БИОФИЗИКА, 2023, том 68, № 1, с. 134-141
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 51-73
ЧИСЛЕННОЕ МОДЕЛИРОВАНИЕ ПРЕДНЕРЕСТОВОЙ И НЕРЕСТОВОЙ
МИГРАЦИЙ ПРЕДСТАВИТЕЛЯ СЕМЕЙСТВА ТЕРПУГОВЫХ РЫБ
(НА ПРИМЕРЕ ОДНОПЕРОВОГО ТЕРПУГА)
© 2023 г. А.Н. Четырбоцкий*, #, А.Н. Вдовин**, В.А. Четырбоцкий***
*Дальневосточный геологический институт ДВО РАН,
просп. 100-летия Владивостока, 159, Владивосток, 690022, Россия
**Тихоокеанский филиал ВНИИ рыбного хозяйства и океанографии («ТИНРО»),
пер. Шевченко, 4, Владивосток, 690091, Россия
***Физический факультет Московского государственного университета имени М.В. Ломоносова,
Ленинские горы, 1, Москва, 119991, Россия
#E-mail: chetyrbotsky@yandex.ru
Поступила в редакцию 05.09.2022 г.
После доработки 25.10.2022 г.
Принята к публикации 27.10.2022 г.
Предложена пространственно-временная модель динамики терпуговых рыб в период их преднере-
стовых и нерестовых массовых миграций. Модель разработана на материале оригинальных верифи-
цированных многолетних наблюдений и промышленных уловов в заливе Петра Великого (Япон-
ское море), запись уравнений динамики плотности самцов/самок и массовых суточных перемеще-
ний рыб выполнена в терминах уравнений переноса. Эти уравнения представлены в форме
модифицированных уравнений Патлака-Келлера-Сегеля, согласно которым поток объектов/суб-
станции направлен вдоль градиентов введенных функций зон стимулов. Полагается, что в предне-
рестовый период происходят морфофизиологическая (биохимическая терморегуляция и ряд других
эндогенных процессов) и поведенческая адаптации к достаточно длительному и энергозатратному
нересту, где стимулом массовых перемещений рыб выступает их пребывание в комфортных для них
в этот период условиях среды. При нересте стимул обусловливает поиск распределенных в при-
брежной области дна водоема удобных и хорошо аэрированных выступов его рельефа. Моделиро-
вание связанных со стимулами действий выполнено на основании сведений о предпочтительных
глубинах преднерестовой зоны рыб и особенностях распределений комфортных для них участков
нереста. Полагается, что интенсивность суточных перемещений пропорциональна их линейному
размеру (чем крупнее рыба, тем быстрее она движется). В уравнениях для этапа нереста учитывается
пространственная конкуренция самцов, что в естественных условиях наблюдается только в окрест-
ности нерестовых участков. Вне этих участков самцы продолжают поиски новых удобных для нере-
ста участков. Для самок стимулом передвижений выступают скопления самцов, сигналом о скопле-
нии которых могут выступать некоторые выделяемые самцами субстанции (например, выделяемая
самцами слизь) или визуальный контакт. Учитывается диффузия распределения рыб и вязкость
среды обитания (диффузия скорости). Начальное распределение рыб задается их усредненным за
многолетний период наблюдений июльским распределением в заливе Петра Великого.
Ключевые слова: миграция рыб, нерест, уравнения Патлака-Келлера-Сегеля, зона адаптации.
DOI: 10.31857/S0006302923010155, EDN: OBCFQP
Характер пространственно-временной после-
них прогрета и аэрирована. Именно здесь проис-
довательности этапов жизненного цикла рыб от-
ходит их приспособление к продолжительному
ражает особенности их эволюционного пути раз-
энергозатратному периоду последующего нере-
вития. Так, в преднерестовый этап поколения не-
ста. К тому же повышенная температура этих глу-
рестовых мигрирующих видов рыб регулярно
бин интенсифицирует процессы дозревания их
преодолевают значительные расстояния, чтобы
половых продуктов. И здесь же рыбы усиленно
затем образовать нерестовые скопления. Для тер-
питаются, компенсируя затраты энергии на фор-
мирование половых продуктов.
пуговых рыб это область первой сотни метров
морских глубин, которая обычно к моменту нача-
Согласно наблюдениям, самцы первыми по-
ла преднерестовой миграции уже достаточно для кидают преднерестовую зону и поднимаются к
134
ЧИСЛЕННОЕ МОДЕЛИРОВАНИЕ ПРЕДНЕРЕСТОВОЙ И НЕРЕСТОВОЙ МИГРАЦИЙ
135
местам нереста, что связано с расчисткой нере-
Pleurogrammus azonus, который является наиболее
стового участка и подготовкой галечного грунта
массовым представителем семейства терпуговых
для строительства нерестового бугра [1]. Перед
Hexagrammidae, включающего в себя три рода и
занятием нерестовых участков самцы прогоняют
9 видов [5].
