БИОХИМИЯ, 2019, том 84, вып. 11, с. 1592 - 1609
УДК 576
ПОИСК ВНУТРИКЛЕТОЧНЫХ СЕНСОРОВ, ВОВЛЕЧЕННЫХ
В ФУНКЦИОНИРОВАНИЕ ОДНОВАЛЕНТНЫХ КАТИОНОВ
КАК ВТОРИЧНЫХ ПОСРЕДНИКОВ
Обзор
© 2019
Е.А. Климанова1*,**, С.В. Сидоренко1, А.М. Тверской1**, А.А. Шиян1,
Л.В. Смольянинова1**, Л.В. Капилевич2, С.В. Гусакова3,
Г.В. Максимов1, О.Д. Лопина1**, С.Н. Орлов1,2,3
1 Московский государственный университет им. М.В. Ломоносова, биологический факультет,
119234 Москва, Россия; электронная почта: klimanova.ea@yandex.ru
2 Национальный исследовательский Томский государственный университет, 634050 Томск, Россия
3 Сибирский государственный медицинский университет Минздрава России, 634050 Томск, Россия
Поступила в редакцию 03.06.2019
После доработки 15.07.2019
Принята к публикации 16.07.2019
Поддержание неравновесного распределения Na+ и K+ между цитоплазмой и внеклеточной средой предпо
лагает наличие сенсоров, реагирующих конформационными переходами в ответ на изменение внутрикле
точной концентрации этих одновалентных катионов. Молекулярная природа сенсоров одновалентных ка
тионов была установлена при изучении структуры Na,K АТPазы, рецепторов, сопряженных с GTP связы
вающими белками, и белков теплового шока. В последнее время было обнаружено, что изменение внутри
клеточной концентрации Na+ и K+ является ключевым фактором регуляции транскрипции и трансляции
соответственно. В данном обзоре мы суммируем основные результаты этих исследований, а также физиоло
гическое и патофизиологическое значение Na+,Ki+ чувствительного механизма регуляции экспрессии
генов.
КЛЮЧЕВЫЕ СЛОВА: натрий, калий, транскрипция, трансляция.
DOI: 10.1134/S032097251911006X
В своих работах, которые привели к полу
внешнего стимула предшествует формированию
чению Нобелевской премии, Earl Wilbur
функционального ответа клетки и нормализует
Sutherland Jr. сформулировал концепцию, со
ся после окончания действия данного стимула.
гласно которой любая молекула может рассмат
(ii) В отсутствие внешнего стимула изменение
риваться в качестве посредника в процессе пере
внутриклеточной концентрация посредника яв
дачи внеклеточных сигналов внутрь клетки, если
ляется достаточным условием для формирования
она удовлетворяет трем основным критериям.
ответа клетки. (iii) Функциональный ответ клет
(i) Изменение внутриклеточной концентрации
ки опосредован взаимодействием посредника с
молекулы кандидата в ответ на воздействие
его высокоселективными внутриклеточными
Принятые сокращения: AMPK - АМР чувствительная протеинкиназа; CaMK - Ca2+/кальмодулин чувствитель
ная протеинкиназа; COX 2 - циклооксигеназа 2 го типа; CREB (cyclic AMP response element binding protein) - белок,
связывающий cAMP и регулирующий транскрипцию генов; EPS (electrical pulse stimulation) - электрическая пульсовая
стимуляция; ERG (early response genes) - гены раннего ответа; GPCRs (G protein coupled receptors) - рецепторы, сопря
женные с G белками; HIF 1 - фактор, индуцируемый гипоксией 1; HRE (HIF response element) - HIF чувствительный
элемент; HSP - белок теплового шока; NaRE - Na чувствительный элемент, регулирующий транскрипцию генов;
NaREВ - неидентифицированный сенсор изменения [Na+]i, активирующий NaRE; NBD - нуклеотидсвязывающий до
мен; NFAT (nuclear factor of activated T cells) - ядерный фактор активированных Т клеток; NFκB (nuclear factor kappa
light chain enhancer of activated B cells) - ядерный фактор «каппа би» активированных В клеток; NMDA - N метил D ас
партатат; SBD - субстратсвязывающий домен; SERCA (skeletal muscle sarcoplasmic /endoplasmic reticulum Ca ATPase) -
Ca ATPаза саркоплазматического/эндоплазматического ретикулума скелетной мускулатуры; UTR - нетранслируемые
участки мРНК; ГМК - гладкомышечные клетки, КТС - кардиотонические стероиды.
* Адресат для корреспонденции.
** Автор является выпускником кафедры биохимии биологического факультета МГУ им. М.В. Ломоносова.
1592
ОДНОВАЛЕНТНЫЕ КАТИОНЫ КАК ВТОРИЧНЫЕ ПОСРЕДНИКИ
1593
сенсорами [1]. Последующие работы показали,
и/или калием в диапазоне, соответствующем
что этим требованиям удовлетворяют такие низ
диапазону изменения их внутриклеточных кон
комолекулярные соединения, как циклические
центраций (что, по видимому, следует рассмат
нуклеотиды (cAMP, cGMP), продукты катабо
ривать как первое указание на наличие белков,
лизма фосфолипидов (диацилглицерол, инози
содержащих высокоселективные участки связы
тол 1,4,5 трисфосфат), а также ионы Ca2+ [2-5].
вания этих катионов), приведены на рис. 1. На
Как и в случае перечисленных выше канони
ряду с мембранными транспортерами были об
ческих посредников, внутриклеточное содержа
наружены и другие белки, активность которых
ние моновалентных ионов временно изменяется
изменялась при варьировании концентрации
в ответ на действие целого ряда стимулов. Так,
одновалентных катионов [22, 23]. В этой части
например, активация Na+о/H+i обмена и, как
нашего обзора мы суммируем сведения, полу
следствие этого, увеличение [Na+]i отмечено
ченные при изучении структуры участков свя
при действии ростовых факторов во всех изучав
зывания одновалентных катионов.
шихся на этот предмет типах клеток [6-11]. В
Na,K5ATРаза. Несмотря на 70 лет, прошед
эритроцитах млекопитающих, мембранный по
ших с момента идентификации первых селек
тенциал которых близок к равновесному хлор
тивных транспортеров Na+ и К+, молекулярная
ному потенциалу (Em = ECl ~ -(8-12) мВ), акти
структура участков связывания этих катионов
вация Са2+ чувствительных К+ каналов приво
наиболее полно изучена только в случае Na,K
дит к (5-8)× уменьшению [K+]i [12], что в свою
ATРазы.
очередь сопровождается сжатием клеток и эрит
В каталитическом цикле Na,K АТРаза попе
розом
- особой формой программируемой
ременно существует в двух конформационных
смерти эритроцитов [13]. В нейронах и астроци
состояниях, одно из которых имеет высокое
тах короткие периоды синаптической активнос
сродство к ионам Na+ (E1), а другое - к ионам
ти приводят к увеличению локальной концен
K+ (Е2). Сопоставление 3D структуры фермента
трации [Na+]i, от ~10 до 50-100 мM, что объяс
в двух конформациях позволило получить ин
няют входом Na+ через ионотропные глутамат
формацию о центрах связывания этих катионов
ные рецепторы или сопряженные с переносом
(рис. 2). Согласно данным рентгеноструктурно
Na+о транспортеры нейромедиаторов [14, 15].
го анализа в конформации Е1 [24], участки свя
Гипоксия клеток гладкой мускулатуры приводит
зывания Na+ (всего их три: I, II и III) располага
к (2-3)× увеличению [Na+]i, что наряду с при
ются внутри трансмембранной части α субъе
ростом содержания фактора транскрипции, ин
диницы (между трансмембранными α спираля
дуцируемого гипоксией (HIF 1), является при
ми М4, М5, M6 и M8) на глубине ~1/3 от толщи
чиной изменения экспрессии генов [16, 17].
ны мембраны и ближе к ее цитоплазматической
В скелетной мускулатуре физические нагрузки
поверхности. В формировании участков связы
сопровождаются диссипацией трансмембран
вания Na+ основное участие принимают следу
ных градиентов Na+ и K+, что приводит к
ющие аминокислоты α субъединицы фермента:
(5-10)× увеличению соотношения [Na+]i/[K+]i
Ala323, Glu779, Asp804, Asp808, Thr772, Ser775,
[18]. Эти данные, а также результаты селектив
Asn776 (участок I); Val322, Ala323, Val325,
ного влияния Na+, K+, Cl- и HCO3- на актив
Glu327, Asp804 (участок II) и Tyr771, Thr772,
ность ферментов и экспрессию генов позволили
Thr774, Ser775, Asp808, Gln923, Asp926 (участок
в 2006 м году Orlov и Hamet [19] сформулиро
III). Участок I локализован в том же положении,
вать гипотезу, рассматривающую одновалент
что и один из участков связывания K+, участок
ные ионы как вторичные посредники. Задачей
II - на 5 Å ближе к цитоплазматической поверх
данного обзора является рассмотрение новых
ности мембраны. Участки I и II находятся вбли
сведений по этому вопросу с особым вниманием
зи цитоплазматической части трансмембранной
к работам, посвященным идентификации внут
спирали M5 и разделены боковой цепью Ser775.
риклеточных сенсоров одновалентных катио
Участок III стерически ограничен (вдоль М5) и
нов. Сведения о вовлечении одновалентных
слишком мал для иона K+.
анионов в процессы внутриклеточной сигнали
Расстояние между участками I и II для Na+
зации рассмотрены в недавних обзорах [20, 21].
составляет 3,2-3,6 Å, что меньше, чем расстоя
ние между двумя участками связывания К+.
Этой длины, по видимому, достаточно для
ИДЕНТИФИЦИРОВАННЫЕ СЕНСОРЫ
смежного размещения двух ионов натрия (ион
ОДНОВАЛЕНТНЫХ КАТИОНОВ
ный радиус 0,95 Å), но не двух ионов калия
(ионный радиус катиона 1,33 Å). Было предпо
Наиболее изученные мембранные транспор
ложено, что связывание Na+ специфично, пос
теры, активность которых регулируется натрием
кольку для связывания этого катиона достаточ
БИОХИМИЯ том 84 вып. 11 2019
1594
КЛИМАНОВА и др.
