БИОФИЗИКА, 2023, том 68, № 4, с. 736-747
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 591.148:593.8(262.5)
БИОЛЮМИНЕСЦЕНЦИЯ ГРЕБНЕВИКА
Pleurobrachia pileus (O.F. Müller, 1776) В ЛЕТНИЙ СЕЗОН
© 2023 г. А.В. Темных*, #, М.И. Силаков*, О.В. Машукова*
*ФИЦ «Институт биологии южных морей им. А.О.Ковалевского РАН»,
просп. Нахимова, 2, Севастополь, 299001, Россия
#E-mail: atemnykh@yandex.ru
Поступила в редакцию 31.12.2022 г.
После доработки 30.03.2023 г.
Принята к публикации 05.04.2023 г.
Способность к биолюминесценции обнаружена у многих морских гидробионтов, в том числе прак-
тически все представители типа гребневиков (Ctenophora) обладают способностью излучать свет.
Свечение видов и биолюминесцентных субстратов, выделенных из них, активно используется в
биологии, экологии и медицине для исследования физиологического состояния организма или вы-
явления неблагоприятных параметров среды. Параметры свечения представителей гребневиков
Черного моря Mnemiopsis leidyi A. Agassiz, 1865 (Lobata) и Beroe ovata Bruguière, 1789 (Beroida) исполь-
зуются в изучении влияния токсикантов, таких, как жидкие углеводороды, полихлорированные би-
фенилы, тяжелые металлы. Способность к биолюминесценция автохтонного обитателя Черного
моря - гребневика Pleurobrachia pileus (O.F. Müller, 1776) - вызывала сомнение у некоторых иссле-
дователей. Целью данной работы являлось подтверждение способности свечения данного вида и
выбор оптимального времени для проведения экспериментов с ним. Было выявлено, что биолюми-
несцентную реакцию на механическую и химическую стимуляцию проявляют все размерные груп-
пы данного вида в любое время суток. При химической стимуляции величины энергии сигнала вы-
ше в несколько раз, чем при механической стимуляции. Выявлено, что оптимальное время для про-
ведения биофизических исследований - период с 20:00 до 6:00, так как энергетические показатели
биолюминесцентного сигнала в это время максимальны в пределах суток.
Ключевые слова: Ctenophora, люминесценция, циркадный ритм, Черное море.
DOI: 10.31857/S0006302923040142, EDN: KMUHYX
Способность испускать свет в результате био-
друг от друга и не закреплялись эволюционно [2].
химической реакции называется биолюминес-
Это привело к тому, что ферменты и субстраты
ценцией и, в отличие от других видов люминес-
этих систем различны и сейчас известно уже о
ценции, таких как флуоресценция, термохеми-
тридцати типах биолюминесцентных систем,
люминесценция и т.д., присуща только живым
только десять из которых хорошо изучены [5].
организмам, имеющим специализированный
Кроме того, отличаются по характеру и люминес-
ферментативный аппарат или биосубстрат [1]. В
центные структуры (фотофоры) и фотоциты -
ходе химической реакции люциферин (субстрат)
клеточные типы, ответственные за эти процессы.
окисляется с помощью молекулярного кислорода
Это влияет на длину волны испускаемого света,
под действием люциферазы (фермента) с образо-
которая находится в синем диапазоне у морских
ванием органических пероксидных соединений,
микроводорослей динофлагеллят, в зеленом - у
распад которых приводит к возбуждению моле-
желетелого зоопланктона [6] и т.д. Соответствую-
кул вещества, ответственных за светоизлучение
щие этим цветам энергии фотонов составляют от
[2].
40-70 ккал на 1 эйнштейн, что значительно пре-
Биолюминесценция встречается у эволюци-
вышает энергию большинства биохимических
онно разнородных групп организмов [2-4], в том
реакций, в том числе и распад высокоэнергетиче-
числе у некоторых бактерий, грибов, водорослей,
ской молекулы АТФ (7 ккал). Большинство мор-
кишечнополостных, червей, моллюсков, насеко-
ских биолюминесцентных организмов в природе
мых и рыб, но не наблюдается у более высокоор-
генерируют световые вспышки в ответ на внеш-
ганизованных животных. Биолюминесцентные
ние, в основном, механические либо химические
системы возникали у организмов независимо
воздействия.
736
БИОЛЮМИНЕСЦЕНЦИЯ ГРЕБНЕВИКА
737
Биолюминесценция является важным элемен-
(Cidippida), как и других видов рода Pleurobrachia
том функционирования пелагических сообществ
(Cidippida), является спорным моментом. Так,
[3, 4, 7]. Это связано с экологической ролью света
для акватории Средиземного моря и берегов Се-
в жизни гидробионтов. Биолюминесценция ис-
верной Америки возможность излучать свет для
пользуется для внутривидовой и межвидовой
P. bachei A. Agassiz, 1860, P. rhodopis Chun, 1879 и
коммуникации, для привлечения жертвы и отпу-
P. pileus не только ставится под сомнение, но и ка-
гивания хищников. Так, для фитопланктонных
тегорически заявляется о невозможности этих
организмов биолюминесценция выполняет за-
видов к биолюминесценции [29]. Другие исследо-
щитную функцию. Светящиеся виды микроводо-
ватели [14, 23, 30] полагают, что P. pileus является
рослей выедаются в меньшей степени, чем несве-
биолюминесцентным видом. Так, проведенные
тящиеся [8]. Это также относится и к некоторым
лабораторные исследования над черноморской
видам копепод, например Metridia lucens Boeck,
популяцией P. pileus в холодное время года пока-
1865 [9]. Кроме того, неравномерное затухание и
зали, что стимулирование химическим и механи-
рассеивание различных длин волн оптического
ческим способом представителей этого вида вы-
спектра в водной среде снижают эффективность
зывает биолюминесцентный отклик [30]. Таким
света на глубинах, превышающих уже первые де-
образом, биолюминесцентный сигнал этого вида
сятки метров [10, 11]. Таким образом, ниже фоти-
также может быть использован для анализа со-
ческого слоя биолюминесценция может служить
стояния организма при влиянии на него различ-
средством обмена информацией между гидро-
ных абиотических и антропогенных факторов. На
бионтами.
данный момент многие аспекты биолюминес-
ценции P. pileus не известны.
В настоящее время уже выявлены зависимости
временных и энергетических параметров свето-
Целью данной работы являлось выявление
излучения от физиологического состояния ряда
энергетических и временных параметров биолю-
организмов [12-14]. Выяснилось, в частности,
минесцентного сигнала и его циркадного ритма у
что влияние может оказывать отклонение темпе-
Pleurobrachia pileus в летний период для выбора
ратуры от оптимальной для жизнедеятельности
оптимального времени проведения исследований
данного организма [15, 16], изменение уровня и
по влиянию различных факторов на его физиоло-
оптического спектра освещенности [17-21], со-
гическое состояние.
лености среды [22] и т.д. При этом, как считается,
естественные колебания перечисленных факто-
МАТЕРИАЛЫ И МЕТОДЫ
ров не являются для организма экстремальными,
поскольку составляют экологическую нишу вида
Биологический материал для исследования
и определяют его популяционный гомеостаз.