более или менее крупных рыб. Вслед за ними сю-
да отправляются и самки для откладывания икры,
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ
которую затем оплодотворяют самцы. Интересно
заметить, что многократное спаривание и увели-
Количественным основанием для построения
чение плотности в скоплениях обеспечивает ре-
модели является система уравнений Патлака-
продуктивный успех в условиях идеального рас-
Келлера-Сегеля [6, 7]. Суть положений ее меха-
пределения самок [2, 3].
низма состоит в принятии положения о направ-
Постановка задач математического моделиро-
ленных массовых перемещениях особей в поле
вания преднерестовых и нерестовых миграций
гетерогенного распределения некоторой функ-
морских рыб не является важной темой предмет-
ции, которая в терминах работ [7, 8] именуется
но ориентированных публикаций, что, по-види-
стимулом или так называемым медленным такси-
мому, обусловлено трудностями выбора универ-
сом. В частности, стимулом для насекомых вы-
сальных (или относительно стандартных) схем
ступают распределения концентрации их пище-
из-за различий в стратегии размножения, этоло-
вого субстрата [7], а для бактерий водоносных
гии нереста и характера миграций. На наш взгляд,
пластов - распределение концентрации их пита-
основная причина такой ситуации состоит в мо-
тельных веществ [9]. Полагается, что поток непо-
бильности перемещений рыб и малости их разме-
средственно самих объектов пропорционален
ров по сравнению с остальными характерными
распределению стимула. Иная ситуация положе-
размерами. Количественные измерения показа-
на в основу моделей [10, 11], где интенсивность
телей у плавающих рыб и обследование нерести-
перемещений пропорциональна градиенту сти-
лищ на порядок сложнее аналогичных исследова-
мула. Подобным образом этот градиент «управля-
ний в сухопутной среде. Между тем актуальность
ет» характером усредненных за сутки скоростей
таких исследований определяется теоретически-
объектов изучения.
ми и прикладными аспектами. Во-первых, актив-
В системе уравнений Патлака-Келлера-Сеге-
ные перемещения рыб дают искаженную оценку
ля допускается, что объект наделен таксисом к
промысловых запасов, что может отразиться на
некоторым характеристикам окружающей среды
рациональной эксплуатации промысловой части
(такими как температура, соленость, рельеф
популяции. Кроме того, полученные результаты
местности и т. д.), то есть он способен восприни-
могут оказаться полезными для краткосрочного
мать некоторый внешний сигнал [12]. Здесь рас-
прогнозирования промысловой обстановки.
сматриваются четыре типа таких распределений,
Цель работы состоит в разработке компактной
посредством которых моделируется направлен-
численной 1D-модели преднерестовой и нере-
ное массовое перемещение нерестовых рыб: гра-
стовой миграций массовых самцов и самок, где в
диент зоны адаптации, градиент зоны нереста и
рамках единого механизма рассматриваются про-
распределение самцов уже непосредственно пе-
блемы преднерестовой приспосабливаемости и
ред нерестовыми участками (в этот период самки
таксис рыб, конкуренция самцов за место нере-
перемещаются вслед за самцами и, кроме того, на
стового участка. Под таксисом здесь понимаются
этих участках самцы конкурируют между собой за
массовые направленные перемещения, обуслов-
оплодотворение икры).
ленные неоднородностью пространственного
При построении модели принимаются такие
распределения некоторого стимула [4]. Для реше-
допущения:
ния поставленных задач вводится понятие этапа
адаптации, по завершения которого самцы и сле-
1. Динамика распределения плотности рыб
дующие за ними самки уходят на нерест. Для са-
следует уравнению переноса, где еще дополни-
мок в качестве стимула их передвижений высту-
тельно учитывается диффузия их плотности и
пают скопления самцов, сигналом о таком скоп-
естественная смертность.
лении могут являться выделяемые ими
2. Вводятся понятия зоны адаптации и зоны
некоторые химических вещества (например, вы-
действия стимулов.
деляемая самцами характерная слизь). Для оцен-
ки адекватности модели и понимания сущности
3. Динамика скорости перемещений пропор-
результатов моделирования выполнен ряд вычис-
циональна градиенту зоны стимула и вязкости
лительных экспериментов на примере терпуга
среды.
