БИОХИМИЯ, 2019, том 84, вып. 12, с. 1801 - 1814
УДК 577.24
В ЧЕМ ЗАКЛЮЧАЕТСЯ СМЫСЛ
АНТАГОНИСТИЧЕСКОЙ ПЛЕЙОТРОПИИ?
Обзор
© 2019
Дж. Миттельдорф1,2
1 Washington University School of Medicine, St. Louis, MO, USA; E mail: aging.advice@gmail.com
2 National Institute for Biological Sciences, Beijing, China
Поступила в редакцию 05.07.2019
После доработки 19.09.2019
Принята к публикации 19.09.2019
Биологическое старение рассматривается как явление, генетически ассоциированное с повышением пло
довитости или выживаемости живых организмов в раннем возрасте. Это интерпретировано как подтверж
дение для преобладающей теории эволюции старения, называемой как антагонистическая плейотропия
(AP). В настоящем обзоре представлено альтернативное объяснение наблюдаемой плейотропии, согласно
которому плейотропия является скорее адаптацией в своем собственном смысле, а не физическим предва
рительным условием для биологической эволюции. Эволюционная теория антагонистической плейотро
пии впервые была предложена 60 лет назад Джорджем Уильямсом. Содержание ее заключается в том, что те
гены, которые выгодны для индивида в раннем возрасте, обладают неизбежными плейотропными эффекта
ми, которые вызывают клеточное старение в дальнейшей жизни. Основным сильным аргументом в пользу
теории антагонистической плейотропии является то, что она объясняет отбор признака, который является
вредным для приспособляемости индивида (старение) без учета его влияния на коллективную выгоду (груп
повой отбор). Слабой стороной теории является то, что компромиссы не кажутся неизбежными. Известны
примеры мутаций, которые вызывают повышение продолжительности жизни без очевидного взимания за
это платы. Факт, заключающийся в том, что дикие типы этих генов вызывают клеточное старение без пре
доставления сопутствующих индивидуальных выгод, по видимому, представляет собой доказательство су
ществования другого механизма отбора, зависимого от выгоды для приспособляемости в результате старе
ния, которое может быть только коллективным, но не индивидуальным. В связи с тем, что теория антаго
нистической плейотропии была выдвинута относительно давно, были предложены различные правдопо
добные механизмы многоуровневого отбора, согласно которым старение возникает как общая адаптация. В
контексте адаптивных теорий старения возникает вопрос: почему антагонистическая плейотропия продол
жает существовать в природе, если ее можно избежать? В настоящей статье предположено, что естествен
ный отбор может действительно благоприятствовать плейотропии как развившейся адаптации. Это проис
ходит потому, что комбинация высокой плодовитости и долгой продолжительности жизни является иску
шением для индивидов и в то же время опасностью для стабильности популяций. Популяции хищников,
которые растут быстрее, чем их жертвы, могут подвергаться риску воздействия хаотической динамики чис
ленности популяции, приводящей к ее вымиранию. Как только устойчивая смесь плодовитости и продол
жительности жизни устанавливается в результате действия многоуровневого естественного отбора, плейот
ропия может помочь убедиться, что она не потеряна. Популяция свободно может сдвигаться от одного сос
тояния (высокая плодовитость/короткая продолжительность жизни) к другому состоянию (более низкая
плодовитость/более высокая продолжительность жизни), как того требуют изменяющиеся условия окружаю
щей среды, без риска превышения предельной численности популяции и ее коллапса. В данном обзоре опи
саны эксперименты с проведением ориентированного на индивида компьютерного моделирования, в кото
рых плейотропия развивается как адаптация на основе группового отбора в соответствии с рядом предпо
ложений и в широком диапазоне пространства параметров.
КЛЮЧЕВЫЕ СЛОВА: старение, запрограммированное старение, антагонистическая плейотропия, группо
вой отбор, многоуровневый отбор.
DOI: 10.1134/S0320972519120054
Антагонистическая плейотропия (AP) явля
веденных демографических исследований про
ется в настоящее время наиболее широко при
цесса старения в дикой природе [3-5]. Теория
нятым объяснением эволюции процесса старе
гласит, что есть гены (или выработанные паттер
ния [1, 2]. Основную конкуренцию ей составля
ны генной экспрессии), ассоциированные с
ет «теория накопления мутаций», которая под
фертильностью и другими элементами приспо
вергнута сомнению в 1990 e гг. в результате про
собленности индивида, которые в дальнейшей
3
1801
1802
МИТТЕЛЬДОРФ
жизни тесным образом связаны с потерей гомео
ни и более медленная скорость плавания в срав
стаза и саморазрушением, чтобы естественный
нении с сестринской популяцией, находящейся
отбор смог принять последнее как неизбежную
даже на расстоянии в несколько сот метров от
плату за первое.
места, кишащего хищниками. Предположи
Клеточное старение представляет дилемму
тельно, что более низкая плодовитость и более
для теории эволюции, так как оно имеет опреде
короткая продолжительность жизни были необ
ленную генетическую основу [6], несмотря на
ходимы для поддержания численности популя
его полностью отрицательное воздействие на
ции в условиях отсутствия давления со стороны
приспособляемость индивида. Преимущество
хищников, но регуляция численности популя
теории антагонистической плейотропии заклю
ции является групповой выгодой, несовмести
чается в том, что она решает эту дилемму внутри
мой с рамками теории антагонистической
контекста бесспорных механизмов отбора, ко
плейотропии [17].
торые укоренились в соперничестве индивидов.
Демографические исследования, в целом,
Эта теория не подразумевает групповой отбор.
также не выявили «платы за размножение».
Недостатком ее является то, что эмпирические
Просматривая царство животных, Finch [18]
доказательства существования плейотропных
пришел к заключению, что размножение часто
связей неравномерны [7]. Были обнаружены
сталкивается с риском немедленной гибели. Од
многочисленные примеры плейотропии, но нет
нако нет доказательств того, что фертильность
доказательств того, что они неминуемы или они
связана с ускоренным старением в дальнейшей
каким либо значительным образом препятству
жизни. Исследования в области демографии че
ют естественному отбору [8]. Хуже того, было
ловека также не выявили отрицательную корре
обнаружено множество генов, которые ускоря
ляцию между женской фертильностью и про
ют процесс старения, для которого не было вы
должительностью жизни женщины [19], иногда
явлено сопутствующей выгоды. В классической
говорится о небольшом положительном эффек
работе по эволюции процесса старения в 1992 г.
те [20, 21]. Некоторые теоретики считают, что
Stearns [9] составил таблицу таких генов многих
отсутствие свидетельств существования платы
видов, и в течение 26 прошедших лет их число
за размножение не может рассматриваться как
вышло за пределы таблицы. Kenyon et al. [10]
свидетельство против рассматриваемой теории,
описали случаи червей и других видов, у кото
так как естественный отбор подделал компро
рых нарушение активности гена, который пре
мисс между плодовитостью и продолжитель
обладает в диком типе, приводит к появлению
ностью жизни на генетическом уровне, а не в
здорового, долгоживущего мутантного потомка.
продолжительности жизни какого либо инди
Неравномерные («пятнистые») доказатель
вида. Демографические доказательства были бы
ства были интерпретированы как частичная
проклятием для теории одноразовой сомы, так
поддержка [11], хотя, по мнению автора настоя
как она основывается на метаболических комп
щего обзора, это несовместимо с предсказания
ромиссах, но не для других версий теории анта
ми теории антагонистической плейотропии [7].
гонистической плейотропии. Но в любом случае
Непоследовательность плейотропии подрывает
многие примеры бесплатных генетических мо
основную логику этой теории, так как она зави
дификаций, которые увеличивают продолжи
сит от положения, заключающегося в том, что
тельность жизни, являются проблемой для тео
издержки и выгоды плейотропии неразделимы
рии антагонистической плейотропии.
друг от друга.
Чтобы понять плейотропные связи между
Противоположные доказательства включают
плодовитостью и биологическим старением, ко
в себя как генетические, так и демографические
торые не соответствуют друг другу, можно пред
примеры.
