БИОФИЗИКА, 2022, том 67, № 6, с. 1143-1157
БИОФИЗИКА КЛЕТКИ
УДК 575.35+501+579+54.027
ПРИМЕНЕНИЕ БАЗОВЫХ ИЗОТОПНЫХ УРАВНЕНИЙ ДЛЯ ОПИСАНИЯ
ДИНАМИКИ МИКРОБИОЛОГИЧЕСКИХ ПРОЦЕССОВ:
ПЕРЕРАСПРЕДЕЛЕНИЕ ДЕЙТЕРИЯ
© 2022 г. В.А. Вавилин*, #, Л.Я. Локшина*
*Институт водных проблем РАН, ул. Губкина, 3, Москва, 119333, Россия
#E-mail: vavilin@iwp.ru
Поступила в редакцию 15.02.2022 г.
После доработки 10.06.2022 г.
Принята к публикации 24.06.2022 г.
С использованием базовых изотопных уравнений, основанных на сохранении баланса дейтерия,
анализируется динамика внутриклеточного окисления метана нитрит-ионом (NDAMO) в двух ла-
бораторных экспериментах с различными начальными концентрациями субстратов и с привлече-
нием изотопных переменных с дейтерием, причем во втором эксперименте начальное количество
воды в закрытом сосуде было уменьшено приблизительно в 2.8 раза. Согласно модели в конце пер-
вого эксперимента метан полностью расходуется при остающемся избытке нитрит-иона, тогда как
в конце второго эксперимента метан остается при полном исчерпании нитрит-иона. В качестве изо-
топных переменных в модели рассматриваются концентрации содержащих единственный атом
дейтерия субстратов (СН4 и NH4+), продукта (Н2О) и биомассы метанотрофных микроорганизмов
ANME (C5H7NO2). Выведены стехиометрические уравнения реакций, описывающие перераспре-
деление дейтерия между субстратами, продуктом реакции (водой) и биомассой. Показано, что
фракционирование изотопов идет в процессе микробиологической реакции в воде, при этом суб-
страты обогащаются дейтерием, а вода и биомасса им обедняются. Если в первом эксперименте, со-
провождающемся небольшим падением содержания дейтерия в воде и биомассе, процесс фракци-
онирования заканчивается при t ≥ 17 ч, то во втором эксперименте он заканчивается при t ≥ 140 ч и
сопровождается значительным падением содержания дейтерия в воде, зависящем от начальной
концентрации воды, содержащей дейтерий. Это происходит из-за разбавления воды в сосуде обед-
ненной дейтерием водой, образующейся в процессе NDAMO. В работе также дана сводка получен-
ных нами в течение ряда лет результатов моделирования динамики одиннадцати биологических
процессов, в которых измеряли в основном стабильные изотопы углерода. Приводятся используе-
мые при моделировании факторы фракционирования изотопов.
Ключевые слова: динамика экологических процессов, кинетический изотопный эффект, базовые
изотопные уравнения, последовательные и параллельные реакции.
DOI: 10.31857/S0006302922060114, EDN: LKFKYL
Кинетический изотопный эффект заключает-
стандарте (hE/lE)ст, который отличается высоким
ся в изменении скорости химических, биологиче-
содержанием более тяжелого изотопа hE [2]:
ских и физических процессов при замене в моле-
куле реагирующего вещества какого-либо атома
h
l
его изотопом [1]. Величина отношения тяжелых
(
E
E
)
h
o
обр
δ
E
=
−1
×1000,
(1)
(hE) и легких (lE) стабильных изотопов R = hE/lE
(
oo
)
h
l
(
E
E
)
в веществе обычно мала, Она традиционно опре-
ст
деляется при помощи разницы отношений hE/lE
где величина δhE называется изотопной сигнату-
в исследуемом образце (hE/lE)обр и определенном
рой (signature). Концентрация субстрата (реаген-
та) с более тяжелым (редким) изотопом, как пра-
Сокращения: NDAMO - реакция окисления метана нитри-
вило, много меньше концентрации субстрата с
том, ANME - анаэробные метан-окисляющие микроорга-
низмы.
более легким изотопом.
1143
1144
ВАВИЛИН, ЛОКШИНА
Уравнение Релея [3], изначально предложен-
линейной химической кинетики n-го порядка по
ное для описания диффузии газов, традиционно
концентрации субстрата S [5], однако, в совре-
применяется и для вычисления фактора фракци-
менной научной литературе по-прежнему часто
онирования (разделения) стабильных изотопов:
считается, что справедлива линейная кинетика
по концентрации субстрата S [6]. При этом кине-
Rt/R0 = (St/S0)1/(α-1),
(2)
тический коэффициент слегка меньше у субстра-
где R0 и Rt - отношение тяжелого и легкого ста-
та с более тяжелым изотопом. Фактор фракцио-
нирования α обычно считается величиной посто-
бильных изотопов в субстрате в моменты времени
янной, что не всегда справедливо [7]. Фактически
0 и t соответственно; S0 и St - начальная и теку-
уравнение Релея (2) является статическим, по-
щая концентрации субстрата; α - фактор фрак-
скольку оно фиксирует концентрацию субстрата
ционирования стабильных изотопов при превра-
только в моменты времени 0 и t и не зависит от
щении субстрата в продукт. Величина α опреде-
динамики концентрации субстрата в промежутке
ляется [4] по наклону прямой линии, полученной
от 0 до t.
в двойных логарифмических координатах, следу-
ющих из уравнения Релея (2).
Базовое изотопное динамическое уравнение
При описании изотопного эффекта в уравне-
для единственного субстрата записывается в сле-
нии (2) уже давно отмечается справедливость не- дующем виде [8]:
h
dhS
1
S dS
=
,
(3)
dt
α
S dt
(3) для субстрата нужно дополнить соответствую-
где hS и S = lS + hS ≈ lS - концентрации субстрата,
щими уравнениями для продукта реакции (4) и
содержащего более тяжелый изотоп, и полного
биомассы (5), осуществляющей саму микробио-
субстрата, содержащего более легкий и тяжелый
логическую реакцию:
изотоп соответственно; dS/dt - скорость измене-
ния концентрации полного субстрата. Уравнение
h
dhP
1
S dP
=
,
(4)
dt
α
S dt
h
dhB
1
S dB
=
,
(5)
dt
α
S dt
диционно функция Моно используется при опи-
где продукт (hP) и биомасса (hB) содержат более
сании динамики анаэробных микробиологиче-
тяжелый изотоп. При этом учитывается, что в
ских процессов
[11,
12]. В монографии
[13]
продукт и биомассу более тяжелый изотоп посту-
обсуждается переход от функции Михаэлиса-
пает из субстрата. Из уравнений (3)-(5) следует
Ментен, известной в ферментативной кинетике,
сохранение суммарного баланса более тяжелого
к функции Моно, известной в микробиологиче-
изотопа в субстрате, продукте и биомассе
ской динамике.
(dhS/dt + dhP/dt + dhB/dt = 0), если материальный
В предложенных нами динамических уравнени-
баланс в системе без изотопов сохраняется
ях (3) - (5) вводится концентрация более тяжелого
(dS/dt + dP/dt + dB/dt = 0). В работе [8] изотопные
(редкого) элемента в субстрате, продукте и биомас-
уравнения для субстрата, продукта и биомассы
типа уравнений (3)-(5) используются как состав-
се. Из уравнения (3) следует, что скорость измене-
ния концентрации субстрата, содержащего более
ные части двух последовательных реакций (пре-
тяжелый изотоп, пропорциональна скорости изме-
вращения нитрата в нитрит и последующее пре-
нения концентрации полного субстрата, умножен-
вращение нитрита в молекулярный азот).
