БИОФИЗИКА, 2022, том 67, № 6, с. 1232-1235
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 577.3
ГИПЕРКАЛЬЦИЕВАЯ МОДЕЛЬ ДЛЯ ВЫЯВЛЕНИЯ
ПОСТСИНАПТИЧЕСКИХ ЭФФЕКТОВ АТФ
© 2022 г. А.Е. Хайруллин*, **, #, С.Н. Гришин*, А.Ю. Теплов*, А.А. Еремеев**,
Т.В. Балтина**, А.У. Зиганшин*
*Казанский государственный медицинский университет, ул. Бутлерова, 49, Казань, 420012, Россия
**Казанский федеральный университет, ул. Кремлёвская, 18, Казань, 420008, Россия
#E-mail: khajrulli@yandex.ru
Поступила в редакцию 27.06.2022 г.
После доработки 27.06.2022 г.
Принята к публикации 11.07.2022 г.
Выявление пре- и постсинаптических эффектов АТФ методологически весьма сложная задача. Ра-
нее мы использовали технику карбахолин-индуцированных сокращений для оценки вовлеченно-
сти Р2-сигнализации в процессы синаптической передачи. Поиск моделей, позволяющих фикси-
ровать постсинаптическое звено пуринергической сигнализации в условиях электрической стиму-
ляции привел нас к идее купировать пресинаптический сегмент АТФ-опосредованной модуляции.
В экспериментах in vitro на выделенных нервно-мышечных препаратах m. soleus и m. EDL крысы
методом изометрической электростимуляционной механомиографии выявлены постсинаптиче-
ские эффекты АТФ на фоне гиперкальциевой среды. Так, влияние АТФ на фоне повышенного
содержания Са2+ проявилось в полуторном усилении сокращения на m. soleus и угнетении сократи-
мости m. EDL, что соотносилось с данными, полученными ранее на карбахолин-вызванных со-
кращениях. Нами продемонстрированы АТФ-зависимые процессы, локализованные в постсинап-
тическом сегменте, способные вносить существенный вклад в реализацию механизмов адаптации в
условиях гипотермии.
Ключевые слова: нервно-мышечный синапс, гиперкальциевая модель, АТФ, сурамин, постсинаптические
эффекты.
DOI: 10.31857/S0006302922060205, EDN: LLXPKJ
Важная роль ионов кальция в регуляции функ-
ных отделах нервной терминали лягушки [5]. Из-
циональной активности почти всех клеток и тка-
менение амплитуды Са2+-транзиента отражает
ней неоспорима. В покое концентрация свобод-
изменение концентрации свободных ионов каль-
ного кальция в цитоплазме крайне мала - при-
ция внутри терминали [6], и ее изменение при
мерно в двадцать тысяч раз ниже, чем во
действии АТФ может свидетельствовать о влия-
внеклеточной среде. Повышение содержания
нии этого пурина на активность пресинаптиче-
ионов кальция до 10-6-10-5 М запускает каскад
ских кальциевых каналов. На нервной терминали
биохимических реакций, в результате которых,
функционируют несколько типов потенциал-за-
например, может произойти экзоцитоз медиато-
висимых кальциевых каналов [7].
ра в синаптическую щель из нервного окончания.
В мионевральных синапсах теплокровных вы-
К настоящему времени безусловно доказано,
является не только пресинаптический, но и пост-
что в нервно-мышечном соединении АТФ моде-
синаптический эффект АТФ [8-10]. При этом в
лирует амплитуду многоквантовых токов, акти-
синапсе «быстрой» мышцы эффект схож по знаку
вируя пресинаптические Р2Y-рецепторы [1-3].
с негативным постсинаптическим, тогда как в
Угнетающее действие АТФ на амплитуду преси-
«медленных» двигательных единицах он, наобо-
наптических токов может быть обусловлено из-
рот, потенцирующий. Является ли постсинапти-
менением активности кальциевых каналов, вход
ческое модулирующее действие АТФ в мио-
кальция через которые запускает процесс экзоци-
невральных синапсах теплокровных таким же
тоза синаптических везикул. Действительно,
Са2+-зависимым?
