БИОХИМИЯ, 2019, том 84, вып. 11, с. 1622 - 1648
УДК 57.052;57.053;57.054
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ И ТРАНСЛЯЦИОННОЕ
ЗНАЧЕНИЕ ЧУВСТВИТЕЛЬНОСТИ ГИППОКАМПА
К ДИСТАНТНОМУ ФОКАЛЬНОМУ ПОВРЕЖДЕНИЮ МОЗГА:
ЦЕНА СТРЕССОРНОГО ОТВЕТА
Обзор
© 2019
Н.В. Гуляева1,2*
1 Институт высшей нервной деятельности и нейрофизиологии РАН,
117485 Москва, Россия; электронная почта: nata_gul@mail.ru
2 Научно(практический психоневрологический центр им. З.П. Соловьева ДЗМ, 115419 Москва, Россия
Поступила в редакцию 07.06.2019
После доработки 24.08.2019
Принята к публикации 24.08.2019
Фокальные поражения мозга (в первую очередь инсульт и черепномозговая травма) с большой вероятностью
индуцируют развитие впоследствии когнитивных и депрессивных расстройств, часто коморбидных. Ассоци"
ация этих осложнений с изменениями гиппокампа (несмотря на отсутствие первичного повреждения этой
структуры), а также отсутствие четкой зависимости между вероятностью развития депрессии и деменции и
тяжестью и локализацией первичного поражения послужили основой для новой гипотезы о дистантном пов"
реждении гиппокампа, как ключевом звене патогенеза когнитивных и психических расстройств.
В соответствии с этой гипотезой избыток кортикостероидов, секретированных после фокального поражения
мозга, особенно у пациентов с аномальным стрессорным ответом в связи с дисфункцией гипоталамо"гипо"
физарно"надпочечниковой оси (ГГНО), взаимодействует с кортикостероидными рецепторами гиппокампа,
индуцируя сигнальные пути, стимулирующие нейровоспаление и последующие события, включая наруше"
ния нейрогенеза и нейродегенерацию. В этой статье проанализированы молекулярные и клеточные механиз"
мы, ассоциированные с регуляторной ролью ГГНО и множественными функциями кортикостероидных ре"
цепторов гиппокампа. Функциональные и структурные повреждения гиппокампа, региона мозга, селектив"
но уязвимого при действии внешних факторов и отвечающего на них повышением секреции цитокинов, сос"
тавляют основу для нарушения когнитивных функций и развития психопатологии. Эта концепция подтвер"
ждается собственными экспериментальными данными, результатами других групп, а также результатами
проспективных клинических исследований постинсультных осложнений. Предложены клинически релевант"
ные биохимические подходы к прогнозу риска постинсультных/посттравматических когнитивных и депрес"
сивных расстройств путем оценки биохимических маркеров индивидуального стрессорного ответа пациен"
та. Обсуждены патогенетически обоснованные пути предупреждения последствий фокального поражения
мозга воздействием на ключевые молекулярные мишени, ассоциированные с дисфункцией гиппокампа.
КЛЮЧЕВЫЕ СЛОВА: гиппокампа, стресс, гипоталамо"гипофизарно"надпочечниковая ось, кортикостеро"
иды, глюкокортикоидный рецептор, минералокортикоидный рецептор, цитокины, инсульт, черепномозго"
вая травма, депрессия, деменция.
DOI: 10.1134/S0320972519110083
Нас убивает не стресс, а наша реакция на него.
Ганс Селье, «The Stress of Life»
Сложная структура гиппокампа (разные по"
личных форм поведенческой пластичности и
ля, септо"темпоральный градиент) и его связи с
ответа на действие факторов внешней среды.
другими ключевыми структурами мозга объяс"
Гиппокамп, важнейшая структура мозга, вовле"
няют его центральное место в реализации раз"
ченная в рабочую и пространственную память, а
Принятые сокращения: 11HSD - 11β"гидроксистероиддегидрогеназа; АКТГ - адренокортикотропный гормон;
БА - болезнь Альцгеймера; Aβ - амилоид β; BDNF - мозговой нейротрофический фактор; ГАМК - гамма"аминомасля"
ная кислота; КРГ - кортикотропин"рилизинг гормон; КС -кортикостероид(ы); ГР - глюкокортикоидный рецептор;
ГГНО - гипоталамо"гипофизарно"надпочечниковая ось; MCI - мягкие когнитивные нарушения; МР - минералокорти"
коидный рецептор; PSD - постинсультная депрессия; TrkB - рецептор тропомиозин"зависимой киназы B; ИЛ - интер"
лейкин; ФНО"α - фактор некроза опухоли α; ЦНС - центральная нервная система.
* Автор является выпускником кафедры биохимии биологического факультета МГУ им. М.В. Ломоносова.
1622
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1623
также в эмоциональные формы поведения жи"
сии. Активация этих путей обеспечивает как
вотных и человека, очень пластичный отдел
быстрый адаптивный ответ, так и последующие
мозга, что, собственно, предсказуемо для клю"
многоуровневые продолжительные изменения
чевой мнемонической структуры. Однако это
в мозге, лежащие в основе долговременной па"
лишь одна сторона медали. Другая сторона, оче"
мяти, формирующей адаптивный опыт. Нор"
видно, отражающая цену такой пластичности,
мальная активность ГГНО (ритмическое и эпи"
обеспечивающей центральную роль в формиро"
зодическое высвобождение КС из надпочечни"
вании памяти и эмоций, - это селективная
ков) необходима для поддержания гомеостаза.
чувствительность гиппокампа к действию мно"
Отрицательные обратные связи, реализуемые
гочисленных стрессорных факторов, включая
КС, включают многочисленные механизмы,
ишемию, судорожную активность, травмы голо"
ограничивающие активацию ГГНО и потенци"
вы, старение и т.п. [1]. Неудивительно, что гип"
ально пагубную избыточную продукцию КС [3]
покамп является важнейшей мишенью действия
(рис. 1, a). Адекватная секреция КС мощно ре"
«гормонов стресса», в первую очередь, корти"
гулируется комплексом нейронных цепей,
костероидов (КС) в мозге. Вместе с нейромеди"
контролирующих быстрые механизмы обрат"
аторами КС регулируют многочисленные функ"
ной связи, включая негеномные эффекты КС,
ции гиппокампа при развитии и во взрослом ор"
обеспечивающие моментальное ингибирова"
ганизме, а дисфункция кортикостероидной ре"
ние секреции гипоталамического кортикотро"
гуляции составляет патогенетическую основу
пин"рилизинг"гормона (КРГ) и гипофизарного
многих болезней мозга, как неврологических,
адренокортикотропного гормона (АКТГ), в то
так и психических.
время как промежуточные и долговременные
КС - класс стероидных гормонов, выполня"
геномные механизмы включают модуляцию
ющих роль ключевых гормонов стрессорного
лимбических сетей и периферических метабо"
ответа, призванных способствовать преодоле"
лических посредников. Нарушение функции
нию стресса. Действуя через специфические
этого механизма обратной связи (например, в
внутриклеточные рецепторы в мозге и на пери"
результате тяжелого и/или хронического стрес"
ферии, КС регулируют поведение, а также мета"
са) приводит к хронически повышенным уров"
болические, сердечно"сосудистые, иммунные и
ням КС, которые оказывают повреждающий
нейроэндокринные функции. Действие КС
эффект на гиппокамп, структуру с высокой
опосредовано глюкокортикоидными (ГР) и ми"
плотностью ГР и МР и по этой причине очень
нералокортикоидными рецепторами (МР),
чувствительную к КС. Хроническая экспози"
представителями суперсемейства ядерных ре"
ция КС вызывает повреждение нервных клеток
цепторов, которые, связавшись с лигандом,
и атрофию дендритов, снижает нейрогенез в
действуют как факторы транскрипции и могут
гиппокампе и нарушает синаптическую плас"
непосредственно модулировать экспрессию ге"
тичность.
нов. Взаимодействуя с другими факторами
Недавно мы предложили новую гипотезу о
транскрипции, КС могут опосредованно регу"
принципиальном участии механизмов стрессор"
лировать активность ряда генов. В последнее
ного ответа (включая дисфункцию ГГНО, взаи"
время описаны мембраносвязанные ГР и МР
модействие секретированных КС с рецепторами
[2], опосредующие быструю негеномную сигна"
в гиппокампе и последующее нейровоспаление)
лизацию и активирующие пути трансдукции
в дистантном повреждении гиппокампа, лежа"
сигнала, такие как протеинкиназные каскады
щем в основе вызванных фокальными пораже"
для модуляции других транскрипционных фак"
ниями мозга (например, постинсультных и
торов и активации или репрессии генов"мише"
посттравматических), когнитивных наруше"
ней. Эти многочисленные механизмы лежат в
ний/деменции и депрессивных расстройств [4].
основе КС"зависимой регуляции важнейших
В данной статье мы представляем критический
функций в самых разных клетках центральной
обзор данных литературы и собственных резуль"
нервной системы (ЦНС) и периферических ор"
татов для уточнения и укрепления молекуляр"
ганов.
ной части этой концепции. Будут представлены
Адекватный ответ на стресс критичен для
данные и логические основания, подтверждаю"
выживания и адаптивного поведения. У млеко"
щие, что феномен дистантной уязвимости гип"
питающих стрессорный ответ опосредован в
покампа тесно связан с функционированием
первую очередь секрецией КС с участием гипо"
ГГНО и, следовательно, с индивидуальной
таламо"гипофизарно"надпочечниковой оси
стресс"реактивностью. Будут рассмотрены био"
(ГГНО), а также реакцией симпатической
химические механизмы КС"зависимого стрес"
нервной системы и высвобожением катехола"
сорного ответа, а также трансляционное значе"
минов при адренергической нейротрансмис"
ние концепции.
БИОХИМИЯ том 84 вып. 11 2019
1624
ГУЛЯЕВА
РЕЦЕПТОРЫ КС, ГГНО
порядок более низкой аффинностью, а также
И СТРЕССОРНЫЙ ОТВЕТ
оказывают быстрые негеномные эффекты на
возбудимость и активацию нейронов в гиппо"
Стресс как таковой является
кампе, гипоталамусе, амигдале и префронталь"
также причиной самого себя и
ной коре, таким образом в течение минут влияя
своим собственным результатом.
на когнитивные функции, адаптивное поведе"
Ганс Селье
ние и нейроэндокринную систему [8, 9]. ГР и
МР, демонстрирующие в гиппокампе высокую
КС оказывают широкий спектр эффектов в
степень колокализации, в основном находятся в
ЦНС: от регуляции транскрипции генов, кле"
цитоплазме без лиганда, а после связывания с
точной сигнализации, модуляции структуры си"
лигандом происходит их транслокация в ядро с
напса, трансмиссии и функции глии до контро"
последующим выполнением функций тран"
ля поведения. De Kloet et al. [2] справедливо рас"
скрипционных факторов. Таким образом, их
сматривают высвобождение КС из надпочечни"
субклеточная локализация является важным
ков в рамках циркадианного ритма и после
компонентом их биологической активности
стресса, как лучший пример интеграции тела и
[10]. Предполагалось, что МР действуют в про"
сознания с окружающей средой. КС координи"
активном режиме для предотвращения наруше"
руют функции клеток и органов во временн
′х
ний гомеостаза, а дополнительная активация ГР
промежутках от миллисекунд до часов, дней и
в реактивном режиме поддерживает восстанов"
даже продолжительности жизни. Несомненно,
ление после стресса. Трансляционным послед"
рецепторы КС демонстрируют удивительную
ствием является то, что дисбаланс МР и ГР
пластичность, связанную с их определяющей
обусловливает поведенческие дефициты и ней"
ролью в судьбе организма. В середине 1960"х гг.
роэндокринные нарушения, усиливающие уяз"
было сделано первое наблюдение, на котором
вимость организма к зависимым от стресса за"
базируется современная концепция действия
болеваниям мозга. МР реагируют при низких
КС в мозге. В лимбической системе мозга были
концентрациях КС, а для дополнительной акти"
открыты рецепторы КС, опосредующие регуля"
вации ГР требуются более высокие концентра"
цию нейронных сетей, обеспечивающих актив"
ции. Было высказано предположение о том, что
ность ГГНО и поведенческую адаптацию. В се"
занятость МР в нейронах гиппокампa важна для
редине 1970"х гг. были идентифицированы 2
стабильности происходящей трансмиссии, ба"
различных типа ядерных рецепторов КС, МР и
зальной активности и чувствительности систе"
ГР, которые координированно играют ключе"
мы стрессорного ответа, поведенческой реак"
вую роль в стероидном контроле ГГНО и пове"
тивности и выбора ответа. Дополнительная пре"
дения [5]. Опосредованное КС действие отрица"
ходящая активация ГР подавляет возбудимость,
тельной обратной связи реализуется активацией
облегчает восстановление после стрессорного
как ГР, так и МР на центральном уровне, в ос"
ответа и способствует сохранению информа"
новном, в гиппокампе (рис. 1, а). ГР широко
ции.
представлены в регионах мозга и периферичес"
Таким образом, баланс эффектов, опосредо"
ких органах, в то время как плотность МР зна"
ванных МР и ГР, представлялся критичным для
чительна только в отдельных структурах мозга, и
долговременного контроля, специфических ас"
гиппокамп является самым важным примером
пектов нейронной активности, способности ре"
такой структуры. Начиная с 1990"х гг., De Kloet
агировать на стресс и поведенческой адаптации,
и его группа прицельно исследуют баланс про"
опосредованных КС [11]. Эти предположения
цессов, опосредованных МР и ГР, в контроле го"
были подтверждены экспериментально в после"
меостаза, как фактора, определяющего «здо"
дующих исследованиях, которые показали, что
ровье и болезнь» [6]. Эффекты КС могут сильно
устойчивость поведения зависит от сбалансиро"
различаться, в зависимости от типа рецепторов,
ванных процессов, опосредованных МР и ГР;
области мозга, типа клеток и физиологического
мозговые МР способствуют процессам оценки,
контекста, причем эти изменения, в конце кон"
поиску типа преодоления стресса, обучению и
цов, зависят от дифференциальных взаимодей"
извлечению памяти; мозговые ГР способствуют
ствий МР и ГР с другими белками, которые оп"
восстановлению, рациональным решениям и
ределяют связывание лигандов, транслокацию в
контекстуальной памяти, при этом транскрип"
ядро и транскрипционную активность [7].
ционные факторы и ко"регуляторы придают
КС, попавшие в мозг, связываются с двумя
специфичность эффектам, опосредованным МР
типами внутриклеточных рецепторов, которые
и ГР [12]. КС осуществляют быстрое негеном"
регулируют транскрипцию связанных с КС ге"
ное воздействие на нейроны поля СА1 гиппо"
нов, высокоаффинным МР и ГР с примерно на
кампa через классические МР, доступные снару"
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1625
а
б
Рис. 1. Регуляция ГГНО, КС и участие рецепторов КС в стрессорном ответе. а - Организация стрессорного ответа в
ГГНО [3, 14, 62, 63, 93]. В пределах основной нейроэндокринной оси стрессор вызывает высвобождение секретагогов
адренокортикотропного гормона (АКТГ), кортикотропин"рилизинг гормона (КРГ) и аргинин"вазопрессина (АВП) из
нейросекреторных нейронов медиального парвоцеллюлярного паравентрикулярного ядра (ПВЯ) гипоталамуса в пор"
тальный кровоток гипофиза на уровне срединного возвышения; пептиды транспортируются через портальные сосуды
в передний гипофиз, обеспечивая доступ к кортикотрофам. Эти секретагоги стимулируют расщепление про"опиомела"
нокортина (ПОМК) до АКТГ и высвобождение АКТГ из гипофиза в системное кровообращение, индуцируя синтез и
высвобождение кортикостероидов (КС), кортизола (у человека и некоторых животных) или кортикостерона (у большин"
ства животных, в т.ч. грызунов) из коркового слоя надпочечников. Базальное высвобождение КС возрастает в несколь"
ко раз после действия стрессорного фактора. КС поступают в кровообращение и осуществляют как периферическое,
так и центральное действие, достигая однотипных рецепторов практически во всех органах и тканях, включая мозг (КС
свободно проходят гемато"энцефалический барьер, ГЭБ). Клеточные эффекты КС опосредованы глюкокортикоидны"
ми рецепторами (ГР) и минералокортикоидными рецепторами (МР). ГР экспрессируются в большинстве структур и
регионов мозга, связанных с ГГНО, в то время как в гиппокампе экспрессированы и МР. Регуляция ГГНО опосредова"
на многоуровневыми петлями обратной связи между структурами ГГНО и мозга, в первую очередь гиппокампа и фрон"
тальной коры. В гипоталамусе ингибиторный ГАМКергический вход в нейроны ПВЯ усиливается посредством возбуж"
дающего гиппокампального выхода, при этом гамма"аминомасляная кислота (ГАМК) ингибирует секрецию кортикот"
рофа. Циркулирующие КС оказывают влияние на гипоталамус, ингибируя последующую секрецию КРГ, и гипофиз,
ингибируя секрецию АКТГ. Циркулирующий АКТГ также оказывает влияние на гипоталамус, ингибируя секрецию
КРГ. Эти петли отрицательной обратной связи приводят к существенному снижению уровня КС в крови после прекра"
щения действия стрессорного фактора. Стресс подавляет экспрессию МР в гиппокампе, а затем снижает экспрессию
ГР в гиппокампе, индуцируя гиперсекрецию КРГ и АВП и, в конце концов, гиперактивность ГГНО;
б - рецепторы КС и нейроэндокринный стрессорный ответ [2, 6, 8, 12, 96, 105]. Разные фазы стресс"реакции, восста"
новления после стресса и адаптации связаны с различными паттернами секреции КС и активации МР и ГР (детали в
тексте)
БИОХИМИЯ том 84 вып. 11 2019
1626
ГУЛЯЕВА
жи плазматической мембраны и обладающие на
лансированно, модулируя широкий спектр ак"
порядок более низким сродством к кортикосте"
тивностей ЦНС.