параметров свечения Pleurobrachia pileus собран
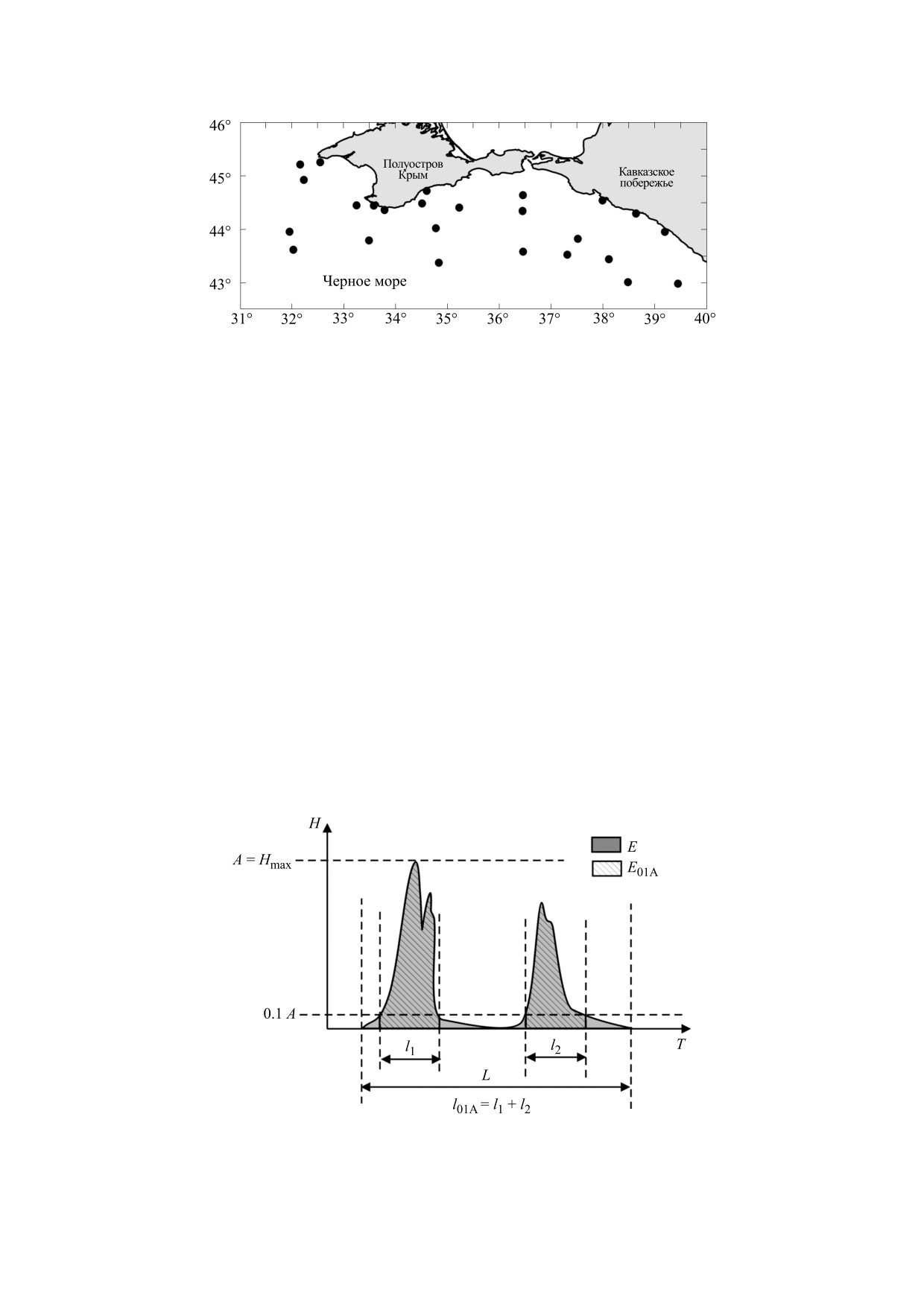
на 26 станциях в научном рейсе №102 НИС «Про-
Способность к биолюминесценции, обнару-
фессор Водяницкий» в период с 10.06.18 г. по
женная у многих морских гидробионтов [2, 4, 18,
01.07.18 г. (рис. 1). Для сбора проб использовали
23], выявлена и у представителей желетелого мак-
планктонную сеть Джеди (ячея 115 мкм, входное
ропланктона - гребневиков (Ctenophora). Прак-
тически все представители гребневиков являются
отверстие 0.1 м2) и макропланктонную сеть Бого-
биолюминесцентными видами с хорошо изучен-
рова-Раса (ячея 400 мкм, входное отверстие
ными параметрами светового сигнала [6, 24].
0.5 м2).
Представители этого типа животных светятся зе-
Из планктонной пробы изымали не помятые,
леными вспышками. В этом случае стимулято-
активные экземпляры Pleurobrachia pileus, изме-
ром, как и у большинства кишечнополостных,
ряли их и ранжировали по размеру в стеклянные
является ион Ca2+, реагирующий с люциферин-
стаканы емкостью 0.5-1.0 л, наполненные мор-
люциферазным пероксидным комплексом
-
ской водой, взятой с поверхности на той же стан-
экворином.
ции и профильтрованной через сито ячеей
35 мкм. Через 1 ч после вылова объекты помеща-
Три вида гребневиков, играющих важную роль
ли в темное помещение. Экспозиция проводи-
в экосистеме Черного моря и являющиеся доми-
лась через 2 ч после пребывания в темноте при
нирующими по биомассе видами, принадлежат к
температуре 21°С.
трем отрядам: Lobata, Beroida, Cidippida. Наибо-
лее полно изучена биолюминесценция двух ви-
Исследование параметров свечения проводи-
дов-вселенцев - Mnemiopsis leidyi A. Agassiz, 1865
ли на лабораторном биофизическом комплексе
(Lobata) и Beroe ovata Bruguière, 1789 (Beroida)
«Свет» [4] методом механической и химической
[24-28]. Параметры свечения этих видов исполь-
стимуляции отдельного объекта, помещенного в
зуются для оценки влияния токсических веществ
теневую камеру комплекса в кювете объемом
на состояние организма в биофизических иссле-
50 мл [14]. Приборный лабораторный комплекс
дованиях [26-28]. Светимость третьего абориген-
«Свет» включал высоковольтный блок питания
ного вида - Pleurobrachia pileus (O. F. Müller, 1776)
(ВС-22), люминескоп, состоящий из приемника
БИОФИЗИКА том 68
№ 4
2023
738
ТЕМНЫХ и др.
Рис. 1. Карта станций научного рейса № 102 НИС «Профессор Водяницкий», на которых выполнялся сбор Pleurobrachia
pileus для проведения эксперимента.
светового излучения (ФЭУ-71) и темновой каме-
Были исследованы 498 объектов размером
ры для объекта, а также регистрирующего устрой-
4.0-21.0 мм (орально-аборальная длина), ранжи-
ства - цифрового интерфейса.
рованных на 9 размерных групп: 4.5 ± 0.5, 6.5 ±
± 0.5, 8.5 ± 0.5, 10.5 ± 0.5, 12.5 ± 0.5, 14.5 ± 0.5,
В качестве механического стимулятора ис-
16.5 ± 0.5, 18.5 ± 0.5, 20.5 ± 0.5 мм. Размеры каж-
пользовали морскую воду (объем 2 мл), вводимую
дой особи определяли под бинокулярным микро-
с помощью поршневого устройства с созданием
скопом МБС-10 с точностью до 0.1 мм. Для визу-
турбулентного течения жидкости в кювете. Хими-
ального удобства в таблицах и графиках группы
ческую стимуляцию проводили добавлением 2 мл
названы по размеру меньших в них особей в мм
96%-го этанола ламинарным течением.
(размерная группа 4.5 ± 0.5 обозначена как груп-
па 4 мм и т.д.). Исследование диапазона измене-
Основными энергетическими параметрами
ния параметров биолюминесцентного сигнала у
биолюминесцентного сигнала являются ампли-
каждой размерной группы проводили в четные
туда (А) - максимальная энергия сигнала в еди-
часы суток: 02:00, 04:00, 06:00, …, 24:00. Экспери-
ницу времени отсчета, общая энергия сигнала (E)
менты по сравнению величин основных парамет-
и суммарная энергия сигнала, превышающая по-
ров биолюминесцентного сигнала между размер-
рог 0.1 от амплитуды (E0.1A). Временными пара-
ными группами проводили в утренние часы (8:00)
метрами выбраны общая длительность сигнала
и темное время суток (22:00).
(L) и суммарная длительность отдельных импуль-
сов биолюминесцентного сигнала (l0.1A) [3, 4, 12]
Детально суточную динамику параметров био-
(рис. 2).
люминесценции исследовали у самых многочис-
Рис. 2. Параметры биолюминесцентного сигнала.
БИОФИЗИКА том 68
№ 4
2023
БИОЛЮМИНЕСЦЕНЦИЯ ГРЕБНЕВИКА
739
Таблица 1. Обозначения анализируемых параметров биолюминесцентного сигнала и их расшифровка
№
Обозначение
Расшифровка
1
Aср.
Средняя по группе одноразмерных особей амплитуда
биолюминесцентного сигнала, мкВт∙см-2
2
Aинд.
Амплитуда биолюминесцентного сигнала отдельной особи в группе,
мкВт∙см-2
3
Lср.
Средняя по группе одноразмерных особей длительность сигнала, с
4
Lинд.
