БИОХИМИЯ, 2019, том 84, вып. 10, с. 1401 - 1409
УДК 577.217.5
ТРЕТИЙ ФАКТОР ИНИЦИАЦИИ ТРАНСЛЯЦИИ
В МИТОХОНДРИЯХ: СТРУКТУРА, ФУНКЦИИ, ВЗАИМОДЕЙСТВИЯ
И РОЛЬ В ЗДОРОВЬЕ И БОЛЕЗНЯХ ЧЕЛОВЕКА
Мини обзор
© 2019 И.В. Чичерин1,2*, M.В. Балева1, С.A. Левицкий1,
Э.Б. Дашинимаев3,4, И.А. Крашенинников1
1 Московский государственный университет им. М.В. Ломоносова, биологический факультет,
119234 Москва, Россия; электронная почта: i.v.chicherin@gmail.com
2 Московский государственный университет им. М.В. Ломоносова,
Институт функциональной геномики, 119234 Москва, Россия
3 Институт биологии развития им. Н.К. Кольцова РАН, 119334 Москва, Россия
4 Российский национальный исследовательский медицинский
университет им. Н.И. Пирогова, 117977 Москва, Россия
Поступила в редакцию 29.03.2019
После доработки 25.04.2019
Принята к публикации 23.06.2019
Митохондрии - важнейшие органеллы эукариотической клетки, обеспечивающие дыхание посредством
работы электрон!транспортной цепи. Экспрессия генов в митохондриях происходит по образцу бактери!
альных систем, однако между ними существует множество эволюционных различий. Одним из них являет!
ся организация процесса инициации трансляции. Данный обзор посвящен третьему фактору инициации
трансляции в митохондриях - белку, играющему ключевую роль в экспрессии генома органелл. Обсуждает!
ся его роль в здоровье и болезнях человека.
КЛЮЧЕВЫЕ СЛОВА: IF3mt, митохондрии, трансляция, рибосомы, болезнь Паркинсона.
DOI: 10.1134/S0320972519100038
По современным оценкам, протеом митохон!
торые характеризуются повышенным содержа!
дрий составляет от 1000 до 1500 индивидуаль!
нием белков, измененной морфологией и пере!
ных белков [1]. Абсолютное большинство из них
стройкой функциональных центров [4]. Другой
(~99%) кодируется в ядре, синтезируется в ци!
особенностью являются структуры мРНК, в ко!
топлазме и затем попадает в органеллы по кана!
торых практически отсутствуют 5'!НТО, что
лам импорта белков. Однако 13 белок!кодирую!
предполагает альтернативный механизм иници!
щих генов не мигрировали в ядро в ходе эволю!
ации трансляции [5]; кроме того, тРНК митохон!
ции и сохранились в митохондриальной ДНК
дрий характеризуются потерей некоторых клас!
[2]. Все они являются центральными компонен!
сических структурных мотивов [6]. Особенно
тами комплексов электрон!транспортной цепи.
впечатляющие различия можно найти на этапе
Митохондрии млекопитающих обладают всем
инициации трансляции. Хорошо известно, что
необходимым оборудованием для экспрессии
данный процесс управляется тремя каноничес!
этих генов.
кими факторами: IF1, IF2 и IF3. Однако мито!
Синтез белка в митохондриях организован
хондрии млекопитающих располагают только
по классической схеме, изученной у бактерий, и
двумя из них, а именно: IF2mt (выполняет роль
включает в себя несколько универсальных эта!
IF1 и IF2) и IF3mt [7]. Оба фактора следуют об!
пов, таких как инициация, элонгация, термина!
щему направлению организации белкового син!
ция и рециклинг [3]. При этом некоторые зна!
теза в митохондриях, являясь гомологами про!
чительные различия в компонентах системы
кариотических факторов, и в то же время имея
позволяют выделить митохондриальную транс!
собственные уникальные черты.
ляцию в отдельную группу. Одним из них явля!
В данном обзоре собрана вся информация о
ется структура митохондриальных рибосом, ко! IF3mt с учетом недавно появившихся структур!
ных и функциональных исследований, показы!
вающих его влияние на здоровье и болезни че!
* Адресат для корреспонденции.
ловека.
3
1401
1402
ЧИЧЕРИН и др.
СТРУКТУРНЫЕ ЭЛЕМЕНТЫ IF3mt
ла исследована только в IF3 прокариот и мито!
ЧЕЛОВЕКА И ИХ РОЛЬ В РАБОТЕ
хондрий человека.
ФАКТОРА
Значительные изменения наблюдались в
N! и C!концевых участках IF3mt: оба из них
IF3 прокариот состоит из двух глобулярных
имели примерно 30 нт удлинения, отсутствую!
доменов, соединенных гибким мостиком [8].
щие у IF3 бактерий (рис. 2). Стоит отметить, что
Белок выполняет несколько функций: 1) препят!
наличие таких удлинений наблюдается у многих
ствует ассоциации свободных 30S и 50S субъ!
белков!участников митохондриального транс!
