БИОХИМИЯ, 2019, том 84, вып. 10, с. 1450 - 1459
УДК 578.23
ИЗМЕНЕНИЯ РЕЦЕПТОРНОЙ СПЕЦИФИЧНОСТИ ВИРУСОВ
ГРИППА ЧЕЛОВЕКА H3N2 В ХОДЕ ДЛИТЕЛЬНОЙ ЦИРКУЛЯЦИИ*
© 2019 А.Г. Гамбарян1**, А. Балиш2, А.И. Климов2
, А.Б. Тузиков3, А.А. Чинарев3,
Г.В. Пазынина3, Н.В. Бовин3
1 ФГБНУ «Федеральный научный центр исследований и разработки
иммунобиологических препаратов им. М.П. Чумакова РАН»,
108819 Москва, Россия; электронная почта: al.gambaryan@gmail.com
2 Отделение по гриппу, Центры по контролю и профилактике
заболеваний, Атланта, GA 30333, США
3 Институт биоорганической химии им. академиков М.М. Шемякина и Ю.А. Овчинникова РАН,
117997 Москва, Россия
Поступила в редакцию 03.04.2019
После доработки 05.06.2019
Принята к публикации 15.06.2019
Ранее было показано, что аминокислоты 155Thr, 158Glu и 228Ser гемагглютинина имеют решающее значе
ние для распознавания Neu5Gc. В данном исследовании показано, что способность связывать рецептор,
терминированный Neu5Gc, также может быть связана с аминокислотой 145: вирусы 1972-1999 гг. с Lys145
связывали Neu5Gc, тогда как вирусы с Asn145 не связывали. Спорадическое появление и исчезновение спо
собности связывать олигосахариды Neu5Gc, а также отсутствие остатков Neu5Gc в составе гликоконъюга
тов человека указывают на неадаптивную природу этого свойства. Ранее показано, что вирусы гриппа A
(H3N2) 1968-1989 гг., в отличие от вирусов H1N1, не различают Neu5Aca2 6Galβ1 4Glc (6'SL) и Neu5Aca2
6Galβ1 4GlcNAc (6'SLN). Вирусы H3N2, изолированные после 1993 г., наподобие вирусов H1N1, приобре
ли способность различать 6'SL и 6'SLN. Мы определили, что преимущественное сродство к 6'SLN постепен
но увеличивалась с 1992 по 2003 г. После 2003 г. вирусы утратили способность связывать ряд сиалозидов,
включая 6'SL, которые являлись хорошими рецепторами для более ранних вирусов H3N2, и только 6'SLN
сохранил высокое сродство к более поздним человеческим вирусам H3N2. Это свойство коррелирует с при
обретением новых сайтов гликозилирования в положениях 122, 133 и 144, а также с заменами Glu190Asp и
Gly225Asp в гемагглютинине. Эти же замены ответственны за рецепторный фенотип вирусов H1N1 челове
ка. Конвергентная эволюция рецепторной специфичности подтипов H1N1 и H3N2 указывает на то, что
6'SLN является оптимальным природным человеческим рецептором для вирусов гриппа.
КЛЮЧЕВЫЕ СЛОВА: вирус гриппа, гемагглютинин, сиалосахариды, рецепторная специфичность, H3N2.
DOI: 10.1134/S0320972519100075
Вирусы гриппа являются РНК вирусами с
ных птиц иногда передаются другим диким и
негативной цепью и сегментированными гено
домашним животным. В редких случаях они
мами. Они содержат восемь сегментов РНК, ко
приспосабливаются к новым видам и образуют
торые кодируют от 10 до 11 белков. Вирусная
устойчивые специфические для хозяина линии.
частица содержит два основных гликопротеина
Считается, что все известные линии вирусов
оболочки: гемагглютинин (HA) и нейраминида
гриппа домашней птицы и млекопитающих,
зу (NA). Естественным резервуаром вирусов
включая людей, происходят от вирусов диких
гриппа являются дикие водные птицы, в кото
водных птиц [1].
рых циркулируют вирусы с 16 антигенными
Сиаловые кислоты являются минимально
подтипами HA и 9 подтипами NA. Вирусы вод необходимыми детерминантами клеточных ре
Принятые сокращения: PBS - фосфатно солевой буфер; BSA - бычий сывороточный альбумин; КЭ - куриные яй
ца с эмбрионом; HA - гемагглютинин; NA - нейраминидаза; РСУ - рецептор связывающий участок; Sia - сиаловая кис
лота (либо Neu5Ac, либо Neu5Gc); Glyc PAA - гликозилированный полиакриламид; Glyc PAA biot - биотинилирован
ный гликозилированный полиакриламид.
biokhimiya, в рубрике «Papers in Press», BM19 109, 26.08.2019.
** Адресат для корреспонденции.
1450
РЕЦЕПТОРНАЯ СПЕЦИФИЧНОСТЬ ВИРУСОВ ГРИППА ЧЕЛОВЕКА Н3N2
1451
цепторов для вирусов гриппа [2, 3]. Сиалилоли
па YDS, 6 STF и специфичным для свиней ре
госахаридные фрагменты гликопротеинов и
цептором Neu5Gc Tn [16].
гликолипидов на клеточной поверхности демон
Позже появилась замена Gly225Asp. Инте
стрируют значительное структурное разнооб
ресно отметить, что Asp190 и Asp225 консерва
разие, и структура углеводной цепочки может
тивны у вирусов H1N1 человека и ответственны
влиять на способность сиаловой кислоты слу
за узнавание рецептора Neu5Acα2-6Galβ1-
жить рецепторной детерминантой вируса грип
4GlcNAc. Было показано, что изоляты H3N2
па. Одной из основных характеристик рецеп
1998 и 1999 годов, в отличие от ранних изолятов,
торной специфичности вирусов гриппа являет
связывали 6'SLN с бóльшим сродством, чем 6'SL
ся способность различать дисахаридные мотивы
[17].
Neu5Acα2-3Gal и Neu5Acα2-6Gal [4, 5]. Виру
Вследствие реконструкции РСУ, адаптация
сы гриппа человека A и B с высоким сродством
вирусов к куриным эмбрионам стала проходить
связывают сахариды, терминированные
по новым схемам аминокислотного замещения
Neu5Acα2-6Gal, и не связывают Neu5Acα2-3Gal
[18]. Масштабный мутационный анализ RBS
терминированные рецепторы [6]. Напротив, ви
вирусов A/WSN/33 (H1N1) и A/Hong Kong/
русы гриппа А птиц, независимо от их субтипа,
1/1968 (H3N2) выявил множество компетент
не связывают Neu5Acα2-6Gal. Они имеют бо
ных по репликации мутантов, еще не обнару
лее высокое сродство к олигосахаридам, терми
женных в природе [19].
нированным Neu5Acα2-3Gal, чем к свободной
Самые ранние вирусы человека H3N2 преи
нейраминовой кислоте (Neu5Ac), что свиде
мущественно связывали короткие разветвлен
тельствует о специфических взаимодействиях
ные сиалилированные гликаны, в то время как
между рецептор связывающим участком (РСУ)
более поздние вирусы связывали длинные поли
HA и галактозой, связанной с сиаловой кисло
лактозаминовые цепи, терминированные сиа
той 2-3 связью. Сравнение аминокислотных
ловой кислотой, и не связывали короткие сиа
последовательностей HA разных субтипов выя
лилированные гликаны [20]. Ранние человече
вило, что остатки в положениях 138, 190, 194,
ские вирусы H3N2 связывали широкий спектр
225, 226 и 228 консервативны в РСУ вирусов
аналогов человеческого рецептора, тогда как
гриппа субтипов H1-H12, выделенных из диких
изоляты 2006-2011 гг. преимущественно связы
уток. В H3 HA ранних вирусов H3N2 человека
вали удлиненные, разветвленные мультивалент
были заменены аминокислоты 226 и 228, а в H1
ные рецепторы. Эти рецепторы с удлиненными
HA вирусов H1N1 человека были заменены ами
ветвями имеют потенциал для увеличения срод
нокислоты 138, 190, 194 и 225 [7, 8].