рону, чем ядерный вариант, участвующий в ней"
МР играют ключевую роль в поддержании
ропротекции. Соответственно, этот «мембран"
циркадианных ритмов АКТГ и кортизола путем
ный рецептор» должен играть важную роль, ког"
модуляции высвобождения КРГ и аргинин"ва"
да уровни КС высоки, т.е. в начальной фазе
зопрессина [14]. Они связываются как с альдо"
стрессорного ответа. De Kloet et al. [2] показали,
стероном, так и с глюкокортикоидами, при этом
что в этой фазе КС способствуют возбудимости
сродство последних к МР выше, чем к тесно
гиппокампа, усиливая эффект других стрессор"
связанным с ними ГР. МР мозга имеют «2 лица»,
ных гормонов. Эти пермиссивные негеномные
условно названные «Соль и Стресс». «Соль»
эффекты могут вносить вклад в быстрые пове"
имеет отношение к регуляции солевого аппети"
денческие ответы и кодирование связанной со
та и реципрокно к возбуждению, мотивации и
стрессом информации. Важно, что КС, как бы"
вознаграждению через сеть альдостерон"
ло показано, модулируют гиппокампальные си"
чувствительных нейронов, экспрессирующих
напсы. Дендритные шипики представляют со"
МР, с проекциями из ядра одиночного пути и
бой постсинаптические структуры синапсов,
циркумвентрикулярных органов, а «Стресс» от"
необходимые для синаптической пластичности
носится к ядерным и мембранным рецепторам
и когнитивной функции. Быстрые негеномные
лимбической системы переднего мозга, которые
модуляции шипиков были описаны после ап"
работают, как переключатель в выборе опти"
пликации кортикостерона (модель острого
мального пути преодоления стресса [15]. Вслед"
стресса в гиппокампе), и эти модуляции, по"ви"
ствие минимального проникновения альдосте"
димому, опосредованы МР [13]. За быстрыми
рона через гемато"энцефалический барьер и от"
эффектами МР следуют более медленные, опос"
сутствия нейронального фермента 11β"гидро"
редованные ГР эффекты, которые облегчают
стероиддегидрогеназы (11HSD) типа 2 (внутри"
подавление временно повышенной возбудимос"
клеточный «привратник») МР в нейроне пол"
ти, восстановление после стрессорного опыта и
ностью заняты лигандом даже при низких физио"
сохранение информации для будущего исполь"
логических уровнях КС и представляют собой
зования (контекстуализация, рационализация и
ключевой фактор формирования высших ког"
сохранение опыта в памяти) [12] (рис. 1, б).
нитивных функций, таких как запоминание,
Эти последовательные фазы когнитивной
обучение и настроение [16]. Активация лимби"
деятельности зависят от синаптической мета"
ческих МР способствует селективному внима"
пластичности, регулируемой координирован"
нию, извлечению памяти и процессам оценки и
ной активацией МР и ГР, включая использова"
при этом ассоциирована с эмоциональными
ние корегуляторов и факторов для контекст"за"
проявлениями, такими как страх и агрессия. Со"
висимой специфичности действия стероидов;
ответственно, повышение концентрации КС ак"
они могут модулироваться генетическими фак"
тивирует ГР в лимбических сетях переднего моз"
торами и (ранним) опытом. Интересно, что вос"
га, лежащих в основе исполнительных функций
палительные ответы на повреждение (см. следу"
и сохранения памяти; эти эффекты ГР вносят
ющий раздел), по"видимому, управляются сход"
вклад в баланс с МР"зависимыми эффектами на
ным образом сбалансированными процессами,
гомеостаз, возбудимость и поведенческую адап"
опосредованными МР и ГР, включая иницииру"
тацию [15]. Таким образом, к ключевым модуля"
ющие, завершающие и прайминговые механиз"
торам функций МР относятся ГР, которые могут
мы, участвующие в стрессе"адаптации [12].
влиять на функционирование МР образованием
Таким образом, различия в распределении и
гетеродимеров и дифференциальными геном"
аффинности ГР и МР, а также их участие как в
ными или негеномными ответами на экспрес"
быстрых негеномных («мембранные» рецепто"
сию генов и активность ферментов 11HSD, оп"
ры КС), так и в медленных, опосредованных ге"
ределяющих доступность активных КС в клетке
нами эффектах на нейроны (внутриклеточные
[17].
рецепторы КС), обеспечивают тонкую и хорошо
Результаты, полученные на линиях генети"
отрегулированную ассоциацию природных
чески модифицированных мышей, подтвержда"
(например, индуцированных стрессом) сдвигов
ют важность ГР в регуляции ГГНО: нарушение
уровня КС со сложным паттерном изменений
активности ГР активирует, а повышенные уров"
нейронной активности, зависимой от времени и
ни белка ГР ингибируют ГГНО. Генетическое
региона мозга, и, следовательно, с определен"
снижение уровня белка ГР и инактивация гена
ными поведенческими, в частности, когнитив"
ГР в мозге снижает тревожное поведение, что
ными и эмоциональными паттернами. Эта сис"
указывает на центральную роль ГР в эмоцио"
тема «один гормон/два рецептора» работает сба"
нальном поведении. У животных с мутациями
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1627
ГР изменены свойства нейронов поля СА1 гип"
Потенциал"чувствительные Ca(2+) каналы
покампа и гиппокамп"зависимая эксплицитная
гиппокампа относятся к наиболее важным ми"
память. Сравнение животных с мутациями МР и
шеням КС. Занятость ГР лигандом поддержива"
ГР свидетельствует о необходимости МР, но не
ет стабильную электрическую активность ней"
ГР для поддержания гранулярных клеток зубча"
ронов гиппокампа. Если уровни КС низкие,
той фасции [18]. Тем не менее, именно ГР счи"
Ca(2+) токи L"типа в клетках поля CA1 гиппо"
тается основным медиатором стрессорного от"
кампа невелики, а когда КС растут, амплитуда
вета процессов пролиферации, дифференци"
Ca(2+) токов L"типа увеличивается, и этот про"
ровки, миграции и функциональной интегра"
цесс требует связывания гомодимеров ГР с
ции новорожденных нейронов гиппокампа.
ДНК. Хроническая перегрузка Ca(2+) может
Экспрессия ГР непосредственно регулирует ба"
повышать уязвимость лимбических клеток к до"
ланс возбуждения и торможения, ключевой для
полнительным провокациям, например, при
нормального нейрогенеза у взрослых. Видимо,
эпилептических или ишемических эпизодах
именно дисбаланс возбуждающих и тормозных
[22]. ГР способны существенно изменять коли"
процессов лежит в основе аберрантной функцио"
чество митохондриальных транскриптов [23].
нальной интеграции новорожденных нейронов,
Регуляция митохондриальной транскрипции
связанной с психическими и пароксизмальны"
стрессом и КС может быть важной для многих
ми заболеваниями мозга [19].
чувствительных к стрессу тканей, включая мозг.
ГР кодируются геном NR3C1. Трансдукция
Целый ряд стрессоров, как физических, так
сигнала КС включает несколько последователь"
и психологических, индуцирует эпигенетичес"
ных стадий, приводящих к трансактивации тар"
кие модификации, которые позволяют объяс"
гетного гена: связывание лиганда, транслока"
нить, каким образом факторы внешней среды,
цию комплекса лиганда с активированным ГР в
которые не влияют на последовательность ДНК,
ядро, связывание ДНК, взаимодействие с ко"
оказывают такие существенные и продолжи"
активатором и задействование функциональных
тельные эффекты на физиологию и поведение.
транскрипционных механизмов. Каждая стадия
В гранулярных нейронах зубчатой фасции обна"
может быть нарушена, и это приведет к измене"
ружен новый быстрый негеномный механизм:
нию передачи сигнала КС. В частности, при
КС через ГР способствуют проведению сигнала
синдроме глюкокортикоидной резистентности
по протеинкиназному пути ERK"MAPK к ниже"
может быть снижен уровень функциональных
лежащим ядерным киназам MSK1 и Elk"1. Ак"
ГР, сродство и связывание КС, нарушена транс"
тивация этого сигнального пути приводит к
локация ГР в ядро, снижено или отсутствовать
фосфорилированию серина10 (S10) и ацетили"
связывание с ДНК и/или посттранскрипцион"
рованию лизина14 (K14) в гистоне H3 (H3S10p"
ные модификации ГР. К настоящему времени
K14ac) с последующей индукцией ранних генов
описано 26 мутаций гена NR3C1 с потерей функ"
c(Fos и Egr(1 [24, 25]. КС, высвобождающиеся в
ции в контексте различных заболеваний, при
рамках стрессорного ответа и действующие че"
этом клинические проявления в основном ассо"
рез ГР, усиливают сигнал через путь
циированы с гиперкортизолизмом без регуля"
ERK1/2/MSK1"Elk"1 и таким образом повыша"
торной петли отрицательной обратной связи на
ют влияние на эпигенетические механизмы и
ГГНО. Некоторые полиморфизмы ГР
экспрессию генов. В контроле эпигенетических
(ER22/23EK, GR(9β) связаны с глюкокортико"
и транскрипционных ответов на психологичес"
идной резистентностью и с более близким к здо"
кий стресс важная роль принадлежит гамма"
ровому метаболическим профилем, в то время
аминомасляной кислоте (ГАМК) [24].
как другие связаны с гиперчувствительностью к
Таким образом, КС оказывают влияние на
КС (N363S, BclI) [20]. Недавно идентифициро"
мозг через геномные механизмы, в которых
ваны геномные локусы, на которых селективно
участвуют как ГР, так и МР, которые непосред"
связываются либо МР, либо ГР; показано, что в
ственно связываются с ДНК, а также через неге"
гиппокампе члены семейства транскрипцион"
номные и эпигенетические механизмы. КС, как
ных факторов NeuroD специфически ассоци"
через геном, так и негеномно, действуют синер"
ированы со связанной с МР ДНК. С использо"
гично с нейротрансмиттерами, нейротрофичес"
ванием мышей со специфическим нокаутом МР
кими факторами и другими медиаторами стрес"
в переднем мозге было показано, что NeuroD
са (рис. 2), определяя тем самым настоящие и
действует пермиссивно, усиливая опосредован"
будущие ответы организма на стрессогенное ок"
ную МР транскрипцию, и, видимо, работает
ружение.
именно этот механизм специфического связы"
Ключевая роль нейротрофических факто"
вания МР против ГР, а не конкуренция за связы"
ров, в первую очередь мозгового нейротрофи"
вание с ДНК [21].
ческого фактора (BDNF) в нейропластичности
БИОХИМИЯ том 84 вып. 11 2019
1628
ГУЛЯЕВА
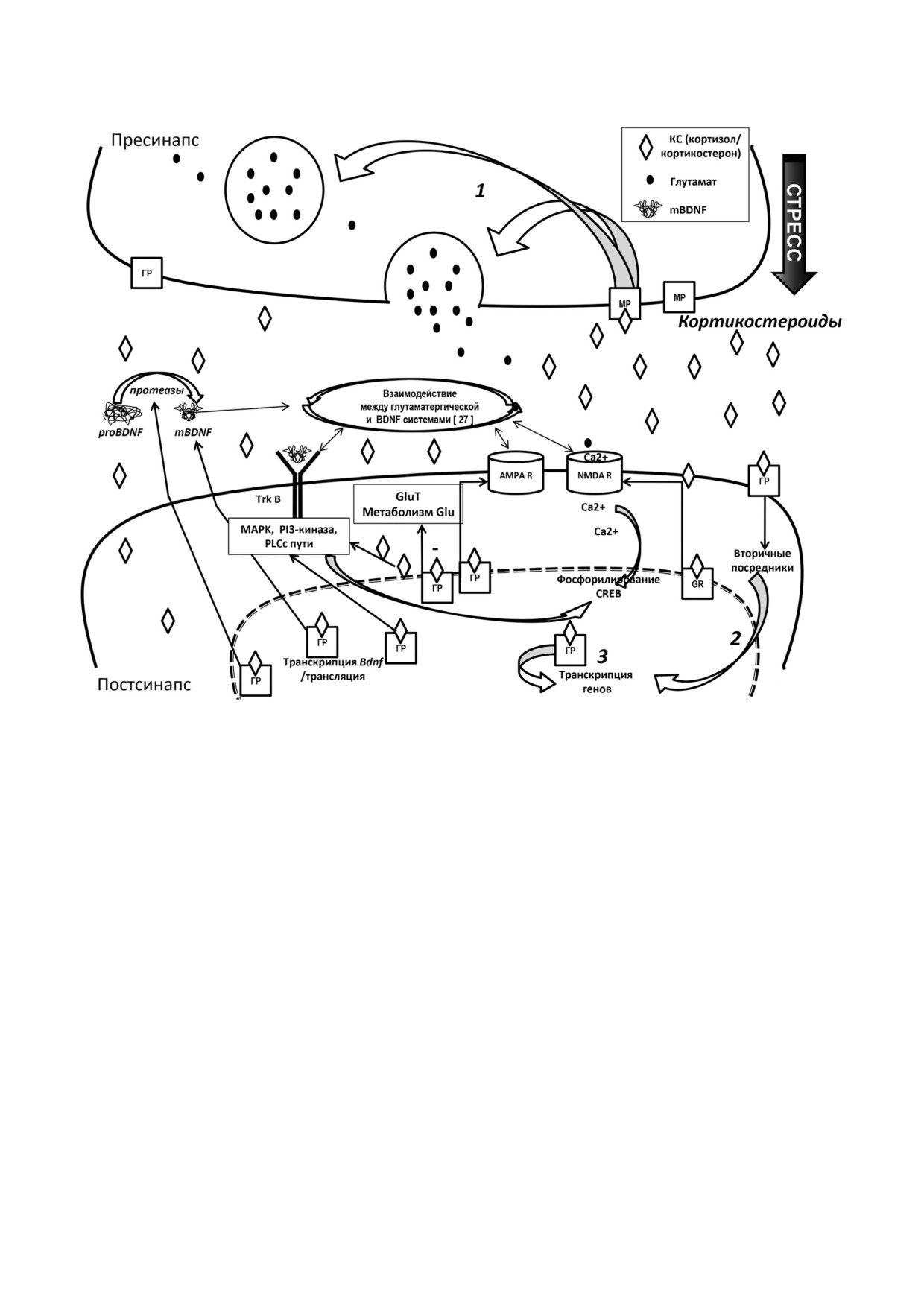
Рис. 2. Механизмы синаптических эффектов КС и взаимодействие с другими системами (глутаматергический синапс,
как пример) [15, 27, 29, 31]. Связывание КС с рецепторами лежит в основе взаимодействия и координации КС/ГГНО со
всеми основными системами, важными для нейрональной пластичности. КС связываются с разным сродством с МР и
ГР, которые существуют в цитоплазматической/ядерной и мембраносвязанной формах и опосредуют, соответственно,
отложенные и быстрые эффекты КС. Эти эффекты задействуют негеномные механизмы: (1) - опосредованы мембран"
ными рецепторами, непрямые геномные механизмы (2) - опосредованы мембранными рецепторами и вторичными по"
средниками и геномные механизмы (3) - опосредованы цитоплазматическими рецепторами, которые поступают в ядро
и действуют как факторы транскрипции. При реализации геномного механизма КС проникает через плазматическую
мембрану, входит в цитозоль и связывается с ГР, тем самым индуцируя гомодимеризацию (комплекс КС-ГР). Быстрые
эффекты КС включают усиление пресинаптического синтеза глутамата через пресинаптические мембранные МР; поз"
же постсинаптические цитозольные ГР регулируют ионотропные рецепторы N"метил"D"аспартата (NMDAR) и α"ами"
но"3"гидрокси"5"метил"4"изоксазолпропионовой кислоты (AMPAR), усиливая их встраивание в постсинаптичекую
мембрану, а также метаболизм и транспорт глюкозы (Glu), снижая активность ее мембранных транспортеров (GluT).
КС регулируют синтез, процессинг, траффик, секрецию и сигналинг мозгового нейротрофического фактора (BDNF)
(детали в тексте). Множественные взаимодействия системы BDNF/TrKB с глутаматной системой (см. детали [27]) вмес"
те с участием КС под контролем обеих систем позволяют сбалансировать и отрегулировать процессы, обеспечивающие
нейропластичность
требует тонко настроенного взаимодействия со"
экспрессия ГР, МР, а также BDNF и его рецеп"
существующих в ЦНС систем BDNF и КС. Сиг"
тора, тропомиозин"зависимой киназы B (TrkB),
нал КС опосредует регуляцию стрессорного от"
что обеспечивает эффективное взаимодействие
вета на поддержание гомеостаза, в то время как
между системами [26]. Продолжительные эф"
сигнал нейротрофинов играет принципиальную
фекты стресса (включая ранний стресс) на КС и
роль в росте нейронов и является ключевым для
сигнальные пути нейротрофинов проявляются в
аксонального наведения и синаптической ин"
уязвимых для стресса регионах гиппокампа. Не"
теграции. В гиппокампе обнаружены высокая
давно мы проанализировали многочисленные
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1629
взаимодействия между системами BDNF и глу"
после обучения сопряжена с усилением связан"
тамата [27], а теперь обсудим, как функциони"
ного с ростом и выживанием пути BDNF/CREB
рование BDNF/TrkB координируется при помо"
(cAMP response element"binding protein), кото"
щи КС в глутаматных синапсах. На самом деле,
рый, как известно, является общим механиз"
синергизм между сигналами TrkB и ГР является
мом, необходимым для формирования долго"
ключевым фактором, определяющим ответ
временной памяти.