ной на текущую пропорцию более тяжелого изотопа
В работах [9, 10] показано, что статическое
в субстрате к полному субстрату и деленной на фак-
уравнение Релея (2) выводится из основного изо-
тор фракционирования изотопов α, который отра-
топного уравнения (3), где нелинейная скорость
жает тот факт, что константа скорости для субстрата
процесса dS/dt для скорости микробиологиче-
с более тяжелым изотопом несколько меньше, чем
ской реакции задается в общем виде. Часто для
та же константа скорости для субстрата с легким
описания скорости биологического процесса ис-
изотопом. При этом концентрация субстрата, со-
пользуется нелинейная функция типа Моно. Тра-
держащего более тяжелый (редкий) изотоп, во мно-
БИОФИЗИКА том 67
№ 6
2022
ПРИМЕНЕНИЕ БАЗОВЫХ ИЗОТОПНЫХ УРАВНЕНИЙ
1145
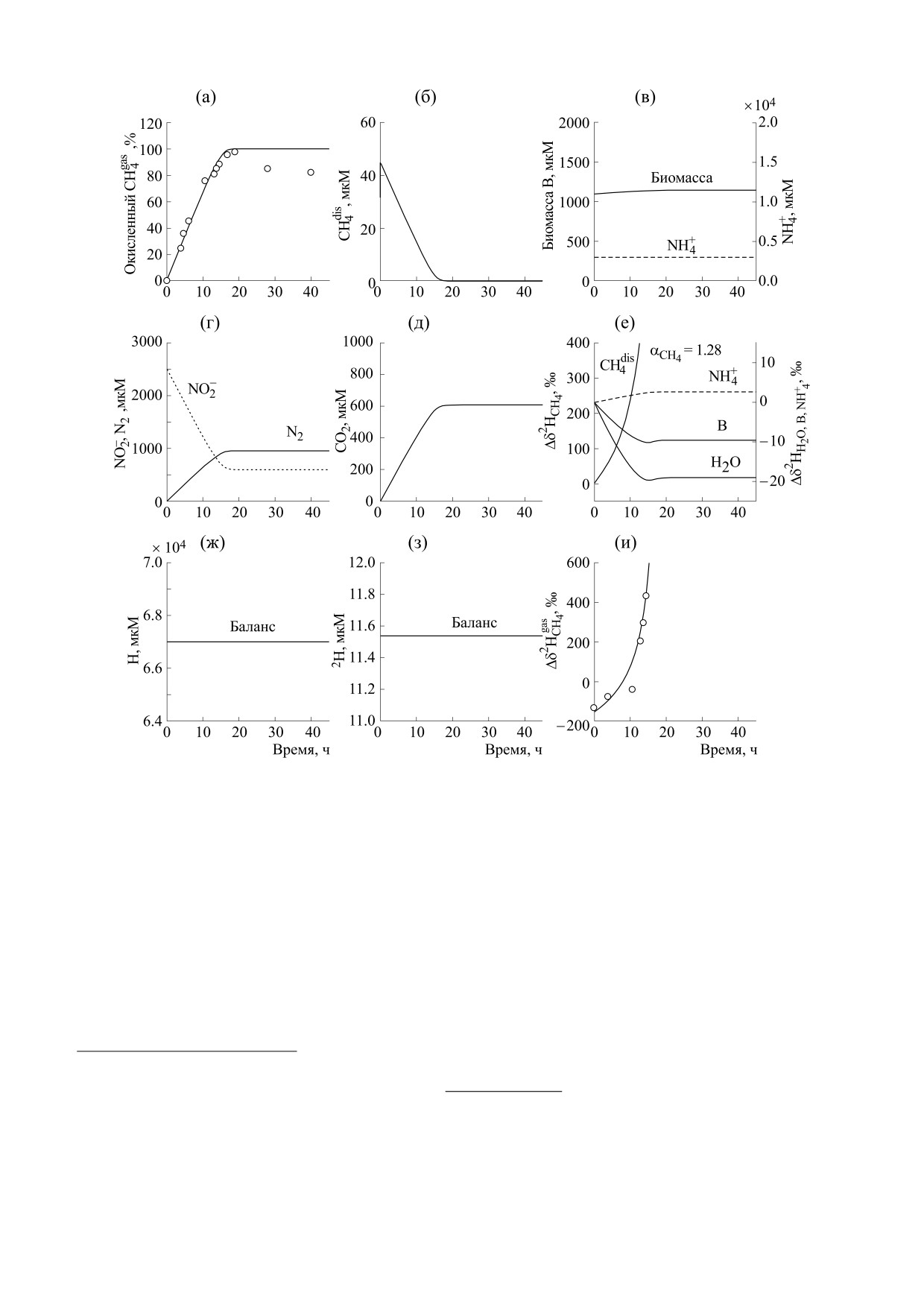
Рис. 1. Результаты расчета по динамической модели, описывающей анаэробное окисление метана нитритом NDAMO,
при начальных концентрациях газообразного метана CH4gas = 788 мкМ, биомассы B = 1100 мкМ, объемах газовой и
жидкой фаз реактора 15.6 и 44.4 мл соответственно (эксперимент R0.35). Символы (кружки) - эксперимент [14],
кривые - модель (10)-(11).
го раз меньше концентрации субстрата, содержаще-
Калибровку динамической модели проводили в
го более легкий изотоп, которая почти равна кон-
два этапа. Сначала калибровали модель с общими
центрации полного субстрата (с легким и тяжелым
переменными без участия изотопов, далее - часть
изотопом).
модели с изотопными переменными. Так, при
Ранее анализ экспериментальных данных при
моделировании процесса окисления метана нит-
моделировании микробиологических процессов
ритом (NDAMO) [10] вычисляли изотопную сиг-
выполнялся нами в основном для углерода 13C.
натуру по углероду для газообразного метана:
13
12
С
gas
/
С
gas
13
3
CH
4
CH
4
δ
С
gas
[‰]
=
10
−1,
(6)
CH
4
0.0112372
БИОФИЗИКА том 67
№ 6
2022
1146
ВАВИЛИН, ЛОКШИНА
13
12
Целью настоящей работы является описание
где
С gas
и
С gas
- содержание тяжелого и лег-
CH
4
CH
4
процесса NDAMO при рассмотрении дейтерия,
кого изотопа углерода в газообразном метане, со-
стабильного изотопа водорода. Для сравнения
ответственно; 13С/12С = 0.0112372 - стандарт для
теоретических кривых с экспериментальными
окаменелости моллюска Belemnitella americana из
данными для газообразного метана вычисляли
формации Пи-Ди, Южная Каролина, США
изотопную сигнатуру по водороду:
(PDB).
2
1
H
gas
/
H
gas
2
3
CH
4
CH
4
δ
H
gas
[‰]
=
10
−1,
(7)
CH
4
0.000150022503
2
1
ние объемов газовой и жидкой фаз равно 2.8). В
где
H gas
и
H gas
- содержание тяжелого и лег-
CH
4
CH
4
работе [15] ранее было вычислено, что начальная
кого изотопа водорода в газообразном метане со-
концентрация газообразного метана составляла
ответственно; 2Н/1Н = 0.000150022503 - венский
около 790 мкМ (R0.35) и 4780 мкМ (R2.8). Необ-
стандарт для морской океанической воды
ходимо отметить, что показанные на рис. 1 и 2
(V-SMOW). Далее теоретическую кривую сравни-
экспериментальные точки соответствуют разным
вали с экспериментальными данными.
сосудам, измерялись они лишь один раз. Соглас-
но авторам работы [14], максимальная удельная
МАТЕРИАЛЫ И МЕТОДЫ
скорость роста биомассы анаэробных метанокис-
Процесс NDAMO. Экспериментальная проце-
ляющих микроорганизмов (ANME) была почти
дура была детально описана ранее [14]. Культура
одинаковой в обоих экспериментах (R0.35 и
Candidatus Microbilis Oxyfera была обогащена в
R2.8).