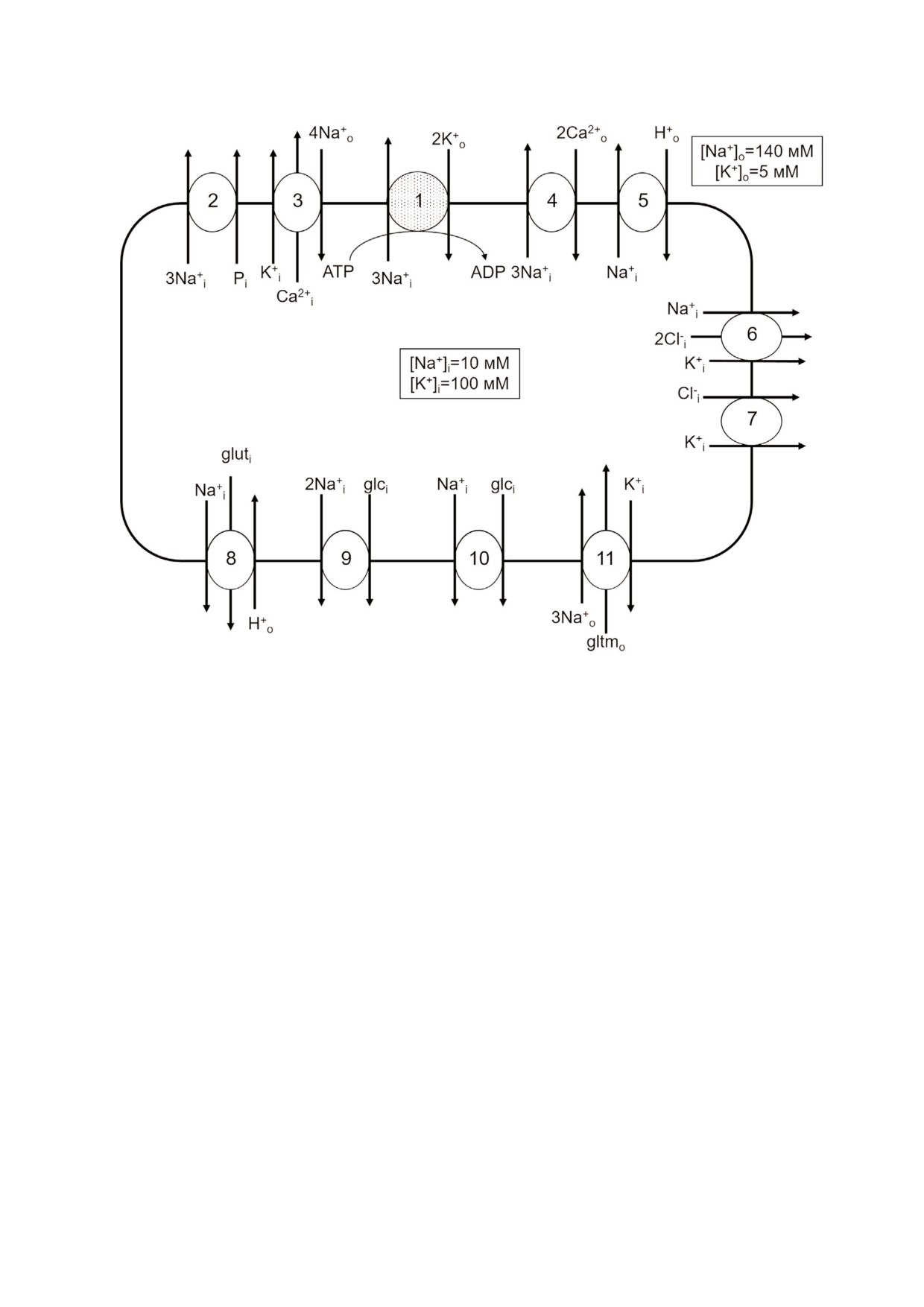
Рис. 1. Мембранные транспортеры, активность которых регулируется внутриклеточным натрием (1, 2, 4-6, 8-10) и кали
ем (3, 6, 7, 11). 1 - Na,K ATPаза; 2 - 3Na+ фосфат котранспорт; 3 - K+ зависимый 4Na+/Ca2+ oбмен (KNCX);
4 - 3Na+/Ca2+ oбмен (NCX); 5 - Na+/H+ oбмен; 6 - Na+,K+,2Cl котранспорт (NKCC); 7 - K+,Cl котранспорт (KCC);
8 - Н+ зависимый Na+,глутамин котранспорт (SNAT3); 9 - 2Na+,глюкоза котранспорт (SLC5A1); 10 - Na+, глюкоза ко
транспорт (SLC5A2); 11 - K+ зависимый 3Na+,глутамат котранспорт. Представлены только те направления функциони
рования транспортеров, которые активируются при увеличении внутриклеточных концентраций Na+ и K+. На примере
плазмы крови и эритроцитов человека приведены значения концентраций натрия и калия во внеклеточных жидкостях и
цитоплазмы покоящейся клетки
но полости с радиусом 2,4 Å, а для связывания
мер полости, достаточной для входа первого ио
К+ необходима полость радиусом 2,8 Å. Именно
на Na+ (но не K+) из цитоплазмы к участку III,
такого размера полость обнаружена в кристалле
будет обеспечен только в том случае, если боко
Na,K АТРазы в конформации Е2 (конформация
вая цепь Ser775 изменяет свое положение. Исхо
с высоким сродством к К+). Участок I наименее
дя из данных рентгеноструктурного анализа,
стерически ограничен, по этой причине он, по
можно полагать, что первый ион Na+ проходит к
видимому, является и К+ связывающим цент
участку III и связывается с ним, но в момент его
ром, поскольку боковые цепи Ser775 и дополни
прохода участки I и II не должны быть оконча
тельно Asn776 участвуют в координации как ио
тельно сформированы (в противном случае Na+
на Na+, так и иона K+. Участки II и III в E1 кон
свяжется с ними). И только после связывания
формации недостаточно велики для связывания
первого иона Na+ с участком III, а второго - с
иона K+. Однако при конформационном пере
участком I, образуется участок связывания II.
ходе Е1 Е2, по видимому, происходит увеличе
Известно, что у другой АТPазы P типа, а
ние полости участка II, и он становится способ
именно у Са АТРазы саркоплазматического ре
ным связывать К+.
тикулума (SERCA, skeletal muscle sarcoplasmic
Для объяснения последовательного и коопе
reticulum/endoplasmic reticulum Ca ATPase, Ca
ративного связывания трех ионов Na+ возможен
ATPaза саркоплазматического/эндоплазмати
следующий сценарий: в Е1 конформации раз
ческого ретикулума скелетных мышц (SERCA))
БИОХИМИЯ том 84 вып. 11 2019
ОДНОВАЛЕНТНЫЕ КАТИОНЫ КАК ВТОРИЧНЫЕ ПОСРЕДНИКИ
1595
участки связывания Са2+ формируются в основ
зывания К+, в отличие от участков связывания
ном теми же аминокислотными остатками, что
Ca2+ в SERCA, расположены на крайне неболь
и в случае катион связывающих участков Na,K
шом расстоянии друг от друга, хотя ионный ра
АТРазы [25]. Важное значение для селективнос
диус K+ гораздо больше, чем у Ca2+ (1,353 и
ти Na,K АТРазы в отношении Na+ играет ори
0,99 Å соответственно) (рис. 3). Предполагается,
ентация цитоплазматической части трансмем
что такое близкое расположение возможно
бранной спирали М5: если спираль М5 выпря
только в случае участков связывания одновалент
мить так же, как в Ca ATPазе, то связывание К+
ных катионов и обусловлено тем, что в обоих
в участке III (исходя из данных рентгенострук
ферментах для координации катионов исполь
турного анализа) представляется возможным.
зуются карбонильные атомы кислорода основ
Действительно, удаление С концевых а.о. в α
ной цепи а.о.. Во вторых, сравнение структур
субъединице Na,K АТРазы, увеличивающих
Na,K ATРазы и SERCA показывает, что у них
подвижность спирали М5, приводит к уменьше
довольно сильно различается координационная
нию сродства к Na+ и к связыванию К+ в участ
геометрия связанных катионов, что свидетель
ке III.
ствует о потенциально низком сродстве однова
Na,K АТРаза относится к семейству АТРаз
лентных катионов к участкам связывания ионов
Р типа, для которых характерна существенная
в SERCA. Кроме того, рассчитанные значения
гомология в первичной последовательности,
валентности мало различаются между Na+ и K+,
сходство в третичной структуре каталитических
свидетельствуя о том, что эти участки связыва
субъединиц, а также в механизме гидролиза АТР
ния не очень селективны по отношению к K+.
и транспорта ионов. Именно по этой причине
Можно заключить, что эти данные еще раз
Toyoshima et al. [26] подробно обсуждают воп
подтверждают идею о том, что Na,K ATРаза -
рос, почему Na,K ATРаза способна связывать
это в первую очередь Na+ насос. В третьих, по
K+ и некоторые другие одновалентные катионы,
ложение и конформация боковых цепей а.о.
в то время как SERCA кроме Са2+ связывает
Na,K ATРазы очень похожи на положения
только H+. На основе данных рентгеноструктур
Ca2+ связывающих участков SERCA, за исклю
ного анализа Na,K ATРазы в E2 конформации
чением Asn783. Этот а.о. Na,K ATРазы форми
с разрешением 2,4 Å можно сделать выводы,
рует водородную связь с Tyr854 на изогнутой
представленные ниже. Во первых, участки свя
части спирали М7, спираль М5 также частично
Рис. 2. Атомарная модель Na+ связывающих участков Na,K ATPазы, представленная перпендикулярно мембране со сто
роны цитоплазмы (a) и параллельно мембране со стороны М6 спирали (б). Обозначения аминокислотных остатков кур
сивом указывают на то, что карбонил главной цепи способствует координации Na+. Зеленые пунктирные линии означа
ют вероятную координацию Na+, оранжевые линии - потенциальные водородные связи (при условии, что карбоксиль
ные группы протонированы). Использованы данные, представленные в статье Kanai et al. [24].
biokhsm/
БИОХИМИЯ том 84 вып. 11 2019
1596
КЛИМАНОВА и др.
Рис. 3. Атомарная модель К+ связывающих участков Na,K ATPазы, представленная со стороны цитоплазмы вдоль
М5 спирали (а) и параллельно мембране (б). Сплошные фиолетовые линии обозначают боковые и основные цепи
Ca2+ координирующих аминокислотных остатков SERCA. Фиолетовые сферы обозначают ионы K+, красные - молеку
лы воды. Голубые сферы показывают положения ионов Ca2+, связанного с SERCA. Двусторонняя стрелка на (б) изобра
жает стерическое столкновение, возникающее между участком связывания K+ I и амидом Asn768 SERCA, которому соот
ветствует Asn783 Na,K ATPазы. Использованы данные, представленные в статье Toyshima et al. [26].
biokhsm/
разворачивается и образует изгиб. Этот излом
ство мембранных белков, которые являются ми
создается благодаря наличию Pro785, который
шенями для трети фармакологических препара
имеет решающее значение для создания прост
тов. 826 этих рецепторов, обнаруженных в гено
ранства и координации К+ за счет взаимодей
ме человека, разделены на 4 семейства (A, B, C и
ствия с карбонильным кислородом основной
F), первичные структуры представителей этих
цепи Thr799. В структуре SERCA место Pro785
семейств сильно различаются. В середине 70 x
занимает Gly770, это приводит к меньшей изог
годов Pert et al. [27] обнаружили, что в гомогена
нутости спирали М5 и, соответственно, к мень
тах мозга увеличение
[Na+]i в диапазоне
шему размеру свободной полости. В результате
10-100 мМ уменьшает связывание агонистов
амид Asn768 SERCA (аналог Asn783 Na,K
опиоидных рецепторов, относящихся к семей
ATPазы) оказывается очень близко к первому
ству А, не оказывая существенного влияния на
участку связывания К+ и к боковой цепи участ
связывание антагонистов этих же рецепторов.
ка II, что не позволяет ионам K+ связаться с эти
Впоследствии это явление было обнаружено и
ми участками SERCA. Все описанное выше по
для адренэргических рецепторов, а также рецеп
казывает, что участки связывания одновалент
торов аденозина, дофамина, брадикинина и ря
ных катионов формируются хотя и похожими
да других GPCRs [28, 29]. При исследовании му
аминокислотными остатками, но они образуют
тированных форм рецепторов было установле
ся только при формировании третичной струк
но, что для появления Na+ зависимой регуля
туры белка, и небольшое изменение этой струк
ции необходимо присутствие аспарагиновой
туры существенно изменяет структуру катион
кислоты во втором трансмембранном домене
связывающих центров.
GPCRs. Эти исследования, а также кристалло
Рецепторы, сопряженные с GTP5связывающи5
графия высокого разрешения позволили уста
ми белками. Рецепторы, сопряженные с GTP
новить структуру высококонсервативного Na+
связывающими белками (G protein coupled
связывающего участка, расположенного внутри
receptors, GPCRs) - самое большое суперсемей
центрального кластера трансмембранного доме
БИОХИМИЯ том 84 вып. 11 2019
ОДНОВАЛЕНТНЫЕ КАТИОНЫ КАК ВТОРИЧНЫЕ ПОСРЕДНИКИ
1597
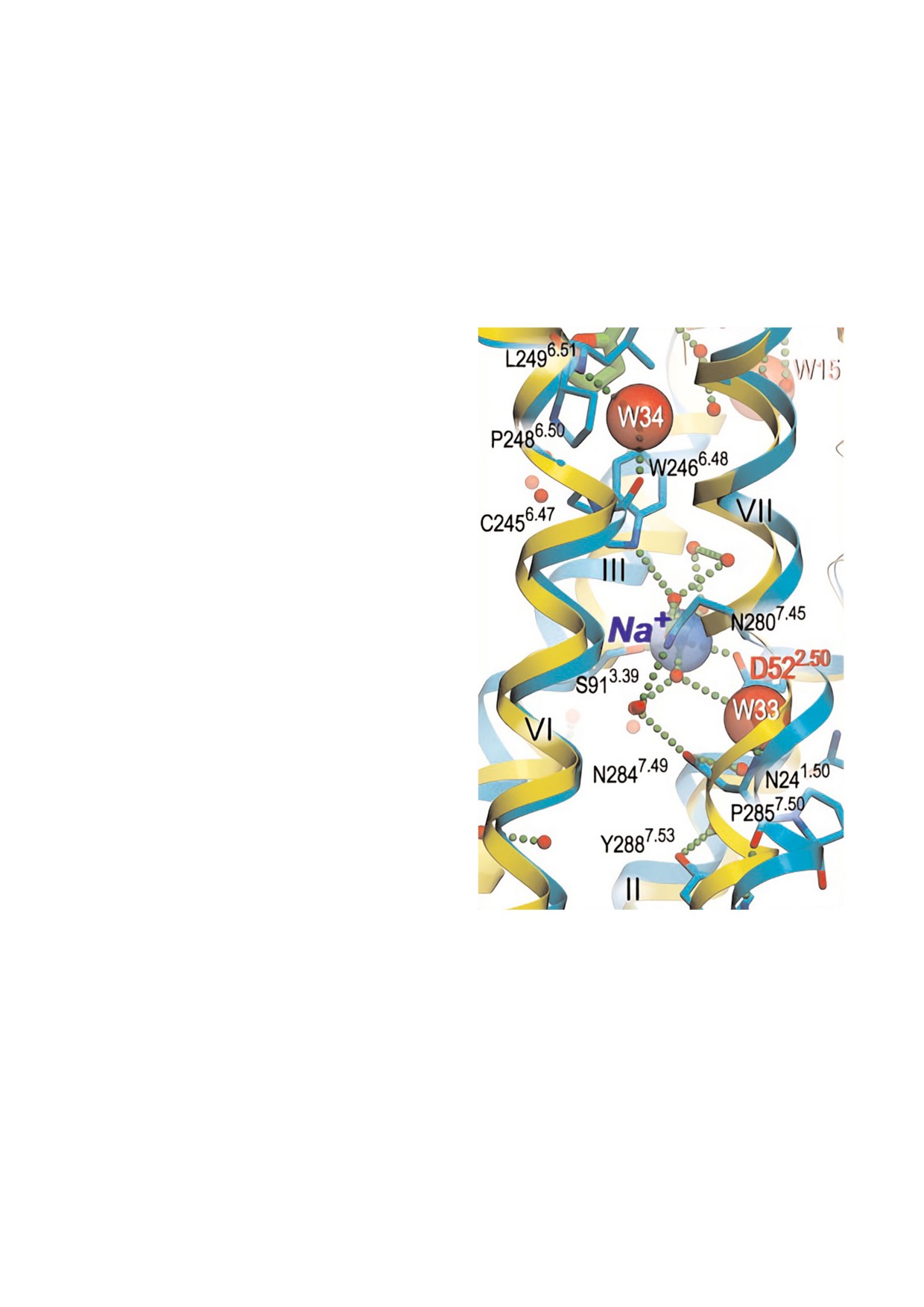
на, состоящего из 7 спиралей (рис. 4). Этот
Asp10 и основной цепью Tyr15 (рис. 5). Кроме
кластер представляет собой часть канала, распо
того, было установлено, что при замене K+ на
ложенного между участками связывания лиган
Na+ последний также координируется ADP,
да и G белка, заполненного молекулами воды. В
Asp10, Tyr15 и находится в том же положении,
канале имеется два узких «горлышка», в кото
что и K+ [34].
