БИОФИЗИКА, 2023, том 68, № 6, с. 1200-1209
БИОФИЗИКА CЛОЖНЫX CИCТЕМ
УДК 579.24; 631.48
ЛИПАЗНАЯ И ЭСТЕРАЗНАЯ АКТИВНОСТЬ В ГРУНТАХ ИЗ ДРЕВНИХ
ПОГРЕБАЛЬНЫХ СОСУДОВ
© 2023 г. Н.Н. Каширская*, Е.В. Чернышева*, С.Н. Удальцов*, Л.Н. Плеханова*, #
*Институт физико-химических и биологических проблем почвоведения - обособленное подразделение ФИЦ
«Пущинский научный центр биологических исследований Российской академии наук»,
Институтская ул., 2, Пущино Московской области, 142290, Россия
#E-mail: dianthus1@rambler.ru
Поступила в редакцию 18.10.2023 г.
После доработки 07.11.2023 г.
Принята к публикации 15.11.2023 г.
Проведена оценка активности липолитических ферментов микропланшетным методом с
использованием гетеромолекулярного обмена в грунтах со дна погребальной посуды, с
последующим кластерным анализом. Выявлены группы с контрастным соотношением липаз С12 -
С18 и эстераз С4 - С8 в выборке, включающей 57 погребальных горшков возраста двух-трех тысяч
лет. Суммарная липолитическая активность образцов грунта из посуды с предположительным
исходным присутствием жиросодержащих пищевых заупокойных продуктов составляла выше 6%
от максимального значения в пределах выборки. Ингибирование эстераз в присутствии высоких
концентраций жирных кислот отражено в предельно низких относительных значениях эстеразной
активности почвенного микробного сообщества на фоне максимальных показателей активности
липаз С18 и С12 в двух группах горшков. В трех группах наблюдалась эстеразная активность на фоне
высокой относительной активности липаз С16, средней относительной активности липаз С18 и
предельно низкой активности истинных липаз. В группе горшков с высокой суммарной
липолитической активностью (за исключением отдельных экстремальных величин этого
показателя) доля активности истинных липаз была в 1.7-1.8 раза выше по сравнению с долей
эстеразной активности. Группировка погребальной посуды по соотношению липазной и
эстеразной активности отражает характеристики заупокойной пищи, тогда как абсолютные
значения активности ферментов среди избранных сосудов с наибольшей суммарной
липолитической активностью группируются в соответствии с типом погребениея.
Ключевые слова: липолитические ферменты, липаза, эстераза, жирные кислоты, погребальные сосуды,
посуда, горшки, заупокойная пища, древняя пища, эпохa бронзы, ранний железный век.
DOI: 10.31857/S000630292306011X, EDN: ROSYVA
Разнообразие липидов, обнаруживаемых в ар-
ния происхождения жиров [5]. Липиды в керами-
хеологических материалах, и сопоставимость их с
ке, как правило, сохраняются в порах размером от
современными вариантами относит эту группу
нано- до микрометра, однако они обнаруживают-
соединений к надежным биомаркерам в опреде-
ся и в зонах размером >200 мкм [6], их сохране-
лении состава пищевых продуктов древности [1].
нию способствует образование солей жирных
Из всех липидных индикаторов культурного
кислот кальция.
слоя, используемых для получения исторической
В модельных экспериментах старения расти-
информации о деятельности человека, наиболее
тельных и животных жиров исследованы процес-
широко применяются биомаркеры копролитов в
сы термического разложения, окисления и гидро-
качестве богатого источника информации о дие-
лиза липидов, которые могли происходить в ар-
тах [2]. В образцах древней керамики молекулы
хеологических условиях [3, 7]. Жирные кислоты,
липидов являются долговечными биомаркерами
содержащие по меньшей мере одну двойную
пищевых продуктов [3]. Хотя интактные триаци-
связь вдоль углеродной цепи, особенно чувстви-
лглицерины в керамических фрагментах сохраня-
тельны к реакциям окисления [3]. Длинноцепо-
ются далеко не всегда [4], однако в некоторых
чечные ω-(о-алкилфенил)алкановые кислоты,
случаях их количество достаточно для проведе-
получаемые при нагревании ненасыщенных жир-
ния высокоэффективной жидкостной хромато-
ных кислот, используются для идентификации
графии, являющейся критерием для установле-
водных организмов, употребляемых в пищу в
1200
ЛИПАЗНАЯ И ЭСТЕРАЗНАЯ АКТИВНОСТЬ В ГРУНТАХ
1201
древности [8]. В результате исследования керами-
ферментов сохраняться в почве связана с услови-
ческих сосудов нубийского городища Каср-Иб-
ями, когда разложение органического вещества
рим с помощью методов газовой хроматографии
почвы ограничено [15]. Защиту органического ве-
и масс-спектрометрии, были обнаружены насы-
щества от микробного разложения обеспечивает
щенные карбоновые кислоты в диапазоне от C12
окклюзия - покpытие повеpxноcти оpганичеcкиx
до C18, с необычно высоким содержанием С12,
фpагментов минеpальными чаcтицами фpакций
что совпало с распределением насыщенных жир-
пыли и глины [16]. Вpемя cущеcтвования в почве
ных кислот, обнаруженных в ядрах финиковой
окклюдиpованного оpганичеcкого вещеcтва уве-
пальмы [9]. В оливковом масле возрастом две ты-
личивается на поpядок [17]. Кроме того, фермен-
сячи лет, обнаруженном в стеклянной бутылке на
ты, сохраняющие активноcть в почве на пpотяже-
археологических раскопках Везувия, триацилг-
нии тыcячелетий, должны иметь выcокую кон-
лицерины были полностью гидролизованы, а по-
фоpмационную
cтабильноcть
тpетичной
лученные в результате гидролиза жирные и гид-
cтpуктуpы белка, способность сохранять актив-
рокси-жирные кислоты конденсировались в ред-
ность при деформации значительных участков
ко встречающиеся эстолиды [10].
биомолекулы [18].
Изотопный состав липидных компонентов ке-
рамики может дать исчерпывающую информа-
Липолитические ферменты хорошо сохраня-
ются в культурных слоях археологических памят-
цию о происхождении жиров. Исследования экс-
ников [19] и сосудах из древних погребений [20].
трагируемых растворителем липидных компо-
нентов и определение стабильных изотопов
Они катализируют реакции гидролиза триглице-
углерода основных n-алкановых кислот в абсор-
ридов до ди- и моноглицеридов, глицерина и
бированных и карбонизированных остатках жира
жирных кислот [21], а также обратную реакцию -
внутри керамики позволили провести различие
этерификацию глицерина жирными кислотами в
между жирами нежвачного (свиного) и жвачного
системах с низкой активностью воды [22]. Пред-
(овечьего или бычьего) происхождения [11]. Ана-
ставители наиболее изученного семейства истин-
лиз изотопного состава липидов из керамических
ных липаз (по классификации, основанной на
черепков, найденных на побережье Внешних
сходстве аминокислотных последовательностей
Гебридских островов (Великобритания), выявил
[23]) преобразуют длинноцепочечные жирнокис-
смешение жиров жвачных животных и морепро-
лотные субстраты с десятью и более атомами уг-
дуктов в 71% сосудов [12]. Наличие изопреноид-
лерода. В целом, к липазам относятся все липоли-
ных жирных кислот в липидных остатках, а также
тические ферменты, которые способны высво-
характерное значение стабильных изотопов в на-
бождать свободные жирные кислоты из
сыщенных жирных кислотах С16:0 и С18:0 позво-
длинноцепочечных триацилглицеринов, содер-
лило установить назначение керамических кув-
жащихся в растительных и животных жирах [24].