БИОФИЗИКА том 68
№ 1
2023
136
ЧЕТЫРБОЦКИЙ и др.
4. Длительность преднерестового адаптацион-
нерест, после которого они направлено следуют
ного этапа t*, по его завершению рыбы поднима-
за самцами.
ются в зону нереста.
7. На границах среды отсутствуют потоки ком-
5. Поскольку перед нерестом самцы прогоня-
понентов системы.
ют от нерестовых кладок других рыб, то на этом
8. Для динамических переменных заданы на-
этапе самцы конкурируют между собой за участ-
чальные распределения.
ки мест кладок.
6. По сравнению с самцами у самок отмечается
Согласно принятым допущениям, запись
несколько иной переход из зоны адаптации на уравнений модели принимает вид
∂
t
M
=∂
x
(U
M
M)
-β
M
M
+
D
∂M xx
M,
∂
t
F
=∂
x
(U
F
F)
-β
F
F
+
D
∂
F xx
F
,
(U )
∂U
=∂
(
k
R
−
k
M)
+
D
∂
U
,
t
M
x
11
12
M xx M
(1)
(U )
∂
U
=∂
(
k
R
+
k
M)
+
D
∂
U
,
t
F
x
21
22
F xx F
∂
x
M
|
a,
b
=∂
x
F
|
a,
b
=
0
и
∂
U
x M a
|
,b
=∂
U
x F a
|
,b
=
0,
M
(x,0)
=
M x)
,
F
(x
,0
)
=
F x)
,
U
(
x
,0)
=
U
(x
,0)
=U x)
0
0
M
F
0
где x и t - пространственная и временная коорди-
висимость отражает неравномерность вклада сти-
наты; M(x,t), UM(x,t), F(x,t), UF(x,t) - плотности
мулов в динамику системы уравнений (1). Коэф-
распределений, скорости самцов и самок; βM и
фициенты k11 и k21 характеризуют степень
βF - коэффициенты естественной смертности;
«привлекательности» стимулов, а k12 и k22 - про-
(U
)
странственную конкуренцию самцов за нересто-
R(x,t) - функция зоны стимулов; DM, DF,
D
,
M
вые участки и направленное движение самок
(U
)
D
- коэффициенты диффузии (первый под-
F
вслед за самцами уже непосредственно при нере-
строчный индекс указывает на соответствующую
сте.
переменную: M - самец, F - самка); {kij, i,j = 1,2} -
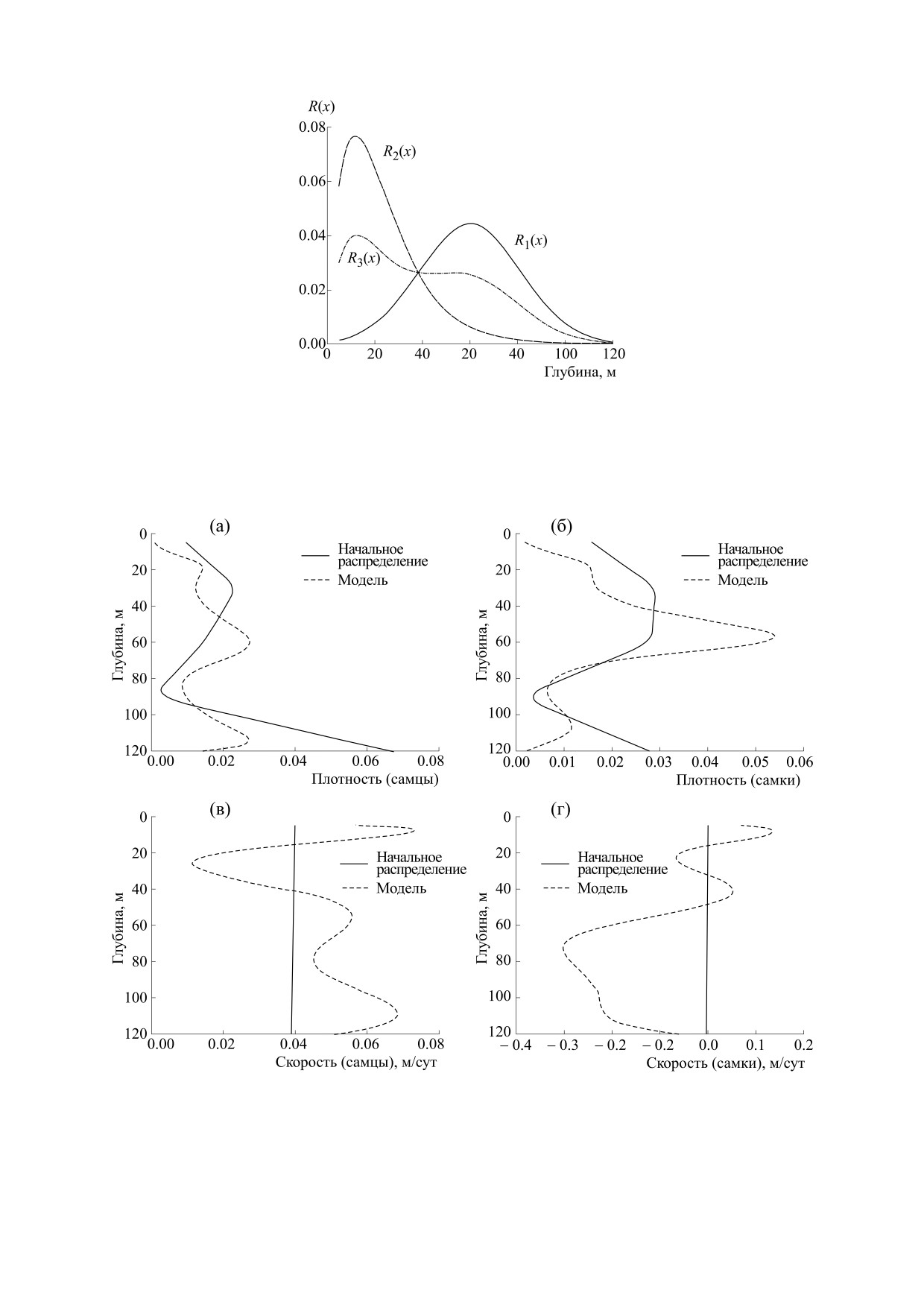
Функции зоны стимулов {Ri, i = 1÷3} опреде-
зависимые от времени неотрицательные коэффи-