ства за счет одновременного связывания с двумя
Человеческие штаммы H1N1 связывают
субъединицами одного тримера гемагглютини
Neu5Acα2-6Galβ1-4GlcNAc трисахарид (6'SLN)
на [21, 22].
с более высоким сродством, чем Neu5Acα2-
В данной работе мы исследовали рецептор
6Galβ1-4Glc (6'SL), в то время как ранние чело
связывающие свойства широкого набора виру
веческие вирусы H3N2 не различали эти две
сов гриппа H3N2 человека, выделенных с 1968
структуры [5, 8, 9, 10]. После 1992 г. вирусы че
по 2003 г.
ловеческого гриппа H3N2 приобрели новые
свойства; а именно, они утратили способность
расти в куриных эмбрионах (КЭ) и агглютини
МЕТОДЫ ИССЛЕДОВАНИЯ
ровать эритроциты кур [11, 12, 13].
В течение 1993-2003 гг. в HA вирусов H3N2
Реактивы. 96 луночные планшеты фирмы
человека появились несколько аминокислотных
«Costar» (США), конъюгат стрептавидина с пе
замен в положениях, которые ранее были кон
роксидазой хрена и о фенилендиамин фирмы
сервативными и ответственными за связывание
«Boehringer Mannheim» (Германия). α Бензил
с рецептором. Так, была обнаружена замена
сиалозид (SBn) и 6'SLN (9 NAc) синтезированы
Glu190Asp [12]. Ile, а затем Val стали преоблада
как описано в [23, 24]. Синтез немеченых гли
ющим остатком в положении 226 H3 HA вместо
козилированных полиакриламидов (Glyc PAA)
Leu, типичного для HA более ранних изолятов
(30 кДа) и биотинилированных полимеров,
H3N2 человека [13-15]. Возникли новые потен
Glyc PAA biot (1000 кДа) были описаны в рабо
циальные сайты гликозилирования в 122, 133 и
тах [25, 26]. Структуры аналогов рецепторов и
144 положениях. Отбор антигенно отличных ва
их сокращенные наименования приведены в
риантов H3N2 может способствовать измене
табл. 1.
нию рецепторной специфичности. Замены ами
Вирусы были получены из коллекции Центра
нокислот 156, 158, 189 и 193 влияют на связыва
по контролю за заболеваниями (США). Вирусы
ние с минорными гликанами человеческого ти
культивировали 48 ч в клетках MDCK в среде
БИОХИМИЯ том 84 вып. 10 2019
6*
1452
ГАМБАРЯН и др.
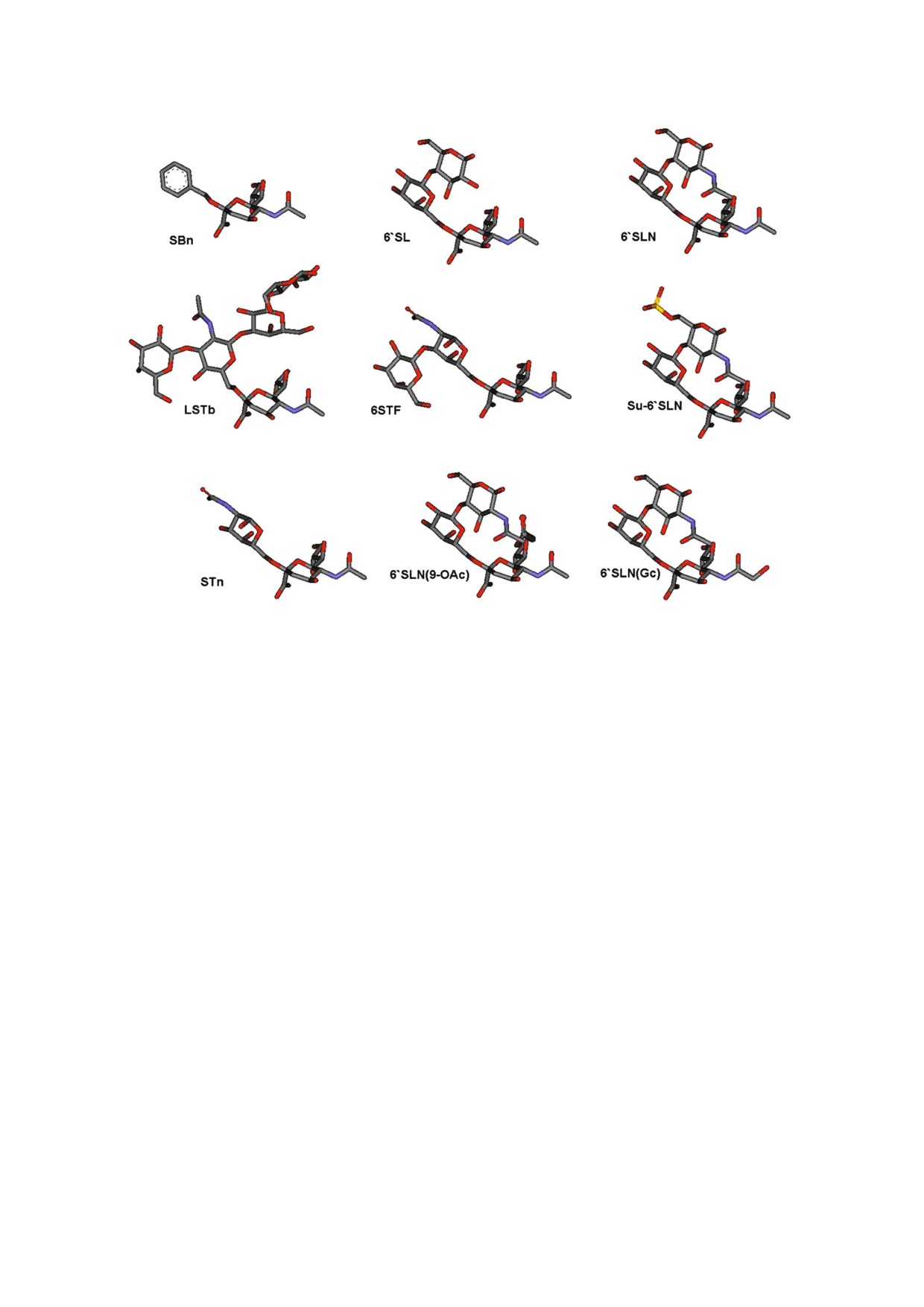
Таблица 1. Структура олигосахаридов, пришитых к полиакриламиду
Биотинилированные полимеры
Обозначения
Neu5Acα2 3Galβ1 4Glcβ PAA biot
3'SL
Neu5Acα2 6Galβ1 4Glcβ PAA biot
6'SL
Neu5Acα2 6Galβ1 4GlcNAcβ PAA biot
6'SLN
Neu5Gcα2 6Galβ1 4GlcNAcβ PAA biot
6`SLN(Gc)
Немеченые полимеры
Neu5Acα OCH2C6H4 PAA
SBn
Neu5Acα2 6Galβ1 4Glcβ PAA
6'SL
Neu5Acα2 6Galβ1 4GlcNAcβ PAA
6'SLN
(Neu5Acα2 62 6G* GN M)2 3,6 M GN GNβ PAA
YDS
Galβ1 3(Neu5Acα2 6)GlcNAcβ1 3Galβ1 4Glc PAA
LSTb
Neu5Acα2 6GalNAcα PAA
STn
Galβ1 3(Neu5Acα2 6)GalNAcα PAA
6STF
Neu5Acα2 6Galβ1 4 (6 O Su)GlcNAcβ PAA
Su 6`SLN
9 N Ac Neu5Acα2 6Galβ1 4GlcNAcβ PAA
6'SLN(9 NAc)
MEM при 37 °С и инактивировали 0,1% ным
бирования связывания фетуина как описано ра
глютаровым альдегидом. Культуральные жид
нее [27]. Анализ основан на конкуренции между
кости осветляли низкоскоростным центрифуги
исследуемым аналогом рецептора и стандарт
рованием, после чего вирусы собирали путем
ным препаратом меченного пероксидазой фету
высокоскоростного центрифугирования, ре
ина за центры связывания на иммобилизован
суспендировали в 0,1 М NaCl, 0,02 М Тris буфе
ном вирусе. Инкубацию проводили в течение 1 ч
ре (рН 7,2), который содержал 50% ный глице
при 4 °С в рабочем буфере. Данные были выра
рин. Хранили при -20 °С.