клетки на стресс [28]. КС изменяют экспрессию
Все вышесказанное относится в основном к
и сигнал BDNF (рис. 2). Поскольку BDNF, не"
глутаматергическим синапсам, которые, как
обходимый для реализации нейропластичности,
было предположено 30 лет назад, тесно связаны
способствует выживанию клеток, усиливает
с КС и повреждением гиппокампа [33]. В насто"
нейрогенез в гиппокампе и клеточную возбуди"
ящее время показаны многочисленные эффек"
мость, было предположено, что специфические
ты стресса на возбуждающую трансмиссию и
отрицательные эффекты КС могут быть опосре"
синаптическую пластичность в гиппокампе, а
дованы снижением экспрессии и сигнала
также критическая роль взаимодействия стресса
BDNF. Suri и Vaidya суммировали данные о вли"
и глутаматергических нейронов гиппокампа в
янии КС на систему BDNF в гиппокампе на
когнитивных и эмоциональных последствиях
различных уровнях, от хорошо документиро"
аверсивных стимулов. Накапливается все боль"
ванных вызванных КС изменений в мРНК
ше свидетельств того, что острый и хронический
BDNF до результатов исследований сигнальной
стресс через высвобождение КС вызывает изме"
трансдукции через рецепторы BDNF [29].
нения глутаматергической нейротрансмиссии в
Изменения уровней BDNF жизненно важ"
гиппокампе и префронтальной коре, влияя, та"
ны. Противодействуя неблагоприятным эффек"
ким образом, на ключевые аспекты когнитивно"
там избыточной сигнальной трансдукции КС,
го процессинга [34]. Дисфункцию глутаматерги"
индуцированной стрессом, нарушения системы
ческой трансмиссии считают центральной в
BDNF, фактора сопротивляемости, вовлечены в
связанных со стрессом психических заболева"
психопатологию, вызванную хроническим
ниях. Как и система BDNF, возбуждающий ней"
стрессом [30]. Хронический иммобилизацион"
ромедиатор глутамат вовлечен, преимуществен"
ный стресс приводит к повышению экспрессии
но, в феномены клеточной и синаптической
мРНК и белка BDNF в некоторых областях моз"
пластичности, а также в феномен памяти. Связи
га, например, в базальной латеральной амигда"
между двумя системами многочисленны и обес"
ле, но снижает экспрессию в других регионах,
печивают взаимную регуляцию глутаматерги"
например, в области CA3 гиппокампа, при этом
ческой системы и системы BDNF. Имеющиеся
плотность дендритных шипиков возрастает или
данные позволяют предположить, что именно
снижается вместе с подобными изменениями
сложная и хорошо скоординированная природа
BDNF [31]. Ввиду мощного влияния BDNF на
этих связей обеспечивает оптимальную синап"
рост дендритных шипиков эти наблюдения поз"
тическую и клеточную пластичность нормаль"
воляют предположить, что фундаментальная
ного мозга. Обе системы связаны с патогенезом
причина направления и степени изменения
депрессии, и нарушение прочных и хорошо сба"
плотности дендритных шипиков в определен"
лансированных связей между ними приводит к
ном отделе мозга при стрессе - это локальное
неблагоприятным изменениям нейронной
изменение уровня BDNF, наиболее вероятно, за
пластичности, лежащим в основе депрессивных
счет вызванного стрессом высвобождения КС,
расстройств и других психических заболеваний
которые поступают в нейрон и изменяют
[34]. Недавно были раскрыты механизмы, опос"
экспрессию гена BDNF. Различия в распределе"
редующие влияние стресса и КС на глутаматную
нии ГР и МР и нижележащих (downstream) си"
трансмиссию, включая эффекты на высвобож"
стем могут лежать в основе противоположных
дение глутамата, глутаматные рецепторы, кли"
эффектов на экспрессию гена BDNF. Другая
ренс и метаболизм глутамата (см. обзор [34]). Во
возможность регуляции - эпигенетический
влиянии стресса на возбуждающие синапсы
контроль транскрипции BDNF (различия в сте"
вовлечен сложный комплекс нейрогормонов и
пени метилирования и ацетилирования) в раз"
нейромедиаторов, среди которых КС играют
ных частях мозга после стресса [31]. Важно, что
ключевую роль. Chaouloff и Groc [35] проанали"
позитивные эффекты активации ГР на консоли"
зировали тонические и базовые эффекты КС на
дацию памяти критически требуют вовлечения
гиппокампальную возбуждающую трансмис"
пути BDNF. Finsterwald et al. [32] предложили
сию, перемещение и экспрессию глутаматных
гипотезу о том, что умеренный стресс способ"
рецепторов и синаптическую пластичность,
ствует формированию прочной долговременной
уделяя особое внимание их временнóму харак"
памяти, поскольку активация ГР гиппокампа
теру (быстрые и преходящие эффекты с после"
БИОХИМИЯ том 84 вып. 11 2019
1630
ГУЛЯЕВА
дующими медленными и долго сохраняющими"
структуры/органа. В гиппокампе избыток КС
ся геномными эффектами). Многие эффекты
может стимулировать провоспалительные гены
КС на глутаматергическую систему опосредова"
]37]. Принимая во внимание, что большие дозы
ны их связыванием с цитозольными МР и ГР, их
кортизола обладают провоспалительным
последующей транслокацией в ядро и регуляци"
действием, ситуация представляется достаточно
ей транскрипции целевых генов, а быстрые эф"
сложной [38] (рис. 3). Феноменологически из"
фекты реализуются через мембранные рецепто"
быточная аккумуляция КС в мозге представля"
ры КС (рис. 2). Этот важнейший «регуляторный
ется тесно связанной с провоспалительными со"
треугольник», объединяющий ГГНО/КС/ре"
бытиями, и такая связь обнаружена в различных
цепторы КС, систему BDNF и глутаматергичес"
моделях депрессии на грызунах [39]. Стресс вы"
кую систему, представляет собой сердцевину
зывает секрецию цитокинов, а цитокины могут
сложной многоуровневой регуляции сигналь"
вызывать гормональные изменения, сходные с
ных и метаболических процессов в гиппокампе,
таковыми после стресса. Продолжительный
лежащих в основе способности организма адап"
стрессорный ответ и избыточная продукция ци"
тироваться к вызовам существования и поддер"
токинов нарушают нейронную пластичность и
живать равновесие нейропластичности, нару"
повышают активность ГГНО, сенситизируя ее
шение которого приводит к различным цереб"
ответ на цитокины и стресс. Стрессовая ситуа"
ральным патологиям.
ция и индуцированное высвобождение КРГ вы"
зывают в мозге провоспалительный ответ, ха"
рактеризующийся комплексным высвобожде"
КС, НЕЙРОВОСПАЛЕНИЕ
нием различных медиаторов воспаления, участ"
И ГИППОКАМП
вующих во множественных взаимодействиях
между нейроэндокринной и нейроиммунной
Все нейродегенеративные заболевания
системами, включая цитокины, простаноиды,
действительно зависят от нейровоспаления.
оксид азота (NO) и факторы транскрипции [40].
Давид Перлмуттер
Катехоламины и КС играют критическую
роль в регуляции цитокинов мозга после стрес"
Более 20 лет назад было установлено, что
са. Нейроэндокринные ответы на психологи"
провоспалительные цитокины, высвобождае"
ческие стрессоры влияют на быструю и длитель"
мые активированными вспомогательными им"
ную регуляцию провоспалительных цитокинов
мунными клетками, оказывают сильный эф"
ИЛ"1β, ФНО"α и ИЛ"6 в гиппокампе, гипотала"
фект на мозг, включая активацию ГГНО, повы"
мусе и префронтальной коре; эта регуляция из"
шение температуры и поведенческую депрес"
меняется во времени при повторных стрессах
сию. Эти эффекты опосредованы цитокинами,
(см. обзор [41]). Центральные катехоламины
которые синтезируются и высвобождаются в
индуцируют секрецию ИЛ"1β из микроглии,
мозге в ответ на цитокины, высвобождаемые на
при этом ИЛ"1β является ключевым фактором
периферии. КС обладают мощным регулятор"
для дальнейшей активации микроглии и прив"
ным действием на синтез цитокинов активиро"
лечения в мозг моноцитов. Как правило, повы"
ванными макрофагами и моноцитами; КС так"
шенные КС ингибируют продукцию цитокинов
же регулируют синтез и действие цитокинов в
в мозге за счет подавления норадренергических
мозге, и это имеет важные функциональные
нейронов голубого пятна и ингибирования сиг"
следствия [36]. Все больше данных свидетель"
нального пути NF"κB (nuclear factor kappa"light"
ствуют, что индуцированные стрессом цитоки"
chain"enhancer of activated B cells). Однако при
ны мозга важны для этиологии практически
повторных стрессах КС и периферические кате"
всех патологий мозга, и что усиленное воспале"
холамины способствуют воспалительным отве"
ние и гиперактивность ГГНО часто связаны, хо"
там на будущие стимулы, инициируя выход мо"
тя молекулярные механизмы этой связи до кон"
ноцитов из костного мозга, подавляя ингибитор"
ца не ясны.
ные рецепторы на микроглии и праймируя ней"
КС уже давно рассматривались как противо"
роиммунные воспалительные ответы, опосредо"
воспалительные и протекторные агенты из"за их
ванные периферическими моноцитами и мак"
способности ингибировать экспрессию генов
рофагами [42] (рис. 3).
провоспалительных медиаторов и других потен"
Гиппокамп селективно чувствителен к ней"
циально пагубных молекул. Тем не менее иссле"
ровоспалению, и это может быть одной из важ"
дования последних лет обнаружили ситуации, в
ных причин вовлеченности этой структуры в па"
которых эти гормоны могут действовать как
тогенез многих психических и неврологических
провоспалительные агенты, в зависимости от их
заболеваний (см. обзор [4]). Стрессорные фак"
дозы, периода экспозиции и анализируемых
торы вызывают зависимые от времени и стресс"
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1631
Рис. 3. Кортикостероиды и нейровоспаление: стресс индуцирует высвобождение гормонов ГГНО и усиливает экспрессию
цитокинов [3, 40, 41, 70, 73]. Стресс активирует ГГНО и симпатическую нервную систему (СНС). КС индуцируют акти"
вацию микроглии, которая высвобождает провоспалительные цитокины, в т.ч. IL"1β и хемокин CCL2 (C"C Motif
Chemokine Ligand 2). Этот цитокиновый ответ, в свою очередь, участвует в развитии реактивного эндотелия региональной
нейро"сосудистой системы. Как КС, так и СНС стимулируют продукцию праймированных миелоидных клеток
CD11+/CD45+ в костном мозге. Высвобождение миелоидных клеток в циркуляцию приводит к доставке этих клеток к ре"
активной нейро"сосудистой сети с последующими адгезией и диапедезом в мозге. Цитокины вызывают гиперкортизоле"
мию путем нарушения регуляции ГГНО, активируя ее напрямую или опосредованно, за счет модификации чувствитель"
ности ГР к КС, что приводит к гиперсекреции КС. Цитокины непосредственно стимулируют активацию ГГНО путем
внутренних и внешних относительно ГГНО механизмов и повышают ее чувствительность к последующим стрессорным
воздействиям. Формируется порочный круг (вверху справа): хроническое нейровоспаление ингибирует функцию ГР, ко"
торый, в свою очередь, усиливает активность провоспалительных цитокинов и усугубляет хроническое нейровоспаление
специфичные паттерны экспрессии цитоки"
терминирующего и праймирующего механиз"
нов/хемокинов в мозге, а связанные с воспале"
мов, участвующих в стрессе/адаптации (рис. 1, б).
нием гены положительно регулируются и демон"
Например, МР в гиппокампе спонтанно гипер"
стрируют уникальные профили экспрессии у са"
тензивных крыс линии SHR связаны с провос"
мок и самцов в зависимости от индивидуаль"
палительным трендом. У этих животных проде"
ных, кооперативных или антагонистических
монстрирован дисбаланс МР и ГР: экспрессия
взаимодействий между рецепторами стероид"
МР возрастает, и активация МР эндогенным
ных гормонов (эстрогеновый и ГР). Все больше
кортикостероном вызывает провоспалительный
свидетельств того, что нарушение баланса и взаи"
каскад, участвующий в активации микроглии,
модействия между ГР и NF"κB, эффектором
оверэкспрессии провоспалительных медиато"
иммунной оси, могут играть ключевую роль в
ров, регуляторном снижении противовоспали"
реализации отрицательных эффектов хроничес"
тельных факторов, что в конечном итоге приво"
кого стресса на настроение и поведение [43].
дит к повреждению ткани гиппокампа [45]. Вве"
В соответствии с гипотезой «баланса МР/ГР» [44]
дение ИЛ"1 вызывает длительное повышение
воспалительный ответ на повреждение опреде"
уровня кортикостерона; ИЛ"1 также влияет на
ляется сбалансированным действием, опосре"
функцию МР в гиппокампе и вызывает сдвиг в
дованным МР и ГР, в качестве инициирующего,
балансе МР/ГР, который может обусловливать
БИОХИМИЯ том 84 вып. 11 2019
1632
ГУЛЯЕВА
продолжительную активацию ГГНО при им"
востепенное значение для предупреждения це"
мунном ответе [11].
ребральных патологий.
Предполагается, что КС индуцируют синтез
и высвобождение из микроглии белка HMGB1
(high mobility group box"1), эндогенного сигнала
КС, КОГНИТИВНЫЕ ФУНКЦИИ
опасности или алармина, который передает сиг"
И ГИППОКАМП
нал через TLR2/TLR4, таким образом осущес"
твляя прайминг NLRP3 инфламмасомы. Когда
У меня не Альцгеймер. У меня парт"таймер.
уровни КС достигают критического порога, они
Глен Кэмпбелл
могут функционировать как алармин, индуци"
руя HMGB1 и подготавливая врожденную им"
Несколько десятилетий назад при исследо"
мунную систему организма (прайминг NLRP3
вании изображения мозга человека с использо"
инфламмасомы) для последующих иммунных
ванием МРТ было показано, что гиппокамп, наи"
изменений; это дает значительное преимущест"
более пластичная и уязвимая структура и ми"
во в выживании за счет усиления центрального
шень КС, в различных условиях подвергается
ответа врожденной иммунной системы и «бо"
селективной атрофии, в т.ч. при синдроме Ку"
лезненного поведения» (sickness response) на им"
шинга, посттравматическом стрессорном рас"
мунные вызовы [46].
стройстве, рецидивирующей депрессии, нор"
В мозге стресс повышает как уровнь КС, так
мальном старении, предшествующем депрес"
и норадреналина, и оба гормона влияют на ге"
сии, и при болезни Альцгеймера (БА). Сморщи"
номные и сигнальные каскады, ответственные
вание гиппокампа обычно сопровождается на"
за модуляцию прочности памяти. Глия - одна из
рушениями декларативной, эпизодической,
основных мишеней для индуцированных стрес"
пространственной и контекстуальной памяти, и
сом изменений, а астроциты, микроглия и оли"
изменения гиппокампа, как считают, являются
годендроциты вносят свой уникальный вклад в
нейронным субстратом для когнитивных изме"
обучение и память. Все 3 типа глии экспресси"
нений, связанных с различными патологиями
руют рецепторы КС и норадреналина и поэтому
[49]. Можно считать доказанной причинно"
являются непосредственными мишенями
следственную связь между продолжительным
действия стрессорных гормонов. Становится
стрессом, повышенными уровнями КС и когни"
понятным, что воспалительные цитокины и им"
тивными/психическими расстройствами, при
муномодуляторные молекулы, высвобождаемые
этом появляется все больше подтверждений
глией при стрессе, могут поддерживать многие
связей между хроническим стрессом/КС и ней"
поведенческие эффекты острого и хроническо"
родегенеративными заболеваниями, такими как
го стресса [47]. Микроглия, основной тип им"
БА и болезнь Паркинсона [50, 51].