среде без кислорода. При проведении двух инку-
Динамическая модель NDAMO. Ранее авторы
бационных экспериментов использовали 60-мл
стеклянные сосуды с объемами газовой и жидкой
работы [16] предложили стехиометрическое урав-
фаз 15.6 и 44.4 мл (R0.35, отношение объемов га-
нение для описания объединенного процесса
зовой и жидкой фаз равно 0.35) и с объемами га-
анаэробного окисления метана и денитрифика-
зовой и жидкой фаз 44.4 и 15.6 мл (R2.8, отноше-
ции нитрит-иона:
3CH4 + 8NO2- + 8H+ → 3CO2 + 4N2 + 10H2O.
(I)
В работе [17] было показано, что процесс анаэ-
для описания фракционирования изотопов угле-
робного окисления метана нитрит-ионом проис-
рода при анаэробном окислении метана, осу-
ходит внутри бактериальной клети, при этом в хо-
ществляемом культурой Candidatus Microbilis
де промежуточных реакций в процессе денитри-
Oxyfera. В настоящей работе это уравнение моди-
фикации нитрит-иона образуется кислород,
фицировано, чтобы описать динамику фракцио-
который и окисляет метан. В работе [10] автором
нирования изотопов водорода в процессе
было предложено стехиометрическое уравнение
NDAMO:
8
20
−
8
20
+
+
-
CH
+
−
Y
NO
+
−
Y
H
+YNH
+YOH
=
4
(
)
2
(
)
4
3
3
3
3
(II)
10
13
4
10
=
YC H NO
+
(
1−
5Y)CO
+
−
Y)HO
+
−
Y)N
,
5
7
2
2
(
2
(
2
3
3
3
3
где Y - коэффициент трансформации суб-
щих реакцию. В анаэробных условиях величи-
страта в биомассу, C5H7NO2 - обобщенная
на Y гораздо меньше, чем в аэробных.
химическая формула бактерий, осуществляю-
БИОФИЗИКА том 67
№ 6
2022
ПРИМЕНЕНИЕ БАЗОВЫХ ИЗОТОПНЫХ УРАВНЕНИЙ
1147
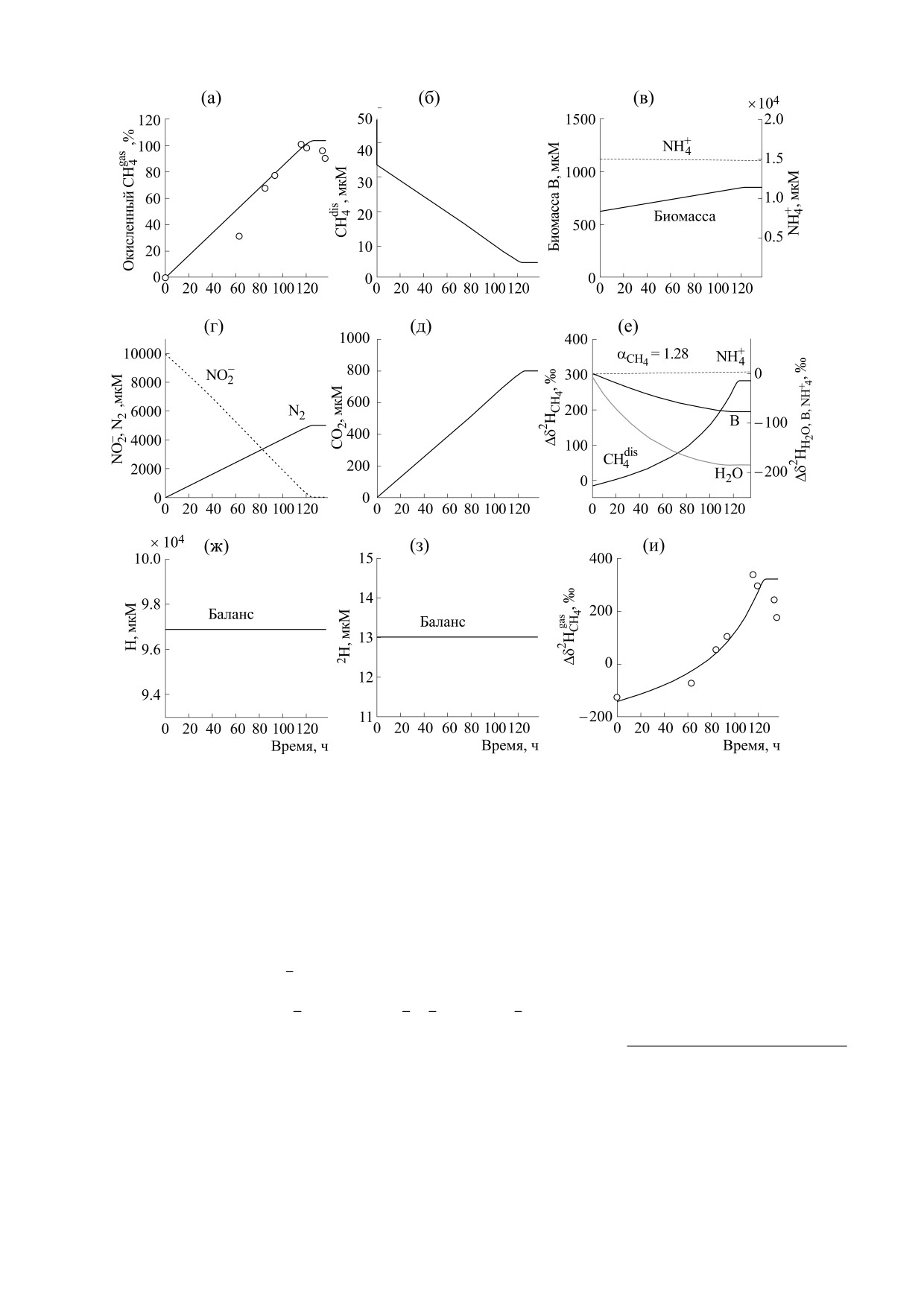
Рис. 2. Результаты расчета по динамической модели, описывающей анаэробное окисление метана нитритом NDAMO
при начальных концентрациях газообразного метана CH4gas = 4780 мкМ, биомассы B = 630 мкМ, объемах газовой и
жидкой фаз реактора 44.4 и 15.6 мл соответственно (эксперимент R2.8). Символы (кружки) - эксперимент [14],
кривые - модель (10)-(11).
Перепишем уравнение (II) в виде, удобном для дальнейшего рассмотрения:
8
20
-
+
CH
+
−
Y
NO
+YNH
=
YC
H
NO
+
(1 −
5Y
)CO
+
4
(
)
2
4
5
7
2
2
3
3
(III)
4
10
2
7
8
23
-
+
-
Y
N
+
+
Y
H
O
+
-
Y
OH
(
)
2
(
)
2
(
)
3
3
3
3
3
3
Следуя стехиометрическому уравнению
образного и растворенного метана запишем в
(III), динамическую модель для общих пере-
виде следующей системы дифференциальных
менных с учетом процессов массообмена газо-
уравнений:
БИОФИЗИКА том 67
№ 6
2022
1148
ВАВИЛИН, ЛОКШИНА
gas
dCH
dis
4
=-
K
(SS-CH
),
4
L
dt
dis
dCH
4
dis
=-R
+
K
(SS-CH
),
L
4
dt
-
dNO
2
8
−
20
=-
Y R,
dt
3
dB
=
YR,
dt
(8)
dN2
4
−10
=
Y R,
dt
3
dCO2
=
(1-
5Y)R,
dt
+
dNH4
=-
YR,
dt
−
+
dH
O
d(
H
2
O
недис
+OH
+
H
)
2
+
7Y
8
-
23Y
10
-16Y
2
=
=
R
+
R
+
R,
dt
dt
3
3
3
dis
-
+
CH
NO
NH
концентрации CH4dis и CDH3dis вычислялись как
4
2
4
где
R
=
ρmB
-
dis
-
SS = KHPCH4 и SSD = KHPCDH3, где KH - кон-
K
CH
+
CH
4
K
-
+
NO
2
K
+
+
NH+
4
4
NO
2
NH
4
скорость окисления растворенного метана в виде
станта Генри, PCH4 = 22.4-6×VL × CH4gas/VG и
нелинейной функции Моно для трех субстратов.
