БИОФИЗИКА, 2023, том 68, № 1, с. 98-107
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 543.4 (045)
ЧАСТОТНЫЕ И ВРЕМЕННЫЕ ХАРАКТЕРИСТИКИ ЗРИТЕЛЬНОЙ
СИСТЕМЫ НАСЕКОМЫХ В ОПТИЧЕСКОМ ДИАПАЗОНЕ
© 2023 г. С.И. Зиенко*, #, В.Л. Жбанова*, ##
*Филиал Национального исследовательского университета «МЭИ» в Смоленске,
Энергетический проезд, 1, Смоленск, 214013, Россия
#E-mail: stanislav-zienko@rambler.ru
##E-mail: vera-zhbanova@eandex.ru
Поступила в редакцию 27.01.22 г.
После доработки 29.06.22 г.
Принята к публикации 14.09.22 г.
Исследованы свойства монохромной, дихромной и трихромной зрительной системы насекомого в
частотной и временной области. В основе метода исследования лежит представление оптической
среды зрительной системы насекомого в виде динамического звена, имеющего вход и выход. Для
аппроксимации спектральных кривых применена функция Гаусса. Исследование инерционных
свойств такого звена во временной области выполнено путем подачи на его вход сигналов в форме
δ-функции или единичной функции. Установившиеся вынужденные колебания на выходе
исследуемого динамического звена определяются при подаче на вход гармонического воздействия.
Предложена методика построения кривых спектральной чувствительности для монохромного,
дихромного и трихромного зрения насекомых в частотной области, установлено, что спектры
обладают свойствами сверхширокополосных сигналов. Обнаружено, что динамическое звено
зрительной системы насекомого обладает свойством усиления оптического сигнала на участках
ультрафиолета и синей части спектра во временной и частотной области. Выявлено, что при частоте
равной и выше частоты диэлектрической релаксации оптическая среда зрительной системы
насекомого не реагирует на электромагнитное излучение. Это явление в наибольшей степени
проявляется в ультрафиолетовой и синей части спектра. Установлено, что показатель
широкополосности оптических сигналов μ изменяется от 0.41 до 1.21, таким образом, зрительная
система насекомого обладает сверхширокополосными свойствами. Показано, что число периодов
световых колебаний в зрительной системе насекомых, имеющей спектр в форме кривой Гаусса,
связано с показателем μ простым соотношением. Выявлено, что при частоте, равной и выше
частоты диэлектрической релаксации, оптическая среда зрительной системы насекомого не
реагирует на электромагнитное излучение. Это явление в наибольшей степени проявляется в
ультрафиолетовой и синей части спектра. Полученные новые знания могут найти применение при
создании матричных твердотельных фотоприемников в ультрафиолетовой и синей части спектра.
Ключевые слова: монохромное зрение, насекомые, спектр, динамическое звено, импульсная
характеристика, переходная характеристика.
DOI: 10.31857/S0006302923010106, EDN: OAIXWK
Насекомые - крупнейшая группа живых орга-
щество фасеточного глаза состоит в том, что та-
низмов, населяющих нашу планету. Несмотря на
кой глаз обладает большим углом обзора. Многим
различный образ жизни устройство зрительной
же млекопитающим, и человеку в том числе, при-
системы почти у всех насекомых одинаково: это
ходится поворачивать голову. Однако за такое
фасеточный глаз [1-4]. Он состоит из омматидий
преимущество глазу насекомого пришлось по-
- отдельных глазков, которые смотрят в различ-
жертвовать резкостью изображения, ведь рез-
ных направлениях. В каждом омматидии есть
кость зависит от диаметра отверстия, через кото-
своя линза; она фокусирует свет на нескольких
рое свет входит в оптическую систему. Острота
фоторецепторных клетках, объединенные в зри-
зрения или разрешающая способность характе-
тельную палочку. Свет, воздействуя на эти клет-
ризует способность глаза различать степень рас-
ки, вызывает последовательность нервных
члененности объектов, в том числе отличать две
импульсов, передаваемых в мозг насекомого по
точки от одной более крупной. Для фасеточного
зрительному нерву. Очевидно, основное преиму-
глаза существует простая зависимость между рас-
98
ЧАСТОТНЫЕ И ВРЕМЕННЫЕ ХАРАКТЕРИСТИКИ
99
стоянием до объекта и числом различимых дета-
лей: чем ближе объект, тем больше деталей видит
насекомое.
Характерно то, что зрительные клетки, как и
твердотельные фотополупроводниковые матери-
алы, поглощают фотоны определенной энергии,
отраженные окружающими предметами или ис-
ходящие непосредственно от источника. Они
воспринимают лучистую энергию в определен-
ном диапазоне длин волн и отличают свет от теп-
лового воздействия лучей. Зрительную клетку фа-
сеточного глаза может возбудить единичный фо-
тон, кроме того, она обладает свойством анализа
поляризации света и восприятия, невидимых че-
ловеком лучей на определенной длине волны.
Цветовое зрение насекомых основано на ис-
пользовании двух или трех фотоприемников с
разными спектральными характеристиками. Под
фотоприемниками понимают определенный тип
фоторецепторов со специфическим зрительным
пигментом. Спектральная чувствительность фо-
топриемника определяется свойствами зритель-
ного пигмента и условиями проникновения света
к фоторецепторам.