жены в терминах констант аффинности (Kaff),
Анализ прямого связывания с Glyc PAA biot
формально эквивалентных константам диссо
(1000 кДа) проводили как описано ранее [8].
циации комплекса вирус/рецепторный аналог.
Планшеты покрывали очищенным вирусом
Константы рассчитывали по концентрациям ос
гриппа с титром
4-8 ед гемагглютинации
татков сиаловой кислоты в растворе.
(100 мкл/лунку) при 4 °С в течение 2 ч с после
Статистика. Расчет констант диссоциации и
дующим промыванием фосфатно солевым бу
величины стандартного отклонения определяли
фером (PBS). После этого добавляли в лунки по
с помощью программы «Excel».
100 мкл блокирующего раствора (PBS, с добав
лением 0,1% ного BSA), инкубировали 1 ч, а за
тем промывали холодным PBS, содержащим
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
0,05% ный Tween 20 (отмывочный раствор).
Glyc PAA biot титровали с двукратными разве
Изучали рецептор связывающие свойства
дениями на рабочем буфере (PBS с 0,01% ным
вирусов гриппа человека H3N2, выделенных и
Tween 20, 0,1% ным BSA и 1 мкМ ингибитора
пассированных в клетках MDCK или куриных
нейраминидазы, оселтамивир карбоксилата),
эмбрионах. Для более адекватного сравнения
добавляли в лунки по 50 мкл и инкубировали
адаптированные к КЭ вирусы в последнем пас
планшеты в течение 2 ч при 4 °С. Начальная
саже выращивали в клетках MDCK.
концентрация Glyc PAA biot составляла 0,5 мкМ
В первой серии экспериментов были ис
по сиаловой кислоте. Планшеты промывали хо
пользованы пять меченых аналогов рецепторов:
лодным отмывочным раствором и инкубирова
меченый пероксидазой фетуин и четыре био
ли со стрептавидин пероксидазой в рабочем бу
тин меченых зонда, Glyc PAA biot (1000 кДа),
фере при 4 °С в течение 1 ч. После промывки ак
(остаток Glyc = 3'SL, 6'SL, 6'SLN и 6'SLN (Gc))
тивность пероксидазы в лунках анализировали с
в тесте прямого связывания (формулы и молеку
использованием субстратного раствора о фени
лярные модели остатков приведены в табл. 1 и
лендиамина.
на рис. 1). Результаты анализа представлены в
Конкурентное связывание. Сродство вируса к
табл. 2. Для удобства чтения данные представле
Glyc PAA (30 кДа) определяли в реакции инги
ны в качественной форме: максимальное связы
БИОХИМИЯ том 84 вып. 10 2019
РЕЦЕПТОРНАЯ СПЕЦИФИЧНОСТЬ ВИРУСОВ ГРИППА ЧЕЛОВЕКА Н3N2
1453
Рис. 1. Структура тестированных сиалосахаридов.
biokhsm/
вание выражается тремя звездочками, в десять
2003 и A/New York/72/2003, утратившие SG144
раз слабее связывание - двумя звездочками,
вторично, частично связывали фетуин. Потеря
низкое связывание - одной звездочкой, а отсут
SG126 восстановливала способность связывать
ствие связывания - знаком минус.
фетуин на следовых уровнях. Следовательно,
Распознавание связи между сиаловой кисло>
изобилие новых сайтов гликозилирования (122,
той и галактозой (α2-3 против α2-6). В соответ
133 и 144) имеет решающее значение для неспо
ствии с ранее полученными данными [10, 17] ни
собности связывать фетуин.
один из вирусов, изолированных в культуре кле
Распознавание типа сиаловой кислоты (N>аце>
ток MDCK не связывался с 3'SL, в то время как
тилнейраминовая или N>гликолилнейраминовая).
все адаптированные к КЭ вирусы связывали его
Способность тестируемых H3N2 вирусов рас
несмотря на то, что они были выращены MDCK
познавать N гликолилнейраминовую кислоту
в последнем пассаже (табл. 2).
очень нерегулярна. Как ранние, так и поздние
Связывание вируса с фетуином. Фетуин явля
вирусы с Trp155, не связывают 6'SLN(Gc). Вирус
ется гликопротеином, который содержит в ос
A/Port Chalmers/73 связывал N ацетилнейрами
новном би и древовидные N гликаны, оканчи
новую и N гликолилнейраминовую кислоты с
вающиеся мотивами Siaα2-3Galβ1-4GlcNAc и
одинаковым сродством, что хорошо согласуется
Siaα2-6Galβ1-4GlcNAc [28]. Все вирусы, выде
с работами Anders et al. и Suzuki, в которых по
ленные до 1999 г., и все адаптированные к КЭ
казано, что вирусы H3N2 с Tyr155 распознают
вирусы связывают фетуин с высоким сродством.
Neu5Gcα2-6Gal [29, 30]. Хорошо связывают
Большинство вирусов после 1999 г. утратили
6'SLN(Gc) также все вирусы 1990 х гг. с сочета
способность связывать фетуин; это свойство
нием Lys145 и His155. Штаммы с заменой
коррелирует с появлением новых сайтов глико
Lys145Asn 6'SLN(Gc) не связывают. Вирусы, вы
зилирования (SG) в 122, 133 и 144 м положени
деленные после 2002 г., не связывают Neu5Gc
ях. Вирусы Texas/41/99 и Alaska/37/99, в которых
рецептор даже при наличии Lys145 и His155.
отсутствовал SG122, и вирусы A/New York/68/
Возможно, это связано с дальнейшей пере
БИОХИМИЯ том 84 вып. 10 2019
1454
ГАМБАРЯН и др.