АТФ обратимо снижала Са2+-ток в перисинапти-
МЕТОДЫ
ческом отделе аксона [4] и уменьшала амплитуду
Исследования проводили на мышечных пре-
Са2+-транзиента, зарегистрированного в различ-
паратах белых лабораторных крыс-самцов массой
1232
ГИПЕРКАЛЬЦИЕВАЯ МОДЕЛЬ
1233
140-180 г, которых содержали в группах по три-
Далее температуру последовательно снижали до
пять особей с водой и пищей ad libitum. Животных
34, 30, 26, 22, 18 и 14°С. При каждой температуре
погружали в наркоз, вводя внутрибрюшинно эта-
добавляли АТФ и оценивали сократительные от-
минал натрия в дозе 40 мг/кг, обескровливали и
веты мышцы через 10 мин после аппликации. Да-
выделяли на задних конечностях m. soleus (кам-
лее ткань инкубировали с сурамином в течение
баловидную мышцу) и m. EDL (длинный разги-
20 мин с последующим добавлением АТФ. Тем-
батель пальцев). Выделенные мышцы фиксиро-
пературу раствора регулировали с помощью водя-
вали вертикально, присоединяя один конец к
ного насоса TE-8A (Techne, Великобритания),
датчику механической активности, и погружали в
быстрое снижение температуры жидкости в водя-
ванночки объемом 10 мл, заполненные раство-
ном насосе проводили добавлением льда.
ром Кребса следующего состава (в мМ): NaCl -
Результаты механомиографических экспери-
118.0, KCl - 4.75, CaCl2 - 2.5, NaHCO3 - 24.8,
ментов на m. soleus и m. EDL крысы анализирова-
KH2PO4 - 1.18, MgSO4·7H2O - 1.18, глюкоза - 11,
ли с применением метода ANOVA. За достовер-
рН 7.4, t = 37.0 ± 0.5°C. Термостат поддерживал
ный принимали уровень значимости менее 0.05.
заданное значение температуры. Мышцы были
Экспериментальные данные представлены как
растянуты начальной нагрузкой в 1 г, далее остав-
среднее арифметическое ± стандартная ошибка
лялись в покое на 30 мин для привыкания к среде.
среднего (n - число нервно-мышечных препара-
тов для механо-миографических экспериментов).
Сокращения регистрировали сначала в нор-
мальном растворе Кребса, а потом в растворе с
повышенным содержанием ионов кальция
РЕЗУЛЬТАТЫ
(7.2 мМ).
Электростимуляцию проводили путем стиму-
При увеличении концентрации внеклеточного
ляции культи нерва, которую помещали в сакшн-
кальция до 7.2 мМ сила сокращения «медленной»
электрод оригинальной конструкции. Для раз-
и «быстрой» двигательных единиц достоверно не
дражения использовали стимулятор MultiStim
изменилась. Так, при 37°С амплитуда сокраще-
D330 (Digitimer Ltd, Великобритания). Сокраще-
ний составила 102 ± 8.2% (n = 11, р > 0.05) для m.
ния мышц вызывали стимуляцией прямоуголь-
soleus и 104 ± 3.9% (n = 11, р > 0.05) для m. EDL от-
ными импульсами частотой 0.1 Гц, длиной 0.5 мс
носительно сокращений при нормальном содер-
и амплитудой 10 В в течение 2 мин. Силу сокра-
жании кальция [12, 13].
щений регистрировали изометрическим датчи-
АТФ в концентрации 100 мкМ при 37°С никак
ком механической активности FSG-01 (Linton,
достоверно не модифицировала силу сокращения
Великобритания), аналоговый сигнал оцифровы-
камбаловидной мышцы (95.8 ± 5.1% от исходных
вали и обрабатывали с помощью системы сбора
значений до подачи агента, n = 11, р > 0.05).
данных Biopack MP100WSW (Biopack, США).
