли упругого стержня и квантовые модели ДНК
ворит о сложности переноса им энергии в биоло-
[42-48]. Образование открытых состояний [49]
гических системах [38]. Помимо солитонов Давы-
можно объяснить на основании вращения азоти-
дова в макромолекулярных структурах могут воз-
стых оснований вокруг сахаро-фосфатного осто-
никать различные типы стабильных состояний,
ва (модель Л.В. Якушевич), либо на основании
которые зависят от геометрии структуры, а также
сдвига азотистых оснований в направлении, пер-
от ее физических свойств [39].
пендикулярном оси вращения молекулы ДНК
(модель Пейярда-Бишопа и другие). Динамика
Другим, не менее интересным методом, явля-
моделей ДНК описывается нелинейными урав-
ется концепция самонапряженных конструкций
нениями, которые допускают аналитическое ре-
или тенсегрити, суть данного метода заключается
в описании поведения биокомплексных систем.
шение в виде солитонов и кинков. Необходимо
отметить, что изучению физических свойств мо-
Данная модель клетки объясняет, как ее механи-
лекулы ДНК посвящено огромное количество
ческое поведение возникает в результате физиче-
публикаций. Так, хорошо изучена температурная
ских взаимодействий между различными систе-
зависимость гибкости ДНК в присутствии сил
мами молекулярных нитей, формирующих ци-
тоскелет [40].
растяжения и расстегивания [50]. Считается, что
двухцепочечная ДНК обладает внутренней жест-
В работе [41] рассматривается математическая
костью на изгиб [51-53]. Одним из важных фак-
модель, воспроизводящая регуляцию экспрессии
торов, влияющих на прочность молекулы явля-
генов на основе связывания лигандов с молеку-
ются термически генерируемые пузырьки денату-
лой ДНК. Макромолекула представлялась иде-
рации
[54]. При этом подобные пузырьки
альной струной для изучения механических воз-
возникают при считывании информации с моле-
мущений, динамика этих процессов описывается
кулы ДНК, поэтому изучение их свойств является
следующим уравнением:
крайне важной и актуальной задачей [55-59]. Од-
2
2
ним из факторов, влияющих на образование пу-
∂
U
F
∂
U
=
+
g(x,t),
(31)
зырьков денатурации является величина торси-
2
2
∂
t
ρ(x)∂
x
онного момента, действующего на двойную спи-
где U(x,t) представляет собой поперечное откло-
раль молекулы [60]. Измерению его величины
нение струны на расстоянии x от конца струны в
также посвящен ряд работ [61-63].
момент времени t, F - натяжение струны, ρ(x) -
Известно, что экспериментальное исследова-
линейная плотность, g(x,t) - удельная внешняя
ние динамики ДНК и возникновения открытых
сила, действующая на струну. Величина F/ρ(x) =
состояний ограничено пространственным разре-
= с2, где c - скорость распространения возмуще-
шением доступных биофизических инструментов
ния в ДНК. Начальное возмущение имеет вид:
[64, 65]. Более полное описание механики ДНК
2
2
можно получить, если объединить данные о ее
−
(
at
+(x−x
0
)
)
/b
g
(x,t)
=
Ae
,
(32)
механических показателях и исследовать ее дви-
жения. Такой подход возможен только в рамках
где A и b - численные параметры. Для определе-
математического моделирования [2]. Поэтому на
ния положения струны использовали разностную
сегодняшний день одним из основных методов
схему, а для оценки - электромеханическую мо-
исследования механических свойств ДНК явля-
дель отталкивания дипольных моментов.
ется математическое моделирование.
Данные методы на сегодняшний день вызыва-
На наш взгляд, наиболее универсальной моде-
ют большой интерес к исследованию свойств не-
лью для исследования динамики азотистых осно-
линейных возбуждений, в частности таких как
ваний, позволяющей учитывать различные внеш-
различные типы солитонов, поляронов или бипо-
ние факторы является модель Л.В. Якушевич без
ляронов, которые могут образовываться в биоло-
упрощений.