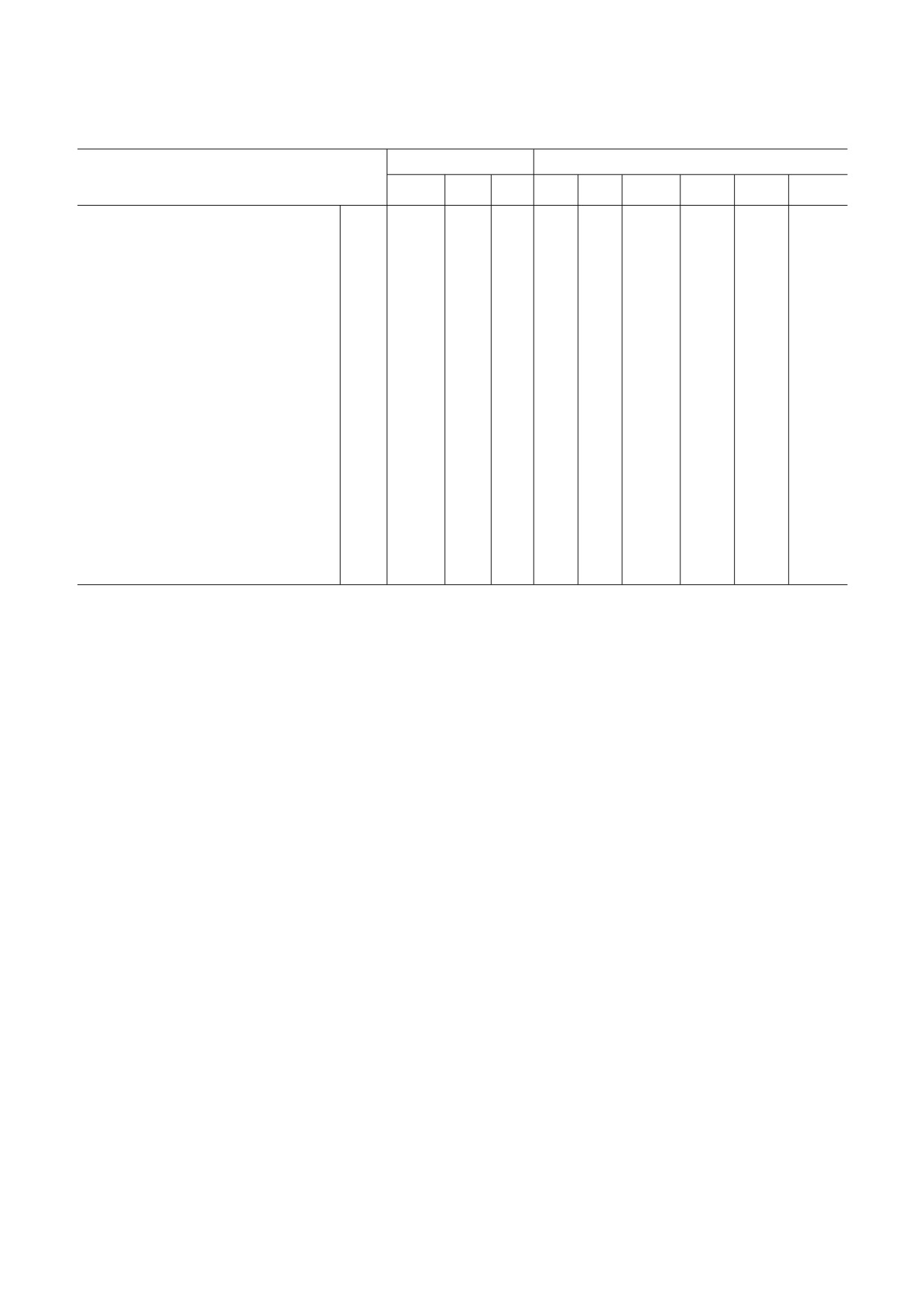
Таблица 2. Связывание вирусов с сиалогликоконьюгатами
Позиция в HA
Сиалогликоконьюгаты
Вирусы
Фет
122
133
144
145
155
190
225
3'SL
6'SL
6'SLN
6'SLN (Gc)
Aichi/2/68
E
Ser
Thr
Glu
Gly
+++
+
+++
+++
-
USSR/039/68
E
Ser
Thr
Glu
Gly
+++
+
+++
+++
-
Canada/228/68
E
Ser
Thr
Glu
Gly
+++
+
+++
+++
-
Taiwan/1/69
E
Ser
Thr
Glu
Gly
+++
+
+++
+++
-
Canada/2/70
E
Ser
Thr
Glu
Gly
+++
+
+++
+++
-
Port Chalmers/1/73
E
Ser
Tyr
Glu
Gly
+++
+
+++
+++
+++
Seoul/23/91
E
Lys
His
Glu
Gly
+++
++
+++
+++
++
Seoul/46/91
E
Lys
His
Glu
Gly
+++
+
+++
+++
++
Kasauli/149/92
E
Lys
His
Glu
Gly
+++
+
+++
+++
++
Umea/01/92
M
Lys
His
Glu
Gly
+++
-
+++
+++
++
Umea/02/92
M
Lys
His
Glu
Gly
+++
-
+++
+++
++
Indonesia/3946/92
M
Lys
His
Glu
Gly
+++
-
+++
+++
++
Qingdao/53/92
E
Asn
His
Glu
Gly
+++
++
++
++
-
Russia/125/93
M
Lys
His
Glu
Gly
+++
-
+++
+++
++
Stockholm/1/93
M
Asn
His
Asp
Gly
+++
-
+++
+++
nt
Georgia/3/93
M
Asn
His
Asp
Gly
+++
-
+++
+++
nt
New York/3/93
M
Asn
His
Asp
Gly
+++
-
+++
+++
nt
Wyoming/1/93
M
Asn
His
Asp
Gly
+++
-
+++
+++
-
Washington/26/93
M
Asn
His
Asp
Gly
+++
-
+++
+++
-
Hong Kong/1/94
M
Lys
His
Asp
Gly
+++
-
+++
+++
++
Hong Kong/55/94
M
Lys
His
Asp
Gly
+++
-
+++
+++
++
Wuhan/359/95
E
Lys
His
Asp
Gly
+++
+
+++
+++
+
Spain/351/95
M
Lys
His
Asp
Gly
+++
-
+++
+++
++
Changwon/9/95
M
Asn
His
Asp
Gly
+++
-
+++
+++
-
Canada/17/96
M
Asn
His
Asp
Gly
+++
-
+++
+++
-
Texas/41/99
M
SG
SG
Lys
His
Asp
Gly
++
-
++
+++
nt
Alaska/37/99
M
SG
SG
Lys
His
Asp
Gly
++
-
++
+++
+
Egypt/8811/99
M
SG
SG
Lys
His
Asp
Gly
+
-
+
+++
nt
California/32/99
E
SG
SG
Lys
His
Asp
Gly
+++
++
++
+++
+
Fujian/411/2002
M
SG
SG
SG
Lys
Thr
Asp
Asp
-
-
-
+++
nt
England/01/21/2003
M
SG
SG
SG
Lys
Thr
Asp
Asp
-
-
-
+++
nt
Wisconsin/32/2003
M
SG
SG
SG
Gln
Thr
Asp
Asp
-
-
-
+++
nt
Hong Kong/797/2003
M
SG
SG
SG
Lys
Thr
Asp
Asp
-
-
-
+++
nt
Nebraska/6/2003
M
SG
SG
SG
Lys
Thr
Asp
Asp
-
-
-
+++
-
Texas/33/2003
M
SG
SG
SG
Lys
Thr
Asp
Asp
-
-
-
+++
-
Texas/39/2003
M
SG
SG
SG
Lys
Thr
Asp
Asp
-
-
-
+++
nt
Texas/40/2003
M
SG
SG
SG
Lys
Thr
Asp
Asp
-
-
-
+++
nt
Texas/42/2003
M
SG
SG
SG
Lys
Thr
Asp
Asp
-
-
-
+++
nt
Wyoming/3e/2003
E
SG
SG
SG
Lys
Thr
Asp
Asp
+++
+
+
+++
nt
Chile/8266e/2003
E
SG
SG
Lys
Thr
Asp
Asp
+++
+
++
+++
-
New York/16/2003
M
SG
SG
SG
Lys
Thr
Asp
Asp
-
-
-
+++
nt
New York/68/2003
M
SG
SG
Lys
His
Asp
Asp
++
-
+
+++
-
New York/72/2003
M
SG
SG
Lys
His
Asp
Gly
++
-
++
+++
-
New York/74/2003
M
SG
SG
SG
Lys
Thr
Asp
Asp
–
-
–
+++
nt
стройкой РСУ, включающей замены аминокис
вые признаки увеличения сродства к 6'SLN по
лот 190, 225 и 226.