PCDH3 = 22.4×10-6VL×CDH3gas/VG - парциаль-
gas
dis
–
При этом
CH
,
CH
,
NO
, N2, CO2,
NH+4,
4
4
2
ные давления метана и метана, содержащего
H2Oнедис, OH- - концентрации соответствующих
2H(D), выраженные в атм; VL, VG - соответствен-
веществ; B - концентрация метанотрофов; ρm -
но объемы жидкости и газа в исследуемом реакто-
их максимальная удельная скорость окисления;
ре в литрах. Из работы [18] можно заключить, что
K
CH
,
K
–
,
K
+
- соответствующие константы
изменение константы Генри для CDH3 не окажет
4
NO
2
NH
4
полунасыщения; KL - коэффициент массообме-
сильного влияния на результаты, поскольку ме-
на между растворенным метаном и метаном в га-
тан мало растворим в воде. В связи с необходимо-
зовой фазе. В системе уравнений (8) рассматрива-
стью рассматривать содержание дейтерия в воде,
ется полная концентрация воды: H2O = H2Oнедис +
в модель в качестве переменной была введена
+ H+ + OH-.
концентрация воды H2O. Оценку величины на-
При описании процесса массообмена метана
чальной концентрации воды проводили следую-
между жидкой и газовой фазами равновесные щим образом:
m
H2O
1
ρ
VH2O H2O
1
4
V
H2O
H
O
=
=
=
5.6
⋅10
(мкмоль/мл)
(9)
2
H
O L
М V М V
H
O L
V
L
2
2
БИОФИЗИКА том 67
№ 6
2022
ПРИМЕНЕНИЕ БАЗОВЫХ ИЗОТОПНЫХ УРАВНЕНИЙ
1149
Таким образом, начальная концентрация воды
лось. Если для рассмотрения динамики 13C необ-
зависит от объема, который занимают остальные
ходимо было учитывать только один субстрат
вещества,
находящиеся
в
сосуде
(CH4), один продукт (CO2) и биомассу микроор-
(V
=
V
-V
). В табл. 2 приведены исполь-
H
2
O
L
вещества
ганизмов (C5H7NO2), содержащие углерод, то для
зуемые в модели начальные концентрации воды
для R0.35 и R2.8. В соответствии с существенно
рассмотрения динамики 2H необходимо принять
большим начальным содержанием веществ в слу-
во внимание уже два субстрата (CH4 и NH4+),
чае R2.8 (причем в модели рассматриваются толь-
один продукт (H2O) и биомассу (C5H7NO2), со-
ко вещества, входящие в стехиометрическое
держащие водород (см. «Приложение»).
уравнение (III)) начальная концентрация воды в
R2.8 много ниже, чем в R0.35.
Допуская различные факторы фракциониро-
Ранее [15] для изотопов водорода в процессе
вания изотопов водорода для субстратов метана
NDAMO кинетический изотопный эффект рас-
(
α
) и аммония (α
+
), можно записать следу-
CH
4
NH
сматривался лишь для одного субстрата - метана.
4
При этом фракционирование изотопов водорода
ющие уравнения для изменения концентраций
в продукте реакции - воде - вообще не вычисля-
изотопных переменных:
dis
dis
dСDH
3
1
СDH
3
1
dis
=-
R
+
L
K SSD -
СDH
3
)
dis
dt
α
CH
α
CH
4
4
gCH4
gas
dСDH
3
1
dis
L
K SSD -
СDH
3
)
dt
=-α
gCH4
dis
+
2H
d
B
R
1
СDH
Y
NDH
3
3
=
+
(10)
dis
+
dt
1+λ
α
CH
α
+
NH
CH
4
4
NH
4
4
dis
+
dHDO
R
1
СDH
3
Y
NDH
3
=
+
dis
+
dt
1+1/ λ
α
α
+
CH
4
CH
4
NH
4
NH
4
+
d
NDH+
3
1
NDH
3
=-
YR
+
dt
α
+
NH
NH
4
4
где
α
- кинетический фактор фракциониро-
по водороду между метаном в газовой фазе и рас-
gCH4
творенным в воде метаном вводится как пара-
вания по водороду в процессе массообмена мета-
на из воды в газовую фазу, соответственно:
метр, величина которого мала (α
=
0.01
[19]).
gCH
4
dis
gas
+
СDH
,
СDH
, 2HB, HDO,
NDH
- концентра-
3
3
4
Для того чтобы баланс в реакциях с изотопами
ции соответствующих реагентов, имеющих лишь
дейтерия сохранялся, необходимо считать, что
один изотоп дейтерия; λ - доля дейтерия, которая
часть выделяющейся воды и биомассы остается
в результате реакции оказывается в воде (см.