рых ширина канала уменьшается за счет Trp246
Приведенные данные показывают принци
и Tyr288 до диаметра чуть меньшего, чем диа
пиальное различие структурной организации
метр молекулы воды (2,4 и 2,0 Å соответствен
участков связывания одновалентных катионов и
но). Связывание лиганда изменяет конформа
Са2+. В самом деле, Са2+ связывающие участки
цию рецептора, что приводит к перекрыванию
канала в этих участках.
В центральном кластере канала, ограничен
ном Trp246 и Tyr288 (расстояние между ними
13 Å), находится ион Na+ и 10 молекул воды, ко
торые целиком заполняют полость кластера.
Этот катион координирован пятью атомами
кислорода от Asp52, Ser91 и 3 х молекул воды.
Кроме того, Na+ формирует водородные связи с
высококонсервативными а.о., включая Asn24,
Trp246, Asn280, Asn284 и Tyr288 [30].
Поскольку доступ Na+ в центральный клас
тер канала возможен с обеих сторон мембраны,
возникает вопрос о том, внеклеточный или
внутриклеточный Na+ необходим для регуляции
активности рецептора. Аллостерическое влия
ние Na+ на связывание лигандов известно по ре
зультатам многих работ. Следует, однако, отме
тить, что в большинстве изученных GPCRs Na+
ингибирует связывание агонистов с IC50 ~50 мM
[29, 31, 32]. Исходя из этих наблюдений, а также
того факта, что значение [Na+]о = 140 мM, роль
внеклеточного Na+ в регуляции активности
GPCRs представляется проблематичной. Воп
рос о вкладе внутриклеточного Na+ в функцио
нирование GPCRs остается нерешенным.
Белки теплового шока. Члены семейства бел
ков теплового шока с молекулярной массой
70 кДа (Hsp70) консервативны почти во всех
живых организмах и вовлечены в регуляцию
многих внутриклеточных процессов, включая
правильное сворачивание синтезирующихся
и/или денатурированных белков для предотвра
щения их агрегации. Hsp70 обладает АТР зави
симой шаперонной активностью. Было установ
лено, что ATРазная активность Hsp70 резко воз
Рис. 4. Центральный кластер трансмембранного домена,
растает в ответ на увеличение концентрации K+
состоящего из 7 спиралей (они отмечены римскими циф
в диапазоне 20-100 мM, в то время как Na+, Li+
рами), рецептора, сопряженного с GTP связывающими
и Cs+ не оказывают существенного влияния на
белками. Структура, связанная с антагонистом, показана
АТРазную активность [33].
синим цветом, структура активного состояния, связанная
с агонистом, - желтым цветом. Молекулы воды представ
Hsp70 состоит из N концевого, нуклеотид
лены в виде красных сфер, а солевые мостики и водород
связывающего домена (NBD), и С концевого,
ные связи - в виде маленьких зеленых сфер. Ион Na+ обо
субстрат связывающего домена (SBD). Данные
значен голубой прозрачной сферой, молекулы воды W33 и
кристаллической структуры NBD Hsp70
W34, стабилизирующие изломы в спиралях VI и VII, -
красными полупрозрачными сферами. Использованы дан
Escherichia coli, связанного с ADP, показали, что
ные, представленные в статье Liuetal et al. [30].
K+ связывающий участок располагается в этом
С цветным вариантом рис. 4 можно ознакомиться в элек
домене. При связывании ион K+ координирует
ся β фосфатной группой ADP, боковой цепью
journal/biokhsm/
БИОХИМИЯ том 84 вып. 11 2019
1598
КЛИМАНОВА и др.
Рис. 5. Взаимодействие между нуклеотид связывающим доменом (NBD) Hsp70 (E. coli) и ADP при отсутствии иона Mg2+.
Ион K+ представлен в виде серой сферы, координационные связи отмечены красными пунктирными линиями. Исполь
зованы данные, представленные в статье Arakawa et al. [34].
biokhsm/
парвальбумина, кальмодулина, тропонина С,
аорты крысы при ингибировании Na,K АТРазы
кальбиндина и ряда других белков содержат так
как уабаином, так и отсутствием в среде инкуба
называемый «EF hand» мотив, организованный
ции К+, сопровождается замедлением апоптоза,
последовательностью из 14 аминокислот [35, 36].
вызванного устранением ростовых факторов
Приведенные выше примеры показывают, что
[40]. Антиапототическое действие ингибиторов
участки связывания одновалентных катионов
Na,K АТРазы было также продемонстрировано
образованы далеко отстоящими друг от друга
на культуре клеток эпителия проксимального
аминокислотами, геометрия которых контроли
отдела нефрона [41], нейронов головного мозга
руется третичной структурой белков. Эта особен
[42] и эндотелия сосудов [43]. В дальнейших
ность не позволяет провести прямое сканирова
экспериментах мы обнаружили, что ингибиро
ние библиотек кДНК с целью выявления потен
вание Na,K АТРазы в ГМК сопровождается
циальных сенсоров Na+ и K+, что было успешно
резким усилением синтеза РНК [44] и появле
использовано для выявления набора Са2+ связы
нием большего количества вновь синтезирован
вающих белков [37]. Следует также отметить, что
ных белков [45-47], что предполагало экспрес
в отличие от белков, содержащих «EF hand» мо
сию генов раннего ответа (early response genes,
тив и насыщающихся при [Ca2+] ~1 мкM, срод
ERG). В самом деле, обработка клеток уабаином
ство участков связывания одновалентных катио
приводила к резкому увеличению экспрессии
нов изменяется в пределах 10-100 мМ, что зат
c-Fos и c-Jun - одних из наиболее изученных
рудняет использование радиоактивных изотопов
представителей семейства ERG [47]. Так как за
для идентификации белков, связывающих одно
щита от апоптоза ингибиторами Na,K ATPазы
валентные катионы в лизатах клеток.
устранялась в присутствии блокаторов синтеза
РНК (актиномицина D) и белка (циклогексами
да) [48], мы предположили, что этот эффект
НАТРИЙ КАК РЕГУЛЯТОР
опосредован регуляцией транскрипции и/или
ТРАНСКРИПЦИИ
трансляции.
Данные, полученные в последние два деся
Сжатие и набухание клеток рассматривают
тилетия, показали, что уабаин и другие кардио
ся как универсальные маркеры двух различных
тонические стероиды (КТС) способны генери
типов клеточной смерти - апоптоза и некроза
ровать внутриклеточные сигналы независимо от
соответственно [38, 39]. Для выяснения меха
ингибирования Na,K ATPазы [49, 50]. Учиты
низма апоптоза мы сопоставили влияние инги
вая это обстоятельство, мы использовали техно
биторов ион транспортирующих систем на из
логию Affymetrix и сравнили действие уабаина и
менения объема и жизнеспособность клеток. К
бескалиевой среды (как двух независимых под
своему удивлению мы обнаружили, что длитель
ходов ингибирования Na,K ATPазы) на тран
ная инкубация гладкомышечных клеток (ГМК)
скриптом ГМК, а также клетки эндотелия чело
БИОХИМИЯ том 84 вып. 11 2019
ОДНОВАЛЕНТНЫЕ КАТИОНЫ КАК ВТОРИЧНЫЕ ПОСРЕДНИКИ
1599
века (human umbilical vein endothelial cells,
5' нетранслируемом участке промотора (5' UTR)
HUVEC) и линии HeLa [51]. При действии ин
гена.
гибитора уабаина или при отсутствии K+, акти
В ряде типов клеток долговременное инги
ватора Na,K ATPазы, изменялась экспрессия
бирование Na,K АТРазы сопровождается при
2+
≥500 генов с высоко достоверной положитель
ростом [Ca2+]i вследствие активации Na+
i
/Ca
o
ной корреляцией, что указывало на диссипацию
обмена и и/или деполяризации мембраны и вхо
ионных градиентов и увеличение соотношения
да Са2+ через потенциал зависимые Са каналы.
[Na+]i/[K+]i как универсальный механизм регу
Так, например, в нашей лаборатории было уста
ляции транскрипции. Было установлено, что 80
новлено, что добавление уабаина сопровождает
Na+i/K+i чувствительных транскриптов генов яв
ся активацией Nai+/Ca2+ обмена в фибробластах
ляются общими как для этих 3 х типов клеток
легких человека [58] и потенциал зависимых
[51], так и для нейронов головного мозга крысы
Са каналов L типа в ГМК аорты крысы [59]. В
и культуры миотубул мыши С2С12 [52], причем
этой связи можно было бы предположить, что
в списке этих универсальных Na+i/K+i чувстви
изменения транскриптома в ответ на увеличе
тельных генов много ERG, включая Fos, FosB,
ние соотношения [Na+]i/[K+]i связаны с прирос
Jun, JunB, Atf3 и Egr-1.
том [Ca2+]i и активацией перечисленных выше
Уже в первых экспериментах мы установили,
Са2+ зависимых механизмов регуляции тран
что защита от апоптоза уабаином сохраняется в
скрипции. Эта гипотеза, однако, противоречит
среде с повышенным содержанием К+, но устра
данным, полученным с использованием хелато
няется при выравнивании трансмембранного
ров кальция. В самом деле, хелаторы внеклеточ
градиента Na+ [40, 53]. Эти результаты указыва
ного Ca2+ и внутриклеточного Ca2+ (EGTA и
ют, что антиапоптотическое действие уабаина
BAPTA AM соответственно) не оказывали вли
обусловлено приростом Na+i, а не потерей K+i.