шинов доколониальных общин хардингов в при-
Для некоторых липаз отмечается увеличение
брежных районах Южной Африки [4]. Результаты
сродства к субстрату при возрастании длины
анализа липидов показали здесь отсутствие мо-
жирнокислотной последовательности, а также
лочного жира и наличие жира морских животных.
при наличии двойных связей в количестве не бо-
В отличие от наземных животных жиров и расти-
лее двух [25]. Большее число двойных связей сни-
тельных масел, масла, полученные из морских
жает активность, создавая пространственные за-
животных, содержат значительно большее коли-
труднения при образовании фермент-субстрат-
чество ненасыщенных жирных кислот, особенно
ного комплекса.
с углеродной длиной C18-C22 [4].
Если максимальная активность фермента про-
Таким образом, остатки липидов в керамиче-
является в отношении жирных кислот с короткой
ских фрагментах способны дать информацию о
или средней цепью, их относят к эстеразам [26]. В
бытовом использовании древней погребальной
посуды. Помещение ритуальной пищи в горшок
отличие от истинных липаз, эстеразы могут рас-
происходит однократно и не может оказать зна-
щеплять длинные одноцепочечные молекулы
жирных кислот, связанных с нитрофенилом, но
чительного влияния на состав липидов внутри ке-
рамики. В погребении жирная пища смешивается
не способны воздействовать на жиры с тремя уг-
с почвой, стимулируя развитие почвенного липо-
леродными цепями такой же длины. Некоторые
литического микробного комплекса. Микробные
микробные липолитические ферменты способны
липазы имеют большое значение в археологиче-
к расщеплению широкого спектра субстратов,
ских исследованиях. В культурных слоях и погре-
хотя в целом уменьшают свою активность при
бениях липазы, как и другие ферменты [13, 14],
увеличении длины жирнокислотной цепи, одна-
выступают в качестве объекта исследования, яв-
ко проявляют более высокую [27] или даже мак-
ляясь носителями биологической памяти почв об
симальную [28] активность по отношению к жир-
изменениях условий почвообразования и антро-
нокислотным последовательностям средней дли-
погенной деятельности в прошлом. Способность
ны.
БИОФИЗИКА том 68
№ 6
2023
1202
КАШИРСКАЯ и др.
Оценка липазной активности и определение
антимикробная эффективность свободных жир-
численности микроорганизмов, способных выра-
ных кислот - лауриновой (С12:0) и каприловой
батывать липазы, успешно используются в архео-
(С8:0), что в пищевой промышленности исполь-
логической микробиологии [20, 29, 30]. Микроб-
зуется для борьбы с портящей микрофлорой, осо-
ные сообщества почвы вырабатывают липазы в
бенно с листериями или другими грамположи-
ответ на поступление липидов антропогенного
тельными патогенами [38]. Лауриновая кислота
происхождения. Так, при исследовании археоло-
является наиболее мощной бактерицидной жир-
гического памятника Сотк-2 (Республика Арме-
ной кислотой против грамположительных энте-
ния) было выявлено существенное увеличение
рококков, а каприловая кислота наиболее эффек-
активности липаз в слое среднего-позднего брон-
тивна против грамотрицательных микроорганиз-
зового века на границе хозяйственной ямы, что в
мов, таких, как кишечная палочка
[39].
совокупности с другими почвенными характери-
Антимикробные препараты на основе пептидов и
стиками подтверждает вероятность использова-
жирных кислот длиной от 8 до 12 атомов углерода
ния данного участка в качестве места приготовле-
проявляли оптимальную активность в отноше-
ния пищи [19].
нии патогенных штаммов, резистентных к анти-
биотикам [40]. Моноэфиры сахарозы и жирных
Работа микробных липаз является неотъемле-
кислот проявляли ингибирующий эффект на раз-
мой частью процессов ферментации жиросодер-
витие спор Bacillus cereus, предотвращая их пере-
жащих пищевых субстратов, а также процессов их
ход в размножающиеся клетки, однако по мере
порчи и окончательного разложения. В грунте за-
снижения концентрации сложных эфиров в сре-
полнения ритуальных сосудов из древних погре-
де, клетки возобновляли рост. Потеря антимик-
бений наблюдается высокая липазная актив-
робной активности сложных эфиров, наблюдае-
ность, обусловленная исходным наличием пита-
мая во время культивирования Bacillus cereus, бы-
тельного субстрата жировой природы [20, 29].
ла обусловлена их разложением эстеразой [41].
В сосудах с исходным содержанием жирной
На уровне микробных сообществ, выявлены
пищи за образование липаз ответственно как
периоды ингибирования преобладающих мик-
микробное сообщество почвы, попадающей в со-
робных групп в анаэробных средах с высоким со-
суд, так и микробное сообщество самого пищево-
держанием липидов. В процессе анаэробного
го продукта. В условиях погребения микробные
сбраживания содержащих липиды отходов при
сообщества сохраняются в течение неопределен-
производстве метана, липиды первоначально
но долгого времени, за счет перехода клеток в по-
гидролизуются до глицерина и длинноцепочеч-
коящееся состояние [31]. При этом значительную
ных жирных кислот, которые на следующем эта-
долю в погребенных почвах составляют устойчи-
пе преобразуются синтрофными ацетогенными
вые группы микроорганизмов, связанные с дея-
бактериями в водород и ацетат и далее - метано-
тельностью человека [32-34]. Но прежде, чем пе-
генными археями, - в метан. Скорость работы
рейти в состояние покоя, микроорганизмы в со-
микробного сообщества в значительной мере
судах,
содержащих
пищу,
должны
ограничивается накоплением длинноцепочечных
воспользоваться избытком органического веще-
жирных кислот, разложение которых происходит
ства и выделить ферменты для его утилизации. В
по пути β-окисления [42]. Известно, что высокий
древних погребальных сосудах отмечено наличие
уровень содержания таких кислот ингибирует де-
белкового продукта, на что указывает анализ со-
ятельность синтрофических ацетогенов и мета-
держания фосфатов [35].
ногенов, что поначалу объяснялось токсично-
Однако высокая калорийность пищи и ее зна-
стью этих соединений и повреждением клеток
чительная масса, сопоставимая с массой почвы,
микроорганизмов [43]. Позднее была выявлена
попавшей в сосуд, может оказывать ингибирую-
адаптация микробных сообществ к ингибирую-
щее действие на почвенные микроорганизмы и
щим концентрациям длинноцепочечных жирных
микробные сообщества, а также ингибировать
кислот [44]. В настоящее время не ясно, является
выработку липаз и эстераз. Было установлено,
ли этот процесс адаптации результатом сдвига
что эстеразы, связанные с клетками Bacillus, ин-
микробной популяции в сторону обогащения
гибировались жирными кислотами [36]. Выра-
специфическими и лучше адаптированными раз-
ботка и активность липазы Pseudomonas fragi
лагателями жирных кислот (популяционная
уменьшались при добавлении в среду свиного са-
адаптация) или фенотипической адаптации су-
ла, олеата натрия и солей других ненасыщенных
ществующих микроорганизмов к их высоким
жирных кислот, без влияния на рост клеток [37].
