БИОФИЗИКА, 2022, том 67, № 6, с. 1220-1223
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 577.3
ПУРИНЕРГИЧЕСКИЕ МЕХАНИЗМЫ АДАПТАЦИИ РАЗЛИЧНЫХ ТИПОВ
ДВИГАТЕЛЬНЫХ ЕДИНИЦ
В УСЛОВИЯХ АЛЛЕРГИЧЕСКОЙ ПЕРЕСТРОЙКИ
© 2022 г. А.Е. Хайруллин*, **, #, А.Ю. Теплов*, С.Н. Гришин*, А.У. Зиганшин*
*Казанский государственный медицинский университет, ул. Бутлерова, 49, Казань, 420012, Россия
**Казанский федеральный университет, ул. Кремлевская, 18, Казань, 420008, Россия
#E-mail: khajrulli@yandex.ru
Поступила в редакцию 27.06.2022 г.
После доработки 27.06.2022 г.
Принята к публикации 11.07.2022 г.
На изолированных двигательных мышцах мыши: «быстрой» (m. EDL) и «медленной» (m. soleus) ме-
тодом изометрической электростимуляционной механомиографии выявлена роль пресинаптиче-
ского звена сигнализации в процессах адаптации к аллергии. Различия в динамике функциональ-
ных характеристик мышц под влиянием экзогенной АТФ у интактных и сенсибилизированных
яичным альбумином животных демонстрируют участие пуринергических механизмов в развитии
приспособительных изменений в пресинаптическом звене «быстрой» мышцы, и меньшее вовлече-
ние этих механизмов на сходной структуре у «медленной» мышцы. В процессе белковой сенсиби-
лизации комплексная работа АТФ зависимых механизмов как пост-, так и пресинаптической мем-
браны обеспечивает адекватную перестройку сократительной функции локомоторных мышц в
условиях аллергии, и у разных мышц эти механизмы существенно различаются.
Ключевые слова: белковая сенсибилизация, аллергия, АТФ, мышь, m. EDL, m. soleus.
DOI: 10.31857/S0006302922060157, EDN: LLXGMU
если у «медленной» мышцы постсинаптический
Изучение механизмов динамики сократитель-
эффект обеспечивался АТФ-зависимыми меха-
ной функции мышечной системы в условиях ал-
лергической перестройки остаются актуальными
низмами, то у «быстрой» мышцы их реализация
осуществлялась каким-то иным путем. Адапта-
в современной спортивной медицине [1, 2]. В
ция перечисленных мышц к аллергическим изме-
частности, как известно, обязательная вакцина-
нениям не ограничивается постсинаптическим
ция спортсменов перед соревнованиями приво-
звеном сигнализации, и мы можем предполагать
дит к изменению функции локомоторных мышц.
иные механизмы реализации этих эффектов.
Ранее нами было показано, что в условиях аллер-
гии различные двигательные мышцы меняют си-
Целью настоящего исследования явилось изу-
лу своего сокращения на холиномиметик in vitro,
чение роли пуринергических механизмов различ-
что указывает на участие в механизмах адаптации
ных двигательных мышц мыши: «быстрой» -
постсинаптического звена сигнализации. Была
m. EDL и «медленной» - m. soleus в процессах
продемонстрирована роль пуринергических ме-
адаптации организма при аллергической пере-
ханизмов в этих процессах. При этом у мышц с
стройке.
различным волоконным составом способность
изменять силу своего сокращения на карбахолин
МЕТОДЫ
после апликации экзогенной АТФ проявляется в
разной степени. Механизмы адаптации в услови-
Подготовительные процедуры. Эксперименты
ях развивающейся аллергии у разных мышц так-
проводили на изолированных нервно-мышечных
же существенно различаются. В частности, если
препаратах m. soleus и m. EDL 24 белых мышей
«медленная» мышца в условиях белковой сенси-
обоего пола, массой 25-32 г. Животных содержа-
билизации увеличивала силу своего карбахолин-
ли в специальных боксах на обычном пищевом
индуцированного сокращения, то у «быстрой»
режиме, со свободным доступом к воде. Опытную
мышцы наблюдалась обратная картина [3, 4]. И
группу животных предварительно сенсибилизи-
ровали овальбумином [3]. В качестве контроля
Сокращение:БС - белковая сенсибилизация.
использовали несенсибилизированных мышей.