без дейтерия (см. «Приложение»). Тогда в модель
«Приложение»). В системе уравнений (10) фактор
для общих переменных (8) следует добавить ма-
кинетического изотопного фракционирования
лые поправки:
БИОФИЗИКА том 67
№ 6
2022
1150
ВАВИЛИН, ЛОКШИНА
gas
dCH
dis
4
=-
K
(SS-CH
),
4
L
dt
dis
+
dCH
1
NDH
3
dis
4
=-R1+
+
K
L
(SS-CH
4
),
+
dt
α
+
NH
NH
4
4
dis
+
dHNO
8
−
20
1
CDH
3
1
NDH
3
2
=-
Y R1+
+
,
dis
+
dt
3
α
α
CH
CH
4
+
NH
4
4
NH
4
dis
+
NDH
dB
1
1
CDH
3
1
Y
3
=
R
Y
+
Y
−
+
,
(
)
dis
+
dt
1+λ α
CH
1+1/λα
+
NH
CH
4
4
4
NH
4
dis
+
NDH
(11)
dN
2
4
−10
1
CDH
3
1
3
=
Y R1+
+
,
dis
+
dt
3
α
α
+
CH
CH
4
NH
4
4
NH
4
dis
+
dCO
1
CDH
1
NDH
3
2
3
=
(1-
5Y)
R1+
+
,
dis
+
d
t
α
CH
α
+
NH
CH
4
4
NH
4
4
+
dis
dNDH3
CDH
1
3
=-
YR
1+
,
dis
dt
α
CH
4
CH
4
dis
dH
2
O
10
−
16Y
10
-16Y
1
1
CD
H
3
=
R
+
−
+
dis
d
t
3
3
1
+1/λα
CH
CH
4
4
+
NDH
10
-
16Y
Y
1
3
+
−
3
1+1/λα
+
NH+
NH
4
4
Изотопные сигнатуры веществ по водороду
+
+
NDH
/4NH
2
3
4
вычисляли, используя величины переменных мо-
δ
H
+
[‰]
=
10
−
1
,
(13)
NH
4
3
дели, по следующим формулам:
0.000150022503
2H
CDH
/4CH
B/7B
2
3
4
2
δ
H
[‰]
=
10
−
1
,
(12)
δ
H
[‰]
=
10
-
1
,
(14)
CH
3
B
3
4
0.000150022503
0.000150022503
2(
2
+
7Y
)
+
(
8
−
23Y
)
HDO/2H
O
2
2
3
10
−16
Y
δ
H
H
O
[‰]
=
10
−
1
(15)
2
0.000150022503
В табл. 1 содержатся кинетические коэффици-
фициента трансформации субстрата в биомассу Y
енты полной динамической модели (10) - (11), а в
имеет существенное значение. ANME являются
табл. 2 - соответствующие начальные значения
медленно растущими микроорганизмами, что
переменных. Из систем дифференциальных
соответствует небольшой величине Y. Ранее в
уравнений (10) и (11) видно, что величина коэф-
работе
[16] сообщалось, что время удвоения
БИОФИЗИКА том 67
№ 6
2022
ПРИМЕНЕНИЕ БАЗОВЫХ ИЗОТОПНЫХ УРАВНЕНИЙ
1151
Таблица 1. Параметры полной динамической модели
Таблица 2. Начальные значения переменных динами-
(8)-(9)
ческой модели (8)-(9)
Переменная
Значение, мкМ
Параметр
Значение
gas
788.3
ρm
0.054 мкМ/сут
CH
4
Y
0.05 мкМ/мкМ
dis
31.5
CH
4
KCH4
2.00 мкМ
HNO2
2500.0
KHNO
43.00 мкМ
B
1100.0
2
KNH
0.01 мкМ
N2
0.01
3
CO2
0.000001
αH
1.31
NH3
3000.0
αgCH
1.01
4
H2O
10000.0
KL
100.0 сут-1
2
dis
0.004
H-СH4
KH
0.923 мкМ/атм
2
gas
0.098
H-СH4
VL
44.4 мл
2H-B
0.136
VG
15.6 мл
2H-H2O
0.372
2H-HNO2
0.310
биомассы Td микроорганизмов ANME равно од-
ной-двум неделям. Возьмем промежуточное зна-
2H-NH3
0.372
чение: Td =
1.5 недели. Используем формулу
Td=ln2/μm, где μm - удельная максимальная
скорость роста биомассы. Из нее найдем, что
[15]. Длительность реакции NDAMO зависит от
μm=Y, ρm=
0.0028 ч-1. Взяв значение ρm =
начальных концентраций СН4, NO2- и биомассы
= 0.054 мкМ·мкМ-1·ч-1, приведенное в табл. 1,
ANME. В процессе окисления метана концентра-
ция биомассы увеличивается. В эксперименте
получим Y ≈ 0.052 мкМ·мкМ-1.
R2.8, несмотря на значительно увеличенную на-
чальную концентрацию NO2- (табл. 2), длитель-
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
ность реакции NDAMO существенно возрастает,
На рис. 1 и 2 показана теоретическая динамика
поскольку начальная концентрация биомассы
процесса NDAMO для двух случаев (R0.35 и
ANME значительно меньше, чем в эксперименте
R2.8). Динамика для R2.8 с измененными объе-
R0.35 (сравни рис. 1в и 2в). Процесс прекращает-
мами газа и жидкости была использована нами
ся, когда концентрация одного из субстратов
для проверки предложенной математической мо-
уменьшается до 0: СН4 в R0.35 и NO2-в R2.8
дели. Видно, что теоретическая модель, включа-
ющая в качестве переменных концентрации ве-
(сравни рис. 1б и 2г).
ществ, содержащих дейтерий, в том числе био-
массу, успешно прошла эту проверку.
Поскольку начальные величины δ2Н для реа-
гентов и биомассы были нам неизвестны, на
В динамической модели учитывается, что про-
рис. 1е и 2е показана разница между текущими и
цесс массообмена метана между газовой и жид-
начальными значениями соответствующих вели-
кой фазами происходит быстро, а микробиологи-
ческая реакция NDAMO, идущая в воде, продол-
чин δ2Н, а именно: Δδ2Н = δ2Н(t) - δ2Н(0). Фрак-
жается достаточно долго. Концентрация
ционирование изотопов водорода прекращается,
растворенного метана является небольшой
когда скорость реакции NDAMO (III) стремится
(рис. 1б и 2б). Еще раз подчеркнем, что показан-
к 0. Фактор кинетического изотопного фракцио-
ные на рис. 1 и 2 экспериментальные точки соот-
нирования водорода для метана в процессе
ветствуют разным сосудам и получены в результа-
NDAMO является одинаковым, равным 1.28 в
те однократного измерения. Поэтому две точки
обоих экспериментах (рис. 1е и 2е). Он гораздо
на рис. 1а при t > 20 ч были исключены из рас-
больше, чем фактор фракционирования в
смотрения, поскольку являлись артефактами
процессе массобмена метана между его газооб-
БИОФИЗИКА том 67
№ 6
2022
1152
ВАВИЛИН, ЛОКШИНА
разной и растворенной формами (α
=
1.28
и
В системе сохраняется баланс атомов водорода
CH
4
H и дейтерия D (рис. 1з и 2з):
α
=
1.01).
gCH
4
dis
gas
+
dCH
dCH
dB
dNH
dH
O
4
4
4
2
4
+
d4
+
7
+
4
+
2
=
0,
dt
dt
dt
dt
dt
dis
gas
2H
+
dCDH
dCDH
d
B
dNDH
dHDO
3
3
3
+
+
+
+
=
0,
dt
dt
dt
dt
dt
где сомножители указывают на количество эле-
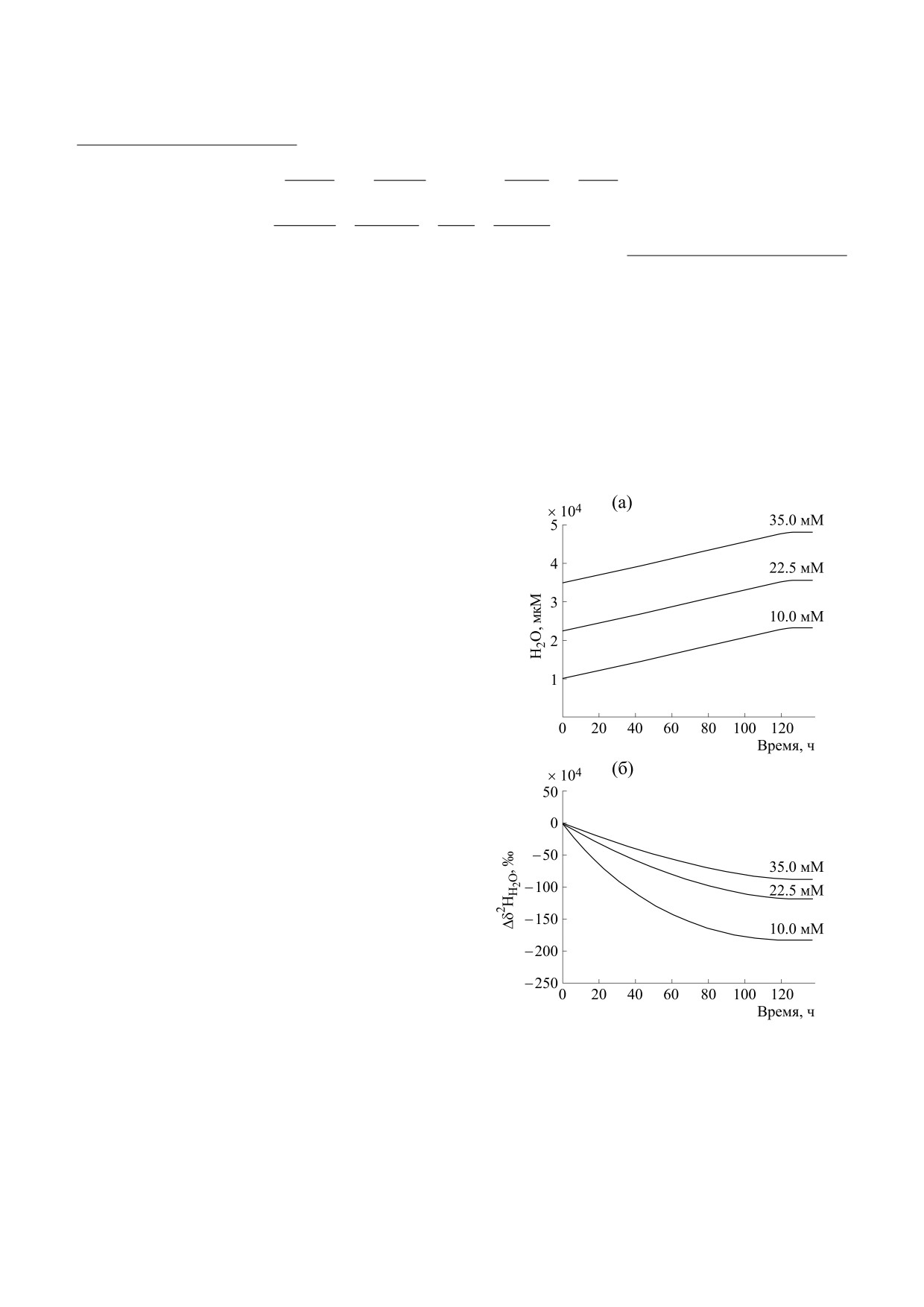
эксперименте он заканчивается при t ≥ 140 ч и
мента водорода в субстратах, биомассе и воде как
сопровождается значительным падением содер-
продукте реакции, тогда как для дейтерия счита-
жания дейтерия в воде, зависящим от начальной
ется, что лишь один его атом присутствует в реа-
концентрации воды, содержащей дейтерий. Это
гентах и биомассе.