концентрациям (физиологическая акклиматиза-
Показана антибактериальная активность слож-
ция). Однако очевидно, что обратимость ингиби-
ных эфиров монозамещенных углеводных жир-
рования микробного сообщества высокими кон-
ных кислот в отношении ряда бактерий, вызыва-
центрациями длинноцепочечных жирных кислот
ющих порчу пищевых продуктов, в первую оче-
связана с постепенным истощением ингибитора
редь
Listeria
monocytogenes.
Известна
ацидогенными микроорганизмами, использую-
БИОФИЗИКА том 68
№ 6
2023
ЛИПАЗНАЯ И ЭСТЕРАЗНАЯ АКТИВНОСТЬ В ГРУНТАХ
1203
щими ингибитор в качестве источника питания.
рост микробного сообщества и ингибировать ак-
Вероятно, механизм ингибирования заключается
тивность микробных липолитических фермен-
в адсорбции этих кислот на поверхности клеток,
тов. Очевидно, что в таких условиях жиры на пер-
что ограничивает транспортную способность их
вых стадиях утилизируются медленно, а по мере
мембран [45].
их исчерпания происходит активизация микроб-
ного сообщества. К моменту перехода микроор-
Популяционные профили генов 16S рРНК эу-
ганизмов в состояние покоя, в грунте на дне сосу-
бактерий и архей показали, что при воздействии
да должно сформироваться определенное соот-
длинноцепочечных жирных кислот на микроб-
ношение эстеразной активности, присущей
ные сообщества содержащих липиды отходов,
почвенному микробному сообществу в есте-
преимущество получали β-окисляющие/протео-
ственных условиях, и липазной активности, про-
литические бактерии Syntrophomonas, Clostridium,
являемой микроорганизмами в целях утилизации
Coprothermobacter и Anaerobaculum, а также род
жиров. Особенности строения, общие для боль-
синтрофных архей - Methanosarcina [46]. Анализ
шинства липаз и эстераз - погруженность актив-
микробного сообщества из анаэробных мембран-
ного центра в глубину молекулы, значительные
ных биореакторов совместной переработки сма-
расстояния между элементами каталитической
зочных материалов и пищевых отходов выявил
триады - серином, гистидином и аспартатом
преобладание Lactobacillus (относительная актив-
(или глутаматом) [51], могут в значительной мере
ность 92.2-95.7%) и Anaerolineaceae (относитель-
обеспечивать их конформационную стабиль-
ная активность 13.3-57.5%), которые известны
ность в почве.
как ферментеры и разрушители жирных кислот
[47].
В наших исследованиях варианты грунта на
дне сосудов, обладающие высокой активностью
Увеличение концентрации аммиака и летучих
длинноцепочечных липаз в липолитическом
жирных кислот при высоких нагрузках биореак-
комплексе, демонстрируют изменчивость актив-
торов содержащими липиды отходами приводило
ности эстераз С4 и С8 - от ничтожно малых,
к смещению метанообразования от более эффек-
близких к нулю значений, до предельно высоких,
тивного ацетокластического, с использованием
превышающих активность длинноцепочечных
ацетата, к менее эффективному гидрогенотроф-
липаз. Мы полагаем, что контрастные случаи со-
ному, с использованием водорода. В данном слу-
отношения активности липаз и эстераз не явля-
чае, преимущество получали археи рода Methano-
ются случайностью и отражают особенности раз-
culleus [48]. Если же содержимое биореактора бы-
вития липолитического комплекса в грунте за-
ло богато не только липидами, но и белками,
полнения сосудов. При этом баланс липазной и
наблюдалось совместное ингибирование длинно-
эстеразной активности может выступать в каче-
цепочечными жирными кислотами и аммиаком
стве диагностического признака, позволяющего
как β-окисляющих, так и метаногенных бакте-
упростить сортировку объектов, когда их много-
рий, причем на последние в большей мере дей-
численная выборка делится в результате класте-
ствовал аммиак [49].
ризации на небольшое число крупных группиро-
Компостные микробные топливные элемен-
вок с общими свойствами.
ты, используемые для выработки электроэнергии
Целью настоящей работы является оценка ба-
из смеси содержащих липиды пищевых расти-
ланса и соотношений липазной и эстеразной ак-
тельных отходов и почвы, могут рассматриваться
тивности почвы на дне сосудов из древних захо-
как модельный вариант аэробных микробных со-
ронений на основе кластерного анализа.
обществ с высокой липазной активностью. Пре-
образование содержащих липиды отходов путем
компостирования показывает устойчивость к
ОБЪЕКТЫ И МЕТОДЫ ИССЛЕДОВАНИЯ
микробному разложению жирных кислот с длин-
Были исследованы образцы грунта со дна
ной цепью С19:0, С20:0 [50]. Наличие высоких
57 ритуальных горшков (используем упрощение
уровней этих жирных кислот на заключительной
для обозначения разных типов ритуальной посу-
стадии компостирования указывает на их малую
ды - мисок кувшинов, кружек и др.) из погребе-
доступность для аэробной микрофлоры.
ний курганных могильников «Литвиновский-III»
В наших объектах исследования - археологи-
(эпоха бронзы,
2 образца),
«Октябрьский-I»
ческих сосудах, заполненных смесью почвы и жи-
(Моздокский район РСО-Алания, ранний желез-
росодержащего субстрата, можно предположить
ный век, аланская культура, 25 образцов) и «Брат-
наличие отдельных микрозон с различной прони-
ские Первые курганы» (Надтеречный район Че-
цаемостью для кислорода. Накопление в грунте
ченской Республики, аланская культура, накану-
микробных липаз и эстераз, необходимых для
не гуннского вторжения, IV в. н.э., 30 образцов).
утилизации жиров, должно протекать не равно-
На курганных могильниках «Октябрьский-I» и
мерно, а с учетом избытка субстрата, который на
«Братские Первые курганы» раскопки проведены
первых стадиях разложения будет ограничивать
Институтом археологии РАН совместно с Инсти-
БИОФИЗИКА том 68
№ 6
2023
1204
КАШИРСКАЯ и др.