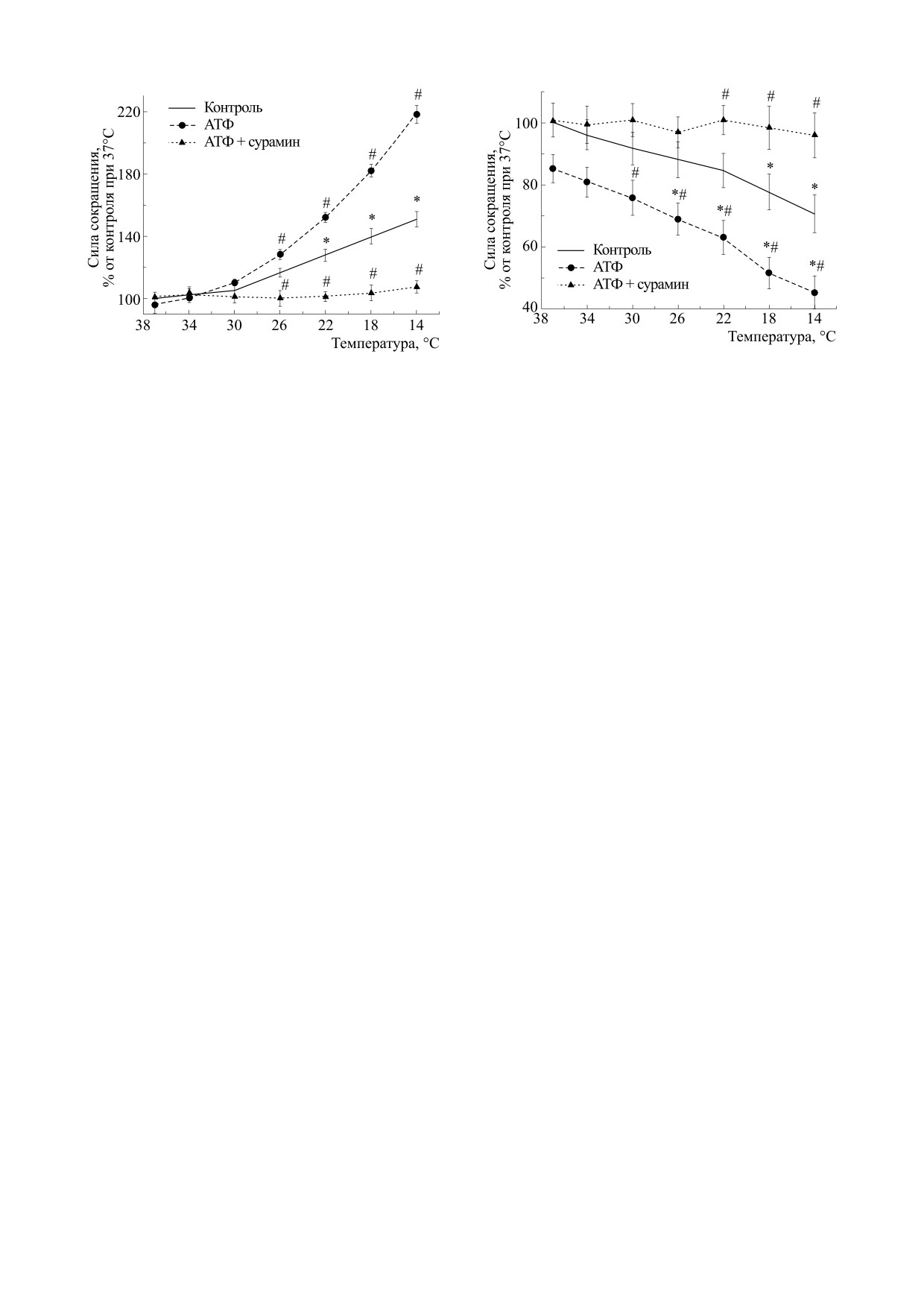
При понижении температуры омывающего
Средние значения всех сокращений, полученных
камбаловидную мышцу раствора эффект экзо-
в течение 2 мин (12 ответов), обрабатывали как
генной АТФ в концентрации 100 мкМ проявлял-
один результат [11]. Сократительные ответы рас-
ся в экспоненциальном повышении силы сокра-
считывали в процентах относительно исходных
щения вплоть до полуторного увеличения при
результатов, полученных в начале эксперимента.