1220
ПУРИНЕРГИЧЕСКИЕ МЕХАНИЗМЫ АДАПТАЦИИ
1221
Условия проведения экспериментов по регистра-
ции параметров сокращения. Мышечный препарат
помещали вертикально в термостатируемых ван-
ночках объемом 10 мл. Один конец мышцы был
зафиксирован, другой при помощи лигатуры кре-
пили к датчику механической активности. Ван-
ночку заполняли модифицированным раствором
Кребса следующего состава (в мМ): NaCl - 118.0,
KCl
- 4.75, CaCl2
- 2.5, NaHCO3
- 24.8,
KH2PO4 - 1.18, MgSO4·7H2O - 1.18, глюкоза -
11.0, рН 7.4 ± 0.1. Условия изометрии достигались
растяжением препарата мышцы в ванночке в те-
чении 20 мин с силой 0.5 г при постоянной пер-
фузии раствором при температуре 37°С. Сократи-
тельные реакции изолированной мышцы вызы-
вались при помощи электрического стимулятора
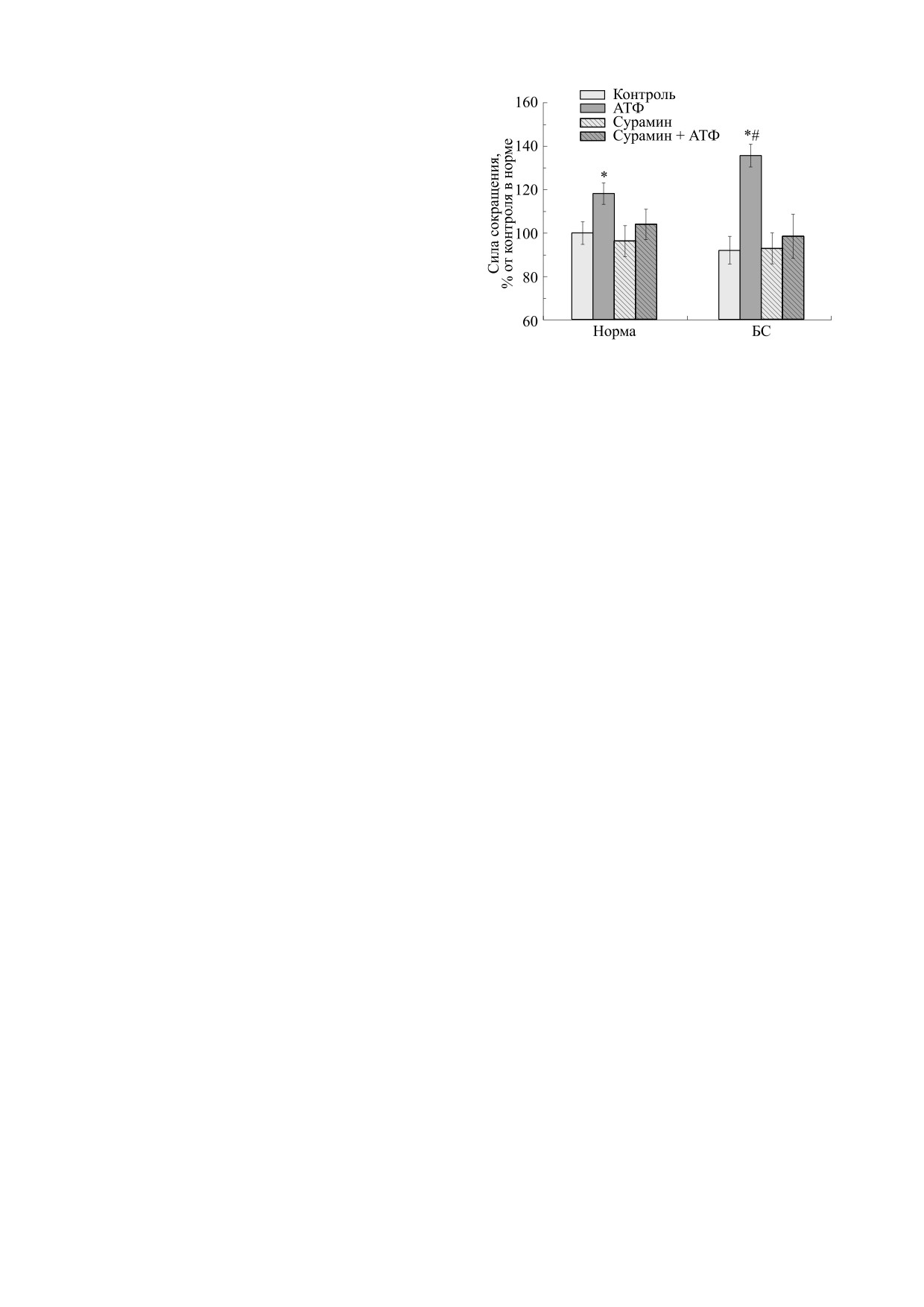
Рис. 1. Сила сокращений m. EDL мыши, вызванных
MultiStim D330 (Digitimer, Великобритания) [5].