мунных клеток в ЦНС, играет ключевую физио"
Пациенты с БА, болезнями Паркинсона и
логическую роль в поддержании гомеостаза, в
Гентингтона имеют хронически повышенный
первую очередь в ответе на стресс и опосредует
уровень кортизола, что указывает на изменения
синаптическую пластичность, обучение и па"
контроля ГГНО. В экспериментальных моделях
мять. Считают, что многие из этих эффектов за"
этих заболеваний хронический стресс или вве"
пускаются стрессорными сигнальными молеку"
денные КС усугубляют как клинические симп"
лами, прежде всего КС. Большинство исследо"
томы, так и нейродегенеративные процессы
ваний показало, что стресс вызывает значимое
[52]. В клинических исследованиях показано,
структурное ремоделирование микроглии и мо"
что при БА повышенный кортизол ассоцииро"
жет усиливать высвобождение из микроглии
ван с худшим уровнем общего когнитивного
провоспалительных медиаторов [39]. КС усили"
функционирования, а также с нарушенными в
вают рамификацию микроглии гиппокампa и
большей степени эпизодической памятью, ис"
могут модулировать возрастные изменения ее
полнительной функцией, языком, простран"
морфологии [48]. Вызванные стрессом измене"
ственной памятью, скоростью переработки ин"
ния микроглии не являются эпифеноменом, но
формации и социальной когницией [53]. Сход"
имеют значение для поведения; микроглия не"
ным образом хронически повышенный корти"
посредственно регулирует определенные аспек"
костерон или введение КС вызывают аномаль"
ты когнитивной функции и эмоций. Поскольку
ное поведение (депрессивноподобное, тревож"
нейровоспаление рассматривается как ключе"
ное) у животных. Более того, у когнитивно здо"
вой механизм повреждения нервных клеток и
ровых людей повышенный уровень кортизола
нейродегенерации, уточнение молекулярных
ассоциирован с повышенным риском когнитив"
механизмов, осуществляемого КС и его рецеп"
ного снижения и БА, а у пациентов с деменцией
торами контроля нейровоспаления, имеет пер"
и мягким когнитивным снижением (MCI) уров"
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1633
ни кортизола в цереброспинальной жидкости
Хронический стресс - фактор риска, моду"
выше, чем у когнитивно здоровых контролей
лирующий функцию микроглии, которая связа"
(см. обзор [53]). Повышенный уровень кортизо"
на с аккумуляцией Aβ, тау"патологией, нейро"
ла в цереброспинальной жидкости может быть
дегенерацией и потерей синапсов при БА, хотя
ассоциирован также с быстрым когнитивным
может играть и благоприятную роль, в частнос"
снижением при MCI.
ти в элиминации Aβ фагоцитозом [56]. Другие
Холинергические нейроны базального пе"
возможные механизмы, лежащие в основе учас"
реднего мозга, которые, как считается, опосре"
тия избытка КС в БА, включают их взаимодей"
дуют когнитивные функции, вовлечены в этио"
ствие с воспалительными медиаторами, нейро"
логию стресса, когнитивного старения, БА и
трансмиттерами и факторами роста.
других нейродегенеративных заболеваний. Эти
Факты о том, что нарушения ГГНО ассоци"
нейроны участвуют в стрессорном ответе и ког"
ированы с нарушениями памяти, а гиперкорти"
ниции и имеют проекции в различных областях
золемия - с атрофией гиппокампа, подтвержде"
неокортекса и лимбических структур, включая
ны многочисленными исследованиями, однако
гиппокамп, первичную мишень КС, связанную
последние данные указывают на более сложную
как с когнитивной функцией, так и с ответом
картину функционирования и патологии ГГНО
ГГНО. Между КС/МР/ГР и холинергической
в приобретении, извлечении и консолидации
системой существует связь, важная для объясне"
памяти. Эти данные включают: существование
ния холинергической нейродегенерации в ба"
«инвертированной U"образной связи» между
зальном переднем мозге в условиях индуциро"
стимуляцией ГР мозга и презентацией памяти;
ванного стрессом ускоренного когнитивного
неодинаковые ответы различных областей гип"
снижения и БА [54].
покампа на стимуляцию КС; потенциальную
Несомненно, высокий уровень кортизола
обратимость атрофии гиппокампa в некоторых
стимулирует различные сигнальные механиз"
условиях, хотя до сих пор неясно, является такая
мы, потенциально участвующие в нейродегене"
атрофия причиной или следствием этих патоло"
рации. КС могут оказывать нейротоксические
гических условий [57].
эффекты на гиппокамп, способствуя окисли"
Важно иметь в виду, что в зубчатой фасции
тельному стрессу и токсичности амилоидного
гиппокампа в течение всей жизни поддержива"
β"пептида (Aβ). БА, наиболее частая форма де"
ется образование новых нейронов. «Взрослый»
менции, характеризуется патологической акку"
нейрогенез тесно связан с функциями гиппо"
муляцией в нейронах гиппокампа и префрон"
кампа, в том числе обучением, памятью, регуля"
тальной коры двух белков, Aβ и гиперфосфори"
цией тревожности и механизмом обратной свя"
лированных форм связанного с микротрубоч"
зи стрессорного ответа. Изменения нейрогенеза
ками тау"белка. Aβ, пептид, высвобождаемый
участвуют в развитии болезней мозга, в первую
синапсами в физиологических условиях, пато"
очередь психических и нейродегенеративных.
логически аккумулируется в структурах мозга,
Стресс влияет на все процессы гиппокампаль"
вовлеченных в механизмы памяти, и представ"
ного нейрогенеза, включая образование, мигра"
ляет собой «токсический признак» БА. Олиго"
цию и выживание новых нейронов, и КС вовле"
мерная форма Aβ, как полагают, влияет на си"
чены в индуцированные стрессом нарушения
наптическую функцию. Все больше данных о
нейрогенеза у взрослых. Считается, что КС ока"
том, что ГР участвует в генерации олигомерно"
зывают отрицательный эффект на пролифера"
го Aβ, указывающих на прочное функциональ"
цию как эмбриональных, так и взрослых ней"
ное взаимодействие между ГР и Aβ в возбужда"
ральных стволовых клеток и предшественников,
ющих синапсах [55]. Несмотря на то, что в кли"
что связано с патофизиологией заболеваний
нических исследованиях до сих пор не доказана
мозга, таких как болезни депрессивного и аутис"
прямая причинно"следственная связь между
тического спектра, а также нарушениями обуче"
КС и патогенезом БА, данные доклинических
ния и памяти [58, 59]. Но это довольно односто"
исследований показывают, что повышенные
ронний взгляд, поскольку в зависимости от ти"
уровни КС ускоряют образование Aβ в живот"
па и силы стресса, а также стресс"реактивности
ных моделях БА, стимулируя амилоидогенный
организма, повышение уровня КС может усили"
путь и одновременно снижая клиренс Aβ через
вать нейрогенез в зубчатой фасции, хотя в дру"
транскрипционные механизмы, в которые вов"
гих ситуациях подавляет его. Эффекты острого
лечен ГР. При этом эффекты стресса на фосфо"
и умеренного стресса на «взрослый» нейрогенез
рилирование тау"белка, видимо, в основном
обычно непродолжительны и бывают быстро
опосредованы рецептором КРГ (CRFR1) и не
преодолены, однако следствием действия хро"
зависят от вызванного стрессом повышения КС
нического и более тяжелого стресса может быть
[51].
длительное снижение нейрогенеза [60]. В этих
8 БИОХИМИЯ том 84 вып. 11 2019
1634
ГУЛЯЕВА
обстоятельствах факторы, которые создают бу"
симого снижения сигнала ГГНО и повышенной
фер против подавляющего влияния повышен"
секреции КРГ из паравентрикулярного ядра и
ных КС, остаются неясными и подлежат иссле"
негипоталамических нейронов, в то время как
дованию [61]. Есть данные о прямых и непря"
гиппокамп в норме ингибирует вызванную
мых эффектах КС на пролиферацию нейраль"
стрессом активацию ГГНО [63] (рис. 1, a). Все
ных стволовых клеток и предшественников и
большее число исследований указывает на ано"
«взрослый» нейрогенез; сформулирована гипо"
мальное функционирование МР и ГР, как на ре"
теза о том, что ритмы и осцилляции КС в про"
шающий компонент патофизиологии депрес"
цессе активности ГГНО могут быть критичны"
сии. При хроническом стресс"индуцированном
ми для нормального нейрогенеза в гиппокампе
ингибировании системой обратной связи уров"
[59].
ни цитозольных ГР в гиппокампе и префрон"
Именно чувствительностью гиппокампа к
тальной коре, структурах, глубоко вовлеченных
стрессу объясняют негативное влияние стресса
в патогенез депрессии, существенно изменяют"
и стрессорных гормонов на когнитивные функ"
ся [40]. Длительные нарушения паттерна высво"
ции человека и животных. Однако за последние
бождения КС и реактивности их рецепторов
20 лет получены новые данные, которые пока"
приводят к структурным и функциональным из"
зывают, что стресс оказывает влияние на целый
менениям нейронов, которые могут вносить
ряд мнемонических корковых и подкорковых
вклад в проявление психопатологии [64].
структур мозга, а не только на гиппокамп [62],
Депрессия часто встречается при инфекци"
но обсуждение этих данных выходит за пределы
онных, аутоиммунных и нейродегенеративных
тематики данной статьи. В любом случае, боль"
заболеваниях, в то же время у больных депрес"
шинство эффектов КС на мозг осуществляется
сией нарушена иммунная функция [65]. Внача"
через МР и ГР, включая сложные и часто проти"
ле считали, что гиперкортизолемия вызывет по"
воположные действия на структуры мозга, во"
давление иммунной функции, но сейчас оче"
влеченные в различные когнитивные функции.
видно, что хронический стресс и депрессия вы"
зывают гипоактивность ГР в иммунных клетках
и лимбических областях мозга. В настоящее
КС, ДЕПРЕССИЯ,
время считают, что депрессия ассоциирована с
НЕЙРОВОСПАЛЕНИЕ
активацией некоторых звеньев клеточного им"
И ГИППОКАМП
мунитета, что приводит к гиперсекреции про"
воспалительных цитокинов и гиперактивности
Если бы мне пришлось определить большую
ГГНО, и такая иммунная активация вызывает
депрессию в одном предложении, я бы описал
«стресс"подобные» изменения поведения и ней"
ее как требующее сильного внешнего стимула
рохимические сдвиги [66]. Усиленное воспале"
генетическое/нейрохимическое расстройство,
ние и гиперактивность ГГНО - два признака
типичным проявлением которого является
большой депрессии часто ассоциированы: счи"
неспособность любоваться закатом.
тается, что развитие депрессии связано с нару"
Роберт М. Запольский
шением аллостаза и воспалительной активацией
иммунной системы. Это приводит к хроничес"
Лимбическая дисфункция и нарушение ре"
кому повышению уровней КС и провоспали"
гуляции ГГНО - ключевые черты аффективных
тельных цитокинов, индуцируя аллостатичес"
расстройств. Депрессия является лидирующей
кую нагрузку и последовательно нейродегенера"
причиной инвалидизации в мире, однако био"
цию, необратимые когнитивные нарушения и
логические и молекулярные механизмы, лежа"
перманентную неработоспособность [67]. По"
щие в ее основе, остаются не до конца понятны"
вышенная чувствительность к воспалительным
ми. Стресс, фактор, предрасполагающий к раз"
и аутоиммунным заболеваниям при депрессии
витию депрессии, приводит к активации ГГНО
связана с нарушениями активности ГГНО, ко"
и высвобождению КС. Кроме депрессии, одно"
торые могут быть связаны с гипокортизолизмом
временные изменения стрессорного ответа и
или глюкокортикоидной резистентностью вслед"
иммунитета считают факторами целого ряда
ствие нарушения локальных факторов, влияю"
связанных со стрессом нейропсихических забо"
щих на доступность и функцию КС, включая ГР
леваний, в том числе тревожных и посттравма"
[68].
тических стрессорных расстройств.
Как воспаление, так и стрессорная актива"
Гиперактивность ГГНО, один из фундамен"
ция ГГНО - нормальные компоненты стрессор"
тальных биологических механизмов большой
ного ответа, но если стресс и гиперкортизоле"
депрессии, является следствием ослабления ин"
мия продолжительны, периферические макро"
гибирования по типу обратной связи КС"зави"
фаги активируются, как и центральная микро"
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1635
глия, вследствие чего нейронные сети повреж"
руя триптофан"кинурениновый путь, приводя"
даются, и их функция нарушается (рис. 3) [69].
щий к синтезу нейротоксичного агониста
Поскольку между эндокринной системой, им"
NMDA"рецептора хинолиновой кислоты и
муной системой и ЦНС существуют многочис"
3"гидроксикинуренина. Тем самым они усили"
ленные взаимосвязи, активация воспалительно"
вают окислительный стресс, способствующий
го ответа может влиять на нейроэндокринные
нейродегенерации и нарушению пластичности
процессы и наоборот.
мозга [69]. Считают, что формируется порочный
Физические, психологические и комбини"
круг, тесно сплетающий между собой нейропси"
рованные стресс"факторы вызывают провоспа"
хические заболевания и воспаление, которые
лительный ответ в мозге и других системах, ха"
усиливают друг друга внутри одного двунаправ"
рактеризующихся комплексным высвобожде"
ленного цикла. Депрессия облегчает воспали"
нием ряда медиаторов воспаления. Провоспа"
тельные реакции, а воспаление стимулирует
лительные цитокины, включая ИЛ"1β, ИЛ"6 и
депрессию и другие нейропсихические заболе"
ФНО"α, вовлечены в этиологию клинической
вания. Потенциальными триггерами хроничес"
депрессии и тревожных расстройств. КС инги"
кого воспаления являются изменения нейроэн"
бируют (через ГР), а катехоламины стимулиру"
докринной регуляции, метаболизма, дие"
ют (через β"адренергические рецепторы) обра"
та/микробиота и негативный поведенческий
зование ИЛ"1β [40]. Воспалительные цитокины
опыт, включая ранний [71]. Сложность механиз"
вовлечены в депрессию за счет их действия на
мов, лежащих в основе депрессии, затрудняет
3 основных пути в мозге: нейроэндокринного,
создание общей концепции ее патогенеза, а сов"
истощения нейромедиаторов и процесса ней"
ременные теории о гиперсекреции КС и серото"
ропрогрессии. За счет нарушения регуляции
нинергической дисфункции не дают достаточ"
ГГНО цитокины напрямую вызывают гиперкор"
ных объяснений о природе этого заболевания.
тизолемию активацией ГГНО и опосредованно,
Поскольку воспалительные и нейродегенера"
за счет изменения чувствительности ГР к корти"
тивные процессы играют важную роль в депрес"
золу, что приводит к его гиперсекреции (рис. 3).
сии, усиленная нейродегенерация при депрес"
Цитокины способствуют истощению централь"
сии может, по крайней мере, частично, быть
ного серотонина, снижая его синтез и повышая
причиной воспалительных процессов, что и
обратный захват. Они также могут вызывать ис"
послужило основой для новой гипотезы, кото"
тощение нейротрофических факторов, в первую
рая, по сути, является продолжением цитокино"
очередь BDNF, который играет нейропротектор"
вой гипотезы. Постулируется, что внешние, в
ную и, следовательно, антидепрессивную роль.
т.ч. психосоциальные стрессоры и внутренние
Цитокины активируют клеточные каскады, вы"
стрессоры, в т.ч. органические воспалительные
зывающие эксайтотоксичность и апоптоз и ин"
нарушения или состояния, могут вызывать деп"
гибируют нейрогенез в гиппокампе [70].
рессию, способствуя развитию воспалительных
Важно отметить, что депрессия коморбидна
процессов в мозге [72].
многим другим состояниям, сопровождающим"
ГР гиппокампа и ГГНО тесно связаны с про"
ся нейродегенерацией. Простое представление,
воспалительными цитокинами и нейровоспале"
что КС индуцируют нейродегенерацию, не дос"
нием. Повышенные уровни провоспалительных
таточно точно; скорее повышенные уровни ци"
цитокинов и функциональная резистентность
токинов, особенно в ситуации глюкокортикоид"
ГР, наиболее широко исследованные факторы
ной резистентности, являются основными фак"
патофизиологии депрессии, образуют пороч"
торами патогенеза, а хронические воспалитель"
ный круг (рис. 3), в котором хроническое нейро"
ные изменения в этой ситуации могут представ"
воспаление ингибирует функцию ГР, что, в свою
лять собой общий фактор, определяющий по"
очередь, повышает активность провоспалитель"
вышенную уязвимость пациентов с депрессией
ных цитокинов и усиливает хроническое нейро"
к последующему развитию нейродегенератив"
воспаление. С другой стороны, нейровоспале"
ных изменений [65]. Пациенты с депрессией де"
ние вызывает дисбаланс между окислительным
монстрируют все кардинальные черты воспале"
стрессом и антиоксидантными системами, ко"
ния, включая повышенный уровень циркулиру"
торые также связаны с депрессией. Цитокины и
ющих индукторов воспаления, активированные
ГР -важные факторы депрессии, однако они
сенсоры и медиаторы воспаления, воздействую"
являются принципиальными компонентами
щие на все ткани. Провоспалительные цитоки"
сложной системы воспалительных и эндокрин"
ны модулируют настроение и когницию, сни"
ных взаимодействий, а не играют независимые
жая уровни моноаминов, активируя нейроэн"
роли [73, 74].
докринные ответы, способствуя эксайтотоксич"
Хронический стресс в ранний период жизни
ности (повышенные уровни глутамата), активи"
и у взрослых снижает экспрессию ГР, в частнос"
БИОХИМИЯ том 84 вып. 11 2019
8*
1636
ГУЛЯЕВА
ти, через эпигенетические механизмы и усили"
ванная КС гибель клеток гиппокампа пол"
вает экспрессию ко"шаперона гена FKBP5, ко"
ностью ответственна за соответствующие ког"
торый ограничивает активность ГР, лимитируя
нитивные и эмоциональные дефициты, спра"
транслокацию рецепторного комплекса в ядро.
ведливо лишь частично [78]. Важную роль в аф"
Другой механизм изменения реактивности ГР -
фективных расстройствах, в частности депрес"
это состояние фосфорилирования рецептора,
сии, играет нейрогенез. Специфическая субпо"
которое регулирует активацию, субклеточную
пуляция нейронов гиппокампа, гранулярные
локализацию и транскрипционную активность.
