БИОФИЗИКА, 2022, том 67, № 6, с. 1269-1276
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 577.354
МОДЕЛЬ ЭФФЕКТА БРОКА-ЗУЛЬЦЕРА
© 2022 г. С.И. Ляпунов*, #, И.И. Шошина**, ***, ##
*Институт общей физики им. А.М. Прохорова РАН, ул. Вавилова, 38, Москва, 119991, Россия
#E-mail: dc.cetsil@gmail.com
**Санкт-Петербургский государственный университет,
Университетская наб., 7-9, Санкт-Петербург, 199034, Россия
##E-mail: shoshinaii@mail.ru
***Сибирский федеральный университет, Свободный просп., 82/1, Красноярск, 660041, Россия
Поступила в редакцию 23.12.2021 г.
После доработки 07.02.2022 г.
Принята к публикации 23.08.2022 г.
Феномен Брока-Зульцера - один из аспектов проблемы субъективной деформации реального
мира, загадки сознания. Эффект Брока-Зульцера проявляется в превышении субъективной
яркости при коротких длительностях стимула. Несмотря на долгую историю изучения, эффект не
имеет удовлетворительной теории механизма его возникновения. В работе предложена модель
формирования эффекта Брока-Зульцера, основанная на теории треморного модуляционного
сигнала в зрительной системе. Также демонстрируется возможность использования эффекта
Брока-Зульцера в качестве маркера и инструмента для оценки функционального состояния,
адаптации и дезадаптации к изменяющимся условиям окружающей среды. Понимание природы
явления имеет исключительное теоретическое и практическое значение для различных областей
знаний.
Ключевые слова: эффект Брока-Зульцера, модель треморного модуляционного сигнала, яркость,
яркость адаптации, теория двух путей, маркер функционального состояния.
DOI: 10.31857/S0006302922060254, EDN: LMRCZK
кость тестового поля - 0.8, 5.3, 11.2, 22.4, 32.6 и
Феномен Брока-Зульцера - один из аспектов
проблемы субъективной деформации реального
40.7 кд/м2; длительность постоянного поля срав-
мира, загадки сознания
[1-10]. А. Брока и
нения 2.0 с. При яркости тестового поля менее
Д. Зульцер [1] сообщили об этом эффекте в 1902 г.
Авторы решали задачу уравнивания кажущейся
яркости (светлоты) световой вспышки заданной
длительности (тестовое поле) с кажущейся ярко-
стью световой вспышки продолжительностью 1-
2 с (поле сравнения) в условиях темновой адапта-
ции. Эффект состоит в превышении кажущейся
яркости тестового поля над полем сравнения
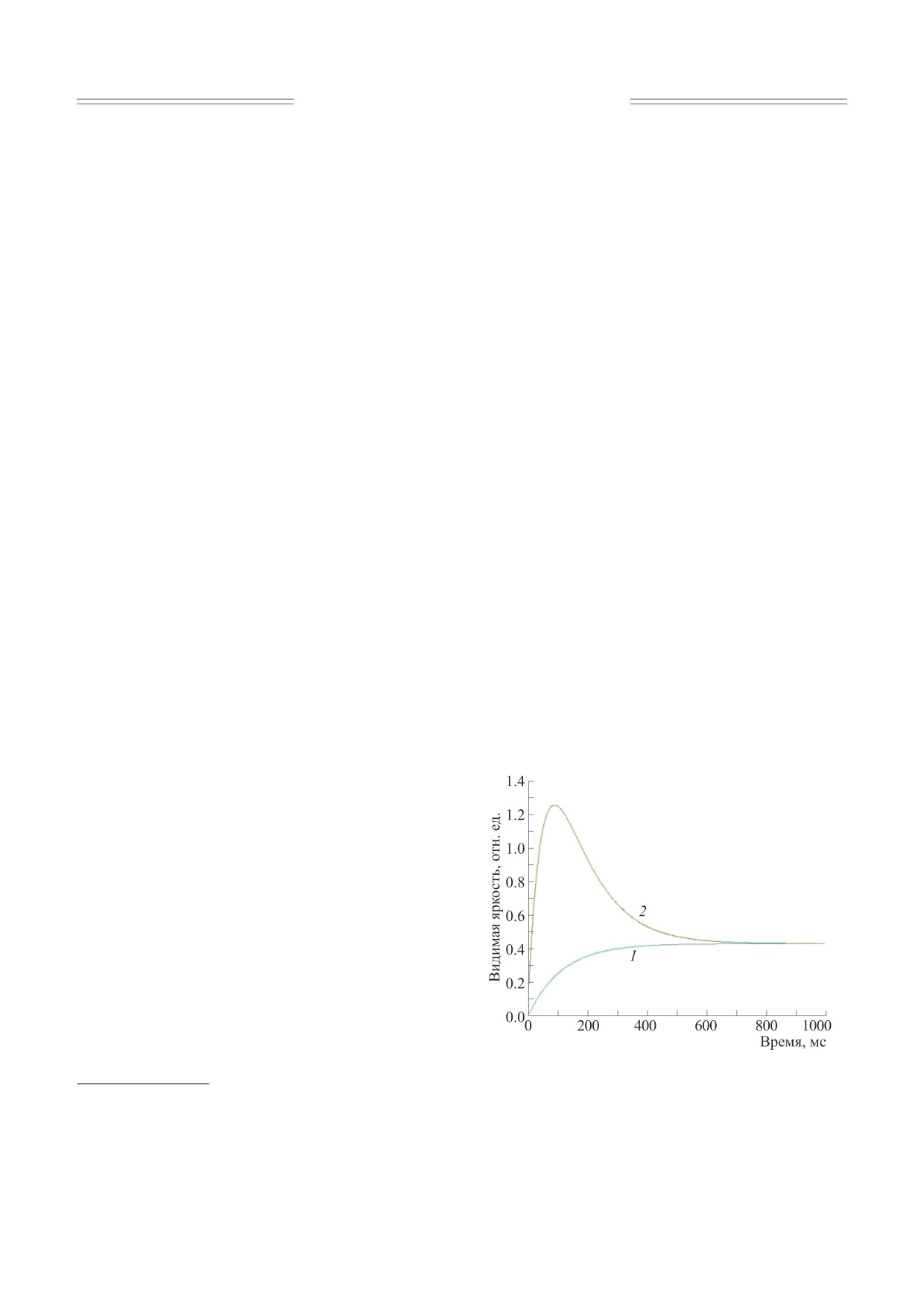
(рис. 1).