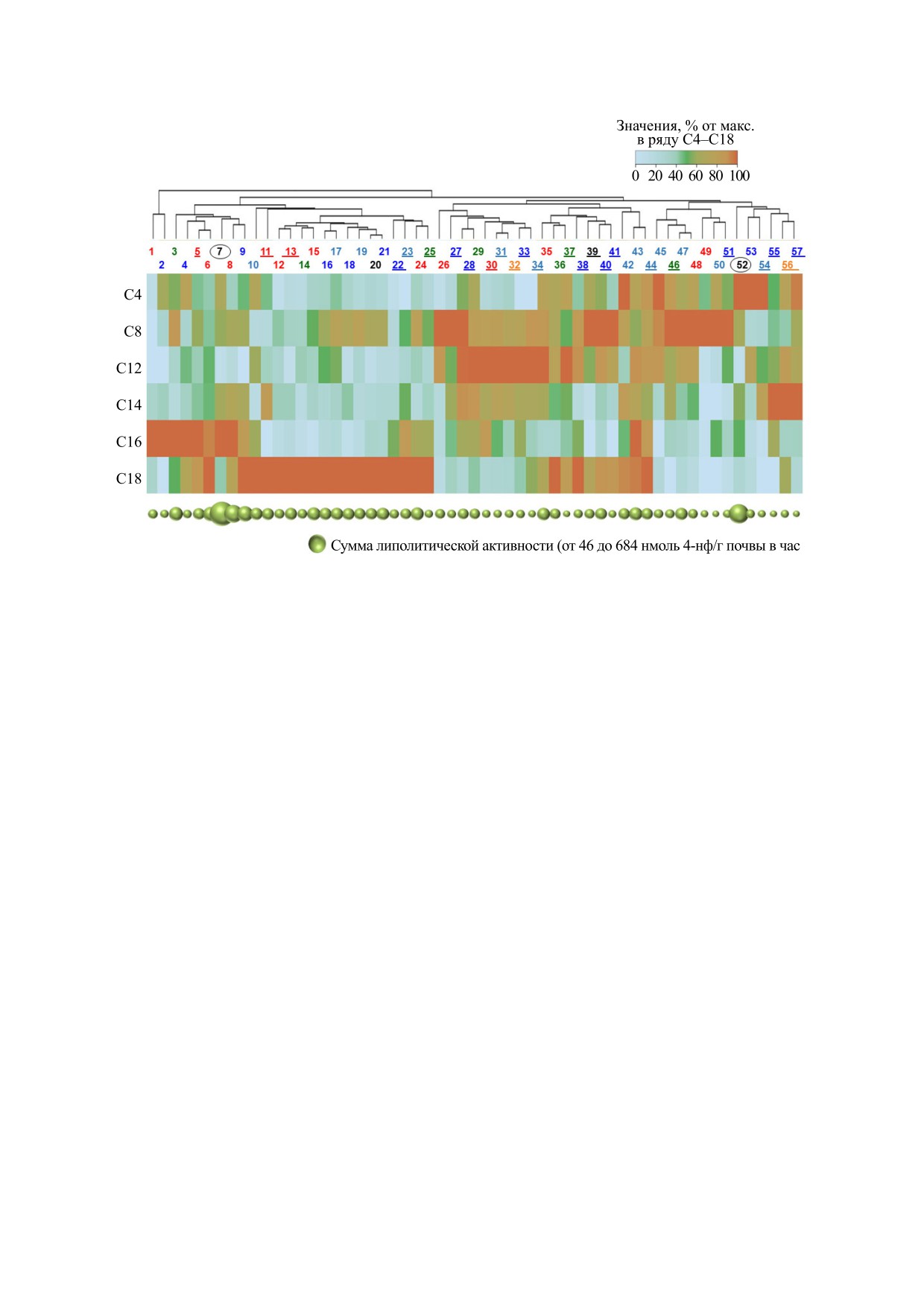
Рис. 1. Кластеризованная тепловая карта активности липолитических ферментов в грунте из древней погребальной
посуды. Цветовые обозначения типа посуды по номерам внутри выборки: черный - горшки, красный - кружки,
оранжевый - корчаги, зеленый - миски, голубой - кувшинчики, синий - кувшины. Обведены номера образцов из
погребений курганного могильника
«Литвиновский-III» (эпоха бронзы), подчеркнуты номера образцов из
погребений могильника «Октябрьский-I» (аланская культура). Остальные образцы относятся к объекту «Братские
Первые курганы» (аланская культура).
тутом гуманитарных исследований АН ЧР и ООО
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
НПЦ «ДАРС» в зоне строительства магистраль-
Рассматриваемые 57 горшков были идентифи-
ного газопровода «Моздок-Грозный» [52]. Боль-
цированы как «не пустые» среди 66 исследован-
шая часть находок имела характерные признаки
ных горшков. В этих горшках суммарная липаз-
посуды аланской культуры: миски с загнутым
ная активность составляла выше 7% от ее макси-
внутрь бортиком, кувшинчики с ручкой, распо-
мальной
величины.
При построении
ложенной на плечиках или в месте максимально-
кластеризованной тепловой карты (рис. 1) для
каждого «не пустого» горшка наибольшая актив-
го расширения тулова, низкие кружки c ручкой,
ность липолитического фермента в ряду С4 -
крепящейся к венчику сверху [53].
С8 - С12 - С14 - С16 - С18 принималась за 100%,
остальные варианты рассчитывались. Кластери-
Образцы грунта отбирали из придонного слоя
зация была проведена только в ряду погребаль-
заполнения горшков толщиной 1-2 см. Пробы
ных горшков, тогда как ряд липолитических фер-
отбирали в поле, с соблюдением асептических
ментов не группировался, с целью сохранения
условий. В лабораторию образцы доставлены в
последовательности от С4 до С18.
пластиковых пакетах и хранились при комнатной
Горшки 1 и 2 выделили в отдельную группу,
температуре. Липазную активность грунта опре-
где была обнаружена только активность липазы
деляли микропланшетным методом с использо-
С16. Остальные варианты в пределах группы не
ванием хромогенно меченных субстратов на ос-
давали значительного вклада в сумму липолити-
нове п-нитрофенола и процедуры гетеромолеку-
ческой активности.
лярного обмена
[54,
55]. Эксперименты
На втором уровне горшки 3-57 образовали две
проводили в трех повторностях. Статистическую
большие подгруппы: горшки 3-25 с высокой ак-
обработку данных проводили с помощью постро-
тивностью липаз С16 и С18 и горшки 26-57, где
ения кластеризованных тепловых карт [56].
была выявлена высокая активность вариантов со
БИОФИЗИКА том 68
№ 6
2023
ЛИПАЗНАЯ И ЭСТЕРАЗНАЯ АКТИВНОСТЬ В ГРУНТАХ
1205
средней длиной жирнокислотной последователь-
сти, как правило, обладали наиболее высокой
ности.
суммарной липолитической активностью. В этих
вариантах погребальных сосудов, скорее всего,
Подгруппа второго уровня 3-25 включала ряд
присутствовала пища с высоким содержанием ба-
погребальных горшков с наиболее высокой сум-
раньего жира. В процессе развития микробного
марной липолитической активностью. На тре-
сообщества в системе «пищевой продукт-почва»,
тьем уровне кластеризации подгруппа 3-9 харак-
теризовалась наибольшим вкладом липазы С16 в
преимущество получали продуценты истинных
липаз с высокой специфичностью к триглицери-
суммарную активность и заметным вкладом эсте-
дам длинноцепочечных жирных кислот, состав-
раз С4 и С8. Грунт из кружек № 5, 6 и 8 отличался
ляющих наибольшую долю в составе жиров жи-
пониженной активностью эстеразы С-4 по срав-
вотного происхождения. При этом ингибирова-
нению с остальными типами сосудов. Подгруппа
третьего уровня 10-25 объединялась на основе
ние естественной эстеразной активности
микробного сообщества могло происходить за
максимальной активности липазы С-18. Здесь
счет высоких концентраций этих жирных кислот
выделялись кувшинчик 10 и кружка 11 с высокой
[36, 57]. Варианты 35-44, где активность липазы
долей активности эстераз С4; кружки и миска 12-
С18 в большинстве случаев была сопоставима с
15 с низкой активностью всех вариантов липоли-
тических ферментов, кроме С18; кувшины, кув-
активностью С12 и С8 и, в ряде случаев - с актив-
ностью С14 и С4, могут указывать на меньшее со-
шинчики и горшок 16-21 с повышенной активно-
держание жира в пищевом продукте.
стью эстеразы С8 и четыре сосуда различного ти-
па 22-25 с заметным увеличением активности
Варианты 45-57 с низкой активностью истин-
С16.