гических макромолекулярных структурах. Кон-
цепция самонапряженных конструкций, описан-
ная в работах [40, 41], совмещает различные
ФИНАНСИРОВАНИЕ РАБОТЫ
подходы к описанию динамики молекулы ДНК, а
также показывает, что помимо статистических
Работа выполнена при финансовой поддержке
методов может быть использована модель элек-
Кубанского научного фонда в рамках научного
тромеханических возмущений.
проекта № Н-21.1/11, гранта Президента Россий-
ской Федерации для государственной поддержки
молодых российских ученых - кандидатов наук
ОБСУЖДЕНИЕ И ВЫВОДЫ
(проект МК-2366.2022.1.4), Российского фонда
На сегодняшний день разработано множество
фундаментальных исследований (проект № 19-
различных нелинейных моделей ДНК, различа-
44-230026)
и государственного задания
ются они в основном своей подробностью: моде-
ЮНЦ РАН № 122020100351-9.
БИОФИЗИКА том 67
№ 6
2022
ОБЗОР МАТЕМАТИЧЕСКИХ МОДЕЛЕЙ
1077
КОНФЛИКТ ИНТЕРЕСОВ
24. A. Wildes, L. Khadeeva, W. Trewby, et al., J. Phys.
Chem. B, 119 (12), 4441 (2015).
Авторы заявляют об отсутствии конфликта
интересов.
25. G. Gaeta, C. Reiss, and M. Peyrard, La Rivista del
Nuovo Cimento, 17, 1 (1994).
26. G. Weber, Europhys. Lett., 73 (5), 806 (2006).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
27. M. Peyrard, S. Cuesta-López, and D. Angelov, J. Phys.
Настоящая статья не содержит каких-либо ис-
Condensed Matter, 21 (3), 34103 (2009).
следований с участием людей или животных в ка-
28. M. Barbi, S. Lepri, M. Peyrard, et al., Phys. Rev. E, 68,
честве объектов исследований.
61909 (2003).
29. T. Dauxois and M. Peyrard, Phys. Rev. E, 51, 4027
СПИСОК ЛИТЕРАТУРЫ
(1995).
30. T. Dauxois, Phys. Rev. A, 159, 390 (1991).
1.
A. L. B. Pyne, A. Noy, K. H. S. Main, et al., Nature
Commun., 12, 1053 (2021).
31. S. Zdravković, J. Nonlinear Math. Phys., 18 (2), 463
2.
M. Manghi and N. Destainville, Phys. Reports, 631, 1
(2011).
(2015).
32. 32. T. Kawahara, J. Phys. Soc. Japan, 35, 1537 (1973).
3.
A. Basu, D. G. Bobrovnikov, Z. Qureshi, et al., Nature,
33. R. K. Dodd, J. C. Eilbeck, J. D. Gibbon, and
589, 462 (2021).
H. C. Morris, Solitons and Nonlinear Wave Equations
4.
S. W. Englander, N. R. Kallenbach, A. J. Heeger, et al.
(Acad. Press, Lond., 1982).
Proc. Natl. Acad. Sci. USA, 77, 7222 (1980).
34. S. Zdravković and M. V. Satarić, Phys. Rev. E, 73,
5.
Л. В. Якушевич, Нелинейная физика ДНК (НИЦ
021905 (2006).
«Регулярная и хаотическая динамика», Ижевский
35. A. S. Davydov, Phys. Scripta, 20 (3-4), 387 (1979).
институт компьютерных исследований, Ижевск,
36. A. S. Davydov, Solitons in molecular systems (Riedel
2007).
Publ. Comp., D., 1985).
6.
F. Hase and M. Zacharias, Nucl. Acids Res., 44, 7100
37. А. В. Савин и О. В. Гендельман, Высокомолекуляр.
(2016).
соединения, 41 (2), 263 (1999).
7.
S. Yomosa, Phys. Rev. A, 27, 2120 (1983).
38. L. Cruzeiro-Hansson, Phys. Rev. A, 45, 4111 (1992).
8.