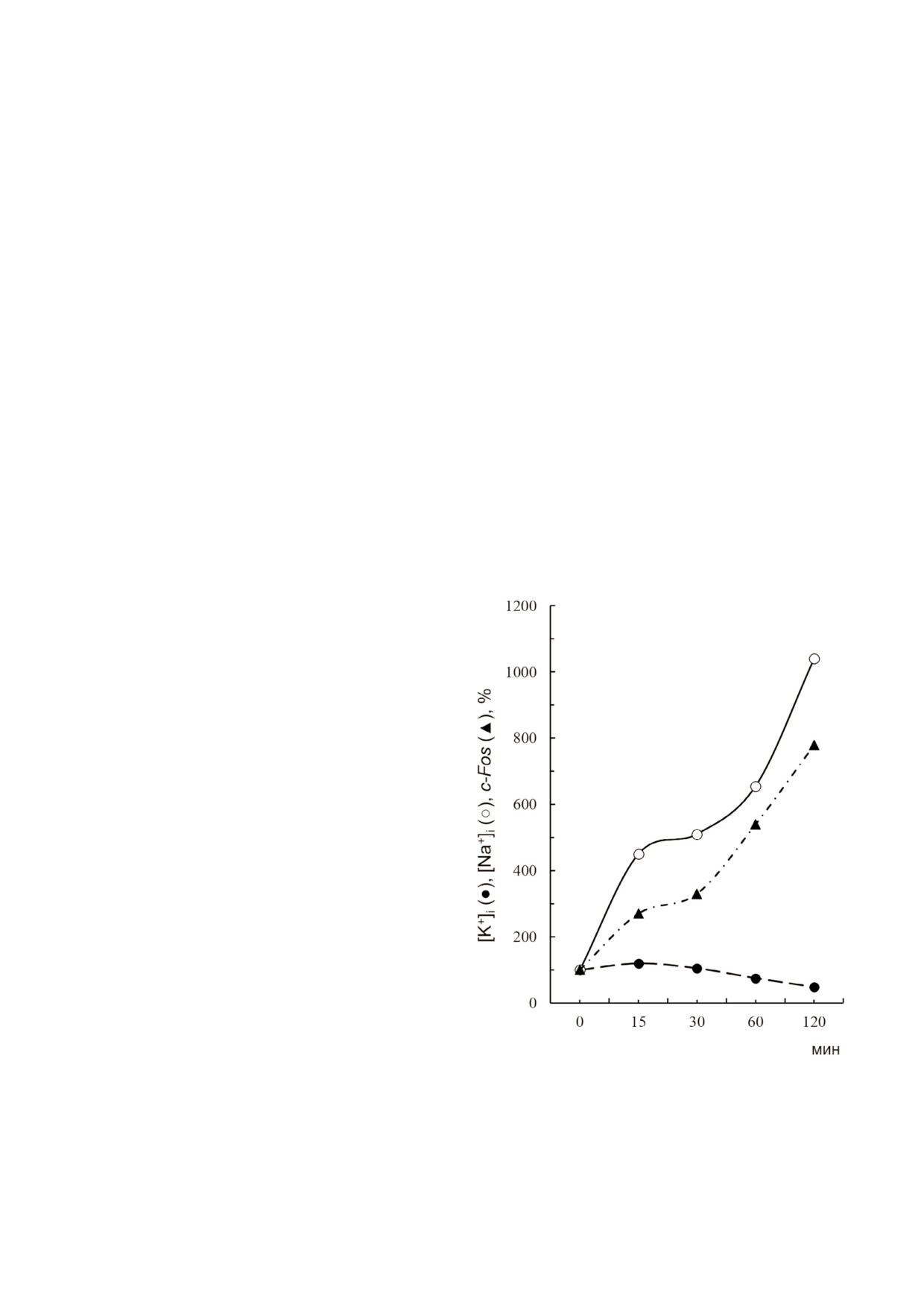
яния на прирост содержания c-Fos в ГМК, выз
Вывод о ведущей роли увеличения Na+i согласу
ванный добавлением уабаина [47]. Более того,
ется с данными по сравнению кинетики
действия уабаина на экспрессию ERG и содер
жание Na+i и K+i в ГМК [47]. В самом деле, 4×
снижение содержания мРНК c-Fos было обна
ружено через 30 мин после добавления уабаина,
когда содержание внутриклеточного Na+ воз
растало 5×, а содержание K+i уменьшалось на
10-15% (рис. 6).
Доказательство Са2+5независимого механизма
проведения сигнала. В многочисленных работах
было обнаружено, что увеличение концентра
ции внутриклеточного кальция ([Ca2+]i) сопро
вождается активацией нескольких сигнальных
систем, лежащих в основе механизма, получив
шего название excitation transcription coupling
[54-57]. Во первых, увеличение [Ca2+]i приво
дит к активации ряда изоформ Ca2+/кальмоду
лин чувствительной протеинкиназы (CaMK),
фосфорилированию ингибитора IkB и трансло
кации NFkB (nuclear factor kappa light chain
enhancer of activated B cells) из цитоплазмы в яд
ро за счет высвобождения фосфо IkB. Во вто
рых, увеличение [Ca2+]i сопровождается дефос
форилированием (Ca2+ кальмодулин) зависи
мой фосфатазы (кальцинейрин), ядерного фак
тора активированных Т клеток (nuclear factor of
activated T cells, NFAT) и его транслокацией в
ядро. В третьих, увеличение [Ca2+]i приводит к
фосфорилированию регулятора транскрипции
Рис. 6. Кинетика изменения содержания внутриклеточно
CREB (cyclic AMP response element binding pro
го калия ([K+]i), натрия ([Na+]i) и мРНК с Fos в клетках
tein). Транслоцированные в ядро NFkB, NFAT и
гладкой мускулатуры аорты в ответ на добавление 1 мМ уа
CREB взаимодействуют с соответствующими
баина. Использованы данные, представленные в статье
участками связывания, локализованными в
Taurinetal et al. [47]
БИОХИМИЯ том 84 вып. 11 2019
1600
КЛИМАНОВА и др.
было обнаружено, что нагрузка клеток хелато
Са2+ хелаторы могут оказывать влияние на
рами кальция увеличивает, а не уменьшает чис
транскрипцию генов через связывание полива
ло генов, экспрессия которых изменяется в от
лентных катионов, поскольку сродство EGTA и
вет на ингибирование Na,K АТРазы [51]. Среди
BAPTA к Mn2+, Zn2+, Cu2+, Co2+ и Fe2+/3+ на нес
универсальных Na+i/K+i чувствительных генов,
колько порядков выше, чем к Ca2+. По этой при
чья экспрессия изменялась более чем 3× в при
чине хелатирование ионов кальция сопровож
сутствии Са2+ хелаторов во всех исследовавших
дается изменением активности ряда ферментов,
ся на этот предмет клетках, были идентифици
регулируемых поливалентными катионами, в
рованы транскрипционные факторы (Fos, Jun,
том числе многочисленных факторов регуляции
Hes1, Nfkbia), интерлейкин 6 (Il-6) и циклоок
транскрипции, содержащих Zn2+ связывающие
сигеназа 2 го типа (СОХ-2 или Ptgs2).
домены [62]. Дополнительным фактом, свиде
При анализе этих данных необходимо отме
тельствующим о непрямом влиянии хелаторов
тить, что хелаторы кальция могут влиять на
Са2+ на метаболизм клетки, в 1981 г. явилась ра
функционирование клеток независимо от их
бота Krishtal et al. [63], в которой показано рез
обеднения кальцием. Так, например, мы обна
кое возрастание проводимости кальциевых ка
ружили, что добавление EGTA к ГМК приводит
налов при хелатировании Са2+.
к возрастанию проницаемости сарколеммы для
В поисках альтернативных подходов для вы
одновалентных катионов [60] и ~10× увеличе
яснения механизма регуляции транскрипции
нию соотношения [Na+]i/[K+]i [59]. В этой связи
Na+i чувствительных генов мы использовали ин
не удивителен тот факт, что содержание 2071
гибиторы Са2+ чувствительных сигнальных сис
транскриптов в ответ на ингибирование Na,K
тем. В этих работах было установлено, что уве
ATPазы изменялось с высокой степенью досто
личение экспрессии Egr1 и Atf3 в ГМК, обрабо
верности (R2 > 0,51; p < 10-12) и в ответ на добав
танных уабаином, не зависит от присутствия
ление Са2+ хелаторов [61]. Кроме изменения
ингибиторов потенциал зависимых Са2+ кана
проницаемости плазматической мембраны
лов и Na+/Ca2+ обмена, а также антагонистов
Рис. 7. Механизмы вовлечения одновалентных катионов и кальция в регуляцию транскрипции Egr1, Atf3, Ptgs2 и Nr4a1 в
клетках гладкой мускулатуры аорты крысы. 1 - Na,K ATPаза; 2 - потенциал зависимые Са2+ каналы; 3 - Na+/Ca2+ об
менник; СаМ - кальмодулин; CaN - кальцинейрин; NaRE - идентифицированный Na+ чувствительный элемент, регу
лирующий транскрипцию генов; и NaREB - не идентифицированный сенсор изменения [Na+]i, активирующий NaRE.
Остальные сокращения приведены в тексте
БИОХИМИЯ том 84 вып. 11 2019
ОДНОВАЛЕНТНЫЕ КАТИОНЫ КАК ВТОРИЧНЫЕ ПОСРЕДНИКИ
1601
кальмодулина и ингибиторов Ca2+/кальмоду
Поиск сенсора, вовлеченного в регуляцию
лин зависимых протеинкиназ и фосфатаз, в то
транскрипции Na+i5чувствительных генов. Рас
время как увеличение экспрессии Ptgs2 и Nr4a1
смотренные выше негативные результаты могут
уменьшалось в присутствии ингибиторов Са2+
быть объяснены следующими альтернативными
зависимой протеинкиназы (CaMKII) и фосфата
механизмами, изложенными ниже. (i) Было об
зы (кальценейрин) соответственно (рис. 7) [59].
наружено, что при относительно низких значе
Совокупность данных, рассмотренных в этом
ниях [Ca2+]i (~100 нM), Na+ взаимодействует с
разделе, свидетельствует об участии Ca2+ неза
Са2+ связывающими центрами кальпаина, вы
висимого механизма в регуляции транскрипции
зывая полумаксимальную активацию этого фер
генов, экспрессия которых изменяется при ин
мента при [Na+]i = 15 мM [66]. Роль Са2+ связы
гибировании Na,K АТРазы и действии других
вающих белков в регуляции транскрипции за
стимулов, приводящих к увеличению [Na+]i.
счет их взаимодействия с одновалентными ка
Канонические 5'5UTR промоторы не вовлече5
тионами остается неизученной. (ii) Фрагмент
ны в регуляцию транскрипции Na+i5чувствитель5
5' UTR, ответственный за Na+i опосредованную
ных генов. К настоящему времени наиболее изу
регуляцию транскрипции c-Fos, находится на
чен механизм регуляции экспрессии генов,
отдаленном от канонического промотора участ
опосредованный взаимодействием ERG и дру
ке. В самом деле, установлено, что реорганиза
гих факторов регуляции транскрипции с участ
ция хроматина влияет на c-Fos через участок,
ками связывания, локализованными в нетранс
граничащий с основанием (-1900) 5' UTR [67].
лируемой 5' UTR последовательности. Исходя
В ряде генов выявлены и более отдаленно лока
из этого, мы предприняли попытку идентифи
лизованные тканеспецифичные альтернатив
цировать Na+ чувствительный элемент регуля
ные промоторы (сложные промоторы, состоя
ции транскрипции (Na+ response element, NaRE)
щие из двух или более независимо функциони
в последовательности 5' UTR Na+ чувствитель
рующих частей, которые расположены перед
ного гена c-Fos. Правомочность такого подхода
одним или разными экзонами гена и обеспечи
подкреплялась данными об отсутствии влияния
вают образование на одном гене молекул мРНК
уабаина на содержание мРНК c-Fos человека,
разных размеров). Так, в случае коактиватора
который был трансфицирован в фибробласты
рецептора активаторов пролиферации перокси
мыши NIH
3T3 после удаления участка
сом Ppargc1a (Ppargc1a) в скелетных мышцах и
(-222)-(-70), входящего в структуру 5' UTR
нервной ткани обнаружены альтернативные
этого гена [64]. Учитывая результаты этих иссле
промоторы, которые расположены еще даль
дований, мы трансфицировали клетки HeLa лю
ше - на 14 й и 587 й тысячах пар нуклеотидов
циферазой под контролем протяженного участ
выше канонического промотора соответственно
ка 5' UTR c-Fos, который ограничен основани
[68, 69]. (iii) Наряду с 5' UTR транскрипция мо
ем (-650) и содержит все наиболее изученные
жет регулироваться последовательностями, рас
элементы промотора, активируемые ростовыми
положенными в интронах или 3' UTR. Такого
факторами и другими каноническими регулято
рода возможности установлены и при изучении
рами транскрипции (рис. 8). В качестве положи
транскрипции c-Fos [70, 71]. В этой связи в на
тельного контроля мы использовали клетки, об
шей лаборатории проводится скрининг генов,
работанные набором ростовых факторов, содер
транскрипция которых изменяется на началь
жащихся в сыворотке. Как и в случае гладкомы
ных этапах диссипации трансмембранного гра
шечных клеток [47], ингибирование Na,K
диента одновалентных катионов. Мы надеемся,
АТРазы в клетках HeLa как уабаином, так и от
что этот подход позволит нам идентифициро
сутствием внеклеточного К+ сопровождается
вать их общие Na+ чувствительные участки
приростом экспрессии c-Fos (рис. 8, а). Тем не
(NaRE) и на основании этих данных установить
менее в отличие от эндогенного c-Fos экспрес
природу Na+ чувствительных белков (NaREВ),
сия люциферазы не регулировалась уабаином,
взаимодействующих с этими участками. (iv) К
но увеличивалась при добавлении ростовых
настоящему времени накоплено большое коли
факторов сыворотки (рис. 8, б) [65]. Получен
чество данных о регуляции экспрессии генов
ные результаты указывают на то, что прирост
при изменении их расположения внутри ядра
содержания c-Fos мРНК в ответ на ингибирова
(т.н. трехмерная модель регуляции транскрип
ние Na,K АТРазы не опосредован участком 5'
ции) [72]. В самом деле, на стадии интерфазы
UTR, взаимодействующим с классическими ре
наиболее конденсированная часть хроматина
гуляторами транскрипции. Альтернативные
(гетерохроматин) находится на периферии ядра,
Са2+ независимые механизмы регуляции Na+i
в то время как менее конденсированная часть
чувствительного транскриптома рассмотрены в
хроматина (эухроматин) локализована внутри
следующем разделе.