единиц, сохраняя пул свободных 30S субъеди!
ляционного аппарата [13]. Считается, что эти
ниц для инициации трансляции; 2) способству!
участки возникли как приспособления белков к
ет взаимодействию кодона с антикодоном в
специфическим условиям работы в органеллах,
P!сайте и формированию 30S инициаторного
что вызывает к ним живой интерес исследовате!
комплекса; 3) обеспечивает правильное поло!
лей в данной области знания.
жение мРНК на 30S субъединице; 4) разрушает
Основная идея о том, как исследовать роль
ложные и неправильно собранные инициатор!
данных удлинений, довольно проста - удалить
ные комплексы [9]. N! и C!концевой домены
их и посмотреть, что получится. Этот подход
взаимодействуют с рибосомой независимо друг
был применен к N! и C!концам IF3mt человека
от друга; в то время как C!домен абсолютно не!
[14]. К удивлению ученых, удаление концов
обходим и способен выполнять всю работу фак!
слегка повышало инициаторную активность
тора, N!домен играет скорее вспомогательную
фактора как с митохондриальными, так и с бак!
роль [10].
териальными рибосомами. Однако, другая ак!
Человеческий IF3mt был впервые обнаружен
тивность белка IF3mt, а именно разборка непра!
в 2002 г. при анализе базы данных библиотек
вильных комплексов, собранных без мРНК,
кДНК экспрессионных фрагментов (EST) чело!
сильно зависела от наличия С!конца. При этом
века, используя известные последовательности
разборка других неправильных комплексов,
IF3 в качестве образцов [11]. Он относится к се!
собранных без инициаторной тРНК и факторов
мейству белков IF3, в состав которого входят го!
трансляции, от N! и C!концов никак не зависе!
мологи прокариот (E. coli, T. thermophilus), хло!
ла. Дальнейшие исследования показали, что
ропластов (A. thaliana, E. gracilis) и митохондрий
N! и C!концевые удлинения IF3mt человека
дрожжей (Aim23 in S. cerevisiae and IF3mt of
участвуют в его взаимодействии с рибосомой
S. pombe) и млекопитающих (H. sapiens, B. tau-
[15]. Так, удаление N! и C!концов лишь незна!
rus). Все члены семейства мало гомологичны по
чительно влияло на взаимодействие IF3mt с 28S
аминокислотной последовательности (от 20 до
субъединицей: такой мутантный белок (IF3mt!
25%), но некоторые консервативные остатки все
ΔNC) освобождался из малой субъединицы и за!
же можно выделить у всех видов (рис. 1). Общие
тем повторно связывался с ней медленнее, чем
черты скорее прослеживаются в доменной орга!
полноразмерный фактор. Одновременно с этим,
низации этих белков, которые подобно прока!
удаление любого из концов усиливало связыва!
риотическим предкам состоят из двух доменов,
ние IF3mt с 39S большой субъединицей. Вероят!
соединенных линкером (рис. 2). Функциональ!
но, поэтому мутант IF3mt!ΔNC обнаруживался
ная роль этих элементов сильно отличалась от
во фракции 55S рибосом и давал сложную кар!
той, что наблюдалась в бактериальном белке, но
тину при их диссоциации, отличную от нор!
было и небольшое сходство [12]. Все они могли
мального белка.
взаимодействовать с 28S субъединицей, но по!
Коровая часть гомолога IF3 из хлоропластов
разному. Сила взаимодействия свободного
E. gracilis (IF3chl) также имеет концевые удлине!
C!домена была близка к таковой полноразмер!
ния, которые больше, чем у IF3mt человека
ного IF3mt, в то время как N!домен взаимодей!
(~150 аминокислот на N!конце и 60 на C-кон!
ствовал с ней в 10 раз слабее. Мостик между до!
це). Подобно IF3mt, удаление этих участков по!
менами человеческого IF3mt оказался не только
вышало активность фактора, что говорит об их
средством соединения, но и важным участни!
регуляторной роли [16]. Совершенно другая си!
ком взаимодействия с малой субъединицей.
туация наблюдалась для гомолога IF3 из мито!
В отличие от бактериального IF3, оба домена и
хондрий дрожжей S. cerevisiae, Aim23p, в кото!
мостик IF3mt необходимы для работы белка.
ром всего лишь одного удлинения (неважно,
Свободный C!домен способен диссоциировать
N! или C!концевого) было достаточно для со!
55S рибосомы, но его активность заметно сни!
хранения активности [17]. При этом мутант
жается по сравнению с полноразмерным бел!
Aim23p!ΔNC был совершенно нефункциона!
ком. На сегодняшний день активность отдель!
лен: он не связывался с рибосомами и не разде!
ных доменов представителей семейства IF3 бы!
лял их на субъединицы, а его фенотип совпадал
БИОХИМИЯ том 84 вып. 10 2019
IF3mt: СТРУКТУРА, ФУНКЦИИ И ВЗАИМОДЕЙСТВИЯ
1403
с фенотипом делеции по гену AIM23. Эти факты
(рис. 2) IF3 полностью нарушали его взаимодей!