За более чем столетний период существования
с момента открытия эффект Брока-Зульцера не-
однократно исследовался и подтверждался как в
классических, так и в измененных условиях ярко-
сти адаптации, угловых размеров, цвета и формы
полей сравнения [2]. Результаты, полученные
Брока и Зульцером [1], показаны в табл. 1. для
следующих условий эксперимента: яркость адап-
тации - темнота; искусственный зрачок - 2.5 мм;
угловой размер тестового поля - 180 угл. мин; яр-
Рис. 1. Восприятие импульсных световых полей в
Сокращения: ТМС - треморный модуляционный сигнал,
соответствии с законом Блонделя-Рэя (кривая 1) и
ГК - ганглиозные клетки.
эффектом Брока-Зульцера (кривая 2).
1269
1270
ЛЯПУНОВ, ШОШИНА
Таблица 1. Результаты, полученные Брока и Зульцером
Эффект/Яркость тестового поля, кд/м2
0.8
5.3
11.2
22.4
32.6
40.7
Коэффициент превышения видимой
1.2
1.9
2.0
4.6
4.9
5.5
яркости над установившимся значением
Длительность импульса тестового поля,
соответствующая максимальному
125
100
62
45
40
32
превышению, мс
ют соизмеримую угловую скорость, что обеспе-
0.2 кд/м2 эффект Брока-Зульцера отсутствует, а
чивает двухкоординатную модуляцию сигнала.
воспринимаемая яркость подчиняется закону
Блонделя-Рэя [2].
В качестве меры треморного модуляционного
сигнала принята нормированная разность кон-
Эффект Брока-Зульцера позволяет отметить
центрации фотореагента в фоторецепторе в край-
ряд особенностей зрительного восприятия:
них точках тремора. Из этого следует, что источ-
- действие суммы нескольких световых им-
ником ТМС являются перепады и градиенты яр-
пульсов не равно действию одного импульса сум-
кости. Постоянная яркость или малые ее
марной длительности, что характеризует нели-
изменения в пределах шума спонтанной генера-
нейность эффекта;
ции импульсов действия (спайков) ганглиозной
- воспринимаемая яркость тестового стимула
клетки или порога возбуждения нейрона дают ну-
левое значение сигнала. На перепаде или гради-
всегда определяется ощущением, которое возни-
енте яркости ТМС может существовать бесконеч-
кает в конце действия стимула, а не эффектив-
но долго. В модели полагается, что именно дли-
ным значением за время действия стимула или за
время инерции;
тельный ТМС является основанием для
автоформирования линий возбуждения, которые
- продолжительность превышения видимой
впоследствии повторяются в высших отделах
яркости над установленным значением может со-
зрительной системы. Адекватность модели под-
ставлять до 2-4 с.
тверждена для центрального и периферического
Несмотря на долгую историю изучения, фено-
зрения в большом диапазоне внешних условий -
мен не имеет удовлетворительной теории меха-
яркости адаптации (10-3-103 кд/м2), угловых раз-
низма его возникновения.
меров (1-300 угловых минут) и формы тестовых
стимулов (диски, кольца Ландольта, миры Фуко,
синусоидальные решетки), времени их предъяв-
МОДЕЛИРОВАНИЕ ЭФФЕКТА
ления (10-5-5 с). Модель не содержит поправоч-
БРОКА-ЗУЛЬЦЕРА
ных коэффициентов и функций во всем диапазо-
Проведем моделирование эффекта Брока-
не внешних условий при восприятии различных
Зульцера с позиций модели контрастной чувстви-
тестовых стимулов в пороговых и надпороговых
тельности зрительной системы, основанной на
условиях.
треморном модуляционном сигнале (далее мо-
Временные показатели эффекта Брока-Зуль-
дель ТМС), подробно изложенной в нескольких
цера определяются показателями фотохимиче-
предыдущих работах [11-14]. В основу модели
ских процессов в рецепторе. Скорость уменьше-
ТМС положен факт исчезновения восприятия
ния концентрации (разложения) фотореагента
при остановке изображения на сетчатке, полу-
пропорциональна освещенности E и текущей
ченный и подробно описанный А.Л. Ярбусом [5].
концентрации с:
Из всех движений, совершаемых глазом, в каче-
стве основных движений в модели ТМС приняты
dc
= K
Ec
(1)
тремор и дрейф. Есть две причины этого выбора.
dt
Во-первых, из всех движений, осуществляемых
Скорость увеличения концентрации (восста-
глазом, тремор и дрейф являются самыми низко-
новления) фотореагента пропорциональна кон-
скоростными (угловая скорость до 0.4 град/с) и не
центрации в неактивной фазе (1 - c):
ухудшают пространственное разрешение и кон-
трастную чувствительность зрительной системы.
dc
=K
(1 -
c
)
(2)
Это означает, что на данных угловых скоростях
2
dt
зрительная система накапливает отношение сиг-
нал/шум без потерь. Во-вторых, тремор и дрейф
Суммарный процесс, описывающий текущую
составляют пару ортогональных движений и име-
концентрацию фотореагента как балансное со-
БИОФИЗИКА том 67
№ 6
2022
МОДЕЛЬ ЭФФЕКТА БРОКА-ЗУЛЬЦЕРА
1271
стояние конкурирующих процессов распада и
приятия. Решение уравнения (3) имеет следую-
восстановления, имеет следующий вид:
щий вид:
c(t) = c0 + (c1 - c0)exp(-(K2 + K1E1)t),
(4)
dc
где концентрация c определяется по формуле
=-(K
E+K
)c+K
(3)
1
2
2
dt
K
2
c=
(5)
Числовые значения коэффициентов фотохи-
K
E+K
1
2
мической реакции вычисляются для круглого
Нормированное приращение концентрации
стимула с размером, равным одной угловой ми-
должно быть больше порога нейронного возбуж-
нуте, и постоянны во всех внешних условиях вос-
дения F:
(c
0
−c t))
(1- exp
(-(K
2
+
K
1
E t))
=
(c
0
−
c
1
)
≥
F
= const.