сравнению с 6'SL наблюдались у вирусов 1993 г.,
Распознавание 6'SL vs 6'SLN. Способность
в которых Glu заменил Asp в положении 190
различать 6'SLN и 6'SL изменялась у вирусов че
(табл. 3). Эта тенденция усилилась после появ
ловека H3N2 с 1968 по 2003 гг. Вирусы, выделен
ления новых сайтов гликозилирования (табл. 2).
ные с 1968 по 1992 г., связывают эти соединения
Вирусы с SG144 и с заменой Gly225/Asp утрати
с одинаковым сродством. У вирусов H3N2, вы
ли способность связывать 6'SL, тем не менее
деленных после 1993 г., сродство 6'SLN выше,
сродство к 6'SLN оставалась высоким.
чем 6'SL. После 2002 г. вирусы, как правило,
Рецепторная специфичность вирусов к Sia(2-6)>
полностью теряли способность связывать 6'SL.
концевым рецепторам с различными сахаридными
A/New York/68/2003 and A/New York/72/2003 без
корами. Рецептор связывающий фенотип виру
SG144 являются исключениями из этого прави
сов H3N2 человека тестировали с использова
ла; они связывают 6'SL на низком уровне. Пер
нием Neu5Acα2-6Gal терминированных ре
БИОХИМИЯ том 84 вып. 10 2019
РЕЦЕПТОРНАЯ СПЕЦИФИЧНОСТЬ ВИРУСОВ ГРИППА ЧЕЛОВЕКА Н3N2
1455
Таблица 3. Сродство (Kaff, μM) вирусов к сиалогликоконьюгатам*
Позиция в НА
Сиалогликоконьюгаты
Вирусы
190
225
226
SBn
6'SL
6'SLN
STn
LSTb
YDS
Aichi/2/68
E
Glu
Gly
Leu
1
1
1
50
50
2
USSR/039/68
E
Glu
Gly
Leu
1
1
1
50
50
2
Canada/228/68
E
Glu
Gly
Leu
1
1
1
50
50
2
Taiwan/1/69
E
Glu
Gly
Leu
1
1
1
50
50
2
Canada/2/70
E
Glu
Gly
Leu
1
1
1
50
50
2
Port Chalmers/1/73
E
Glu
Gly
Leu
4
2
2
100
100
2
Seoul/23/91
E
Glu
Gly
Leu
1
1
1
30
30
1
Seoul/46/91
E
Glu
Gly
Leu
2
2
2
30
100
10
Kasauli/149/92
E
Glu
Gly
Leu
1
1
1
30
30
1
Umea/01/92
M
Glu
Gly
Leu
0,3
0,3
0,3
50
50
1
Umea/02/92
M
Glu
Gly
Leu
0,3
0,3
0,3
50
50
1
Indonesia/3946/92
M
Glu
Gly
Leu
0,3
0,3
0,3
50
50
1
Qingdao/53/92
E
Glu
Gly
Leu
1
1
1
3
3
3
Stockholm/1/93
M
Asp
Gly
Leu
0,3
0,3
0,2
1
2
2
Wyoming/1/93
M
Asp
Gly
Leu
0,2
0,2
0,1
0.2
0,2
0,2
Hong Kong/55/94
M
Asp
Gly
Leu
0,2
0,5
0,2
1
0,5
1
Spain/351/95
M
Asp
Gly
Leu
0,2
0,2
0,1
0,2
0,2
0,2
Wuhan/359/95
E
Asp
Gly
Ile
1
1
0,4
10
10
5
Texas/41/99
M
Asp
Gly
Val
1
1
0,1
50
50
2
Alaska/37/99
M
Asp
Gly
Val
1
2
0,1
100
100
3
New York/72/2003
M
Asp
Gly
Val
3
3
0,1
100
100
2
Wyoming/3/2003
M
Asp
Asp
Ile
>50
>50
1
nt
nt
nt
Texas/33/2003
M
Asp
Asp
Val
>50
>50
0,5
nt
nt
nt
New York/68/2003
M
Asp
Asp
Ile
>50
50
0,5
40
40
5
* Значения Kaff формально эквивалентны константам диссоциации комплексов вирусов с рецепторными аналогами. Чем
выше значения Kaff, тем ниже сродство. Приводятся данные, усредненные из трех экспериментов. Стандартные ошибки
не превышали 60% средних значений.
цепторов с различным строением предлежащей
нием фрагмента Galb с фрагментом аминокис
цепочки (табл. 1, рис. 1). α Бензилсиалозид
лотной цепи 222-226 в RBS.
(SBn) и 6'SLN (9 NAc) синтезированы; осталь
Все протестированные вирусы показали вы
ные сиалосахариды встречаются в природных
сокое сродство к 6'SLN. Сродство к 6'SLN со
гликопротеинах или гликолипидах и были вы
держащему двухантенному олигосахариду YDS
делены из биологического сырья. Для конкурен
было ниже, чем к 6'SLN. Разница увеличивается
тного анализа были использованы немеченые
с годом изоляции и может быть объяснена сте
сиалогликополимеры с низкой мол. массой
рическим перекрыванием второй антенны YDS
(30 кДа) [25]. В табл. 3 приводятся значения Kaff,
с углеводными фрагментами HA вблизи RBS.
формально эквивалентные константам диссо
Результаты конкурентного анализа совпадают с
циации рецепторных аналогов к вирусам. Все
результатами анализа прямого связывания.
вирусы, связывались с 6'SLN и 6 O Su 6'SLN с
Вирусы, изолированные до 1999 г., имели
одинаковым сродством, и ни один из протести
почти равное сродство к 6'SL и 6'SLN, тогда как
рованных вирусов не связывал с 6'SLN (9 NAc)
после 1999 г. сродство вирусов к 6'SL понижено,
и 6 STF. Поэтому данные для 6 O Su 6'SLN,
вплоть до полного отсутствия связывания.
6'SLN (9 NAc) и 6 STF в таблице не приводятся.
Сродство к STn и LSTb было одинаковым для
Равное сродство к 6'SLN и его сульфатиро
всех протестированных вирусов. Оно был очень
ванному аналогу 6 O Su 6'SLN неудивительно,
низким для вирусов с 1968 по 1992 г. (вплоть до
поскольку сульфогруппа, присоединенная к O 6
вируса Umea/02/92). Вирусы Qingdao/53/92,
N ацетилглюкозамина, направлена из рецептор
Stockholm/1/93, Wyoming/1/93, Spain/351/95
связывающего участка в воду. Также неудиви
связывают STn и LSTb почти так же хорошо, как
тельно отсутствие связывания с 6'SLN (9 NAc),
и
6'SLN. Все эти вирусы имеют замену
поскольку 9 NAc группа стерически не поме
Glu 190Asp. У вируса Wuhan/359/95 (с заменой
щается между C 9 Neu5Ac и поверхностью ре
Leu 226Ile) сродство к этим олигосахаридам
цептор связывающего кармана в области 183,
резко понижено. Более поздние вирусы, с Ile
190 и 228 аминокислот [3]. Отсутствие связыва
или Val226, а также SG133 и SG144, практически
ния с 6 STF может быть объяснено перекрыва
не связывали STn и LSTb.