говорят о различиях в механизмах работы среди
ствие с 30S малой субъединицей и способность
представителей семейства IF3.
диссоциировать 70S рибосомы. Подобно этому,
мутации в спирали 3 C!домена (рис. 2) IF3mt ан!
нулировали диссоциативную активность факто!
ВЗАИМОДЕЙСТВИЕ IF3mt ЧЕЛОВЕКА
ра, однако мутантный белок по!прежнему мог
С 28S СУБЪЕДИНИЦЕЙ
связываться с 28S субъединицей в противопо!
ложность бактериальному гомологу. Это гово!
По данным экспериментов in vitro, механизм
рит о том, что две указанные активности прост!
действия IF3mt отличен от такового у прокарио!
ранственно разделены в IF3mt, но не в IF3 бак!
тического IF3 [18]. Мутации ключевых амино!
терий. Скорее всего, они осуществляются пос!
кислот в этих белках имеют сильно разные эф!
редством двух разных функциональных сайтов
фекты. Так, мутации в спирали 4 C!домена
IF3mt.
Рис. 1. Множественное выравнивание аминокислотных последовательностей Aim23p S. cerevisiae, IF3mt H. sapiens, IF3
E. coli and IF3 T. thermophilus в программе Clustal Omega. Звездочка обозначает полностью консервативный остаток; две
точки - консервативность в группе с сильно схожими свойствами; одна точка - консервативность в группе со слабо по!
хожими свойствами. Красным цветом показаны гидрофобные аминокислоты, синим - кислые, фиолетовым - основные,
зеленым - глицин и аминокислоты, содержащие !OH, !SH, =NH группы.
journal/biokhsm/
БИОХИМИЯ том 84 вып. 10 2019
3*
1404
ЧИЧЕРИН и др.
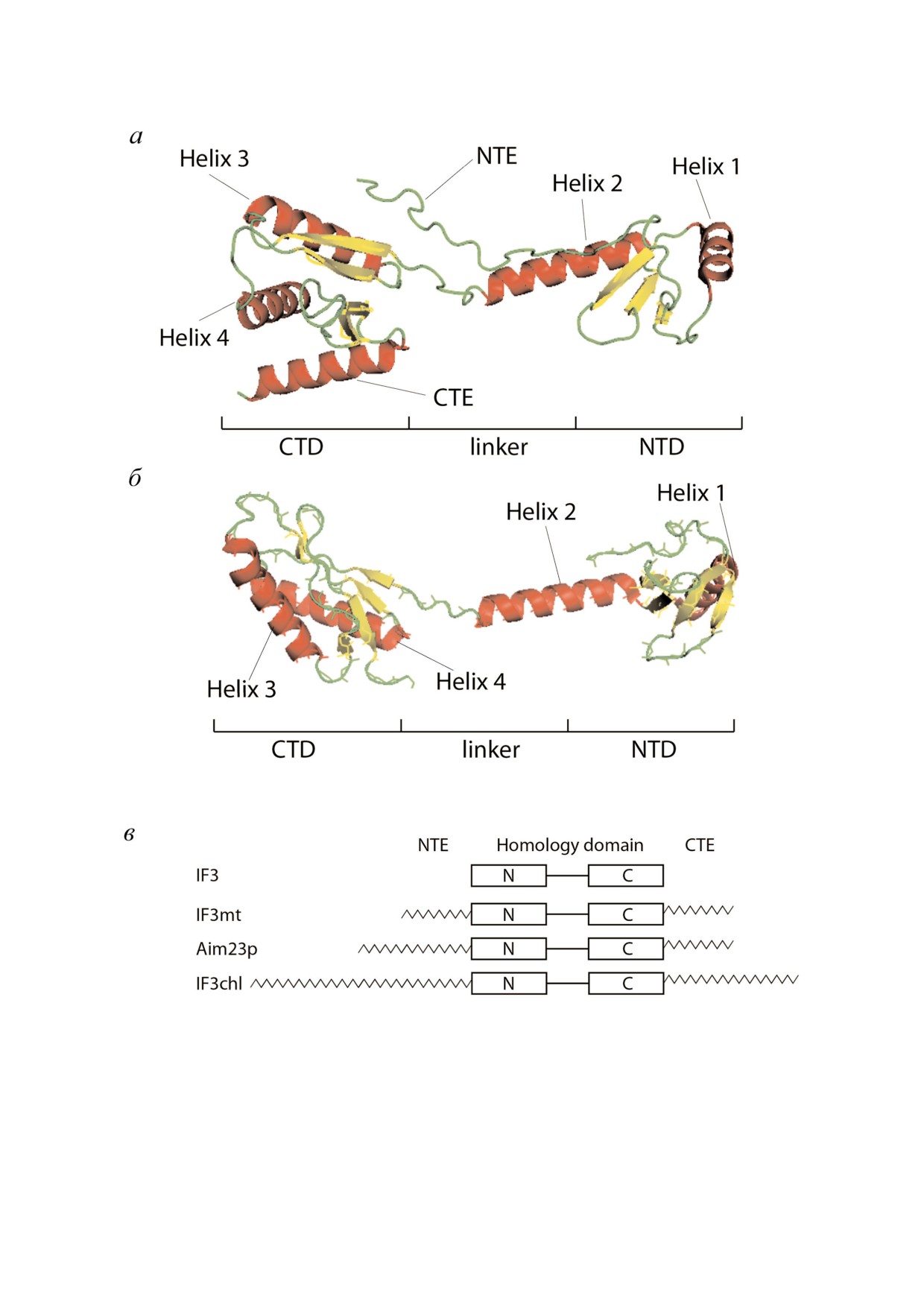
Рис. 2. а - Структура IF3mt H. sapiens (PDB ID: 6NEQ); б - структура IF3 T. thermophilus (PDB ID: 5LMN). α!Спирали по!