(6)
c
0
c
0
Переход от начального значения c0 к значению
dc(t)/c0 = dc(∞)/c0(1 - exp(-(K2 + K1E)t)),
(11)
c1 не происходит мгновенно и требует определен-
TMC(t) = TMCmax(1 - exp(-(K2 + K1E)t)),
(12)
ного времени для достижения балансного состоя-
ния конкурирующих процессов распада и восста-
SN(t) = 1 - exp(-(K2 + K1E)t).
(13)
новления. При этом время распада фотореагента
в необратимое состояние характеризуется значе-
Максимальное, предельное значение ТМС со-
нием порядка одной миллисекунды и переходные
ответствует перепаду яркости физического сти-
процессы в основном определяются временем
мула в крайних точках тремора, преобразованно-
восстановления фотореагента.
му в пространство концентрации фотореагента, а
Если соотнести приращение концентрации
функция SN(t) описывает временную динамику
для бесконечно большого и текущего времени,
накопления отношения «сигнал/шум» в зритель-
можно получить выражение для относительного
ной системе. Данная гипотеза, принятая в моде-
порога восприятия от времени предъявления сти-
ли, была убедительно подтверждена при описа-
мула. Для сравнения представлены выражение
нии порогового восприятия разнообразных
(7), полученное согласно модели ТМС, и выраже-
стимулов при неограниченном времени наблюде-
ние (8) для закона Блонделя-Рея и Блоха-Шар-
ния. Однако при рассмотрении реальных и мо-
пантье:
дельных переходных процессов обнаруживается
некоторое расхождение, которое позволяет свя-
1
K
=
,
(7)
зать их между собой и, возможно, поможет соеди-
u
(1- exp( (
2
+
1
) ))
нить биофизические экспериментальные данные
t
p
K
u
=
,
(8)
(
t
p
+ 0,21
)
где tp - время предъявления стимула.
Значение освещенности E в плоскости сетчат-
ки, создаваемой яркостью L, находим по следую-
щей зависимости:
E = πτLDp2/4f2,
(9)
где Dp - диаметр зрачка глаза, определяемый
по формуле Крауфорда:
Dp = 5 - 3th(0.4lgLa).
(10)
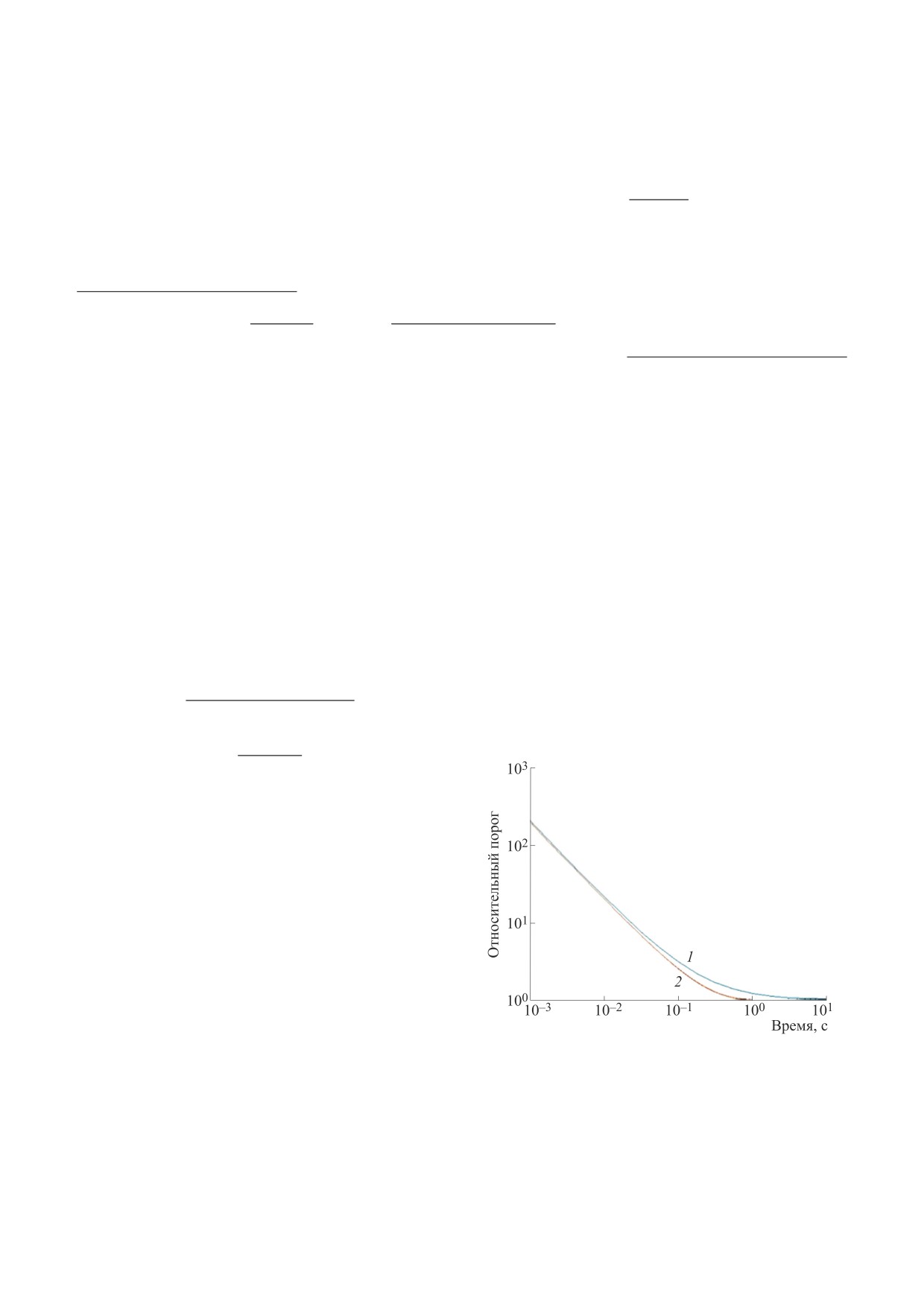
Результаты расчета зависимости относитель-
ного порога восприятия от времени предъявле-
ния стимула представлены на рис. 2.
Выражение (6) имеет важную физическую и
физиологическую трактовку. Физическая трак-
Рис. 2. Зависимость относительного порога восприятия
товка проливает свет на формирование отноше-
от времени предъявления стимула: кривая 1 - закон
ния сигнал/шум в зрительной системе. Преобра-
Блонделя-Рэя и Блоха-Шарпантье; кривая
2
-
зуем выражение следующим образом:
результаты расчета по уравнению (7).