БИОХИМИЯ том 84 вып. 10 2019
1456
ГАМБАРЯН и др.
Ранее было показано, что бензилсиалозид
сиаловой кислоты не могла дать вирусу избира
(табл. 1) хорошо ингибирует связывание виру
тельное преимущество. Спорадическое появле
сов H3N2 человека с природными рецепторами
ние этой способности, по видимому, является
[31, 32]. Действительно, для всех протестиро
случайным результатом аминокислотных замен
ванных вирусов, кроме трех (три последних
в гипервариабельных областях антигенных сай
пункта, табл. 3), сродство к SBn равно сродству
тов.
к 6'SLN. В то же время три вируса с заменой
Gly225Asp в HA, а именно Wyoming/3/2003,
Ранние вирусы H3N2 человека не были
Texas/33/2003 и NewYork/68/2003 утратили
адаптированы к определенному сиалилолигоса
сродство к SBn и 6'SL. Вероятно, объемный гид
хариду в респираторном эпителии человека.
рофильный Asp полностью нарушает гидрофоб
Первоначальные замены в HA: Gln226Leu и
ное взаимодействие бензильного кольца с
Gly228Ser обеспечивают взаимодействия с ме
Val226.
тиленовой группой галактозы, смежной с 2 6Sia.
Миметики рецепторов общей структуры
Neu5Acα OCH2 R, содержащие гидрофобный
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
агликон, такие как бензилсиалозид и даже α ме
тилсиалозид, имеют хорошее сродство к ранним
В данной работе была рассмотрена эволю
вирусам H3N2 [31]. Если асиаловая часть анало
ция рецепторной специфичности вирусов H3N2
га углеводного рецептора стерически не препят
человека 1968-2003 гг. в контексте замен амино
ствовала связыванию, различные соединения с
кислот в РСУ и строения рецепторных сиалоса
2-6 связанной сиаловой кислотой обладали
харидных цепочек. Две замены Gln226Leu и
практически равным сродством к вирусам [9,
Gly228Ser отличают HA ранних вирусов челове
31]. По мере появления в человеческой популя
ка от консенсусной последовательности вирусов
ции иммунной прослойки к H3N2 вирусам, у
гриппа птиц. Эти замены повысили сродство
вирусов возникли новые сайты гликозилирова
вируса к Neu5Acα2 6Gal и снизили сродство к
ния в HA в положениях 126 и 246, что снижало
Neu5Acα2-3Gal, но предпочтение первого ва
нейтрализующее действие антител. Но при этом
рианта не было абсолютным - сродство к неза
некоторые природные рецепторы могли стано
якоренному 3'SL было всего в 5 раз ниже, чем к
вится недоступными для вируса. Так, способ
6'SL [31, 32]. Комплекс концевого рецептора
ность связываться с ганглиозидами GM3 и 3 SPG
Neu5Acα2 3Gal с HA вируса Aichi/2/68 был ус
снижалась с приобретением сайтов гликозили
пешно закристаллизован и получена полная
рования в положении 126 и исчезала с приобре
трехмерная структура этого комплекса [33].
тением 246SG [7].
Ранние вирусы человека H3N2 связывались с
Когда аминокислота Glu190, консерватив
ганглиозидами, терминированными Neu5Acα2-
ная для вирусов гриппа птиц, была заменена на
3Gal; эта способность впоследствии уменьши
Asp, вирусам стали доступны по меньшей мере
лась и была полностью утеряна [7].
два новых природных сиалосахара (STn и LSTb).
По данным Anders и Suzuki, способность
Вирусы с Asp190 и Leu226 связывают их с высо
распознавать форму Neu5Gc нейраминовой
ким сродством. После замены Leu226Ile/Val эта
кислоты зависит от аминокислот в положениях
способность была потеряна. Поскольку амино
155, 158 и 228 [29, 30]. В нашем исследовании
кислоты 190 и 225 тесно контактируют с конце
способность связываться с Neu5Gc рецептором
вой частью рецептора - сиаловой кислотой и
для вирусов 1992-1999 гг. коррелировала с па
прилегающей областью галактозы, они не могли
рой аминокислот 145 и 155: вирусы с Lys145 и
напрямую влиять на удаленные остатки моноса
His155 связывали Neu5Gc рецептор, в то время
харидов. Вероятно, замены в этих положениях
как вирусы с Asn145, His155 и Lys145, Thr155 не
слегка разворачивают сиаловую кислоту, что
связывали (табл. 2). Предполагаемое располо
приводит к более или менее успешному распо
жение Lys145 относительно N гликолильной
ложению STn и LSTb в РСУ.
группы N гликолилнейраминовой кислоты по
Одновременно с тем, что STn и LSTb стано
казано на рис. 2. По видимому, аминогруппа
вятся доступными для вируса, 6'SLN становится
Lys145 была способна образовывать водородную
рецептором с наибольшим сродством, и даль
связь с N гликолильной группой.
нейшая эволюция усилила эту тенденцию.
Нерегулярное появление и исчезновение
Сродство к STn, LSTb и 6'SL исчезало один за
сродства к Neu5Gc фрагменту указывает на не
другим, а сродство к 6'SLN оставалось высоким.
адаптивный характер этого свойства. Neu5Gc
После замены Gly225Asp первоначальный меха
фрагмент отсутствует на клетках человека; сле
низм распознавания рецепторов (посредством
довательно, способность связывать эту форму
гидрофобного взаимодействия 226 аминокисло
БИОХИМИЯ том 84 вып. 10 2019
РЕЦЕПТОРНАЯ СПЕЦИФИЧНОСТЬ ВИРУСОВ ГРИППА ЧЕЛОВЕКА Н3N2
1457
Рис. 2. Предполагаемое расположение Lys145 в сайте связывания рецептора H3 HA. Помечены Trp153, Lys145 и N глико
лильная группа сиаловой кислоты (SA). Lys145 вместо Ser145 и N гликолильная группа вместо N ацетильной были встро
ены в модель с помощью программного обеспечения DS ViewerPro 5.0, Accelrys Inc., с использованием трехмерной моде
ли Х 31 HA в комплексе с пентасахаридом LSTc [33].
journal/biokhsm/
ты с метиленовой группой галактозы) был утра
низм ответственен за дискриминационное свя
чен, а РСУ вирусов подвергся глубокой рекон
зывание 6'SL/6'SLN для вирусов H3N2. Такая
струкции. Вместо Leu226 и Ser228 в РСУ виру
конвергентная эволюция рецепторной специ
сов H3N2 после 2003 г. мы обнаруживаем
фичности подтипов H1N1 и H3N2 предполагает,
Asp190, Asp225, Val226 и Ser228, которые отлича
что 6'SLN является оптимальным природным
ют эти вирусы от вирусов гриппа птиц.
человеческим рецептором для вирусов гриппа.
Таким образом, в ходе эволюции вирусов
H3N2 и H1N1 человека их рецепторный фено
Финансирование. Работа выполнена при под
тип становился все более похожим, и аминокис
держке РФФИ (гранты 02 04 48109, 01 04 49300
лотный состав РСУ стал ближе, чем в начале
и 14 04 00547 и 17 04 00148) и МНТЦ (грант 5
циркуляции вирусов H3N2. Вирусы человека
2464).