Для различения цветов необходимо присут-
ствие максимум двух фотоприемников. Однако
фасеточные глаза термитов имеют только один
фотоприемник, поэтому они неспособны к цве-
товому зрению. В связи с этим условно выделяют
три типа зрения: монохромное, дихромное и три-
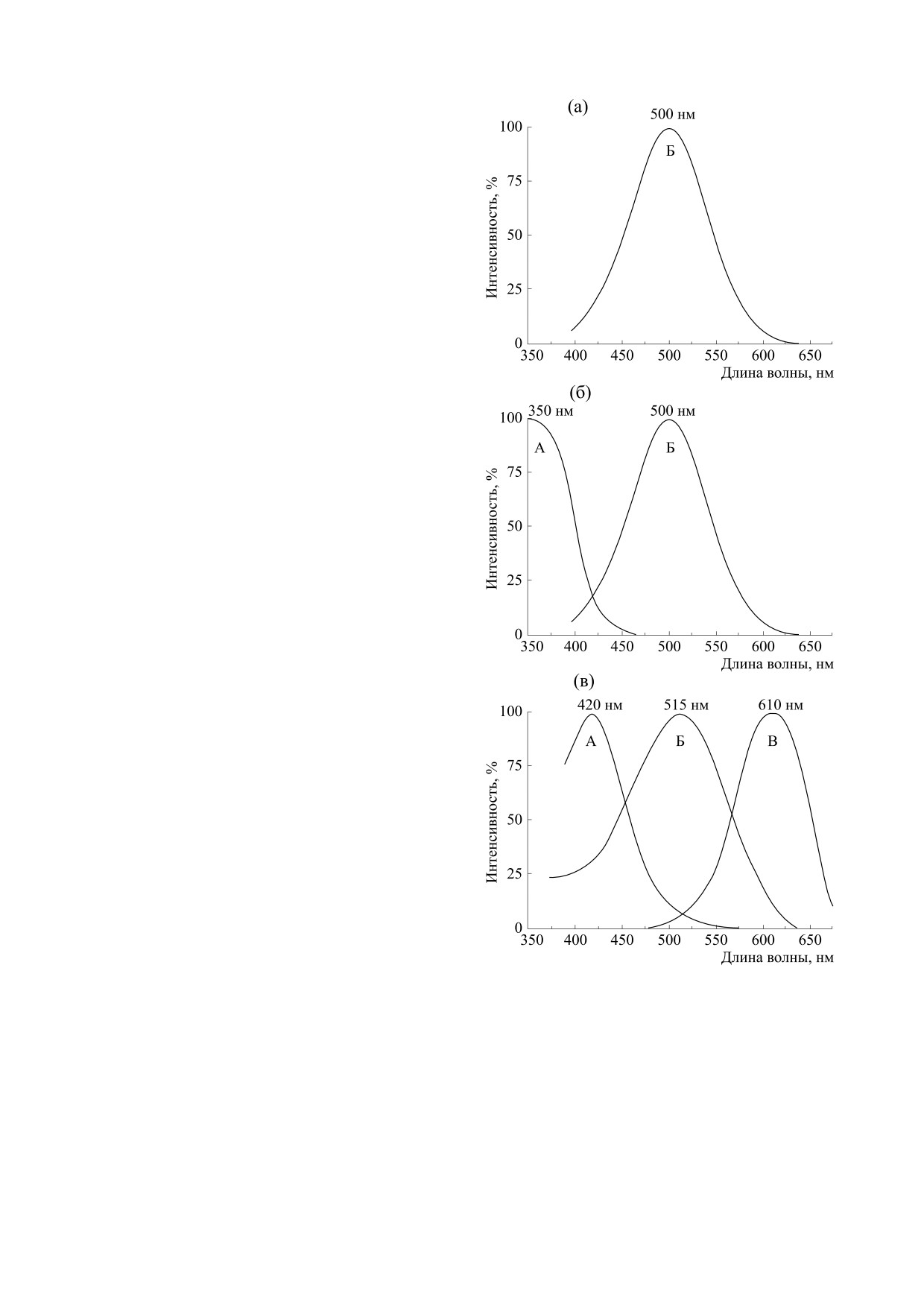
хромное. Их графики, построенные по данным
работы [3], представлены на рис. 1б,в.
Максимальная чувствительность единствен-
ного фотоприемника термитов проявляется при
излучении с длинной волны около
500 нм
(рис. 1а). У жуков-бронзовок в наличии два фото-
приемника, поэтому они способны к цветовому
зрению (рис. 1б). Существуют насекомые, кото-
рые способны различать цвета только одной
(верхней или нижней) половинкой фасеточного
глаза. Так, у стрекозы единственный фотоприем-
ник верхней половины глаза обнаруживает мак-
симум светочувствительности в области 420 нм, в
нижней его половине найдены два фотоприемни-
ка с пиками чувствительности при 515 и 619 нм
(рис. 1в).
Для упрощения анализа на рис. 1 выделены
три характерных участка длин волн: А - первый,
Б - второй и В - третий. Анализ кривых, изобра-
женных на рис. 1, показывает, что спектральные
Рис. 1. Графики относительной спектральной чув-
кривые на участках Б по форме близки к друг дру-
ствительности глаз насекомых разных типов зрения:
монохромного (а), дихромного (б) и трихромного (в).
гу и, следовательно, насекомые разного проис-
хождения имеют одинаковую светочувствитель-
ность. Участок А на рис. 1б расположен в ультра-
большинства насекомых есть ультрафиолетовый,
фиолетовом диапазоне, невидимом для человека.
синий и зеленый фоторецепторы.
В то же время у насекомых спектр для красного
цвета, в отличие от глаза человека, здесь отсут-
Особенностью света, в котором протекает
ствует. В настоящее время установлено, что у
процесс зрительного восприятия насекомыми,
БИОФИЗИКА том 68
№ 1
2023
100
ЗИЕНКО, ЖБАНОВА
не исключение. При решении поставленных за-
дач в исследовании будет применяться необходи-
мый математический аппарат, в том числе - ли-
нейный.