ложить другое объяснение. В контексте теорий
Есть (генетические) примеры, когда клеточ
адаптивного старения [7, 22-24] антагонисти
ное старение было развито, несмотря на факт
ческая плейотропия рассматривается как воз
существования определенной, бесплатной аль
можный адаптивный признак, который сохра
тернативы. Наиболее известным таким приме
няет долгосрочные выгоды для сообщества, ко
ром является приспособленность популяции
торые приносит процесс старения от потери до
тринидадских гуппи, которая описана в работе
кратковременной власти силы индивидуального
[12], но также имеются и многие другие приме
отбора. Эта перспектива предполагает, что меха
ры [13-16]. В частности, Reznick [13] обнару
низм клеточного старения был выработан в ре
жил, что у этих пресноводных рыбок, находя
зультате долгосрочного действия многоуровне
щихся в условиях низкой опасности со стороны
вого отбора как некий компромисс между выго
хищников, развивается более низкая фертиль
дой для индивида в связи с высокой продолжи
ность, более короткая продолжительность жиз
тельностью жизни и риском для сообщества
БИОХИМИЯ том 84 вып. 12 2019
АНТАГОНИСТИЧЕСКАЯ ПЛЕЙОТРОПИЯ
1803
из за хаотичной популяционной динамики. Со
ческое мышление, чтобы представить себе, что
четание высокой продолжительности жизни и
естественный отбор мог быть таким мудрым,
повышенной рождаемости приводит к повыше
как защищать клеточное старение от действия
нию скорости роста численности популяций,
краткосрочного отбора, так как он несет риск
которая настолько быстра по сравнению со ско
вымирания в результате превышения числен
ростью восстановления видов, служащих в каче
ности популяции. До настоящего времени автор
стве источника пищи. Превышение численнос
отвечал на это с приведением: 1) бесспорного
ти популяции становится явно выраженным,
примера параллельного случая, в котором есте
тяжелым, приводящим к хаотичной популяци
ственный отбор привел к созданию такого спо
онной динамике и ее вымиранию
[7,
25].
соба защиты от краткосрочного преимущества,
Плейотропия может являться развившимся ме
и 2) простой компьютерной модели, рассматри
ханизмом, чтобы избежать эту Трагедию общест
вающей групповой отбор и индивидуальный от
венного достояния (Tragedy of the сommons) [26].
бор в их естественном балансе, при котором
Быстро протекающий эффективный индивиду
можно видеть, что плейотропия активно разви
альный отбор всегда способствует повышению
вается как адаптация.
плодовитости и продолжительности жизни, но
эти факторы всегда находятся в обратной зави
симости друг от друга и поэтому удается избе
ПРЕЦЕДЕНТЫ
жать опасности для популяции. В популяции
могут сохраниться различные стратегии с ком
Бесспорным примером выступает эволюция
бинациями (долгая продолжительность жиз
связи между полом и размножением, которое
ни/низкая плодовитость) или (короткая про
возникло примерно во время Кембрийского
должительность жизни/высокая плодовитость ),
взрыва. Если бы обмен генами был необязатель
но популяция при этом защищена от «мошен
ным, тогда был бы сильным индивидуальный
ников», которые могут взять на себя управление
отбор, чтобы избежать обмена генами. Половое
популяцией с неустойчивой комбинацией из
размножение требует затрат времени и энергии.
долгой продолжительности жизни и высокой
Половое размножение разбивает комбинации
плодовитости, подталкивающей популяцию к
генов, которые были ранее проверены и извест
перенаселенности и исчезновению. Поддержа
но, что они эффективно работают вместе. В слу
ние продукта продолжительности жизни и пло
чае наиболее приспособившихся организмов с
довитости под контролем важно для демографи
наиболее адаптированными комбинациями ге
ческой стабильности популяций и экосистем [7,
нов есть плата за обмен генов с конкурентами.
27, 28], и антагонистическая плейотропия как
Пол стал связанным с размножением опреде
приобретенный в результате эволюции признак
ленно для того, чтобы сделать его обязательным.
обеспечивает выгоду от подавления появления
В случае большинства многоклеточных видов
мошенников.
животных и растений имеются мощные генети
Могут возникнуть возражения, что плейот
ческие и анатомические барьеры, предотвраща
ропия присутствует повсюду в геноме и нет не
ющие эволюционный возврат к бесполому
обходимости в существовании какого то эволю
размножению. Даже у организмов-гермафро
ционного механизма, чтобы объяснить проис
дитов существует физические препятствия для
хождение плейотропии в этом особом случае.
самооплодотворения. Связь между размноже
Однако универсальная плейотропия берет нача
нием и полом настолько для нас привычна, что
ло от экономики природы, повторного исполь
мы можем забыть о том, что сама связь должна
зования старых инструментов для решения но
иметь адаптивное значение, иначе она не могла
вых задач. Плейотропия возникает, когда эво
бы развиться.
люция продвигается по пути наименьшего соп
Половое размножение под видом конъюга
ротивления. Антагонистическая плейотропия
ции впервые развилось у одноклеточных прос
требует эволюционного объяснения, так как она
тейших, и на протяжении сотен миллионов лет
не является примером двойного использования
оно представляло собой вид активности, неза
одного признака, а скорее представляет собой
висимый от размножения [29]. Размножение
вынужденное принятие связи, которая является
происходит в виде клонирования клеток, и по
дорогостоящей для приспособляемости инди
ловое размножение здесь приобретает форму
вида.
конъюгации. Половое размножение не несет
С точки зрения обычной популяционной ге
адаптивной ценности для индивида, и первым
нетики, возникает другое возражение, заключа
механизмом, который был выработан для усиле
ющееся в том, что «естественный отбор не име
ния процесса обмена генами, был механизм
ет предвидения». Это выглядит как телеологи
клеточного старения - раннего способа прог
БИОХИМИЯ том 84 вып. 12 2019
3*
1804
МИТТЕЛЬДОРФ
раммированного старения. У жгутиковых линии
котором необходимо сказать, что он не обладает
клонов клеток со временем теряют свою жиз
немедленным воздействием на приспособляе
неспособность и для их восстановления необхо
мость организма, но который модулирует дина
дима конъюгация. Обязательная связь механиз
мику естественного отбора в ходе эволюции.
мов восстановления с процессом конъюгации
Эволюция способности к эволюционированию
является примером развившейся плейотропии
не соответствует стандартным моделям популя
[30].
ционной генетики [32], но тем не менее мы зна
С появлением многоклеточных форм жизни
ем, что она имеет место [33, 34]. В дополнение к
данный способ делать половое размножение
половому размножению, к широко распростра
обязательным перестал быть доступным. Весьма
ненным признакам способности к эволюциони
примечательно [31], что половое размножение
рованию относятся различия в скоростях мута
стало анатомически привязано к репродукции,
ции, иерархическая структура генома, сохране
усиливая тем самым императив делиться генами
ние биоразнообразия, старение и собственно ге
при каждом жизненном цикле. Среди теорети
нетический код [33, 35, 36]. Принимаем мы или
ков процесса эволюции в течение десятилетий
не приемлем предполагаемые механизмы отбо
обсуждалась цель полового размножения. Но
ра адаптации способности к эволюционирова
для обеспечения связи между полом и размно
нию, мы все равно должны принять сам факт,
жением может быть только одна мыслимая цель,
что способность к эволюционированию каким
и это обеспечение совместного использования
то образом развилась.
генов путем эффективного блокирования спол
зания к бесполому способу размножения. При
чина, по которой половое размножение стало
ЛОГИСТИЧЕСКАЯ МОДЕЛЬ ЗАДЕРЖКИ
предпосылкой для репродукции, заключается в
РОСТА ПОПУЛЯЦИИ
том, что избирательное преимущество полового
размножения достается только демону, а расхо
Компьютерная модель эволюционного про
ды несут только индивиды. Без существования
исхождения антагонистической плейотропии
такой тесной связи между полом и размножени
основывается на метапопуляции локальных
ем половое размножение легко могло быть утра
субпопуляций, каждая из которых находится
чено в результате эффективного действия меха
под управлением отложенной логистической
низма индивидуального отбора, а сообщество
динамики [25, 27, 37]. Логическое уравнение яв
утратило бы свое разнообразие и способность
ляется наиболее предпочтительной моделью для
адаптироваться к постоянно меняющимся усло
описания поведения популяций при условии
виям окружающей среды.
фиксированной несущей емкости (carrying
Таким образом, эволюция облигатного по
capacity):
лового размножения обеспечивает предшест
венника для механизма, который, как полагает
dx/dt = ax(xss x).
автор, несет ответственность за эволюцию
плейотропии. Как половое размножение, так и
Решения логистического дифференциаль
старение приносят групповую выгоду и индиви
ного уравнения всегда устойчивы для популя
дуальные издержки. Оба процесса подвергаются
ции x, восстанавливающейся плавно и эффек
опасности исчезновения до тех пор, пока они не
тивно до стационарного уровня xss. Но в реаль
защищены от быстрого и эффективного воздей
ной действительности, обратная связь, касаю
ствия индивидуального отбора. Половое разм
щаяся устойчивого стационарного уровня попу
ножение защищено от действия индивидуаль
ляции, возникает не сразу. Для животных, в ок
ного отбора путем разделения средств размно
ружающей их среде всегда имеется источник
жения между двумя индивидами таким образом,
пищи. В течение длительного периода времени
что для того чтобы произошло размножение,
источник пищи обновляется, однако на корот
каждый индивид должен поделиться генами.