электрической стимуляцией у интактных («Норма»,
n = 14) и сенсибилизированных («БС», n = 10) живот-
Прямоугольные электрические импульсы часто-
ных, в отсутствие и в присутствии АТФ (100 мкМ) и
той 0.1 Гц и длительностью 0.5 мс наносились в
сурамина (100 мкМ). Результаты представлены в виде
течении двух минут (12 пиков). Среднее значение
M ± m в % от исходных величин, принятых за 100%;
силы всех 12 сокращений анализировали как
* - р < 0.05 от контроля в норме, # - р < 0.05 относи-
один результат. Силу сокращения оценивали в
тельно эффектов в норме.
граммах. Ответы записывали с помощью изомет-
рического механического датчика FSG-01 (Linton
Instrumentation, Великобритания) и аналого-
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
цифрового преобразователя MP100WSW (Bi-
«Быстрая» мышца в условиях электростимуля-
opack, США) [6, 7]. Результаты исследования на-
ции снижала силу своего сокращения при белко-
блюдали на мониторе и хранили на персональном
вой сенсибилизации (БС). Если у несенсибили-
компьютере.
зированной мыши она составляла 302.4 ± 15.7 мг
(n = 14), то в опытной серии - 278.5 ± 19.3 мг
Эффекты пуринергических агонистов и антаго-
(n = 10).
нистов. В начале эксперимента сократительные
Действие экзогенной АТФ
(100 мкМ) на
ответы мышц на электрическую стимуляцию ре-
m. EDL обеих групп животных демонстрировало
гистрировались дважды с интервалом 5 мин (для
следующую динамику силы сокращения. У не-
оценки стабилизации ответов). Раствор АТФ до-
сенсибилизированных мышей этот показатель
бавляли в концентрации 100 мкМ в ванночку с
возрастал до 357.4 ± 14.8 мг, то есть на 18.2%
раствором Кребса, через 10 мин инкубации запи-
(n = 14, p < 0.05); у сенсибилизированных живот-
сывали сократительные ответы на электрическую
ных - до 410.3 ± 15.7 мг, то есть на 47.3% относи-
стимуляцию. Затем ткань промывали несколько
тельно контроля (n = 10, p < 0.05, рис. 1). Более
раз и инкубировали в течение 20 мин антагони-
выраженное увеличение силы сокращения
стом (сурамином в концентрации 100 мкМ). По-
«быстрой» мышцы у второй группы животных
сле этого добавляли агонист (АТФ в концентра-
(18.2% против 47.3%; p < 0.05) свидетельствует об
ции 100 мкМ) и сократительные ответы записы-
участии пуринергических механизмов пресинап-
вали повторно. Все сократительные ответы
тических образований в процессах адаптации.
вычисляли в процентах от исходных сокращений.
Инкубация мышцы с сурамином (100 мкМ) пол-
ностью предупреждала влияние АТФ на силу со-
Достоверность различий. Статистическую об-
кращений.
работку данных проводили с помощью програм-
мы для статистической обработки SPSS Statistics.