циенты пропорциональности. Их временная за- ляются таким представлением:
*
R x), при
t
<
t
,
1
*
R x)
=
R
(
x)
, самцы при
t
>
t
,
2
*
3
R x)
=
[
1
R x)
+
2
R x)] /2, самки при
t
>
t
,
где характер распределения каждого из них в наи-
что обычно выражается в окраске отдельных ча-
большей степени соответствует наблюдениям;
стей тела рыб. В частности, у терпуга - это напо-
R1(x) характеризует общую для самцов и самок
минающая поднятое забрало черная маска [13].
зону стимула на этапе адаптации, а R2(x) и R3(x) -
Численное решение системы уравнений (1)
их зоны стимулов переходов на этап нереста. От-
выполняется матричной прогонкой, вычисли-
личное от нуля начальное движение рыб U0(x) ≠ 0
тельная схема которой следует записи
обусловлено гормональным фоном их организма,
t)
(t+1)
t)
(t+1)
t)
(t+1)
t)
A
i
Y
i−1
-
B
i
Y
i
+C
i
Y
i+1
=-F
i
,
где i -номер узла схемы (I = 1 ÷ n, N - число уз-
(t+1)
(M)
(F )
'
Y
i
=
(M
i,t+1
,F
i,t+1
,U
it+1
,U
i,t+1
)
,i
=
2
÷
N.
t)
t)
t)
лов);
{A
i
,B
i
,C
i
i
= 2 ÷ N}
- набор матриц 4 × 4;
Элементы матриц следуют выражениям
БИОФИЗИКА том 68
№ 1
2023
ЧИСЛЕННОЕ МОДЕЛИРОВАНИЕ ПРЕДНЕРЕСТОВОЙ И НЕРЕСТОВОЙ МИГРАЦИЙ
137
t)
Δt
(M)
Δt
Δt
(F )
Δt
Δt
(M)
Δt
(F)
A
=
diag
U
+
D
,
U
+
D
,,
D
,
D
i
i−1/2,t
2
M
i-1/2,t
2
F
2
U
2
U
Δx
Δx
Δx
Δx
Δx
Δx
Δt
(M)
Δt
Δt
(F)
Δt
B
1,1
=
U
i+1/2,t
+1+
2
D
M
+β Δ
M
t,
B
2,2
=
U
i+1/2,t
+1+
2
D
F
+β Δ
F
t
2
2
Δx
Δx
Δx
Δx
Δt
(M)
Δt
(F )
B
=
1+
2
D
+,
B
=
1+
2
D
3,3
2
U
4,4
2
U
Δ
x
Δx
,
Δt
(M)
Δt
(F )
(2)
B
=
1+
2
D
+,
B
=
1+
2
D
3,3
2
U
4,4
2
U
Δ
x
Δx
Δt
Δt
A
=
k
,
B
=-A
,
A
=-
k
,
B
=-A
3,1
1,2
3,1
3,1
4,1
2,2
4,1
4,1
Δx
Δx
t)
(M)
Δ
t
(F
)
Δt
'
F
=
(M
,
F
,U
+
k
ΔR,
U
+
k
ΔR
)
i
i,t+1
i,t+
1
it+1
1,1
i
i,t
+1
2,1
i
Δ
x
Δx
где функция diag(·) - функция построения диаго-
гнормальному распределению [14, 15]. Примени-
нали матрицы, Δt и Δx - размеры вычислитель-
тельно для R(x) здесь уместно особенно подчерк-
ной сетки.