ПОСТРОЕНИЕ КРИВЫХ СПЕКТРАЛЬНОЙ
ЧУВСТВИТЕЛЬНОСТИ ЗРЕНИЯ
НАСЕКОМЫХ В ДИАПАЗОНЕ ЧАСТОТ
Вначале рассмотрим процесс построения кри-
вой спектральной чувствительности для моно-
хромного зрения насекомых. Переход от шкалы
длин волн к шкале энергии выполняли по форму-
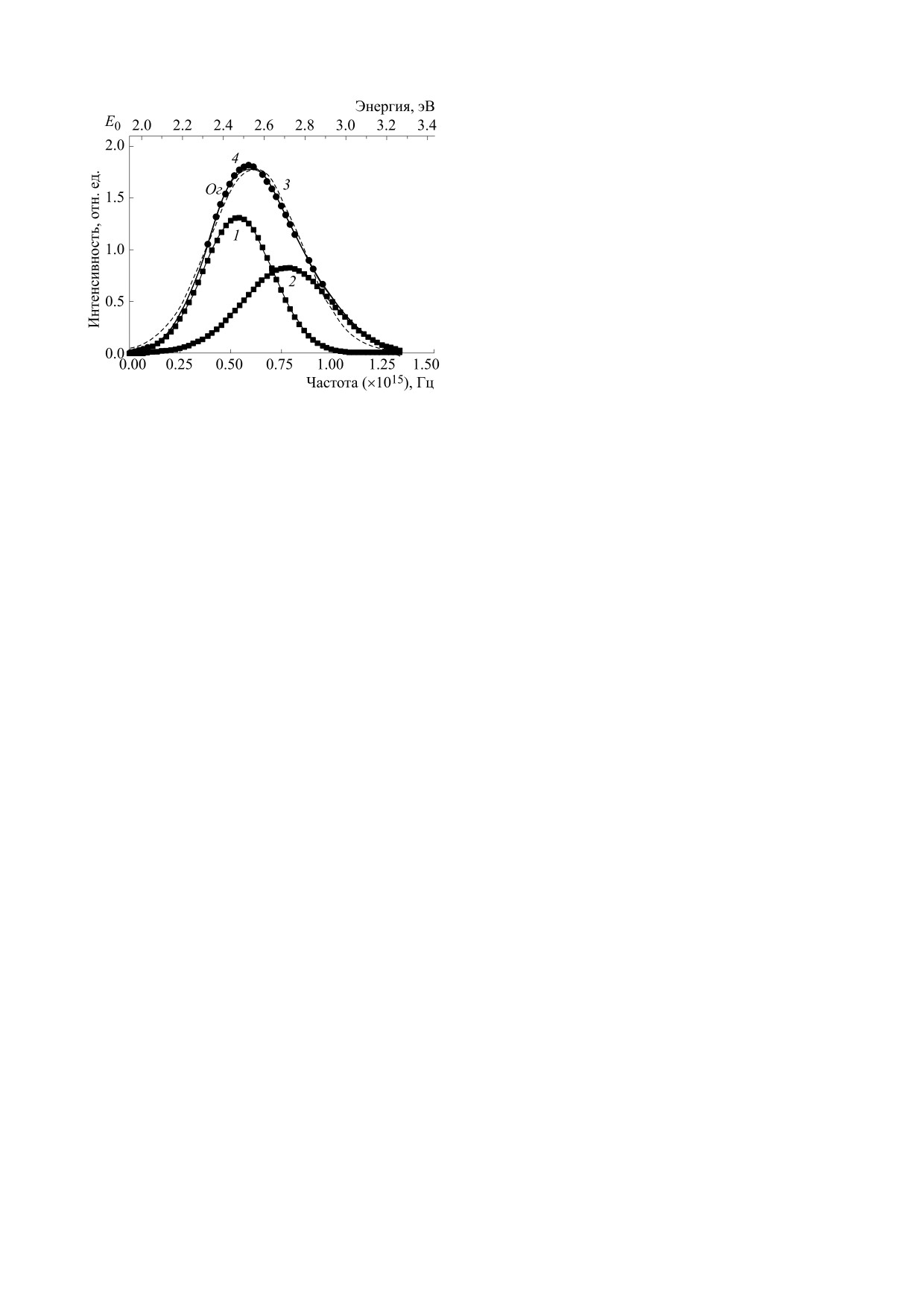
ле E = 1240/λ, где λ в нм и E в эВ (рис. 2). Затем на-
ходили начальное значение энергии E0, при кото-
ром интенсивность равна нулю. В данном случае
E0 = 1.945 эВ.
После этого переходили к шкале частот. Ее на-
Рис. 2. Дифференциальная функция распределения
ходили следующим образом. Из шкалы энергии E
монохромного зрения насекомого в диапазоне частот
вычитали энергию E0, в результате получали шка-
G(f) (сплошная кривая); аппроксимация функции
G(f) кривыми Гаусса (кривые 1 и 2); Ог - огибающая
лу локальной энергии E - E0. Затем учитывали
спектра; 3 - аппроксимация одной кривой Гаусса,
4 - групповое время задержки.
переход от энергии E к энергии E - E0 с помощью
коэффициента m = Eп/(Eп - E0), где Eп - энергия,
соответствующая пику спектральной кривой;
являются его малые временные интервалы. На-
рассчитывали частоту по формуле: f = mv. Здесь
пример, для длины волны 600 нм (оранжевый
v = (E - E0)/h, h = 4.1∙10-15 эВ∙с - постоянная
цвет), период колебаний равен ~2 фс, что соот-
Планка. Окончательно имеем f = (m/4.1)∙(E -
ветствует фемтосекундному масштабу времени.
Это означает фактически полную реализацию
- E0)∙1015 Гц. В расчете за единицу частоты при-
возможностей оптического сигнала. Один период
нимали величину равную 1015 Гц [7, 8]. По такой
оптического колебания - предельная длитель-
же методике строили частотные спектры для ди-
ность светового импульса и одновременно пре-
хромного (кривая Б) и трихромного (кривая В)
дельная «скорость» оптического отклика матери-
зрения насекомых (рис. 3а,б).
альной среды.
Что касается спектров дихромного (кривая А,
Оптика импульсов из малого числа колебаний
рис. 1б) и трихромного (кривые А и Б, рис. 1в)
светового поля интересна как с фундаменталь-
зрения, они содержат фононную компоненту.
ной, так и с прикладной точки зрения. Надо
Для ее устранения переходили от шкалы длин
иметь в виду, что фемтосекундные оптические
волн к шкале частот. В качестве примера рассмот-
импульсы, содержат внутри себя всего несколько
рим процесс преобразования кривой А (рис. 1б).
периодов электромагнитных колебаний [5, 6]. В
На рис. 4 показана спектральная кривая А в ча-
литературе эти вопросы практически не рассмат-
стотном диапазоне, нормированная по площади,
ривались. Между тем решение этих задач имеет
равной единице. Для того чтобы вычесть линей-
большой научный и практический интерес.
ный наклонный фон из исходного спектра, зада-
Цель работы заключается в исследовании зри-
вали значение функции в виде уравнения пря-
тельной системы насекомых различного проис-
мой: y = kx + b. Для данных указанных на рис. 4,
хождения в частотной и временной области. В
значение k = 2.9 и b = -0.52. В этом случае базовая
рамках цели были поставлены следующие задачи:
линия имеет вид пунктирного отрезка прямой 1.
- построение кривых спектральной чувстви-
Результат вычитания, выполненный с помощью
тельности зрения насекомых в диапазоне частот;
математического пакета Origin [9], представлен
на рис. 3а в виде кривой А. Аналогичным образом
- исследование импульсных и переходных ха-
строили спектральные кривые А и В, изображен-
рактеристик динамического звена;
ные на рис. 1в.
- изучение частотных характеристик динами-
Спектры, изображенные на рис. 2 и 3а,б, нор-
ческого звена.