Длительность сигнала отдельной особи в группе, с
5
Eср.
Средняя по группе одноразмерных особей энергия сигнала, мкВт∙с∙см-2
6
Eинд.
Энергия сигнала отдельной особи в группе, мкВт∙с∙см-2
ленных экземпляров размером 10 мм каждые два
ставлены на рис 3, приведены данные для размер-
часа в течение 24 ч.
ной группы 16 мм.
Для анализа полученных результатов состав-
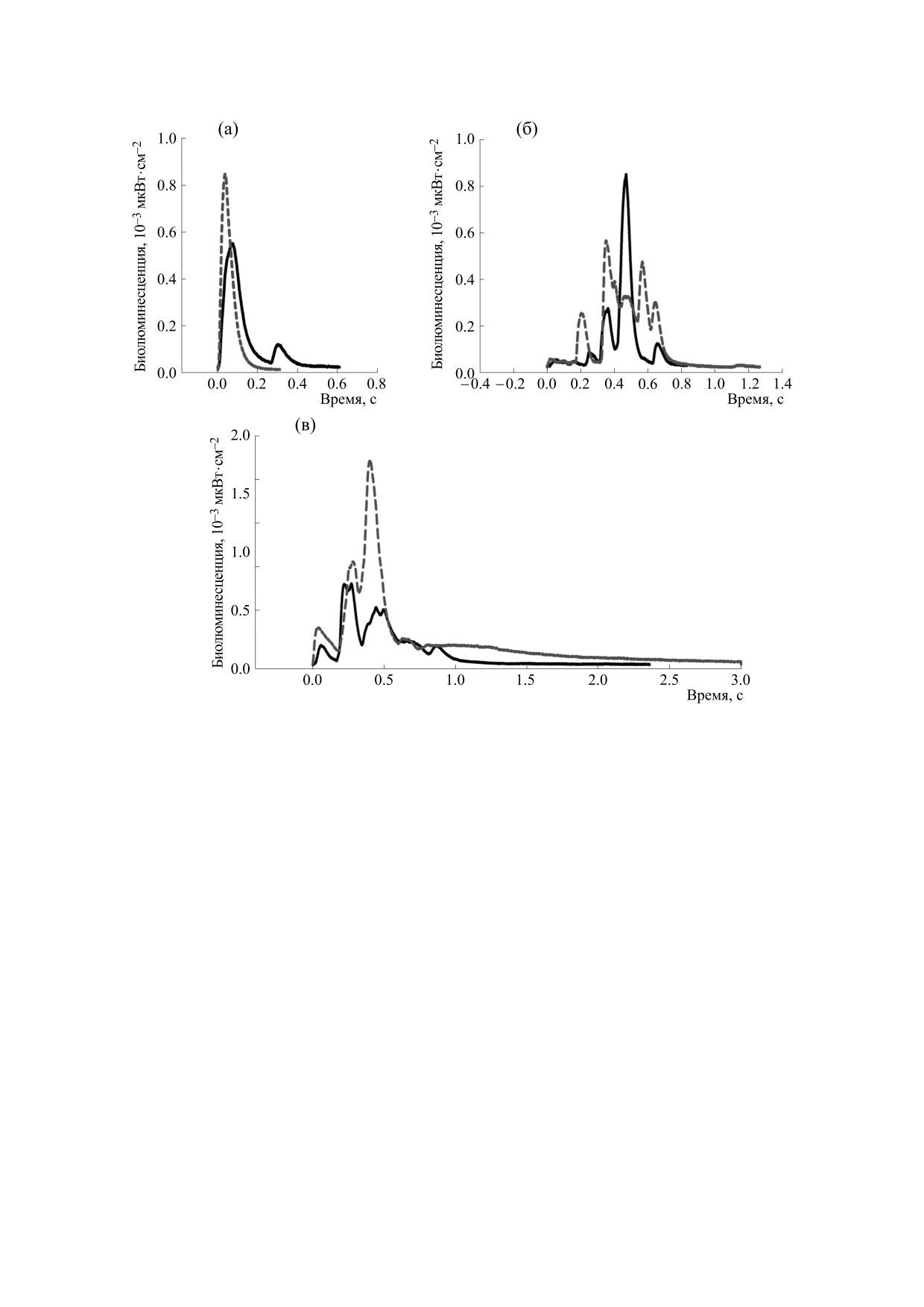
При механической стимуляции наблюдали два
ляли матрицы данных, вычисляли минимальные,
типа сигнала P. pileus, в первом случае регистри-
максимальные и средние величины энергетиче-
ровали резкий фронт нарастания амплитуды до
ских и временных параметров в каждой размер-
максимума, иногда с последующим вторым, су-
ной группе, а также медианы амплитуд размер-
щественно меньшим пиком, после чего шел та-
ных групп в программе MS Excel 2007 с помощью
кой же резкий краткий по времени спад (рис. 3а).
надстройки «Анализ данных».
Во втором случае сначала регистрировалось сла-
Расшифровка обозначений приведена в
бое увеличение амплитуды, низкие величины ко-
табл. 1.
торой сохранялись некоторое время, затем им-
пульс большой длительности со множественны-
Достоверность различия средних величин
ми пиками-вершинами (рис. 3б).
каждого параметра биолюминесцентного сигнала
между двумя видами стимуляции оценивали пу-
При химической стимуляции, после которой
тем вычисления критерия достоверности разли-
особь погибала, сигнал по форме напоминал вто-
чия:
рой упомянутый выше тип при механической
стимуляции, однако без первичного слабого уве-
x1
-
x
2
t
=
,
личения, амплитуда сразу достигала существен-
2
2
S
+
S
ных величин, при этом последний импульс часто
x1
x2
имел пологий и длительный фронт затухания
где t - критерий достоверности различия,
x
-
1
(рис. 3в).
среднее значение параметра при механической
Четкая связь между формой импульса и разме-
стимуляции,
x
- среднее значение параметра
2
рами экземпляров не обнаружена.
при химической стимуляции,
S
x
- средняя
1
Амплитуда и длительность биолюминесцентного
ошибка значение параметра при механической
сигнала у разноразмерных групп. Было выявлено,
стимуляции,
S
- средняя ошибка значение па-
x2
что величины амплитуды биолюминесцентного
раметра при химической стимуляции.
сигнала различаются при механическом и
Если значение t < 2, то различие между двумя
химическом типах стимуляции. Средняя по
выборками считается недостоверным, если t ≥ 2,
всем размерным группам амплитуда Аср при ме-
то различие между двумя выборками достоверно
ханической стимуляции колебалась от 0.2 до
на 95%.
1.4∙10-3 мкВт∙см-2 (табл. 1). При этом в группах с
Расчет доверительных интервалов проводили
размерами 4-5 мм и 8-19 мм величина Аср нахо-
по доверительной вероятности. Ширина довери-
дилась в пределах (0.6-0.9)∙10-3 мкВт∙см-2, а ми-
тельного интервала при доверительной вероятно-
нимальное и максимальное значение Аср зареги-
сти 0.95 составила 1.96‧S
, где
S
- средняя
x
x
1
стрированы в группе самых крупных особей и
ошибка значения параметра.
группе 6-7 мм соответственно. При химической
стимуляции минимальная по группам средняя
РЕЗУЛЬТАТЫ
амплитуда Аср зарегистрирована в группе с наи-
меньшими размерами - 1.21, максимальная -
Форма биолюминесцентного импульса. Форма и
типы биолюминесцентного сигнала у индивидуу-
2.26∙10-3 мкВт∙см-2 - в группе с размерами 6-7 мм.
мов P. pileus при обоих видах стимуляции пред-
У самых крупных экземпляров амплитуда сигнала
БИОФИЗИКА том 68
№ 4
2023
740
ТЕМНЫХ и др.
Рис. 3. Формы биолюминесцентных импульсов P. pileus: (а) и (б) - при механическом типе стимуляции, (в) - при
химическом типе стимуляции. Сплошной и пунктирной линией обозначены разные формы сигнала для особей
размером 16 мм.
от 1.0 с (для группы 20-21 мм) до 2.4 с (для группы
у отдельных особей достигала 8.3∙10-3 мкВт∙см-2, но
4-5 мм), во всех остальных размерных группах
при наличии в группе слабо высветившихся осо-
бей средняя величина сигнала в этой группе со-
Lср составляла 1.6-1.9 с.