ком промежутке времени источники пищи мо
Процесс старения защищен от влияния индиви
гут быть значительно истощены в результате не
дуального отбора с помощью многочисленных и
контролируемого потребления. Пока еще мож
различных по характеру своего действия спосо
но найти пищу, размножение может происхо
бов генетической плейотропии, связывающей
дить беспрепятственно, не обращая внимания
продолжительность жизни с существенными
на надвигающуюся нехватку. Популяция может
затратами на приспособляемость.
продолжать превышать свой устойчивый уро
Подобно половому размножению антаго
вень численности до тех пор, пока острая нех
нистическая плейотропия представляет собой
ватка продовольствия не приведет к массовому
признак способности к эволюционированию, о
вымиранию. Таким образом, доступность пищи
БИОХИМИЯ том 84 вып. 12 2019
АНТАГОНИСТИЧЕСКАЯ ПЛЕЙОТРОПИЯ
1805
обеспечивает основу для логистической дина
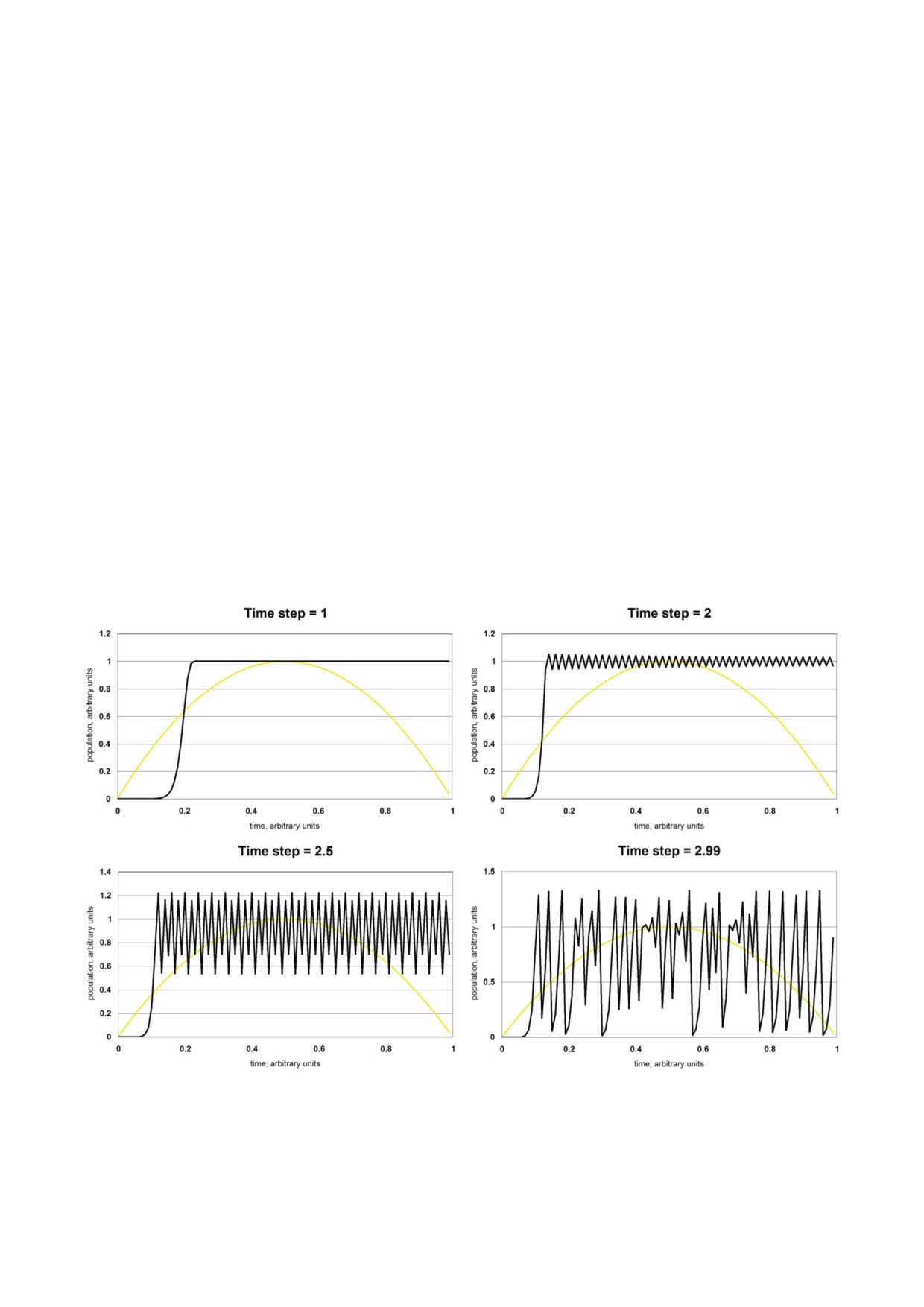
вести себя хаотически. При значениях >3 чис
мики популяции, при которой восстановитель
ленность популяции зашкаливает и затем она
ный член в уравнении обычно задерживается во
вымирает за один цикл. (рис. 1).
времени. В отличие от дифференциальной фор
Эвристическое поведение популяций с отло8
мы логистического уравнения, решения логис
женной логистической обратной связью. Значе
тического уравнения с временной задержкой
ние задержки является параметром окружаю
могут быть либо стабильными, либо флуктуиру
щей среды, не поддающимся генетическому
ющими, либо хаотичными.
контролю. Но скорость роста численности по
На первых этапах развития теории хаоса,
пуляции зависит от плодовитости и продолжи
дифференциальное логистическое уравнение с
тельности жизни, признаков, которые оба были
дискретным временем было первым уравнени
выработаны в ходе эволюции. Мы ожидаем, что
ем, которое было использовано для демонстра
индивидуальный отбор будет способствовать
ции хаотической динамики [38, 39]. Выбор меж
повышению плодовитости и продолжительнос
ду гладким поведением и колеблющимся или
ти жизни, создавая все бóльшие скорости роста
хаотичным поведением предсказывается пара
численности на уровне популяции. По мере то
метром хаоса, определяемым как продукт диск
го, как растет скорость повышения численности
ретного временного шага Δt и (логарифмичес
популяции, значение параметра хаоса прибли
кой) скорости роста численности популяции.
жается к 3, и локальная популяция разрушает
Если величина параметра хаоса является незна
сама себя.
чительной, то происходит плавное восстановле
В случае метапопуляции любое локальное
ние численности популяции до ее стационарно
вымирание будет носить временный характер,
го уровня. Если этот параметр превышает значе
поскольку популяция может повторно занять
ние 2, то наблюдаются некоторые колебания.
место обитания, переместившись в него из дру
Как только параметр хаоса достигает значения 3,
гой территории, на которой не произошло вы
то кривая численности популяции начинает
мирания. Территория, с которой переселяются
Рис. 1. Временной график дискретного логистического уравнения. По мере того, как приращение времени Δt превысит
значение, равное примерно 2,5, поведение решения изменяется от периодического на хаотичный.
biokhsm/
БИОХИМИЯ том 84 вып. 12 2019
1806
МИТТЕЛЬДОРФ
основатели новой популяции вместо вымершей
цев, и они снова постоянно возникают на гра
для повторного посева вымершего сайта, веро
фике.) Резко, примерно при времени = 12 000,
ятно, будет иметь более низкую плодовитость и
одно возобновление роста численности популя
продолжительность жизни просто потому, что
ции осуществляется агентом, эволюциониро
они не уничтожили сами себя. По мере того, как
вавшим для поддержания стабильного темпа
этот процесс вымирания и повторного зарожде
роста, и в течение некоторого времени популя
ния повторяется, метапопуляция эволюциони
ция временно стабильна. Однако вскоре инди
рует в сторону скорости роста вблизи порога ха
видуальный отбор снова способствует тому, что
оса. Динамика вымираний и повторных зарож
повышается рождаемость и продолжительность
дений держит темпы роста под контролем, поэ
жизни индивидов, и снова наступает этап глубо
тому случаи вымирания стали нечастыми.
кой цикличности. В глобальном масштабе мета
Конечно, индивидуальный отбор способ
популяция выживает только потому, что отдель
ствует как более долгой продолжительности
ные участки не исчезают сразу, но очаги ста
жизни, так и более высокой рождаемости. Ус
бильности остаются редкими и временными.