ядра [73]. Именно внутри эухроматина находит
6 БИОХИМИЯ том 84 вып. 11 2019
1602
КЛИМАНОВА и др.
Рис. 8. Изменение содержания в клетках HeLa мРНК эндогенного с Fos (a) и трансфецированной плазмиды с люцифе
разой под контролем промотора 5’ UTR c-Fos (б) при добавлении ростовых факторов (FBS) и ингибировании Na,K
ATPазы уабаином (ouabain) и в среде, не содержащей калия (K+ free). Значения, полученные в отсутствии указанных сти
мулов, приняты за 100%. Использованы данные, приведенные в статье Haloui et al. [65]
ся основное количество «фабрик транскрип
увеличением соотношения [Na+]i/[K+]i, c изме
ции» (transcription factories), которые визуализи
нением экспрессии генов [75].
руются по включению меченного флуоресцент
ной меткой уридин 5' трифосфата [74]. Пере
группировка гетеро и эухроматина контролиру
КАЛИЙ КАК РЕГУЛЯТОР ТРАНСЛЯЦИИ
ется такими эпигенетическими факторами как
метилирование ДНК и метилирование и ацети
50 лет назад было установлено, что синтез
лирование хроматина. Роль увеличения соотно
белка в экстрактах клеток прокариот снижается
шения [Na+]i/[K+]i в регуляции пространствен
при уменьшении концентрации K+ [76]. Позд
ной организации гетеро и эухроматина остается
нее этот факт был обнаружен в клетках эукариот
недостаточно исследованной. Однако в послед
животного и растительного царства (см. обзор
нее время появилась информация о том, что ионы
Orlov и Hamet [19]). В этой связи неудивитель
К+ и Na+ по разному стабилизируют четырехспи
но, что в большинстве изученных типов клеток,
ральные, богатые гуанином структуры, так на
включая фибробласты [77], клетки эпителия
зываемые G квадруплексы. G4 квадруплексы
[78] и ретикулоциты [79], ингибирование Na,K
специфически связывают ионы одно и двухва
ATPазы сопровождается уменьшением синтеза
лентных металлов внутри полости, образован
белков без воздействия на трансмембранный
ной карбонильными группами остатков гуани
перенос меченых аминокислот, внутриклеточ
на, при этом катионы теряют свою гидратную
ное содержание ATP и сборку рибосом. На при
оболочку. Максимальный стабилизирующий
мере лизатов ретикулоцитов было установлено,
эффект наблюдается для K+, а Na+ оказывает ме
что полумаксимальное увеличение синтеза гло
нее выраженное стабилизирующее действие.
бина в средах, содержащих 60, 90 и 125 мM Na+,
Характерно, что G квадруплексы часто встреча
наблюдается при [K+] = 15, 25 и 40 мМ соответ
ются в промоторах. Дальнейшие исследования
ственно [79]. В связи с этим можно предполо
позволят установить, связано ли изменение ста
жить, что Na+ (в отличие от К+) не активирует
бильности таких G квадруплексов, вызванное
процесс синтеза белка, но конкурирует с К+ за
БИОХИМИЯ том 84 вып. 11 2019
ОДНОВАЛЕНТНЫЕ КАТИОНЫ КАК ВТОРИЧНЫЕ ПОСРЕДНИКИ
1603
места связывания с сенсором, вовлеченным в
ГМК аорты крысы, что контрастировало с
регуляцию трансляции. В самом деле, было об
(7-8)× увеличением синтеза РНК, измеренного
наружено, что в отличие от K+ и Rb+, Na+, Li+ и
по включению [3H] уридина (рис. 9). В присут
Cs+ не оказывают достоверного влияния на ак
ствии ростовых факторов наблюдалось 10× и 2×
тивность очищенной тирозил тРНК синтетазы
увеличение синтеза РНК и белка соответствен
человека [80].
но. В этом случае уабаин вызывал 2× снижение
Следует обратить внимание и на тот факт,
синтеза белка на фоне 2× увеличения синтеза
что обнаруженное в интактных клетках подавле
РНК [44].
ние синтеза белка в ответ на уменьшение [К+]i
Для объяснения этого явления могут быть
может быть отчасти компенсировано за счет
предложены 3 гипотезы. (i) Содержание K+i
увеличения [Na+]i и активации рассмотренного
чувствительного сенсора, вовлеченного в регу
выше Na+ зависимого механизма регуляции
ляцию трансляции, тканеспецифично и сниже
транскрипции генов, в том числе генов, вовле
но в клетках гладкой мускулатуры. В самом де
ченных в процессы элонгации при трансляции
ле, в отличие от тирозил тРНК синтетазы мле
[81, 82]. Так, например, мы установили, что ин
копитающих К+ не оказывает существенного
гибирование Na,K ATРазы кардиотоническими
влияния на активность этого фермента, изоли
стероидами в клетках эндотелия человека соп
рованного из бактерий [84, 85]. (ii) K+i чувстви
ровождается
3× уменьшением содержания
тельный механизм регуляции трансляции огра
мРНК фактора элонгации трансляции EIF5
ничен определенным набором генов [86]. (iii)
[83]. Следует также отметить, что относитель
Ингибирование трансляции гена, вызванное
ный вклад К+ чувствительного механизма
падением [K+]i, может быть компенсировано за
трансляции в регуляцию экспрессии генов но
счет рассмотренного выше механизма Na+i зави
сит тканеспецифический характер. Действи
симой активации транскрипции как самого это
тельно, при отсутствии ростовых факторов не
го гена, так и ЕRG, вовлеченных в регуляцию
обнаружено достоверного влияния 24 часового
его транскрипции. В настоящее время мы про
ингибирования Na,K ATPазы уабаином на
водим экспериментальную проверку этих гипо
включение [3H] лейцина во фракцию белков
тез.
Рис. 9. Влияние уабаина (1 мМ, 24 ч) на синтез РНК (а) и белка (б) в клетках гладкой мускулатуры аорты крысы. В части
экспериментов среда инкубации содержала 1% сыворотку быка (CS). Использованы данные, приведенные в статье
Orlov et al. [44]
БИОХИМИЯ том 84 вып. 11 2019
6*
1604
КЛИМАНОВА и др.
ФИЗИОЛОГИЧЕСКОЕ
новлено, что изменения транскрипции 1018 ге
И ПАТОФИЗИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ
нов, вызванные EPS, сохранялись в присут
Na+i,K+i5ЧУВСТВИТЕЛЬНОГО МЕХАНИЗМА
ствии никардипина. В отличие от изменений
РЕГУЛЯЦИИ ЭКСПРЕССИИ ГЕНОВ
ионного баланса EPS не оказывала влияния на
фосфорилирование ацетил CoA карбоксилазы
В этом разделе мы ограничимся только нес
и фактора Unc 51, используемых в качестве
колькими примерами, демонстрирующими зна
маркеров активации АМРК. Среди EPS
чение Na+i,K+i чувствительного механизма регу
чувствительных генов было выявлено ~300
ляции экспрессии генов в формировании функ
транскриптов, чье содержание также изменя
ционального ответа клеток.
лось при добавлении уабаина в концентрациях,
Секреция миокинов. В 2000 х гг. было уста
вызывающих увеличении [Na+]i/[K+]i в диапазо
новлено, что скелетные мышцы функциониру
не, который соответствовал выявленному в
ют как эндокринный орган, продуцирующий по
клетках, подвергнутых электростимуляции. Со
мере увеличения физической нагрузки цитоки
вокупность полученных данных указывает на
ны и другие низкомолекулярные белки, полу
участие Na+i чувствительного, Ca2+ и АМРК
чившие название миокинов. Предполагается,
независимого механизма сопряжения возбужде
что в основе этого явления лежат 3 механизма
ния и транскрипции в регуляции экспрессии ге
[57]. (i) Возбуждение скелетной мускулатуры
нов в клетках скелетной мускулатуры.
сопровождается активацией Ca2+ каналов L ти
Формирование клеточной памяти. В конце
па, увеличением [Ca2+]i и изменениями тран
прошлого и начале нынешнего столетия было
скрипции генов по механизмам, рассмотрен
показано, что при возбуждении нейронов го
ным в разделе «Доказательство Са2+ независи
ловного мозга резко возрастает содержание
мого механизма проведения сигнала». (ii) Физи
мРНК Erg1 и c-Fos [92-94], которые при сравне
ческая нагрузка приводит к увеличению соотно
нии действия уабаина и бескалиевой среды на
шения АМР/АТР и активации AMPK, вовле
клетки ГМК и электрически невозбудимых тка
ченной в регуляцию транскрипции генов. (iii)
ней были классифицированы как Na+i,K+i
Интенсивная физическая нагрузка сопровожда
чувствительные гены раннего ответа [51, 52]. В
ется локальным снижением парциального дав
опытах на Egr1+/- мышах было обнаружено рез
ления кислорода (pO2), увеличением содержа
кое ухудшение их способности к обучению [95],
ния индуцируемого гипоксией фактора HIF 1α
что указывало на ключевую роль изменений
и активацией гетеродимером HIF 1α/HIF 1β
транскриптома нейронов, опосредованных
элемента HRE, обнаруженного в 5' UTR промо
этим геном, в формировании долгосрочной па
торах ряда pO2 чувствительных генов [18, 87]. В
мяти (см. обзор Thiel et al. [96]). Было установле
нашей работе были получены данные о сущест
но, что увеличение транскрипции Egr1 и других
вовании альтернативного сигнального механиз
ЕRG при возбуждении нейронов по крайней
ма, реализуемого через активацию транскрип
мере частично опосредовано активацией рецеп
ции в ответ на увеличение [Na+]i.
торов N метиl D аспартата (NMDA). Так как
Культивируемые клетки скелетной мускула
активация NMDA рецепторов сопровождается
туры, подвергнутые импульсной электростиму
увеличением [Ca2+]i, была сформулирована ги
ляции (electrical pulse stimulation, EPS), рассмат
потеза о решающей роли Ca2+ опосредованных
риваются как наиболее адекватная in vitro мо
сигнальных систем в формировании долгосроч
дель сокращающей скелетной мускулатуры [88].
ной памяти нейронов [97].
В нашей лаборатории было установлено, что,
Следует, однако, отметить, что проницае
как и в экспериментах in vivo [89], 4 х часовая
мость NMDA рецепторов для одновалентных
электростимуляция миотубул мыши линии
катионов выше, чем для Са2+ (PNa~PK>>PCa), и
C2C12 приводит к ~5× увеличению соотноше
их активация может приводить к увеличению
ния [Na+]i/[K+]i [90]. Учитывая отсутствие ги
соотношения [Na+]i/[K+]i. Действительно, уве
поксии в такого рода экспериментах, мы иссле
личение [Na+]i отмечено как при электрическом
довали роль Са2+ и АМРК зависимой сигналь
[98], так и при фармакологическом [99] возбуж
ной системы в изменениях транскриптома кле
дении нейронов головного мозга. С использова
ток С2С12 [91]. Было установлено, что наряду с
нием селективных флуоресцентных индикато
накоплением Na+ и потерей K+, EPS сопровож
ров внутриклеточного Na+ и Са2+ было показа
дается ритмическими осцилляциями [Ca2+]i и
но, что при возбуждении пирамидальных ней
изменением содержания 3215 транскриптов.