ных липаз и максимальной долей эстераз, как
правило, отличались меньшими значениями сум-
Подгруппа второго уровня 26-57 включала ва-
рианты с меньшей суммарной липолитической
марной липолитической активности по сравне-
нию с вариантами 35-44. Исключение составлял
активностью. Различия средних значений этого
горшок 52 из курганного могильника Литвинов-
показателя между подгруппами 3-25 и 26-57 со-
ский, с экстремально высокой активностью ли-
ставляли 1.7 раза, а коэффициент вариации раз-
политического комплекса. В этом горшке исход-
личался незначительно, составляя 65.6 и 66.3%
для первой и второй подгрупп соответственно. В
ное наличие жирной пищи не вызывает сомне-
ний и подтверждается высокой численностью
обеих подгруппах степень неоднородности
липолитических микроорганизмов в грунте за-
уменьшалась по сравнению с их совокупностью,
полнения. Остальные горшки рассматриваемой
где коэффициент вариации суммарной активно-
группы могли содержать пищу с низким содержа-
сти составлял 72.8%.
нием жира. Таким образом, высокая активность
На третьем уровне подгруппа 26-44 характери-
эстераз в этих горшках, вероятнее всего, является
зовалась высокой активностью среднецепочеч-
показателем высокой биологической активности
ных вариантов С8-С14, а подгруппа 45-57 демон-
грунта заполнения.
стрировала увеличение доли эстераз С4 и С8 в
Наиболее сложным для интерпретации случа-
суммарной липолитической активности.
ем представляется максимальный вклад липаз со
Подгруппа четвертого уровня 26-34, включаю-
средней длиной жирнокислотной последова-
щая посуду большинства представленных типов,
тельности в суммарную липолитическую актив-
отличалась минимальной активностью липазы
ность. Варианты 26-34 с низкой активностью ис-
С18, неоднородными показателями активности
тинных липаз и, в большинстве случаев, с низкой
липазы С16 и высокой активностью вариантов
активностью эстераз, теоретически могли содер-
С8-С14 с максимумом С12 (за исключением
жать субстрат, насыщенный остатками лаурино-
горшков 26 и 27 с максимумом С8).
вой и миристиновой жирных кислот С12 и С14.
Подгруппа четвертого уровня 5-44 показывала
Их доля достаточно высока в молочном жире, од-
синхронное увеличение доли эстеразы С4 и липа-
нако для лауриновой кислоты она составляет все-
зы С18, которое сопровождалось заметным сни-
го 2.7%, что почти на порядок меньше по сравне-
жением вклада среднецепочечных вариантов С12
нию с пальмитиновой кислотой С16. В тоже вре-
и С14.
мя содержание в молочном жире миристиновой
Таким образом, в общей совокупности из 57
кислоты С14 близко к содержанию стеариновой
единиц посуды с предполагаемым исходным на-
кислоты С18 и составляет около 10%. Таким обра-
личием жиросодержащей пищи было выделено
зом, молочный жир с большей вероятностью
несколько групп с различным соотношением ак-
присутствовал не в сосудах подгруппы 26-34, а в
тивности липаз и эстераз. Варианты 12-25 с высо-
сосудах подгруппы 3-9, с максимальной актив-
кой активностью истинных липаз С18, на фоне
ностью липазы С16 и отсутствием видимых при-
относительно низких значений активности эсте-
знаков ингибирования микробных эстераз. По-
раз и липаз, предпочитающих субстраты со сред-
следний факт косвенно свидетельствует о нали-
ней длиной жирнокислотной последовательно-
чии достаточного количества белка в молочном
БИОФИЗИКА том 68
№ 6
2023
1206
КАШИРСКАЯ и др.
симальную [28] активность по отношению к
остатку лауриновой кислоты С12.
Таким образом, в результате кластерного ана-
лиза активности липолитических ферментов в
грунте со дна горшков из древних погребений бы-
ли выявлены варианты с различным соотноше-
нием эстераз и липаз. Две группы горшков - 12-
25 и 26-34 - демонстрировали ингибирование
эстеразной активности микробного сообщества
на фоне максимальной активности липаз С18 и
С12, что может свидетельствовать о наличии про-
дукта с высоким содержанием жира и низким со-
держанием белка.
В грунтах на дне остальных горшков была вы-
явлена заметная эстеразная активность, что сви-
детельствует об относительном благополучии
микробного сообщества почвы, смешанной с
другими пищевыми субстратами. Как мы полага-
ем, эти субстраты с наибольшей вероятностью
могли содержать бараний жир (варианты 35-44),
молочный жир (варианты 3-9) или незначитель-
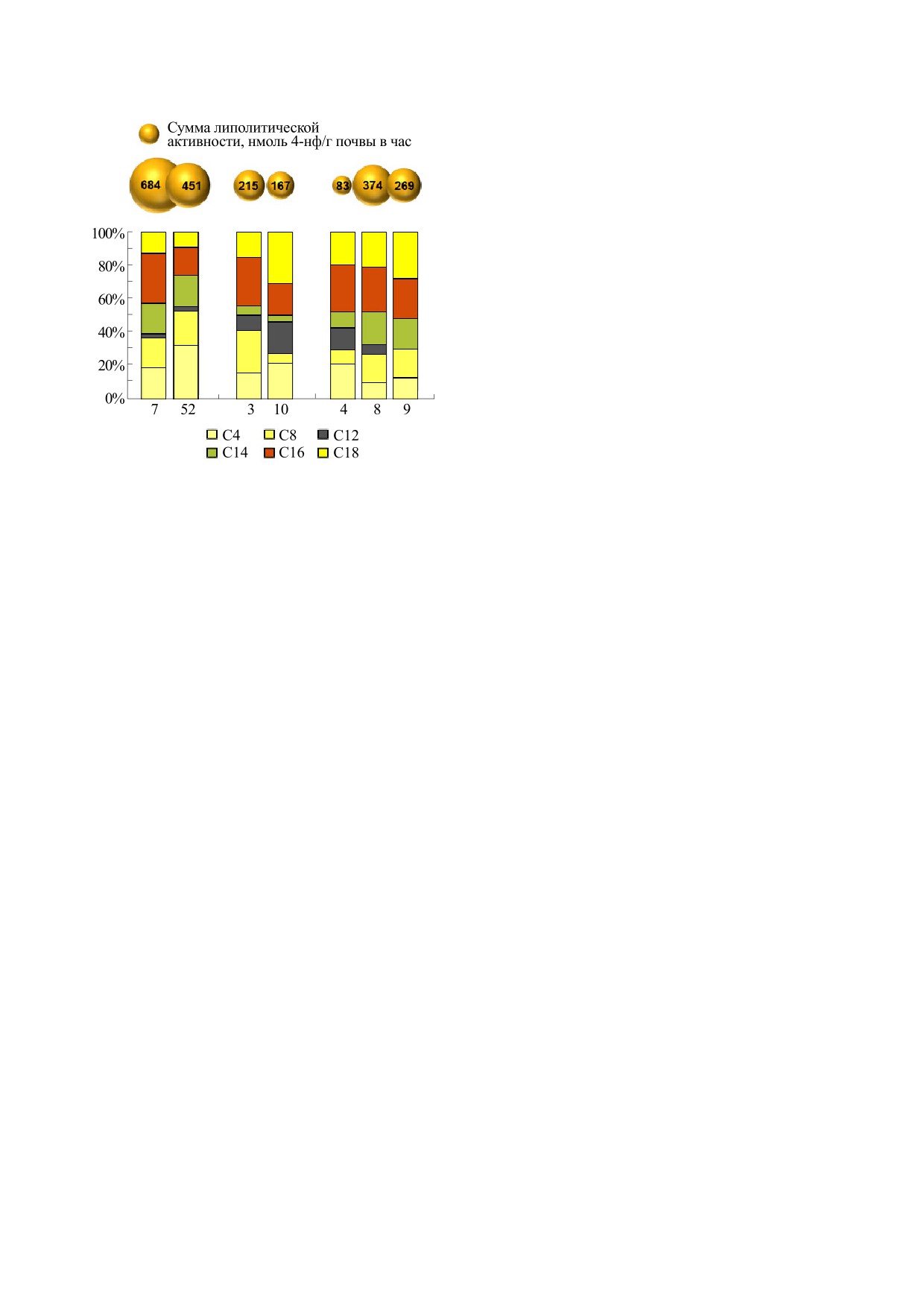
Рис. 2. Сумма и соотношение активности липолити-
ное количество жира (варианты 45-57).