тойчивая чистая скорость роста, достаточно вы
Для сравнения, на рис. 2, b показана типичная
сокая для того, чтобы оправиться от падения
картина прогрессии в случае, когда плейотро
численности популяции, и достаточно низкая
пии позволено развиваться. Начальное движе
чтобы избежать порога хаоса, может быть дос
ние в сторону хаоса выглядит подобным обра
тигнута как в случае низкой плодовитости или
зом, но медленнее. Популяция вступает в ста
короткой продолжительности жизни или при
дию глубокой цикличности. Затем, при значе
любом сочетании этих двух признаков. Поэтому,
нии t, примерно равным 14 000, одно из повтор
с точки зрения перспективы группового отбора,
ных возобновлений популяции происходит при
эти две стратегии конкурентно способны. Одна
участии агента, эволюционировавшего для
ко быстрее размножаться и быстрее умирать
обеспечения стабильной скорости роста. В это
имеет преимущество на уровне индивидуально
время стабильность популяции еще не наруше
го отбора. В случае стабильной популяции хоро
на благодаря развитию плейотропии. Скорость
шо известно, что стратегией победы здесь явля
роста численности популяции дальше не растет,
ется тот, у кого более высокий параметр Мальту
так как каждое увеличение плодовитости урав
са r [40], и это соответствует сочетанию высокой
новешивается соответствующим снижением
плодовитости и короткой продолжительности
продолжительности жизни и наоборот. Впослед
жизни.
ствии динамика популяции остается стабильной
В действительности, многие факторы оказы
на протяжении многих тысяч временных шагов
вают влияние на оптимальное сочетание факто
(с правого края рисунка, не показано).
ра продолжительности жизни и плодовитости,
Динамика численности метапопуляции в
включая случайную смертность, родительскую
представленной модели. Я предположил [7] и
заботу, стабильность факторов окружающей
другие исследователи согласились со мной [28],
среды и физические ограничения. Не надо ожи
что динамика численности локальной популя
дать, что более короткая продолжительность
ции может быть быстрой [41], и локальные вы
жизни при более высокой плодовитости всегда
мирания могут быть сильной и быстродейству
может быть оптимальным сочетанием.
ющей силой отбора [42], по эффективности
В соответствии с представленной в настоя
сравнимой с индивидуальным отбором, но дви
щей работе компьютерной моделью, на рис. 2
гающейся в обратном направлении. Это обеспе
представлены два примера динамики популя
чивает основу для наиболее правдоподобного
ции на отдельных участках. На рис. 2, a, можно
сценария для эволюции клеточного старения.
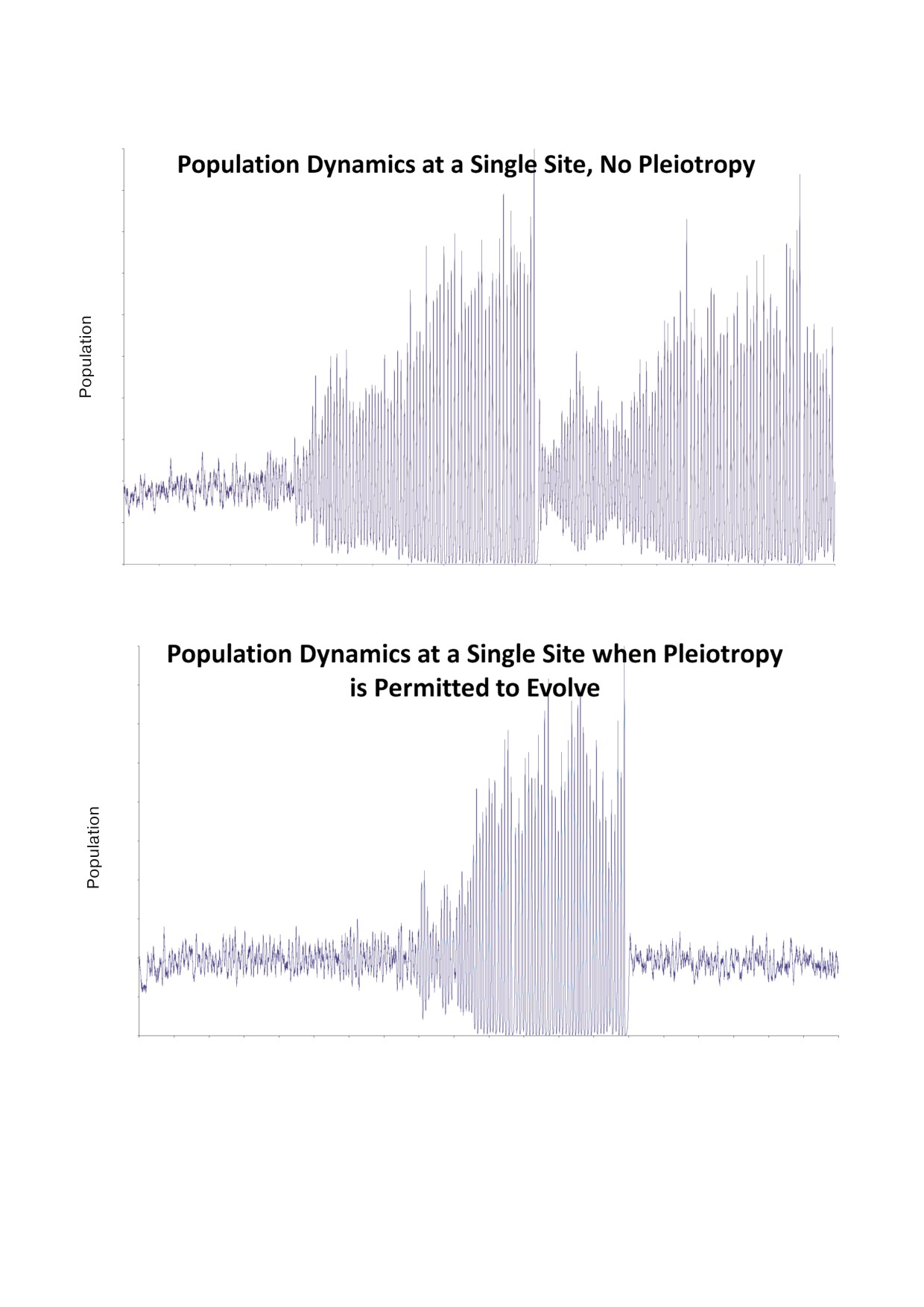
видеть типичную прогрессию развившейся
Даже в случае отсутствия плейотропии (выше,
плейотропии. Продолжительность жизни и пло
рис. 2) рост рождаемости и продолжительности
довитость были инициализированы с низкими
жизни сдерживается за счет постоянных неп
значениями, и поэтому сначала рост численнос
рекращающихся вымираний. Каждый раз, когда
ти популяции населения был медленным по
популяция сама себя обрекает на вымирание,
сравнению с временем задержки. По мере уве
она снова заселяется переселенцами с участка,
личения плодовитости и продолжительности
который находится не очень далеко от само
жизни динамика численности становится нес
убийственного пути ускоренного роста числен
табильной. Начиная примерно с t = 5000, коле
ности. Агент, повторно заселяющий участок, ве
бания численности популяции становятся
роятно, будет иметь более низкую плодовитость
экстремальными и часто приводят к вымира
и продолжительность жизни, чем бывшие (вы
нию. (Популяции, чья численность падает до
мершие) его обитатели. Эта динамика, в целом,
нуля, быстро возобновляются за счет переселен
способствует поддержанию продолжительности
БИОХИМИЯ том 84 вып. 12 2019
АНТАГОНИСТИЧЕСКАЯ ПЛЕЙОТРОПИЯ
1807
a
500
450
400
350
300
250
200
150
100
50
0
1
5001
10001
15001
20001
Time
b
500
450
400
350
300
250
200
150
100
50
0
1
5001
10001
15001
20001
Time
Рис. 2. Кривые роста численности популяции на сайте одной субпопуляции показывают основную динамику отбора внут
ри метапопуляции. a - Без плейотропии; b - с плейотропией.
biokhsm/
БИОХИМИЯ том 84 вып. 12 2019
1808
МИТТЕЛЬДОРФ
жизни и рождаемости под постоянным контро
преобладающим в метапопуляции. Вариант (a).
лем, хотя и на уровнях, которые находятся чуть
Здесь, конечно, есть недооценка силы группо
выше линии неустойчивости. Сам по себе каж
вого отбора, потому что это дает преимущество
дый участок (сайт) нестабилен, но в глобальном
участкам, на которых происходит расцвет попу
масштабе метапопуляция может поддерживать
ляции много раз со стационарной устойчивой
себя бесконечно.
популяцией. В действительности, нужно время,
Когда опция развивающейся плейотропии
чтобы найти новое место обитания, созревшее
была добавлена в модель, у индивидуальных
для ее колонизации, и популяция, которая рас
участков появился ресурс для сдерживания их
цветает и погибает, вряд ли будет успешно экс
самоубийственного марша к более высоким
портироваться. В варианте (b) вероятно, есть пе
темпам роста. Сайты, которые развивают связь
реоценка силы группового отбора, потому что
между более высокой рождаемостью и более ко
он обладает излишним весом в пользу стабиль
роткой продолжительностью жизни, могут из
ных участков в сравнении с нестабильными, ко
бежать повторных рисков и исчезновения и ос
торые могут иметь гораздо большие популяции
таваться стабильными в течение длительного
для их перемещения.