мировали по площади, равной единице. Другими
Следует учесть, что в-основном биологиче-
словами, выполнялось условие нормировки:
ские системы имеют нелинейную зависимость
согласно психофизиологическому закону Вебе-
∞
G( f )df
=
1.
(1)
ра-Фехнера. И зрительная система насекомых -
0
БИОФИЗИКА том 68
№ 1
2023
ЧАСТОТНЫЕ И ВРЕМЕННЫЕ ХАРАКТЕРИСТИКИ
101
Рис. 3. Дифференциальные функции распределения дихромного (а) и трихромного (б) зрения насекомых в диапазоне
частот.
Важным параметром спектральной линии яв-
Численные значения спектральных кривых,
ляется ее ширина, которая является мерой ее не-
изображенных на рис. 2 и 3а,б, и вспомогатель-
монохроматичности. Если Δf/fm << 1, то колеба-
ных данных (E0 и m), используемых при их по-
ния во времени описываются гармонической
строении, представлены в табл. 1.
функцией.
Сигналы в диапазоне 0.25 ≤ μ ≤ 2 называют
сверхширокополосными. При этом сверхширо-
Вместе с тем, когда Δf и fm сопоставимы по ве-
кополосные сигналы с μ = 2 относятся к видео-
личине, в оптике нет четкого определения харак-
сигналам, с 0.1 ≤ μ ≤ 0.2 - к широкополосным и с
терной величины их отношения. В то же время в
μ ≤ 0.001 - к узкополосным.
радиотехнических приложениях для оценки ши-
Узкополосные и сверхширокополосные сиг-
рины спектра вводится показатель широкополос-
налы существенно отличаются друг от друга. Уз-
ности, который определяется из следующего со-
кополосные синусоидальные сигналы обладают
отношения [10]:
уникальными свойствами. При таких широко ис-
пользуемых преобразованиях, как сложение, вы-
Δf
f
макс
-
f
мин
μ=
=
,
(2)
читание, интегрирование, дифференцирование
f
(f
+
f
)/2
их форма остается прежней. Здесь и далее под
макс
мин
m
формой понимается закон изменения сигнала во
где fm, fмин и fмакс - средняя, минимальная и мак-
времени. Преобразованные сигналы могут разли-
симальная частоты энергетического спектра,
чаться только амплитудой и сдвигом во времени.
Δf = fмакс- fмин - ширина полосы частот сигнала
В противоположность им у сверхширокополос-
на его полувысоте.
ного сигнала при указанных (и других) преобра-
Таблица 1. Основные параметры спектральных кривых насекомых в частотной области
Тип зрения
Участок
fm, ×1015 Гц
Δf, ×1015 Гц
Н, отн. ед.
E0, эВ
m
μ
fr, ×1015 Гц
Монохромное
А
0.60
0.52
1.77
1.94
4.61
0.86
0.81
А
0.57
0.34
2.91
1.80
2.27
0.60
0.43
Дихромное
Б
0.60
0.57
1.68
1.94
4.61
0.9
0.89
А
0.76
0.33
2.96
1.80
8.64
0.43
0.37
Трихромное
Б
0.55
0.65
1.43
2.15
3.80
1.18
0.80
В
0.49
0.61
1.50
1.95
5.00
1.21
0.69
БИОФИЗИКА том 68
№ 1
2023
102
ЗИЕНКО, ЖБАНОВА
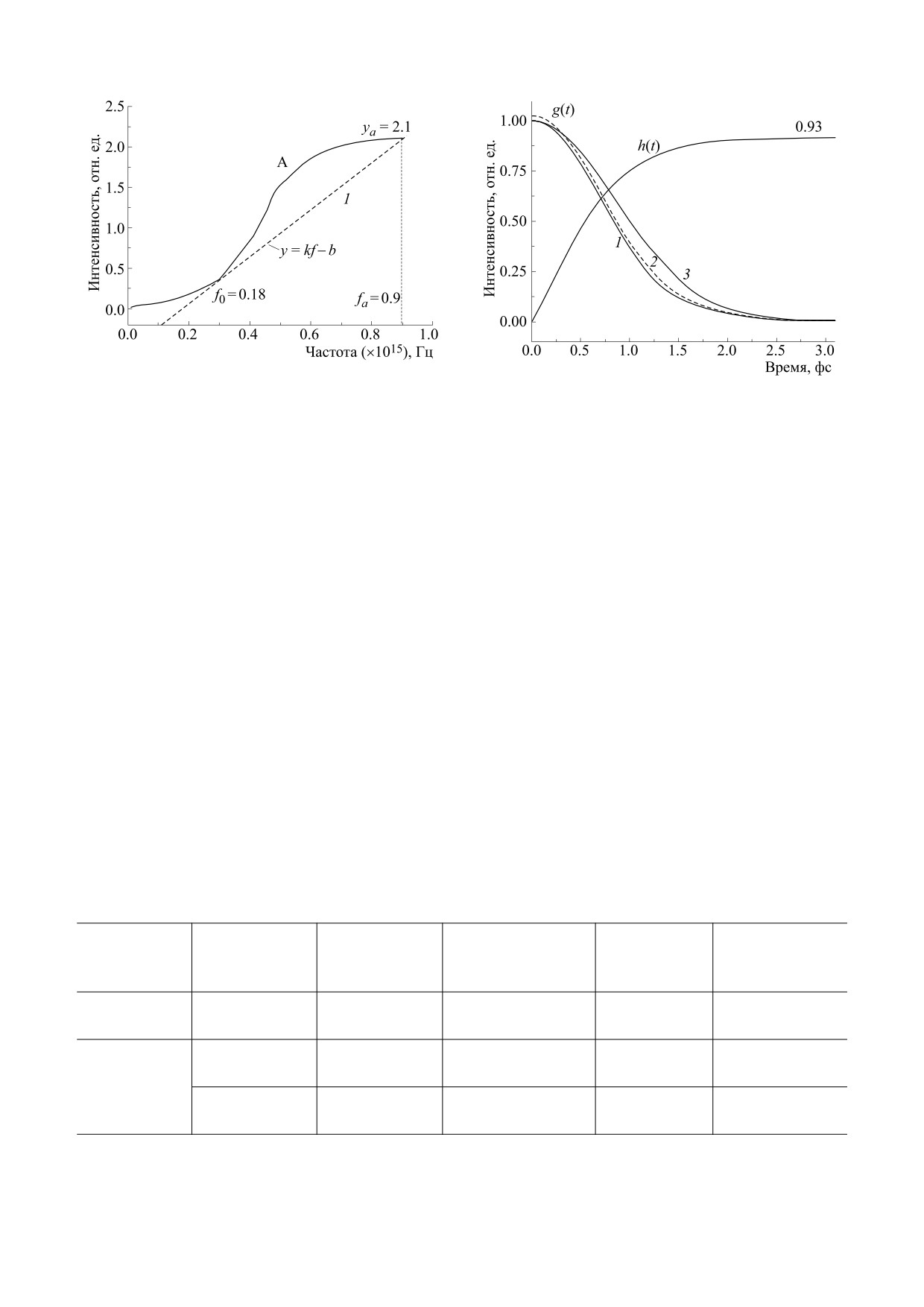
Рис. 4. Задание базовой линии (пунктир) в виде
линейной функции.