нуть именно смысловой характер указанных рас-
пределений: в первом случае это распределение
обусловлено влиянием многих слабо взаимозави-
ВЫЧИСЛИТЕЛЬНЫЕ ЭКСПЕРИМЕНТЫ
симых случайных величин, каждая из которых
Детализация функций и коэффициентов си-
вносит малый вклад относительно общей суммы;
стемы уравнений (1) здесь выполнена на основа-
основание формирования логнормального рас-
нии усредненных многолетних наблюдений
пределения случайных величин составляет про-
преднерестовых и нерестовых миграций однопе-
цесс, где действие каждого добавочного фактора
рового терпуга. Учитываются также наблюдае-
на случайную величину пропорционально ее до-
мые условия его жизненного цикла. Так, R(x) из
бавочному уровню [16]. По-видимому, такая си-
выражений (2) в зоне адаптации [x1,x2] оказыва-
туация отражает эволюцию рыб. Согласно ска-
ется близким к нормальному распределению, а в
занному, здесь принимается следующая аппрок-
прибрежной зоне нереста [x0,x3] - близким ло-
симация этих функций
2
-2
*
R x)
=
A
exp-a·(x
-
x
)
(x
-
x
)
, при
t
<
t
,
1
1
1
2
1
*
R(
x,
t)
=
2
R x)
=
A
2
xexp(
−bx), самцы при
t
>
t
,
(3)
*
3
R x)
=
[
1
R x)
+
2
R x)] /2, самки при
t
>
t
,
где коэффициенты А1, А2 и а вычисляются на ос-
которая по своей форме во многом подобна этому
представлению.
новании наблюдений. В частности, для терпуга
интервал зоны нагула и адаптации рыб х1 = 40 м,
В последующих вычислениях для каждого из
распределений {Ri(x), i = 1÷3} выполнена единич-
х2 = 90 м, а зоны нереста х0 = 5 м, х3 = 25 м [17];
ная нормировка (сумма его членов приведена к
коэффициенты А1 = 1 ед. (единиц) и а =0.25 оце-
единице). Нормированные распределений зон
нивали подбором; коэффициенты А2
=
стимулов модели (1) представлены на рис. 1.
= 14.8147 ед.·м-1 и b = 0.085 м-1 оценивали мето-
Границы изменения аргументов функций зон
стимулов расширены на весь диапазон глубин,
дом наименьших квадратов на основании линей-
что не соответствует принятым на практике пред-
ного соотношения, которое получено после взя-
ставлениям (обычно указываются конкретные
тия логарифма от обеих частей выражения для
значения [13]). Такая ситуация обусловлена не-
R2(x). Для представления R2(x) вместо логнор-
значительными отличиями этих функций от нуля
мального представления функции зоны стимула
вне указанных интервалов, так и нахождения рыб
используется более простая кривая Рикера [18],
в других глубинных пределах. Так, определенная
БИОФИЗИКА том 68
№ 1
2023
138
ЧЕТЫРБОЦКИЙ и др.
Рис. 1. Нормированные распределения зон стимулов в модели (1).
Рис. 2. (а)-(г) - Плотности модельных распределений и скорости перемещений: на 60-е сутки моделирования.