H1N1 и H3N2 2003-2009 гг. имеют множество
Благодарности. С глубоким прискорбием со
сайтов гликозилирования в верхней части HA и
общаем, что наш коллега Александр Климов
Asp в положениях 190 и 225; и те, и другие раз
скончался.
личают 6'SL и 6'SLN. Ранее была показана роль
Конфликт интересов. Авторы заявляют об от
Asp190 и Asp225 в селективном сродстве вирусов
сутствии конфликта интересов.
H1N1 к 6'SLN [10]. Кристаллографический ана
Соблюдение этических норм. Данная статья
лиз молекулы H1 HA выявил водородные связи
не содержит исследований, в которых в качестве
между Asp190 и азотом группы GlcNAc 3 в мо
объектов использовали людей или животных.
лекуле 6'SLN [34]. Вероятно, подобный же меха
БИОХИМИЯ том 84 вып. 10 2019
1458
ГАМБАРЯН и др.
СПИСОК ЛИТЕРАТУРЫ
1.
Webster, R.G., Bean, W.J., Gorman, O.T., Chambers,
15.
Bush, R.M., Bender, C.A., Subbarao, K., Cox, N.J., and
T.M., and Kawaoka, Y. (1992) Evolution and ecology of
Fitch, W.M. (1999) Predicting the evolution of human
influenza A viruses, Microbiol. Rev., 56, 152-179.
influenza A, Science, 286, 1921-1925, PMID: 10583948.
2.
Paulson, J.C. (1985) Interactions of animal viruses with
16.
Wang, X., Ilyushina, N.A., Lugovtsev, V.Y., Bovin, N.V.,
cell surface receptors, in The receptors. Vol. 2 (Conn, M.,
Couzens, L.K., Gao, J., Donnelly, R.P., Eichelberger, M.C.,
ed.), pp. 131-219. Academic Press, Orlando, FL.
and Wan, H. (2017) Amino acids in hemagglutinin anti
3.
Wiley, D.C., and Skehel, J.J. (1987) The structure and
genic site B determine antigenic and receptor binding dif
function of the hemagglutinin membrane glycoprotein of
ferences between A(H3N2)v and ancestral seasonal H3N2
influenza virus, Ann. Rev. Biochem., 56, 365-394, doi: 10.
influenza viruses, J. Virol., 91, doi: 10.1128/JVI.01512 16.
1146/annurev.bi.56.070187.002053.
17.
Mochalova, L., Gambaryan, A., Romanova, J., Tuzikov, A.,
4.
Rogers, G.N., and D’Souza, B.L., (1989) Receptor bind
Chinarev, A., Katinger, D., Katinger, H., Egorov, A., and
ing properties of human and animal H1 influenza virus iso
Bovin, N. (2003) Receptor binding properties of modern
lates, Virology, 173, 317-322.
human influenza viruses primarily isolated in Vero and
5.
Connor, R.J., Kawaoka, Y., Webster R.G., and Paulson, J.C.
MDCK cells and chicken embryonated eggs, Virology, 313,
(1994) Receptor specificity in human, avian, and equine
473-480.
H2 and H3 influenza virus isolates, Virology, 205, 17-23,
18.
Stevens, J, Chen, L.M., Carney, P.J., Garten, R., Foust, A.,
doi: 10.1006/viro.1994.1615.
Le, J., Pokorny, B.A., Manojkumar, R., Silverman, J.,
6.
Gambaryan, A.S., Tuzikov, A.B., Piskarev, V.E.,
Devis, R., Rhea, K., Xu, X., Bucher, D.J., Paulson, J.C.,
Yamnikova, S.S., Lvov, D.K., Robertson, J.S., Bovin, N.V.,
Cox, N.J., Klimov, A., and Donis, R.O. (2010) Receptor
and Matrosovich, M.N. (1997) Specification of receptor
specificity of influenza A H3N2 viruses isolated in mam
binding phenotypes of influenza virus isolates from differ
malian cells and embryonated chicken eggs, J. Virol., 84,
ent hosts using synthetic sialylglycopolymers: non egg
8287-8299, doi: 10.1128/JVI.00058 10.
adapted human H1 and H3 influenza A and influenza B
19.
Wu, N.C., Xie, J., Zheng, T., Nycholat, C.M., Grande, G.,
viruses share a common high binding affinity for
Paulson, J.C., Lerner, R.A., and Wilson, I.A.
(2017)
6’ sialylN (acetyllactosamine), Virology, 232, 345-350,
Diversity of functionally permissive sequences in the recep
doi: 10.1006/viro.1997.8572.
tor binding site of influenza hemagglutinin, Cell Host
7.
Matrosovich, M.N., Gambaryan, A.S., Teneberg, S.,
Microbe, 21, 742-753, doi: 10.1016/j.chom.2017.05.011.
Piskarev, V.E., Yamnikova, S.S., Lvov, D.K., Robertson, J.S.,
20.
Gulati, S., Smith, D.F., Cummings, R.D., Couch, R.B.,
and Karlsson, K.A. (1997) Avian influenza A viruses differ
Griesemer, S.B., St George, K., Webster, R.G., and Air, G.M.
from human viruses by recognition of sialyloligosaccha
(2013) Human H3N2 influenza viruses isolated from 1968
rides and gangliosides and by a higher conservation of the
to 2012 show varying preference for receptor substructures
HA receptor binding site, Virology,
233,
224-234,
with no apparent consequences for disease or spread, PLoS
doi: 10.1006/viro.1997.8580.
One, 8, 66325, doi: 10.1371/journal.pone.0066325.
8.
Matrosovich, M., Tuzikov, A., Bovin, N., Gambaryan, A.,
21.
Yang, H., Carney, P.J., Chang, J.C., Guo, Z., Villanueva, J.M.,
Klimov, A., Castrucci, M.R., Donatelli, I., and Kawaoka, Y.
and Stevens, J. (2015) Structure and receptor binding pref
(2000) Early alterations of the receptor binding properties
erences of recombinant human A(H3N2) virus hemagglu
of H1, H2, and H3 avian influenza virus hemagglutinins
tinins, Virology, 477, 18-31, doi: 10.1016/j.virol.2014.
after their introduction into mammals, J. Virol., 74,
12.024.
8502-8512, doi: 10.1128/JVI.74.18.8502 8512.2000.
22.
Peng, W., de Vries, R.P., Grant, O.C., Thompson, A.J.,
9.
Gambaryan, A.S., Piskarev, V.E., Yamskov, I.A.,
McBride, R., Tsogtbaatar, B., Lee, P.S., Razi, N., Wilson, I.A.,
Sakharov, A.M., Tuzikov, A.B., Bovin, N.V., Nifant’ev, N.E.,
Woods, R.J., and Paulson, J.C. (2017) Recent H3N2 virus
and Matrosovich, M.N. (1995) Human influenza virus recog
es have evolved specificity for extended, branched human
nition of sialyloligosaccharides, FEBS Lett., 366, 57-60.
type receptors, conferring potential for increased avidity,
10.
Gambaryan, A.S., Robertson, J.S., and Matrosovich, M.N.
Cell Host Microbe, 21, 23-34, doi: 10.1016/j.chom.
(1999) Effects of egg adaptation on the receptor binding
2016.11.004.
properties of human influenza A and B viruses, Virology,
23.
Bovin, N.V., Korchagina, E.Yu., Zemlyanukhina, T.V.,
258, 232-239, doi: 10.1006/viro.1999.9732.
Byramova, N.E., Galanina, O.E., Zemlyakov, A.E.,
11.
