БИОХИМИЯ, 2019, том 84, вып. 10, с. 1500 - 1510
УДК 577.171.6
СИГНАЛЬНЫЕ ПУТИ ПРОЛАКТИНА, ДЕТЕРМИНИРУЮЩИЕ
ЕГО ПРЯМЫЕ ЭФФЕКТЫ НА ПОЧКУ В МОДЕЛИ
ХОЛЕСТАЗА БЕРЕМЕННЫХ
© 2019 П.А. Абрамичева1*, Т.А. Балакина1, И.А. Морозов2,
Т.А. Щелкунова1, О.В. Смирнова1
1 Московский государственный университет им. М.В. Ломоносова, биологический факультет,
119991 Москва, Россия; электронная почта: abramicheva.polina@gmail.com
2 Институт молекулярной биологии им. В.А. Энгельгардта РАН,
119991 Москва, Россия
Поступила в редакцию 29.01.2019
После доработки 21.06.2019
Принята к публикации 21.06.2019
Холестаз беременных - патология беременности, связанная с нарушением оттока желчи и водно солевым
обменом. Пролактин - это один из важнейших регуляторов водно солевого баланса. Изучено изменение
экспрессии длинной и короткой изоформ рецептора пролактина (ПРЛР) и посредников пролактинового
сигналинга в коре и внешнем мозговом слое почки крысы в модели холестаза беременных и группах срав
нения с помощью иммуноблоттинга и ОТ ПЦР. Показано, что в условиях холестаза беременных в реализа
ции эффектов пролактина принимают участие обе изоформы ПРЛР. Доказаны прямые эффекты пролакти
на на почку, так как: 1) экспрессия мРНК обеих изоформ ПРЛР в почке изменяется при различных физио
логических состояниях (при холестазе и холестазе беременных) в зависимости от уровня пролактина; 2) по
казан рост уровня экспрессии pSTAT5, как ключевого посредника сигналинга длинной изоформы ПРЛР
при холестазе беременных; 3) продемонстрирован рост экспрессии мРНК терминаторов пролактинового
сигналинга SOCS3 и PIAS3, являющихся молекулярными мишенями длинной изоформы ПРЛР и имеющих
STAT чувствительные элементы в промоторах своих генов, в условиях преобладания длинной изоформы
ПРЛР; 4) обнаружено снижение экспрессии мРНК GALT, как молекулярной мишени короткой изоформы
ПРЛР, в условиях относительного роста ее содержания во внешнем мозговом слое почки.
КЛЮЧЕВЫЕ СЛОВА: короткая и длинная изоформы пролактинового рецептора, STAT5, SOCS3, PIAS3,
холестаз беременных, гиперпролактинемия, почка.
DOI: 10.1134/S0320972519100117
Холестаз беременных - патология беремен
вень пролактина существенно повышен по
ности, сопровождающаяся не только повыше
сравнению с выявляемым при нормально про
нием уровней билирубина, желчных кислот и
текающей беременности [4], что также подтвер
печеночных ферментов в крови, но и нарушени
ждено нашей лабораторией на эксперименталь
ями водно солевого обмена. Данное заболева
ной модели холестаза беременных [7]. Пролак
ние характеризуется неблагоприятным прогно
тин регулирует >300 функций организма, кото
зом для плода, связанным с поступлением к
рые можно сгруппировать в две большие катего
плоду токсических продуктов обмена от матери
рии: связанные с регуляцией размножения и с
[1-5]. Известно, что уровень пролактина в плаз
поддержанием гомеостаза [8-10]. Если роль
ме крови значительно возрастает в течение бере
пролактина в регуляции репродуктивной функ
менности, что объясняется стимуляцией мате
ции хорошо изучена, то исследования его учас
ринских лактотрофов гипофиза плацентарными
тия в регуляции процессов метаболизма, в част
эстрогенами [6]. При холестазе беременных уро
ности водно солевого обмена у млекопитаю
Принятые сокращения: ПРЛР - рецептор пролактина, JAK2 - нерецепторная тирозинкиназа 2 семейства Janus,
STAT5A - трансдуктор сигнала и активатор транскрипции 5А, STAT5B - трансдуктор сигнала и активатор транскрипции 5В,
FOXO3 - транскрипционный фактор семейства Forkhead 3, GALT - галактоза 1 фосфат уридилтрансфераза, CIS - суп
рессор цитокинового сигналинга, SOCS3 - супрессор цитокинового сигналинга 3, PIAS3 - белковый ингибитор
активированного STAT, HPRT - гипоксантингуанинфосфорибозилтрансфераза, GAPDH - глицеральдегидфосфат
дегидрогеназа.
* Адресат для корреспонденции.
1500
СИГНАЛЬНЫЕ ПУТИ ПРОЛАКТИНА
1501
щих, только начинают развиваться. Нами было
inducible SH 2 domain proteins), SOCS3, относя
показано, что суточный клиренс натрия возрас
щийся к семейству SOCS белков (Suppressors of
тает в условиях гиперпролактинемии при холес
Cytokine Signaling), и PIAS3 (protein inhibitor of
тазе беременных по сравнению с нормальными
activated STAT), причем SOCS3 может подавлять
животными [11, 12].
сигналинг и короткой изоформы ПРЛР. Все пе
В тканях млекопитающих, регулирующих
речисленные терминаторы пролактинового сиг
гомеостаз воды и солей, обнаружен высокий
налинга обладают STAT чувствительными эле
уровень экспрессии рецептора пролактина
ментами. Например, в промоторе гена CIS со
(ПРЛР). В нашей лаборатории с помощью мето
держится 4 STAT5 связывающих сайта. Есть
да иммуногистохимии было показано, что уро
данные о том, что у мышей с нокаутом по
вень ПРЛР в разных отделах почки по разному
STAT5A и STAT5B исчезает экспрессия CIS в
меняется в модели холестаза беременных: в
яичнике [18].
проксимальных канальцах он не отличается от
Цель нашего исследования - установить, яв
контрольных групп, в дистальных канальцах
ляются ли эффекты пролактина в коре и мозго
уровень ПРЛР падает, а в канальцах мозгового
вом слое почки прямыми в норме и в модели хо
слоя - растет [13].
лестаза беременных. О возможности прямых
ПРЛР относится к классу 1 суперсемейства
эффектов мы судили по ряду параметров: 1) на
рецепторов цитокинов. Наиболее хорошо опи
личию и уровню экспрессии мРНК короткой и
саны изоформы, обнаруженные у крысы: длин
длинной изоформ ПРЛР; 2) возможности роста
ная (80-85 кДа), короткая (45 кДа) и мутантная
фосфорилирования белка STAT5 в условиях ги
средняя форма (65 кДа), обнаруженная в про
перпролактинемии в модели холестаза беремен
лактин зависимой линии T клеточной лимфо
ных при контроле стабильности экспрессии
мы Nb2 крысы [14, 15]. Считается, что большин
мРНК STAT5А и STAT5В; 3) по наличию измене
ство известных путей передачи сигнала связано
ний экспрессии мРНК терминаторов пролакти
с длинной изоформой ПРЛР и обусловлено ак
нового сигналинга, таких как CIS, SOCS3 и
тивацией нерецепторной тирозинкиназы 2 се
PIAS3, являющихся молекулярными мишенями
мейства Janus (JAK2) и последующим фосфори
длинной изоформы ПРЛР и имеющих STAT
лированием, главным образом транскрипцион
чувствительные элементы в промоторах своих
ных факторов STAT5А и STAT5В (Signal Trans
генов; 4) по наличию изменений экспрессии
ducers and Activators of Transcription), и трансло
мРНК молекулярных мишеней короткой изо
кацией димеризованных фосфорилированных
формы ПРЛР, таких как FOXO3 и GALT.