Рис. 5. Модуль (кривая 1) импульсной характеристи-
ки и мнимая компонента (кривая 2) комплексной им-
пульсной характеристики монохромного зрения на-
секомого. Сплошная линия - численный расчет,
зованиях изменяются не только параметры, но и
пунктир - аппроксимация кривой Гаусса.
форма [11, 12]. В табл. 1 приведены рассчитанные
по формуле (3) значения μ для разных типов зре-
ния насекомых. Можно видеть, что величина μ
t
находится в диапазоне 1.21 ≥ μ ≥ 0.43, и по опреде-
ht)
=
g t)dt
(3)
0
лению зрительная система насекомых разного ти-
па обладает свойствами сверхширокополосных
Импульсную (временную) характеристику на-
сигналов.
ходят по формуле обратного преобразование Фу-
рье:
ИМПУЛЬСНЫЕ И ПЕРЕХОДНЫЕ
ХАРАКТЕРИСТИКИ ДИНАМИЧЕСКОГО
*
∞
g
(t)
=
G f)exp(-2πft)df
(4)
ЗВЕНА
0
Оптическую среду зрительной системы насе-
Вначале решение интеграла (5) выполнено
комого удобно представить для анализа в виде ди-
численным методом для монохромного зрения
намического звена, имеющего вход и выход. Во
(функция G(f), рис. 2). Результаты расчета для мо-
временной области для исследования инерцион-
дуля импульсной характеристики приведены на
ных свойств такого звена выполняют путем пода-
рис. 5 (кривая 1). Численный метод дает высокую
чи на его вход сигналов в форме δ-функции
точность расчета, но не позволяет установить
или единичной функции 1(t). Реакцию звена на
влияние параметров частотного спектра (fm и Δf)
δ-функцию называют импульсной (временной)
на временные характеристики переходного про-
характеристикой g(t), а на единичную функцию
цесса. Для установления их влияния аппрокси-
1(t) - переходной характеристикой h(t). Функции
мируем функцию G(f) одной кривой (кривая 3 на
g(t) и h(t) связаны между собой соотношением:
рис. 2) и двумя кривыми Гаусса (пики 1 и 2 на
Таблица 2. Параметры аппроксимации частотного спектра монохромной системы зрения
Число кривых
Номер кривой на
Площадь,
Пик, ×1015 Гц
Ширина,
Амплитуда,
Гаусса
рис. 2
отн. ед.
×1015 Гц
отн. ед.
1
3
0.99
0.62
0.44
1.78
2
1
0.56
0.54
0.41
1.31
2
0.44
0.76
0.49
0.82
БИОФИЗИКА том 68
№ 1
2023
ЧАСТОТНЫЕ И ВРЕМЕННЫЕ ХАРАКТЕРИСТИКИ
103
рис. 2) [13]. Параметры аппроксимации приведе-
тельности переходного процесса монохромного
ны в табл. 2.
зрения насекомых, равной от 0.8 до 2.5 периодов
Когда спектр описывается одной кривой Гаус-
колебаний света.
са (2), модуль импульсной (временной) характе-
В общем случае интеграл (4) с учетом соотно-
ристики g(t) может быть получен из решения со-
шения (6) не имеет аналитического решения.
отношений (2) и (4) в аналитическом виде [14]:
Численное решение его с данными, представлен-
ными в табл. 1, приведено в виде графика h(t) на
g(t) = Cexp(-at2),
(5)
рис. 5. Можно видеть, что значение переходной
где коэффициент С = 1 - амплитуда импульсной
характеристики меняется от 0 до максимального,
равного h(∞) = 0.93. В тоже время при t = ∞ мож-
характеристики, параметр а = 3.5Δf2. Из соотно-
но получить для h(∞) простое соотношение:
шения (6) можно видеть, что форма импульсной
h(∞) = С∙0.47/Δf. Коэффициент С - имеет раз-
(временной) характеристики определяется
мерность - число фотонов в единицу времени
шириной спектра Δf и не зависит от пиковой
(одну фемтосекунду). Коэффициент передачи
частоты. Расчетная графическая зависимости
(по мощности) динамического звена находится
g(t), построенная с учетом данных табл. 2, приве-
как в работе [18]:
дены на рис. 5 в виде кривой 3.
kmax = h(∞)/1(t) = C·0.47/Δf,
(9)
В случае аппроксимации спектра двумя кри-
где 1(t) - единичная функция или функция Хеви-
выми Гаусса импульсная (временная) характери-
сайда.
стика имеет вид [15-17]:
Из соотношения (10) следует, что когда шири-
2
2
на спектра не превышает критического значении
g(t)
=
{A t)1
+
2
A t)
+
2A t)1
2
A t) cos (2πΩt)}1/2,(6)
Δfкр = 0.47∙1015 Гц, коэффициент передачи боль-
где A1(t) = C1exp(-α1t2), A2(t) = C2exp(-α2t2), α1 =
ше единицы. Следовательно, в такой ситуации
= 3.5Δf12, α2 = 3.5Δf22, Ω = f2 - f1.
динамическое звено обладает свойством усиле-
ния оптического сигнала. Согласно данным
Результаты расчета по формуле (7) с учетом
табл. 1 этому условию удовлетворяет участки А
данных табл. 2 представлены на рис. 5 в виде кри-
дихромного и трихромного спектра глаза насеко-
вой 2. В данном случае результаты аналитическо-
мых.