БИОФИЗИКА том 68
№ 1
2023
ЧИСЛЕННОЕ МОДЕЛИРОВАНИЕ ПРЕДНЕРЕСТОВОЙ И НЕРЕСТОВОЙ МИГРАЦИЙ
139
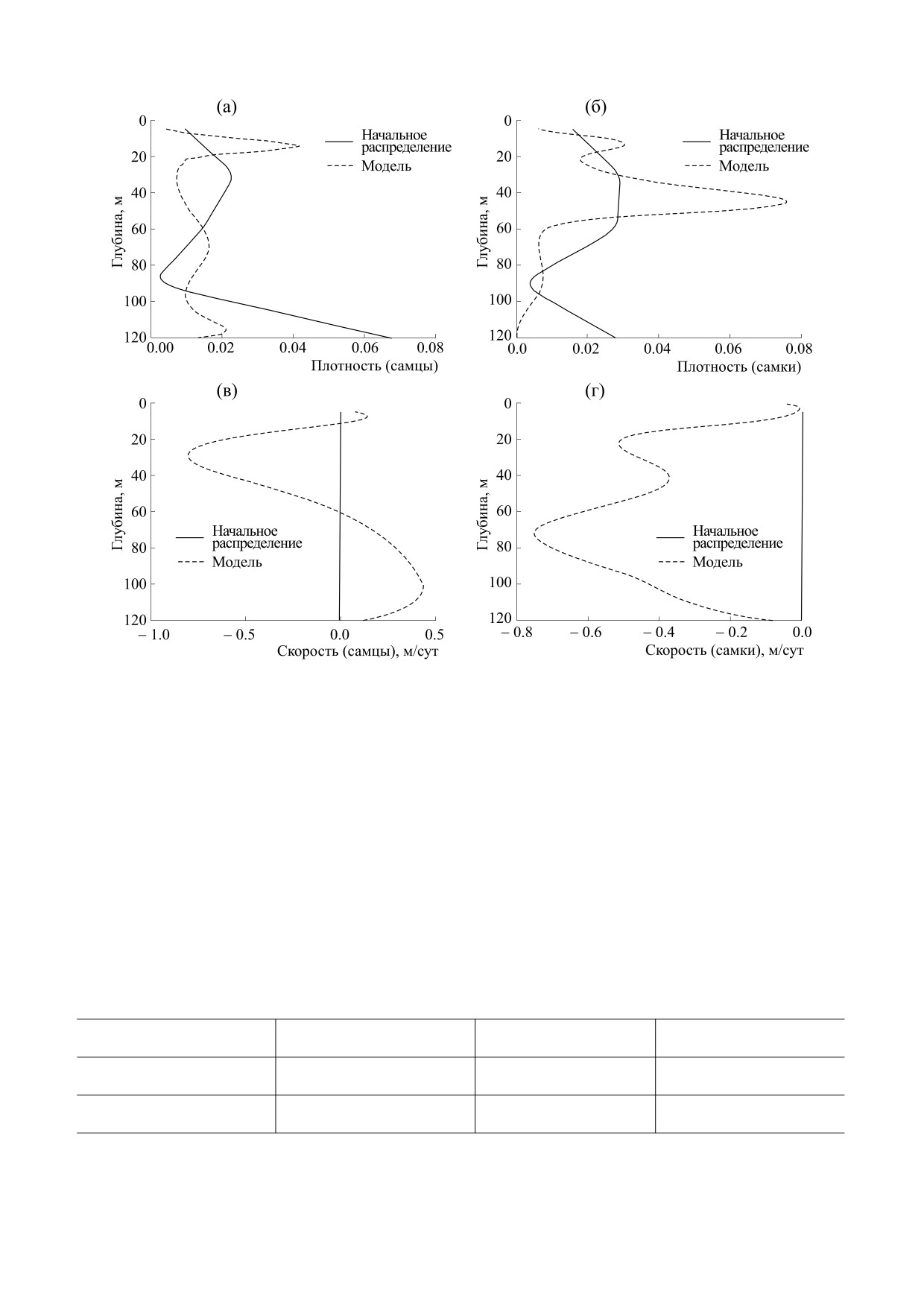
Рис. 3. (а)-(г) -Плотности модельных распределений и скорости перемещений: на на 110-е сутки моделирования.
доля половозрелых рыб не участвует в нересте,
чение плотности в районе 20 м и направление
что связано с патологиями половых продуктов.
суточных перемещений/скоростей самцов. На
60-е сутки самки в основном еще пребывают в зо-
Численные значения параметров модели (1)
не их преднерестовой адаптации (рис. 2б,г). Тогда
представлены в табл. 1. Плотности модельных
же начинается их перемещение в зону нереста. На
распределений и скорости перемещений, соот-
110-е сутки (рис. 3) основная часть самцов нахо-
ветствующие разным периодам моделирования,
дится в таком интервале глубин нереста, где отме-
представлены на рис. 2 и 3.
чается низкая скорость их перемещений. Для са-
Анализ представленных на графиках кривых
мок отмечается аналогичная картина. Имеется
также определенное запаздывание по временам
дает следующую предметную интерпретацию.