БИОФИЗИКА том 67
№ 6
2022
1272
ЛЯПУНОВ, ШОШИНА
Представленные зависимости и иллюстрации
модели ТМС позволяют описать большое коли-
чество экспериментальных данных на уровне
«фоторецептор - ганглиозная клетка» и перейти
к моделированию эффекта Брока-Зульцера.
Рассмотрим основные выражения модели
ТМС для условий экспериментов Брока-Зульце-
ра. Концентрации с0 соответствует яркости адап-
тации Lad. Концентрация с1 соответствует ярко-
сти тестирующего поля L1. Согласно модели ТМС
сигнал формируется на границе стимула, поле
суммирования сигнала равно размеру тестового
поля N, выраженному в угловых минутах, время
накопления сигнала tau определяется частотой
тремора. Выражение для ТМС имеет следующий
вид:
Pp(t) = (c0 - c1)N(1 - exp(-(K2 + K1E1)t)).
(14)
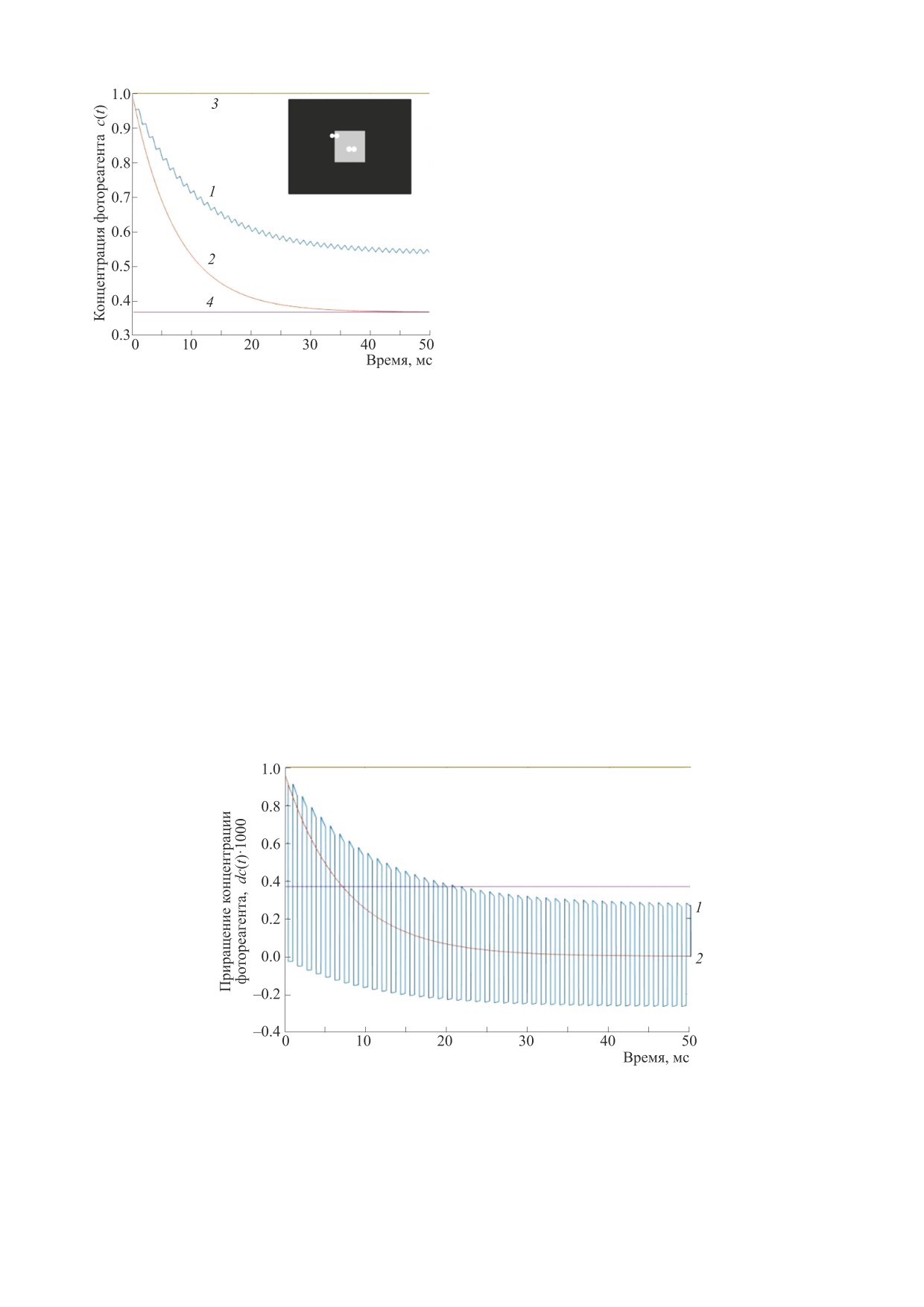
Рис. 3. Временная динамика концентрации фотореа-
гента на частоте тремора: 1 - при треморной модуляции
Поскольку при постоянной яркости ТМС ра-
на границе стимула, 2 - при треморной модуляции в по-
вен нулю, то выражение для ТМС в тестовом поле
ле стимула, 3 - уровень концентрации адаптации, 4 -
уровень концентрации стимула.
пропорционально площади (N - 2)2 и также рав-
но нулю:
и модели протекающих в сетчатке процессов с
Ps(t) = (c0 - c1)(N -2)2(1 - exp(-(K2 + K1E1)t)). (15)
контрастной чувствительностью зрительной си-
Но в тестовом поле есть временной сигнал,
стемы.
обусловленный переходным процессом от уровня
На рис. 3 представлены переходные процессы
c0 к уровню с1 (далее сигнал переходного процес-
концентрации фотореагента на границе и в поле
са). Логично предположить, что глаз не в состоя-
стимула при треморной модуляции сигнала. По-
нии определить источник сигнала и суммирует все
ложения фоторецептора на яркостном тесте в
временные изменения. Сигнал переходного про-
крайних точках тремора показаны парами кругов.
цесса равен производной от переходного процесса
умноженной на временной интервал tau и пропор-
На рис. 4 представлены результаты дифферен-
цирования данных предыдущего графика с вре-
ционален площади тестового поля (N - 2)2. Соглас-
менным шагом 0.1 мс, для наглядности усилен-
но формуле (12) выражение для сигнала переход-
ные в 1000 раз.