STAT5 белков в ядро, где они взаимодействуют
со специфическими STAT чувствительными
элементами (GAS элементами) в промоторах
МЕТОДЫ ИССЛЕДОВАНИЯ
генов мишеней [8, 16]. Функции и сигналинг
короткой изоформы ПРЛР пока мало изучены.
Модели животных. Работа выполнена на
Известно, что она выступает в роли доминант
группах белых беспородных половозрелых са
ного негативного регулятора, образуя гетероди
мок крыс (масса 250-300 г): интактных («нор
меры с длинной изоформой ПРЛР, тем самым
ма», n = 16), с гиперпролактинемией, вызываю
подавляя проведение сигнала через длинную
щей ложную беременность («пролактин», n = 15),
изоформу. К молекулярным посредникам сиг
с холестазом («холестаз», n = 16) и с моделью хо
налинга короткой изоформы относят тран
лестаза беременных (обструктивный холестаз в
скрипционный фактор FOXO3 (Forkhead
сочетании с гиперпролактинемией - «холестаз +
Transcription Factor 3), который стимулирует
пролактин», n = 19). Гиперпролактинемию мо
экспрессию гена фермента метаболизма галак
делировали с помощью трансплантации гипо
тозы GALT (галактоза 1 фосфат уридилтрансфе
физа крысы донора под капсулу почки крысы
раза). Halperin et al. [17] показали, что у транс
реципиента (производился надрез капсулы поч
генных мышей, экспрессирующих только ко
ки у крысы реципиента, параллельно из турец
роткие изоформы ПРЛР, под действием пролак
кого седла извлекали гипофиз крысы донора;
тина происходит подавление FOXO3, и, соотве
гипофиз помещали под капсулу почки), что
тственно, ингибирование экспрессии GALT в
приводило к повышению продукции гомоло
яичниках, в результате чего происходит уско
гичного пролактина [7, 19-21], холестаз - путем
ренная инициация роста фолликулов яичников
перевязки общего желчного протока. Животных
и их последующая гибель.
содержали в стандартных условиях вивария при
К основным транскрипционным факторам,
естественном световом режиме и свободном
подавляющим проведение сигнала через длин
доступе к воде и пище. Приживаемость гипофи
ную изоформу ПРЛР, относят CIS (cytokine
за оценивали по визуальным критериям - по
БИОХИМИЯ том 84 вып. 10 2019
1502
АБРАМИЧЕВА и др.
степени васкуляризации данного транспланта
низировали в соответствующем объеме реагента
та. Индукция гиперпролактинемии при транс
Trizol (“Invitrogen”, США) с целью дальнейшего
плантации гипофиза в модели холестаза бере
выделения суммарной РНК. Препарат обраба
менных и при изолированной гиперпролакти
тывали ДНКазой I RQ1 DNase («Promega»,
немии доказана нами ранее.
США). Для удаления ДНКазы I РНК очищали
Фидченко и соавт. показали [7], что в груп
на колонках c использованием набора RNeasy
пах «пролактин» (n = 6) и «холестаз + пролак
Plus Mini Kit («Qiagen», США), после чего про
тин» (n = 17) при успешной приживаемости
водили реакцию обратной транскрипции с по
трансплантата гипофиза концентрация пролак
мощью набора ImProm IITM Reverse Transcrip
тина составляла 12,90 ± 2,93 и 48,80 ± 5,78 нг/мл
tion System («Promega», США). Во всех случаях
и была достоверно (p < 0,05) выше, чем в соот
процедуры выполняли по инструкции произво
ветствующем контроле - «норма» (n = 10) -
дителя. Для проведения ПЦР в реальном време
2,30 ± 0,74 нг/мл, «холестаз» (n= 31) - 11,8 ± 2,65 нг/мл.
ни использовали амплификатор Bio Rad CFX96
Кроме того, концентрация пролактина при хо
(«Bio Rad», Канада) и наборы реакционных
лестазе была достоверно (p < 0,05) выше по
смесей («Синтол», Россия), праймеры (таблица)
сравнению с нормой.
добавляли до конечной концентрации 160 нМ.
Через 2 недели после проведенной операции
Режимы амплификации: 95 °C - 5 мин; 95 °C -
внешний мозговой слой и кору почки выделяли
10 с; 58 °C - 17 с; 72 °C - 20 с, 40 циклов; кривая
с целью получения материала для определения
плавления 72-95 °C, инкремент 0,5 °C - 5 с.
уровня мРНК изоформ ПРЛР, вторичных по
Подбор праймеров производили с помощью
средников и терминаторов пролактинового сиг
программы Beacon Designer 7, а также с исполь
налинга и белка STAT5 и pSTAT5.
зованием баз данных NCBI, BLAT Search. В ка
ОТ6ПЦР. Кору и внешний мозговой слой
честве генов домашнего хозяйства использовали
почки отделяли при помощи скальпеля, гомоге гены
глицеральдегидфосфатдегидрогеназы
Последовательности олигонуклеотидов, использованные в ПЦР
Название гена
Название
Последовательность олигонуклеотида
Длина ПЦР
олигонуклеотида
продукта, п.н.
Длинная изоформа пролактинового
PRLR for
5' CTGGGCAGTGGCTTTGAAG 3'
131
рецептора
PRLR(L) rev
5' CCAAGGCACTCAGCAGCTCT 3'
Короткая изоформа пролактинового
PRLR for
5' CTGGGCAGTGGCTTTGAAG 3'
143
рецептора
PRLR(S) rev
5' AAGGGCCAGGTACAGATCCA 3'
Трансдуктор сигнала и активатор
STAT5A for
5' ATTCACCACTCGGGATTTCTC 3'
186
транскрипции 5А
STAT5A rev
5' ACAAACTCAGGGACCACTTG 3'
Трансдуктор сигнала и активатор
STAT5B for
5' GAACACCCGCAATGATTATAGC 3'
159
транскрипции 5В
STAT5B rev
5' TTCTCTTCCGTGACCGACTC 3'
Транскрипционный фактор семейства
FOXO3 for
5' TGTCCTACGCTGACCTGATC 3'
196
Forkhead 3
FOXO3 rev
5' TTCTGAACCCGCATGAAACG 3'
Галактоза 1 фосфат
GALT for
5' AAGGAAGGTTCACAGTAGATGG 3'
185
уридилтрансфераза
GALT rev
5' GACTTTCAACTCTCCAGGGTAG 3'
Супрессор цитокинового сигналинга
CIS for
5' ATGAACCGAAGGTGCTAGAC 3'
106
CIS rev
5' TCACTGGCTGTAATGGAACC 3'
Супрессор цитокинового сигналинга 3
SOCS3 for
5' CCTGGACTCCTATGAGAAAGTG 3'
200
SOCS3 rev
5' CCTATCTTCTGCTGGGCTAAC 3'
Белковый ингибитор активированного
PIAS3 for
5' AGGAGCCAAGTGCGATTATAC 3'
194
STAT
PIAS3 rev
5' TGATGTTGATTGGACGACTAGG 3'
Гипоксантингуанинфосфорибозил
HPRT for
5' GCTATAAGTTCTTTGCTGACCTG 3'
149
трансфераза (ГПРТ)
HPRT rev
5' ATCTCCACCAATAACTTTTATGTCC 3'
Глицеральдегидфосфатдегидрогеназа
GAPDH for
5' GCAAGAGAGAGGCCCTCAG 3'
74
(ГАФД)
GAPDH rev
5' TGTGAGGGAGATGCTCAGTG 3'
БИОХИМИЯ том 84 вып. 10 2019
СИГНАЛЬНЫЕ ПУТИ ПРОЛАКТИНА
1503
(GAPDH) и гипоксантинфосфорибозилтранс
XRS plus («Bio Rad», Канада). Относительный
феразы (HPRT). Данные нормировали на гео
уровень белка оценивали методом денситомет
метрическое среднее пороговых циклов генов
рии с использованием программы ImageLab™
домашнего хозяйства, т.к. данный метод расчета
3.0 Software («Bio Rad», Канада).
позволяет нивелировать индивидуальные раз
Статистическая обработка данных. Статисти
личия между генами домашнего хозяйства в од
ческую обработку полученных результатов про
ной группе животных [22] по формуле:
водили с помощью однофакторного дисперси
онного анализа (one way ANOVA) с поправками
на множественное сравнение (критерий
Holm-Sidak) и корреляционного анализа (кри
,
терий Pearson) в программе GraphPad Prizm 7
(«GraphPad Software Inc.», США).