казаны красным, β!листы - желтым, петли - зеленым; в -доменная организация IF3 T. thermophilus (IF3), митохондрий
дрожжей S. cerevisiae (Aim23p), митохондрий H. sapiens (IF3mt) и хлоропластов E. gracilis (IF3chl).
journal/biokhsm/
БИОХИМИЯ том 84 вып. 10 2019
IF3mt: СТРУКТУРА, ФУНКЦИИ И ВЗАИМОДЕЙСТВИЯ
1405
IF3 прокариот связывается с 30S субъедини!
те. Таким образом, этот структурный элемент
цей своим C!доменом рядом с P! сайтом и ко!
отвечает за одну из активностей IF3mt, а имен!
дон!антикодоновым дуплексом (рис. 3), а N!до!
но: удаление инициаторной тРНК, связанной в
мен взаимодействует с боковым выступом и
Р!сайте в отсутствие мРНК. Согласно более
инициаторной тРНК [19]. Первые попытки точ!
ранним экспериментам, именно эта активность
но установить место посадки IF3mt на 28S субъ!
теряется, если удалить С!концевое удлинение
единице были сделаны с помощью химической
[18].
сшивки белков с последующим масс!спектро!
метрическим анализом [20]. Эти эксперименты
показали, что IF3mt расположен рядом с 4 бел!
МИТОХОНДРИАЛЬНЫЙ IF3 УЧАСТВУЕТ
ками, имеющими гомологов у бактерий
В РАБОТЕ МЫШЦ МЛЕКОПИТАЮЩИХ
(MRPS5, MRPS9, MRPS10 и MRPS18!2) и 4 дру!
гими белками, специфичными для митохон!
Недавние работы показали, что экспрессия
дрий (MRPS29, MRPS32, MRPS36 и PTCD3).
генов митохондрий млекопитающих тесно свя!
Такой же набор партнеров был обнаружен для
зана с режимом питания и физической актив!
укороченной версии белка без N!домена. Эти
ностью. Одна из них посвящена анализу транс!
наблюдения позволили установить, что IF3mt
ляции мРНК, кодирующих компоненты аппа!
расположен в области сочленения шеи, головы
рата белкового синтеза митохондрий в мыши!
и бокового выступа 28S субъединицы, подобно
ной модели ожирения, спровоцированного вы!
бактериальному гомологу. При этом его контак!
сококалорийной диетой [23]. Ожирение являет!
ты с рибосомой сильно изменились и перестали
ся широко распространенной проблемой во
включать в себя некоторые белки, гомологи ко!
всем мире, особенно в странах с высоким уров!
торых прочно взаимодействовали с IF3.
нем жизни. Оно развивается вследствие упо!
Данные, полученные методом электронной
требления большого количества сладкой и жир!
крио!микроскопии высокого разрешения, поз!
ной пищи и вызывает проблемы с сердцем, поч!
волили точно определить положение IF3mt на
ками и кровеносными сосудами. Исследователи
28S субъединице [21]. В целом, оно оказалось
сравнили содержание нескольких митохондри!
похоже на положение IF3 на 30S субъединице
альных белков (IF2mt, IF3mt, TUFM, TACO1,
прокариот. N!домен IF3mt NTD располагался
цитохром b и ND4) в мышцах физически актив!
близко к борозде, отделяющей боковой выступ
ных мышей и мышей, ведущих малоподвижный
от тела 28S субъединицы (рис. 3), подобно N!до!
образ жизни в двух разных режимах питания:
мену IF3. При этом в бактериальной системе
нормальный и с повышенным содержанием жи!
этот домен не имеет ни одного контакта с рибо!
ров и холестерина. Результаты экспериментов
сомными белками или РНК [22], но N!домен
показали, что количество IF3mt значительно
IF3mt взаимодействовал с одним белком (uS11)
повышалось при физических нагрузках у мы!
и одной спиралью 12S рРНК (h23). Междомен!
шей, питающихся высококалорийной пищей.
ный мостик IF3mt расположен на краю боково!
Похожий эффект наблюдался и при нормаль!