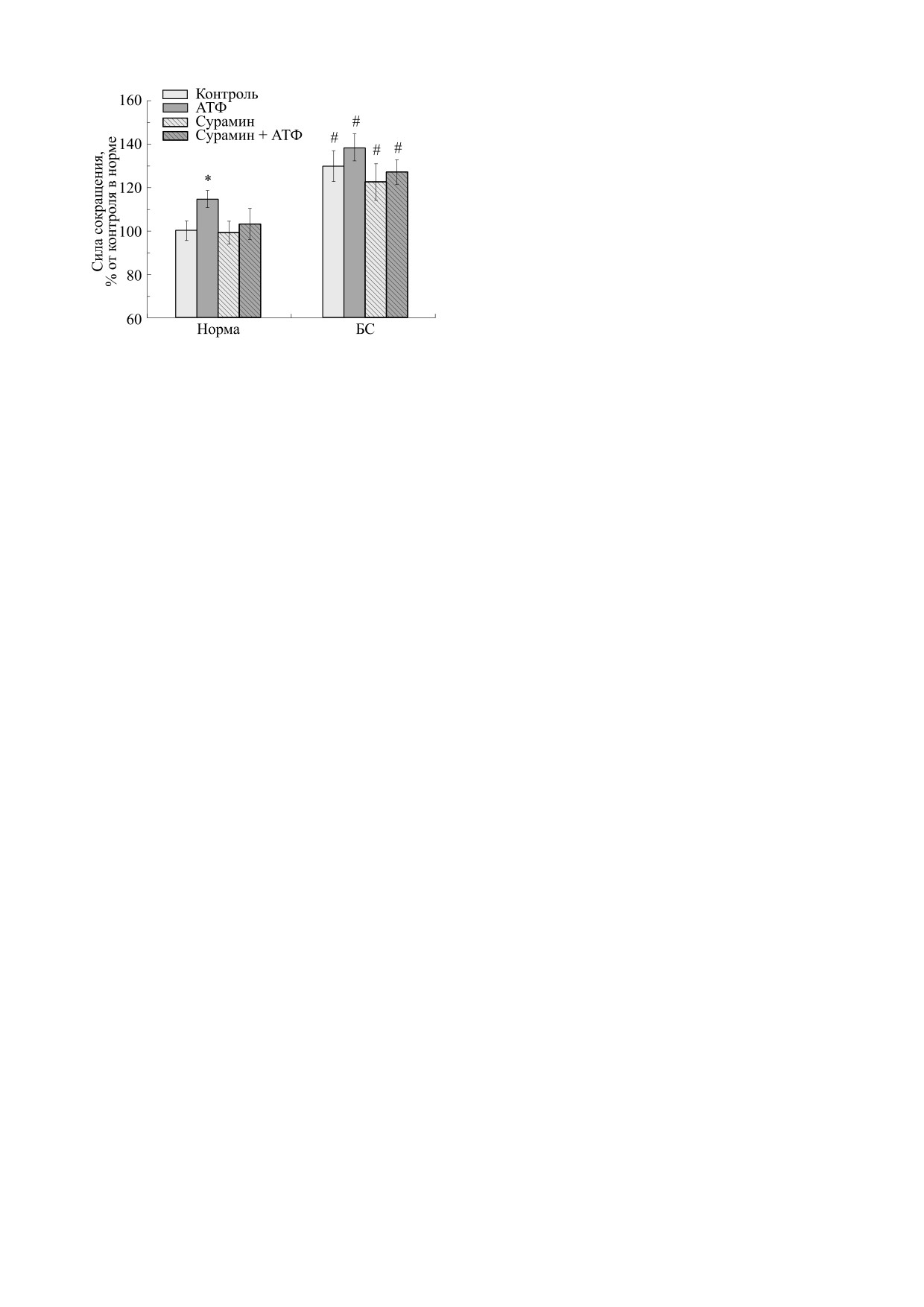
«Медленная» мышца в условиях электрости-
Проверку соответствия полученных данных нор-
муляции увеличивала силу своего сокращения
мальному распределению проводили с помощью
при БС. Если у несенсибилизированной мыши
критерия Колмогорова. Рассчитывали средние
она была 411.5 ± 18.5 мг (n = 14), то в опытной се-
арифметические анализируемых параметров и
рии - 533.3 ± 29.2 мг (n = 10).
стандартную ошибку. Статистическую значи-
Влияние экзогенной АТФ на m. soleus обеих
мость наблюдаемых изменений оценивали с по-
групп животных на различия в силе сокращения
мощью критерия Стьюдента для независимых и
носило потенцирующий характер. Мы получили
попарно сопряженных выборок. Различия рас-
следующие значения. У несенсибилизированных
сматривали как значимые при p < 0.05.
мышей показатель силы сокращения возрастал до
БИОФИЗИКА том 67
№ 6
2022
1222
ХАЙРУЛЛИН и др.