клетки зубчатой фасции, постоянно функцио"
Фосфорилирование ГР представляет собой важ"
нирующей нейрогенной ниши, более чувстви"
ный механизм взаимодействия между трансдук"
тельна к избытку КС, хотя механизмы этой уни"
цией нейротрофического сигнала и ГР"зависи"
кальной уязвимости до сих пор не ясны. Таким
мой транскрипцией, объединяющих два важных
образом, КС через МР и ГР влияют не только на
фактора психических заболеваний. Одним из
гибель клеток, но и на их рождение и дифферен"
нижележащих относительно ГР генов, который
цировку, при этом занятость МР необходима
может принимать участие в связанных со стрес"
для их выживания. На модели хронического
сом изменениях, является ген сыворотко" и
непредсказуемого стресса было показано, что
глюкокортикоид"индуцируемой киназы
1
МР"зависимый нейрогенез тесно связан с тре"
(SGK1); ее экспрессия существенно возрастает
вого"подобным поведением [79]. В то время как
после хронического стресса у грызунов, а также
избыточная занятость ГР лигандом в отсутствии
в крови нелеченных пациентов с депрессией.
активации МР может вызывать гибель нейро"
Повышение активности SGK1 в итоге может
нов, дополнительная занятость МР обычно при"
снижать нейрогенез в гиппокампе и участвовать
водит к менее тяжелым и потенциально обрати"
в структурных аномалиях, отмеченных у паци"
мым последствиям, ограничивающимся атро"
ентов с депрессией [75]. Стресс динамически
фией дендритов и потерей синаптических кон"
регулирует сети ко"экспрессии ГР"зависимых
тактов. Причиной снижения объема гиппокам"
основных генов риска большой депрессии, а из"
па при депрессии может быть, опосредованное
менения в структуре корреляций функциональ"
ГР, существенное повреждение зубчатой фасции
ных генетических вариантов, составляющих от"
[80].
вечающую на КС ко"экспрессионную сеть, воз"
можно, вносят вклад в риск заболеваний, свя"
занных с ранней детской травмой. Часть генети"
ДЕПРЕССИЯ, ДЕМЕНЦИЯ
ческих вариантов может быть связана с риском
И ПОВРЕЖДЕННЫЙ ГИППОКАМП:
депрессии за счет изменения на сетевом уровне
КОНСТРУКЦИЯ «ДВА В ОДНОМ»
эффектов раннего и взрослого социального
опыта на образование и структуру нейронных
Мне показалось, что в какой"то момент
сетей [76].
это было как будто у меня два заболевания, од"
Аллостатическая перегрузка, которая может
но - болезнь Альцгеймера, а второе -
иметь место при хроническом стрессе, способна
знание о том, что у меня болезнь Альцгеймера.
изменять ГГНО путем эпигенетичеких модифи"
Терри Пратчетт
каций генов в гиппокампе, гипоталамусе и дру"
гих отвечающих на стресс регионах мозга [77].
Депрессия тесно связана с когнитивными
Эпигенетические процессы могут играть важ"
нарушениями и нейродегенерацией, а потеря
ную роль в этиологии стресс"зависимых психи"
нейронов является общей чертой большой деп"
ческих заболеваний, таких как большое депрес"
рессии и деменции. Клиницисты рассматрива"
сивное и тревожные расстройства. Психологи"
ют депрессию, как продром и компонент БА;
ческие стрессогенные события оказывают дол"
она может также быть триггером начинающейся
говременное влияние на поведение путем изме"
БА [81]. Обсуждается, что либо существует об"
нения функционирования гиппокампa за счет
щее патологическое событие, лежащее в основе
глутаматергических и вызванных КС изменений
и большой депрессии и БА, либо депрессия мо"
в эпигенетической регуляции транскрипции ге"
жет сенситизировать мозг ко второму событию
нов, которые модулируются локальными
(«hit»), которое и запускает БА. Депрессия мо"
ГАМКергическими интернейронами и лимби"
жет ускорять старение мозга за счет изменения
ческими афферентными входами [24].
метилирования ДНК, повышения уровня кор"
Влияние КС на гиппокамп"зависимые про"
тизола, но снижения дигидроэпиандростерона,
цессы обучения и памяти не подлежит сомне"
и таким образом увеличивает риск БА. Обнару"
нию, хотя, согласно данным последних иссле"
женные до настоящего момента гены"предикто"
дований, представление о том, что индуциро"
ры БА (например, APOEε4) не являются факто"
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1637
рами риска депрессии, а гены, вовлеченные в
рацию поврежденных нейронных сетей. Нейро"
депрессию (например, SLC6A4), не являются
токсические конечные продукты триптофан"ки"
факторами риска БА, поэтому общая генетичес"
нуренинового пути, в частности, хинолиновая
кая предрасположенность представляется мало"
кислота, аккумулируются в астроцитах и нейро"
вероятной [82]. Система КС нарушена и при БА,
нах как при депрессии, так и при деменции. Та"
и при депрессии; КС оказывают значимое ней"
ким образом, усиленная нейродегенерация,
родегенеративное действие на гиппокамп, об"
сниженные нейропротекция и нейрорепарация
ласть раннего отложения Aβ, а редкие генети"
являются общими патологическими признака"
ческие варианты ферментов, регулирующих КС
ми большой депрессии и деменции, и это позво"
(например, 11HSD), предрасполагают к БА. КС
ляет объяснить, почему депрессия часто являет"
влияют на нейрогенез и пластичность в гиппо"
ся прелюдией к деменции [84].
кампе и, следовательно, ухудшают эпизодичес"
Все эти данные и их интерпретация связаны
кую память, основной симптом БА, и эмоцио"
с поиском общих звеньев в молекулярных меха"
нальный статус, основной симптом депрессии.
низмах деменции и депрессии, а также возмож"
Гиппокамп взрослого мозга содержит ство"
ных связей между нарушениями КС и хроничес"
ловые клетки и нейрональные предшественни"
ким воспалением с изменениями структуры
ки, которые в течение всей жизни сохраняют
мозга, которые могли бы запустить нейродеге"
способность самообновления и дифференци"
неративные изменения, ассоциированные с БА
ровки в различные клеточные линии, такие как
и деменциями другого типа, а также с большой
нейроны, астроциты и олигодендроциты. Важ"
депрессией. В целом правильные, эти рассужде"
но, что эти клетки вносят существенный вклад в
ния не содержат основного важного утвержде"
вышеупомянутые функции гиппокампа, в то
ния: структура гиппокампа определяет вовлече"
время как разные стрессоры и КС влияют на
ние этого региона мозга как в эмоциональное
пролиферацию, дифференцировку и судьбу
поведение, так и в когнитивную функцию. Гип"
этих клеток [83]. Нарушения КС при депрессии
покамп - не гомогенная область мозга, и имен"
ингибируют нейрогенез, однако вклад снижен"
но его сложное устройство лежит в основе важ"
ного нейрогенеза в БА до сих пор обсуждается.
ной роли и функциональной плейотропности
Тем не менее, как КС, так и цитокины снижают
гиппокампа. Говоря проще, несложная парадиг"
уровень BDNF, вовлеченного в гиппокампаль"
ма «когнитивная функция контролируется дор"
ный нейрогенез, депрессию и деменцию, и это
сальным (задним) гиппокампом, а эмоции
укрепляет мнение о том, что депрессия с нару"
контролируются вентральным (передним) гип"
шенными КС может быть фактором риска БА.
покампом» приписывает когнитивные рас"
Хроническое воспаление, центральное звено
стройства/деменцию в основном дорсальному, а
в патогенезе большой депрессии, может пред"
развитие эмоциональных нарушений, включая
располагать пациентов с депрессией к последу"
депрессивное поведение, по крайней мере на
ющим нейродегенеративным изменениям. В
начальном этапе, ассоциировано с вентральным
настоящее время имеются клинические данные
гиппокампом (см. обзор [4]). Иными словами, в
о том, что депрессия - это частый предшествен"
здоровом мозге гиппокамп работает, как систе"
ник БА и может быть ранним проявлением де"
ма «два в одном», контролируя когницию и эмо"
менции до появления явных когнитивных сим"
ции, поэтому повреждение гиппокампа может
птомов [81]. Цитокины, включая ИЛ"1β, ИЛ"6 и
стать основой и для когнитивных, и для эмоцио"
ФНО"α, повышаются в крови в некоторых слу"
нальных расстройств. Этот простой взгляд мо"
чаях депрессии, а повышенные цитокины - это
жет быть базисом как для понимания коморбид"
известный фактор риска БА, при этом как цито"
ности депрессии и деменции, так и для исследо"
кины, так и КС потенциально оказывают про"
вания причинно"следственных связей между
воспалительное действие в мозге [81, 82]. Пред"
двумя патологиями и механизмов их ассоциа"
полагается, что прогрессирование от депрессии
ции. Гипотеза, которую мы предложили ранее
к деменции может быть результатом активации
[4], является логическим продолжением такого
макрофагов в крови и микроглии в мозге, сопро"
взгляда: механизмы нарушенного стрессорного
вождающейся высвобождением провоспали"
ответа (включая взаимодействие высвобождае"
тельных цитокинов. Эти цитокины стимулиру"
мых КС с гиппокампальными рецепторами и
ют каскад воспалительных изменений (повыше"
последующие воспалительные события) участ"
ние простагландина E2, оксида азота и дополни"
вуют в дистантном повреждении гиппокампа,
тельных провоспалительных цитокинов) и ги"
лежащем в основе отсроченной деменции и деп"
персекрецию кортизола. Последний ингибирует
ресии, индуцированных фокальным поражени"
синтез белка, за счет этого снижая синтез ней"
ем мозга (например, инсультом или черепно"
ротрофических факторов и предотвращая репа"
мозговой травмой).
БИОХИМИЯ том 84 вып. 11 2019
1638
ГУЛЯЕВА
АНОМАЛЬНЫЙ СТРЕССОРНЫЙ ОТВЕТ
ное высвобождение кортизола в течение дли"
И ПЕРИОДЫ СТРЕССА У ПАЦИЕНТОВ
тельного периода) были более подвержены ког"
ПОСЛЕ ФОКАЛЬНЫХ ПОРАЖЕНИЙ МОЗГА
нитивному снижению после острого инсульта
или транзиторной ишемической атаки [89].
Стресс - это не состояние души...
Можно было бы предположить, что предыду"
он измеряем и опасен.
щий уровень стресса и есть наиболее важный
Роберт М. Запольский
фактор, однако результаты следующей работы
показали, что высокий вечерний уровень корти"
Наше видение зависимых от КС механиз"
зола в слюне после инсульта позволяет предска"
мов, лежащих в основе когнитивных и эмоцио"
зать худший когнитивный исход, который, оче"
нальных расстройств после фокального повреж"
видно, имеет отношение к нарушенной актив"
дения мозга, подтверждается клиническими
ности ГГНО [90]. Эти данные подтверждают
данными. Результаты продолжающегося иссле"
важность индивидуальных особенностей функ"
дования TABASCO (Tel"Aviv Brain Acute Stroke
ционирования ГГНО для развития постинсульт"
Cohort) заслуживают детального обсуждения в
ных когнитивных нарушений.
связи с нашей концепцией [4]. TABASCO -
Постинсультная депрессия (PSD) - наибо"
проспективное когортное исследование, в кото"
лее частое психическое нарушение после ин"
рое последовательно набираются пациенты с
сульта, которое независимо коррелирует с нега"
первым умеренным инсультом. Его дизайн был
тивным клиническим исходом. Идентификация
разработан, чтобы оценить ассоциации между
специфических биомаркеров была бы полезна
демографическими, психологическими, воспа"
для повышения чувствительности диагностики
лительными, биохимическими, нейроимиджин"
PSD и раскрытия патофизиологических меха"
говыми и генетическими маркерами, измерен"
низмов заболевания. В соответствии с данными
ными в острой фазе и в последующем долговре"
проведенного недавно мета"анализа, выявлены
менном периоде [85]. Несмотря на то, что ин"
следующие кандидаты для разделения пациен"
сульт является основной причиной когнитив"
тов с PSD и пациентов после инсульта без деп"
ных нарушений и деменции, механизмы этих
рессии: гиперкортизолемия и сниженный ответ
постинсультных осложнений в полной мере не
кортизола при пробуждении, повышенные ран"
ясны.
ние маркеры воспаления (высокочувствитель"
В исследовании TABASCO объем гиперин"
ный C"реактивный белок, ферритин, неоптерин
тенсивности белого вещества и объем серого ве"
и глутамат), провоспалительные цитокины в
щества были независимо ассоциированы с ког"
сыворотке крови (ФНО"α, ИЛ"1β, ИЛ"6, ИЛ"18,
нитивными нарушениями, в то время как новое
интерферон"γ), а также сниженные провоспа"
детектированное ишемическое поражение не
лительные/ противовоспалительные соотноше"
было маркером когнитивного нарушения и рис"
ния (ФНО"α/ИЛ"10, ИЛ"1β/ИЛ"10, ИЛ"6/
ка постинсультного когнитивного нарушения
ИЛ"10, ИЛ"18/ИЛ"10, интерферон"γ/ИЛ"10);
[86]. Неясно почему пациенты со сходными ра"
концентрация BDNF в сыворотке крови; гено"
диологически инфарктами могут иметь различ"
типы BDNF met/met; повышенный статус мети"
ные клинические проявления - от полного ис"
лирования промотора BDNF [84]. Важно, что
чезновения симптомов до серьезных неврологи"
симптомы депрессии после инсульта или тран"
ческих последствий [87]. Независимо от объема
зиторной ишемической атаки повышают риск
поражения, повышенный воспалительный от"
развития когнитивного и функционального
вет был связан с более тяжелым течением забо"
снижения в двухлетнем периоде [91]. В ситуа"
левания [87]. Пациенты с небольшим или уме"
ции, когда гиппокамп хронически перегружен
ренным инсультом и предсуществующими пов"
КС, он может быть праймирован или даже пов"
реждениями белого вещества более уязвимы к
режден перед фокальным поражением мозга.
развитию когнитивных нарушений, независимо
Исследование TABASCO показало предсказа"
от их новых ишемических поражений [88]. Все
тельную ценность средней диффузивности гип"
вышеуказанное подтвержает наше представле"
покампа; предположительно эти изменения
ние о том, что когнитивные нарушения в буду"
предшествуют и вносят вклад в волюметричес"
щем могут быть ассоциированы с дистантным
кие изменения и сдвиги коннективности в гип"
повреждением гиппокампа, которое не обяза"
покампе и потенциально могут быть использо"
тельно сильно зависит от тяжести первичного
ваны, как маркер для ранней идентификации
поражения мозга, но скорее связано с индиви"
пациентов с риском развития когнитивных на"
дуальным ответом ГГНО пациента. Пациенты с
рушений или деменции [92]. Все эти данные
более высокой концентрацией кортизола в во"
подтверждают значение функционирования
лосах (очевидно, отражающей более интенсив"
ГГНО у конкретного пациента в развитии опо"
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1639
средованных повреждением гиппокампa пос"
новным фактором, обусловливающим дистант"
тинсультных когнитивных и эмоциональных
ное повреждение гиппокампа, которое лежит в
расстройств.
основе постинсультных и посттравматических
Хроническая стресс"индуцированная акти"
когнитивных и депрессивных расстройств. Пос"
вация ГГНО имеет много вариантов (хроничес"
ле фокального поражения мозга развиваются
кая базальная гиперсекреция, сенситизирован"
две разные последовательности событий. Пер"
ный стрессорный ответ и даже истощение над"
вая связана непосредственно с фокальным пов"
почечников), а ее проявление зависит от таких
реждением ткани мозга, а вторая ассоциирована
факторов, как хроничность, интенсивность,
со стрессом и последующей активацией ГГНО,
частота и модальность стресса. Важно, что ин"
избыточной секрецией КС и стимуляцией гип"
дивидуальный ответ на острый или хроничес"
покампальных ГР и МР, вызывающих дистант"
кий стресс зависит от множества факторов,
ные функциональные и структурные изменения
включая генетические, ранний жизненный
в гиппокампе (нейровоспаление, нарушения
опыт, внешние условия, пол и возраст. Кон"
нейрогенеза и синаптической пластичности,
текст, в котором действует стрессор, определяет,
нейродегенерацию и т.д.) [4].
будет ли индивидуальный ответ на острый или
Почему дисфункция ГГНО и аномальный
хронический стресс адаптивным или дезадап"
стрессорный ответ так важны в пред" и пост"ин"
тивным (патологическим)
[93]. Вызванная
сультном периодах (рис. 4, a)? Потенциальный,
стрессом избыточная секреция КС является ос"
предшествующий инсульту стресс, связанный с
Рис. 4. Периодизация стрессорной нагрузки после фокальных поражений мозга и дистантное повреждение гиппокампа.
а - Фазы потенциальных стрессорных нагрузок до и после фокального поражения мозга (детали в тексте); б - порочный
круг дисфункции ГГНО - важная причина атрофии гиппокампа (детали в тексте).
biokhsm/
БИОХИМИЯ том 84 вып. 11 2019
1640
ГУЛЯЕВА
продолжительными периодами повышения
требуют экспериментального исследования, од"
уровня кортизола, может запускать сигнальные
нако несколько подходов, вытекающих из извест"
и метаболические изменения в гиппокампе, ко"
ных механизмов, представляются понятными, и
торые могут усугубиться в последующие стрес"
ряд потенциальных специфических мишеней
сорные периоды. Предшествующая кортизоль"
для лечения можно обозначить (рис. 5).