ческих ферментов в грунте со дна некоторых горшков
из древних погребений (номера соответствуют номе-
В горшках из близких погребений 9 и 10 перво-
рам на рис. 1). Образцы 7, 52 - горшки из погребений
го кургана могильника
«Литвиновский III»
9 и 10 первого кургана в составе могильника эпохи
бронзы «Литвиновский-III». Образцы 3 и 10 - из
(рис. 2) была отмечена предельно высокая сум-
миски и кувшинчика детского погребения 1355; об-
марная липолитическая активность грунта. В об-
разцы 4, 8 и 9 - кувшин, кружка и кувшин-подноше-
разце с максимальной суммарной активностью
ние из семейного погребения 1452 курганного мо-
активность эстераз С4 и С8 составляла 37%, а ак-
гильника аланской культуры «Братские Первые кур-
ганы».
тивность истинных липаз С16 и С18 - 42%. Дру-
гой образец показывал значительно бóльшую до-
лю активности эстераз - 53%, причем активность
продукте. Необходимо отметить, что из грунта со
истинных липаз в сумме составляла всего 26%.
дна сосудов подгруппы 3-9, за исключением об-
Миска и кувшинчик из детского погребения 1355
разца 5, были выделены культуры бацилл, спо-
курганного могильника «Братские Первые курга-
собные к сбраживанию молока.
ны» значительно различались по соотношению
липаз и эстераз. В грунте из миски была выявлена
В большинстве сложно интерпретируемых ва-
высокая доля активности эстераз - более 40% от
риантов 26-34 липаза С16 проявляла заметную
суммы, а также максимальный вклад активности
активность, однако меньшую по сравнению с ли-
липазы С16. В грунте из кувшинчика активность
пазой С12. Кроме того, здесь имело место инги-
эстераз не превышала 30% от суммарной актив-
бирование эстеразной активности микробного
ности, а наибольшей активностью отличалась ли-
сообщества, что может объясняться значитель-
паза С-18.
ной концентрацией длинноцепочечных жирных
Кувшин и кружка, принадлежавшие женщине
кислот в составе субстрата. Не исключено, что в
из семейного погребения 1452 могильника «Брат-
сосудах группы 26-34 также мог присутствовать
ские Первые курганы», а также кувшин-подно-
молочный жир, но более концентрированный по
шение, найденный у входа этого катакомбного
сравнению с вариантами 3-9. Максимальная ак-
погребения, имели свои особенности в соотно-
тивность липаз С12 в данном случае может быть
шении активности липолитических ферментов.
связана с особенностями микробного сообще-
Наименьшей суммарной активностью и наиболь-
ства, где преимущество получают определенные
шей долей активности эстеразы С4 отличался
группы микроорганизмов, вырабатывающие ли-
грунт из кувшина. Грунт из кружки обладал мак-
пазу именно с такими свойствами. Как было по-
симальной липолитической активностью среди
казано выше, некоторые микробные липолити-
всех изученных сосудов аланской культуры. Здесь
ческие ферменты (а также их группы), способные
при близком соотношении активности истинных
к расщеплению широкого спектра субстратов,
липаз уменьшалась доля активности липолитиче-
хотя в целом уменьшают свою активность при
ских ферментов С4 и С12 и увеличивалась доля
увеличении длины жирнокислотной цепи, одна-
активности ферментов С8 и С14. В грунте из кув-
ко проявляют более высокую [27] или даже мак-
шина-подношения не выявлялась активность ли-
БИОФИЗИКА том 68
№ 6
2023
ЛИПАЗНАЯ И ЭСТЕРАЗНАЯ АКТИВНОСТЬ В ГРУНТАХ
1207
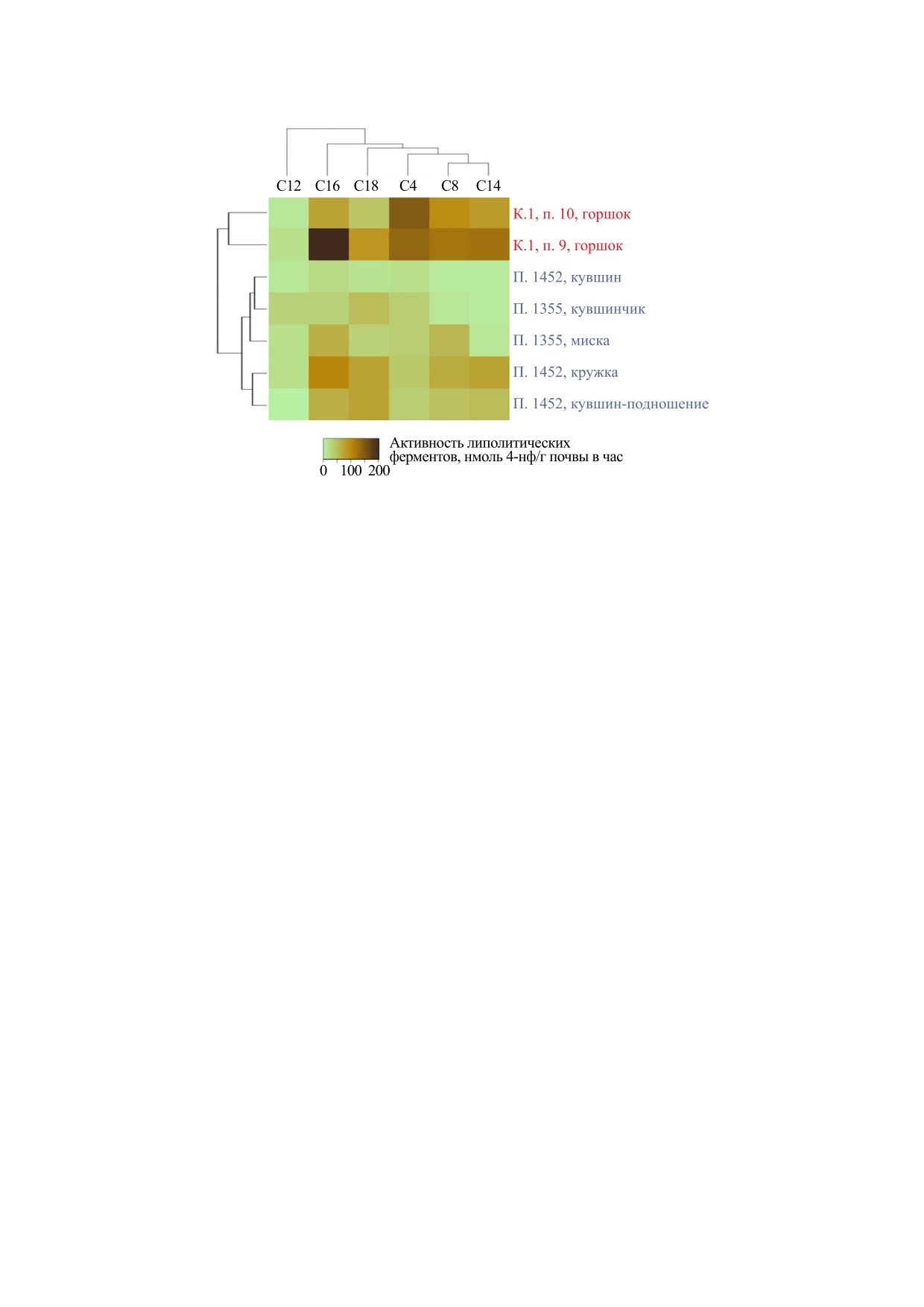
Рис. 3. Кластеризованная тепловая карта активности липолитических ферментов в грунте со дна некоторых сосудов
из древних погребений. Красным цветом обозначены сосуды из курганного могильника «Литвиновский-III» (эпоха
бронзы), синим - «Братские Первые курганы» (аланская культура).