синаптическом образовании. Результаты экспе-
риментов показывают, что в
«быстрых» и
«медленных» мышцах мыши в условиях БС вклю-
чаются различные механизмы адаптации. Сенси-
билизация разнонаправленно влияет на силу со-
кращения различных мышц in vitro. Если у «быст-
рой» мышцы она незначительно снижалась, то у
«медленной» мышцы существенно возрастала.
Сравнение силы сокращения до и после апли-
кации АТФ демонстрирует работу пуринергиче-
ских механизмов, которые существенно различа-
ются у разных мышц, как у контрольных, так и у
сенсибилизированных животных.
Невысокая лабильность силы сокращения
«медленной» мышцы в условиях электростимуля-
Рис. 2. Сила сокращений m. soleus мыши, вызванных
ции до и после воздействия АТФ у контрольных и
электрической стимуляцией, интактных («Норма»,
сенсибилизированных мышей свидетельствует о
n = 14) и сенсибилизированных («БС», n = 10) живот-
малом вовлечении пуринергических механизмов
ных в отсутствие и в присутствии АТФ (100 мкМ) и
пресинаптической мембраны в процессах адапта-
сурамина (100 мкМ). Результаты представлены в виде
ции этой мышцы к аллергии.
M ± m в % от исходных величин, принятых за 100%;
Соотношение результатов, полученных на
* - р < 0.05 от контроля в норме, # - р < 0.05 относи-
тельно эффектов в норме.
двух моделях инициации сокращения - электро-
стимуляцией и холиномиметиком [3] позволяет
отделить участие пост- и пресинаптической мем-
471.1 ± 16 мг (n = 14, p < 0.05). У сенсибилизиро-
браны в процессах адаптации. В частности, у
ванных животных до 568.7 ± 25.5 мг (n = 10,
«медленной» мышцы в условиях аллергической
p > 0.05, рис. 2). Различия в АТФ-опосредован-
перестройки показана работа АТФ-зависимых
ном потенцировании силы сокращений до 14.5%
механизмов постсинаптической мембраны.
в норме и до 8.6% при сенсибилизации не носят
статистически достоверного характера. Инкуба-
«Быстрая» мышца в условиях электростимуля-
ция мышцы с сурамином (100 мкМ) полностью
ции незначительно снижала силу своего сокра-
предупреждала влияние АТФ на силу сокраще-
щения, что обеспечивается механизмами преси-
наптической мембраной. «Медленная» мышца,
ний.
напротив, в условиях электростимуляции суще-
Таким образом, у m. EDL в процессе сенсиби-
ственно увеличивает силу своего сокращения.
лизации подключаются пуринергические меха-
Но адаптация в условиях развивающейся аллер-
низмы адаптации в пресинаптическом звене сиг-
гии реализуется за счет работы постсинаптиче-
нализации. В противоположность этому, у m. so-
ской мембраны и АТФ-зависимые механизмы
leus в сходных структурах данные механизмы
пресинаптической мембраны в адаптации не
задействованы незначительно либо вовсе не во-
участвуют.
влекаются.
Полученные методом непрямой электрости-
муляции данные дополняют общую схему изме-
ОБСУЖДЕНИЕ
нений синаптических структур различных двига-
Существуют данные, подтверждающие нали-
тельных мышц, что обеспечивает развитие слож-
чие постсинаптических и пресинаптических эф-
ных механизмов адаптации при аллергической
фектов пуринов [8]. Определенную сложность в
перестройке организма.
исследования вносит тот факт, что в физиологи-
ческих условиях влияние АТФ и метаболитов в
ВЫВОДЫ
нервно-мышечном синапсе минимально, но ста-
новится более выраженным в некоторых патофи-
Механизмы адаптации у «быстрых» и «медлен-
зиологических ситуациях, таких как стресс [9],
ных» локомоторных мышц мыши в условиях ал-
гипогравитация [10], травмы [11], ишемия или ги-
лергической перестройки организма имеют су-
потермия [6, 7, 12, 13]. Кроме того, существуют
щественные различия. В условиях электростиму-
данные, демонстрирующие разнонаправленность
ляции увеличение силы сокращения m. EDL
действия АТФ в нервно-мышечном синапсе раз-
in vitro сенсибилизированной мыши после влия-
ных типах двигательных единиц.