ставила Аср = 1.79∙10-3 мкВт∙см-2 (табл. 2).
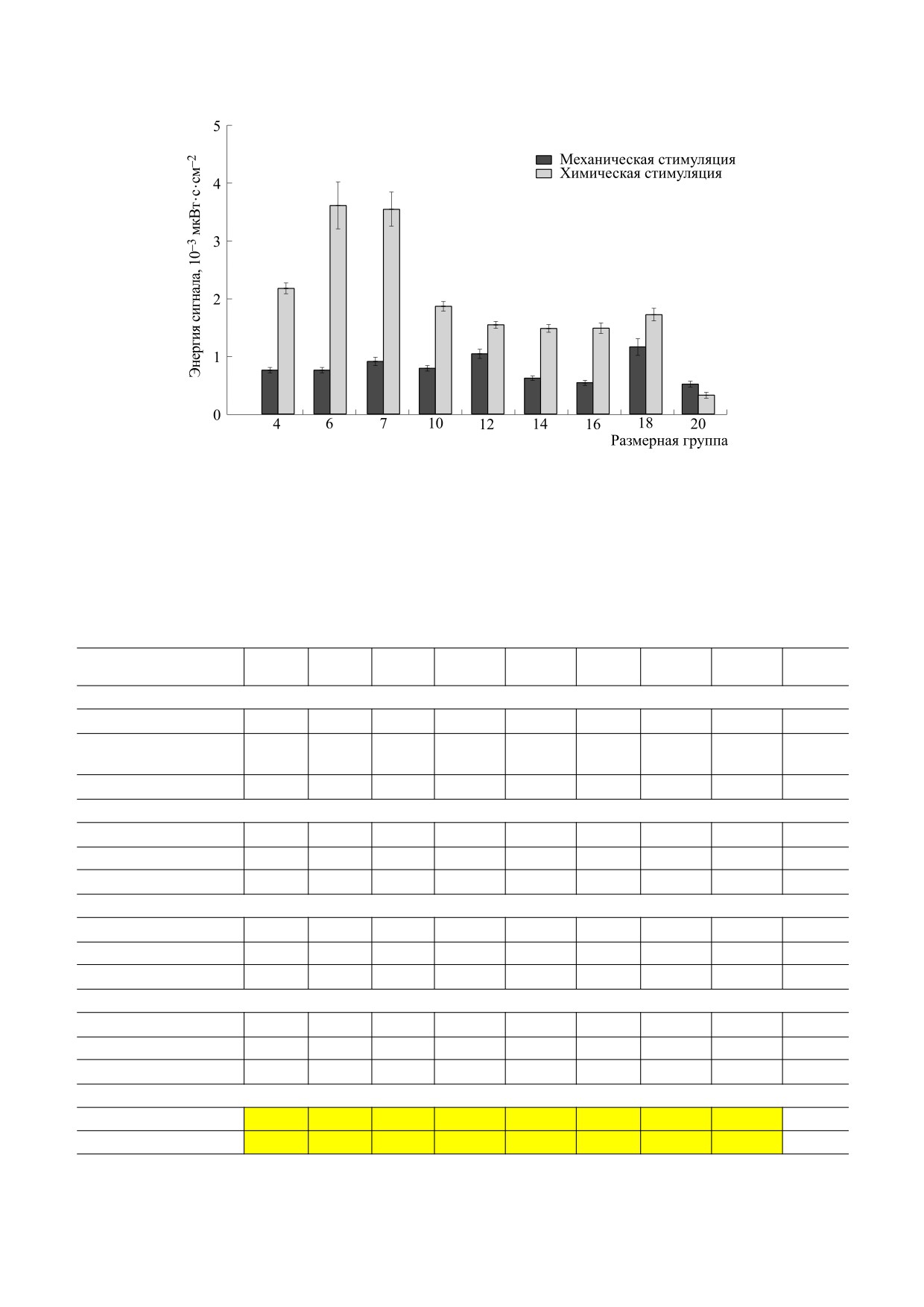
Энергия биолюминесцентного сигнала при
Средняя для всех размерных групп длитель-
механической стимуляции у разных групп слабо
ность сигнала Lср при механической стимуляции
отличалась друг от друга (рис. 4). Минимальные
значения зарегистрированы у группы самых
варьировала от 1.3 до 2.8 с (табл. 3). Наибольшие
крупных особей, наибольшие - у экземпляров
величины средней длительности сигнала зареги-
размером 18-19 мм. Энергия биолюминесцнтно-
стрированы у размерных групп с самыми малень-
го сигнала при химической стимуляции, напро-
кими и самыми крупными размерами, а мини-
тив, сильно различалась по группам. Наибольшие
мальная - у группы с размерами 6-7 мм. Макси-
мальная индивидуальная длительность сигнала
средние значения, выявленные у особей разме-
зарегистрирована в группе с размерами 16-17 мм
ром 6-7 мм, превышали минимальные, зареги-
(Lинд.max = 7 с).
стрированные у группы самых крупных P. pileus, в
семь раз. Величины энергии биолюминесцентно-
При химическом типе стимуляции макси-
го отклика при химической стимуляции, за ис-
мальные длительности сигнала в разных группах
ключением группы 20-21 мм, превышали тако-
достигали Lинд.max = 5-6 с, средняя по каждой
вые при механической стимуляции в полтора-че-
размерной группе длительность Lср изменялась
тыре раза (рис. 4).
БИОФИЗИКА том 68
№ 4
2023
БИОЛЮМИНЕСЦЕНЦИЯ ГРЕБНЕВИКА
741
Таблица 2. Амплитуда биолюминесцентного сигнала, ×10-3 мкВт см-2
Размерная группа
4
6
8
10
12
14
16
18
20
Механическая стимуляция
Среднее, Aср
0.59
1.37
0.80
0.73
0.76
0.89
0.62
0.94
0.20
Стандартная
0.09
0.18
0.11
0.07
0.09
0.09
0.08
0.20
0.03
ошибка
Медиана
0.45
0.88
0.41
0.56
0.57
0.75
0.54
0.74
0.16
Максимум, Aинд.max
1.21
4.81
3.34
2.56
2.81
2.47
1.60
2.72
0.35
Химическая стимуляция
Среднее, Aср
1.21
2.26
2.08
1.44
2.11
1.36
1.62
2.00
1.79
Стандартная
0.25
0.31
0.24
0.15
0.29
0.18
0.28
0.44
0.88
ошибка
Медиана
0.94
1.63
1.38
0.84
1.32
0.77
0.91
0.99
0.31
Максимум,
4.30
8.41
7.10
6.56
7.10
5.63
6.35
6.59
8.31
Aинд.max
Критерий достоверности различия
2.3
2.4
4.9
4.3
4.5
2.4
3.5
2.2
1.8
Параметры сигнала у разноразмерных групп в
химической (табл. 4). Наибольшие величины ам-
светлое и темное время суток. Были проведены ис-
плитуды отмечены у групп с размерами 12-13 мм
следования параметров биолюминесцентного
(утром) и 18-19 мм (ночью) при химической сти-
сигнала у разных размерных групп в утреннее
муляции.
время после восхода солнца (8:00) и ночное время
В светлое время суток при механической сти-
суток (22:00).
муляции средняя по всем группам амплитуда со-
ставляла 0.8∙10-3 мкВт∙см-2, максимальная была
Амплитуда биолюминесцентного сигнала у
зарегистрирована в группе 6-7 мм, и составила
всех групп при механической стимуляции и в
утреннее, и в ночное время была ниже, чем при
1.4∙10-3 мкВт∙см-2, в этой же группе наблюдали
Таблица 3. Длительность биолюминесцентного сигнала, с
Размерная группа
4
6
8
10
12
14
16
18
20
Механическая стимуляция
Среднее, Lср
2.37
1.25
1.85
1.87
1.71
1.36
2.08
1.76
2.82
Стандартная
0.51
0.18
0.25
0.23
0.26
0.21
0.41
0.44
0.87
ошибка
Медиана
1.43
0.78
0.97
0.81
0.81
0.69
1.02
0.94
2.10
Максимум, Lинд.max
5.99
4.99
5.99
5.99
5.99
5.99
6.99
5.19
5.61
Химическая стимуляция
Среднее, Lср
2.39
1.93
1.75
1.76
1.56
1.77
1.67
1.72
0.97
Стандартная
0.40
0.25
0.19
0.16
0.18
0.22
0.28
0.39
0.37
ошибка
Медиана
1.79
1.27
1.14
1.33
1.07
1.16
1.15
0.95
0.54
Максимум, Lинд.max
4.99
5.71
5.59
5.64
4.99
5.26
5.77
5.99
2.96
Критерий достоверности различия
0.0
2.2
0.3
0.4
0.5
1.3
0.8
0.1
2.0
БИОФИЗИКА том 68
№ 4
2023
742
ТЕМНЫХ и др.