го расчета хорошо согласуются с данными чис-
ленного анализа (кривая 1, рис. 5). Это связано с
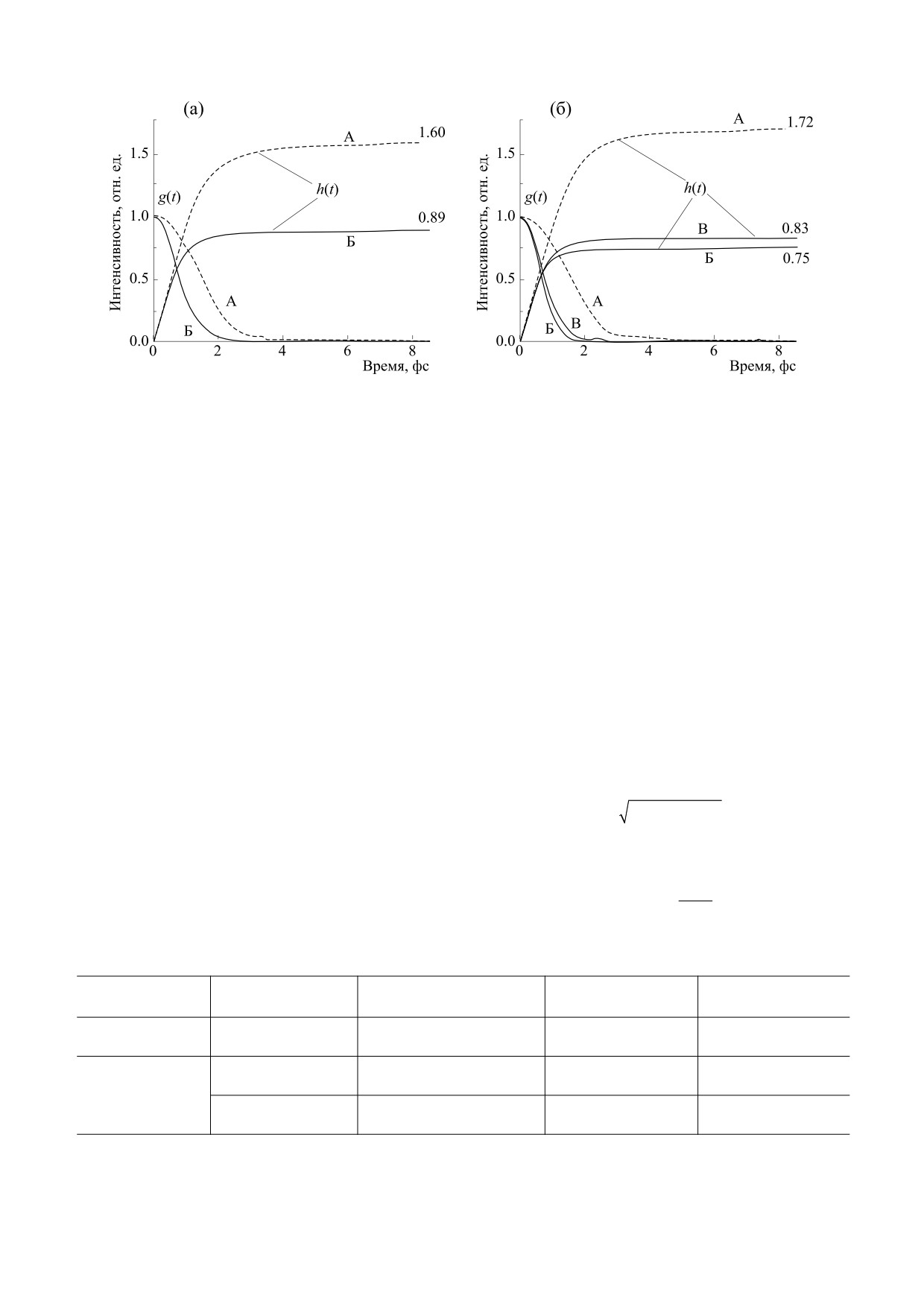
Для исследования данного явления было вы-
тем, что, как показывает анализ уравнения (7),
полнено численное решение интеграла (5) для
форма кривой g(t) зависит не только от
участков А и Б дихромного (рис. 3а) и участков А,
ширины спектра Δf, но от разности пиковых ча-
Б и В трихромного зрения (рис. 3б). Результаты
стот Ω = f2 - f1. В то же время аппроксимация
расчета в виде графиков g(t) представлены на
спектра одной кривой Гаусса позволяет получить
рис. 6а,б. Затем по известным значениям g(t) вы-
простое аналитическое соотношение
(6) для
полняли численное решение интеграла (4). Гра-
оценки длительности переходного процесса в оп-
фические зависимости h(t) приведены там же на
тической среде глаза насекомого.
рис. 6а,б.
Импульсная (временная) характеристика по
Численные значения kmax для участков А и Б
физическому смыслу отражает нелинейные свой-
дихромного и участков А, Б и В трихромного зре-
ства процесса перехода глаза насекомого из за-
ния на основании данных численного расчета
крытого состояния в открытое состояние. Соот-
(рис. 6а,б) представлены в табл. 3. В ней также
ношение (6) с учетом соотношения (2) можно
приведены величина времени нарастания tn, из-
привести к виду
меренное на уровне 0.1,…,0.9 от максимального
значения для каждого из участков. Здесь также
g(t) = exp(-3.5μ2(t/Tm)2,
(7)
приведены данные для монохромного зрения.
где Tm = 1/fm - период колебаний. Длительность
Анализ данных табл. 3 показывает, что участ-
переходного процесса tп оценим на уровне 0.03,
кам А дихромного и трехмерного зрения, в отли-
затем, полагая tп = nT0, где n - число периодов ко-
чие от других, соответствуют повышенные значе-
ния kmax, равные 1.69 и 1.72. Это указывает на су-
лебаний, находим связь числа колебаний с пока-
зателем широкополосности:
ществование в зрительной системе насекомых
механизма усиления оптического излучения в
n = 1/μ.
(8)
ультрафиолетовой и синей части спектра. Дан-
В соответствии с данными табл. 1 минималь-
ный результат согласуется с результатами иссле-
ное значение показателя широкополосности со-
дования, опубликованные в научной литературе
ставляет μmin ≈ 0.4 и максимальное - μmax ≈ 1.2.
[1]. В ней отмечается высокая чувствительность
Согласно выражению (9) это соответствует дли-
фасеточных глаз к ультрафиолету и синему свету
БИОФИЗИКА том 68
№ 1
2023
104
ЗИЕНКО, ЖБАНОВА
Рис. 6. Импульсные (временные) g(t) и переходные характеристики h(t) для дихромного (а) и трихромного зрения (б).