ная нагрузка может быть легко отслежена по его
Одновременная оценка нервных, эндокрин"
уровню в волосах [94]. Фокальное поражение
ных и иммунных биомаркеров с использовани"
мозга, стрессогенный фактор, индуцирует по"
ем современных неинвазивных методов широко
вышение КС, усиливающее сигнализацию через
обсуждается [68] и важна для понимания функ"
ГР и МР. Последующий период лечения и вос"
ционирования ГГНО, участия воспаления и
становления следует рассматривать, как время
нейротрофических факторов. Это может помочь
хронического физического и психологического
оценить риск последующего развития когни"
стресса, в частности, вызванного осознанием
тивных и депрессивных расстройств. Кортизол в
пациентом своего болезненного состояния. С
волосах рассматривается как перспективный
учетом возможного предшествующего наруше"
маркер изменений активности ГГНО, вызван"
ния функции ГГНО и высокого уровня кортизо"
ных стрессом, соматическими и психическими
ла, ассоциированного с аномальной стресс"ре"
заболеваниями. Например, у пациенток с боль"
активностью, психическим заболеванием, гене"
шой депрессией уровень кортизола в волосах
тическим фоном, эти множественные хроничес"
был существенно ниже, а в сыворотке крови вы"
кие стрессорные факторы влияют на пациента с
ше, чем в контрольной группе. Снижение кор"
фокальным поражением мозга (в первую оче"
тизола в волосах у этих пациенток указывает на
редь инсультом или травмой), потенциально ин"
сниженную функцию ГГНО за предшествую"
дуцируя или последовательно усугубляя дистант"
щий месяц [94]. Таким образом, оценка «акку"
ное повреждение гиппокампа. В норме гипота"
мулированного» и системного уровня КС дает
ламический КРГ активирует высвобождение
информацию о дисфункции ГГНО. Определе"
АКТГ из гипофиза и последующую секрецию
ние концентрации хемокинов и их рецепторов
КС надпочечниками, при этом кортизол инги"
является важным показателем активации им"
бирует свою собственную секрецию через меха"
мунной и нейроэндокринной систем. Актив"
низм отрицательной обратной связи (рис. 1).
ность этих систем отражает тяжесть заболевания
Когда уровень кортизола хронически повышен и
и дает важную информацию об эффективности
функция гиппокампа изменена, гиппокамп"за"
лечения антидепрессантами [67].
висимое замедление ГГНО нарушено [53], и уро"
Важнейшим принципом оценки ГГНО и ме"
вень кортизола продолжает нарастать, вызывая
ханизмов стрессорного ответа должно быть при"
дальнейшее повреждение гиппокампа. Пороч"
менение умеренной провокации. В нашем пре"
ный круг дисфункции ГГНО, лежащий в основе
дыдущем исследовании показано, что пациент"
атрофии гиппокампа, представлен на рис. 4, б.
ки с большой депрессией демонстрировали по"
вышенную активность ГГНО, а также повышен"
ные уровни ИЛ"6 и цилиарного нейротрофичес"
ДИСТАНТНОЕ ПОВРЕЖДЕНИЕ
кого фактора (CNTF) в крови [95]. В этой груп"
ГИППОКАМПА ПОСЛЕ ФОКАЛЬНОГО
пе обнаружено вызванное стрессом повышение
ПОРАЖЕНИЯ МОЗГА: ЕСТЬ ЛИ
уровней глюкозы и ФНО"α, отсутствовавшее в
ВОЗМОЖНОСТЬ ПРЕДОТВРАТИТЬ?
контрольной группе (в качестве стресса исполь"
зовали короткий умеренный когнитивный тест).
Подход победителя: наверно,
Эти данные подтвердили нарушение стресс"ре"
это трудно, но возможно.
активности при большой депрессии и свиде"
Подход лузера: наверно, это
тельствовали о том, что при депрессии реакция
возможно, но слишком трудно.
простых метаболических и провоспалительных
Уильям Джеймс
показателей на умеренную стрессогенную наг"
рузку может быть показательной. Понимание
Мы обсудили механизмы вызванного КС
механизмов взаимодействия между КС и BDNF
дистантного повреждения гиппокампа, а также
перспективно для идентификации потенциаль"
причины когнитивных и эмоциональных
ных терапевтических мишеней [29]. BDNF сы"
расстройств, вызванных этим повреждением.
воротки - стресс"чувствительный показатель,
Следующий принципиальный вопрос - как
который существенно повышается после стрес"
предотвратить или купировать эти последствия.
са, а затем снижается до восстановления исход"
Конечно, некоторые важные звенья этих пато"
ного уровня. Linzet et al. [30] обнаружили анта"
генетических механизмов остаются неясными и
гонистические взаимоотношения между BDNF
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1641
Рис. 5. «Магический ящик» механизмов многоуровневого стрессорного ответа. Адекватный пластичный ответ мозга на
каждом уровне определяется взаимодействием между основными системами, ГГНО/КС, глутаматергической, нейротро"
финовой и цитокиновой, а также их тонко настроенной взаимной регуляцией. В нормальных условиях влияние естест"
венного стресса на любое звено вызывает благоприятные ко"регуляторные изменения других процессов, стимулируя мно"
гоуровневые адаптивные изменения. Если стрессорный фактор избыточно сильный и вызванную им «поломку» механиз"
ма не удается с легкостью компенсировать адаптивными изменениями, механизм начинает неправильно функциониро"
вать, вызывая патологические изменения, в том числе молекулярные события, приводящие к дисфункции клеток мозга
и, в конце концов, к нарушениям поведения, которые являются отличительной особенностью неврологических и психи"
ческих заболеваний. Звездочками отмечены потенциальные фармакологические мишени для предотвращения или лече"
ния аномального функционирования системы
и кортизолом; более высокие пики BDNF после
Полногеномный скрининг гиппокампа на гены,
психосоциального стресса были ассоциированы
экспрессия которых зависит от КС, позволяет
с более резким восстановлением кортизола, а
определить потенциальные мишени для препа"
величина стрессорного ответа кортизола была
ратов, поддерживающих репаративные возмож"
связана с более крутым восстановлением BDNF
ности, имеющиеся в больном мозге [5].
после стресса. Важно, что ответ мозга на стресс
Другим подходом может быть использование
зависит от индивидуального генетического фо"
известной ассоциации между депрессией и ког"
на и его взаимодействия с событиями, происхо"
нитивными нарушениями. Tene et al. [91] пред"
дившими в жизни индивидуума [96]. Например,
положили, что скрининг депрессии у пациен"
на кортизол и эмоциональность/когницию вли"
тов, переживших инсульт или транзиторные
яют гаплотипы гена МР на базе rs5522 и
ишемические атаки, является инструментом для
rs2070951. Гаплотип МР 1 (GA) модерирует эф"
идентификации пациентов, которые наиболее
фекты (ранних) стрессоров и репродуктивного
вероятно будут иметь более тяжелый когнитив"
цикла. Гаплотип МР 2 (CA) способствует функ"
ный и функциональный исход. Панель специ"
циональному варианту, который защищает жен"
фических биомаркеров может быть полезна в
щин от депрессии за счет ассоциации с опти"
качестве параклинической основы для диагнос"
мистическим, устойчивым фенотипом
[97]. тики постинсультной депресии и выяснения
БИОХИМИЯ том 84 вып. 11 2019
1642
ГУЛЯЕВА
различных аспектов ее многофакторного пато"
тальных исследований), и это внушает опти"
генеза, оптимизации терапевтических меропри"
мизм. Очевидно, что трансляция экспримен"
ятий и оценки эффективности лечения [84]. К
тальных результатов и проведение соответству"
сожалению, до сих пор проводится недостаточ"
ющих клинических исследований приблизит
но обширных, тщательных и полноценных про"
нас к окончательной цели - предупреждению
должительных исследований для более четкой
депрессии и деменции после фокального пора"
идентификации подтипов депрессии, которые
жения мозга. Антиглюкокортикоидные агенты в
могут быть главным предиктором последующе"
определенных условиях обладают терапевтичес"
го когнитивного снижения. Это бы позволило
ким потенциалом, например, при заболеваниях,
применить специфическую терапию в период
характеризующихся ранними повреждениями
критических эпизодов этих подтипов и сокра"
гиппокампа и нарушениями памяти, как MCI,
тить риск когнитивных последствий [82].
БА и другие типы деменции [97].
Таким образом, первый вывод состоит в том,
Первый антагонист МР, спиронолактон, был
что мы достаточно легко можем оценивать дис"
разработан более 60 лет назад для лечения пер"
функцию ГГНО и индивидуальную стресс"реак"
вичного альдостеронизма и патологического
тивность пациента, используя тестовую стрес"
отека. С тех пор знание о функциях МР развива"
сорную провокацию, и это практический под"
лось вместе с клиническими подтверждениями
ход к оценке риска дистантного повреждения
терапевтической значимости антагонистов МР
гиппокампа и потенциального развития пост"
для предотвращения нежелательной активации
инсультных или посттравматических когнитив"
МР [100]. Между разработкой первого и второ"
ных и депрессивных расстройств.
го, МР"селективного, поколения антагонистов
Гиперактивность ГГНО может присутство"
МР прошло ~40 лет [101]. Еще через 20 лет поя"
вать при хронических заболеваниях, влияя на
вилось третье поколение антагонистов, однако
эндокринную, сердечно"сосудистую и нервную
до сих пор большое число различных типов кле"
систему (деменция, депрессия), особенно при
ток, экспрессирующих МР, и их разнообразные
коморбидных состояниях. Спектр молекул, спо"
клеточно"специфичные действия, а также уни"
собных взаимодействовать с различными уров"
кальные сложные взаимодействия МР на моле"
нями ГГНО, экспоненциально растет, они име"
кулярном уровне не были учтены. Обсуждается
ют мишени на разных уровнях, от супрагипота"
разработка четвертого поколения селективных
ламических до пострецепторных механизмов, и
антагонистов МР, а также специфических аго"
включают агенты, действующие на супрахиаз"
нистов МР с учетом структурных особенностей
матическое ядро, рецептор CRH"R1, стероидо"
ткани и лиганд"специфичной активации МР
генез в надпочечниках, ГР и периферичес"
[101].
кие/центральные ферменты 11HSD типа 1. Эта
Введение антагонистов МР в стандартную
область исследования быстро развивается с
терапию сердечной недостаточности, заболева"
целью продвижения терапевтических стратегий
ний почек, метаболического синдрома и диабе"
для борьбы с гиперактивностью ГГНО и сниже"
та постепенно возрастает вслед за клинически"
ния бремени связаных со стрессом заболеваний
ми испытаниями, которые демонстрируют оче"
[98]. Согласно данным многочисленных иссле"
видные преимущества этих препаратов. Кроме
дований, дисфункция рецепторов КС вовлечена
блокирования пагубных эфффектов МР в серд"
в патогенез аффективных расстройств. Ряд
це, сосудах и почках, антагонисты МР действу"
контрольных испытаний еще предстоит провес"
ют на МР в регуляторных гемодинамических
ти, однако уже есть подтвержения клинически
центрах мозга, снижая избыточные симпатичес"
благоприятных эффектов при использовании
кие влияния, высвобождение вазопрессина,
снижающих КС препаратов или антагонистов
аномальное функционирование барорецепто"
рецепторов КС у определенных пациентов с
ров и уровни циркулирующих и тканевых про"
депрессией. Методами генной инженерии полу"
воспалительных цитокинов. Тем не менее рас"
чены мыши со специфическими изменениями
сматривая потенциальное применение антаго"
индивидуальных генов, участвующих в проведе"
нистов МР, нужно иметь в виду, что МР вовлече"
нии сигнала рецепторами КС и регуляции
ны в когнитивные функции и память [101].
стрессорных гормонов. Идентификция и де"
Опосредованная МР трансдукция сигнала в
тальное описание этих молекулярных путей, в
мозге считается протекторным фактором при
конце концов, должно привести к развитию но"
развитии психопатологии, в частности, аффек"
вых нейрофармакологических стратегий [99].
тивных расстройств. Поэтому активация МР
Фармакологические инструменты для моду"
может быть средством для облегчения депрес"
ляции МР и ГР, а также снижения уровней КС
сии и когнитивной дисфункции. Соответствен"
доступны (хотя, в основном для эксперимен"
но, было показано, что агонист МР, флудрокор"
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1643
тизон, увеличивает эффективность антидепрес"
а затем в клинике. Hовые соединения, которые
сантов и улучшает память и исполнительные
будут разработаны, должны быть более специ"
функции у молодых пациентов с депрессией.
фичными в отношении определенных рецепто"
КС координируют через МР сети, определяю"
ров КС и тканеспецифичными.
щие индивидуальное преодоление стресса, и это
Понимание роли иммунной системы и вос"
дополняется эффектами с участием более низ"
паления у пациентов с большой депрессией или
коаффинных ГР, вовлеченных в последующее
когнитивным дефицитом необходимо для раз"
формирование адаптации к стрессу. При регуля"
работки эффективных средств, направленных
ции соотношения МР/ГР именно МР является
на процессы воспаления, для снижения симпто"
важнейшим фактором, способствующим устой"
матики пациента и коморбидности. Поскольку
чивости [97].
большинство антидепрессантов оказывают спе"
Низкоаффинные ГР, широко экспрессиро"
цифические противовоспалительные эффекты,
ванные в организме, активируются только при
восстановление сниженного за счет воспали"
возрастании уровня КС (при циркадианных
тельных процессов нейрогенеза может быть не"
максимумах и ответе на стресс). ГР модулируют
посредственно связано с эффективностью лече"
функционирование и выживание нейронов че"
ния антидепрессантами [72].
рез геномные и негеномные механизмы, и их
Альтернативным подходом к предотвраще"
транскрипционная регуляторная активность
нию стрессорного повреждения гиппокампа мо"
тесно связана с секреторными паттернами КС
жет быть анализ известных свойств препаратов,
[52]. Генетические и средовые факторы могут
которые используют в клинике по иным показа"
нарушить функцию ГР [68]. Недавно показано,
ниям; целесообразно найти препараты с желае"
что КС-ГР могут оказывать нейротрофическое
мыми свойствами и исследовать их потенциал.
действие; для потенциального использования
Например, имеется целый ряд препаратов, ко"
ГР, как мишени любой терапевтической страте"
торые позиционируются как адаптогены для
гии, при нейродегенеративных заболеваниях
стабилизации физиологических процессов и
нужно знать конкретные модификации как в
поддержания гомеостаза. Адаптогенная концеп"
ГГНО, так и в активности ГР и найти пути ис"
ция в фармакологии требует дополнительных
пользования их протекторных свойств [52]. В
клинических и доклинических исследований, и
экспериментах на грызунах мифепристон (RU
поэтому не принята в строгой современной тер"
486), антагонист ГР, благоприятно влиял на
минологии, тем не менее ряд препаратов, разре"
чувствительность к хроническому стрессу [102].
шенных в клинике, обладает подтвержденными
Интересно, что проведение сигнала ГР в интер"
адаптогенными свойствами. В частности, пре"
нейронах специфически значимо для самок, что
параты, содержащие нейропептиды, например,
может быть важным для направленной на ГР
церебролизин, плейотропны и оказывают адап"
гендер"специфичной терапии связанных со
тогенные эффекты на молекулярном уровне
стрессом аффективных состояний [103].
[104]. Оптимальным представляется проведение
В настоящее время в российской клиничес"
мета"анализа адаптогенных свойств препаратов,
кой практике широко используются минимум
используемых для лечения поражений мозга (в
2 антагониста рецепторов КС. Оба обладают
частности, после инсульта и травмы), найти на"
широкой специфичностью в отношении раз"
иболее перспективные из них и эксперимен"
личных стероидных рецепторов, и ни один из
тально исследовать их влияние на ГГНО при
них не используется для лечения когнитивных и
действии различных стрессорных факторов. По"
аффективных расстройств. Спиронолактон,
этому третий вывод состоит в предположении,
синтетический 17"спиронолактоновый КС с ка"
что какие"либо из многочисленных препаратов
лий"сберегающей, диуретической, антигипер"
с адаптогенными свойствами и уже используе"
тензивной и антиандрогенной активностями
мые в клинике, могут обладать потенциалом для
конкурентно ингибирует МР. Мифепристон -
нормализации ГГНО и других механизмов
антагонист ГР и прогестероновых рецепторов.
стрессорного ответа. Большинство эффектив"
Важно, что эти антагонисты рецепторов КС бе"
ных препаратов можно обнаружить скринингом
зопасны при лечении различных патологий в
в эксперименте и верифицировать в доклини"
клинике. Поэтому второй вывод состоит в том,
ческих и клинических исследованиях.