где R - условные единицы экспрессии мРНК, E -
эффективность амплификации, Ct - соответ
ствующий пороговый цикл, HPRT - ген, коди
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
рующий HPRT, GAPDH - ген, кодирующий
GAPDH.
Установлено, что во внешнем мозговом слое
Вестерн6блоттинг. Методика получения ли
почки экспрессия мРНК обеих изоформ непро
затов тканей почки и особенности электрофоре
порционально возрастает относительно нормы в
тического разделения белков описаны в нашей
группе «холестаз» с преобладанием короткой,
работе ранее [11]. Для детекции STAT5 и pSTAT5
как доминантной негативной. В условиях холес
использовали поликлональные антитела кроли
таза беременных соотношение экспрессии изо
ка (ab68465, 1 : 1000 и ab97734, 1 : 1000 соответ
форм изменяется в сторону длинной в связи с
ственно («Abcam», Канада)), для детекции рефе
непропорциональным снижением экспрессии
ренсного белка GAPDH - моноклональные ан
обеих изоформ ПРЛР относительно группы «хо
титела мыши, конъюгированные с пероксида
лестаз» (рис. 1, а, и 2, а). В коре почки экспрес
зой хрена (ab9482, 1 : 15000, («Abcam», Канада)).
сия длинной изоформы ПРЛР возрастает при хо
В качестве вторичных антител использовали по
лестазе беременных относительно нормы (рис. 1, б).
ликлональные антитела козы против иммуно
Статистически значимых изменений экспрессии
глобулинов кролика, конъюгированные с перок
короткой изоформы ПРЛР в коре не обнаружено
сидазой хрена (1 : 6000, («Bio Rad», Канада)).
(рис. 2, б). В коре соотношение экспрессии
Антитела, связавшиеся с мембраной, детектиро
мРНК длинной изоформы ПРЛР и короткой
вали с помощью набора для оценки хемолюми
возрастает при холестазе беременных относи
несценции Clarity™ Western ECL Substrate («Bio
тельно группы «холестаз» и «норма» в связи с
Rad», Канада) на приборе Endolab ChemiDoc
постепенным ростом длинной изоформы ПРЛР.
а
б
Рис. 1. Экспрессия мРНК длинной изоформы ПРЛР у крыс исследуемых групп во внешнем мозговом слое (а) и коре (б)
почки, n - количество животных. В связи с ограниченностью материала для исследования конкретное количество живот
ных в отдельных экспериментах отличается от суммарно указанного количества. Данные по группам на этом и последу
ющих рисунках представлены в виде среднего ± S.D. ** p < 0,01, **** p < 0,0001 - Статистически значимые различия по
сравнению с группой «норма», #### p < 0,0001 - по сравнению с группой «холестаз» (one way ANOVA, тест Sidak)
БИОХИМИЯ том 84 вып. 10 2019
1504
АБРАМИЧЕВА и др.
а
б
Рис. 2. Экспрессия мРНК короткой изоформы ПРЛР у крыс исследуемых групп во внешнем мозговом слое (а) и коре (б)
почки. *** p < 0,001 - Статистически значимые различия по сравнению с группой «норма», ### p < 0,001 - по сравнению
с группой «холестаз» (one way ANOVA, тест Sidak)
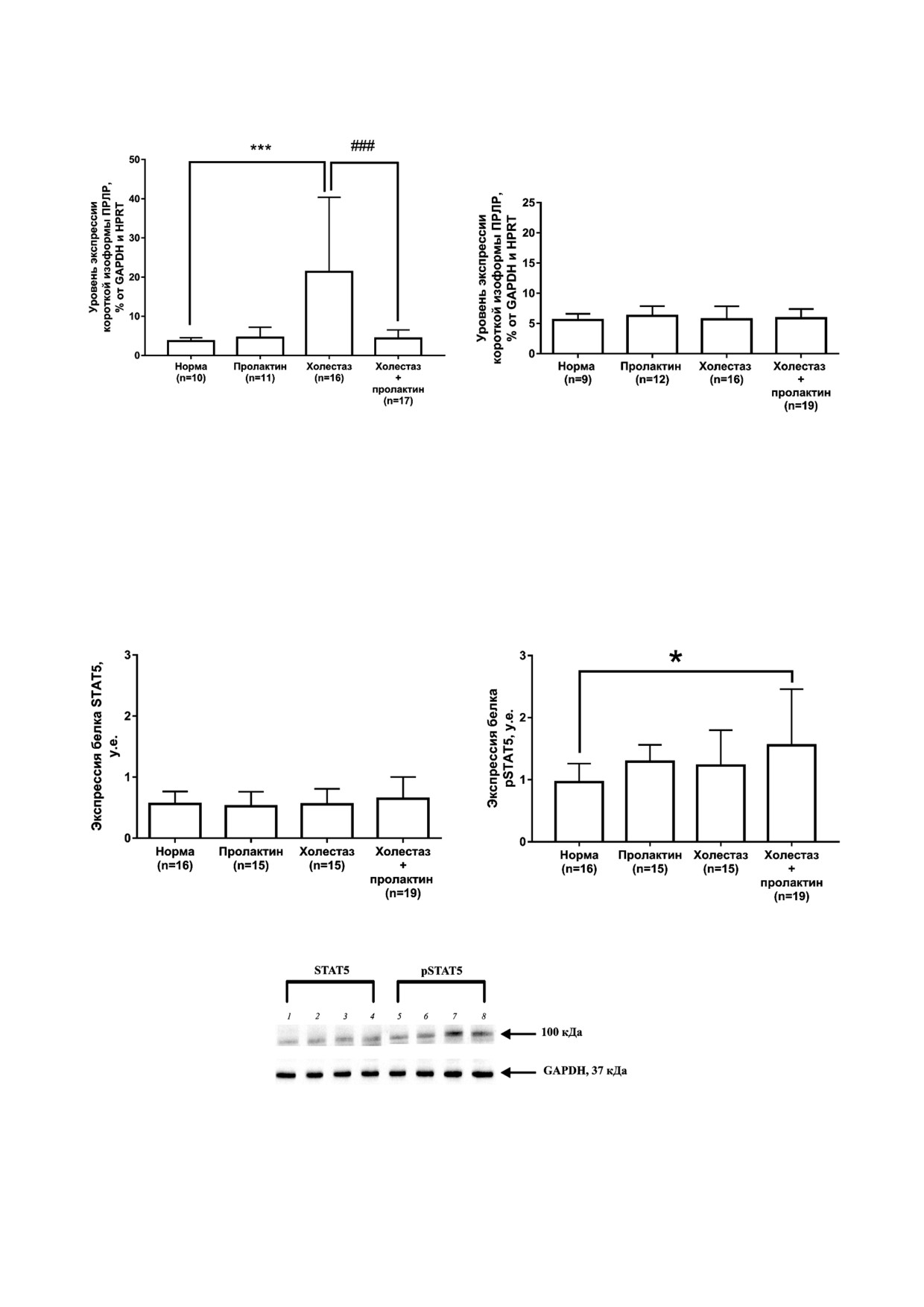
Была проанализирована экспрессия белка
слое почки, чего не наблюдается в коре почки.
STAT5 и его фосфорилированной формы pSTAT5.