Численные значения kmax указаны напротив соответствующих кривых h(t).
и проявляется у всех насекомых. Исключение
рактеристики являются энергетическими харак-
красных лучей из видимой части спектра не обя-
теристиками системы, которые устанавливают
зательно, например, дневные бабочки и мухи ре-
связь между спектрами входного и выходного
агируют на красный цвет [2, 3].
сигналов, представляющих прямое преобразова-
ние Фурье от функции времени g(t).
Важное значение для переходного процесса
имеют потери энергии ΔЕ, Их величину находили
По известной зависимости g(t) вычисляли
с помощью равенства Парсеваля [19]:
комплексную частотную характеристику r*(f).
∞
2
Для этого использовали формулу прямого преоб-
ΔE= g
(t)dt.
(10)
0
разование Фурье [7, 20]:
Численные значения энергии ΔЕ для различ-
∞
ных типов зрения приведены в табл. 3. Анализ
*
r
(f)
=
g(t)exp(-2πjft)dt
(11)
этих данных показывает, что эффект усиления
0
света на участке А дихромного и трихромного
Функция (12) содержит вещественную a(f) и
зрения сопровождается заметными потерями
мнимую части b(f).
энергии равными ~1.11 и ~1.20 эВ. На других
k*(f) = a(f) + j·b(f).
(12)
участках спектра подобные потери, примерно, в
два раза меньше.
Модуль функции (13) вычисляли по формуле
2
2
k( f )
=
a
(f)
+
b
(f ).
(13)
ЧАСТОТНЫЕ ХАРАКТЕРИСТИКИ
ДИНАМИЧЕСКОГО ЗВЕНА
Величину фазового сдвига находили по фор-
Частотные характеристики описывают уста-
муле:
новившиеся вынужденные колебания на выходе
b f )
исследуемого динамического звена при подаче на
φ
(f )
= -arctg
(14)
вход гармонического воздействия. Частотные ха-
a f )
Таблица 3. Численные параметры временных характеристик
Тип зрения
Участок
kmax
t н, фс
ΔE, эВ
Монохромное
Б
0.93
1.35
0.63
А
1.60
2.10
1.11
Дихромное
Б
0.88
1.00
0.59
БИОФИЗИКА том 68
№ 1
2023
ЧАСТОТНЫЕ И ВРЕМЕННЫЕ ХАРАКТЕРИСТИКИ
105
Рис. 7. Частотные характеристики зрительной системы насекомого: (а) - амплитудно-частотные характеристики
(участок А дихромного зрения), 1 - вещественная составляющая спектра a(f), 2 - мнимая составляющая спектра b(f),
3 - модуль r(f), 4 - групповое время запаздывания; (б) - амплитудные фазочастотные характеристики, (участок А), 1 -
дихромное зрение, 2 - трихромное зрение.
Для оценки временных процессов используют
зрительной системы насекомого. Тот факт, что на
понятие группового времени запаздывания. В об-
рис. 7б амплитудные фазочастотные характери-
щем случае групповое время запаздывания опре-
стики не охватывают начала координат, свиде-
деляется первой производной фазового спектра
тельствуют об устойчивости режима усиления.
(15) со знаком минус:
Значения частоты диэлектрической релаксации fr
для зрительных систем насекомых различного
1
dφ
τ(f )
=-
⋅
(15)
происхождения приведены в табл. 1. Из сопостав-
360
df
ления этих данных можно видеть, что дихромное
Здесь фазовый угол φ(f) измеряется в угловых
и трихромное зрения на участке А имеют частоту
градусах, нормирующий множитель имеет вели-
f
r, которая по величине меньше пиковой частоты
чину 360°. Графические амплитудно-частотные
f
m, тогда как другие типы зрения имеют обратную
характеристики и амплитудные фазочастотные
картину.
характеристики зрительной системы насекомого,
Численное значение модуля частотной харак-
приведены на рис.
7а и
7б соответственно.
теристики r(0) находим из соотношения (12), по-
Кривая 4, изображенная на рис. 7а, показывает
лагая в нем частоту f = 0:
зависимость группового времени запаздывания
от частоты. При частоте, равной частоте диэлек-
∞
трической релаксации fr, производная dφ/df в со-
r(0)
=
g(t)dt
(16)
0
отношении (16) равна нулю, оптическая среда
зрительной системы насекомого не реагирует на
Соотношение (17) совпадает с интегралом (4),
электромагнитное излучение. По существу, ча-
если в нем положить t = ∞. Тогда r(0) = kmax и,
стота fr представляет собой предельное значение
следовательно, эффект усиления света у дихром-
частоты, при которой оптическая среда не реаги-
ного и трихромного зрения (участок А) проявля-
рует на оптическое излучение. При этом веще-
ется и в частотной области. Это явление можно
ственная составляющая комплексного спектра
объяснить, если рассмотреть закон Бугера-Лам-
(кривая 1, рис. 7а) принимает нулевое значение,
берта для поглощения света
т. е. активные потери энергии отсутствует, а мо-
дуль r(f) (кривая 3, рис. 7а) становится, как следу-
I = I0exp(-ax),
(17)
ет из формулы (14), равным мнимой компоненте
(кривая 2, рис. 7а). В последнем случае зритель-
где а - положительный показатель поглощения;
ная система насекомого проявляет емкостные
x - толщина поглощающего слоя; I0 - интенсив-
свойства. Поэтому в точке a (рис. 7б) происходит
ность света, входящего в среду; I - интенсивность
завершение кривой годографа радиус-вектора
света, прошедшего слой толщиной x.
БИОФИЗИКА том 68
№ 1
2023
106
ЗИЕНКО, ЖБАНОВА
Впервые особенности среды с отрицательным
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
поглощением света рассмотрел советский физик
Настоящая статья не содержит каких-либо ис-
В.А. Фабрикант. Им было показано, что для та-
следований с участием людей или животных в ка-
кой среды закон (18) имеет другой вид:
честве объектов исследований.
I = I0exp(|a|x),
СПИСОК ЛИТЕРАТУРЫ
где |а| - положительная величина, что соответ-
1.
В. П. Тыщенко, Физиология насекомых (Высшая
ствует не ослаблению, а усилению света по мере
школа, М., 1986).
продвижения его через вещество. Иначе говоря, в
2.