что лиганды рецепторов КС доступны, и неко"
В данной области еще много вопросов, кото"
торые из них доказали свою безопасность в кли"
рые нужно выяснить, и новое понимание проб"
нике (хотя и используются по показаниям, не
лемы дает возможность по"новому взглянуть на
связанным с когнитивными и аффективными
патофизиологию и потенциально новые сред"
расстройствами). Доступные соединения можно
ства для терапии связанных со стрессом когни"
исследовать в соответствующих экспериментах,
тивных и психических расстройств. В частнос"
БИОХИМИЯ том 84 вып. 11 2019
1644
ГУЛЯЕВА
ти, более глубокое понимание действия КС мог"
ным высвобождением КС, активацией ГГНО и
ло бы привести к более рациональной стратегии
гиппокампальных рецепторов КС. Иными сло"
лечения таких заболеваний [105]. Эпиграфом к
вами, мы предположили, что дистантное пов"
этой главе выбраны слова Уильяма Джеймса,
реждение гиппокампа и последующие когнитив"
выдающегося мыслителя конца 19 века, «отца
ные/аффективные расстройства после различ"
американской психологии». Давайте следовать
ных поражений мозга являются не прямым пос"
подходу победителя: наверно, трудно предот"
ледствием первичного поражения, а в значи"
вратить отложенные опосредованные гиппо"
тельной степени результатом стрессорной наг"
кампом расстройства после повреждения мозга,
рузки и взаимодействием КС со специфически"
но это возможно на основании известных нам
ми рецепторами в гиппокампе. Это дистантное
механизмов и тех механизмов, которые еще бу"
повреждение ассоциировано с деменцией и деп"
дут открыты в ближайшем будущем. Ключевой
рессией, развивающимися после фокального по"
вывод - утверждение о том, что нужно прово"
ражения. Очевидно, что индивидуальная стресс"
дить поиск потенциальных средств, предотвра"
реактивность и есть ключевой фактор, определя"
щающих дистантное повреждение гиппокампа
ющий риск развития патологических событий. В
после фокального поражения мозга, а также
этой работе мы постарались: а) построить основу
развитие отложенных когнитивных и эмоцио"
для более глубокого понимания системы много"
нальных нарушений. Перспективы такого поис"
уровневого взаимодействия и взаимозависимых
ка выглядят весьма оптимистично.
механизмов опосредованного гиппокампом
стрессорного ответа; б) описать возможные при"
чины и природу нарушения регуляции этой
ЗАКЛЮЧИТЕЛЬНЫЕ ЗАМЕЧАНИЯ
сложной системы; в) показать, как нарушение
функционирования этой системы влияет на дея"
Здоровье начинается
тельность гиппокампа и как оно влияет на ког"
с понимания болезни.
нитивный и эмоциональный статус человека;
Испанская пословица
г) предположить, какие патогенетически обос"
нованные подходы могут быть использованы для
Поскольку дисфункция гиппокампа после
предотвращения или репарации такой дисфунк"
фокального поражения мозга тесно связана с
ции гиппокампа. «Магический ящик» регуляции
развитием когнитивных нарушений, включая
стрессорного ответа (рис. 5) содержит многочис"
деменцию и коморбидную депрессию, важно
ленные механизмы, интегрирующие множество
понимать конкретные механизмы дистантного
систем (ГГНО/КС/рецепторы КС, нейротрофи"
повреждения гиппокампа. Имеющиеся данные
ны и их рецепторы, воспалительные механизмы,
указывают на наличие общих факторов, лежа"
нейромедиаторы и их рецепторы и многие дру"
щих в основе этого феномена, которые не зави"
гие) на различных уровнях, и это, если система
сят строго от природы и локализации первично"
разбалансирована, осложняет превентивные/те"
го повреждения.
рапевтические мероприятия, поскольку ключе"
Дистантное повреждение гиппокампа, свя"
вая мишень не всегда очевидна. Однако тонкая
занное с первичным поражением других облас"
взаиморегуляция систем и оперативные меха"
тей мозга, описано много лет назад (см. обзор
низмы нейропластичности [106] дают надежду,
[4]), однако до сих пор не было предложено ра"
что починка одного ключевого звена может по"
циональной концепции для объяснения селек"
мочь отремонтировать всю машину.
тивной уязвимости гиппокампа в этих ситуаци"
ях. Недавно мы предложили новый взгляд, кото"
рый способствует объяснению, почему гиппо"
Финансирование. Работа поддержана Рос"
камп так избирательно чувствителен к повреж"
сийским фондом фундаментальных исследова"
дению такого типа [4]. Мы предположили, что
ний (грант № 18"00"00125) (стресс, депрессия) и
этот феномен может быть опосредован относи"
Программой президиума Российской академии
тельно высокой уязвимостью к нейровоспале"
наук «Фундаментальные основы технологии
нию, связанной с нарушением локального мета"
физиологических адаптаций (дистантное пов"
болизма и сигнальной функции КС. Мы рас"
реждение гиппокампа)».
смотрели, по крайней мере, две различные ли"
Конфликт интересов. Автор заявляет об отсут"
нии событий после фокального повреждения
ствии конфликта интересов.
мозга (инсульта или травмы): первая связана с
Соблюдение этических норм. Настоящая
прямым первичным поражением ткани мозга, а
статья не содержит каких"либо исследований с
вторая - с опосредованными ГГНО событиями
участием людей или использованием животных
стрессорного ответа, в первую очередь избыточ"
в качестве объектов исследований.
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1645
СПИСОК ЛИТЕРАТУРЫ
1.
McEwen, B.S. (1996) Gonadal and adrenal steroids regu"
between excitation and inhibition? Cell. Mol. Life Sci., 71,
late neurochemical and structural plasticity ofthe hip"
2499-2515.
pocampus via cellular mechanisms involving NMDA
20.
Vitellius, G., Trabado, S., Bouligand, J., Delemer, B., and
receptors, Cell. Mol. Neurobiol., 16, 103-116.
Lombès, M. (2018) Pathophysiology of glucocorticoid sig"
2.
De Kloet, E.R., Han, F., and Meijer, O.C. (2008) From the
naling, Ann. Endocrinol. (Paris), 79, 98-106.
stalk to down under about brain glucocorticoid receptors,
21.
Van Weert, L.T.C.M., Buurstede, J.C., Sips, H.C.M.,
stress and development, Neurochem. Res., 33, 637-642.
Mol, I.M., Puri, T., Damsteegt, R., Roozendaal, B.,
3.
Uchoa, E.T., Aguilera, G., Herman, J.P., Fiedler, J.L.,
Sarabdjitsingh, R.A., and Meijer, O.C. (2019) Mechanistic
Deak, T., and de Sousa, M.B. (2014) Novel aspects of glu"
insights in NeuroD potentiation of mineralocorticoid
cocorticoid actions, J. Neuroendocrinol., 26, 557-572.
receptor signaling, Int. J. Mol. Sci., 20, doi: 10.3390/
4.
Gulyaeva, N.V. (2019) Functional neurochemistry of the
ijms20071575.
ventral and dorsal hippocampus: stress, depression,
22.
Joëls, M., and Karst, H. (2012) Corticosteroid effects on
dementia and remote hippocampal damage, Neurochem.
calcium signaling in limbic neurons, Cell. Calcium, 51,
Res., 44, 1306-1322.
277-283.
5.
De Kloet, E.R. (2000) Stress in the brain, Eur. J.
23.
Lapp, H.E., Bartlett, A.A., and Hunter, R.G. (2019) Stress
Pharmacol., 405, 187-198.
and glucocorticoid receptor regulation of mitochondrial
6.
De Kloet, E.R., Vreugdenhil, E., Oitzl, M.S., and Joëls,
gene expression, J. Mol. Endocrinol., 62, R121-R128.
M. (1998) Brain corticosteroid receptor balance in health
24.
Reul, J.M. (2014) Making memories of stressful events: a
and disease, Endocr. Rev., 19, 269-301.
journey along epigenetic, gene transcription, and signaling
7.
Meijer, O.C., Buurstede, J.C., and Schaaf, M.J.M. (2019)
pathways, Front. Psychiatry, 5, 5.
Corticosteroid receptors in the brain: transcriptional
25.
Mifsud, K.R., Gutièrrez"Mecinas, M., Trollope, A.F.,
mechanisms for specificity and context"dependent effects,
Collins, A., Saunderson, E.A., and Reul, J.M. (2011)
Cell. Mol. Neurobiol., 39, 539-549.
Epigenetic mechanisms in stress and adaptation, Brain
8.
Joëls, M.
(2018) Corticosteroids and the brain,
Behav. Immun., 25, 1305-1315.
J. Endocrinol., 238, R121-R130.
26.
Bennett, M.R., and Lagopoulos, J. (2014) Stress and trau"
9.
Groeneweg, F.L., Karst, H., de Kloet, E.R., and Joëls, M.
ma: BDNF control of dendritic"spine formation and
(2011) Rapid non"genomic effects of corticosteroids and
regression, Prog. Neurobiol., 112, 80-99.
their role in the central stress response, J. Endocrinol., 209,
27.
Gulyaeva, N.V. (2017) Interplay between Brain BDNF and
153-167.
glutamatergic systems: a brief state of the evidence and
10.
Nishi, M., and Kawata, M. (2006) Brain corticosteroid
association with the pathogenesis of depression,
receptor dynamics and trafficking: implications from live
Biochemistry (Moscow), 82, 301-307.
cell imaging, Neuroscientist, 12, 119-133.
28.
Leal, G., Bramham, C.R., and Duarte, C.B. (2017) BDNF
11.
De Kloet, E.R, Oitzl, M.S., and Schöbitz, B. (1994)
and hippocampal synaptic plasticity, Vitam. Horm., 104,
Cytokines and the brain corticosteroid receptor balance:
153-195.
relevance to pathophysiology of neuroendocrine"immune
29.
Suri, D., and Vaidya, V.A. (2013) Glucocorticoid regula"
communication, Psychoneuroendocrinology, 19, 121-134.
tion of brain"derived neurotrophic factor: relevance to hip"
12.
De Kloet, E.R., Meijer, O.C., de Nicola, A.F.,
pocampal structural and functional plasticity,
de Rijk, R.H., and Joëls, M. (2018) Importance of the
Neuroscience, 239, 196-213.
brain corticosteroid receptor balance in metaplasticity,
30.
Linz, R., Puhlmann, L.M.C., Apostolakou, F., Mantzou,
cognitive performance and neuro"inflammation, Front.
E., Papassotiriou, I., Chrousos, G.P., Engert, V., and
Neuroendocrinol., 49, 124-145.
Singer, T. (2019) Acute psychosocial stress increases serum
13.
Murakami, G., Hojo, Y., Kato, A., Komatsuzaki, Y.,
BDNF levels: an antagonistic relation to cortisol but no
Horie, S., Soma, M., Kim, J., and Kawato, S. (2018) Rapid
group differences after mental training, Neuropsychophar(
nongenomic modulation by neurosteroids of dendritic
macology, doi: 10.1038/s41386"019"0391"y.
spines in the hippocampus: androgen, oestrogen and corti"
31.
Daskalakis, N.P., De Kloet, E.R., Yehuda, R., Malaspina,
costeroid, J. Neuroendocrinol., 30, doi: 10.1111/jne.12561.
D., and Kranz, T.M. (2015) Early life stress effects on glu"
14.
Giordano, R., Pellegrino, M., Picu, A., Bonelli, L., Balbo, M.,
cocorticoid"BDNF interplay in the hippocampus, Front.
Berardelli, R., Lanfranco, F., Ghigo, E., and Arvat, E.
Mol. Neurosci., 8, 68.
(2006) Neuroregulation of the hypothalamus"pituitary"
32.
Finsterwald, C., and Alberini C.M. (2014) Stress and glu"
adrenal (HPA) axis in humans: effects of GABA", miner"
cocorticoid receptor"dependent mechanisms in long"term
alocorticoid", and GH"Secretagogue"receptor modula"
memory: from adaptive responses to psychopathologies,
tion, Sci. World J., 6, 1-11.
Neurobiol. Learn. Mem., 112, 17-29.
15.
Joëls, M., and de Kloet, E.R. (2017) 30 Years of the min"
33.
Sapolsky, R.M. (1990) Glucocorticoids, hippocampal
eralocorticoid receptor: the brain mineralocorticoid recep"
damage and the glutamatergic synapse, Prog. Brain Res.,
tor: a saga in three episodes, J. Endocrinol., 234, T49-T66.
86, 13-23.
16.
Le Menuet, D., and Lombès, M. (2014) The neuronal
34.
Popoli, M., Yan, Z., McEwen, B.S., and Sanacora, G.
mineralocorticoid receptor: from cell survival to neurogen"
(2011) The stressed synapse: the impact of stress and glu"
esis, Steroids, 91, 11-19.
cocorticoids on glutamate transmission, Nat. Rev.
17.
Odermatt, A., and Kratschmar, D.V. (2012) Tissue"specif"
Neurosci., 13, 22-37.
ic modulation of mineralocorticoid receptor function by
35.
Chaouloff, F., and Groc, L. (2011) Temporal modulation
11β"hydroxysteroid dehydrogenases: an overview, Mol.
of hippocampal excitatory transmission by corticosteroids
Cell. Endocrinol., 350, 168-186.
and stress, Front. Neuroendocrinol., 32, 25-42.
18.
Kellendonk, C., Gass, P., Kretz, O., Schütz, G., and
36.
Goujon, E., Layé, S., Parnet, P., and Dantzer, R. (1997)
Tronche, F. (2002) Corticosteroid receptors in the brain:
Regulation of cytokine gene expression in the central ner"
gene targeting studies, Brain Res. Bull., 57, 73-83.
vous system by glucocorticoids: mechanisms and function"
19.
Saaltink, D.J., and Vreugdenhil, E. (2014) Stress, gluco"
al consequences, Psychoneuroendocrinology, 22, Suppl. 1,
corticoid receptors, and adult neurogenesis: a balance
S75-S80.
БИОХИМИЯ том 84 вып. 11 2019
1646
ГУЛЯЕВА
37.
Duque, Ede A., and Munhoz, C.D. (2016) The pro"
an acute functional cross"talk between amyloid"β and glu"
inflammatory effects of glucocorticoids in the brain, Front.
cocorticoid receptors at hippocampal excitatory synapses,
Endocrinol. (Lausanne), 7, 78.
Neurobiol. Dis., 118, 117-128.
38.
Pariante, C.M.
(2017) Why are depressed patients
56.
Bisht, K., Sharma, K., and Tremblay, M.È. (2018) Chronic
inflamed? A reflection on 20 years of research on depres"
stress as a risk factor for Alzheimer’s disease: roles of
sion, glucocorticoid resistance and inflammation, Eur.
microglia"mediated synaptic remodeling, inflammation,
Neuropsychopharmacol., 27, 554-559.
and oxidative stress, Neurobiol. Stress, 9, 9-21.
39.
Walker, F.R., Nilsson, M., and Jones, K. (2013) Acute and
57.
Pomara, N., Greenberg, W.M., Branford, M.D., and
chronic stress"induced disturbances of microglial plasticity,
Doraiswamy, P.M. (2003) Therapeutic implications of
phenotype and function, Curr. Drug Targets,
14,
HPA axis abnormalities in Alzheimer’s disease: review and
1262-1276.
update, Psychopharmacol. Bull., 37, 120-134.
40.
Gądek"Michalska, A., Tadeusz, J., Rachwalska, P., and
58.
Numakawa, T., Odaka, H., and Adachi, N. (2017) Actions
Bugajski, J. (2013) Cytokines, prostaglandins and nitric
of brain"derived neurotrophic factor and glucocorticoid
oxide in the regulation of stress"response systems,
stress in neurogenesis, Int. J. Mol. Sci., 18, doi: 10.3390/
Pharmacol. Rep., 65, 1655-1662.
ijms18112312.
41.
Johnson, J.D., Barnard, D.F., Kulp, A.C., and Mehta, D.M.
59.
Fitzsimons, C.P., Herbert, J., Schouten, M., Meijer, O.C.,
(2019) Neuroendocrine regulation of brain cytokines after
Lucassen, P.J., and Lightman, S. (2016) Circadian and
psychological stress, J. Endocr. Soc., 3, 1302-1320.
ultradian glucocorticoid rhythmicity: implications for the
42.
Deak, T., Quinn, M., Cidlowski, J.A., Victoria, N.C.,
effects of glucocorticoids on neural stem cells and adult
Murphy A.Z., and Sheridan, J.F. (2015) Neuroimmune
hippocampal neurogenesis, Front. Neuroendocrinol., 41,
mechanisms of stress: sex differences, developmental plas"
44-58.
ticity, and implications for pharmacotherapy of stress"
60.
Lucassen, P.J., Oomen, C.A., Naninck, E.F., Fitzsimons, C.P.,
related disease, Stress, 18, 367-380.
van Dam, A.M., Czeh, B., and Korosi, A.
(2015)
43.
Bekhbat, M., Rowson, S.A., and Neigh, G.N. (2017)
Regulation of adult neurogenesis and plasticity by (early)
Checks and balances: the glucocorticoid receptor and
stress, glucocorticoids, and inflammation, Cold Spring
NFκB in good times and bad, Front. Neuroendocrinol., 46,
Harb. Perspect. Biol., 7, a021303.
15-31.
61.
Schoenfeld, T.J., and Gould, E. (2012) Stress, stress hor"
44.
De Kloet, E.R., and Joëls, M. (2017) Brain mineralocorti"
mones, and adult neurogenesis, Exp. Neurol., 233, 12-21.
coid receptor function in control of salt balance and stress"
62.
Lupien, S.J., and Lepage, M. (2001) Stress, memory, and
adaptation, Physiol. Behav., 178, 13-20.
the hippocampus: can’t live with it, can’t live without it,
45.
Brocca, M.E., Pietranera, L., de Kloet, E.R., and De
Behav. Brain Res., 127, 137-158.
Nicola, A.F. (2019) Mineralocorticoid receptors, neuroin"
63.
Herman, J.P., Ostrander, M.M., Mueller, N.K., and
flammation and hypertensive encephalopathy, Cell. Mol.
Figueiredo, H. (2005) Limbic system mechanisms of stress
Neurobiol., 39, 483-492.
regulation: hypothalamo"pituitary"adrenocortical axis,
46.
Frank, M.G., Weber, M.D., Watkins, L.R., and Maier, S.F.
Prog. Neuropsychopharmacol. Biol. Psychiatry.,
29,
(2015) Stress sounds the alarmin: the role of the danger"
1201-1213.
associated molecular pattern HMGB1 in stress"induced
64.