Рис. 4. Средняя энергия биолюминесцентного сигнала каждой размерной группы при разных способах стимуляции.
максимальную индивидуальную амплитуду сиг-
ла незначительно выше утренней величины и со-
нала Аинд.max=3.8∙10-3 мкВт∙см-2.
ставила 0.9∙10-3 мкВт∙см-2. Максимальная сред-
няя и индивидуальная величины амплитуды, как
В темное время суток при механической сти-
и утром, зарегистрированы в группе 6-7 мм. В
муляции средняя по всем группам амплитуда бы- группе с самыми крупными экземплярами ам-
Таблица 4. Амплитуда биолюминесцентного сигнала, ×10-3 мкВт∙см-2
Размерная группа
4
6
8
10
12
14
16
18
20
Механическая стимуляция (8 ч)
Среднее, Aср
0.65
1.42
0.76
0.57
1.22
0.76
1.10
-
-
Стандартная
0.02
0.12
0.05
0.02
0.05
0.03
0.00
-
-
ошибка
Максимум, Aинд.max
1.21
3.76
2.80
2.77
1.95
1.20
1.14
-
-
Механическая стимуляция (22 ч)
Среднее, Aср
0.45
1.54
0.76
1.04
0.84
0.53
0.43
0.83
0.33
Стандартная ошибка
0.00
0.05
0.04
0.04
0.03
0.02
0.02
0.05
0.00
Максимум, Aинд.max
0.45
4.81
4.39
3.76
3.76
1.21
1.24
1.89
0.35
Химическая стимуляция (8 ч)
Среднее, Aср
1.18
1.49
1.31
1.42
2.43
1.69
1.16
-
-
Стандартная ошибка
0.03
0.07
0.04
0.03
0.12
0.02
0.10
-
-
Максимум, Aинд.max
2.12
3.76
2.97
4.10
4.34
1.93
3.11
-
-
Химическая стимуляция (22 ч)
Среднее, Aср
1.41
1.96
1.48
0.79
2.15
1.13
1.63
2.27
1.57
Стандартная ошибка
0.00
0.04
0.04
0.02
0.08
0.04
0.04
0.16
0.12
Максимум, Aинд.max
1.41
3.64
3.76
2.73
7.10
4.08
3.47
5.48
2.73
Критерий достоверности различия при разных типах стимуляции
8 ч
16.0
0.5
8.9
22.5
9.4
26.0
0.6
-
-
22 ч
1.0
6.5
13.5
5.7
15.2
13.2
25.3
8.5
10.7
БИОФИЗИКА том 68
№ 4
2023
БИОЛЮМИНЕСЦЕНЦИЯ ГРЕБНЕВИКА
743
Таблица 5. Длительность биолюминесцентного сигнала, с
Размерная группа
4
6
8
10
12
14
16
18
20
Механическая стимуляция (8 ч)
Минимум, Lинд.min
0.11
0.14
0.37
0.85
0.65
0.68
0.54
-
-
Максимум, Lинд.max
4.99
2.43
5.99
6.99
4.94
5.99
1.33
-
-
Среднее, Lср
2.24
2.32
5.05
4.80
4.63
5.47
5.96
-
-
Механическая стимуляция (22 ч)
Минимум, Lинд.min
0.53
0.36
0.26
0.23
0.18
0.68
0.27
0.44
0.21
Максимум, Lинд.max
0.53
4.99
4.99
4.99
4.99
4.99
4.99
2.43
0.60
Среднее, Lср
0.53
1.22
1.34
1.01
1.88
2.03
3.43
1.14
0.41
Химическая стимуляция (8 ч)
Минимум, Lинд.min
0.03
0.02
0.03
0.04
0.02
0.13
0.01
-
-
Максимум, Lинд.max
0.21
0.38
0.30
0.41
0.43
0.19
0.31
-
-
Среднее, Lср
0.12
0.15
0.13
0.14
0.24
0.17
0.12
-
-
Химическая стимуляция (22 ч)
Минимум, Lинд.min
0.14
0.04
0.26
0.15
0.35
0.42
0.23
0.23
0.32
Максимум, Lинд.max
0.14
0.36
5.99
9.35
4.96
9.98
6.15
5.51
0.54
Среднее, Lср
0.14
0.20
1.85
3.28
1.58
3.13
1.68
2.76
0.43
Критерий достоверности различия
8 ч
0.89
2.30
0.19
1.28
0.39
1.01
2.05
-
-
22 ч
1.00
0.90
0.74
2.54
0.37
1.00
1.33
0.97
0.12
плитуда была минимальной по сравнению с дру-
сигнала составляла 1.6 с, при этом максимальные
гими размерными группами.
величины наблюдали в группе с размерами 16-
17 мм, средняя длительность в которой составила
При химическом типе стимуляции утром и но-
3.4 с, максимальная индивидуальная - 5 с. При
чью средняя по группам амплитуда составляла
химическом типе стимуляции средняя по груп-
1.5∙10-3 мкВт∙см-2, максимальные величины при
пам длительность сигнала составляла 2.14 с. Мак-
этом наблюдались в размерной группе 12-13 мм.
симальная длительность биолюминесцентного
Таким образом, при химической стимуляции
сигнала отдельных индивидуумов Lинд.max ночью
ночью зарегистрированы более высокие величи-
достигала 5 и 10 с при механической и химиче-
ны индивидуальной амплитуды, чем в утреннее
ской стимуляции, соответственно; в среднем по
время, однако на средних величинах это не отрази-
группам максимумы зарегистрированы для групп
лось, и они были равны утренним, либо ниже их.
с размерами 14-17 мм.