Grassauer, A., Egorov, A.Y., Ferko, B., Romanova, J.,
Ivanov, A.E., Zubov, V.P., and Mochalova, L.V. (1993)
Katinger, H., and Muster, T. (1998) A host restriction
Synthesis of polymeric neoglycoconjugates based on N sub
based selection system for influenza haemagglutinin trans
stituted polyacrylamides, Glycoconj. J., 10, 142-151.
fectant viruses, J. Gen. Virol., 79, 1405-1409, doi: 10.1099/
24.
Shilova, N.V., Galanina, O.E., Pochechueva, T.V.,
0022 1317 79 6 1405.
Chinarev, A.A., Kadykov, V.A., Tuzikov, A.B., and Bovin, N.V.
12.
Nobusawa, E., Ishihara, H., Morishita, T., Sato, K., and
(2005) High molecular weight neoglycoconjugates for solid
Nakajima, K. (2000) Change in receptor binding specifici
phase assays, Glycoconj. J., 22, 43-51, doi: 10.1007/
ty of recent human influenza A viruses (H3N2): a single
s10719 005 0280 y.
amino acid change in hemagglutinin altered its recognition
25.
Gambaryan, A.S., and Matrosovich, M.N. (1992) A solid
of sialyloligosaccharides, Virology,
278,
587-596,
phase enzyme linked assay for influenza virus receptor
doi: 10.1006/viro.2000.0679.
binding activity, J. Virol. Methods, 39, 111-123.
13.
Medeiros, R., Escriou, N., Naffakh, N., Manuguerra J.C.,
26.
Hayase, T., Rice, K.G., Dziegielewska, K.M., Kuhlen
and van der Werf, S. (2001) Hemagglutinin residues of
schmidt, M., Reilly, T., and Lee, Y.C. (1992) Comparison
recent human A (H3N2) influenza viruses that contribute
of N glycosides of fetuins from different species and human
to the inability to agglutinate chicken erythrocytes,
alpha2 HS glycoprotein, Biochemistry, 31, 4915-4921.
Virology, 289, 74-85, doi: 10.1006/viro.2001.1121.
27.
Anders, E.M., Scalzo, A.A., Rogers, G.N., and White, D.O.
14.
Fitch, W.M., Bush, R.M., Bender, C.A., and Cox, N.J.
(1986) Relationship between mitogenic activity of influen
(1997) Long term trends in the evolution of H(3) HA1
za viruses and the receptor binding specificity of their
human influenza type A, Proc. Natl. Acad. Sci. USA, 94,
hemagglutinin molecules, J. Virol., 60, 476-482.
7712-7718, doi: 10.1073/pnas.94.15.7712.
28.
Suzuki, Y. (2001) Host mediated variation and receptor
БИОХИМИЯ том 84 вып. 10 2019
РЕЦЕПТОРНАЯ СПЕЦИФИЧНОСТЬ ВИРУСОВ ГРИППА ЧЕЛОВЕКА Н3N2
1459
binding specificity of influenza viruses, Adv. Exp. Med.
thetic and natural sialosides. Virology, 196, 111-121,
Biol., 491, 445-451.
doi: 10.1006/viro.1993.1459.
29. Byramova, N.E., Tuzikov, A.B., and Bovin, N.V. (1992)
32. Pritchett, T.J., Brossmer, R., Rose, U., and Paulson, J.C.
A simple procedure for the synthesis of the methyl and
(1987) Recognition of monovalent sialosides by influenza
benzyl glycosides of Neu5Ac and
4 epi Neu5Ac,
virus H3 hemagglutinin, Virology, 160, 502-506.
Carbohydr. Res., 237, 161-175, doi: 10.1016/S0008 6215
33. Eisen, M.B., Sabesan, S., Skehel, J.J., and Wiley, D.C.
(92)84240 S.
(1997) Binding of the influenza A virus to cell surface
30. Matrosovich, M.N., Mochalova, L.V., Marinina, V.P.,
receptors: structures of five hemagglutinin-sialyloligosac
Byramova, N.E., and Bovin, N.V. (1990) Synthetic poly
chride complexes determined by X ray crystallography,
meric sialoside inhibitors of influenza virus receptor binding
Virology, 232, 19-31, doi: 10.1006/viro.1997.8526.
activity, FEBS Lett., 272, 209-212.
34. Gamblin, S.J, Haire, L.F, Russell, R.J, Stevens, D.J, Xiao, B.,
31. Matrosovich, M.N., Gambaryan, A.S., Tuzikov, A.B.,
Ha, Y., Vasisht, N., Steinhauer, D.A., Daniels, R.S., Elliot, A.,
Byramova, N.E., Mochalova, L.V., Golbraikh, A.A.,
Wiley, D.C., and Skehel, J.J. (2004) The structure and
Shenderovich, M.D., Finne, J., and Bovin, N.V. (1993)
receptor binding properties of the 1918 influenza hemag
Probing of the receptor binding sites of the H1 and H3
glutinin, Science,
303,
1838-1842, doi:
10.1126/
influenza A and influenza B virus hemagglutinins by syn
science.1093155.
CHANGES IN THE RECEPTOR BINDING PROPERTIES OF H3N2
VIRUSES IN PROLONGED CIRCULATION IN HUMANS
A. S. Gambaryan1*, A. Balish2, A. I. Klimov2, A. B. Tuzikov3,
A. A. Chinarev3, G. V. Pazynina3, and N. V. Bovin3
1 Chumakov Federal Scientific Center for Research and Development
of ImmuneKandKBiological Products, Russian Academy of Sciences,
108819 Moscow, Russia; EKmail: al.gambaryan@gmail.com
2 Influenza Branch, Centers for Disease Control and Prevention,
Atlanta, GA 30333, USA
3 Shemyakin-Ovchinnikov Institute of Bioorganic Chemistry,
Russian Academy of Sciences, 117997 Moscow, Russia
Received April 3, 2019
Revised June 5, 2019
Accepted June 15, 2019
It was previously shown that amino acids 155Thr, 158Glu, and 228Ser of H3N2 virus hemagglutinin (HA) are crucial
for recognition of Neu5Gc. In this study, we have shown that the ability of the virus to bind the Neu5Gc terminated
receptor can also depend on the amino acid 145 of HA: virus isolates of 1972-1999 with Lys145 in HA could bind to
the receptor, whereas those with Asn145 could not. Sporadic appearance and disappearance of the ability to bind
Neu5Gc terminated oligosaccharides as well as the absence of Neu5Gc residues in human glycoconjugates indicate
the non adaptive nature of this property. It was previously shown that the H3N2 virus isolates of 1968-1989, unlike
H1N1 viruses, do not distinguish between Neu5Acα2 6Galβ1 4Glc (6'SL) and Neu5Acα2 6Galβ1 4GlcNAc
(6′SLN). The H3N2 viruses isolated after 1993 have acquired an ability to distinguish between 6′SL and 6′SLN, sim
ilar to H1N1 viruses. We determined that predominant affinity of H3N2 viruses for 6′SLN gradually increased from
1992 to 2003. After 2003, H3N2 viruses lost an ability to bind a number of sialosides including 6′SL that were good
receptors for the earlier H3N2 isolates, and only 6′SLN retains high affinity for the later human H3N2 isolates. This
property correlates with acquisition of new glycosylation sites at positions 122, 133 and 144 as well as with Glu190Asp
and Gly225Asp replacements in HA. These replacements are also responsible for the receptor binding phenotype of
human H1N1 viruses. Convergent evolution of receptor specificity of the H1N1 and H3N2 subtypes indicates that
6'SLN is an optimal natural human receptor for influenza viruses.
Keywords: influenza virus, hemagglutinin, sialooligosaccharides, receptor specificity, H3N2
БИОХИМИЯ том 84 вып. 10 2019