происходит из-за разбавления воды в сосуде
В процессе микробиологической реакции,
обедненной дейтерием водой, образующейся в
идущей в воде, вследствие кинетического изотоп-
процессе NDAMO (рис. 3).
ного эффекта происходит изменение доли дейте-
В табл. 3 приводятся значения факторов фрак-
рия в реагирующих молекулах: субстраты (рас-
ционирования α для одиннадцати биологических
творенный метан и аммоний) обогащаются дей-
процессов, исследованных нами ранее. Среди
терием, а продукт реакции (вода) и биомасса им
них динамика роста водорослей (изотопы азота),
обедняются. Если в эксперименте R0.35 концен-
трация биомассы лишь немного возрастает
(рис. 1в), то в эксперименте R2.8 концентрация
биомассы возрастает существенно (рис. 2в). Зна-
чительное возрастание концентрации биомассы в
процессе NDAMO (сравни рис. 2в и 1в) приводит
к значительному изменению величины ее сигна-
туры Δδ2НВ (сравни рис. 2е и 1е). Также при не-
большой начальной концентрации воды (табл. 2)
значительно падает и содержание дейтерия в воде
(сравни рис. 2е и 1е).
Увеличение содержания дейтерия в метане
(рис. 2е) прекращается, когда реакция NDAMO
заканчивается (рис. 2г). Резкое увеличение вели-
чины δ2Н в метане (рис. 1е и 2е) происходит
вследствие резкого уменьшения его концентра-
ции. Для аммония, концентрация которого изме-
няется мало (рис. 1в и 2в), незначительно изменя-
ется и величина разности
Δδ2H
+
(рис. 1е и 2е).
NH
4
В целом использование концентраций ве-
ществ, содержащих дейтерий (субстратов CDH3 и
NDH3+, продукта HDO и биомассы C5DH6NO2), в
качестве изотопных переменных с сохранением
баланса водорода и дейтерия позволило описать
динамику процесса NDAMO. Согласно форму-
лам (12)-(15), изменение соответствующих вели-
чин Δδ2Н зависит от отношения концентраций
веществ, содержащих и не содержащих дейтерий.
В эксперименте динамика сигнатуры по водо-
роду измерялась лишь для метана, что позволило
определить фактор фракционирования H в мета-
Рис.
3. Теоретические кривые изменений общей
не (δ2H
). Если в первом эксперименте, сопро-
2
CH
концентрации воды (а) и величин
Δδ
H
(б) в
4
H2
0
вождающемся небольшим падением содержания
процессе NDAMO при разных начальных
дейтерия в воде и биомассе, процесс фракциони-
концентрациях воды
(10,
22.5 и
35 мМ) для
рования заканчивается при t ≥ 17 ч, то во втором
эксперимента R2.8.
БИОФИЗИКА том 67
№ 6
2022
ПРИМЕНЕНИЕ БАЗОВЫХ ИЗОТОПНЫХ УРАВНЕНИЙ
1153
Таблица 3. Исследуемые процессы, константа скорости и фактор фракционирования стабильных изотопов
Максимальная
удельная
скорость роста
Фактор
Процесс
биомассы
фракционирован
Примечания
(начальная
ия изотопов
концентрация
биомассы) *
Аммоний:
0.095 ч-1
αN = 1.02
9.3 мкМ
1. Рост водорослей Thalassiosira
Нитрат:
Для смеси N-субстратов высокая
pseudonana на отдельном N-
0.071 ч-1
αN
= 1.0047
концентрация аммония ингибирует
субстрате и их смеси (данные
3.2 мкМ
потребление нитрата и мочевины
[20], модель [21])
Мочевина:
0.084 ч-1
αN = 1.0008
4.0 мкМ
Этанол:
Изотопные уравнения для субстрата,
2. Денитрификация нитрата в
0.72 сут-1
αN = 1.0132
продукта и биомассы используется как
азот почвенными
0.0025 мМ
составные части двух последовательных
микроорганизмами с
реакций. Промежуточная концентрация
органическими субстратами
Глюкоза:
нитрита была небольшой для
глюкозой и этанолом (данные
1.64 сут-1
αN = 1.0175
экспериментов с этанолом и большой
[22], модель [8])
0.0004 мМ
для экспериментов с глюкозой
3. Метанизация ацетата (данные
[23], модель [24]):
Merthanosarcina-
ceae:
αС = 1.02
-1
0.36 сут
Идет конкуренция за ацетат между двумя
а) при низкой концентрации
0.0017 г·л-1
группами ацетокластических
аммония 0.18 г·л-1
Methanosaetaceae:
метаногенов
0.2 сут-1
αС = 1.005
0.0057 г·л-1
Methanosarcina-
ceae:
αС = 1.02
0.054 сут-1
0.0017 г·л-1
Синтрофы,
Происходит замещение
окисляющие
ацетокластического метаногенеза на
б) при высокой концентрации
ацетат:
αС = 1.02
водородотрофный метаногенез. При
аммония 5.4 г·л-1
0.08 сут-1
этом Methanosaetaceae полностью
0.00011 г·л-1
исчезают к концу реакции
Водородотрофн
ые метаногены:
αС = 1.065
2 сут-1
0.00000011 г·л-1
Фракционирование изотопов идет в
4. Аэробное окисление метана
процессе микробного окисления
(данные [25], модель [26]):
растворенного метана
Methylocadmium
CH4
Окисление метана по рибулезо-
α
= 1.015
gracile:
C
а) рибулезо-фосфатный путь
фосфатному пути идет гораздо
0.176 ч-1
= 1.11
αCH4H
медленнее, чем по сериновому
0.2 мМ
БИОФИЗИКА том 67
№ 6
2022
1154
ВАВИЛИН, ЛОКШИНА
Таблица 3. Продолжение
Cu - в избытке:
αCH4C
= 1.02
Methylosinus spo-
В сериновом пути рост биомассы
rum:
αCH4H
= 1.15
б) сериновый путь
увеличивается с возрастанием
1.12 ч-1
Cu - лимитирует:
концентрации CO2
0.025 мМ
αCH4C
= 1.017
αCH4H
= 1.16
Ацетокластичес-
кие метаногены:
αAc-метаногенез
= 1.01
0.0625 сут-1
0.0028 г·л-1
5. Образование метана и
Синтрофы,
Доминирует водородотрофный
углекислого газа в донных
окисляющие
метаногенез, поскольку ацетат
ацетат:
Ac-окисление
отложениях тропических озер
α
= 1.01
переходит в H2 и CO2 из-за
(озеро Jatoba) (данные [27],
0.05 сут-1
синтрофного окисления ацетата
модель [28])
0.0034 г·л-1
Водородотроф-
ные метаногены:
H2/СO2-метаногенез
αC
1.5 сут-1
= 1.072
0.000023 г·л-1
Ацетокластическ
ие метаногены:
αAc-метаногенез =
0.02 сут-1
1.008
6. Образование метана в торфе
Отношение 13C/12C в CO2 определяет
0.0073 г·л-1
бореального мезотрофного
динамические изменения в скоростях
Водородотроф-
болота (данные [29], модель [30])
отдельных реакций
ные метаногены:
=
αH2/СO2-метаногенез
0.125 сут-1
1.075
0.0011 г·л-1
Кислотогенез:
1.0 сут-1
= 1.002
αКислотогенез
0.00000023 г·л-1
Анаэробное
дыхание:
=
αАнаэробное дыхание
2.5 сут-1
1.002
0.00000056 г·л-1
Анаэробное дыхание и кислотогенез
Ацетокластическ
7. Образование метана в
конкурируют за мономер целлюлозы
ий метаногенез:
=
αAc-метаногенез
тропическом озере Ladario с
Высокое соотношение CO2/CH4 с
0.075 сут-1
1.005
задержкой из-за анаэробной
задержкой образования метана
0.00028 г·л-1
респирации (данные [27],
интерпретировалось как
модель [31])
Синтрофное
доминирование анаэробного дыхания
окисление
в начале процесса.