ного процесса с учетом функции SN(t) имеет вид:
Рис. 4. Временная динамика приращения концентрации фотореагента: 1 - при треморной модуляции на границе стимула,
2 - при треморной модуляции в поле стимула.
БИОФИЗИКА том 67
№ 6
2022
МОДЕЛЬ ЭФФЕКТА БРОКА-ЗУЛЬЦЕРА
1273
Pt(t) = (c0 - c1)(N - 2)2exp(-(K2 + K1E1)t)(K2 + K1E1)tau(1 - exp(-(K2 + K1E1)t)).
(16)
Поскольку сравнение сигналов должно проис-
гласно формуле (16)). Первым фактором, опреде-
ходить на равном временном интервале, то дан-
ляющим вес Pt, является разница в концентраци-
ный интервал принимается равным периоду тре-
ях фотореагента, соответствующая яркости адап-
морных колебаний, что для нормальной частоты
тации и яркости тестового поля. Следует
тремора 87 Гц составляет 11.5 мс:
отметить, что это не разница в яркости, а именно
tau = 1/ftr.
(17)
разница в концентрации, так как установившееся
значение концентрации в диапазоне яркости
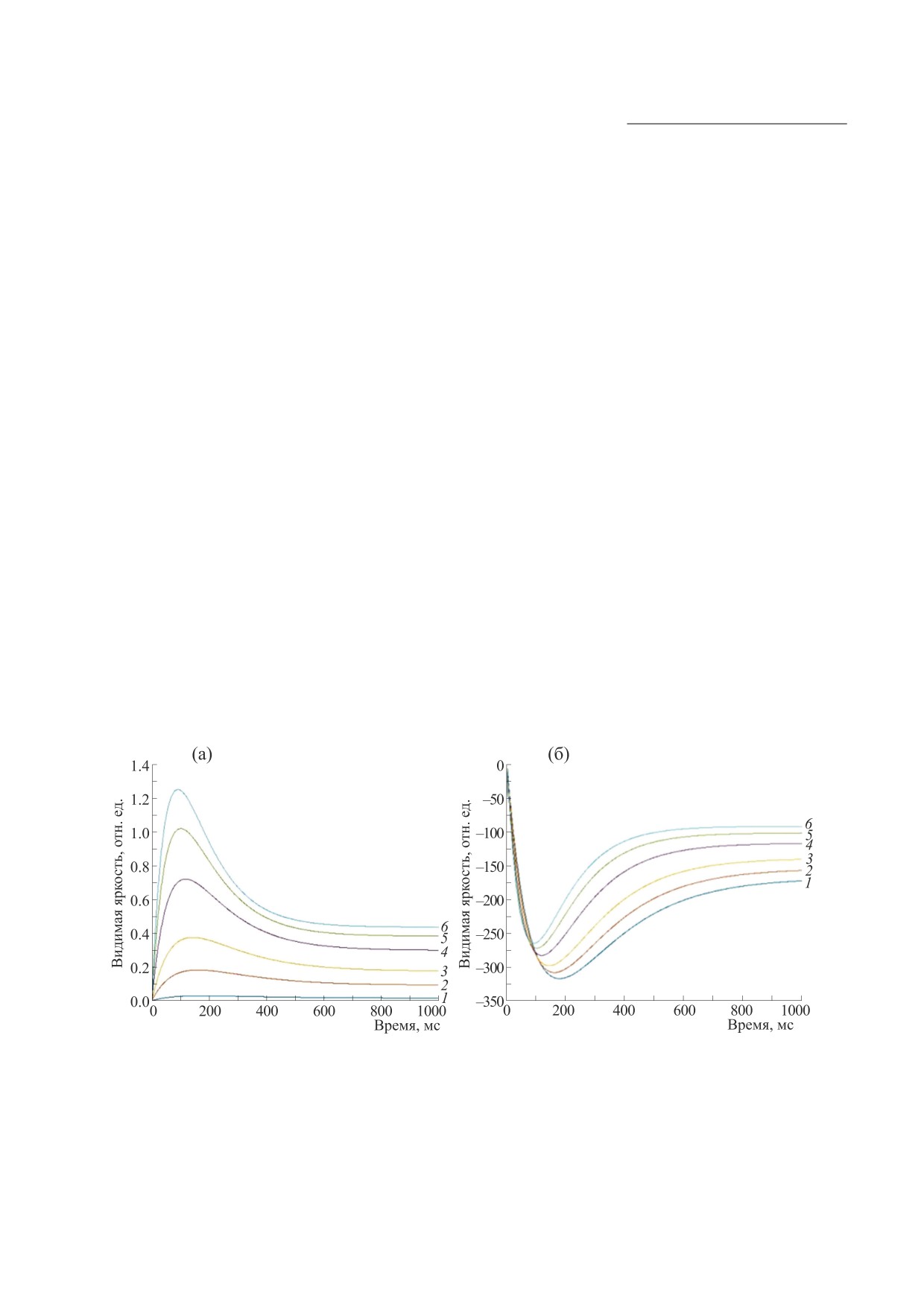
На рис. 5 представлены результаты расчетов
эффекта Брока-Зульцера на основе модели ТМС
0.001-1.0 кд/м2 практически не меняется. Следо-
по формулам (14-17). Эффект Брока-Зульцера
вательно, согласно модели ТМС, глубина адапта-
также можно наблюдать для темных тестовых по-
ции к темноте не является значимым фактором.
лей на светлом фоне (рис. 5в). Его эксперимен-
Зависимость концентрации флотационного аген-
тально наблюдал Г. Дж. Бартон с коллегами [15] в
та от яркости адаптации с учетом функции зрачка
1977 г., но не нашел объяснения данному эффек-
глаза показана на рис. 6. Границы существования
ту. В этих условиях эффект характеризуется мень-
эффекта лежат в диапазоне яркости
1.0-
шими значениями видимого снижения яркости и
1000.0 кд/м2.
незначительным смещением вправо от миниму-
мов.
Второй фактор - угловой размер тестового по-
Таким образом, с точки зрения модели ТМС
ля и поля сравнения. Результаты моделирования
эффект Брока-Зульцера определяется соотноше-
зависимости эффекта Брока-Зульцера от углово-
нием треморного модуляционного сигнала Pp,
го размера тестового поля показаны на рис. 7.