следования самок на нерест.
Для самцов 60-е сутки (рис. 2а,в) являются про-
должением их движения из зоны адаптации в
Сопоставление кривых на рис. 2 и 3 показыва-
зону нереста, на что указывает повышенное зна- ет, что длительность введенного здесь адаптаци-
Таблица 1. Численные значения параметров модели
Δt = 1 сутки
DM = 0.95 м2 сутки-1
D(U)M = 2.1 м2 сутки-1
k11 = 9.8 сутки-1
Δх = 5 м
DF = 0.9 м2 сутки-1
D(U)F = 2.1 м2 сутки-1
k21 = 9.8 сутки-1
βM = 10-3 сутки-1
βF = 10-3 сутки-1
k12 = 10-5 сутки-1
k22 = 0.7 сутки-1
БИОФИЗИКА том 68
№ 1
2023
140
ЧЕТЫРБОЦКИЙ и др.
онного этапа для самок на 20-30% выше, чем у
БЛАГОДАРНОСТИ
самцов. Представляется, что это связано с осо-
Авторы выражают признательность д.б.н.
бенностями формирования половых продуктов
В.В. Суханову и к.б.н. В.Е. Гомелюк за многочис-
самок, массы гонад которых в несколько раз пре-
ленные обсуждения и консультации по рассмат-
восходят масса гонад самцов [19].
риваемым в работе вопросам.
Изменение направлений суточных скоростей
(с плюса на минус и обратно) отражает рыскание
КОНФЛИКТ ИНТЕРЕСОВ
рыб в зонах их адаптации и стимулов, которое вы-
звано неоднородностью морской среды и гетеро-
Авторы заявляют об отсутствии конфликта
генностью популяционной структуры.
интересов.
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
ЗАКЛЮЧЕНИЕ
Настоящая работа не содержит описания ис-
Разработана компактная 1D численная модель
следований с использованием людей и животных
преднерестовой и нерестовой миграций рыб, ко-
в качестве объектов.
торая основана на уравнениях системы Патлака-
Келлера-Сегеля, введен этап преднерестовой
адаптации рыб, введена зон действия стимулов. В
СПИСОК ЛИТЕРАТУРЫ
модели в рамках единого механизма реализуются
1.
В. В. Абрамов, Зоологич. журн., 32 (6), 1198 (1953).
следующие процессы: преднерестовая адаптация
2.
С. И. Павлов, Изв. Самарского науч. центра РАН,
рыб, конкуренция самцов за участки нереста; на
11 (1), 34 (2009).
этапе нереста самки направлено следуют за сам-
3.
A. Toleuchanov, M. Panfilov, and A. Karatauev, in
цами (таксис). Разработан механизм коммуника-
Communications in Computer and Information Science.
ции, согласно которому самцы первыми следуют
Mathematical Modeling in Technological Processes
на нерестовые участки, что стимулирует последу-
(Springer, 2015), Vol. 549, pp. 177-189.
ющее перемещение самок.
4.
Ю. Н. Тютюнов, А. Д. Загpебнева, Ф. А. Cуpков и
А. И. Азовcкий, Биофизика, 54, (3), 508 (2009).
Следуя положениям модели, массовые пере-
5.
N. Takajoshi, Res. Popul. Ecd., 35 (1), 45 (1993).
мещения рыб обусловлены фиксированными в
водных глубинах градиентами зоны действий
6.
В. Н. Говорухин, А. Б. Моргулис и Ю. В. Тютюнов,
Докл. РАН, 372 (6), 730 (2000).
стимулов, в качестве которых выступают ком-
фортные для преднерестовой адаптации глубины,
7.
E. F. Keller and L. A. Segel, J. Theor. Biol., 30, 225
(1971).
хорошо аэрированные и удобные для нереста вы-
ступы участков дна, распределения самцов. Ана-
8.
А. В. Никитина, Изв. ЮФУ. Технические науки,
№ 7 (96), 173 (2009).
лиз результатов показывает: при нерестовой ми-
грации самцы перемещаются быстрее самок, хотя
9.
Y. V. Tyutyunova, L. I. Titovab, and I. N. Senina, Ecol.
Complexity 31, 170 (2017).
и дольше на находятся на нерестилищах; самые
медленные перемещения отмечены для промежу-
10.
В. Н. Говорухин и А. Д. Загребнева, Компьютер-
ные исследования и моделирование, 12 (4), 831
точной зоны, где проходит адаптация; непрерыв-
(2020).
ность переходов между этапами; длительность
11.
А. Б. Моргулис и К. К. Ильин, Вестн. ВГУ. Сер.