ацетата:
= 1.005
αAc-окисление
0.1 сут-1
0.000056 г·л-1
Водородотроф-
ный метаногенез:
=
αH2/СO2-метаногенез
1.75 сут-1
1.05
0.000000011 г·л-1
БИОФИЗИКА том 67
№ 6
2022
ПРИМЕНЕНИЕ БАЗОВЫХ ИЗОТОПНЫХ УРАВНЕНИЙ
1155
Таблица 3. Окончание
Начальные
концентрации
ANME и метана
в газовой фазе:
В проточной системе с постепенным
8. Формирование анаэробного
увеличением входящей концентрации
1.2 г·л-1 , 3.3 г·л-1
αS = 1.040
консорциума метан-окисляющих
метана идет увеличение концентрации
архей (ANME) и сульфат-
ANME в консорциуме микроорганизмов
2.0 г·л-1, 8.0 г·л-1
αS = 1.024
редуцирующих бактерий (DSS)
и уменьшение фактора
(данные [32], модель [33])
фракционирования изотопа серы в
2.4 г·л-1, 14 г·л-1
αS = 1.023
сульфате
3.0 г·л-1, 26 г·л-1
αS = 1.022
3.5 г·л-1, 44 г·л-1
αS = 1.018
Начальные
концентрации
ANME:
9. Линейное окисление метана
Hydrate Ridge:
сульфатом в трех периодических
αС = 1.012
Наименьший фактор фракционирования
8.4 мМ
реакторах, иноукулированных
изотопа углерода в метане получен при
морскими донными
наибольшей концентрации анаэробных
Amon Mud Vol-
отложениями (данные [7],
метанотрофных микроорганизмов
cano:
αС = 1.02
модель [33])
1.2 мМ
Black Sea:
αС = 1.04
1.28 мМ
Гидролиз/кислот
αКислотогенез
=
огенез
1.002
целлюлозы:
0.013 сут-1
10. Продукция метана и
Ацетокластическ
αAc-метаногенез =
Продукция метана в три раза больше, чем
углекислого газа в образцах
ий метаногенез
продукция углекислого газа.
1.005
торфа западносибирского
0.105 сут-1
Доминирование ацетокластического
эвтрофного болота (данные и
метаногенеза над водородотрофным из-
модель [34])
Водородотрофн
H2/СO2-метаногенез
за активности гомоацетогенов
αC
ый метаногенез:
= 1.08
0.912 сут-1
Гомоацетогенез:
αГомоацетогенез =
0.912 сут-1
1.08
αмассообмен
=
Концентрация растворенного метана
лимитирует общую скорость его
Анаэробное
1.001
окисления. Идет обогащение метана
окисление
11. Анаэробное окисление метана
более тяжелым углеродом 13С. Фактор
метана
нитритом (данные [14], модель
фракционирования изотопа углерода в
нитритом:
[10]
αCокисление
=
метане при его микробиологическом
0.016 сут-1
1.030
окислении гораздо больше, чем в
0.014 г·л-1
процессе массообмена между
растворенной и газовой формами метана
Примечание. * - Величина максимальной удельной скорости роста биомассы μm зависит от начальной концентрации
биомассы B0. Чем больше B0, тем ниже величина μm.
БИОФИЗИКА том 67
№ 6
2022
1156
ВАВИЛИН, ЛОКШИНА
денитрификация нитрата почвенными микроор-
БЛАГОДАРНОСТИ
ганизмами (изотопы азота), метанизация ацетата
Авторы признательны В.С. Брезгунову за цен-
при различных концентрациях аммония (изото-
ные советы при описании динамики фракциони-
пы углерода), образование метана в донных отло-
рования стабильных изотопов и Д.С. Ремизову за
жениях тропических озер (изотопы углерода),
предложенный метод оценки стехиометрических
анаэробное окисление метана сульфатом и фор-
коэффициентов в использованных химических
мирование консорциума метан-окисляющих ар-
уравнениях.
хей и сульфатредуцирующих бактерий в морских
экосистемах (изотопы серы и углерода). В цити-
руемых в табл. 3 математических моделях исполь-
ФИНАНСИРОВАНИЕ РАБОТЫ
зовался лишь один субстрат, содержащий разные
Работа выполнена согласно плану Института
стабильные изотопы, прежде всего, углерода 13С в
водных проблем РАН (номер темы Госзадания
целлюлозе [21, 28, 30, 34] и ацетате [24] с образо-
AAA-A18-118022090104-8).
ванием СН4 и СО2, изотопы азота 15N в азотсо-
держащих субстратах [21] и нитрите [9], транс-
КОНФЛИКТ ИНТЕРЕСОВ
формирующемся в закись азота и молекулярный
азот, изотопы серы 35S в сульфате [33], трансфор-
Авторы заявляют об отсутствии конфликта
мирующемся в сероводород через образование
интересов.
нуль-валентной серы. При моделировании про-
цессов аэробного [26] и анаэробного окисления
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
метана сульфатом [10] наряду с изотопом 13С ис-
Настоящая статья не содержит каких-либо ис-
пользовался и изотоп дейтерия 2Н. Однако при
следований с участием людей или животных в ка-
этом не учитывалось, что помимо метана могут
честве объектов исследований.
быть и другие дейтерийсодержащие субстраты.
СПИСОК ЛИТЕРАТУРЫ
ЗАКЛЮЧЕНИЕ
1. Э. М. Галимов, Изотопы углерода в нефтегазовой
Любая математическая модель основана на не-
геологии (Наука, М., 1973).
избежном упрощении свойств реального объекта.
2. H. Craig, Geochim. Cosmochim. Acta, 12, 133 (1957).
Измерение изотопных переменных позволяет по
3. J. W. C. Rayleigh, Philos. Mag., 42, 493 (1896).
возможности заглянуть внутрь «черного ящика».
4. A. Mariotti, J. C. Germon, P. Hubert, et al., Plant and
При этом математическое описание динамики
Soil, 62, 413 (1981).
микробиологических процессов, учитывающее
5. Л. Меландер и У. Содерс, Скорости реакций изо-
изотопы, может быть довольно сложным, по-
топных молекул (Мир, М., 1983).
скольку оно всегда должно опираться на сохране-
ние баланса химических элементов, в том числе
6. S. Bakkaloglu, D. Lowry, R. E. Fisher, et al., Waste
их изотопов, и становится оправданным, если
Managem., 132, 162 (2021).
при этом открываются новые знания о системе.
7. T. Holler T., G. Wegener, K. Knittel, et al., Env. Micro-
Совместное описание нелинейной динамики об-
biol. Rep., 1, 370 (2009).
щих и изотопных переменных позволило стан-
8. V. A. Vavilin and S. V. Rytov, Chemosphere, 134, 417
дартным способом математически анализировать
(2015).
последовательные и параллельные метаболиче-
9. V. A. Vavilin, L. Y. Lokshina, and S. V. Rytov, Isotopes
ские пути превращений различных субстратов в
in Environmental and Health Studies, 58, 44 (2022).
природных и техногенных химико-биологиче-
10. V. A. Vavilin, Env. Dyn. Glob. Climate Change, 12 (2),
ских системах.
(2021). DOI: 10.17816/edgcc58932
Показано, что в процессе микробиологиче-
11. D. J. Batstone, J. Keller, I. Angelidaki, et al., Anaerobic
ской реакции NDAMO, идущей в воде, вслед-
Digestion Model No.1 (ADM1) (IWA Press, Padstow,
ствие кинетического изотопного эффекта проис-
Cornwall, UK, 2002).
ходит изменение доли дейтерия в реагирующих
12. Y. Liu, Appl. Microbiol. Biotechnol., 73 (6), 1241
молекулах, при этом субстраты обогащаются дей-
(2007).
терием, а продукт (Н2О) и биомасса им обедня-
13. В. А. Вавилин и В. Б. Васильев, Математическое
ются.
моделирование процессов биологической очистки
сточных вод активным илом (Наука, М., 1979).
В обобщающей табл. 3 приведены факторы
фракционирования соответствующих изотопов
14. O. Rasigraf, C. Vogt, H. H. Richnow, et al., Geochim.
для одиннадцати экологических процессов с при-
Cosmochim. Acta, 89, 256 (2012).
мечаниями к «механизму» рассматриваемых про-
15. V. A. Vavilin and S. V. Rytov, Ant. Leewenh., 104, 1097
цессов.
(2013).
БИОФИЗИКА том 67
№ 6
2022
ПРИМЕНЕНИЕ БАЗОВЫХ ИЗОТОПНЫХ УРАВНЕНИЙ
1157
16.
A.A. Raghoebarsing, A. Pol, K.T. van de Pas-Schoo-
25. S. Feisthauer, C. Vogt, J. Modrzynski, et al., Geochim.
nen, et al., Nature, 440, 918 (2006).
Cosmochim. Acta, 75, 1173 (2011).
17.
K. F. Etwig, M. K. Butler, D. Le Paslier, et al., Nature,
26. V. A. Vavilin, S. V. Rytov, N. Shim, and C. Vogt, Iso-
464, 543 (2010).
topes in Environmental and Health Studies, 52, 185
(2016).
18.
M. F. Costa Gomes and J.-P. Grolier, Phys. Chem.
27. R. Conrad, M. Noll, P. Claus, et al., Biogeosciences, 8,
Chem. Phys., 3, 1047 (2001).
795 (2011)
19.
M. Knox, P. D. Quay, and D. Wilbur, Geophys. Res.,
28. V. Vavilin, S. Rytov, and R. Conrad, Ecol. Modell.,
97 (C12), 20335 (1992).
363, 81 (2017).
20.
N. A. D. Waser, P. J. Harrison, B. Nielsen, et al., Lim-
29. P. Galand., K. Yrjälä, and R. Conrad, Biogeosciences,
nol. Oceanogr., 43 (2), 215 (1998).
7, 3893 (2010).
21.
V. A. Vavilin, S. V. Rytov, and L. Y. Lokshina, Ecol.
30. V. A. Vavilin, S. V. Rytov, and L. Y. Lokshina, Isotopes
Modell., 279, 45 (2014).
in Environmental and Health Studies, 54, 475 (2018).
22.
G. Vidal-Gavilan, A. Folch, N. Otero, et al., Appl.
31. V. A. Vavilin, S. V. Rytov, and L. Y. Lokshina, Ecol.
Geochem., 32, 153 (2013).
Modell., 386, 59 (2018).
23.
J. Grossin-Debattista, Docteur these (Universite Bor-
32. C. Deusner, T. Holler, G. L Arnold, et al., Earth Plan-
et. Sci. Lett., 399, 61 (2014).
bordeaux1.fr/pdf/2011/GROSSIN-DEBATTIS-
33. V. A. Vavilin, L. Y. Lokshina, and S. V. Rytov, Wat. Sci.
TA_JULIEN_2011.pdf
Technol., 79 (11), 2056 (2019).
24.
V. A. Vavilin and S. V. Rytov, Isotopes in Environmen-
34. L. Y. Lokshina, V. A. Vavilin, Y. Litti, et al., Wat. Re-
tal and Health Studies, 53, 135 (2017).
sour., 46 ( Suppl.), S110 (2019).
Application of the Basic Isotope Equations for Description of the Dynamics
of Microbiological Processes: Deuterium Redistribution
V.A. Vavilin* and L.Y. Lokshina*
*Water Problems Institute, Russian Academy of Sciences, ul. Gubkina 3, Moscow, 119333 Russia
Basic equations used to describe isotope dynamics that take into account the retention of deuterium equilib-
rium, were helpful in two laboratory experiments for analyzing the dynamics of nitrite-dependent anaerobic
methane oxidation (NDAMO) at different initial concentrations of the substrates and deuterium isotope vari-
ables. Notably, in the second experiment, the initial amount of water in a closed vessel was reduced to about
2.8 times. Methane is completely consumed but nitrite ions are still present in excessive amounts at the end
of the first experiment, while methane is present but nitrite ions are completely exhausted at the end of the
second experiment. In the model, concentrations of the substrates containing a single deuterium atom (СН4
and NH4+), concentration of the product (Н2О), and concentration of the biomass of methanotrophic mi-
croorganisms ANME (C5H7NO2) are taken as isotope variables. Stoichiometric reaction equations are de-
rived to describe the redistribution of deuterium between the reaction substrates, product (water), and bio-
mass. The isotope fractionation is shown to proceed in the course of a microbiological reaction in water. As
a result, the substrates become enriched in deuterium, as water and biomass become depleted of deuterium.
In the first experiment accompanied by a slight drop in the content of deuterium in biomass and water, the
fractionation process ends after t ≥ 17 h. In the second experiment, the content of deuterium in water and bio-
mass decreases significantly and fractionation ends after t ≥ 140 h. This is due to the dilution of the water in
the vessel with the deuterium-depleted water generated by the NDAMO process. This paper also provides a
summary of the results of our study on the modeling of the dynamics of eleven biological processes, in which
mainly stable carbon isotopes have been measured, based on the information gathered in a longitudinal study.
The isotope fractionation factors used in the simulation are presented.
Keywords: dynamics of ecological processes, kinetic isotope effect, basic isotope equations, serial and parallel re-
actions
БИОФИЗИКА том 67
№ 6
2022