сформированного на контурах тестового поля и
При уменьшении размеров тестового поля на-
площадного сигнала переходного процесса поля
блюдается уменьшение коэффициента превыше-
сравнения Pt.
ния видимой яркости, при этом максимум кри-
вой смещается вправо. Для яркости тестового по-
Такой взгляд значительно проясняет и упро-
щает физическую природу процессов, происхо-
ля 40.7 кд/м2 значительное увеличение эффекта
дящих в зрительной системе при формировании
начинается при угловых размерах тестового поля,
эффекта Брока-Зульцера, и может внести свой
начиная с 60-90 угловых минут. По результатам
вклад в теорию зрительного восприятия. Проана-
моделирования для получения выраженного эф-
лизируем результаты моделирования. Интенсив-
фекта Брока-Зульцера следует использовать те-
ность эффекта Брока-Зульцера определяется
стовые поля с угловым размером 120-180 угловых
весом составляющей временной области Pt (со-
минут.
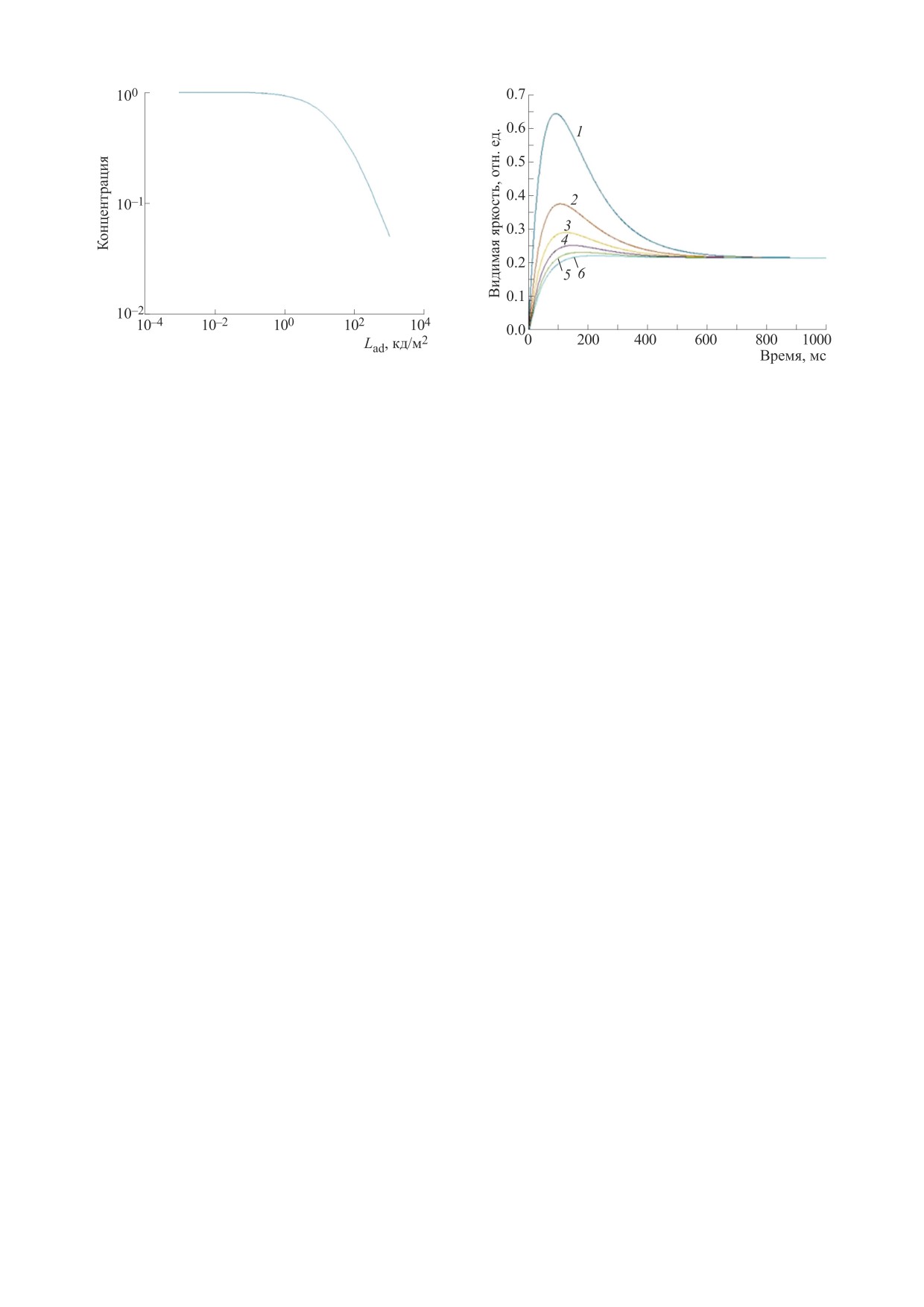
Рис. 5. Результаты моделирования эффекта Брока-Зульцера на основе модели ТМС. Зависимость эффекта Брока-
Зульцера от яркости тестового поля: (а) - яркость адаптации 0.001 кд/м2, (б) - яркость адаптации 1000 кд/м2. Яркость
тестового поля по табл. 1: кривая 1 - 0.8 кд/м2, кривая 2 - 5.3 кд/м2, кривая 3 - 11.2 кд/м2, кривая 4 - 22.4 кд/м2,
кривая 5 - 32.6 кд/м2, кривая 6 - 40.7 кд/м2.
БИОФИЗИКА том 67
№ 6
2022
1274
ЛЯПУНОВ, ШОШИНА
Рис. 6. Зависимость концентрации флотационного
Рис. 7. Зависимость эффекта Брока-Зульцера от
агента от яркости адаптации с учетом функции зрачка
углового размера тестового поля. Обозначения:
глаза.
яркость тестового поля 40.7 кд/м2; размер поля от 30
до 180 угловых минут: кривая 1 - 180 угловых минут,
кривая 2 - 150, кривая 3 - 120, кривая 4 - 90, кривая
ЭФФЕКТ БРОКА-ЗУЛЬЦЕРА КАК МАРКЕР
5 - 60, кривая 6 - 30 угловых минут.
ФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ
Зависимость эффекта Брока-Зульцера от яр-
чатого ядра таламуса (LGN) и далее до слоев 4Cα
кости, длительности, размера и пространствен-
и 6 области V1 зрительной коры [18-22]. Клетки
ной локализации тестового поля при централь-
этого пути реагируют на включение стимула, мер-
ной и периферической стимуляции представляет
цание и доминируют на периферии сетчатки, тем
интерес для обоснования результатов исследова-
самым обеспечивая периферическое зрение.
ний механизмов зрительного восприятия с пози-
ций теории двух потоков [16, 17]. Теория основа-
Парвоцеллюлярная система берет начало от
на на представлении о структурно-функциональ-
малых ГК сетчатки - Р-клеток, аналогов тониче-
ной организации сетчатки и классификации
ских Х-клеток, наиболее плотно расположенных
ганглиозных клеток (ГК). Подавляющее боль-
в центральной ямке, с небольшими рецептивны-
шинство ГК сетчатки дает начало двум основным
ми полями, низкой скоростью аксональной про-
потокам передачи информации из каудальных во
водимости, несущих информацию о красно-зеле-
фронтальные зоны коры [18]. Речь идет о магно-
ной части спектра к P-слоям латерального колен-
целлюлярной и парвоцеллюлярной системах,
чатого ядра таламуса (LGN), которые, в свою
нейроны которых формируют соответственно
очередь, проецируются на 4Cß и 6 слои V1 обла-
дорсальную (теменную) и вентральную (височ-
сти зрительной коры [18, 20-22]. Клетки P-пути
ную) систему передачи информации. Примерно
имеют низкую чувствительность к контрасту (они
70% всей популяции ГК сетчатки составляют
реагируют при насыщении контраста более 16%),
нейроны парвоцеллюлярной системы (Р-систе-
при этом они специфичны к восприятию высо-
мы) и 10% - магноклеточной системы (М-систе-
ких пространственных (выше 7,0 цикл/градус) и
мы). Свойства ГК, которые образуют эти систе-
низких временных частот [18-21].
мы, согласуются с факторами, определяющими
Отростки нейронов магноцеллюлярной систе-
возникновение эффекта Брока-Зульцера. По-
мы из первичной зрительной коры дают проек-
этому представляет интерес использование эф-
ции в основном в область V5/MT экстрастриар-
фекта Брока-Зульцера для изучения свойств и
ной коры, которая специфична к восприятию
функций этих систем. О свойствах нейронов в
движения. Проекции нейронов P-системы на-
рассматриваемых системах известно следующее.
правлены в область V4 нижней височной коры,
Магноцеллюлярная система берет начало от
детекторные нейроны которой обеспечивают
крупных ГК сетчатки (аналогов фазических
восприятие деталей и объектное зрение. Нейро-
Y-клеток) с большими рецептивными полями,
ны магносистемы, отростки которых направля-
высокой чувствительностью к контрасту (ниже
ются преимущественно к дорзальному потоку,
16%) и специфичностью для высоких временных
обрабатывают информацию о стереопсисе, кон-
и низких пространственных частот [19], которые
туре, положении в пространстве, движении и его
передают широкополосный, в основном ахрома-
направлении [23], обеспечивают пространствен-
тический сигнал на M-слои латерального колен-
ное зрение [24, 25] и глобальное описание изоб-
БИОФИЗИКА том 67
№ 6
2022
МОДЕЛЬ ЭФФЕКТА БРОКА-ЗУЛЬЦЕРА
1275
ражения [20, 24]. Нейроны парвоцеллюлярной
можность использования эффекта Брока-Зуль-
системы, которые формируют вентральный путь,
цера в качестве маркера и инструмента для оцен-
играют ведущую роль в передаче информации о
ки функционального состояния и степени рассо-
цвете, процессах различения отдельных предме-
гласования между магно- и парвоцеллюлярной
тов и деталей объектов, то есть локальном анали-
системами.
зе поля зрения [23, 24]. Подводя итог, важно от-
метить, что системы обеспечивают разный уро-
ВЫВОДЫ
вень детализации и скорости передачи
информации [25]: M-система - менее 100 мс, P-си-
Нами предложена модель формирования эф-
стема - более 100 мс.
фекта Брока-Зульцера, основанная на теории
Экспериментальные данные показывают, что
треморного модуляционного сигнала в зритель-
ной системе. Этот взгляд значительно проясняет
М- и Р-системы не являются полностью незави-
и упрощает представления о физической природе
симыми [26, 27], поэтому согласованность взаи-
модействия этих систем имеет фундаментальное
процессов, происходящих в зрительной системе
значение [28]. Эффект Брока-Зульцера может
при формировании эффекта Брока-Зульцера.
стать инструментом, который позволит не только
Показана возможность использования эффекта
Брока-Зульцера в качестве маркера и инструмен-
изучать функциональное состояние этих систем в
та оценки функционального состояния, адапта-
разных условиях, но также, что немаловажно,
служить маркером дисбаланса в их взаимодей-
ции организма к изменяющимся условиям окру-
ствии. На сегодняшний день контрастная чув-
жающей среды. Понимание природы этого явле-
ствительность нейронов этих систем в разном
ния имеет исключительное теоретическое и
практическое значение для различных областей
диапазоне пространственных частот наиболее ак-
знаний физики, философии, психологии, меди-
тивно используется для оценки состояния магно-
и парвоцеллюлярной системы.
цины и др.
Результаты исследований контрастной чув-
ствительности при адаптации к экстремальным
ФИНАНСИРОВАНИЕ РАБОТЫ
воздействиям [29] и на моделях психопатологии
Это исследование не получало какого-либо
[28, 30-31] указывают на изменение характера
специального гранта от финансирующих
взаимодействия магно- и парвоцеллюлярной си-
агентств в государственном, коммерческом или
стем. Взаимодействие магно- и парвоклеточной
некоммерческом секторах.
систем обеспечивает целостность восприятия,
как неотъемлемое условие построения мозгом
объективной внутренней картины внешнего ми-
КОНФЛИКТ ИНТЕРЕСОВ
ра, определяющей поведение. При адаптации к
Авторы заявляют об отсутствии явных и по-
экстремальным воздействиям и срыве адаптации
тенциальных конфликтов интересов, связанных с
в условиях первого психотического эпизода кон-
публикацией данной статьи.