введенного здесь адаптационного этапа на 20-
Физика. Математика, № 2, 114 (2019).
30% выше, чем у самцов. Результаты численных
12.
А. Б. Моргулис, Вестн. ЮУрГУ. Сер. «Математика.
экспериментов показали непрерывность перехо-
Механика. Физика», 11 (3), 28 (2019).
дов между этапом преднерестовой адаптации рыб
13.
В. Н. Гомелюк, Вопросы ихтиологии, 27 (6), 991
и этапом нереста.
(1987).
Согласно построенной модели функции зоны
14.
R. De Graaf and P. Dam, in Salish Sea Ecosystem Conf.
адаптации и зон стимулов нереста определены на
(2014) p.
всем интервале глубин. Такое допущение приво-
Day1/6.
дит к разнонаправленности суточных скоростей,
15.
W. E. Ricker, Computation and Interpretation of Biologi-
которые интегрально обусловливают основное
cal Statistics of Fish Populations. Bull. 191 (Ottawa,
направление к нерестовой зоне. Принятия такого
1975).
допущения оказывается отражением природного
16.
А. Н. Колмогоров, в сб. Теория вероятностей и ма-
факта пропуска нереста части половозрелых рыб.
тематическая статистика (Наука, М., 1986).
17.
А. Н. Вдовин, Изв. ТИНРО, 123, 16 (1998).
Представляется, что введение в рассмотрение
функций распределений стимулов имеет опреде-
18.
M. D. Robarts and J. F. Piatt, J. Fish Biol., 54 (5), 1050
ленное применение при моделировании про-
(2005).
странственно-временной динамики биологиче-
19.
А. Н. Вдовин и А. Н. Четырбоцкий, Труды ВНИ-
ских объектов.
РО, 170, 26 (2018).
БИОФИЗИКА том 68
№ 1
2023
ЧИСЛЕННОЕ МОДЕЛИРОВАНИЕ ПРЕДНЕРЕСТОВОЙ И НЕРЕСТОВОЙ МИГРАЦИЙ
141
Numerical Modeling of Pre-Spawning and Spawning Migrations of the Representative
of the Family Hexagrammidae: The Case of the Arabesque Greenling
A.N. Chetirbotsky*, A.N. Vdovin**, and V.A. Chetirbotsky***
*Far East Geological Institute, Far Eastern Branch of the Russian Academy of Sciences,
prosp. 100-letiya Vladivostoka 159, Vladivostok, 690022 Russia
**Pacific Branch of Research Institute of Fishery and Oceanography (“TINRO”),
per. Shevchenko 4, Vladivostok, 690091 Russia
***Faculty of Physics, Lomonosov Moscow State University, Leninskie Gory 1/2, Moscow, 119991 Russia
This paper presents a spatio-temporal model of greenling dynamics during pre-spawning and spawning mass
migrations. The developed model is based on original verified long-term observations and data on industrial
fishing in the Peter the Great Bay (Sea of Japan), and the equations of the dynamics of the density of
males/females and the mass daily movements of fish are written in terms of the transfer equations. These
equations are written in the form of the modified Patlak-Keller-Segel equations, according to which the
f low of objects/substances is directed along the gradients of stimulus introduced. It is believed that in the pre-
spawning season, adaptation in morpho-physiological (biochemical thermoregulation and a number of other
endogenous processes) and behavioral responses of fish to sufficiently long and energy-consuming spawning
can occur, where the stimulus for mass movements of fish is optimal environmental conditions for spawning
events. During the spawning period, selected sites will be found in convenient and well-aerated embayments
located at the bottom of reservoir in the coastal area. Modeling of stimuli-related movements is performed
based on information about the preferred water depths of the fish's pre-spawning area and relevant features
relating to a selection of suitable bedding sites. It is assumed that the intensity of daily motion is proportional
to their linear size (the larger the fish becomes, the faster it is). The equations for the spawning stage take into
account the spatial competition of males, but in natural conditions it is observed only in the vicinity of spawn-
ing areas. Being away from these areas, males continued to look for new sites good for the spawn. For females,
their movement is provoked by males that assembled in schools, the signal of which can be certain chemical
elements released by males (for example, mucus secretion from males) or visual contact. The diffusion of fish
distribution and the viscosity of the habitat (velocity diffusion) are taken into account. The initial distribution
of fish is given according to the average distribution of fish in July over a long-term observation period in the
Peter the Great Bay.
Keywords: fish migration, spawning, Patlak-Keller-Segel equations, adaptation zone
БИОФИЗИКА том 68
№ 1
2023