ронов вход Na+ предшествует увеличению
Добавление никардипина, блокатора Ca2+ кана
[Ca2+]i. Было также установлено, что в отличие
лов L типа, устраняло осцилляции [Ca2+]i, не
от быстрой нормализации [Ca2+]i, увеличение
влияя на соотношение [Na+]i/[K+]i. Было уста
[Na+]i, вызванное возбуждением нейронов, нор
БИОХИМИЯ том 84 вып. 11 2019
ОДНОВАЛЕНТНЫЕ КАТИОНЫ КАК ВТОРИЧНЫЕ ПОСРЕДНИКИ
1605
мализуется в более длительном временном диа
действие гипоксии на соотношение [Na+]i/[K+]i
пазоне [100].
и транскрипцию Fos, Atf3, Ptgs2 и Per2 и резко
В последних работах мы сопоставили
снижала прирост содержания Klf10, Edn1, Nr4a1
действие уабаина на соотношение [Na+]i/[K+]i и
и Hes1. Трансфекция гладкомышечных клеток
транскриптом культивируемых нейронов голов
Hif-1a siRNA уменьшала прирост содержания
ного мозга крысы [101]. Среди [Na+]i/[K+]i
мРНК Vegfa, Edn1, Klf10 и Nr4a1, регистрируе
чувствительных регуляторов транскрипции мы
мого в условиях гипоксии, но не влияла на
обнаружили увеличение экспрессии Npas4, Fos,
экспрессию Fos, Atf3, Ptgs2 и Per2 [16, 17]. Эти
Junb, Crem, Atf3, Klf4 и Crebzf. Ранее активация
результаты свидетельствуют, что Na+i/K+i опосре
транскрипции этих ERG была выявлена в ней
дованный, Hif 1α независимый механизм регу
ронах, как при их электростимуляции, так и при
ляции транскрипции вносит существенный
воздействии нейротрансмиттеров
[93,
94,
вклад в регуляцию экспрессии генов в условиях
102-104]. В ряде работ было также продемон
гипоксии.
стрировано участие этих генов в процессах обу
чения и формирования пластичности синапсов
Данные, изложенные в нашем обзоре, пока
[105, 106]. В настоящее время в нашей лаборато
зывают, что активность ряда ферментов, а также
рии проводится изучение относительной роли
процессы транскрипции и трансляции регули
Ca2+ опосредованных и Ca2+ независимых сиг
руются при изменении концентрации внутри
нальных систем, активирующихся при диссипа
клеточного Na+ и K+ в физиологически сущест
ции трансмембранных градиентов одновалент
вующем диапазоне, что позволяет рассматри
ных катионов в нейронах головного мозга и во
вать одновалентные катионы в качестве вторич
влеченных в изменения транскриптома в ответ
ных посредников. Следует, однако, отметить,
на электрическую и фармакологическую стиму
что в отличие от Са2+ и других вторичных по
ляцию.
средников, молекулярная организация участков
Гипоксия. Как уже отмечалось выше, сниже
связывания Na+ и K+, образованных третичной
ние деградации HIF 1 в ответ на уменьшение
структурой белка, изучена только для ограни
pO2 и взаимодействие HIF 1 с чувствительным к
ченного числа сенсоров, рассмотренных в раз
гипоксии HRE рассматривается как основной
деле «Идентифицированные сенсоры одновалент
механизм регуляции транскрипции в условиях
ных катионов». Наряду с очевидным фундамен
гипоксии целого ряда генов, среди которых на
тальным значением продолжение этих исследо
иболее изучены Vegfa, Edn1, Klf10 и Nr4a1 [107,
ваний может иметь и существенные практичес
108]. Следует отметить, что 5' UTR ряда pO2
кие приложения. Так, например, было обнару
чувствительных генов, включая Egr1, Atf3, Ptgs2,
жено, что процесс дифференцировки миофиб
Il6, Ppp1r5, Hes1, Nfkbiz, Txnip, Adamts1, Egr 3,
робластов, который активируется при добавле
Cxcl2 и Hsp70, не содержит HRE (см статью
нии к культуре фибробластов легких человека
Koltsova et al. [51]). В этой связи мы обратили
ростового фактора (tumour transforming growth
внимание на то, что Egr1, Atf3, Ptgs2, Il6, Ppp1r5,
factor TGFb1), трансформирующего опухоль,
Hes1, Nfkbiz и Txnip попадают в разряд генов, чья
подавляется КТС [58]. Инъекция КТС также ин
транскрипция в гладкомышечных клетках воз
гибировала процесс накопления миофиброб
растает в ответ на увеличение соотношения
ластов в легких экспериментальных животных,
[Na+]i/[K+]i [51]. С целью дальнейшего изучения
рассматриваемых в качестве адекватной модели
механизма этого явления мы сопоставили
идиопатического фиброза легких человека
действие на транскриптом ГМК ингибиторов
[109]. Было установлено, что это действие КТС
Na,K ATPазы и гипоксии в сочетании с глюкоз
опосредовано ингибированием Na,K ATPазы,
ным голоданием. В этих условиях гипоксия соп
увеличением [Na+]i и последующей активацией
ровождалась 3× увеличением [Na+]i и 2× увели
транскрипции Ptgs2, кодирующего циклоокси
чением
[K+]i. Как ингибирование Na,K
геназу второго типа (СОХ 2) [58], на фоне по
ATPазы, так и гипоксия приводили к однонап
давления транскрипции субъединицы TGFBR2
равленному изменению транскрипции десятков
[58]. Как в экспериментах in vitro, так и in vivo
генов, включая Cyp1a1, Fos, Atf3, Klf10, Ptgs2,
было также показано, что КТС ингибирует реп
Nr4a1, Per2 и Hes1. Устранение трансмембран
ликацию вируса гриппа за счет уменьшения
ного градиента одновалентных катионов в среде
[K+]i и подавления K+i чувствительного звена
со сниженным содержанием Na+ и увеличен
трансляции [78]. Следует, однако, отметить, что
ным содержанием K+ устраняло действие уабаи
фармакологическое использование КТС в дозах,
на как на соотношение [Na+]i/[K+]i, так и на
увеличивающих соотношение [Na+]i/[K+]i, зат
экспрессию перечисленных выше генов. Анало
руднено ввиду их токсического действия [110].
гичная процедура также полностью устраняла
В этой связи представляется перспективным
БИОХИМИЯ том 84 вып. 11 2019
1606
КЛИМАНОВА и др.
разработка лекарственных препаратов, обеспе
00063, разделы «Идентифицированные сенсоры
чивающих тканеспецифическое воздействие на
одновалентных катионов», «Натрий как регуля
активность сенсоров одновалентных катионов,
тор транскрипции» и «Калий как регулятор
вовлеченных в регуляцию транскрипции и
трансляции»).
трансляции.
Благодарности. Авторы выражают благодар
ность профессору медицинского колледжа
г. Олбани (США) А.А. Монгину за прочтение
Финансирование. Работа выполнена при фи
обзора и сделанные критические замечания.
нансовой поддержке Российского научного
Конфликт интересов. Авторы заявляют, что у
фонда (грант № 16 15 10026 п, раздел «Физио
них нет конфликта интересов.
логическое и патофизиологическое значение
Соблюдение этических норм. Все примени
Na+i,K+i чувствительного механизма регуляции
мые международные, национальные и/или инс
экспрессии генов») и Российского фонда фун
титуциональные принципы ухода и использова
даментальных исследований (грант № 18 04 ния животных были соблюдены.
СПИСОК ЛИТЕРАТУРЫ
1.
Sutherland, E.W. (1972) Studies on the mechanism of hor
17.
Orlov, S.N., Birulina, Y.G., Smaglii, L.V., and Gusakova,
mone action, Science, 177, 401-408.
S.V. (2017) Vascular smooth muscle as an oxygen sensor:
2.
Robison, G.A., Butcher, R.W., and Sutherland, E.W.
role of elevation of the [Na+]i/[K+]i, in Hypoxia and
(1971) Cyclic AMP, Academic Press, New York.
Human Diseases (Zheng, J., and Zhou, C., eds.),
3.
Lincoln, T.M., and Cornwell, T.L. (1993) Intracellular
IntechOpen, Vol. 4, pp. 73-90, doi: 10.5772/65384.
cyclic GMP receptor proteins, FASEB J., 7, 328-338.
18.
Kapilevich, L.V., Kironenko, T.A., Zaharova, A.N.,
4.
Carafoli, E. (2002) Calcium signaling: a tale for all seasons,
Kotelevtsev, Yu.V., Dulin, N.O., and Orlov, S.N. (2015)
Proc. Natl. Acad. Sci. USA, 99, 1115-1122.
Skeletal muscle as an endicrine organ: role of
5.
Berridge, M.J. (1993) Inositol triphosphate and calcium
[Na+]i/[K+]i mediated excitation-transcription coupling,
signalling, Nature, 361, 315-325.
Genes Dis., 2, 328-336.
6.
Grinstein, S., Smith, J.D., Benedict, S.H., and Gelfand, E.W.
19.
Orlov, S.N., and Hamet, P. (2006) Intracellular monova
(1989) Activation of sodium hydrogen exchange by mito
lent ions as second messengers, J. Membr. Biol., 210,
gens, Curr. Topics Membr. Transport, 34, 331-343.
161-172.
7.
Prasad, K.V.S., Severini, A., and Kaplan, J.G. (1987) Sodium
20.
Shekarabi, M., Zhang, J., Khanna, A.R., Ellison, D.H.,
ion fluxes in proliferating lymphocytes: an early component of
Delpire, E., and Kahle, K.T. (2017) WNK kinase signaling
mitogenic signal, Arch. Biochem. Biophys., 252, 515-525.
in ion homeostasis and human disease, Cell Metab., 25,
8.
Wakabayashi, S., Shigekawa, M., and Poyssegur, J. (1997)
285-299.
Molecular physiology of vertebrate Na+/H+ exchanger,
21.
Wilson, C.S., and Mongin, A.A. (2019) The signaling role
Physiol. Rev., 77, 51-74.
for chloride in the bidirectional communication between
9.
Marakhova, I.I., Vereninov, A.A., Toropova, F.V., and
neurons and astrocytes, Neurosci. Lett., 689, 33-44.
Vinogradova, T.A. (1998) Na,K, ATPase pump in activat
22.
Page, M.J., and Di Cera, E. (2006) Role of Na+ and K+ in
ed human lymphocytes: on the mechanisms of rapid and
enzyme function, Physiol. Rev., 86, 1049-1092.
long term increase in K influxes during the initiation of
23.
Linden, D.J., Smeyne, M., and Connor, J.A. (1993) Induc
phytohemagglutinin induced proliferation, Biochim.
tion of cerebellar long term depression in culture requires
Biophys. Acta, 1368, 61-72.
postsynaptic action of sodium ions, Neuron, 11, 1093-1100.
10.
Burns, C.P., and Rozengurt, E. (1984) Extracellular Na+
24.
Kanai, R., Ogawa, H., Vilsen, B., Cornelius, F., and
and initiation of DNA synthesis: role of intracellular pH
Toyoshima, C. (2013) Crystal structure of Na+ bound
and K+, J. Cell Biol., 98, 1082-1089.
Na+,K+ ATPase preceding the E1P state, Nature, 502,
11.
Hamet, P., Orlov, S.N., DeBlois, D., Sun, Y., Kren, V., and
201-206.
Tremblay, J. (2004) Angiotensin as a cytokine implicated in
25.
Sweadner, K.J., and Donnet, C. (2001) Structural similar
accelerated cellular turnover, Springer Verlag, N.Y., pp. 71-98.
ities of Na,K ATPase and SERCA, the Ca2+ ATPase of the
12.
Bennekou, P. and Christophersen, P. (2003) Ion channels,
sarcoplasmic reticulum, Biochem. J., 356, 685-704,
Springer, Berlin, pp. 139-152.
doi: 10.1042/0264 6021:3560685.
13.
Schneider, J., Nicolay, J.P., Foller, M., Wieder, T., and
26.
Toyoshima, C., Kanai, R., and Cornelius, F. (2011) First
Lang, F. (2007) Suicidal erythrocyte death following cellu
crystal structure of Na+,K+ ATPase: new light on the old
lar K+ loss, Cell Physiol. Biochem., 20, 35-44.
est ion pump, Structure, 19, 1732-1738, doi: 10.1016/
14.
Rose, C.R., and Konnerth, A. (2001) NMDA receptor
j.str.2011.10.016.
mediated Na+ signals in spines and dendrites, J. Neurosci.,
27.
Pert, C.B., Pasternak, G., and Snyder, S.H. (1973) Opiate
21, 4207-4214.
agonists and antagonists discriminated by receptor binding
15.
Verkhratsky, A., Noda, M., Parpura, V., and Kirischuk, S.
in brain, Science, 182, 1359-1361.
(2013) Sodium fluxes and astroglial function, Adv. Exp.
28.