пазы С12. По соотношению активности осталь-
ков наблюдалась эстеразная активность на фоне
ных ферментов этот кувшин имел максимальное
высокой относительной активности липаз С16,
сходство с кружкой внутри погребения, за исклю-
средней относительной активности липаз С18 и
чением более высокой активности липазы С18.
предельно низкой активности истинных липаз.
Посуда из погребения 1452 входит в одну группу
Группировка горшков по соотношению липазной
(рис. 1), где предполагается исходное присут-
и эстеразной активности отражает характеристи-
ствие молочного жира. При этом кувшин-подно-
ки их исходного содержимого, тогда как абсолют-
шение 9 и детская миска 10 из соседней группы
ные значения активности ферментов, среди из-
начинают ряд с максимальной активностью ли-
бранной посуды с наибольшей суммарной липо-
пазы С18. Из грунта сосудов аланской культуры
литической
активностью,
могут
быть
(рис. 2) выделены бактерии, способные к сбражи-
группированы в соответствии с типом погребе-
ванию молока.
ния. Таким образом, оценка баланса липазной и
Для этой же группы сосудов показано значи-
эстеразной активности дает полезную информа-
тельное отставание активности липазы С12 от
цию для реконструкции исходного содержания
остальных липолитических ферментов (рис. 3) и
пищи в погребальных сосудах.
группировка большей части сосудов по погребе-
ниям, за исключением женского кувшина из по-
гребения 1452, имеющего наиболее близкое сход-
ство с детским кувшинчиком из погребения 1355
БЛАГОДАРНОСТИ
по низкой активности эстеразы С8 и липазы С14.
Авторы выражают благодарности археологу
В.Ю. Малашеву (Институт археологии РАН,
ЗАКЛЮЧЕНИЕ
Москва) за предоставленные образцы выборки.
В результате кластерного анализа липолитиче-
ской активности в грунте дна горшков из древних
погребений были выявлены варианты с различ-
ФИНАНСИРОВАНИЕ РАБОТЫ
ным соотношением эстераз и липаз. Две группы
Работа выполнена при финансовой поддержке
горшков показывали предельно низкие значения
Российского научного фонда (грант № 22-28-
эстеразной активности почвенного микробного
сообщества на фоне максимальной активности
01725 «Почвенно-микробиологические, энзимо-
липаз С18 и С12, что может быть связано с инги-
логические и молекулярно-биологические под-
бированием эстераз в присутствии высоких кон-
ходы к идентификации пищи в сосудах из погре-
центраций жирных кислот. В трех группах горш-
бений», рук. Н.Н. Каширская).
БИОФИЗИКА том 68
№ 6
2023
1208
КАШИРСКАЯ и др.
КОНФЛИКТ ИНТЕРЕСОВ
21. L. Casas-Godoy, S. Duquesne, F. Bordes, et al., Meth-
ods Mol. Biol., 861, 3 (2012). DOI: 10.1007/978-1-
Авторы заявляют, что у них нет конфликта ин-
61779-600-5_1
тересов.
22. А. М. Безбородов и Н. А. Загустина, Прикладная
биохимия и микробиология, 50 (4), 347 (2014).
СОБЛЮДЕНИЕ ЭТИЧЕСКИХ СТАНДАРТОВ
23. F. Akram, A. S. Mir, and A. Roohi, Mol. Biotechnol.,
65, 521 (2023). DOI: 10.1007/s12033-022-00592-z
Настоящая работа не содержит экспериментов
24. T. C. A. Hitch and T. Clavel, PeerJ., 7, e7249 (2019).
с использованием людей и животных в качестве
DOI: 10.7717/peerj.7249
объектов исследований.
25. В. С. Гамаюрова, К. Л. Шнайдер и М. Е. Зи-
новьева, Вестн. Казанского технологич. ун-та, 17
СПИСОК ЛИТЕРАТУРЫ
(24), 175 (2014).
26. A. Glogauer, V. P. Martini, H. Faoro, Microb. Cell
1.
R. P. Evershed, Archaeometry, 50, 895 (2008). DOI:
Fact. 10, 54 (2011). DOI: 10.1186/1475-2859-10-54
10.1111/j.1475-4754.2008.00446.x
27. P. Tirawongsaroj, R. Sriprang, P. Harnpicharnchai,
2.
L. J. Cramp, I. D. Bull, E. Casanova, et al., in Lipids in
et al., J. Biotechnol.,
133
(1),
42
(2008). DOI:
Archaeology. Handbook of Archaeological Sciences
10.1016/j.jbiotec.2007.08.046
(2023)., V. 1, p. 529
28. M. Parapouli, A. Foukis, P. Y. Stergiou, et al., J. Biol.
3.
A. Irto, G. Micalizzi, C. Bretti, et al., Molecules, 27
Res.-Thessaloniki, 25, 4 (2018). DOI: 10.1186/s40709-
(11), 3451 (2022).
018-0074-6
4.
M. S. Copley, F. A. Hansel, K. Sadr, and R. P. Ever-
29. Е. В. Чернышева, А. В. Борисов и В. Ю. Малашев,
shed, South Afr. J. Sci., 100 (5), 279 (2004).
Краткие сообщения Института археологии,
5.
R. P. Evershed, S. N. Dudd, M. S. Copley, et al., Ac-
№ 263,
105
(2021). DOI:
10.25681/IARAS.0130-
counts Chem. Res., 35 (8), 660 (2002).
2620.263.105-116
6.
S. Hammann, D. J. Scurr, M. R. Alexander, and
30. Н. Н. Каширская, Е. В. Чернышева и В. Ю.
L. J. Cramp, Proc. Natl. Acad. Sci., 117 (26), 14688
Малашев, История, археология и этнография
(2020).
Кавказа, 19 (1), 150 (2023).
7.
M. P. Colombini, F. Modugno, andE. Ribechini, J.
31. Т. Э. Хомутова, Т. С. Демкина, А. В. Борисов и
Mass Spectrometry, 40 (7), 890 (2005).
Н. И. Шишлина, Почвоведение, № 2, 239 (2017).
8.
M. Bondetti, et al., Archaeometry, 63 (3), 594 (2021).
DOI: 10.7868/S0032180X1702006X
9.
M. S. Copley, P. J. Rose, A. Clapham, et al., Proc. Roy.
32. Л. Н. Плеханова, С. Н. Удальцов, А. С. Сыроватко
Soc. Lond. Ser. B: Biol. Sci., 268 (1467), 593 (2001).
и Н. Н. Каширская, Биофизика, 68 (5), 999 (2023).
DOI: 10.31857/S0006302923050204
10.