Средняя длительность сигнала по всем груп-
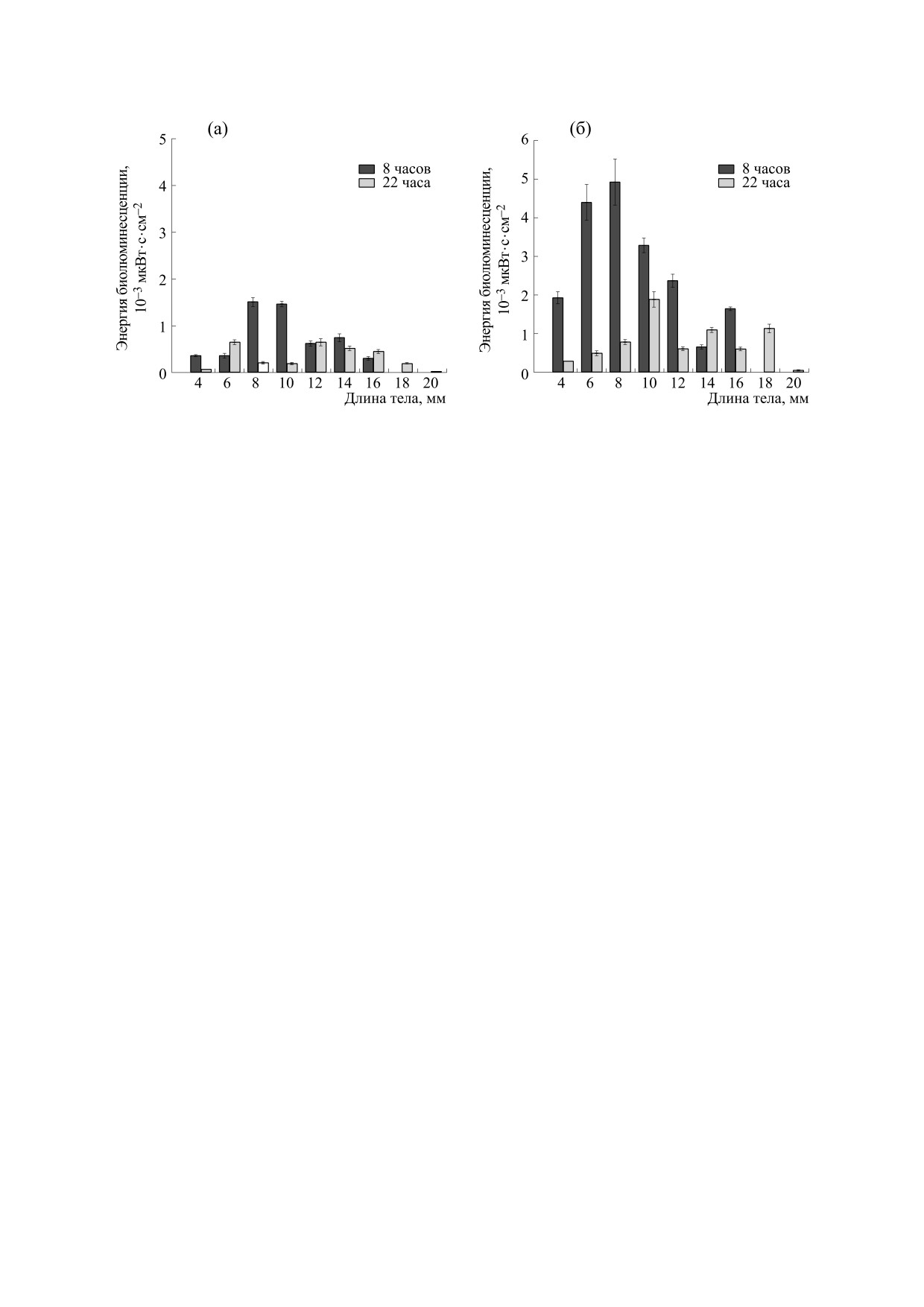
Средняя энергия сигнала в утреннее время
пам утром при механическом типе стимуляции
составляла. 3.1 с (максимум отмечен в группе с
при механической стимуляции составляла
размерами 8-9 мм - 4.5 с). При химическом типе
1.0∙10-3 мкВт∙с∙см-2, при этом максимальные ве-
стимуляции средняя длительность составляла
личины отмечены в группах с размерами 8-11 мм
3.3 с, максимальная среди особей длительность
(рис. 5а). Величины энергии сигнала при химиче-
сигнала также зарегистрирована в группе с разме-
ской стимуляции были значительно выше, чем
рами 8-9 мм (Lинд.max = 11.9 с), а максимальная
при механическом типе стимуляции. Средняя по
средняя по группе длительность отмечена для
группам энергия при химической стимуляции
размеров 16-17 мм (табл. 5).
утром составила 3.1∙10-3 мкВт∙с∙см-2, а макси-
мальная индивидуальная энергия сигнала, заре-
Почти для всех групп утром длительность сиг-
гистрированная в группе с размерами 8-9 мм, -
нала значительно (в два-четыре раза) превышала
таковую в ночное время как при механической
22.6∙10-3 мкВт∙с∙см-2 (средняя по этой размерной
стимуляции, так и при химической (табл. 5).
группе составила 4.9∙10-3 мкВт∙с∙см-2 и также
В ночное время при механическом типе стиму-
была максимальной среди средних других раз-
ляции средняя по всем группам длительность мерных групп (рис. 5б)).
БИОФИЗИКА том 68
№ 4
2023
744
ТЕМНЫХ и др.
Рис. 5. Средняя энергия биолюминесцентного сигнала утром (8:00) и ночью (22:00) при различных видах стимуляции:
(а) - механической, (б) - химической.
В ночное время энергия сигнала была ниже,
нической стимуляции регистрировали с 04:00 до
средняя величина при механической стимуляции
16:00, длительность сигнала отдельных особей в
эти часы достигала 5-7 с. Средняя длительность
составила 0.4∙10-3 мкВт∙c∙см-2, а максимальная
сигнала в течение суток составляла 2 с.
отмечена в группе 12-13 мм (средняя энергия сиг-
нала составила 0.7∙10-3 мкВт∙c∙см-2, индивиду-
При химическом типе стимуляции в течение
суток средние величины амплитуды в основном
альная достигала 3.7∙10-3 мкВт∙c∙см-2).
были выше, чем при механическом типе, наи-
При химической стимуляции средняя энергия
большие величины зафиксированы в
06:00,
сигнала Eср ночью также была ниже, чем в 8 часов
10:00 и в полночь
(3.0-3.8)∙10-3 мкВт∙см-2),
утра, и в итоге средняя по группам величина со-
амплитуда отдельных экземпляров достигала
ставила 1.0∙10-3 мкВт∙c∙см-2.
6.6∙10-3 мкВт∙см-2 (в 24:00).
Длительность и энергия сигнала ночью в боль-
Наибольшие величины длительности сигнала
шинстве размерных групп были ниже утренних
при химической стимуляции отмечены в 8:00 и
величин, эта закономерность нарушалась иногда
22:00 (у отдельных индивидуумов достигая 8.0-
лишь для групп с размерами 14-17 мм.
9.4 с), а среднее время составило 2.26 с. Как и при
Суточная динамика параметров биолюминес-
механической стимуляции, суточная динамика
ценции у группы размером 10 мм. Детально суточ-
длительности сигнала в основном находилась в
ную динамику параметров биолюминесценции
противофазе с изменениями амплитуды. С 18:00
исследовали у экземпляров размерами 10 мм, экс-
до 02:00 длительность превышала таковую при
перименты по высвечиванию проводили каждые
механическом типе стимуляции.
два часа в течение 24 ч. Было выявлено, что
Средняя энергия биолюминесцентного сигна-
при механическом типе стимуляции амплитуда
ла в течение суток при механической стимуляции
биолюминесцентного сигнала достигала наи-
больших значений в период с 18:00 до 6:00, в это
составляла 0.7∙10-3 мкВт∙с∙см-2, при химической -
время ее средние величины достигали Aср = 0.9-
1.7∙10-3 мкВт∙с∙см-2. Максимальная индивиду-
альная величина энергии при механической
2.9∙10-3 мкВт∙см-2, а индивидуальные
-
стимуляции составляла 6.0∙10-3 мкВт∙с∙см-2 и
Aинд.max =
4.3∙10-3 мкВт∙см-2. Максимум Aср
зарегистрирована в 18:00, при химической -
зарегистрирован в 2:00 (рис. 5) и составлял
14.0∙10-3 мкВт∙c∙см-2 (в 22:00).
3.0∙10-3 мкВт∙см-2.
Суточная динамика длительности сигнала в
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
основном находилась в противофазе с изменени-
ями амплитуды, так, высоким величинам ампли-
Несмотря на утверждения об отсутствии био-
туды соответствовали низкие величины длитель-
люминесценции у P. pileus [29], для Черного моря
ности, и наоборот, при минимальных амплитудах
ранее было показано, что особи размером 7-9 мм
отмечались максимальные величины длительно-
являются биолюминесцентными, по крайней ме-
сти сигнала (рис. 6). Высокие величины при меха-
ре, в холодный сезон [30].