времени. Если индивиды, у которых мутации,
При низких значениях миграции, плейотро
затрагивающие продолжительность жизни и
пия почти всегда развивается до значений меж
фертильность, находятся в обратной зависимос
ду -0,5 и -0,7. В случае более высоких значений
ти, могут доминировать в местной популяции,
миграции, развития плейотропии не происхо
то эта популяция может избежать опасности
дит. Этого следовало ожидать, так как значи
постоянного нахождения на грани хаоса. Преи
тельно высокие скорости миграции эффективно
мущество стабильной популяции реализуется
превращают метапопуляцию в унифицирован
как большая вероятность того, что ее члены ста
ную популяцию, и здесь имеет значение только
нут новыми основателями, которые повторно
индивидуальный отбор.
заселят участки, находившиеся в нестабильном
При очень высоких значениях миграции,
состоянии вплоть до вымирания популяций.
оптимальна так называемая стратегия «расточи
тельности»: сочетание высокой продолжитель
ности жизни и высокой рождаемости приводит
РЕЗУЛЬТАТЫ МОДЕЛИРОВАНИЯ
к большим короткоживущим пикам численнос
ти популяции, за которыми следует несколько
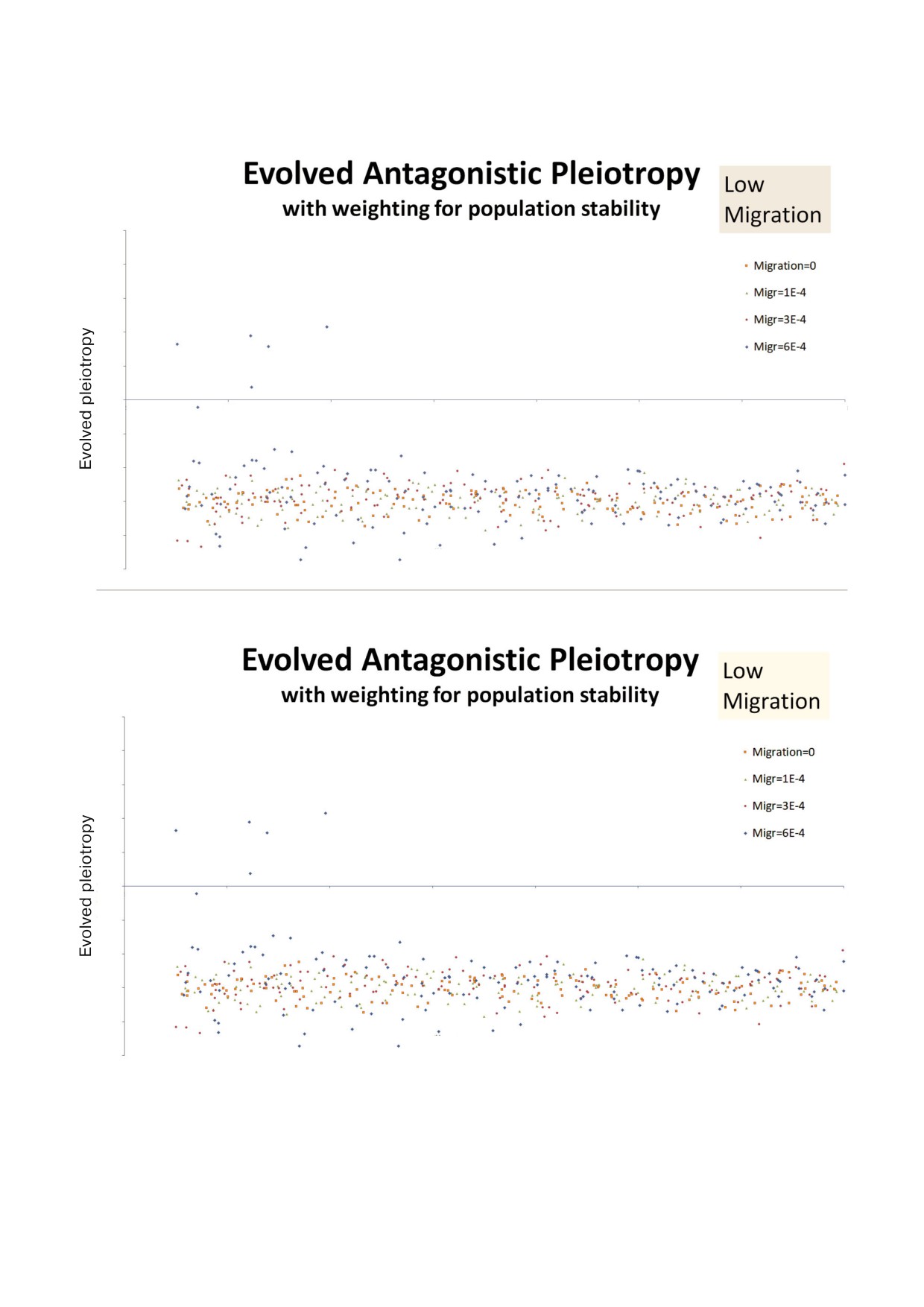
В случае широкого диапазона параметров
циклов локальной экстинкции. Если эта страте
плейотропия развивается до значений от -0,5 до
гия захватывает достаточно много участков, она
-0,7. Это относится к шкале, на которой 0 озна
может поддерживать себя за счет постоянных
чает, что плодовитость и продолжительность
повторных заселений. Стратегия расточитель
жизни изменяются независимо друг от друга и
ства развивается при положительных значениях
между ними нет связи, а значение, равное -1 оз
параметра плейотропии, в противоположность
начает существование четкой обратной корре
антагонистической плейотропии. В варианте (a)
ляции. Знак минус означает, что более высокая
стратегия расточительства работала достаточно
продолжительность жизни связана с более низ
хорошо для высоких значений параметра задер
кой рождаемостью. Модель также допускает по
жки, но очень редко проявлялась при низких
ложительные значения плейотропии, что озна
значениях. В варианте (b) дела параметра расто
чает, что продолжительность жизни и рождае
чительства шли почти всегда плохо. Отрица
мость будут расти и падать параллельно.
тельная плейотропия (AP) развивалась в более,
Результаты были записаны для диапазона
чем 95% запусков, но иногда случался выброс, в
параметра задержки: для пяти значений пара
котором расточительная стратегия брала верх.
метра миграции и двух способов отбора агента
На рис. 3 представлены результаты медлен
основателя, который заселяет каждый участок
ной миграции: 3(a) - это повторное заселение
после вымирания на нем предшественников, а
участка случайным агентом (вариант (a)); 3(b) -
именно: в варианте (a) индивид отбирается слу
это повторное заселение участка с алгоритмом,
чайным образом из метапопуляции, и каждый
который дает больше веса мало изменчивым
индивид имеет равный вес. В варианте (b) пред
сайтам. Следует отметить, что оба результата
почтительны сайты, которые менее изменчивы,
почти всегда остаются в области эволюциони
и поэтому индивиды из стабильных сайтов име
ровавшей отрицательной плейотропии. Но в ва
ют больше шансов на то, чтобы повторно засе
рианте (a) «низкая миграция» означает 0 или
лить новый участок. Важность этого выбора
1 × 10-4. В варианте (a) запуски программы со
очевидна, потому что только при распростране
значением миграции, равным 3 × 10-4 и 6 × 10-4,
нии от участка к участку генотип может стать
в обоих случаях приводили к развитию плейо
БИОХИМИЯ том 84 вып. 12 2019
АНТАГОНИСТИЧЕСКАЯ ПЛЕЙОТРОПИЯ
1809
a
1.0
0.8
0.6
0.4
0.2
0
5
10
15
20
25
30
35
40
−0.2
-0.4
-0.6
-0.8
Logistic delay
-1.0
b
1.0
0.8
0.6
0.4
0.2
0
5
10
15
20
25
30
35
40
−0.2
-0.4
-0.6
-0.8
-1.0
Logistic delay
Рис. 3. Результаты развития плейотропии при низких значениях скорости миграции. а -Каждый агент имел равную воз
можность мигрировать; b - предпочтение в миграции и повторном заселении отдано более стабильным сайтам.
biokhsm/
БИОХИМИЯ том 84 вып. 12 2019
1810
МИТТЕЛЬДОРФ
тропии при низких значениях параметра за
2) каждый агент должен погибнуть в пожи
держки, и затем переходили к отсутствию
лом возрасте, когда его возраст превысит про
плейотропии или положительной плейотропии
должительность жизни, определенную соответ
при более высоких значения параметра задерж
ствующим геном;
ки. В то же время в варианте (b) поведение «низ
3) каждый агент может погибнуть в результа
кая миграция» продолжается при 6 × 10-4, с нес
те перенаселенности с вероятностью, которая
колькими выбросами только на уровне 6 × 10-4.