14°С. Так, при этой температуре на 10-й минуте
Через 30 мин после фиксирования ткани про-
сила сокращения составила 214.6 ± 5.7% (n = 12,
водили контрольную стимуляцию мышц дважды
р < 0.05) от значения этого параметра до подачи
с интервалом в 5 мин, удостоверившись в ста-
АТФ при 37°С (рис. 1).
бильности сократительных ответов, начинали
экспериментальные процедуры.
Неселективный антагонист Р2-рецепторов су-
рамин в концентрации 100 мкМ отменял не толь-
В ванночку с раствором Кребса добавляли
ко потенцирующий эффект экзогенной АТФ, но
100 мкМ АТФ и оценивали сократительные отве-
и гипотермия-ассоциированные изменения со-
ты мышцы через 10 мин. Далее ткань инкубиро-
кратимости m. soleus.
вали с неселективным антагонистом Р2-рецепто-
ров сурамином (100 мкМ) в течение 20 мин с по-
При увеличении до 7.2 мМ концентрации вне-
следующим добавлением АТФ и вновь
клеточного кальция АТФ в концентрации
регистрировали сократительные ответы.
100 мкМ при 37°С угнетала силу сокращения
Все полученные данные рассчитывали в про-
мышцы длинного разгибателя пальцев
центах относительно исходных результатов, по-
(85.2 ± 5.6% от исходных значений до подачи
лученных в начале эксперимента при 37°С.
АТФ, n = 11, р > 0.05).
Влияние температуры на сократительную ак-
При понижении температуры омывающего
тивность m. soleus и m. EDL оценивали в экспе-
исследуемую мышцу раствора проявлялось уси-
риментах с АТФ и сурамином при инициирова-
ление ингибиторного эффекта экзогенной АТФ в
нии сокращения как электрическим полем, так и
концентрации 100 мкМ. Так, при этой температу-
аппликацией карбахолина при температуре 37°С.
ре на 10-й минуте сила сокращения составила
БИОФИЗИКА том 67
№ 6
2022
1234
ХАЙРУЛЛИН и др.
Рис. 1. Эффекты АТФ при повышенной концентра-
Рис. 2. Эффекты АТФ при повышенной концентрации
ции Са2+ (7.2 мМ) на силу вызванных электрическим
Са2+ (7.2 мМ) на силу вызванных электрическим током
током сокращений m. soleus в контроле и при аппли-
сокращений m. EDL в контроле и при аппликации
кации сурамина в концентрации 100 мкМ при раз-
сурамина в концентрации 100 мкМ., при различных
личных температурных режимах; n = 8 - 14; * - р <
температурных режимах; n = 8 - 14; * - р < 0.05 от
< 0.05 от эффекта при 37°С; # - р < 0.05 от контроля.
эффекта при 37°С; # - р < 0.05 от контроля.
43.1 ± 4.6% (n = 12, р < 0.05) от значения этого па-
мых механизмов в различных отделах мио-
раметра до подачи АТФ (рис. 2).
неврального соединения. Использованная экспе-
Сурамин (100 мкМ) в инкубационной среде
риментальная модель позволяет выявлять
предупреждал не только влияние АТФ на мышцу,
преимущественно постсинаптические эффекты
пуринов, которые становятся более выражены в
но и частично ингибиторный эффект гипотермии
на сокращения m. EDL.
условиях гипотермии [16].