Joëls, M., Pasricha, N., and Karst, H. (2013) The interplay
neuroinflammatory priming, Brain Behav. Immun., 48,
between rapid and slow corticosteroid actions in brain, Eur.
1-7.
J. Pharmacol., 719, 44-52.
47.
Pearson"Leary, J., Osborne, D.M., and McNay, E.C.
65.
Zunszain, P.A., Anacker, C., Cattaneo, A., Carvalho, L.A.,
(2016) Role of glia in stress"induced enhancement and
and Pariante, C.M. (2011) Glucocorticoids, cytokines and
impairment of memory, Front. Integr. Neurosci., 9, 63.
brain
abnormalities
in
depression,
Prog.
48.
Van Olst, L., Bielefeld, P., Fitzsimons, C.P., de Vries, H.E.,
Neuropsychopharmacol. Biol. Psychiatry, 35, 722-729.
and Schouten, M. (2018) Glucocorticoid"mediated modu"
66.
Leonard, B. (2000) Stress, depression and the activation of
lation of morphological changes associated with aging in
the immune system, World J. Biol. Psychiatry, 1, 17-25.
microglia, Aging Cell, 17, e12790.
67.
Ogіodek, E., Szota, A., Just, M., Moś, D., and
49.
McEwen, B.S. (1997) Possible mechanisms for atrophy of
Araszkiewicz, A. (2014) The role of the neuroendocrine
the human hippocampus, Mol. Psychiatry, 2, 255-262.
and immune systems in the pathogenesis of depression,
50.
Vyas, S., Rodrigues, A.J., Silva, J.M., Tronche, F.,
Pharmacol. Rep., 66, 776-781.
Almeida, O.F., Sousa, N., and Sotiropoulos, I. (2016)
68.
Silverman, M.N., and Sternberg, E.M.
(2012)
Chronic stress and glucocorticoids: from neuronal plastic"
Glucocorticoid regulation of inflammation and its func"
ity to neurodegeneration, Neural Plast., 2016, 6391686.
tional correlates: from HPA axis to glucocorticoid receptor
51.
Libro, R., Bramanti, P., and Mazzon, E.
(2017)
dysfunction, Ann. N. Y. Acad. Sci., 1261, 55-63.
Endogenous glucocorticoids: role in the etiopathogenesis
69.
Leonard, B.E. (2018) Inflammation and depression: a
of Alzheimer’s disease, Neuro Endocrinol. Lett., 38, 1-12.
causal or coincidental link to the pathophysiology? Acta
52.
Vyas, S., and Maatouk, L. (2013) Contribution of gluco"
Neuropsychiatr., 30, 1-16.
corticoids and glucocorticoid receptors to the regulation of
70.
Makhija, K., and Karunakaran, S. (2013) The role of
neurodegenerative processes, CNS Neurol. Disord. Drug
inflammatory cytokines on the aetiopathogenesis of
Targets, 12, 1175-1193.
depression, Aust. N. Z. J. Psychiatry, 47, 828-839.
53.
Ouanes, S., and Popp, J. (2019) High cortisol and the risk
71.
Bauer, M.E., and Teixeira, A.L. (2019) Inflammation in
of dementia and Alzheimer’s disease: a review of the liter"
psychiatric disorders: what comes first? Ann. N. Y. Acad.
ature, Front. Aging Neurosci., 11, 43.
Sci., 437, 57-67.
54.
Paul, S., Jeon, W.K., Bizon, J.L., and Han, J.S. (2015)
72.
Maes, M., Yirmyia, R., Noraberg, J., Brene, S., Hibbeln, J.,
Interaction of basal forebrain cholinergic neurons with the
Perini, G., Kubera., M., Bob, P, Lerer, B., and Maj, M. (2009)
glucocorticoid system in stress regulation and cognitive
The inflammatory & neurodegenerative (I&ND) hypothesis of
impairment, Front. Aging Neurosci., 7, 43.
depression: leads for future research and new drug develop"
55.
Kootar, S., Frandemiche, M.L., Dhib, G., Mouska, X.,
ments in depression, Metab. Brain Dis., 24, 27-53.
Lorivel, T., Poupon"Silvestre, G., Hunt, H., Tronche, F.,
73.
Kim, Y.K., Na, K.S., Myint, A.M., and Leonard, B.E.
Bethus, I., Barik, J., and Marie, H. (2018) Identification of
(2016) The role of pro"inflammatory cytokines in neuroin"
БИОХИМИЯ том 84 вып. 11 2019
БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДИСТАНТНОГО ПОВРЕЖДЕНИЯ ГИППОКАМПА
1647
flammation, neurogenesis and the neuroendocrine system
89. Ben Assayag, E., Tene, O., Korczyn, A.D., Shopin, L.,
in major depression, Prog. Neuropsychopharmacol. Biol.
Auriel, E., Molad, J., Hallevi, H., Kirschbaum, C.,
Psychiatry, 64, 277-284.
Bornstein, N.M., Shenhar"Tsarfaty, S., Kliper, E., and
74.
Barnard, D.F., Gabella, K.M., Kulp, A.C., Parker, A.D.,
Stalder, T. (2017) High hair cortisol concentrations predict
Dugan, P.B., and Johnson, J.D. (2019) Sex differences in
worse cognitive outcome after stroke: results from the
the regulation of brain IL"1β in response to chronic stress,
TABASCO prospective cohort study, Psychoneuroendo(
Psychoneuroendocrinology, 103, 203-211.
crinology, 82, 133-139.
75.
Cattaneo, A., and Riva, M.A. (2016) Stress"induced
90. Tene, O., Hallevi, H., Korczyn, A.D., Shopin, L., Molad, J.,
mechanisms in mental illness: a role for glucocorticoid sig"
Kirschbaum, C., Bornstein, N.M., Shenhar"Tsarfaty, S.,
nalling, J. Steroid Biochem. Mol. Biol., 160, 169-174.
Kliper, E., Auriel, E., Usher, S., Stalder, T., and Ben
76.
Zimmermann, C.A., Arloth, J., Santarelli, S., Löschner,
Assayag, E. (2018) The price of stress: high bedtime salivary
A., Webe, P, Schmidt, M.V., Spengler, D., and Binder, E.B.
cortisol levels are associated with brain atrophy and cogni"
(2019) Stress dynamically regulates co"expression net"
tive decline in stroke survivors. Results from the TABAS"
works of glucocorticoid receptor"dependent MDD and
CO prospective cohort study, J. Alzheimer’s Dis., 65,
SCZ risk genes, Transl. Psychiatry, 9, 41, doi: 10.1038/
1365-1375.
s41398"019"0373"1.
91. Tene, O., Shenhar"Tsarfaty, S., Korczyn, A.D., Kliper, E.,
77.
Gray, J.D., Kogan, J.F., Marrocco, J., and McEwen, B.S.
Hallevi, H., Shopin, L.,Auriel, E., Mike, A., Bornstein,
(2017) Genomic and epigenomic mechanisms of glucocor"
N.M., and Assayag, E.B. (2016) Depressive symptoms fol"
ticoids in the brain, Nat. Rev. Endocrinol., 13, 661-673.
lowing stroke and transient ischemic attack: is it time for a
78.
Sousa, N., and Almeida, O.F. (2002) Corticosteroids:
more intensive treatment approach? Results from the
sculptors of the hippocampal formation, Rev. Neurosci.,
TABASCO cohort study, J. Clin. Psychiatry, 77, 673-680.
13, 59-84.
92. Kliper, E., Ben Assayag, E., Korczyn, A.D., Auriel, E.,
79.
Chen J., Wang, Z.Z., Zhang, S., Chu, S.F., Mou, Z., and
Shopin, L., Hallevi, H., Shenhar"Tsarfaty, S., Mike, A.,
Chen, N.H. (2019) The effects of glucocorticoids on
Artzi, M., Klovatch, I., Bornstein, N.M., and Ben Bashat, D.
depressive and anxiety"like behaviors, mineralocorticoid
(2016) Cognitive state following mild stroke: a matter of
receptor"dependent cell proliferation regulates anxiety"
hippocampal mean diffusivity, Hippocampus, 26, 161-169.
like behaviors, Behav. Brain Res., 362, 288-298.
93. Herman, J.P., McKlveen, J.M., Ghosal, S., Kopp, B.,
80.
Li, Y., Qin, J., Yan, J., Zhang, N., Xu, Y., Zhu, Y.,
Wulsin, A., Makinson, R., Scheimann, J., and Myers, B.
Sheng, L., Zhu, X., and Ju, S. (2018) Differences of phys"
(2016) Regulation of the hypothalamic"pituitary"adreno"
ical vs. psychological stress: evidences from glucocorticoid
cortical stress response, Compr. Physiol., 6, 603-621.
receptor expression, hippocampal subfields injury, and
94. Pochigaeva, K., Druzhkova, T., Yakovlev, A., Onufriev, M.,
behavioral abnormalities, Brain Imaging Behav.,
Grishkina, M., Chepelev, A., Guekht, A., and Gulyaeva, N.
doi: 10.1007/s11682"018"9956"3.
(2017) Hair cortisol as a marker of hypothalamic"pituitary"
81.
Leonard, B.E. (2007) Inflammation, depression and
adrenal Axis activity in female patients with major depres"
dementia: are they connected? Neurochem. Res., 32,
sive disorder, Metab. Brain Dis., 32, 577-583.
1749-1756.
95. Druzhkova, T., Pochigaeva, K., Yakovlev, A., Kazimirova, E.,
82.
Herbert, J., and Lucassen, P.J. (2016) Depression as a risk
Grishkina, M., Chepelev, A., Guekht, A., and Gulyaeva, N.
factor for Alzheimer’s disease: genes, steroids, cytokines
(2019) Acute stress response to a cognitive task in patients
and neurogenesis - what do we need to know? Front.
with major depressive disorder: potential metabolic and
Neuroendocrinol., 41, 153-171.
proinflammatory biomarkers, Metab. Brain Dis., 34,
83.
Kino, T. (2015) Stress, glucocorticoid hormones, and hip"
621-629.
pocampal neural progenitor cells: implications to mood
96. Joëls, M., Karst, H., and Sarabdjitsingh, R.A. (2018) The
disorders, Front. Physiol., 6, 230.
stressed brain of humans and rodents, Acta Physiol. (Oxf.),
84.
Levada, O.A., and Troyan, A.S. (2018) Poststroke depres"
223, e13066.
sion biomarkers: a narrative review, Front. Neurol., 16, 577.
97. De Kloet, E.R., Otte, C., Kumsta, R., Kok, L., Hillegers, M.H.,
85.
Ben Assayag E., Korczyn, A.D., Giladi, N., Goldbourt, U.,
Hasselmann H., Kliegel, D., and Joëls, M. (2016) Stress
Berliner, A.S, Shenhar"Tsarfaty, S., Kliper, E., Hallevi, H.,
and depression: a crucial role of the mineralocorticoid
Shopin, L., Hendler, T., Baashat, D.B., Aizenstein, O.,
receptor, J. Neuroendocrinol., 28, doi: 10.1111/jne.12379.
Soreq, H., Katz, N., Solomon, Z., Mike, A., Usher, S.,
98. Martocchia, A., Curto, M., Toussan, L., Stefanelli, M.,
Hausdorff, J.M., Auriel, E., Shapira, I., and Bornstein, N.M.
and Falaschi, P. (2011) Pharmacological strategies against
(2012) Predictors for poststroke outcomes: the Tel Aviv
glucocorticoid"mediated brain damage during chronic dis"
Brain Acute Stroke Cohort (TABASCO) study protocol,
orders, Recent Pat. CNS Drug Discov., 6, 196-204.
Int. J. Stroke, 7, 341-347.
99. Müller, M., Holsboer, F., and Keck, M.E. (2002) Genetic
86.
Molad, J., Hallevi, H., Korczyn, A.D., Kliper, E., Auriel, E.,
modification of corticosteroid receptor signalling: novel
Bornstein, N.M., and Ben Assayag, E. (2019) Vascular and
insights into pathophysiology and treatment strategies of
neurodegenerative markers for the prediction of post"
human affective disorders, Neuropeptides., 36, 117-131.
stroke cognitive impairment: results from the TABASCO
100. Sutanto, W., and de Kloet, E.R. (1991) Mineralocorticoid
study, J. Alzheimer’s Dis., doi: 10.3233/JAD"190339.
receptor ligands: biochemical, pharmacological, and clini"
87.
Molad, J., Ben"Assayag, E., Korczyn, A.D., Kliper, E.,
cal aspects, Med. Res. Rev., 11, 617-639.
Bornstein, N.M., Hallevi, H., and Auriel, E.
(2018)
101. Gomez"Sanchez, E.P. (2016) Third"generation mineralo"
Clinical and radiological determinants of transient symp"
corticoid receptor antagonists: why do we need a fourth? J.
toms associated with infarction (TSI), J. Neurol. Sci., 390,
Cardiovasc. Pharmacol., 67, 26-38.
195-199.
102. Wang, W., Liu, L., Yang, X., Gao, H., Tang, Q.K., Yin, L.Y.,
88.
Kliper, E., Ben Assayag, E., Tarrasch, R., Artzi, M.,
Yin, X.Y., Hao, J.R., Geng, D.Q., and Gao, C. (2019)
Korczyn, A.D., Shenhar"Tsarfaty, S., Aizenstein, O.,
Ketamine improved depressive"like behaviors via hip"
Hallevi, H., Mike, A., Shopin, L., Bornstein, N.M., and
pocampal glucocorticoid receptor in chronic stress
Ben Bashat, D. (2014) Cognitive state following stroke: the
induced"susceptible mice, Behav. Brain Res., 364, 75-84.
predominant role of preexisting white matter lesions, PLoS
103. Scheimann, J.R., Mahbod, P., Morano, R., Frantz, L.,
One, 9, e105461.
Packard, B., Campbell, K., and Herman, J.P. (2018)
БИОХИМИЯ том 84 вып. 11 2019
1648
ГУЛЯЕВА
Deletion of glucocorticoid receptors in forebrain
growth factor system in the aging rat brain, Restor. Neurol.
GABAergic neurons alters acute stress responding and pas"
Neurosci., 35, 571-581.
sive avoidance behavior in female mice, Front. Behav.
105. Joëls, M. (2008) Functional actions of corticosteroids in
Neurosci., 12, 325.
the hippocampus, Eur. J. Pharmacol., 583, 312-321.
104. Stepanichev, M., Onufriev, M., Aniol, V., Freiman, S.,
106. Gulyaeva, N.V. (2017) Molecular mechanisms of neuro"
Brandstaetter, H., Winter, S., Lazareva, N., Guekht, A.,
plasticity: an expanding universe, Biochemistry (Moscow),
and Gulyaeva, N. (2017) Effects of cerebrolysin on nerve
82, 237-242.
BIOCHEMICAL MECHANISMS AND TRANSLATIONAL RELEVANCE
OF HIPPOCAMPAL VULNERABILITY TO DISTANT FOCAL BRAIN INJURY:
THE PRICE OF STRESSWRESPONSE
Review
N. V. Gulyaeva1,2*
1 Department of Functional Biochemistry of the Nervous System,
Institute of Higher Nervous Activity and Neurophysiology, Russian Academy of Sciences,
117485 Moscow, Russia; E(mail: nata_gul@mail.ru
2 Moscow Research and Clinical Center for Neuropsychiatry,
Healthcare Department of Moscow, 115419 Moscow, Russia
Received June 7, 2019
Revised August 24, 2019
Accepted August 24, 2019
Focal brain injuries (in particular, stroke and traumatic brain injury) induce with high probability the development of
delayed (months, years) cognitive and depressive disturbances which are frequently comorbid. The association of
these complications with hippocampal alterations (in spite of the lack of a primary injury of this structure), as well as
the lack of a clear dependence between the probability of depression and dementia development and primary damage
severity and localization served as the basis for a new hypothesis on the distant hippocampal damage as a key link in
the pathogenesis of cognitive and psychiatric disturbances. According to this hypothesis, the excess of corticosteroids
secreted after a focal brain damage, in particular in patients with abnormal stress"response due to hypothalamo"pitu"
itary"adrenal axis dysfunction, interacts with corticosteroid receptors in the hippocampus inducing signaling path"
ways which stimulate neuroinflammation and subsequent events including disturbances in neurogenesis and hip"
pocampal neurodegeneration. In this article, the molecular and cellular mechanisms associated with the regulatory
role of the hypothalamo"pituitary"adrenal axis and multiple functions of brain corticosteroid receptors in the hip"
pocampus are analyzed. Functional and structural damage to the hippocampus, a brain region selectively vulnerable
to external factors and responding to them by increased cytokine secretion, forms the basis for cognitive function dis"
turbances and psychopathology development. This concept is confirmed by our own experimental data, results of
other groups and by prospective clinical studies of post"stroke complications. Clinically relevant biochemical
approaches to predict the risks and probability of post"stroke/post"trauma cognitive and depressive disturbances are
suggested using the evaluation of biochemical markers of patients’ individual stress"response. Pathogenetically justi"
fied ways for preventing these consequences of focal brain damage are proposed by targeting key molecular mecha"
nisms underlying hippocampal dysfunction.
Keywords: hippocampus, stress, hypothalamo"pituitary"adrenal axis, corticosteroids, glucocorticoid receptor, mineralo"
corticoid receptor, cytokines, stroke, traumatic brain injury, depression, dementia
БИОХИМИЯ том 84 вып. 11 2019