пропорциональна сайту популяции, как он был
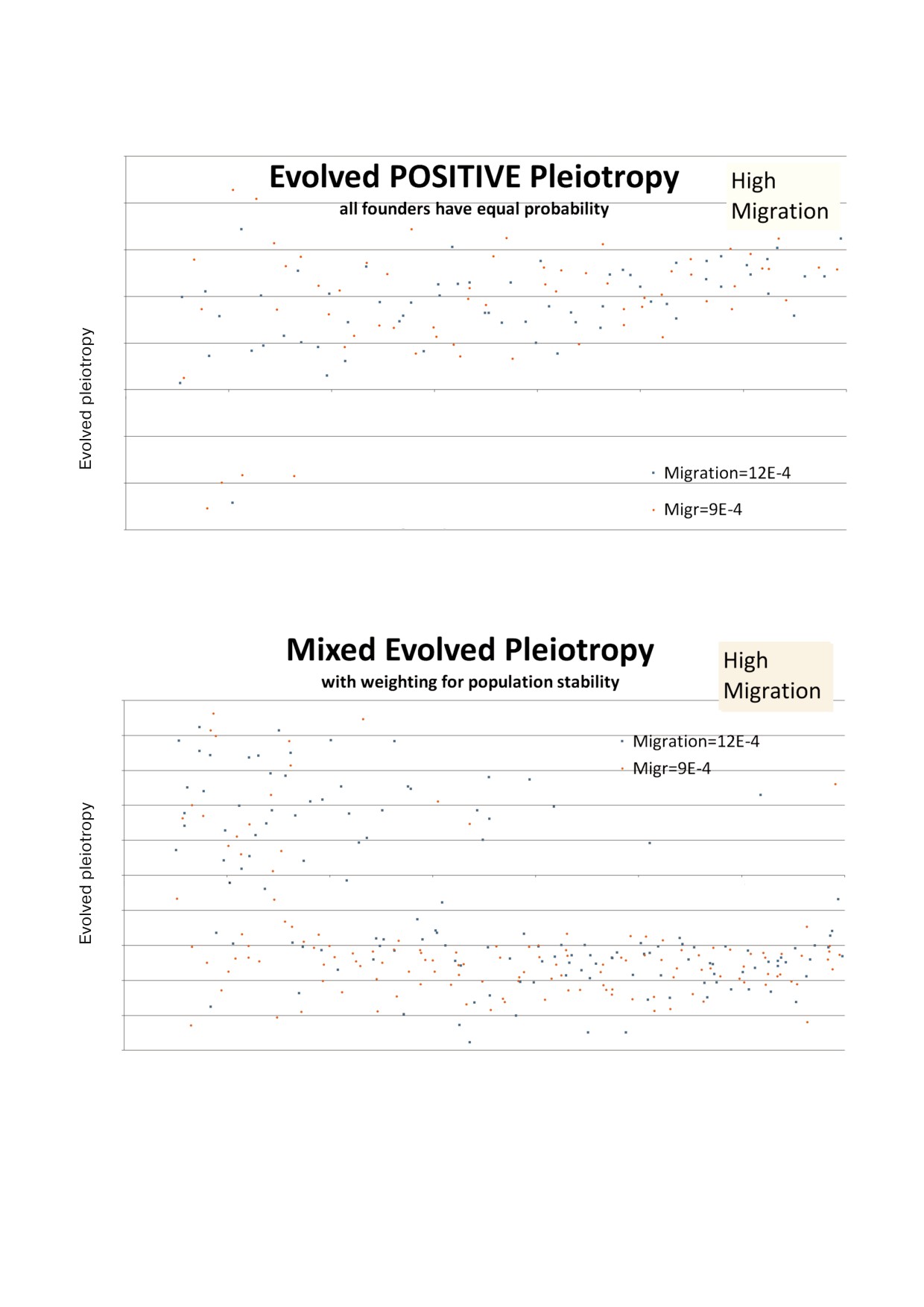
На рис. 4 представлены результаты изучения
равен времени D временного шагов в прошлом,
высоких значений миграции. В варианте (a) ан
где D равен задержке, параметру модели. (Более
тагонистическая плейотропия не развивалась.
точно, вероятность равна [отложенная pop]/100*
В варианте (b) антагонистическая плейотропия
[среднее значение фертильности]). 100 - это ста
продолжает развиваться большинство времени
ционарная популяция, всегда одна и та же для
при значении параметра миграции, равном
всех сайтов и моментов времени. Фертильность
9 × 10-4, и почти половину времени при значе
Avg - это среднее значение фертильности для
нии, равном 12 × 10-4.
всех агентов, присутствующих на сайте. Эта фор
мула предназначена для обозначения того, что
на каждом сайте средняя для конкретного про
ПОДРОБНОЕ ОПИСАНИЕ МОДЕЛИ
межутка времени рождаемость равна скорости
гибели популяции при значении, равном 100.
Ориентированная на индивида модель со
держит 100 сайтов, каждый из которых управля
Когда численность популяции на сайте пос
ется отложенной логистической динамикой
тоянно изменяется вплоть до ее исчезновения,
численности популяции с стационарным чис
то этот сайт заселяется одним случайным аген
лом агентов популяции, равным 100. Логисти
том с одного из оставшихся сайтов (A). В одном
ческая задержка (измеряемая в временных ша
из вариантов агент-основатель выбирается с ве
гах) является параметром модели. Агенты имели
совым коэффициентом для ресурса сайта. Пред
три подвергающихся эволюции гена:
почтительны те сайты, которые продержались
- ген продолжительности жизни. Клеточное
дольше, не подвергаясь колебаниям численнос
старение является шаговой функцией, и агент
ти популяции до ее исчезновения (B). Обосно
резко погибает при достижении им возраста в
вание этой функции заключается в том, что ра
соответствии с генетически отведенным лими
ди эффективности производимых вычислений
том времени;
мы немедленно повторно заселяем освободив
- ген плодовитости. Вероятность (0 < p < 1)
шийся сайт. Однако на самом деле этот процесс
создания (мутированной) копии-клона на каж
является стохастическим и требует времени, от
дом временном шаге;
давая предпочтение тем сайтам, которые длятся
- ген плейотропии. Выражается в виде числа
некоторое время.
от -1 до 1, которое влияет на изменения генов
Необязательно, но есть вероятность (для
фертильности и продолжительности жизни. Для
каждого сайта в каждом временном шаге) для
значения плейотропия, равного 0, мутации, зат
случайной миграции индивидов из другого сай
рагивающие плодовитость и продолжитель
та. Мигрирующий агент также может быть выб
ность жизни, независимы друг от друга. При
ран случайным образом или, как указано выше,
значении параметра плейотропии, равном ±1,
он может быть выбран с весовым коэффициен
изменения плодовитости и продолжительности
том, который предпочитает участки, которые
жизни тесно связаны друг с другом. Повышение
просуществовали дольше. Скорость миграции
рождаемости всегда приводит к пропорцио
является одним из параметров, влияющих на от
нально более высокой (более короткой) продол
носительную силу группового и индивидуаль
жительности жизни. В случае промежуточных
ного отбора. Каждый запуск программы был
значений параметра плейотропия, есть пропор
инициализирован для 25 000 временных шагов,
циональная смесь сцепленных и независимых
чтобы позволить фертильности, продолжитель
мутаций.
ности жизни и плейотропии развиваться мимо
Во время каждого промежутка времени (вре
точки, где начальные условия были потеряны из
менного шага):
памяти системы. Эта сложная система с непред
1) каждый агент может воспроизводиться в
сказуемым поведением, поэтому она в реаль
результате клонирования с вероятностью, кото
ности не достигает стационарного состояния в
рая определяется геном его фертильности. Гены
течение этих временных рамок или вообще ког
потомства подвергаются небольшим, случай
да либо. После 25 000 временных шагов система
ным и связанным друг с другом мутациям;
начинает усредняться по трем разным времен
БИОХИМИЯ том 84 вып. 12 2019
АНТАГОНИСТИЧЕСКАЯ ПЛЕЙОТРОПИЯ
1811
a
1.0
0.8
0.6
0.4
0.2
0
5
10
15
20
25
30
35
40
−0.2
-0.4
-0.6
Logistic delay
b
1.0
0.8
0.6
0.4
0.2
0
5
10
15
20
25
30
35
40
−0.2
-0.4
-0.6
-0.8
-1.0
Logistic delay
Рис. 4. Результаты развития плейотропии при высоких скоростях миграции. а - Каждый агент имел равную возможность
мигрировать; b - предпочтение в вопросе миграции и повторного заселения было дано более стабильным сайтам.
biokhsm/
БИОХИМИЯ том 84 вып. 12 2019
1812
МИТТЕЛЬДОРФ
ным шкалам. После того, как три средних зна
старения, а как свидетельство в пользу теорий
чения согласуются, обычно это достигается че
программированного старения, в которых оно
рез 40 000-50 000 временных шагов, операция
имеет адаптивное значение на популяционном
заканчивается и происходит запись результатов.