S. Yomosa, Phys. Rev. A, 30, 474 (1984).
39. A. A. Gogolin, V. I. Mel'Nikov, and É. I. Rashba, J.
9.
S. Homma and S. Takeno, Progr. Theor. Phys. 72, 679
Exp. Theor. Phys., 42, 254 (1975).
(1984).
40. D. E. Ingber, J. Cell Sci., 116, 1397 (2003).
10.
L. V. Yakushevich, Phys. Lett. A, 136, 413 (1989).
41. Ю. Д. Нечипуренко, Р. В. Полозов и Д. Ю. Нечи-
11.
M. I. Drobotenko, S. S. Dzhimak, A. A. Svidlov, et al.,
пуренко, в кн. Математика. Компьютер. Образо-
Biophysics, 63 (2), 177 (2018).
вание, под ред. Г. Ю. Ризниченко (М.-Ижевск,
12.
G. Gaeta, Phys. Lett. A, 143 (4-5), 227 (1990).
НИЦ «Регулярная и хаотическая динамика»,
13.
G. Gaeta, Phys. Lett. A, 168 (5-6), 383 (1992).
Ижевский институт компьютерных исследований,
2006), сс. 392-402.
14.
L. V. Yakushevich, A. V. Savin, and L. I. Manevitch,
Phys. Rev. E, 66 (1), 16614 (2002).
42. S. K. Nomidis, M. Caraglio, M. Laleman, et al., Phys.
Rev. E, 100 (2), 22402 (2019).
15.
A. A. Basov, M. I. Drobotenko, A. A. Svidlov, et al.,
Molecules, 25 (16), 3753 (2020).
43. S. K. Nomidis, E. Skoruppa, E. Carlon, et al., Phys.
Rev. E, 99 (3), 032414 (2019).
16.
A. A. Svidlov, M. I. Drobotenko, A. A. Basov, et al.,
Physics of Wave Phenomena, 29 (2),180 (2021).
44. S. K. Nomidis, J. Hooyberghs, G. Maglia et al., J. Phys.
17.
A. A. Svidlov, M. I. Drobotenko, A. A. Basov, et al.,
Condensed Matter, 30 (30), 304001 (2018).
Int. J. Mol. Sci., 22 (15),7873 (2021).
45. G. Torrellas and E. MacIá, Phys. Lett., Section A:
18.
A. A. Svidlov, M. I. Drobotenko, A. A. Basov, et al.,
General, Atomic and Solid State Physics 376 (45), 3407
Entropy, 23 (11), 1446 (2021).
(2012).
19.
S. Dzhimak, A. Svidlov, A. Elkina, et al., Int. J. Mol.
46. A. Zeida, M. R. MacHado, P. D. Dans, et al., Phys.
Sci., 23 (8), 4428 (2022).
Rev. E - Statistical, Nonlinear, and Soft Matter Phys-
ics, 86 (2), 21903 (2012).
20. F. Hase and M. Zacharias, Nucl. Acids Res., 44 (15),
7100 (2016).
47. A. K. Mazur and M. Maaloum, Phys. Rev. Lett., 112
(6), 68104 (2014).
21. M. Peyrard and A. R. Bishop, Phys. Rev. Lett., 62, 2755
(1989).
48. M. Zoli, J. Phys. Condensed Matter, 29 (22), 225101
(2017).
22. T. Dauxois, M. Peyrard, and A. R. Bishop, Phys. Rev.
E, 47, 684 (1993).
49. A. K. Mazur, Phys. Rev. E, 95 (6), 62417 (2017).
23. A. Wildes, N. Theodorakopoulos, J. Valle-Orero, et al.,
50. T. Pal and S. M. Bhattacharjee, Phys. Rev. E, 93 (5),
Phys. Rev. Lett., 106 (4), 48101 (2011).
52102 (2016).
БИОФИЗИКА том 67
№ 6
2022
1078
БЕЖЕНАР и др.