Katritch, V., Fenalti, G., Abola, E.E., Roth, B.L.,
Med. Biol., 961, 295-305.
Cherezov, V., and Stevens, R.C. (2014) Allosteric sodium
16.
Koltsova, S.V., Shilov, B., Burulina, J.G., Akimova, O.A.,
in class A GPCR signaling, Trends Biochem. Sci., 39,
Haloui, M., Kapilevich, L.V., Gusakova, S. V., Tremblay, J.,
233-244.
Hamet, P., and Orlov, S.N. (2014) Transcriptomic changes
29.
Strasser, A., Wittmann, H. J., Schneider, E.H., and
triggered by hypoxia: evidence for HIF 1a independent,
Seifert, R. (2015) Modulation of GPCRs by monovalent
[Na+]i/[K+]i mediated excitation transcription coupling,
cations and anions, Naunyn Schmied. Arch. Pharmacol.,
PLoS One, 9, e110597.
388, 363-380, doi: 10.1007/s00210 014 1073 2.
БИОХИМИЯ том 84 вып. 11 2019
ОДНОВАЛЕНТНЫЕ КАТИОНЫ КАК ВТОРИЧНЫЕ ПОСРЕДНИКИ
1607
30.
Liu, W., Chun, E., Thompson, A.A., Chubukov, P., Xu, F.,
46.
Orlov, S.N., and Hamet, P. (2004) Apoptosis vs oncosis:
Katritch, V., Han, G.W., Roth, C.B., Heitman, L.H.,
role of cell volume and intracellular monovalent cations,
Ijzerman, A.P., Cherezov, V., and Stevens, R.C. (2012)
Adv. Exp. Med. Biol., 559, 219-233.
Structural basis for allosteric regulation of GPCRs by sodi
47.
Taurin, S., Dulin, N.O., Pchejetski, D., Grygorczyk, R.,
um ions, Science, 337, 232-236.
Tremblay, J., Hamet, P., and Orlov, S.N. (2002) c Fos expres
31.
Fenalti, G., Giguere, P.M., Katritch, V., Huang, X.P.,
sion in ouabain treated vascular smooth muscle cells from rat
Thompson, A.A., Cherezov, V., Roth, B.L., and Stevens, R.C.
aorta: evidence for an intracellular sodium mediated, calci
(2014) Molecular control of d opioid receptor signalling,
um independent mechanism, J. Physiol., 543, 835-847.
Nature, 506, 191-196.
48.
Orlov, S.N., Taurin, S., Thorin Trescases, N., Dulin, N.O.,
32.
Gutierrez de Teran, H., Massink, A., Rodriguez, D., Liu, W.,
Tremblay, J., and Hamet, P. (2000) Inversion of the intra
Han, G.W., Joseph, J.S., Katritch, I., Heitman, L.H., Xia, L.,
cellular Na+/K+ ratio blocks apoptosis in vascular smooth
Ijzerman, A.P., Cherezov, V., Katritch, V., and Stevens, R.C.
muscle cells by induction of RNA synthesis, Hypertension,
(2013) The role of sodium ion binding site in the allosteric
35, 1062-1068.
modulation of the A2A adenosine G protein coupled
49.
Schoner, W., and Scheiner Bobis, G. (2007) Endogenous
receptor, Structure, 21, 2175-2185.
and exogenous cardiac glycosides: their role in hyperten
33.
O’Brien, M.C., and McKay, D.B. (1995) How potassium
sion, salt metabolism, and cell growth, Am. J. Physiol. Cell
affects the activity of the molecular chaperone Hsc70.
Physiol., 293, C509-C536.
I Potassium is required for optimal ATPase activity, J. Biol.
50.
Orlov, S.N., Klimanova, E.A., Tverskoi, A.M.,
Chem., 270, 2247-2250.
Vladychenskaya, E.A., Smolyaninova, L.V., and Lopina, O.D.
34.
Arakawa, A., Handa, N., Shirouzu, M., and Yokoyama, S.
(2017) Na+i,K+i dependent and independent signaling trig
(2011) Biochemical and structural studies on the high
gered by cardiotonic steroids: facts and artifacts, Molecules,
affinity of Hsp70 for ADP, Protein Sci., 20, 1367-1379.
22, E635, doi: 10.3390/molecules22040635.
35.
Орлов С.Н. (1987) Кальмодулин, Общие проблемы фи-
51.
Koltsova, S.V., Trushina, Y., Haloui, M., Akimova, O.A.,
зико-химической биологии. ВИНИТИ, 8, 5-212.
Tremblay, J., Hamet, P., and Orlov, S.N. (2012) Ubiquitous
36.
Permyakov, E.A., Uversky, V.N., and Permyakov, S.E.
[Na+]i/[K+]i sensitive transcriptome in mammalian cells:
(2017) Parvalbumin as a pleomorphic protein, Curr. Protein
evidence for Ca2+ independent excitation-transcription
Pept. Sci., 18, 780-794.
coupling, PLoS One, 7, e38032.
37.
Heizmann, C.W., and Hunziker, W. (1991) Intracellular
52.
Klimanova, E.A., Sidorenko, S.V., Smolyaninova, L.V.,
calcium binding proteins: more sites than insights, Trends
Kapilevich, L.V., Gusakova, S.V., Lopina, O.D., and
Biochem. Sci., 16, 98-103.
Orlov, S.N. (2019) Ubiquitous and cell type specific tran
38.
Okada, Y., Maeno, E., Shimizu, T., Dezaki, K., Wang, J.,
sciptomic changes triggered by dissipation of monovalent
and Morishima, S. (2001) Receptor mediated control of
cation gradients in rodent cells: physiological and patho
regulatory volume decrease (RVD) and apoptotic volume
physiological implications, Curr. Top. Membr.,
83,
decrease (AVD), J. Physiol., 532, 3-16.
107-149, doi: 10.1016/bs.ctm.2019.01.006.
39.
Orlov, S.N., Platonova, A.A., Hamet, P., and Grygorczyk, R.
53.
Orlov, S.N., Thorin Trescases, N., Pchejetski, D., Taurin, S.,
(2013) Cell volume and monovalent ion transporters:
Farhat, N., Tremblay, J., Thorin, E., and Hamet, P. (2004)
their role in the triggereing and progression of the cell
Na+/K+ pump and endothelial cell survival: [Na+]i/[K+]i
death machinery, Am. J. Physiol. Cell Physiol., 305,
independent necrosis triggered by ouabain, and protection
C361-C372.
against apoptosis mediated by elevation of [Na+]i, Pflugers
40.
Orlov, S.N., Thorin Trescases, N., Kotelevtsev, S.V.,
Arch., 448, 335-345.
Tremblay, J., and Hamet, P. (1999) Inversion of the intra
54.
Alonso, M.T., and Garcia Sancho, J. (2011) Nuclear Ca2+
cellular Na+/K+ ratio blocks apoptosis in vascular smooth
signalling, Cell Calcium, 49, 280-289.
muscle at a site upstream of caspase 3, J. Biol. Chem., 274,
55.
Taurin, S., Hamet, P., and Orlov, S.N. (2003) Na/K pump
16545-16552.
and intracellular monovalent cations: novel mechanism of
41.
Zhou, X., Jiang, G., Zhao, A., Bondeva, T., Hirzel, P., and
excitation-transcription coupling involved in inhibition of
Balla, T. (2001) Inhibition of Na,K ATPase activates PI3
apoptosis, Mol. Biol., 37, 371-381.
kinase and inhibits apoptosis in LLC PK1 cells, Biochem.
56.
Santana, L.F. (2008) NFAT dependent excitation-tran
Biophys. Res. Commun., 285, 46-51.
scription coupling in heart, Circ. Res., 103, 681-683.
42.
Isaev, N.K., Stelmashook, E.V., Halle, A., Harms, C.,
57.
Gundersen, K. (2011) Excitation-transcription coupling
Lautenschlager, M., Weih, M., Dirnagl, U., Victorov, I.V.,
in skeletal muscle: the molecular pathways of exercise, Biol.
and Zorov, D.B. (2000) Inhibition of Na+,K+ ATPase
Rev., 86, 564-600.
activity in cultured cerebellar granule cells prevents the
58.
La, J., Reed, E.B., Koltsova, S.V., Akimova, O.A.,
onset of apoptosis induced by low potassium, Neurosci.
Hamanaka, R.B., Mutlu, R.B., Orlov, S.N., and
Lett., 283, 41-44.
Dulin, N.O. (2016) Regulation of myofibroblast differenti
43.
Trevisi, L., Visentin, B., Cusinato, F., Pighin, I., and
ation by cardiac glycosides, Am. J. Physiol. Lung Cell. Mol.
Luciani, S. (2004) Antiapoptotic effect of ouabain on
Physiol., 310, L815-L823.
human umbilical endothelial cells, Biochem. Biophys. Res.
59.
Smolyaninova, L.V., Koltsova, S.V., Sidorenko, S.V., and
Commun., 321, 716-721.
Orlov, S.N. (2017) Augemented gene expression triggered
44.
Orlov, S.N., Taurin, S., Tremblay, J., and Hamet, P. (2001)
by Na+,K+ ATPase inhibition: role of Ca2+ mediated and
Inhibition of Na+,K+ pump affects nucleic acid synthesis
independent excitation-transcription coupling, Cell
and smooth muscle cell proliferation via elevation of the
Calcium, 68, 5-13.
[Na+]i/[K+]i ratio: possible implication in vascular remod
60.
Orlov, S.N., Aksentsev, S.L., and Kotelevtsev, S.V. (2005)
eling, J. Hypertens., 19, 1559-1565.
Extracellular calcium is required for the maintenance of
45.
Taurin, S., Seyrantepe, V., Orlov, S.N., Tremblay, T. L.,
plasma membrane integrity in nucleated cells, Cell
Thibaut, P., Bennett, M.R., Hamet, P., and Pshezhetsky, A.V.
Calcium, 38, 53-57.
(2002) Proteome analysis and functional expression identi
61.
Koltsova, S.V., Tremblay, J., Hamet, P., and Orlov, S.N.
fy mortalin as an anti apoptotic gene induced by elevation
(2015) Transcriptomic changes in Ca2+ depleted cells: role
of [Na+]i/[K+]i ratio in cultured vascular smooth muscle
of elevated intracellular [Na+]/[K+] ratio, Cell Calcium, 58,
cells, Circ. Res., 91, 915-922.
317-324.
БИОХИМИЯ том 84 вып. 11 2019
1608
КЛИМАНОВА и др.
62.
Matt, T., Martinez Yamout, M.A., Dyson, H.J., and
translation machinery, Am. J. Physiol. Lung Cell. Mol. Physiol.,
Wright, P.E. (2004) The CBP/p300 XAZ1 domain in its
316, L1094-L1106, doi: 10.1152/ajplung.00173.2018.
native state is not a binding patner of MDM2, Biochem. J.,
79.
Cahn, F., and Lubin, M. (1978) Inhibition of elongation
381, 685-691.
steps of protein synthesis at reduced potassium concentra
63.
Krishtal, O.A., Pidoplichko, V.I., and Shakhovalov, Y.A.
tions in reticulocytes and reticulocyte lysate, J. Biol. Chem.,
(1981) Conductance of the calcium channel in the mem
253, 7798-7803.
brane of snail neurones, J. Physiol., 301, 423-434.
80.
Austin, J., and First, E.A. (2002) Potassium functionally
64.
Nakagawa, Y., Rivera, V., and Larner, A.C. (1992) A role
replaced the second lysine of the KMSKS signaure
for Na/K ATPase in the control of human c-fos and c-jun
sequence in human tyrosyl tRNA synthetase, J. Biol.
transcription, J. Biol. Chem., 267, 8785-8788.
Chem., 277, 20243-20248.
65.
Haloui, M., Taurin, S., Akimova, O.A., Guo, D. F.,
81.
Jennings, M.D., and Pavitt, G.D. (2010) eIF5 is a dual
Tremblay, J., Dulin, N.O., Hamet, P., and Orlov, S.N.
function GAP and GDI for eukariotic translational con
(2007) Na+i induced c-Fos expression is not mediated by
trol, Small GTPases, 1, 118-123.
activation of the 5’ promoter containing known transcrip
82.
Cao, J., He, L., Lin, G., Hu, C., Dong, R., Zhang, J.,
tional elements, FEBS J., 274, 3257-3267.
Zhu, H., Hu, Y., Wagner, C.R., He, Q., and Yang, B.
66.
Ono, Y., Ojimam, K., Torii, F., Takaya, E., Doi, N.,
(2014) Cap dependent translation initiation factor,eIF4E,
Nakagawa, K., Hata, S., Abe, K., and Sorimachi, H.
is the target for ouabain mediated inhibition of HIF 1a,
(2010) Skeletal muscle specific calpain is an intracellular
Biochem. Pharmacol., 89, 20-30.