уровне.
В действительности, в соответствии с наши
ми результатами, плейотропия иногда сущест
ОБСУЖДЕНИЕ
вует, иногда нет. В некоторых запусках нашей
программы побеждает стратегия расточитель
В то время, когда в 1957 г. Вильямс [1] выдви
ности. В то же время в действительности страте
нул теорию старения, основанную на антаго
гия расточительности ограничивается микроор
нистической плейотропии, его представления о
ганизмами, за редкими, но впечатляющими
генетике соответствовали классической пара
исключениями, вроде нашествия саранчи [44],
дигме Менделя [43]. Естественно было предпо
кроликов в Австралии [45] или других инвазив
ложить, по аналогии с человеческими машина
ных видов [46].
ми, что гены обычно соответствуют признакам,
Когда мы наблюдаем плейотропию, можем
и множественные функции для одного гена бы
ли мы определить, является ли она навязанной
ли исключением из правила. Теперь же мы зна
или развитой? Автор утверждает, что навязанная
ем, что биологические организмы устроены не
плейотропия неизбежна. Это полезный приз
так, как человек, и что множественные перек
нак, который логически подразумевает долго
рывающиеся функции гена являются правилом,
срочную деградацию. Можно привести ряд час
а не исключением. Мы не должны удивляться
то упоминаемых примеров проявления плейо
тому, что гены, регулирующие продолжитель
тропии у человека:
ность жизни, имеют другие функции, потому
• некоторые гены имеют важное значение
что большинство генов имеют более одной
для развития организма. Однако они не могут
функции. Что удивительно, несмотря на высо
быть выключены полностью или выключены в
кую стоимость процесса клеточного старения в
конце жизни с последствиями, которые ускоря
дикой природе [4], эволюция оказалась неспо
ют старение. В их число входит MTOR [47] и не
собна обойти плейотропию путем разделения
которые стероидные гормоны. Самым печально
выгоды от платы. С точки зрения других компо
известным примером является фолликул сти
нентов приспособленности организма, сила в
мулирующий гормон (FSH), содержание кото
одной области не влечет за собой необходимых
рого после наступления менопаузы повышается
затрат в другой. Мы не ожидаем, что животные,
без видимой цели, кроме увеличения риска сер
которые могут летать, должны иметь ухудшен
дечного приступа у женщин [48]. Все эти факто
ное зрение, или что животные с острым обоня
ры, которые находятся под эпигенетическим
нием должны платить цену в виде уменьшения
контролем, являются примерами устранимой
скорости передвижения или физической силы.
плейотропии;
Я предполагаю, что может быть адаптивное зна
• инсулин и инсулиноподобный фактор
чение для поддержания антагонистической свя
роста (IGF) зависимая передача сигнала необ
зи между более высокой фертильностью и более
ходима для роста и развития. В то же время она
короткой продолжительностью жизни.
также повышает риск возникновения рака и
Традиционно, каждый раз плейотропия наб
способствует сокращению продолжительности
людается в случае с геном, который влияет на
жизни [49]. Как и в случае с TOR, это, кажется,
продолжительность жизни. Этот факт был интер
можно лучше объяснить эпигенетическим прог
претирован как подтверждение теории Вильям
раммированием, чем неизбежным компромис
са и теории плейотропии. Однако мы уже виде
сом;
ли, что плейотропия может быть навязана в ка
• воспаление представляет собой первую
честве предварительного условия или же она
линию обороны организма против микробов и
может эволюционировать постфактум. Она мо
основное средство для деградации поврежден
жет быть обязательной или она может быть нео
ной ткани. Тем не менее, воспаление является
бязательной. Теория Вильямса применима к на
наиболее изученным механизмом, который ус
вязанной, обязательной плейотропии. Если
коряет старение и увеличивает риск всех заболе
плейотропия эволюционирует, т.е. естествен
ваний в пожилом возрасте. Ассоциированное со
ный отбор проходит в пользу плейотропии, тог
старением воспаление может быть интерпрети
да это имеет совсем другое значение. Выявлен
ровано как плейотропное следствие воспаления
ные примеры эволюционирующей плейотропии
[50], но воспаление никак не влияет на старение
должны быть интерпретированы не как поддер
в течение первых десятилетий жизни. Таким об
жка теорий антагонистической плейотропии
разом, это можно было бы также рассматривать
БИОХИМИЯ том 84 вып. 12 2019
АНТАГОНИСТИЧЕСКАЯ ПЛЕЙОТРОПИЯ
1813
как тело, использующее механизм самозащиты
заключается в том, что старение вызвано неиз
для его применения саморазрушения в конце
бежными компромиссами.
жизни;
Навязанная антагонистическая плейотро
• недавно паттерны метилирования, ассо
пия - это объяснение тому, почему был вырабо
циированные с ускоренным старением, были
тан механизм старения, несмотря на его цену
также ассоциированы с экспрессией теломера
для индивида. Развившаяся антагонистическая
зы, которая поддерживает клетки в юношеском
плейотропия - это защитный признак, группо
состоянии [51]. По видимому, нет никакой ло
вая адаптация, которая была выстроена поверх
гической причины, по которой эти две функции
более ранней адаптации самого процесса кле
должны быть связаны, поэтому это кандидат,
точного старения.
который следует понимать как пример эволю
ционирующей плейотропии.
Эти и другие примеры были рассмотрены
Конфликт интересов. Автор заявляет об отсут
Leroi et al. [8], которые задаются вопросом, по
ствии конфликта интересов.
чему в биологии животных было выявлено так
Соблюдение этических норм. В данном обзоре
мало примеров антагонистической плейотро
отсутствуют исследования, в которых в качестве
пии. Однако они не ставят под сомнение фунда
экспериментальных моделей были использова
ментальные основы этой теории, суть которых
ны люди или животные.
СПИСОК ЛИТЕРАТУРЫ
1.
Williams, G. (1957) Pleiotropy, natural selection, and the
16.
Williams, P.D., Day, N., Fletcher, Q., and Rowe, L. (2006)
evolution of senescence, Evolution, 11, 398-411.
The shaping of senescence in the wild, Trends Ecol. Evol.,
2.
Rose, M. (1991) Evolutionary biology of aging. Oxford
21, 458-463.
University Press, Oxford, England.
17.
Williams, G. (1966) Adaptation and natural selection.
3.
Promislow, D.E. (1991) Senescence in natural populations of
Princeton University Press.
mammals: a comparative study, Evolution, 45, 1869-1887.
18.
Finch, C.E. (1990) Longevity, senescence and the genome.
4.
Ricklefs, R. (1998) Evolutionary theories of aging: confirma
University of Chicago Press, Chicago.
tion of a fundamental prediction, with implications for the
19.
Jasienska, G. (2017) Costs of reproduction, health, and life
genetic basis and evolution of life span, Am. Nat., 52, 24-44.
span in women, in the Arc of life: evolution and health across
5.
Bonduriansky, R., and Brassil, C.E. (2002) Senescence:
the life course (Jasienska, G., Sherry, D.S., and Holmes,
rapid and costly ageing in wild male flies, Nature, 420, 377.
D.J., eds.) Springer New York, New York, NY,
6.
Guarente, L., and Kenyon, C. (2000) Genetic pathways that
p. 159-176.
regulate ageing in model organisms, Nature, 408, 255-262.
20.
Gavrilova, N.S., Gavrilov, L.A., Semyonova, V.G., and
7.
Mitteldorf, J. (2016) Aging is a group selected adaptation:
Evdokushkina, G.N. (2004) Does exceptional human
theory, evidence, and medical implications. CRC Press.
longevity come with a high cost of infertility? Testing the
8.
Leroi, A.M., Bartke, A., De Benedictis, G., Franceschi,
evolutionary theories of aging, Ann. N.Y. Acad. Sci., 1019,
C., Garther, A., Gonos, E.S., and Fedes, M.E. (2005)
513-517.
What evidence is there for the existence of individual genes
21.
Mitteldorf, J. (2010) Female fertility and longevity, Age
with antagonistic pleiotropic effects? Mech. Ageing Dev.,
(Dordr.), 32, 79-84, doi: 10.1007/s11357 009 9116 1.
126, 421-429.
22.
Longo, V.D., Mitteldorf, J., and Skulachev, V.P. (2005)
9.