Существует данные, что гипотермия угнетает
кальциевый ток через L-тип каналов [17]. Извест-
ОБСУЖДЕНИЕ
но, что повышение концентрации внеклеточного
кальция приводит к ряду пресинаптических эф-
Высвобождение нейромедиатора из нервных
фектов, в том числе и к устранению пресинапти-
окончаний позвоночных модулируется пурина-
ческого ингибиторного действия АТФ [2, 8, 18]. С
ми: действуя через пуриновые рецепторы, они из-
другой стороны, не обнаружено какого-либо зна-
меняют квантовый состав [2-6]. Но общеизвест-
но, что при этом как квантовый состав, так и ки-
чительного действия Са2+ на постсинаптические
нетика секреции нейротрансмиттера напрямую
холинорецепторы [19].
зависят от уровня [Ca2+]i в пресинаптических
В наших экспериментах при повышенном со-
держании внеклеточного кальция при обычной
окончаниях [14, 15].
электростимуляции воспроизводился потенци-
Нами было показано в периневральном отве-
рующий эффект АТФ, который можно наблюдать
дении, что именно АТФ, а не его метаболит аде-
при базовом перфузирующем растворе только
нозин (как считалось ранее) ингибирует кальцие-
при карбахолин-вызванных сократительных от-
вый вход в терминаль мотонейрона [2-4]. АТФ
ветах [12, 13]. Это косвенно подтверждает постси-
оказывает пресинаптическое ингибиторное дей-
наптический характер действия данного пурина.
ствие на квантовый состав как через активацию
Как известно, АТФ кроме выполнения в организ-
фосфолипазы А2, так и посредством синтеза пе-
ме роли макроэрга является комедиатором си-
роксида водорода [2]. В наших экспериментах по-
наптической передачи [4].
следовательные продукты активации фосфоли-
Если при нормотермии АТФ не оказывает зна-
пазы А2 арахидоновая кислота и простагландин
чимого эффекта на силу сокращения «медлен-
Е2 так же, как и АТФ, на одну шестую снижали
ной» мышцы, то гипотермия приводила к значи-
амплитуду кальциевой составляющей.
тельному приросту к силе сокращения и потен-
цирующему действию АТФ.
В представленной работе показана возмож-
ность выявления постсинаптических эффектов
Различие динамики показателей, характеризу-
АТФ в условиях непрямой электростимуляции на
ющих постсинаптическое звено демонстрирует
фоне гиперкальциевой среды. Дана дифференци-
комплексную картину участия пуринов в адапта-
рованная оценка степени участия АТФ-зависи-
ции мионеврального соединения к гипотермии.
БИОФИЗИКА том 67
№ 6
2022
ГИПЕРКАЛЬЦИЕВАЯ МОДЕЛЬ
1235
ВЫВОДЫ
2.
E. Sokolova, S. Grishin, A. Shakirzyanova, et al., Eur.
J. Neurosci., 18, 1254 (2003).
Приведенные в данной статье данные опреде-
3.
С.Н. Гришин и А. У. Зиганшин, Биол. мембраны,
ленно доказывают, что гиперкальциевая модель
30, (4), 243 (2013).
может использоваться для вычленения постси-
4.
A. U. Ziganshin, C. H. V. Hoyle, A. E. Khairullin, and
наптических эффектов АТФ в работах на нервно-
S. N. Grishin, Int. J. Mol. Sci., 21 (17), 6423 (2020).
мышечных препаратах. Верификация данной мо-
5.
S. Grishin, A. Shakirzyanova, and A. Giniatullin, Eur.
дели как раз и дала доказательства того, что пост-
J. Neurosci., 21, 1271 (2005).
синаптические эффекты АТФ - как потенцирую-
6.
E. Khaziev, A. Golovyahina, and E. Bukharaeva, Bio-
щий в «медленной» мышце, так и ингибирующий
NanoSci, 7, 254 (2017).
в «быстрой» мышце -являются кальций-незави-
7.
A. N. Tsentsevitsky, D. V. Samigullin, L. F. Nurullin,
симыми.
et al., in Frogs: Genetic Diversity, Neural Development
and Ecological Implications (Nova Science Publishers,
ФИНАНСИРОВАНИЕ РАБОТЫ
Inc., New York, 2014), pp. 179-194.
Работа выполнена в рамках программы
8.
С. Н. Гришин, Биол. мембраны, 31 (3), 155 (2014).