БИОФИЗИКА том 68
№ 4
2023
БИОЛЮМИНЕСЦЕНЦИЯ ГРЕБНЕВИКА
745
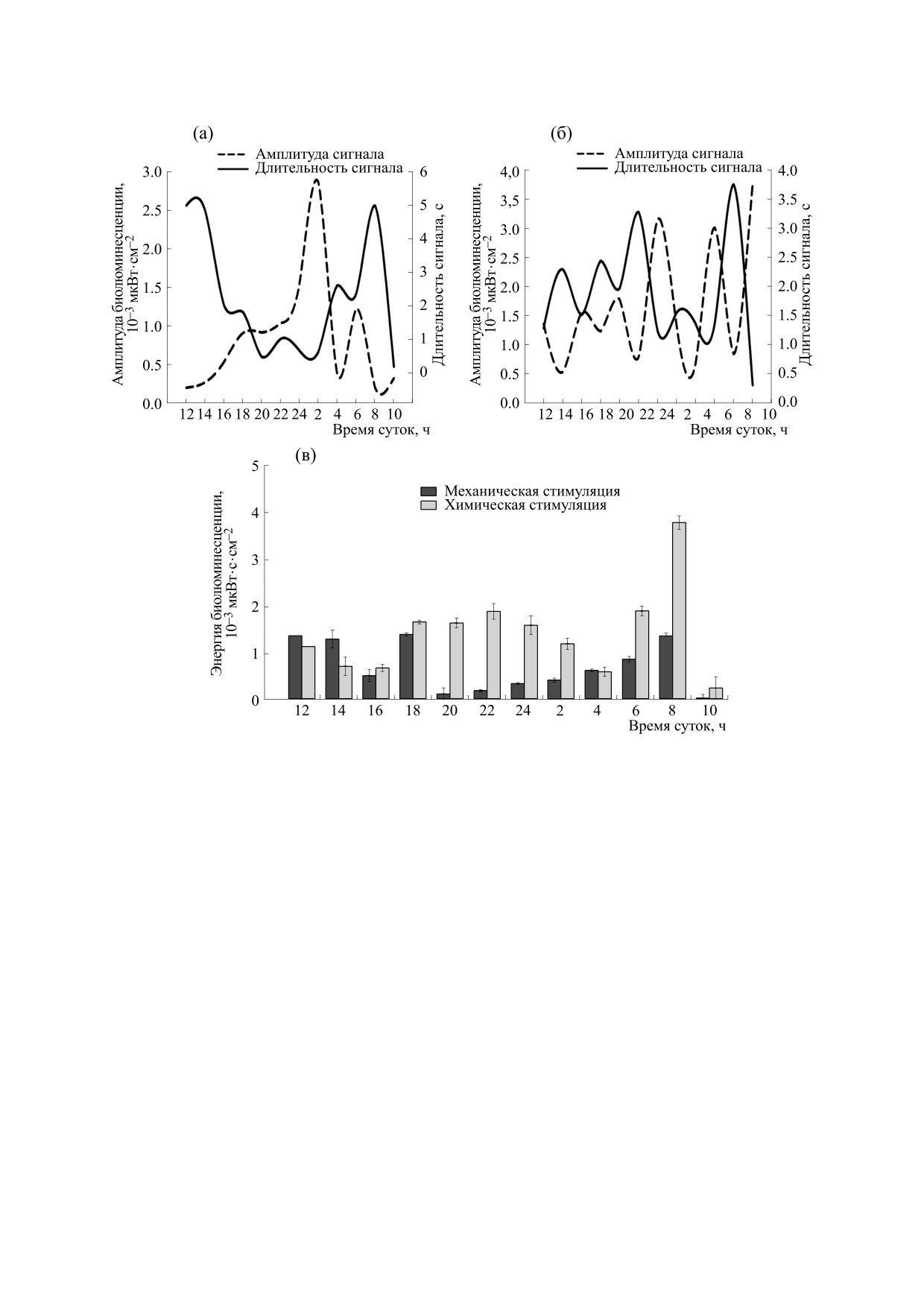
Рис. 6. Суточная динамика средних по исследуемой размерной группе (длина тела 10 мм) величин амплитуды и
длительности при механической (а) и при химической (б) стимуляции; энергии (в) при механическом и химическом
типе стимуляции.
По нашим данным, в летний сезон наблюда-
они походили на первый импульс, хотя интенсив-
лось три типа биолюминесцентного сигнала
ность излучения у них была в несколько раз
P. pileus, первые два регистрировали при механи-
меньше.
ческой стимуляции, третий - при химической
Анализируя параметры биолюминесценции
(см. рис. 3), Форма сигналов отличалась от тако-
P. pileus в дневное и ночное время, можно заме-
вого у других представителей ктенофор - B. ovata
тить, что при механической стимуляции практи-
и M. leidyi. Так, биолюминесцентный сигнал при
чески нет отличий в значениях амплитуды и
механической стимуляции у P. pileus представлял
явной зависимости длительности сигнала от раз-
собой один или несколько объединенных четко
мера особи. При химической стимуляции наблю-
выраженных импульсов (рис. 3а,б), у других ви-
дается слабый рост этих параметров при увеличе-
дов гребневиков импульсы слиты в единый сиг-
нии размера особи. В темное время суток (22:00)
нал, выглядящий как острые зубцы гребенки, с
выявлен незначительный спад значений ампли-
постепенным нарастанием до максимального
туды при механической стимуляции и рост
значения и спадом. При химической стимуляции
длительности сигнала при увеличении размера
у P. pileus последний импульс часто имел, в отли-
особи.
чие от B. ovata, пологий и длительный (1.5-2.5 с)
В результате наших экспериментов выявлено,
фронт спада. У M. leidyi при химической стимуля-
что среди групп с разными размерами и возрас-
ции могли появляться дополнительные импуль-
том особей максимальная амплитуда биолюми-
сы после небольшой паузы (0.1-0.5 с), по форме
несцентного сигнала, как индивидуальная, так и
БИОФИЗИКА том 68
№ 4
2023
746
ТЕМНЫХ и др.
средняя по группе, при механической и химиче-
представлена в Черном море и встречается не
ской стимуляции отмечена в размерной группе
только в глубоководных слоях [31], но и в бухтах и
6-7 мм. Что касается длительности сигнала, мак-
на небольших глубинах [32], тем самым доступ
симальные величины отмечены у самых малень-
к особям данного вида не затруднен, что позволя-
ких (4-7 мм) и самых крупных (16-21 мм) особей.
ет его использовать в экспериментах. Размеры
Это позволяет выбрать эти размерные группы для
P. pileus позволяют исследовать влияние различ-
проведения экспериментальных исследований
ных факторов на биолюминсценцию всех размер-
по влиянию токсических веществ.
ных групп этого вида, в то время как проведение
исследований с особями B. ovata и M. leidyi более
В суточной динамике амплитуды замечена
60 мм затруднено инструментально.
следующая закономерность: при механической
стимуляции в темное время суток (с 18:00 до 6:00)
и при химической стимуляции с 20:00 до 10:00 от-
ВЫВОДЫ
мечаются повышенные величины, максимумы
Было выявлено, что биолюминесцентную ре-
регистрировали с 22:00 до 02:00 при механиче-
акцию P. pileus в летнее время на механическую и
ской и в полночь при химической стимуляции.
химическую стимуляцию проявляют все размер-
Таким образом, оптимальным временем (при по-
ные группы данного вида. В летний сезон при хи-
стоянной температуре 21°С на всех этапах экспе-
мическом типе стимуляции в среднем у всех раз-
римента) в летний сезон для проведения экспери-
мерных групп амплитуда была выше примерно в
ментальных работ по воздействию на физиологи-
два раза, чем при механическом. Наибольшие
ческое состояние вида с использованием анализа
значения амплитуды биолюминесцентного сиг-
параметров биолюминесцентного отклика явля-
нала выявлены у особей размера 6-7 мм. Наи-
ется темное время суток с 20:00 до 6:00.
большая длительность зарегистрирована у особей
Сравнивая значения основных параметров
P. pileus размером 4-7 и 16-21 мм. Величины
биолюминесцентного сигнала P. pileus с B. ovata
амплитуды биолюминесцентного сигнала в
[14, 20] и M. leidyi [14], можно заметить сходство в
темное время суток выше, чем в утреннее время.
суточной динамике амплитуды биолюминес-
На основании полученных результатов можно
центного сигнала. Так, у B. ovata минимум прихо-
заключить, что биолюминесценцию вида
дится на 10:00, а максимум -на 01:00, у M. leidyi
P. pileus можно использовать в исследованиях
минимум выпадает на 7:00-10:00, а максимум на
влияния различных факторов на физиологию ор-
13:00 и 01:00, что соответствует и минимуму, и
ганизма. Оптимальным временем проведения
максимуму в значениях амплитуд у P. pileus.
экспериментов в летнее время является период с
На данный момент для Черного моря список
20:00 до 6:00. Уровень биолюминесцентного сиг-
биолюминесцентов насчитывает 8 видов бакте-
нала отдельной особи P. pileus ниже, чем у гребне-
риопланктона родов Vibrio и Photobacterium, 36 ви-
виков-вселенцев Черного моря B. ovata и M. leidyi,
дов фитопланктона (родов Ceratium, Gonyaulax,
на один-два порядка и сравнима с некоторыми
Lingolodinium, Noctiluca, Protoperidinium и Pyrocys-
представителями динофлагеллят, такими, как
tis) и 9 видов зоопланктона [4]. Биолюминесцен-
Noctiluca scintillans и некоторыми видами рода
ция большинства видов зоопланктона для Черно-
Protoperidinium, но при этом значительно выше,
го моря подтверждена визуально, инструменталь-
чем у остальных светящихся видов фитопланкто-
но получены результаты только для гребневиков.