Stearns, S.C. (1992) The evolution of life histories. Oxford
Programmed and altruistic ageing, Nat. Rev. Genet., 6,
University Press, Oxford, New York, p. 249.
866-872.
10.
Arantes Oliveira, N., Berman, J.R., and Kenyon, C.
23.
Kirkwood, A., Thomas, B.L., and Melov, S. (2011) On the
(2003) Healthy animals with extreme longevity, Science,
programmed/non programmed nature of ageing within the
302, 611.
life history, Curr. Biol., 21,701-707.
11.
Le Bourg, E. (2001) A mini review of the evolutionary the
24.
Libertini, G. (2015) Non programmed versus programmed
ories of aging. Is it the time to accept them? Dem. Res., 4,
aging paradigm, Curr. Aging Sci., 8, 56-68.
1-28.
25.
Mitteldorf, J., and Goodnight, C. (2012) Post reproduc
12.
Bassar, R.D., Ferriere, R., López Sepulcre, A.,
tive life span and demographic stability, Oikos, 121,
Marshall, M.C., Travis, J., Pringle, C.M., and Reznick,
1370-1378.
D.N. (2012) Direct and indirect ecosystem effects of evo
26.
Hardin, G. (1968) The tragedy of the commons, Science,
lutionary adaptation in the Trinidadian guppy (Poecilia
162, 1243-1248.
reticulata), Am. Nat., 180, 167-185.
27.
Mitteldorf, J. (2006) Chaotic population dynamics and the
13.
Reznick, D., Nunney, L., and Tessier, A. (2000) Big houses,
evolution of aging: proposing a demographic theory of
big cars, superfleas and the costs of reproduction, Trends
senescence, Evol. Ecol. Res., 8, 561-574.
Ecol. Evol., 15, 421-425.
28.
Galván, I., and Møller, A.P. (2018) Dispersal capacity
14.
Selman, C., Tuller, J.M.A., and Wieser, D.
(2009)
explains the evolution of lifespan variability, Ecol. Evol., 8,
Ribosomal protein S6 kinase 1 signaling regulates mam
4949-4957.
malian life span, Science, 326,140-144.
29.
Clark, W.R. (1998) Sex and the origins of death. Oxford
15.
Spitze, K. (1991) Chaoborus predation and life history evo
University Press, Oxford.
lution in Daphnia pulex: temporal pattern of population
30.
Mitteldorf, J., and Barja, G. (2019) The evolutionary ori
diversity, fitness, and mean life history, Evolution, 45, 82-92.
gin of cell senescence in ciliates. Forthcoming? (in press)
БИОХИМИЯ том 84 вып. 12 2019
1814
МИТТЕЛЬДОРФ
31.
Bell, G. (1982) The masterpiece of nature: the evolution and
43. Bateson, W., and Mendel, G. (1913) Mendel’s principles of
genetics of sexuality. University of California Press, Berkeley.
heredity. Cambridge University Press, Cambridge, UK.
32.
Smith, J.M. (1989) Evolutionary genetics. Oxford University
44. Lockwood, J.A., and Debrey, L.D. (1990) A solution for
Press, New York.
the sudden and unexplained extinction of the rocky moun
33.
Wagner, G.P., and Altenberg, L. (1996) Complex adaptations
tain grasshopper (Orthoptera: Acrididae), Environ.
and the evolution of evolvability, Evolution, 50, 967-976.
Entomol., 19, 1194-1205.
34.
Pigliucci, M. (2008) Is evolvability evolvable? Nat. Rev.
45. Stead, D.G. (1935) The rabbit in Australia. Winn, Sydney.
Genet., 9, 75.
46. Human, K.G., and Gordon, D.M. (1997) Effects of argen
35.
Kirschner, M., and Gerhart, J. (1998) Evolvability, Proc.
tine ats on invertebrate biodiversity in Northern California,
Natl. Acad. Sci. USA, 95, 8420-8427.
Conserv. Biol., 11, 1242-1248.
36.
Pepper, J.W. (2003) The evolution of evolvability in genet
47. Blagosklonny, M.V., and Hall, M.N. (2009) Growth and
ic linkage patterns, Biosystems, 69, 115-126.
aging: a common molecular mechanism, Aging (Albany
37.
Trubitsyn, A. (2006) Evolutionary mechanisms of species
NY), 1, 357-362.
specific lifespan, Adv. Gerontol., 19, 13-24.
48. Chu, M.C., Rath, K.M., Huie, J., and Taylor, H.S. (2003)
38.
May, R.M. (1974) Biological populations with nonoverlap
Elevated basal FSH in normal cycling women is associated
ping generations: stable points, stable cycles, and chaos,
with unfavourable lipid levels and increased cardiovascular
Science, 186, 645-647.
risk, Hum. Reprod., 18, 1570-1573.
39.
Mandelbrot, B.B. (1983) The fractal geometry of nature,
49. Sonntag, W.E., Lynch, C.,Thornton, P., Khan, A.,
W.H. Freeman & Co., New York, p. 173.
Bennett, S., and Ingram, R. (1999) Pleiotropic effects of
40.
Leroi, A., Chippindale, A.K., and Rose, M.R. (1994)
growth hormone and insulin like growth factor (IGF) 1 on
Long term evolution of a genetic life history trade off in
biological aging: inferences from moderate caloric restricted
Drosophila: the role of genotype by environment interac
animals, J. Gerontol. Ser. A: Biol. Sci. Med. Sci., 54, 521-538.
tion, Evolution, 48, 1244-1257.
50. Franceschi, C., and Campisi, J. (2014) Chronic inflamma
41.
Turcotte, M.M., Reznick, D.N., and Hare, J.D. (2011)
tion (inflammaging) and its potential contribution to age
The impact of rapid evolution on population dynamics in
associated disease, J. Gerontol. Ser. A: Biol. Sci. Med. Sci.,
the wild: experimental test of eco evolutionary dynamics,
69, 4-9.
Ecol. Lett., 14, 1084-1092.
51. Lu, A.T., Xue, L., Salfati, E.L., Chen, B.H., Ferrucci, L.,
42.
Carroll, S.B. (2005) Endless forms most beautiful: the new
Levy, D., McRae, A.F., Marione, R.E., and Visscher, P.M.
science of evo devo and the making of the animal kingdom.
(2018) GWAS of epigenetic aging rates in blood reveals a
W.W. Norton & Company.
critical role for tert, Nat. Commun., 9, 387.
WHAT IS ANTAGONISTIC PLEIOTROPY?
Review
J. Mitteldorf1,2
1 Washington University School of Medicine, St. Louis, MO, USA; E mail: aging.advice@gmail.com
2 National Institute for Biological Sciences, Beijing, China
Received July 5, 2019
Revised September 19, 2019
Accepted September 19, 2019
Biological aging has been observed to be genetically associated with enhancement of fertility or survivability early in
life. This has been interpreted as corroboration for the prevailing theory for evolution of aging, called Antagonistic
Pleiotropy (AP). The present article offers an alternative explanation for observed pleiotropy, in which pleiotropy is
an adaptation in its own right, rather than a physical precondition of biological evolution. The AP evolutionary the
ory was first proposed 60 years ago by George Williams. Its premise is that the same genes that offer individual bene
fits early in life have unavoidable pleiotropic effects that cause senescence later in life. A major strength of AP theory
is that it explains selection of a trait that is detrimental to individual fitness (aging) without reference to collective
benefit (group selection). A weakness of the theory is that the tradeoffs do not appear to be unavoidable; there are
known examples of mutations that lead to longer lifespan without apparent cost. The fact that wild types of these genes
cause senescence without offering concomitant individual benefit seems to be evidence for another selective mecha
nism, dependent on a fitness benefit of aging which can only be collective, not individual. Since AP theory’s early
acceptance, plausible mechanisms of multilevel selection have been proposed, in which aging emerges as a commu
nal adaptation. In the context of adaptive theories of aging, the question arises: why does antagonistic pleiotropy per
sist in nature if it is avoidable? I propose herein that natural selection may actually favor pleiotropy as an evolved adap
tation. This is because the combination of high fertility and long lifespan is a temptation for individuals, but a danger
for the stability of populations. Predator populations that grow faster than their prey can recover risk chaotic popula
tion dynamics, leading to extinction. Once a sustainable mix of fertility and longevity has been established by multi
level natural selection, pleiotropy can help to assure that it is not lost. The population is free to shift from (high fertil
ity/short lifespan) to (lower fertility/longer lifespan) as varying environmental conditions demand, without risking
population overshoot and collapse. I describe herein experiments with an individual based computer simulation in
which pleiotropy evolves as a group selected adaptation under a range of assumptions and in a broad swath of para
meter space.
Keywords: aging, senescence, programmed aging, antagonistic pleiotropy, group selection, multilevel selection
БИОХИМИЯ том 84 вып. 12 2019