Na+ dependent protease, J. Biol. Chem.,
285,
83.
Klimanova, E.A., Tverskoi, A.M., Koltsova, S.V.,
22986-22998.
Sidorenko, S.V., Lopina, O.D., Tremblay, J., Hamet, P.,
67.
Herrera, R.E., Nordheim, A., and Stewart, A.F. (1997)
Kapilevich, L.V., and Orlov, S.N. (2017) Time and dose
Chromatin structure analysis of the human c Fos promot
dependent actions of cardiotonic steroids on transcriptome
er reveals a centrally positioned nucleosome, Chromosoma,
and intracellular content of Na+ and K+: a comparative
106, 284-292.
analysis, Sci. Rep., 7, 45403, doi: 10.1038/srep45403.
68.
Chinsomboon, J., Ruas, J., Gupta, R.K., Thom, R.,
84.
Fisher, P.A., Goodman, J.G., and Kull, F.L.
(1976)
Shoag, J., Rowe, G.C., Sawada, N., Raghuram, S., and
Thyroid ribonucelic acid iodopeptides. Comparison of
Arany, Z. (2009) The transcriptional coactivator PGC
thyrosyl complex II and thyrosyl tRNA, Biochemistry, 15,
1alpha mediates exercise induced angiogenesis in skeletal
4065-4070.
muscle, Proc. Natl. Acad. Sci. USA, 106, 21401-21405.
85.
Quivy, J.P., and Chroboczek, J. (1988) Tyrpsyl tRNA syn
69.
Soyal, S.M., Felder, T.K., Auer, S., Hahne, P., Oberkofler, H.,
thetase from wheat germ, J. Biol. Chem.,
263,
Witting, A., Paulmichl, M., Landwehrmeyer, G.B., Weydt,
15277-15281.
P., and Patsch, W. (2012) A greatly extended PPARGC1A
86.
Dever, T.E. (2002) Gene specific regulation by general
genomic locus encodes several new brain specific isoforms
translation factors, Cell, 108, 545-556.
and influences Huntington disease age of onset, Hum. Mol.
87.
Pedersen, B.K., and Febbraio, M.A. (2008) Muscle as an
Genet., 21, 3461-3470.
endocrine organ: focus on muscle derived interleukin 6,
70.
Mechti, N., Piechaczyk, M., Blanchard, J.M., Jeanteur, P.,
Physiol. Rev., 88, 1379-1406.
and Lebleu, B. (1991) Sequence requirements for prema
88.
Nikolic, N., Gordens, S.W., Thoresen, G.H., Aas, V.,
ture transcription arrest within the first intron of the mouse
Eckel, J., and Eckardt, K. (2017) Electrical pulse stimula
c-fos gene, Mol. Cell. Biol., 11, 2832-2841.
tion of cultured skeletal muscle cells as a model for in vitro
71.
Coulon, V., Veyrune, J. L., Tourkine, N., Vie, A.,
exercise - possibilities and limitations, Acta Physiol., 220,
Hipskind, R.A., and Blanchard, J. M. (1999) A novel cal
310-331.
cium signaling pathway targets the c-fos intragenic tran
89.
Murphy, K.T., Nielsen, O.B., and Clausen, T. (2008)
scriptional pausing site, J. Biol. Chem., 274, 30439-30446.
Analysis of exercise induced Na+ K+ exchange in rat
72.
Lanctot, C., Cheutin, T., Cremer, M., Cavalli, G., and
skeletal muscle, Exp. Physiol., 93, 1249-1262.
Cremer, T. (2007) Dynamic genome architecture in the
90.
Danilov, K., Sidorenko, S.V., Milovanova, K., Klimanova, E.A.,
nuclear space: regulation of gene expression in three
Kapilevich, L.V., and Orlov, S.N. (2017) Electrical pulse
dimensions, Nature Rev. Genet., 8, 104-115.
stimulation decreases electrochemical Na+ and K+ gradi
73.
Trinkle Mulchany, L., and Lamond, A.I. (2008) Nuclear
ents in C2C12 myotubes, Biochem. Biophys. Res. Commun.,
functions in space and time: Gene expression in a dynam
493, 875-878.
ic, constrained environment, FEBS Lett., 582, 1960-1970.
91.
Sidorenko, S.V., Klimanova, E.A., Milovanova, K.,
74.
Maharana, S., Sharma, D., Shi, X., and Shivashankar, G.V.
Lopina, O.D., Kapilevich, L.V., Chibalin, A.V., and Orlov, S.N.
(2012) Dynamic organization of transcription compar
(2018) Transciptomic changes in C2C12 myotubes trig
ments is dependent in functional nuclear architecture,
gered by electrical stimulation: role of Ca2+ mediated and
Biophys. J., 103, 851-859.
Ca2+ independent signaling and elevated [Na+]i/[K+]i
75.
Dolinnaya, N.G., Ogloblina, A.M., and Yakubovskaya, M.G.
ratio, Cell Calcium, 76, 72-86.
(2016) Structure, properties, and biological relevance of
92.
Bakowski, D., and Parekh, A.B. (2002) Monovalent cation
the DNA and RNA G quadruplexes: overview 50 years
permeability and Ca2+ block of the store operated Ca2+
after their discovery, Biochemistry (Moscow),
81,
current ICRAC in rat basophylic leukemia cells, Pfluger
1602-1649.
Arch. Eur. J. Physiol., 443, 892-902.
76.
Lubin, M., and Ennis, H.L. (1964) On the role of intracel
93.
Hunt, S.P., Pini, A., and Evan, G. (1987) Induction of c
lular potassium in protein synthesis, Biochim. Biophys.
Fos like protein in spinal cord neurons following sensory
Acta, 80, 614-631.
stimulation, Nature, 328, 632-634.
77.
Ledbetter, M.L.S., and Lubin, M. (1977) Control of pro
94.
Cole, A.J., Saffen, D.W., Baraban, J.M., and Worley, P.F.
tein synthesis in human fibroblasts by intracellular potassi
(1989) Rapid increase of an immediate early gene
um, Exp. Cell Res., 105, 223-236.
meseenger RNA in hippocampal neurons by synaptic
78.
Amarelle, L., Katzen, J., Shigemura, M., Welch, L.C.,
NMDA receptor activation, Nature, 340, 474-476.
Cajigas, H., Peteranderl, C., Celli, D., Herold, S.,
95.
Jones, M.W., Errington, M.L., French, P.J., Fine, P.J.,
Lecuona, E., and Sznajder, J.L. (2019) Cardiac glycosides
Bliss, T.V.P., Garel, S., Charney, P., Bozon, B., Laroche,
decrease influenza virus replication by inhibiting cell protein
S., and Davis, S. (2001) A requirement for the immediate
БИОХИМИЯ том 84 вып. 11 2019
ОДНОВАЛЕНТНЫЕ КАТИОНЫ КАК ВТОРИЧНЫЕ ПОСРЕДНИКИ
1609
early response gene Zif268 in the expression of late LTP
103. Barry, D.N., and Commins, S. (2017) Temporal dynamics
and long term memories, Nat. Neurosci., 4, 289-296.
of emmediate early gene expression during cellular consol
96. Thiel, G., Mayer, S.I., Muller, I., Stefano, L., and Rossler,
idation of spatial memory, Behav. Brain Res., 327, 44-53.
O.G. (2010) Egr 1 - a Ca2+ regulated trabscription factor,
104. Zhu, S., Tai, C., MacVicar, B.A., Jia, W., and Cynader, M.S.
Cell Calcium, 47, 397-403.
(2009) Glutamatergic stimulation triggers rapid Krupple
97. Alberini, C.A. (2009) Transcription factors in long term
Like factor 4 expression in neurons and the overexpression
memory and synaptic plasticity, Physiol. Rev.,
89,
of KLF4 sensitizes neurons to NMDA induced caspase 3
121-145.
activity, Brain Res., 1250, 49-62.
98. Bennay, M., Langer, J., Meier, S.D., Kafitz, K.W., and
105. Flavell, S.W., and Greenberg, M.E. (2008) Signaling
Rose, C.R. (2008) Sodium signals in cerebellar Purkinje
mechanisms linking neuronal activity to gene expressoin
neurons and bergmann glial cells evoked by glutamatergic
and plasticity of the nervous system, Annu. Rev. Neurosci.,
synaptic transmission, Glia, 56, 1138-1149.
31, 563-590.
99. Jaffe, D.B., Johnston, D., Lasser Ross, N., Lisman, J.E.,
106. Malik, A.N., Vierbuchen, T., Hemberg, M., Rubin, A.A.,
Miyakawa, H., and Ross, W.N. (1992) The spread of Na+
Ling, E., Couch, C.H., Stroud, H., Spiegel, I., Farth, K.,
spikes determines the pattern of dendritic Ca2+ entry into
Harmin, D.A., and Greenberg, M.E. (2014) Genome wide
hippocampal neurones, Nature, 357, 244-246.
identification and characterization of functional activity
100. Baeza Lehnert, F., Saab, A.S., Gutierrez, R., Larenas, V.,
dependent enhancers, Nat. Neurosci., 17, 1330-1339.
Diaz, E., Horn, M., Vargas, M., Hosli, L., Stobart, J.,
107. Semenza, G.L. (2003) Targeting HIF 1 for cancer therapy,
Hirrlinger, J., Weber, B., and Barros, L.F. (2019) Non
Nat. Rev. Cancer, 3, 721-732.
canonical control of neuronal energy status by the Na+
108. Wood, I.S., Perez de Heredia, F., Wang, B., and Trayhurn, P.
pump, Cell Metab., 29, 1-13.
(2009) Cellular hypoxia and adipose tisue dysfunction in
101. Smolyaninova, L.V., Shiyan, A.A., Kapilevich, L.V.,
obesity, Proc. Nutr. Soc., 68, 370-377, doi: 10.1017/
Lopachev, A.V., Fedorova, T. N., Klementieva, T.S.,
S0029665109990206.
Moskovtsev, A.A., Kubatiev, A.A., and Orlov, S.N. (2019)
109. Orlov, S.N., La, J., Smolyaninova, L.V., and Dulin, N.O.
Transcriptomic changes triggered by ouabain in rat cere
(2019) Na+,K+ ATPase as a target for treatment of tissue
bellum granule cells: role of a3 and a1 mediated signaling,
fibrosis, Curr. Med. Chem.,
26, doi:
10.2174/
PLoS One, in press.
0929867324666170619105407.
102. Coba, M.P., Valor, L.M., Kopanitsa, M.V., Afinowi, N.O.,
110. Akimova, O.A., Tverskoi, A.M., Smolyaninova, L.V.,
and Grant, S.G. (2008) Kinase networks integrate profiles
Mongin, A.A., Lopina, O.D., La, J., Dulin, N.O., and
of N methyl D aspartate receptor mediated gene
Orlov, S.N. (2015) Critical role of the a1 Na+,K+ ATPase
expression in hippocampus, J. Biol. Chem.,
283,
subunit in insensitivity of rodent cells to cytotoxic action of
34101-34107.
ouabain, Apoptosis, 20, 1200-1210.
SEARCH FOR INTRACELLULAR SENSORS INVOLVED IN THE FUNCTION
OF MONOVALENT CATIONS AS SECONDARY MESSENGERS
Review
E. A. Klimanova1*,**, S. V. Sidorenko1, A. M. Tverskoy1**, A. A. Shiyan1, L. V. Smolyaninova1**,
L. V. Kapilevich2, S. V. Gusakova3, G. V. Maksimov1, O. D. Lopina1**, and S. N. Orlov1,2,3
1 Lomonosov Moscow State University, Faculty of Biology, 119234 Moscow, Russia; E-mail: klimanova.ea@yandex.ru
2 National Research Tomsk State University, 634050 Tomsk, Russia
3 Siberian State Medical University, 634050 Tomsk, Russia
Received June 3, 2019
Revised July 15, 2019
Accepted July 16, 2019
Maintaining the non equilibrium distribution of Na+ and K+ between the cytoplasm and the extracellular medium
suggests the presence of sensors that undergo conformational transitions in response to a change in the intracellular
concentration of these monovalent cations. Molecular nature of the sensors of monovalent cations was established
studying Na,K ATPase structure, receptors associated with GTP binding proteins, and heat shock proteins. Recently,
it has been found that changes in Na+ and K+ intracellular concentrations are a key factor in the transcription and
translation regulation, respectively. This review summarizes main results of these studies with an emphasis on the
physiological and pathophysiological value of the Na+i,K+i sensitive mechanism of gene expression regulation.
Keywords: sodium, potassium, transcription, translation
БИОХИМИЯ том 84 вып. 11 2019