Уровень экспрессии белкового продукта STAT5
При холестазе беременных увеличивается
(рис. 3, а и в) и уровень мРНК STAT5A и STAT5B
экспрессия белкового продукта pSTAT5 (рис. 3, б
не изменяется в обоих слоях почки в условиях
и в) по сравнению с нормой во внешнем мозговом
этой патологии (p > 0,05, на рис. не показано).
а
б
в
Рис. 3. Уровень экспрессии белка STAT5 (а и в) и pSTAT5 (б и в) во внешнем мозговом слое почки у крыс исследуемых
групп. * p < 0,05 - Статистически значимые различия по сравнению с группой «норма» (one way ANOVA, тест Sidak). Но
мера дорожек: 1 и 5 - норма, 2 и 6 - пролактин, 3 и 7 - холестаз, 4 и 8 - холестаз + пролактин
БИОХИМИЯ том 84 вып. 10 2019
СИГНАЛЬНЫЕ ПУТИ ПРОЛАКТИНА
1505
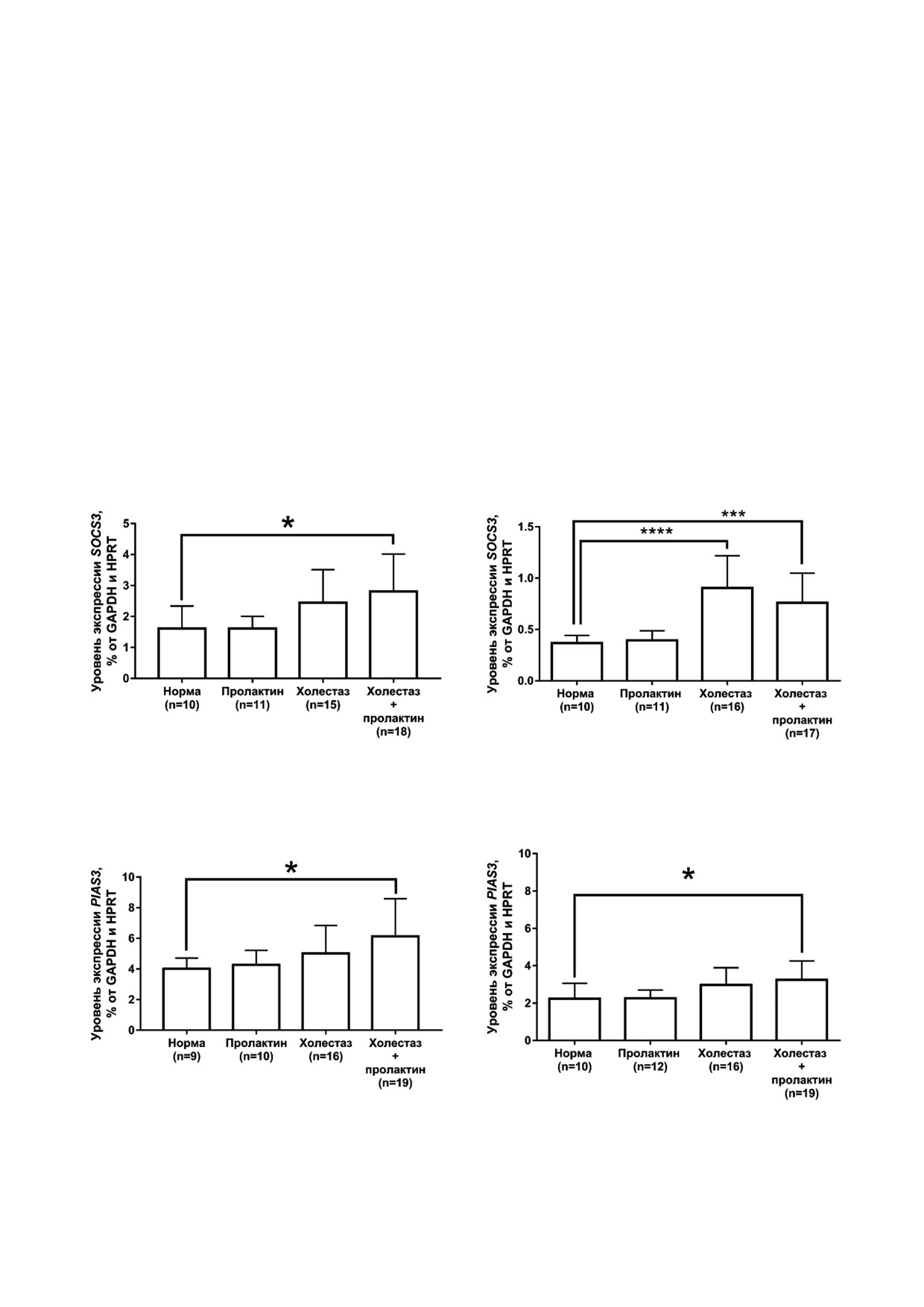
Во внешнем мозговом слое почки экспрес
слоях почки (p > 0,05, на рис. не показано). На
сия генов SOCS3 (рис. 4, а) и PIAS3 (рис. 5, а)
ми было показано наличие положительной кор
возрастает при холестазе беременных по сравне
реляции между экспрессией GALT и FOXO3
нию с интактными животными. В коре в усло
(p < 0,01; R2 = 0,39) в условиях холестаза бере
виях холестаза беременных также возрастает
менных и между экспрессией GALT и короткой
экспрессия как SOCS3 (рис. 4, б), так и PIAS3 от
изоформой ПРЛР у интактных крыс (p < 0,05;
носительно нормы (рис. 5, б), кроме того,
R2 = 0,555).
экспрессия SOCS3 возрастает в группе «холес
таз» относительно интактных крыс. Для CIS не
было выявлено изменений в экспрессии ни в од
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
ной из анализируемых структур почки (p > 0,05,
на рис. не показано).