M. J. Klowden, Physiological Systems in Insects.
усиливающей среде показатель поглощения сре-
Academ. Press, 2007. 688 p.
ды становится отрицательным. Этим объясняется
3.
Nation J. L. Insect Physiology and Biochemistry, 2nd
то, что подобную среду иногда называют средой с
Edition (CRC Press, 2008).
отрицательным показателем поглощения. Это
4.
B.-M. Song and Ch.-H. Lee, Front Neural Circuits,
означает, что в такой среде лавинообразно воз-
12, 16 (2018). DOI: 10.3389/fncir.2018.00016
растает число фотонов за счет актов вынужденно-
5.
В. Г. Беспалов, С. А. Козлов, А. Н. Петров и др.
го излучения.
Фемтосекундная оптика и фемтотехнология (Уни-
верситет ИТМО, СПб., 2018).
6.
Ю. А. Шполянский, Автореф. дисс. … докт. физ-
ВЫВОДЫ
мат. наук (Университет ИТМО, СПб., 2010).
1. Предложена методика построения кривых
7.
С. И. Зиенко и В. Л. Жбанова, Прикладная физи-
спектральной чувствительности для монохром-
ка, № 3, 46 (2021). DOI: 10.51368/1996-0948-2021-3-
ного, дихромного и трихромного зрения насеко-
39-46
мых в частотной области.
8.
S. I. Zienko and V. L. Zhbanova, Scientific and Tech-
nical Journal of Information Technologies, Mechanics
2. Установлено, что показатель широкополос-
and Optics, 21 (6), 828 (2021). DOI: 10.17586/2226-
ности оптических сигналов μ изменяется от 0.41
1494-2021-21-6-828-836
до 1.21 и, по определению (2), зрительная система
9.
О. П. Исакова, Ю. Ю. Тарасевич и Ю. И. Юзюк,
насекомого обладает сверхширокополосными
Обработка и визуализация данных физических экспе-
свойствами.
риментов с помощью пакета Origin (Книжный дом
«Либриком», М., 2009).
3. Показано, что число периодов световых ко-
лебаний в зрительной системе насекомых, имею-
10.
О. В. Лазоренко и Л. Ф. Черногор, Радиофизика и
радиоастрономия, 13 (2), 166 (2008).
щей спектр в форме кривой Гаусса, связано с по-
казателем μ простым соотношением: n = 1/μ.
11.
Ultra-wideband Radar Technology, Ed. by D. James and
P. E. Taylor (CRC Press, Boca Raton, 2000).
4. Обнаружено динамическое звено зритель-
12.
Э. В. Семенов, Автореф. дисс. … докт. техн. наук
ной системы насекомого, обладающее свойством
(М., 2012).
усиления оптического сигнала на участках уль-
13.
А. В. Коваленко, С. М. Вовк и Е. Г. Плахтий,
трафиолета и синей части спектра во временной и
Журн. прикладной спектроскопии, 88 (2), 297
частотной области.
(2021).
5. Выявлено, что при частоте равной и выше
14.
Я. Б. Зельдович и А. Л. Мышкис, Элементы при-
частоты диэлектрической релаксации оптическая
кладной математики (Наука, М., 1972).
среда зрительной системы насекомого не реаги-
15.
S. I. Zienko and D. S. Slabkovsky, Technical Physics
рует на электромагнитное излучение. Это явле-
Letters, 45 (46), 537 (2019).
ние в наибольшей степени проявляется в ультра-
16.
S. I. Zienko and D. S. Slabkovsky, Optics and Spectros-
фиолетовой и синей части спектра.
copy, 127 (3), 564 (2019).
17.
С. И. Зиенко и Д. С. Слабковский, Прикладная
6. Полученные новые знания могут найти при-
физика, № 1, 36 (2020).
менение при создании матричных твердотельных
18.
М. П. Туманов, Теория линейных систем автома-
фотоприемников в ультрафиолетовой, и синей
тического управления: Учебное пособие (МГИЭМ,
части спектра.
М., 2005).
19.
И. С. Гоноровский, Радиотехнические цепи и сигна-
лы (Радио и связь, М., 1986).
КОНФЛИКТ ИНТЕРЕСОВ
20. В. В. Пасынков и В. С. Сорокин, Материалы элек-
Авторы заявляют об отсутствии конфликта
тронной техники, 3-е изд. (Издательство «Лань»,
интересов.
СПб.: 2001).
БИОФИЗИКА том 68
№ 1
2023
ЧАСТОТНЫЕ И ВРЕМЕННЫЕ ХАРАКТЕРИСТИКИ
107
Frequency and Time Domain Characteristics of the Insect Visual System in Optic Flow
S.I. Zienko* and V.L. Zhbanova*
*Branch of National Research University "Moscow Power Engineering Institute" in Smolensk,
Energeticheskii proezd 1, Smolensk, 214013 Russia
Frequency and time domain properties of the monochromatic, dichromatic, and trichromatic visual system
in an insect have been studied. The research method is based on the representation of the optical flow field
of the insect visual system in the form of a dynamic link with an input and output. The Gaussian function was
used to approximate the spectral curves. The study of the inertial properties of such a link in the time domain
was performed by applying signals to its input in the form of a δ-function or a unit function. The steady-state
forced oscillations at the output of the investigated dynamic link are determined when a harmonic effect is
applied to the input. A technique for constructing spectral sensitivity curves for monochrome, dichromatic,
and trichromatic vision of insects in the frequency domain is proposed; it has been established that the spectra
exhibit the properties of ultra wideband signals. It has been found that the dynamic link of the insect visual
system is capable of amplifying the optical signal over ultraviolet and blue parts of the spectrum in the time
and frequency domain. It has been revealed that at a frequency equal to or higher than the frequency of di-
electric relaxation, the optical flow field of the insect's visual system does not sense electromagnetic radiation.
This phenomenon is most pronounced in ultraviolet and blue parts of the spectrum. It has been established
that the optical signal wideband index μ varies from 0.41 to 1.21; thus, the visual system of an insect has ultra
wideband properties. It has been shown that the number of periods of light oscillations in the visual system of
insects, which has a spectrum in the form of a Gaussian curve, is related to the exponent μ by a simple rela-
tionship. New knowledge gained can be used for creation of matrix solid-state photodetectors in ultraviolet
and blue parts of the spectrum.
Keywords: monochrome vision, insects, spectrum, dynamic link, impulse response, transient response
БИОФИЗИКА том 68
№ 1
2023