трастная чувствительность зрительной системы
изменяется в диапазоне низких пространствен-
ных частот, к восприятию которых более специ-
СПИСОК ЛИТЕРАТУРЫ
фичны нейроны магноклеточной системы, фор-
1. A. Broca and D. Sulzer, J. Physiol. Pathol. Générale, 4,
мирующей дорзальный путь. Анализ полученных
632 (1902).
данных с позиций теории треморно-модуляцион-
ного сигнала (ТМС), которая также раскрывает
2. Р. М. Мещерский, Эффект Брока-Зульцера (Нау-
ка, М., 1985).
природу эффекта Брока-Зульцера, позволяет
предположить, что смещение баланса в сторону
3. S. M. Berman and A. L. Stewart, Biol. Cybernetics, 34,
активности магноклеточной системы приводит к
171 (1979).
увеличению поля суммирования сигнала и увели-
4. D. M. Eagleman, J. E. Jacobson, and T. J. Sejnowski,
чению коэффициента превышения видимой яр-
Nature, 428 (6985), 854 (2004).
кости, т. е. увеличению эффекта Брока-Зульце-
5. А. Л. Ярбус, Роль движений глаз в процессе зрения
ра. Другими словами, чем более выражен сдвиг в
(Наука, М., 1965).
сторону активности магноклеточной системы,
6. R. Knight, C. Mazzi, and S. Savazzi, Exp. Brain Res.,
тем выше и влево будет смещен график, отражаю-
233 (12), 3527 (2015).
щий эффект Брока-Зульцера. Таким образом,
7. T. Jaeger and C. Kraemer, Perceptual Motor Skills, 51
полученные физиологические данные [28] интер-
(3), 945 (1980).
претируются нами с позиций модели треморно-
модуляционного сигнала, что позволяет объяс-
8. A. Gorea and C. W. Tyler, Proc. Nat. Acad. Sci. USA,
нить движение точки максимальной контрастной
110 (15), 1330 (2013).
чувствительности в экстремальных условиях
9. H. Rieiro, S. Martinez-Conde, A. P. Danielson, et al.,
функционирования и продемонстрировать воз-
Proc. Natl. Acad. Sci. USA, 109 (48), 19828 (2012).
БИОФИЗИКА том 67
№ 6
2022
1276
ЛЯПУНОВ, ШОШИНА
10. V. Kalnysh and A. Shvets, Ukr. J. Occupational Health,
22. W. H. Merigan and J. H. R. Maunsell, Ann. Rev. Neu-
3 (48), 10 (2016).
rosci., 16, 369 (1993).
11. S. I. Lyapunov, J. Optical Technol., 85 (2), 100 (2018).
23. M. S. Livingston and D. H. Hubel, Science. 240, 740
12. S. I. Lyapunov, J. Optical Technol., 84 (1), 16 (2017).
(1988).
13. S. I. Lyapunov, 84 (9), 613 (2017а).
24. D. J. Calderone, M. J. Hoptman, A. Martinez, et al.,
14. S. I. Lyapunov, J. Optical Technol., 81 (6), 349 (2014).
Cereb. Cortex, 23, 1849 (2013).
15. G. J. Barton, S. Nagshineh, K.H. Ruddock, Biol. Cu-
25. E. Kaplan and E. Benardete, Progr. Brain Res., 134, 17
bern. 27 (4), 189 (1977).
(2001).
16. L. G. Ungerleider and M. Mishkin, In Analysis of Visual
26. E. H. F. de Haan, S. R. Jackson, and T. Schenk, Cor-
Behavior, Ed. by D. J. Ingle, M. A. Goodale, and
tex, 98, 1 (2018).
R. J. W. Mansfield (MIT Press, Cambridge, 1982),
27. E. Freud, M. Behrmann, and J. C. Snow, Open Mind:
pp. 549-586.
Discoveries in Cognitive Science. 4, 40 (2020).
17. M. A. Goodale and A. D. Milner, Trends Neurosci., 15
28. И. И. Шошина и Ю. Е. Шелепин, Механизмы гло-
(1), 20 (1992).
бального и локального анализа зрительной информа-
18. J. J. Nassi and E. M. Callaway, Nat. Rev. Neurosci., 10
ции при шизофрении (ВВМ, СПб., 2016).
(5), 360 (2009).
29. I. I. Shoshina, I. S. Sosnina, K. A. Zelenskiy, et al.,
19. A. M. Derrington and P. Lennie, J. Physiol., 3 (57), 219
Biophysics, 65 (4), 681 (2020).
(1984).
20. D. H. Hubel and T. N. Wiesel, J. Comp. Neurol., 146
30. I. I. Shoshina, Y. E. Shelepin, E. A. Vershinina, and
(4), 421 (1972).
K. O. Novikova, Human Physiol., 41 (3), 251 (2015).
21. E. Kaplan and R. M. Shapley, Proc. Natl. Acad. Sci.
31. I. I. Shoshina, Y. E. Shelepin, S. A. Konkina, et al.,
USA, 83, 2755 (1986).
Neurosci. Behav. Physiol., 44, 244 (2014).
A Model of the Broca-Sulzer Effect
S.I. Lyapunov* and I.I. Shoshina**, ***
*Prokhorov General Physics Institute, Russian Academy of Sciences, ul. Vavilova 38, Moscow, 119991 Russia
**Saint-Petersburg State University, Universitetskaya nab. 7-9, St. Petersburg, 199034 Russia
***Siberian Federal University, Svobodny prosp. 82/1, Krasnoyarsk, 660041 Russia
The Broca-Sulzer phenomenon is one of the aspects of the problem of individual differences in the percep-
tion of visual world, the mystery of consciousness. The Broca-Sulzer effect occurs when brightness enhance-
ment is observed under the stimulus condition over a short period. Although this effect has been studied for
a long time, there is no satisfactory theory of the occurrence mechanism. In this paper, a model for the for-
mation of the Broca-Sulzer effect based on the theory of the tremor signal modulation in the visual system
is proposed. Also, this paper demonstrates the possibility of applying the Broca-Sulzer effect as a marker and
a tool for assessing the functional state, adaptation and maladaptation to changing environmental conditions.
Understanding the nature of the phenomenon has exceptional theoretical and practical significance for var-
ious fields of knowledge.
Keywords: Broca-Sulzer effect, model of tremor modulation signal, brightness, brightness of adaptation, theory of
two pathways, marker of functional state
БИОФИЗИКА том 67
№ 6
2022