Нами было установлено, что во внешнем
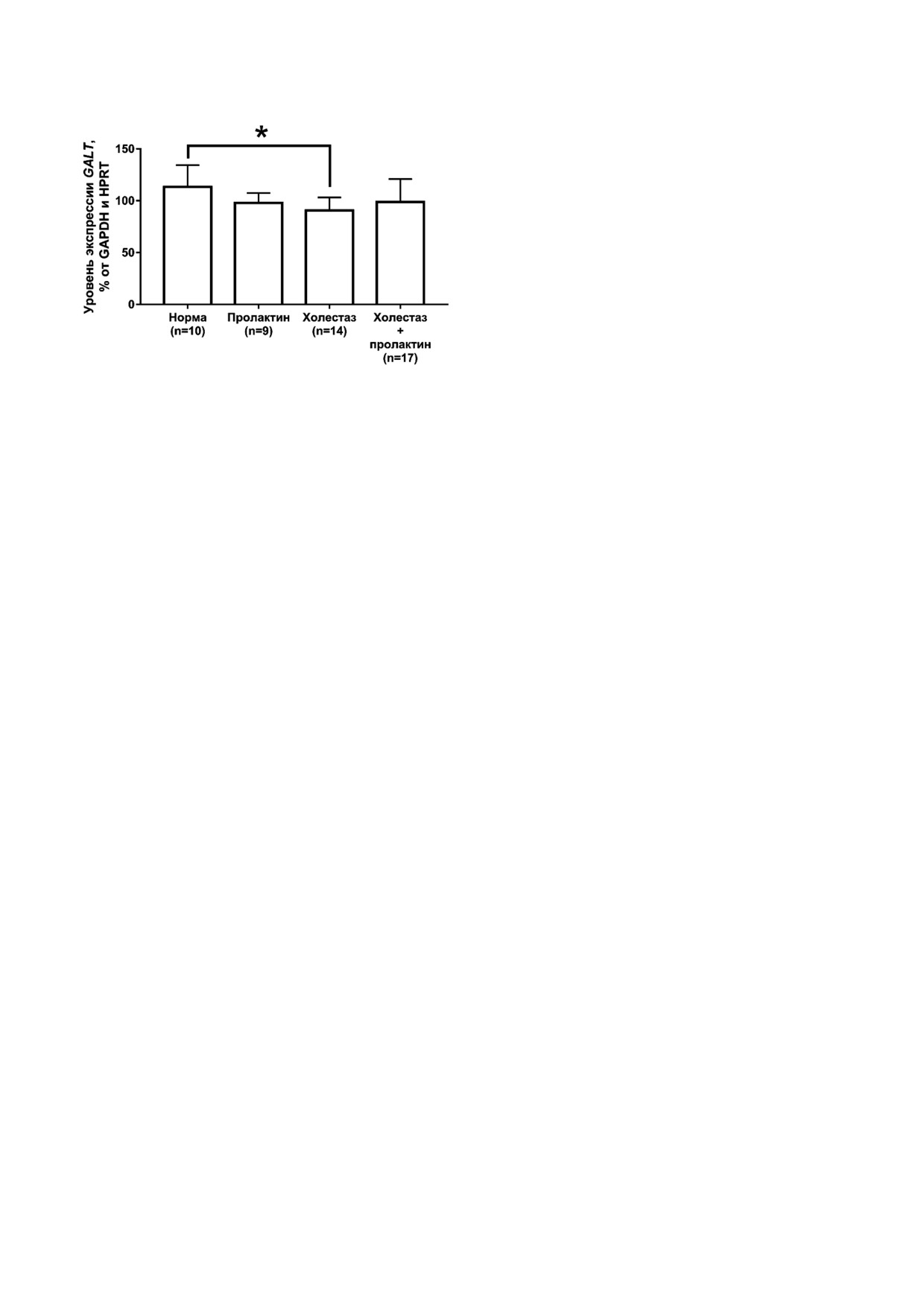
Что касается участников сигнального каска
мозговом слое почки при холестазе экспрессия
да короткой изоформы ПРЛР, то экспрессия
длинной изоформы ПРЛР возрастает в 3 раза,
мРНК GALT снижается при холестазе по сравне
короткой - в 5,8 раз, а в коре растет экспрессия
нию с нормой только во внешнем мозговом слое
только длинной изоформы - в 1,4 раза по срав
почки (рис. 6). Для FOXO3 не было обнаружено
нению с интактными животными. Заслуживает
статистически значимых изменений в обоих
внимания сам факт резкого увеличения
а
б
Рис. 4. Уровень экспрессии мРНК SOCS3 у крыс исследуемых групп во внешнем мозговом слое (а) и коре (б) почки. * p < 0,05,
*** p < 0,001, **** p < 0,0001 - Статистически значимые различия по сравнению с группой «норма» (one way ANOVA, тест
Sidak)
а
б
Рис. 5. Уровень экспрессии мРНК PIAS3 у крыс исследуемых групп во внешнем мозговом слое (а) и коре (б) почки. * p < 0,05 -
Статистически значимые различия по сравнению с группой «норма» (one way ANOVA, тест Sidak)
БИОХИМИЯ том 84 вып. 10 2019
1506
АБРАМИЧЕВА и др.
Полученные нами данные свидетельствуют в
пользу того, что GALT и FOXO3 вовлечены в
сигнальный каскад короткой изоформы ПРЛР
не только в яичнике, но и в почке. Наличие по
ложительной, а не отрицательной корреляции
между GALT и короткой изоформой ПРЛР мо
жет быть связано с тем, что, возможно, наблю
дается компенсаторный рост экспрессии GALT
в условиях его подавления, так как известны
участки промотора GALT, c которыми взаимо
действуют позитивные регуляторы экспрессии
этого гена [28]. Интересно отметить, что в дру
гих пролактин зависимых нерепродуктивных
тканях также изменяется соотношение экспрес
сии изоформ ПРЛР при холестазе по сравнению
Рис. 6. Уровень экспрессии мРНК GALT во внешнем моз
говом слое почки у крыс исследуемых групп. * p < 0,05 -
с нормой. Ранее в нашей лаборатории было по
Статистически значимые различия по сравнению с груп
казано, что в печени самок крыс в условиях
пой «норма» (one way ANOVA, тест Sidak)
обструктивного холестаза наблюдается сниже
ние экспрессии доминирующей короткой изо
чувствительности почки к пролактину в услови
формы ПРЛР и повышение длинной [29]. Кро
ях холестаза. Это может быть связано со смеще
ме того, в условиях холестаза у самок крыс в
нием функций пролактина в сторону регуляции
клетках желчного протока (холангиоцитах) про
водно солевого обмена, в том числе стимуляции
демонстрировано значительное увеличение
экскреции натрия и бикарбонатов [11, 12] и де
экспрессии длинной изоформы по сравнению с
токсикации [23].
нормой, а также присутствие короткой изофор
Кроме того, при холестазе во внешнем моз
мы [30]. Видимо, такие реципрокные изменения
говом слое почки наблюдается непропорцио
соотношения экспрессии изоформ ПРЛР в поч
нальный рост короткой изоформы ПРЛР. Учи
ке и печени в условиях холестаза позволяют ор
тывая, что короткая изоформа выполняет роль
ганизму лучше справляться с выведением ток
доминантного негативного регулятора в резуль
сичных продуктов обмена через почки.
тате образования гетеродимеров с длинной изо
Нами показано, что во внешнем мозговом
формой [24, 25], можно думать, что собственные
слое почки в условиях холестаза беременных
эффекты короткой изоформы в условиях холес
экспрессия обеих изоформ снижается относи
таза особенно хорошо выражены. Но нами по
тельно группы «холестаз», и начинает преобла
лучены данные об изменении экспрессии GALT
дать экспрессия длинной изоформы. Такое из
и ее корреляции с FOXO3 только в условиях хо
менение паттерна экспрессии изоформ свиде
лестаза беременности, но не холестаза. Вероят
тельствует об усилении проведения сигнала че
но, короткая изоформа в почках выполняет
рез длинную изоформу ПРЛР. Механизм акти
свою самостоятельную роль, значение которой
вации длинной изоформы ПРЛР под действием
возрастает в условиях патологии. На данный мо
молекулы пролактина включает димеризацию
мент об индивидуальных функциях короткой
двух молекул рецептора, взаимное фосфорили
изоформы ПРЛР известно только для яичников
рование и активацию двух молекул JAK2, фос
мыши: группой Devi было выявлено ингибиро
форилирование остатков тирозина доменов ре
вание MAPK, передающей сигнал от короткой
цептора box 2, образование сайтов докинга
изоформы, за счет ее ассоциации с DUPD1 (dual
STAT5, включающих SH2 домены, фосфорили
specific phosphatase) - фосфатазой, дефосфори
рование STAT5 под действием JAK2. Гомодиме
лирующей участников каскада, а также ингиби
ры pSTAT5 транслоцируются в ядро, где активи
рование транскрипционных факторов Sp1 и
руют гены, имеющие в своем промоторе STAT
FOXO3 и гена фермента метаболизма галактозы
чувствительные элементы [31, 32]. В наших
GALT, экспрессию которого стимулирует
опытах функциональная активность длинной
FOXO3 [17, 26, 27]. Нами было показано сниже
изоформы подтверждается фосфорилированием
ние экспрессии мРНК GALT в группе «холестаз»
ключевого посредника проведения ее сигнала -
относительно нормы во внешнем мозговом слое
STAT5 в условиях холестаза беременных относи
почки, а также наличие корреляций между
тельно интактных животных (изменений в
экспрессией GALT и FOXO3 в условиях холеста
экспрессии мРНК и белка STAT5 нами не было
за беременных и между экспрессией GALT и ко
обнаружено). На данный момент в литературе
роткой изоформой ПРЛР у интактных крыс.