«Стратегическое академическое лидерство»
9.
A. E. Khairullin, A. Yu. Teplov, S. N. Grishin, et al.,
(ПРИОРИТЕТ-2030) Казанского федерального
Biophysics, 64, 812 (2019).
университета, а также за счет средств субсидии,
10.
A. E. Khairullin, A. U. Ziganshin, and S. N. Grishin,
выделенной Казанскому государственному меди-
Biophysics, 65, 858 (2020).
цинскому университету Минздрава России на
11.
R. A. Eshpay, A. E. Khairullin, R. G. Karimova, et al.,
проведение научных исследований в рамках Про-
Genes and Cells, 10 (4), 123 (2015).
граммы развития Университета.
12.
A. U. Ziganshin, A. E. Khairullin, A. Y. Teplov, et al.,
Muscle & Nerve ,59 (4), 509 (2019).
КОНФЛИКТ ИНТЕРЕСОВ
13.
A. U. Ziganshin, A. E. Khairullin, V. V. Zobov, et al.,
Muscle & Nerve, 55 (3), 417 (2017).
Авторы заявляют об отсутствии конфликта
14.
B. Katz and R. Miledi, Proc. Roy. Soc. Lond. B. Biol.
интересов.
Sci., 16 (161), 496 (1965).
15.
D. Samigullin, E. A. Bukharaeva, E. E. Nikolsky, and
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
F. Vyskočil, Physiol. Res., 54 (1), 129 (2005).
Все манипуляции с экспериментальными жи-
16.
A. E. Khairullin, A. U. Ziganshin, and S. N. Grishin,
вотными проведены в соответствии с Европей-
Biochemistry (Moscow). Suppl. Ser. A: Membrane and
ской конвенцией по защите позвоночных живот-
Cell Biology, 11 (1), 1 (2017).
ных, используемых в научных исследованиях.
17.
A. Findsen, J. Overgaard, and T. H. Pedersen, J. Exp.
Biol., 219, 2340 (2016).
18.
G. Burnstock, T. R. Arnett, and I. R. Orriss, Purinergic
СПИСОК ЛИТЕРАТУРЫ
Signal, 9 (4), 541 (2013).
1. J. G. Nicholls, A. R. Martin, B. G. Wallace, and
19.
R. E. Poage and S. D. Meriney, Physiology & Behavior,
P. A. Fuchs, Sinauer Associates, Inc. 672 (2001).
77 (4-5), 507 (2002).
A High Calcium Level-Based Model for Identifying Postsynaptic Effects of ATP
A.E. Khairullin*, **, S.N. Grishin*, A.Yu. Teplov*, A.A. Eremeev**,
T.V. Baltina**, and A.U. Ziganshin*
*Kazan State Medical University, ul. Butlerova 49, Kazan, 420012 Russia
**Kazan Federal University, Kremlevskaya ul. 18, Kazan, 420008 Russia
To identify the pre- and postsynaptic effects of ATP is indeed a methodological challenge. In our previous
study, the role of P2 receptor signaling in synaptic transmission processes was evaluated using carbachol-in-
duced skeletal muscle contractions. The search for models that can record the postsynaptic side of purinergic
signaling during the application of electrical stimulation led to the idea of controlling the presynaptic terminal
of ATP-mediated modulation. In in vitro experiments, the electromyograms and mechanomyograms during
isometric contractions of isolated nerve-muscle preparations of rat soleus and extensor digitorum longus
(EDL) muscles revealed postsynaptic effects of ATP in presence of high intracellular calcium level. Thus, the
effects of ATP in presence of increased Ca2+ content were seen through contraction of soleus muscles that
started to contract quicker by half and inhibition of contractility of EDL muscles; it was in accord with the
data obtained earlier on carbachol-induced contractions. We have demonstrated ATP-dependent processes
in the postsynaptic side that may contribute significantly to adaptation mechanisms in hypothermia.
Keywords: neuromuscular synapse, hypercalcium model, ATP, suramin, postsynaptic effects
БИОФИЗИКА том 67
№ 6
2022