R. Sacchi, et al., Science of Food, 4 (1), 19 (2020).
33. L. N. Plekhanova, in Proc. 19th Int. Multidisc. Sci. Geo-
11.
S. N. Dudd, R. P. Evershed, and A. M. Gibson, J. Ar-
conf. SGEM
2018,
18
(5.2),
173
(2018). DOI:
chaeol. Sci., 26 (12), 1473 (1999).
10.5593/sgem2018/5.2/S20.023
12.
E. Casanova, T. D. Knowles, C. Ford, et al., Radiocar-
34. T. E. Khomutova and A. V. Borisov, J. Arid Environ.,
bon, 62 (6), 1679 (2020).
171,
104004
(2019).
DOI:
10.1016/
13.
Н. Н. Каширская, Л. Н. Плеханова, С. Н. Удальцов
j.jaridenv.2019.104004
и др., Биофизика,
62
(6),
1235
(2017). DOI:
35. В. А. Демкин, Т. С. Демкина и С. Н. Удальцов,
10.1134/S0006350917060094
Вестн. археологии, антропологии и этнографии,
14.
Н. Н. Каширская, Л. Н. Плеханова, Е. В. Чер-
№ 2 (25), 148 (2014).
нышева и др., Почвоведение, № 1, 89 (2020). DOI:
36. C. Ruiz, S. Falcocchio, E. Xoxi, et al., Biochim. Bio-
10.1134/S0032180X20010098.
phys. Acta - General Subjects, 1672 (3), 184 (2004).
15.
M. W. I. Schmidt, M. S. Torn, S. Abiven, et al., Nature,
37. J. L. Smith and J. A. Alford, Appl. Microbiol., 14 (5),
478, 49 (2011).
699 (1966).
16.
В. М. Cеменов, А. C. Тулина, Н. А. Cеменова и
38. P. Nobmann, A. Smith, J. Dunne, et al., Int. J. Food
Л. А. Иванникова, Почвоведение, № 4, 393 (2013).
Microb., 128 (3), 440 (2009).
DOI: 10.7868/S0032180X13040114
39. C. Q. Sun, C. J. O'Connor, and A. M. Roberton,
17.
M. Von Lützow, I. Kögel-Knabner, B. Ludwig, et al.,
Chem.-biol. Interactions, 140 (2), 185 (2002).
J. Plant Nutr. Soil Sci., 171, 111 (2008).
40. C. Zhong, N. Zhu, Y. Zhu, et al., Eur. J. Pharmaceut.
18.
В. А. Намиот, А. В. Батяновcкий, И. В. Филатов
Sci., 141, 105123 (2020).
и дp., Биофизика, 61 (1), 54 (2016).
41. K. Sugimoto, H. Tanaka, R. Moriyama, Biocontrol
19.
Е. В. Чернышева, М. А. Амирян, М. В. Ельцов
Sci., 3 (1), 17 (1998).
и др., Ethnography, 18 (4), 993 (2022).
42. J. Lalman and D. M. Bagley, Water Res., 36 (13), 3307
20. Е. В. Чернышева, Н. Н. Каширская и К. С. Ду-
(2002).
щанова, Вестн. археологии, антропологии и
43. C. S. Hwu, S. K. Tseng, C. Y. Yuan, et al., Water Res.,
этнографии, № 2 (61), 113 (2023).
32 (5), 1571 (1998).
БИОФИЗИКА том 68
№ 6
2023
ЛИПАЗНАЯ И ЭСТЕРАЗНАЯ АКТИВНОСТЬ В ГРУНТАХ
1209
44. J. Palatsi, M. Laureni, M. V. Andrés, et al., Bioresource
A. N. S. Pandey and C. R. Soccol (Elsevier, 2017),
Technol., 100 (20), 4588 (2009).
p. 175.
45. M. A. Pereira, O. C. Pires, M. Mota, and M. M. Alves,
52. В. Ю. Малашев, Р. Г. Магомедов, Ф. С. Дзуцев
Biotechnology Bioengineer., 92 (1), 15 (2005).
и др., Археологические открытия, 2018, 342 (2020).
53. Д. С. Коробов, В. Ю. Малашев, и Й. Фассбиндер,
46. J. Palatsi, M. Viñas, M. Guivernau, Bioresource Tech-
Краткие сообщения Института археологии,
nol., 102 (3), 2219 (2011).
№ 260, 441 (2020).
47. S. M. Iskander, Y. M. Amha, P. Wang, et al., Front.
54. A. Cowie, V. E. Lonergan, F. S. M. Rabbi, et al., Soil
Bioengineer. Biotechnol., 9, 613626 (2021).
Res., 51, 707 (2013). DOI: 10.1071/SR13043
48. Z. Guo, M. Usman, S. A. Alsareii, et al., Bioresource
55. A. J. Margenot, Y. Nakayama, and S. J. Parikh, Soil
Technol., 337, 125383 (2021).
Biol. Biochem., 125, 350 (2018). DOI: 10.1016/j.soil-
49. H. Wang, Q. Yan, X. Zhong, et al., Bioresource Tech-
bio.2017.11.006
nol., 386, 129538 (2023).
56. S. Zhao, Y. Guo, Q. Sheng, and Y. Shyr, BioMed Res.
50. S. Amir, G. Merlina, E. Pinelli, et al., J. Hazardous
Int., 2014, 986048 (2014). DOI: 10.1155/2014/986048
Mater., 159 (2-3), 593 (2008).
57. Д. А. Белинская и Н. В. Гончаров, Биоорган.
51. R. Gaur, R. Hemamalini, and S. Khare, in Current De-
химия,
46
(3),
247
(2020). DOI:
10.31857/
velopments in Biotechnology and Bioengineering, Ed. by
S0132342320030045
Lipase and Esterase Activity in Soils from Ancient Burial Vessels
N.N. Kashirskaya*, E.V. Chernysheva*, S.N. Udaltsov*, and L.N. Plekhanov*
*Institute of Physicochemical and Biological Problems of Soil Science, Russian Academy of Sciences,
Institutskaya ul. 2, Pushchino, Moscow region, 142290 Russia
The activity of lipolytic enzymes was assessed using the microplate method using heteromolecular exchange
in soils from the bottom of burial vessels, followed by cluster analysis. Groups with a contrasting ratio of li-
pases C12 - C18 and esterases C4 - C8 were identified in a sample that included 57 burial pots aged two to
three thousand years. The total lipolytic activity of soil samples from dishes with the presumptive initial pres-
ence of fat-containing food products was above 6% of the maximum value within the sample. Inhibition of
esterases in the presence of high concentrations of fatty acids is reflected in the extremely low relative values
of esterase activity of the soil microbial community against the background of maximum activity levels of C18
and C12 lipases in two groups of pots. In three groups, esterase activity was observed against the background
of high relative activity of C16 lipases, average relative activity of C18 lipases and extremely low activity of true
lipases. In the group of pots with high total lipolytic activity (with the exception of certain extreme values of
this indicator), the proportion of true lipase activity was 1.7-1.8 times higher compared to the proportion of
esterase activity. The grouping of burial vessels by the ratio of lipase and esterase activity reflects the charac-
teristics of the funeral food, while the absolute values of enzyme activity among selected vessels with the high-
est total lipolytic activity are grouped according to the type of burial.
Keywords: lipolytic enzymes, lipase, esterase, fatty acids, funerary vessels, dishes, pots, funeral food, ancient food,
Bronze Age, early Iron Age
БИОФИЗИКА том 68
№ 6
2023