не встречаются данные о том, какую роль в про
БИОХИМИЯ том 84 вып. 10 2019
СИГНАЛЬНЫЕ ПУТИ ПРОЛАКТИНА
1507
лактиновом сигналинге в почках играют STAT5
препятствует PIAS3, как и в случае с STAT5 [51].
белки. Выделяют две изоформы STAT5
-
PIAS3 играет важную роль в иммунном ответе:
STAT5A и STAT5B, которые кодируются 2 мя
известно, что он контролирует экспрессию та
различными генами [33, 34]. Считается, что
ких транскрипционных факторов, как MITF
STAT5A играет ключевую роль в пролактиновом
(microphthalmia associated transcription factor) и
сигналинге, в то время как STAT5В необходим
NF kB, влияющих на клеточное выживание и
для поддержания полового диморфизма в ско
воспалительный ответ [51, 52]. Возможно, рост
рости роста тела, экспрессии генов печени и
экспрессии SOCS3 и PIAS3, продемонстриро
других функций, связанных с поддержанием
ванный нами при холестазе беременных и в ко
иммунитета [35-39].
ре, и во внешнем мозговом слое почки, может
Помимо снижения экспрессии длинной изо
способствовать провоспалительному статусу
формы ПРЛР в условиях холестаза беременных
почки в условиях этой патологии.
во внешнем мозговом слое по сравнению с груп
пой «холестаз», мы наблюдали рост экспрессии
Полученные нами данные свидетельствуют в
терминаторов пролактинового сигналинга
пользу того, что в реализации эффектов пролак
SOCS3 и PIAS3 в данной экспериментальной
тина в модели холестаза беременных участвуют
группе крыс по сравнению с интактными кры
и короткая, и длинная изоформы ПРЛР во
сами. В коре в условиях холестаза беременных
внешнем мозговом слое почки, а в коре - глав
также возрастает экспрессия как SOCS3, так и
ным образом длинная изоформа. В целом следу
PIAS3 относительно нормы, кроме того, экс
ет заключить, что нами впервые получены дока
прессия SOCS3 возрастает в группе «холестаз»
зательства наличия прямых эффектов пролакти
относительно интактных крыс. Эти транскрип
на на почку: 1) выявлено наличие экспрессии
ционные факторы обладают STAT чувствитель
обеих изоформ ПРЛР и изменение их экспрес
ными элементами в промоторах своих генов, т.е.
сии в зависимости от физиологического состоя
в случае роста STAT5 их экспрессия возрастает,
ния и разного уровня пролактина; 2) показан
и они способствуют подавлению пролактиново
рост фосфорилирования белка STAT5, как клю
го сигналинга через длинную изоформу (в слу
чевого посредника сигналинга длинной изо
чае SOCS3 - и короткую). SOCS3 ингибирует
формы в условиях уменьшении доминирования
проведение сигнала за счет связывания с про
короткой изоформы ПРЛР; 3) продемонстриро
ксимальным сайтом киназы Janus класса 2, с ко
ван рост экспрессии мРНК терминаторов про
торой сопряжены и короткая, и длинная изо
лактинового сигналинга, являющихся молеку
формы ПРЛР [40, 41], а PIAS3 взаимодействует
лярными мишенями длинной изоформы ПРЛР
с ДНК связывающим доменом STAT5, что пре
и имеющих STAT чувствительные элементы в
пятствует связыванию STAT5 с таргетными ге
промоторах своих генов, в условиях преоблада
нами [42, 43]. Известно, что белки семейства
ния длинной изоформы; 4) обнаружено сниже
SOCS, в том числе SOCS3, вовлечены в иммун
ние экспрессии мРНК GALT, как молекулярной
ный ответ, являясь негативными регуляторами
мишени короткой изоформы ПРЛР в условиях
цитокинового сигналинга [18, 44]. Есть данные
относительного роста ее содержания.
о том, что в условиях такой патологии, как вос
Кроме того, следует обратить внимание на
палительное заболевание кишечника (IBD -
то, что в условиях такой патологии, как холес
Inflammatory Bowel Disease), смоделированной
таз, экспрессия изоформ ПРЛР в почках суще
на мышах, был показан рост мРНК SOCS3 по
ственно возрастает по сравнению с нормой. Это
сравнению с нормой [45]. Дело в том, что SOCS3
может быть связано не только с реализацией
ингибирует также JAK2/STAT3 путь, через ко
эволюционно древних осморегуляторных функ
торый проходит сигнал IL 6 и IL 22 в эпители
ций пролактина, но, возможно, имеет отноше
альных клетках тонкого кишечника (данные ин
ние к усилению детоксицирующей функции
терлейкины способствуют пролиферации эпи
почки и к регуляции пролактином ее воспали
телиальных клеток и заживлению ран) [44].
тельного ответа.
Прямые эффекты SOCS3 на сигналинг с участи
ем STAT3 были показаны на эпителиальных
клетках кожи, где избыточная экспрессия
SOCS3 вызывала затрудненное заживление ран,
Конфликт интересов. Авторы заявляют, что у
усиливающее воспаление [46-48]. Известно,
них нет конфликта интересов.
что STAT3 экспрессируется в почке и связан с
Соблюдение этических норм. Все хирургичес
развитием хронических почечных заболеваний,
кие и экспериментальные протоколы были
фиброзом и провоспалительным ответом [49,
одобрены комитетом биоэтики биологического
50]. Связыванию STAT3 с таргетными генами
факультета МГУ.
БИОХИМИЯ том 84 вып. 10 2019
1508
АБРАМИЧЕВА и др.
СПИСОК ЛИТЕРАТУРЫ
1.
Gabzdyl, E.M., and Schlaeger, J.M. (2015) Intrahepatic
form of prolactin receptor, J. Biol. Chem.,
266,
сholestasis of pregnancy, J. Perinat. Neonatal Nurs., 29,
20110-20117.
41-50, doi: 10.1097/JPN.0000000000000077.
16.
Bernard, V., Bouilly, J., Beau, I., Broutin, I., Chanson, P.,
2.
Lo, J.O., Shaffer, B.L., Allen, A.J., Little, S.E., Cheng,
Young, J., and Binart, N. (2016) Germline prolactin recep
Y.W., and Caughey, A.B. (2015) Intrahepatic cholestasis of
tor mutation is not a major cause of sporadic prolactinoma
pregnancy and timing of delivery, J. Matern. Fetal Neonatal
in humans, Neuroendocrinology, 103, 738-745, doi: 10.
Med.,
28,
2254-2258, doi:
10.3109/14767058.2014.
1159/000442981.
984605.
17.
Halperin, J., Devi, S. Y., Elizur, S., Stocco, C., Shehu, A.,
3.
Simjak, P., Parizek, A., Vitek, L., Cerny, A., Adamcova, K.,
Rebourcet, D., and Gibori, G. (2008) Prolactin signaling
Koucky, M., and Starka, L. (2015) Fetal complications due
through the short form of its receptor represses forkhead
to intrahepatic cholestasis of pregnancy, J. Perinat. Med.,
transcription factor FOXO3 and its target gene galt causing
43, 133-139, doi: 10.1515/jpm 2014 0089.
a severe ovarian defect, Mol. Endocrinol., 22, 513-522,
4.
Geenes, V., and Williamson, C. (2009) Intrahepatic
doi: 10.1210/me.2007 0399.
cholestasis of pregnancy, World J. Gastroenterol., 15,
18.
Krebs, D.L., and Hilton, D.J. (2001) SOCS proteins: neg
2049-2066, doi: 10.3748/wjg.15.2049.
ative regulators of cytokine signaling, Stem Cells, 19,
5.
Lee, N.M., and Brady, C.W. (2009) Liver disease in preg
378-387.
nancy, World J. Gastroenterol., 15, 897-906, doi: 10.3748/
19.
Moreno Carranza, B., Goya Arce, M., Vega, C., Adan,
wjg.15.897.