51. A. Noy and R. Golestanian, Phys. Rev. Lett., 109 (22),
59. S. Park, O.-C. Lee, X. Durang, et al., J. Korean Phys.
228101 (2012).
Soc., 78 (5), 408 (2021).
52. A. K. Mazur, Phys. Rev. Lett., 116 (15), 158101 (2016).
60. D. Salerno and A. Tempestini, I. Mai, Phys. Rev. Lett.,
53. A. K. Mazur, Phys. Rev. Lett., 111 (17), 179801 (2013).
109 (11), 118303 (2012).
54. G. Altan-Bonnet, A. Libchaber, and O. Krichevsky,
61. H. Qu, Y. Wang, C.-Y. Tseng, et al., Phys. Rev. X, 1 (2),
Phys. Rev. Lett., 90 (13), 138101 (2003).
1 (2011).
55. O.-C. Lee, J.-H. Jeon, and W. Sung, Phys. Rev. E -
Statistical, Nonlinear, and Soft Matter Physics, 81 (2),
62. D. S. Sanchez, H. Qu, D. Bulla, et al., Phys. Rev. E -
21906 (2010).
Statistical, Nonlinear, and Soft Matter Physics, 87 (2),
22710 (2013).
56. D.S. Sanchez, H. Qu, D. Bulla, et al., Phys. Rev. E -
Statistical, Nonlinear, and Soft Matter Physics, 87 (2),
63. J. Wang, H. Qu, and G. Zocchi, Phys. Rev. E - Statis-
22710 (2013).
tical, Nonlinear, and Soft Matter Physics, 88 (3), 32712
57. O.-C. Lee and W. Sung, Phys. Rev. E - Statistical,
(2013).
Nonlinear, and Soft Matter Physics, 85 (2), 021902
64. A. L. B. Pyne, A. Noy, K. H. S. Main, et al., Nat. Com-
(2012).
mun. 12, 1053 (2021).
58. B. S. Alexandrov, L. T. Wille, K. O. Rasmussen, et al.,
Phys. Rev. E - Statistical, Nonlinear, and Soft Matter
65. S. S. Dzhimak, M. I. Drobotenko, A. A. Basov, et al.,
Physics 74, 050901 (2006).
Dokl. Biochem. Bioph. 483, 359 (2018).
Review of Mathematical Models Describing the Mechanical Motion
in a DNA Molecule
M.V. Bezhenar*, **, A.A. Elkina*, **, J.L. Hernandez Caceres***, M.G. Baryshev*, **,
А.O. Sulima**, S.S. Dzhimak*, **, and V.A. Isaev**
*Southern Scientific Center of the Russian Academy of Sciences, prosp. Chekhova 41, Rostov-on-Don, 344006 Russia
**Kuban State University, Stavropolskaya ul. 149, Krasnodar, 350040 Russia
***Cuban Center for Neurosciences Avenida 25 esquina a 198, Playa, La Habana, Cuba
Various mechanical models of DNA reproducing its structure are considered. The evolution of the mechan-
ical model of the DNA molecule proposed by S.V. Englander, who suggested that nonlinear effects can play
an important role in DNA dynamics, is described in more detail. This paper discusses a mechanical model of
a DNA molecule that takes into account the rotational motion of nitrogenous bases around the sugar-phos-
phate chain, as well as the influence of external periodic action on the dynamics of the molecule. It is shown
that when the model is numerically solved without simplifications, the DNA molecule begins to oscillate un-
der periodic external influence, and the specificity of the molecule’s oscillations depends on the sequence of
nucleotides in the chain. It is important to note that with the help of this mathematical model, it is possible
to calculate the frequency and amplitude of vibrations of a single DNA site. Calculations show that zones of
open states appear more often at gene boundaries and in regions with a predominance of A-T base pairs.
Keywords: mechanical model of a DNA molecule, binding energy, hydrogen bonds, stacking, torsion moment, open
states
БИОФИЗИКА том 67
№ 6
2022