го выступа 28S субъединицы и тянется от спи!
ном режиме питания, но был не так сильно вы!
рали h23 12S рРНК к P!сайту. В отличие от бак!
ражен. Количество IF2mt при физических уп!
териального мостика, служащего лишь соединя!
ражнениях увеличивалось в условиях нормаль!
ющим элементом, мостик IF3mt формирует
ной диеты, однако снижалось, если мышей кор!
множество специфических контактов с нуклео!
мили высококалорийной пищей. Количество
тидами 12S рРНК (h23). C!концевой домен
TUFM не изменялось при физических упражне!
IF3mt расположен на боковом выступе малой
ниях в нормальных условиях, но сильно повы!
субъединицы рядом с P!сайтом (рис. 3) и имеет
шалось в условиях, моделирующих ожирение.
множество взаимодействий с участками 12S
Содержание TACO1, цитохрома b и ND4 возрас!
rRNA, включая спирали h24, h44 и h45. Низкое
тало у физически активных мышей в обоих ре!
разрешение в области N!концевого удлинения
жимах питания.
не позволило точно определить его структуру,
Эти открытия подтверждены в другой рабо!
однако, по имеющимся данным очевидно, что
те, посвященной изучению связи между физи!
оно контактирует со спиралью h24 12S рРНК,
ческими упражнениями и уровнем факторов
увеличивая сродство IF3mt к малой субъедини!
митохондриальной трансляции в мышцах мле!
це. Оказалось, что C!концевое удлинение IF3mt
копитающих [24]. Результаты показали повыше!
совсем не взаимодействует с 28S субъединицей.
ние содержания большинства факторов, участ!
Предположительно, оно проникает в межсубъ!
вующих во всех стадиях процесса: инициации
единичное пространство рибосомы и препят!
(IF2mt и IF3mt), элонгации (mtEFTs, mtEFTu, и
ствует связыванию инициаторной тРНК в P!сай!
mtEFG1), терминации (mtRF1L) и рециклинга
БИОХИМИЯ том 84 вып. 10 2019
1406
ЧИЧЕРИН и др.
Рис. 3. а - Положение IF3mt H. sapiens на 28S субъединице миторибосомы, вид со стороны межсубъединичной поверх!
ности (PDB ID: 6NEQ); б - положение IF3 T. thermophilus на 30S субъединице в такой же ориентации (PDB ID: 5LMN).
Малая субъединица показана серым, IF3 и IF3mt - синим.
journal/biokhsm/
БИОХИМИЯ том 84 вып. 10 2019
IF3mt: СТРУКТУРА, ФУНКЦИИ И ВЗАИМОДЕЙСТВИЯ
1407
(mtRRF1 и mtRRF2) - у активных мышей. Это
ных генах, кодирующих факторы биогенеза и
хорошо коррелировало с экспрессией генов ми!
работы митохондрий, такие как NDUFV2,
тохондриальной ДНК. Кроме того, работа пока!
PGC!1α, HSPA9, LRPPRC, MTIF3, POLG1,
зала метаболическую неоднородность мышц.
TFAM, TFB1M и TFB2M, вызывают предраспо!
Значительные различия наблюдались между
ложенность к болезни [25]. Современная модель
красными и белыми мышцами, последние из
участия митохондрий в развитии БП предпола!
которых имели пониженные экспрессию мито!
гает, что нарушения в работе органелл вызывает
хондриальных генов и содержание факторов
окислительный стресс и повышение количества
митохондриальной трансляции. Мышцы по!
активных форм кислорода, что, в свою очередь,
разному реагировали на физические нагрузки.
приводит к деградации нейронов и создает
Так, масса мышцы soleus у физически активных
предпосылки для болезни.
мышей была выше, чем у малоподвижных, од!
Методами генетики человека была проана!
нако мышцы plantaris в обеих группах весили
лизирована группа из 192 пациентов, в результа!
одинаково. Указанные выше различия в содер!
те чего была обнаружена однонуклеотидная за!
жании митохондриальных факторов были выяв!
мена C798>T в гене IF3MT, показывающая
лены в plantaris, тогда как в soleus их не наблюда!
сильную аллельную ассоциацию с болезнью
лось, за исключением IF2mt, уровень которого
Паркинсона [26]. Эти данные полностью были
повышался и mtRRF2, которого, к удивлению,
подтверждены в другой работе с большим раз!
стало меньше после физических нагрузок. Фи!
мером выборки, состоящей из 2434 пациентов
зиологические и молекулярные механизмы, оп!
[27]. Удивительно, что этот полиморфизм не вы!
ределяющие столь различное поведение мышц
зывает аминокислотной замены в белке
при физических нагрузках, в настоящее время
(Asp266>Asp), что говорит о том, что молекуляр!
неясны. Авторы исследования считают, что при!
ные основы патологии возникают скорее на
чина может заключаться в различном участии
уровне мРНК, чем на уровне белка. Действи!
мышц в данном специфическом упражнении
тельно, моделирование in silico показало, что за!