N., Triebel, J., Lopez Barrera, F., and Clapp, C. (2013)
6.
Ranta, T., Unnerus, H.A., Rossi, J., and Seppala, M.
Prolactin promotes normal liver growth, survival, and
(1979) Elevated plasma prolactin concentration in
regeneration in rodents: effects on hepatic IL 6, suppressor
cholestasis of pregnancy, Am. J. Obstet. Gynecol., 134, 1-3.
of cytokine signaling 3, and angiogenesis, Am. J. Physiol.,
7.
Фидченко Ю.М., Кушнарева Н.С., Смирнова О.В.
305, R720-R726, doi: 10.1152/ajpregu.00282.2013.
(2013) Влияние пролактина на водно солевой обмен у
20.
Asad, M., Shewade, D.G., Koumaravelou, K., Abraham,
самок крыс в модели холестаза беременных, Бюлле+
B.K., Balasinor, N., and Ramaswamy, S. (2001) Effect of
тень экспериментальной биологии и медицины, 156,
hyperprolactinaemia as induced by pituitary homografts
767-770.
under kidney capsule on gastric and duodenal ulcers in
8.
Goffin, V., and Touraine, P. (2015) The prolactin receptor
rats, J. Pharm. Pharmacol., 53, 1541-1547, doi: 10.1211/
as a therapeutic target in human diseases: browsing new
0022357011777918.
potential indications, Expert Opin. Ther. Targets, 19,
21.
Ose, K., Miyata, K., Yoshioka, K., and Okuno, Y. (2009)
1229-1244, doi: 10.1517/14728222.2015.1053209.
Effects of hyperprolactinemia on toxicological parameters
9.
Marano, R.J., and Ben Jonathan, N. (2014) Minireview:
and proliferation of islet cells in male rats, J. Toxicol. Sci.,
Extrapituitary prolactin: an update on the distribution, reg
34, 151-162, doi: 10.2131/jts.34.151.
ulation, and functions, Mol. Endocrinol., 28, 622-633,
22.
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van
doi: 10.1210/me.2013 1349.
Roy, N., De Paepe, A., and Speleman, F. (2002) Accurate
10.
Ignacak, A., Kasztelnik, M., Sliwa, T., Korbut, R.A.,
normalization of real time quantitative RT PCR data by
Rajda, K., and Guzik, T.J. (2012) Prolactin not only lac
geometric averaging of multiple internal control genes,
totrophin. A «new» view of the «old» hormone, J. Physiol.
Genome Biol.,
3,
1-12, doi:
10.1186/gb 2002 3 7
Pharmacol., 63, 435-443.
research0034.
11.
Abramicheva, P., Balakina, T., Bulaeva, O., Guseva, A.,
23.
Aleksandrova, M.I., Sirotina, N.S., and Smirnova, O.V.
Lopina, O., and Smirnova, O. (2017) Role of Na+/K+
(2015) Possible recovery of manifestation of prolactin
ATPase in natriuretic effect of prolactin in a model of
receptor and some of its target proteins in the liver and kid
cholestasis of pregnancy, Biochemistry (Moscow), 82,
ney cells of female rats after relief of cholestasis complicat
632-641, doi: 10.1134/S000629791705011X.
ed and not complicated by hyperprolactinemia, Bull. Exp.
12.
Булаева О.А., Абрамичева П.А., Балакина Т.А., Смир
Biol. Med., 159, 361-364, doi: 10.1007/s10517 015 2963 0.
нова О.В. (2016) Роль пролактина в регуляции биоди
24.
Yonezawa, T., Chen, K.H., Ghosh, M.K., Rivera, L., Dill,
намики бикарбонатов у самок крыс в модели холеста
R., Ma, L., and Walker, A.M. (2015) Anti metastatic out
за беременных, Бюллетень экспериментальной биоло+
come of isoform specific prolactin receptor targeting in
гии и медицины, 162, 559-562.
breast
cancer,
Cancer
Lett.,
366,
84-92,
13.
Александрова М.И., Кушнарева Н.С., Смирнова О.В.
doi: 10.1016/j.canlet.2015.06.010.
(2012) Особенности манифестации рецептора пролак
25.
Tan, D., Johnson, D.A., Wu, W., Zeng, L., Chen, Y.H.,
тина в ткани почки самок крыс при холестазе: влия
Chen, W.Y., and Walker, A.M. (2005) Unmodified pro
ние гиперпролактинемии, Бюллетень эксперимен+
lactin (PRL) and S179D PRL initiated bioluminescence
тальной биологии и медицины, 153, 434-437.
resonance energy transfer between homo and hetero pairs
14.
Boudreau, R.T.M., Sangster, S.M., Johnson, L.M., and
of long and short human PRL receptors in living human
Dauphinee, S. (2002) Implication of 4 phosphoprotein and
cells, Mol. Endocrinol., 19, 1291-1303, doi: 10.1210/me.
the rapamycin sensitive mammalian target of rapamycin
2004 0304.
pathway in prolactin receptor signaling, J. Endocrinol.,
26.
Devi, Y.S., Seibold, A.M., Shehu, A., Maizels, E.,
173, 493-506.
Halperin, J., Le, J., and Gibori, G. (2011) Inhibition of
15.
Ali, S., Pellegrini, I., and Kelly, P.A. (1991) A prolactin
MAPK by prolactin signaling through the short form of its
dependent immune cell line (Nb2) expresses a mutant
receptor in the ovary and decidua: involvement of a novel
БИОХИМИЯ том 84 вып. 10 2019
СИГНАЛЬНЫЕ ПУТИ ПРОЛАКТИНА
1509
phosphatase, J. Biol. Chem., 286, 7609-7618, doi: 10.
40.
Tonko Geymayer, S., Goupille, O., Tonko, M., Soratroi, C.,
1074/jbc.M110.166603.
Yoshimura, A., Streuli, C., and Doppler, W.
(2002)
27.
Devi, Y.S., Shehu, A., Stocco, C., Halperin, J., Le, J.,
Regulation and function of the cytokine inducible SH 2
Seibold, A.M., and Gibori, G. (2009) Regulation of tran
domain proteins, CIS and SOCS3, in mammary epithelial
scription factors and repression of Sp1 by prolactin signal
cells, Mol. Endocrinol., 16, 1680-1695.
ing through the short isoform of its cognate receptor,
41.
Tomic, S., Chughtai, N., and Ali, S. (1999) SOCS 1, 2, 3:
Endocrinology, 150, 3327-3335, doi: 10.1210/en.2008
selective targets and functions downstream of the prolactin
1719.
receptor, Mol. Cell. Endocrinol., 158, 45-54, doi: 10.1016/
28.
Elsas, L.J., Lai, K., Saunders, C.J., and Langley, S.D.
S0303 7207(99)00180 X.
(2001) Functional analysis of the human galactose 1
42.
Rycyzyn, M.A., and Clevenger, C.V. (2002) The intranu
phosphate uridyltransferase promoter in Duarte and LA
clear prolactin/cyclophilin B complex as a transcriptional
variant galactosemia, Mol. Genet. Metab., 72, 297-305.
inducer, Proc. Natl. Acad. Sci. USA, 99, 6790-6795,
29.
Bogorad, R.L., Smyslova, V.S., Smirnov, A.N., Rubtsov,
doi: 10.1073/pnas.092160699.
P.M., and Smirnova, O.V. (2002) The ratio of prolactin
43.
Russell, D.L., and Richards, J.S. (1999) Differentiation
receptor isoforms in rat hepatocytes: the effect of obstruc
dependent prolactin responsiveness and stat (signal trans
tive cholestasis, Mol. Biol. (Mosk.)., 36, 91-93.
ducers and activators of transcription) signaling in rat ovar
30.