на Черного моря.
Сравнивая параметры биолюминесценции оди-
ночного экземпляра P. pileus и исследованных ра-
нее биолюминесцентов [4], можно увидеть, что
ФИНАНСИРОВАНИЕ РАБОТЫ
амплитуда биолюминесцентного сигнала этого
Работа выполнена в рамках темы Государ-
вида - (0.8-4.3)∙10-3 мкВт∙см-2 - близка к ам-
ственного задания № 121041400077-1 «Функцио-
плитуде Noctiluca scintillans (Macartney) Kofoid &
нальные, метаболические и токсикологические
аспекты существования гидробионтов и их по-
Swezy, 1921 (до 4∙10-3 мкВт∙см-2) и некоторым ви-
пуляций в биотопах с различным физико-хими-
дам рода Protoperidinium ((1.1-2.5)∙10-3 мкВт∙см-2),
ческим режимом». Проведенные исследования
но превосходит на один-два порядка
были выполнены в Центре коллективного поль-
остальные виды фитопланктона, на семь по-
зования «НИС Профессор Водяницкий» Феде-
рядков выше значения амплитуды бактерий
рального исследовательского центра «Институт
(4.95∙10-10 мкВт∙см-2 у Vibrio indicus) и на два-три
биологии южных морей имени А.О. Ковалевско-
порядка меньше, чем у других гребневиков (до
го РАН».
1.0 мкВт∙ см-2 у M. leidyi).
Виды B. ovata и M. leidyi активно используются
КОНФЛИКТ ИНТЕРЕСОВ
в экспериментальных биофизических исследова-
ниях по выявлению влияния токсических ве-
Авторы заявляют об отсутствии конфликта
ществ на организм [26-28]. P. pileus также широко
интересов.
БИОФИЗИКА том 68
№ 4
2023
БИОЛЮМИНЕСЦЕНЦИЯ ГРЕБНЕВИКА
747
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
15. P. B. Tett, J. Mar. Boil. Ass. UK, 49 (1), 245 (1969).
16. Ю. Н. Токарев, Дис. … канд. биол. наук (ИнБЮМ,
Настоящая работа не содержит описания ис-
Севастополь, 1990).
следований с использованием людей и животных
17. Э. П. Битюков, Зоол. журн., 47(1), 36 (1968).
в качестве объектов.
18. И. И. Гительзон, Л. А. Левин, Р. Н. Утюшев и др.,
Биолюминесценция океана (Гидрометеоиздат,
СПИСОК ЛИТЕРАТУРЫ
СПб., 1992).
1.
А. И. Журавлев, В. А. Веселовский и Н. Н. Коще-
19. П. В. Евстигнеев и Э. П. Битюков, Экология моря,
енко, Труды МОИП, 21, 19 (1965).
24, 87 (1986).
2.
S. H. D. Haddock, M. A. Moline, and J. F. Case, Annu.
20. О. В. Машукова, Экология моря, 79, 15 (2009).
Rev. Mar. Sci., 2 (1), 443 (2010).
21. О. В. Машукова, в сб. Экологические проблемы Азо-
3.
J. G. Morin, Bull. Marine Sci., 33 (4), 787 (1983).
во-Черноморского региона и комплексное управление
4.
Ю. Н. Токарев, Основы биофизической экологии
прибрежной зоной: Материалы молодежной научно-
гидробионтов (ЭКОСИ-Гидрофизика, Севасто-
практической конференции (ЭКОСИ-Гидрофизи-
поль, 2006).
ка, Севастополь, 2014), сс. 89-94.
5.
O. Shimomura, Bioluminescence: chemical principles
22. П.В. Евстигнеев, Экология моря 42, 31 (1992).
and method (World Scientific, 2012).
23. E. N. Harvey, Bioluminescence (Acad. Press, N.-Y.,
6.
S. H. D. Haddock and J. F. Case, Marine Biol., 133,
1952).
571 (1999).
24. Ю. Н. Токарев, О. В. Машукова и В. И. Василенко,
7.
Э. П. Битюков, В. И. Василенко, И. М. Серикова
Экология моря, 76, 61 (2008).
и др., Экология моря, 46, 19 (1996).
25. Yu. N. Tokarev and O. V. Mashukova, Luminescence,
8.
W. E. Esaias and H. C. Curl, Limnol. Oceanogr., 17
14, 351 (2016).
(6), 901 (1972).
26. O. Mashukova, Y. Tokarev, and E. Skuratovskaya,
9.
С. А. Пионтковский и С. А. Серегин, Поведение ко-
Ecol. Montenegrina, 14, 109 (2017).
пепод (ЭКОСИ-Гидрофизика, Севастополь, 2006).
27. О. В. Машукова и М. И. Силаков, Труды Карадаг-
10.
В. Б. Цейтлин и Ю. А. Рудяков, Докл. АН СССР, 38
ской научной станции им. Т. И. Вяземского - при-
(4), 978 (1989).
родного заповедника РАН, 4 (16), 58 (2020).
11.
E. Pennisi, Science, 235 (3), 1060 (2012).
28. О. В. Машукова, М. И. Силаков, Л. В. Малахова
и др., Системы контроля окружающей среды, 3
12.
П. В. Евстигнеев, в сб. Биологические ресурсы водо-
(45), 39 (2021).
емов в условиях антропогенного воздействия (Наук.
думка, Киев, 1985), сс. 29-31.
29. S. H. D. Haddock and J. F. Case, Biol. Bull., 189 (3),
356 (1995).
13.
Э. П. Битюков, П. В. Евстигнеев и Ю. Н. Токарев,
Гидробиол. журн., 29 (4), 27 (1993).
30. О. В. Машукова, Вестн. Прикаспия, 3 (22), 15
(2018).
14.
Ю. Н. Токарев, П. В. Евстигнеев и О. В. Машуко-
ва, Планктонные биолюминесценты Мирового океа-
31. E. Mutlu and F. Bingel, Marine Biol., 135, 589 (1999).
на: видовое разнообразие, характеристики светоиз-
32. М. Е. Виноградов, М. В. Флинт и Э. А. Шушкина,
лучения в норме и при антропогенном воздействии
в сб. Современное состояние экосистемы Черного
(Нижняя Ореанда, Симферополь, 2016).
моря (Наука, М., 1987), сс. 172-186.
Bioluminescence of Ctenophore Pleurobrachia pileus (O. F. Müller, 1776)
in the Summer Period
A.V. Temnykh*, M.I. Silakov*, and O.V. Mashukova*
*A.O. Kovalevsky Institute of Biology of the Southern Seas, Russian Academy of Sciences,
prosp. Nakhimova 2, Sevastopol, 299001 Russia
The capability of bioluminescence has been found in many marine hydrobionts, including almost all repre-
sentatives of the phylum Ctenophora. The glow of species and bioluminescent substrates isolated from them
is actively used in biology, ecology, and medicine to study the physiological state of an organism or to identify
unfavorable environmental parameters. The parameters of luminescence of the Black Sea ctenophores Mne-
miopsis leidyi A. Agassiz, 1865 (Lobata) and Berce ovata Bruguière, 1789 (Beroida) are used to study the effects
of toxicants, such as liquid hydrocarbons, PCBs, and heavy metals. Some researchers casted doubt on the ca-
pability of the Black Sea autochthonous inhabitant, ctenophore Pleurobrachia pileus (O. F. Müller, 1776) to
produce bioluminescent light. The objective of this work was to ensure scientific support for the light-pro-
ducing ability of species under study and to choose the optimal time for conducting experiments on the fac-
tors that affect bioluminescence. It was found that mechanical and chemical stimulation triggered a biolumi-
nescent response in all size groups of species under study at any time of day. Upon chemical stimulation of
the studied species, the signal energy value was several times greater than that observed after mechanical stim-
ulation. It was revealed that the optimal time interval for conducting biophysical research is the time from
8 pm to 6 am, because the values of luminescence activity are the greatest in this time interval within a day.
Keywords: Ctenophora, luminescence, circadian rhythm, the Black Sea
БИОФИЗИКА том 68
№ 4
2023