(бег в колесе). Эти работы показали, что здо!
мена C798>T SNP изменяет структуру мРНК
ровье и работа мышц млекопитающих зависит
IF3MT в области экзона 5, придавая ей бóльшую
от работы их митохондрий и IF3mt играет важ!
стабильность [28]. В той же работе продемон!
ную роль в этих процессах.
стрировано статистически значимое снижение
содержания мРНК IF3MT в клетках с заменой
C798>T. Эти наблюдения не объясняют пол!
МИТОХОНДРИАЛЬНЫЙ IF3 УЧАСТВУЕТ
ностью роль IF3mt в развитии БП, но задают
В РАЗВИТИИ БОЛЕЗНИ ПАРКИНСОНА
направление исследования. Одним из возмож!
ных объяснений является снижение количества
Болезнь Паркинсона (БП) является вторым
IF3mt, что может приводить к окислительному
по распространенности нейродегенеративным
стрессу митохондрий и тем самым создавать
расстройством, клиническими симптомами ко!
предпосылки для развития болезни.
торого являются тремор, гипокинезия, мышеч!
ная ригидность и постуральная неустойчивость.
Оно связано с когнитивными и психиатриче!
ПЕРСПЕКТИВЫ БУДУЩЕГО
скими проблемами, нарушением обоняния, сна
и требует постоянного присмотра за пациентом
Выше мы обсудили множество исследова!
[25]. Несмотря на то, что развитие БП сложно и
ний IF3mt in vitro. Удивительно, что ни одного
включает в себя множество факторов, таких как
детального исследования in vivo до сих пор не
генетика, возраст и факторы окружающей сре!
проведено с этим белком. При этом они могли
ды, известно, что генетическими причинами бо!
бы дать ценную информацию о гене IF3MT и
лезни могут быть гены, кодирующие белки ми!
показать неожиданные результаты, как это слу!
тохондрий [25]. Развитие БП является сложным
чилось с гомологичным белком митохондрий
процессом и результатом взаимодействия инди!
дрожжей S. cerevisiae, Aim23p [29]. В настоящее
видуальных генетических факторов, возраста и
время неясно, является ли ген абсолютно необ!
окружающей среды. Основными молекулярны!
ходимым для жизни культур клеток и целых ор!
ми причинами, вызывающими болезнь, счита!
ганизмов млекопитающих. По данным глобаль!
ются окислительный стресс, нарушение работы
ных скринингов, ген не является необходимым,
митохондрий, воспаления, нарушения в работе
однако в литературе не описано ни одной нока!
системы транспорта и деградации клетки, а так!
утной линии или модели.
же ошибки в сворачивании белков. Неудиви!
Роль IF3mt в болезни Паркинсона практи!
тельно, что мутации и полиморфизмы в ядер!
чески не исследована. Следующим шагом в этом
БИОХИМИЯ том 84 вып. 10 2019
1408
ЧИЧЕРИН и др.
направлении может стать оценка количества
сово поддержана гос. заданием ИБР РАН (0108!
белка в клетках с заменой C798>T методом вес!
2019!004).
терн!блоттинга и анализа митохондриальных
Конфликт интересов. Авторы заявляют, что
параметров, таких как поглощение кислорода,
исследование проводили при отсутствии каких!
активность комплексов дыхательной цепи и ми!
либо коммерческих или финансовых отноше!
тохондриальная трансляция.
ний, которые могли бы привести к потенциаль!
ному конфликту интересов.
Соблюдение этических норм. Настоящая
Финансирование. Работа выполнена при фи!
статья не содержит описания каких!либо иссле!
нансовой поддержке РФФИ (грант
18!74!
дований с использованием людей и животных в
00016). Работа Дашинимаева Э.Б. была финан!
качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Calvo, S.E., and Mootha, V.K. (2010) The mitochondrial
13.
Brown, A., Amunts, A., Bai, X.C., Sugimoto, Y.,
proteome and human disease, Annu. Rev. Genomics Hum.
Edwards, P.C., Murshudov, G., Scheres, S.H., and
Genet., 11, 25-44, doi: 10.1146/annurev!genom!082509!
Ramakrishnan, V. (2014) Structure of the large ribosomal
141720.
subunit from human mitochondria, Science, 346, 718-722,
2.
Anderson, S., Bankier, A.T., Barrell, B.G., de Bruijn, M.H.,
doi: 10.1126/science.1258026.
Coulson, A.R., Drouin, J., Eperon, I.C., Nierlich, D.P.,
14.
Bhargava, K., and Spremulli, L.L. (2005) Role of the N!
Roe, B.A., Sanger, F., Schreier, P.H., Smith, A.J., Staden, R.,
and C!terminal extensions on the activity of mammalian
and Young, I.G. (1981) Sequence and organization of the
mitochondrial translational initiation factor 3, Nucl. Acids
human mitochondrial genome, Nature, 290, 457-465,
Res., 33, 7011-7018, doi: 10.1093/nar/gki1007.
doi: 10.1038/290457a0.
15.