ния экзогенной АТФ в сравнении с контролем
Модель электростимуляционной механомио-
более выражено. Адаптация «быстрой» мышцы в
графии позволяет анализировать процессы в пре-
описанных условиях реализуется через пуринер-
БИОФИЗИКА том 67
№ 6
2022
ПУРИНЕРГИЧЕСКИЕ МЕХАНИЗМЫ АДАПТАЦИИ
1223
гические механизмы пресинаптической мем-
2. D. Shrikrishna, R. J. Tanner, J. Y. Lee, et al., Chest,
браны.
146 (4), 932 (2014).
3. A. Y. Teplov, S. N. Grishin, M. A. Mukhamedyarov,
et al., Exp. Physiol., 94 (2), 264 (2009).
ФИНАНСИРОВАНИЕ РАБОТЫ
4. А. Ю. Теплов, А. М. Фархутдинов, О. В. Теплов
Работа выполнена в рамках программы
и др., Бюл. эксперим. биологии и медицины, 150
«Стратегическое академическое лидерство»
(9), 262 (2010).
(ПРИОРИТЕТ-2030) Казанского федерального
университета, а также за счет средств субсидии,
5. L. P. Boulet and P. M. O'Byrne, New Engl. J. Med.,
выделенной Казанскому государственному меди-
372 (7), 641 (2015).
цинскому университету Минздрава России на
6. A. U. Ziganshin, A. E. Khairullin, V. V. Zobov, et al.,
проведение научных исследований в рамках Про-
Muscle & Nerve, 55 (3), 417 (2017).
граммы развития Университета.
7. A. U. Ziganshin, A. E. Khairullin, A. Y. Teplov, et al.,
КОНФЛИКТ ИНТЕРЕСОВ
Muscle & Nerve, 59 (4), 509 (2019).
Авторы заявляют об отсутствии конфликта
8. A. U. Ziganshin, A. E. Khairullin, C. H. V. Hoyle, and
интересов.
S. N. Grishin, Int. J. Mol. Sci., 21, 6423 (2020).
9. A. Ziganshin, R. Kamaliev, A. Gabdrakhmanov, et al.,
Int. J. Pharmacol., 14 (8), 1198 (2018).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
10. A. E. Khairullin, S. N. Grishin, and A. A. Eremeev,
Все манипуляции с экспериментальными жи-
Biophysics, 64 (5), 828 (2019).
вотными проведены в соответствии с Европей-
ской конвенцией по защите позвоночных живот-
11. A. E. Khairullin, D. V. Efimova, V. A. Markosyan et al.,
ных, используемых в научных исследованиях.
Biophysics, 66 (3), 483 (2021).
12. A. E. Khairullin, A. Y. Teplov, S. N. Grishin, et al.,
СПИСОК ЛИТЕРАТУРЫ
Biophysics, 64, 812 (2019).
1. R. Kjøbsted, N. Munk-Hansen, J. B. Birk, et al., Dia-
13. A. E. Khairullin, A. U. Ziganshin, and S. N. Grishin,
betes, 66 (3), 598 (2017).
Biophysics, 65, 858 (2020).
Purinergic Mechanisms of Adaptation of Different Types of Motor Units
in the Conditions of Allergic Reorganization
A.E. Khairullin*, **, A.Yu. Teplov*, S.N. Grishin*, and A.U. Ziganshin*
*Kazan State Medical University, ul. Butlerova 49, Kazan, 420012 Russia
**Kazan Federal University, Kremlevskaya ul. 18, Kazan, 420008 Russia
The role for presynaptic signaling was found in the processes of adaptation to allergies during isometric con-
tractions of mouse EDL (fast) and soleus (slow) muscles evoked by electrical stimulation and mechanomy-
ography. Differences in the dynamics of functional characteristics of muscles under the influence of exoge-
nous ATP in intact and egg albumin-sensitized animals demonstrate that purinergic mechanisms are involved
more in the development of adaptive changes in the presynaptic side of the "fast" muscle than in the «slow»
muscle. In the process of protein sensitization, the complex work of ATP-dependent mechanisms of the post-
synaptic membrane and the presynaptic membrane ensures an adequate reorganization of the contractile
function of locomotor muscles in case of allergy and these mechanisms differ significantly in different mus-
cles.
Keywords: protein sensitization, allergy, ATP, mouse, m. EDL, m. soleus
БИОФИЗИКА том 67
№ 6
2022