Bogorad, R.L., Ostroukhova, T.Y., Orlova, A.N., Rubtsov,
ian cells, Mol. Endocrinol., 13, 2049-2064, doi: 10.1210/
P.M., and Smirnova, O.V. (2006) Prolactin receptors in rat
mend.13.12.0389.
cholangiocytes: Regulation of level and isoform ratio is sex
44.
Li, Y., Haar, C., Peppelenbosch, M.P., and van der Woude,
independent, Biochemistry (Moscow),
71,
178-184,
C.J. (2012) SOCS3 in immune regulation of inflammatory
doi: 10.1134/S0006297906020106.
bowel disease and inflammatory bowel disease related can
31.
Ben Jonathan, N., LaPensee, C.R., and LaPensee, E.W.
cer, Cytokine Growth Factor Rev., 23, 127-138, doi: 10.
(2008) What can we learn from rodents about prolactin in
1016/j.cytogfr.2012.04.005.
humans? Endocrinol. Rev., 29, 1-41, doi: 10.1210/er.2007
45.
Suzuki, A., Hanada, T., Mitsuyama, K., Yoshida, T.,
0017.
Kamizono, S., Hoshino, T., and Yoshimura, A. (2001)
32.
Bole Feysot, C., Goffin, V., Edery, M., Binart, N., and
CIS3/SOCS3/SSI3 plays a negative regulatory role in
Kelly, P.A. (1998) Prolactin (PRL) and its receptor:
STAT3 activation and intestinal inflammation, J. Exp.
actions, signal transduction pathways and phenotypes
Med., 193, 471-482, doi: 10.1084/jem.193.4.471.
observed in PRL receptor knockout mice, Endocrinol.
46.
Linke, A., Goren, I., Bo, M.R., Pfeilschifter, J., and
Rev., 19, 225-268.
Frank, S. (2010) The suppressor of cytokine signaling
33.
Hennighausen, L., and Robinson, G.W.
(2008)
(SOCS) 3 determines keratinocyte proliferative and
Interpretation of cytokine signaling through the transcrip
migratory potential during skin repair, J. Invest. Dermatol.,
tion
factors,
Genes
Dev.,
22,
711-721,
130, 876-885, doi: 10.1038/jid.2009.344.
doi: 10.1101/gad.1643908.
47.
Linke, A., Goren, I., Bo, M.R., Pfeilschifter, J., and
34.
Copeland, N.G., Gilbert, D.J., Schindler, C., Zhong, Z.,
Frank, S. (2010) Epithelial overexpression of SOCS 3 in
Wen, Z., Darnell, J.E., and Ihle, J.N. (1995) Distribution
transgenic mice exacerbates wound inflammation in the
of the mammalian Stat gene family in mouse chromo
presence of elevated TGF b 1, J. Invest. Dermatol., 130,
somes, Genomics, 29, 225-228, doi: 10.1006/geno.1995.
866-875, doi: 10.1038/jid.2009.345.
1235.
48.
Ivory, C.P.A., Wallace, L.E., McCafferty, D. M., and
35.
Smirnov, A.N. (2009) Hormonal mechanisms of sex differ
Sigalet, D.L. (2008) Interleukin 10 independent anti
entiation of the liver: the modern concepts and problems,
inflammatory actions of glucagon like peptide 2, Am. J.
Ontogenez, 40, 334-354.
Physiol. Gastrointest. Liver Physiol., 295, G1202-G1210,
36.
Endo, T., Sasaki, A., Minoguchi, M., Joo, A., and
doi: 10.1152/ajpgi.90494.2008.
Yoshimura, A. (2003) CIS1 interacts with the Y532 of the
49.
Ni, J., Shen, Y., Wang, Z., Shao, D.C., Liu, J., Fu, L.J.,
prolactin receptor and supresses prolactin dependent
and Lu, L.M. (2014) Inhibition of STAT3 acetylation is
STAT5 activation, J. Biochem., 133, 109-113.
associated with attenuated renal fibrosis in the obstructed
37.
Matsumoto, A., Seki, Y., Kubo, M., Ohtsuka, S., Suzuki, A.,
kidney, Acta Pharmacol. Sin., 35, 1045-1054, doi: 10.
Hayashi, I., Tsuji, K., Nakahata, T., Okabe, M., Yamada, S.,
1038/aps.2014.42.
and Yoshimura, A. (1999) Suppression of STAT5 functions
50.
Ma, J.Q., Ding, J., Xiao, Z.H., and Liu, C.M. (2014)
in liver, mammary glands, and T cells in cytokine
Ursolic acid ameliorates carbon tetrachloride induced
inducible SH2 containing protein 1 transgenic mice,
oxidative DNA damage and inflammation in mouse kidney
Mol. Cell. Biol., 19, 6396-6407, doi: 10.1128/MCB.19.9.
by inhibiting the STAT3 and NF κB activities, Int.
6396.
Immunopharmacol., 21, 389-395, doi: 10.1016/j.intimp.
38.
Liu, X., Robinson, G.W., Wagner, K.U., Garrett, L.,
2014.05.022.
Wynshaw Boris, A., and Hennighausen, L. (1997) Stat5a is
51.
Yagil, Z., Nechushtan, H., Kay, G., Yang, C.M., Kemeny,
mandatory for adult mammary gland development and lac
D.M., and Razin, E. (2010) The enigma of the role of
togenesis, Genes Dev., 11, 179-186, doi: 10.1101/gad.11.
Protein inhibitor of Activated STAT3 (PIAS3) in the
2.179.
immune response, Trends Immunol., 31, 199-204, doi: 10.
39.
Ormandy, C.J., Camus, A., Barra, J., Damotte, D., Lucas,
1016/j.it.2010.01.005.
B., Buteau, H., and Kelly, P.A. (1997) Null mutation of the
52.
Shuai, K. (2006) Regulation of cytokine signaling pathways
prolactin receptor gene produces multiple reproductive
by PIAS proteins, Cell Res., 16, 196-202, doi: 10.1038/sj.
defects in the mouse, Genes Dev., 11, 167-178, doi: 10.
cr.7310027.
1101/gad.11.2.167.
БИОХИМИЯ том 84 вып. 10 2019
1510
АБРАМИЧЕВА и др.
EXPRESSION OF PROLACTIN RECEPTOR ISOFORMS, MEMBERS
OF PROLACTIN SIGNALLING, AND FACTORS OF RECEPTOR
CYCLE TERMINATION IN THE KIDNEY IN THE CHOLESTASIS
OF PREGNANCY MODEL
P. A. Abramicheva1*, Т. А. Balakina1, I. A. Morozov2,
T. A. Schelkunova1, and O. V. Smirnova1
1 Lomonosov Moscow State University, Faculty of Biology, 199991 Moscow,
Russia; E+mail: abramicheva.polina@gmail.com
2 Engelhardt Institute of Molecular Biology,
Russian Academy of Sciences, 119991 Moscow, Russia
Received January 29, 2019
Revised June 21, 2019
Accepted June 21, 2019
Expression of the long and short isoform of prolactin receptor (PrlR) and mediators of prolactin signaling in cortex
and outer medulla of rat kidney in the model of cholestasis of pregnancy and in control animals was studied. The
direct effects of prolactin on kidney have been proven, since: 1) mRNA expression of both isoforms of PrlR in kidney
changes under different physiological conditions and different levels of prolactin; 2) expression level of pSTAT5 as a
key mediator of signaling of the long PrlR isoform increases in cholestasis of pregnancy; 3) mRNA expression of the
SOCS3 and PIAS3 prolactin signaling terminators increases under predominance of the long isoform; 4) expression
of GALT mRNA as a molecular target of the short PrlR isoform decreases in the outer medulla of kidney.
Keywords: short and long isoforms of prolactin receptor, STAT5, SOCS3, PIAS3, cholestasis of pregnancy, hyperpro
lactinemia, kidney
БИОХИМИЯ том 84 вып. 10 2019