Haque, M.E., Grasso, D., and Spremulli, L.L. (2008) The
3.
Christian, B.E., and Spremulli, L.L. (2012) Mechanism of
interaction of mammalian mitochondrial translational ini!
protein biosynthesis in mammalian mitochondria,
tiation factor 3 with ribosomes: evolution of terminal
Biochim. Biophys. Acta, 1819, 1035-1054, doi: 10.1016/
extensions in IF3mt, Nucl. Acids Res., 36, 589-597, doi: 10.
j.bbagrm.2011.11.009.
1093/nar/gkm1072.
4.
Amunts, A., Brown, A., Toots, J., Scheres, S.H., and
16.
Yu, N.J., and Spremulli, L.L. (1998) Regulation of the
Ramakrishnan, V. (2015) The structure of the human mito!
activity of chloroplast translational initiation factor 3 by
chondrial ribosome, Science, 348, 95-98, doi: 10.1126/sci!
NH2! and COOH!terminal extensions, J. Biol. Chem., 273,
ence.aaa1193.
3871-3877, doi: 10.1074/jbc.273.7.3871.
5.
Montoya, J., Ojala, D., and Attardi, G. (1981) Distinctive
17.
Derbikova, K., Kuzmenko, A., Levitskii, S., Klimontova,
features of the 5'!terminal sequences of the human mito!
M., Chicherin, I., Baleva, M.V., Krasheninnikov, I.A., and
chondrial mRNAs, Nature, 290, 465-470, doi: 10.1038/
Kamenski, P. (2018) Biological and evolutionary signifi!
290465a0.
cance of terminal extensions of mitochondrial translation
6.
Salinas!Giege, T., Giege, R., and Giege, P. (2015) tRNA
initiation factor 3, Int. J. Mol. Sci., 19, 1-15, doi: 10.3390/
biology in mitochondria, Int. J. Mol. Sci., 16, 4518-4559,
ijms19123861.
doi: 10.3390/ijms16034518.
18.
Christian, B.E., and Spremulli, L.L. (2009) Evidence for
7.
Kuzmenko, A., Atkinson, G.C., Levitskii, S., Zenkin, N.,
an active role of IF3mt in the initiation of translation in
Tenson, T., Hauryliuk, V., and Kamenski, P.
(2014)
mammalian mitochondria, Biochemistry, 48, 3269-3278,
Mitochondrial translation initiation machinery: conserva!
doi: 10.1021/bi8023493.
tion and diversification, Biochimie,
100,
132-140,
19.
Hussain, T., Llacer, J.L., Wimberly, B.T., Kieft, J.S., and
doi: 10.1016/j.biochi.2013.07.024.
Ramakrishnan, V. (2016) Large!scale movements of IF3
8.
Biou, V., Shu, F., and Ramakrishnan, V. (1995) X!ray crys!
and tRNA during bacterial translation initiation, Cell, 167,
tallography shows that translational initiation factor IF3
133-144, doi: 10.1016/j.cell.2016.08.074.
consists of two compact alpha/beta domains linked by an
20.
Haque, M.E., Koc, H., Cimen, H., Koc, E.C., and
alpha!helix, EMBO J., 14, 4056-4064.
Spremulli, L.L. (2011) Contacts between mammalian
9.
Petrelli, D., LaTeana, A., Garofalo, C., Spurio, R., Pon, C.L.,
mitochondrial translational initiation factor 3 and riboso!
and Gualerzi, C.O. (2001) Translation initiation factor
mal proteins in the small subunit, Biochim. Biophys. Acta,
IF3: two domains, five functions, one mechanism? EMBO
1814, 1779-1784, doi: 10.1016/j.bbapap.2011.09.013.
J., 20, 4560-4569, doi: 10.1093/emboj/20.16.4560.
21.
Koripella, R.K., Sharma, M.R., Haque, M.E., Risteff, P.,
10.
Ayyub, S.A., Dobriyal, D., and Varshney, U.
(2017)
Spremulli, L.L., and Agrawal, R.K. (2019) Structure of
Contributions of the N! and C!terminal domains of initia!
human mitochondrial translation initiation factor 3 bound
tion factor 3 to its functions in the fidelity of initiation and
to the small ribosomal subunit, Science, 12, 76-86,
antiassociation of the ribosomal subunits, J. Bacteriol.,
doi: 10.1016/j.isci.2018.12.030.
199, 1-12, doi: 10.1128/JB.00051!17.
22.
Lopez!Alonso, J.P., Fabbretti, A., Kaminishi, T., Iturrioz,
11.
Koc, E.C., and Spremulli, L.L. (2002) Identification of
I., Brandi, L., Gil!Carton, D., Gualerzi, C.O., Fucini, P.,
mammalian mitochondrial translational initiation factor 3
and Connell, S.R. (2017) Structure of a 30S pre!initiation
and examination of its role in initiation complex formation
complex stalled by GE81112 reveals structural parallels in
with natural mRNAs, J. Biol. Chem., 277, 35541-35549,
bacterial and eukaryotic protein synthesis initiation path!
doi: 10.1074/jbc.M202498200.
ways, Nucl. Acids Res., 45, 2179-2187, doi: 10.1093/
12.
Haque, M.E., and Spremulli, L.L. (2008) Roles of the N!
nar/gkw1251.
and C!terminal domains of mammalian mitochondrial ini!
23.
Lee, D.E., Brown, J.L., Rosa, M.E., Brown, L.A., Perry,
tiation factor 3 in protein biosynthesis, J. Mol. Biol., 384,
R.A., Washington, T.A., and Greene, N.P.
(2016)
929-940, doi: 10.1016/j.jmb.2008.09.077.
Translational machinery of mitochondrial mRNA is pro!
БИОХИМИЯ том 84 вып. 10 2019
IF3mt: СТРУКТУРА, ФУНКЦИИ И ВЗАИМОДЕЙСТВИЯ
1409
moted by physical activity in Western diet!induced obese
27. Behrouz, B., Vilarino!Guell, C., Heckman, M.G., Soto!
mice, Acta Physiol. (Oxf.), 218, 167-177, doi: 10.1111/
Ortolaza, A.I., Aasly, J.O., Sando, S., Lynch, T., Craig, D.,
apha.12687.
Uitti, R.J., Wszolek, Z.K., Ross, O.A., and Farrer, M.J.
24. Yokokawa, T., Kido, K., Suga, T., Isaka, T., Hayashi, T.,
(2010) Mitochondrial translation initiation factor 3 poly!
and Fujita, S. (2018) Exercise!induced mitochondrial bio!
morphism and Parkinson’s disease, Neurosci. Lett., 486,
genesis coincides with the expression of mitochondrial
228-230, doi: 10.1016/j.neulet.2010.09.059.
translation factors in murine skeletal muscle, Physiol. Rep.,
28. Anvret, A., Ran, C., Westerlund, M., Thelander, A. C.,
6, 1-13, doi: 10.14814/phy2.13893.
Sydow, O., Lind, C., Hakansson, A., Nissbrandt, H.,
25. Gaweda!Walerych, K., and Zekanowski, C. (2013) The
Galter, D., and Belin, A. C. (2010) Possible involvement of
impact of mitochondrial DNA and nuclear genes related to
a mitochondrial translation initiation factor 3 variant caus!
mitochondrial functioning on the risk of Parkinson’s dis!
ing decreased mRNA levels in Parkinson’s disease,
ease, Curr. Genomics, 14, 543-559, doi: 10.2174/1389202!
Parkinsons Dis., 2010, 1-5, doi: 10.4061/2010/491751.
914666131210211033.
29. Kuzmenko, A., Derbikova, K., Salvatori, R., Tankov, S.,
26. Abahuni, N., Gispert, S., Bauer, P., Riess, O., Kruger, R.,
Atkinson, G.C., Tenson, T., Ott, M., Kamenski, P., and
Becker, T., and Auburger, G. (2007) Mitochondrial trans!
Hauryliuk, V.
(2016) Aim!less translation: loss of
lation initiation factor 3 gene polymorphism associated
Saccharomyces cerevisiae mitochondrial translation initia!
with Parkinson’s disease, Neurosci. Lett., 414, 126-129,
tion factor mIF3/Aim23 leads to unbalanced protein syn!
doi: 10.1016/j.neulet.2006.12.053.
thesis, Sci. Rep., 6, 1-9, doi: 10.1038/srep18749.
MITOCHONDRIAL TRANSLATION INITIATION FACTOR 3:
STRUCTURE, FUNCTIONS, INTERACTIONS, AND IMPLICATION
IN HUMAN HEALTH AND DISEASE
Mini review
I. V. Chicherin1,2*, M. V. Baleva1, S. A. Levitskii1,
E. B. Dashinimaev3,4, and I. A. Krasheninnikov1
1 Lomonosov Moscow State University, Faculty of Biology, 119234 Moscow, Russia;
E-mail: i.v.chicherin@gmail.com
2 Institute of Functional Genomics, Lomonosov Moscow State University,
119234 Moscow, Russia
3 Koltzov Institute of Developmental Biology, Russian Academy of Sciences,
119334 Moscow, Russia; E-mail: dashinimaev@gmail.com
4 Pirogov Russian National Research Medical University,
117977 Moscow, Russia
Received March 29, 2019
Revised April 25, 2019
Accepted June 23, 2019
Mitochondria are essential organelles of a eukaryotic cell, providing the respiratory function by means of the electron
transfer chain. Mitochondrial gene expression is organized in bacterial!like manner, however multiple evolutionary
deviations are observed between the two systems. One of them resides in the translation initiation machinery. The cur!
rent review is dedicated to mitochondrial translation initiation factor 3, which plays a key role in protein synthesis in
the organelles. Its involvement in human health and disease is discussed.
Keywords: IF3mt, mitochondria, translation, ribosome, Parkinson’s disease
БИОХИМИЯ том 84 вып. 10 2019