БИОХИМИЯ, 2019, том 84, вып. 12, с. 1832 - 1838
УДК 577.24
СРАВНЕНИЕ НЕОТЕНИЧЕСКИХ ПРИЗНАКОВ
Heterocephalus glaber И Homo sapiens
Мини обзор
© 2019
Н.А. Попов, В.П. Скулачев*
НИИ физико химической биологии им. А.Н. Белозерского,
Московский государственный университет им. М.В. Ломоносова,
119991 Москва, Россия; электронная почта: skulach@belozersky.msu.ru
Поступила в редакцию 31.07.2019
После доработки 24.09.2019
Принята к публикации 24.09.2019
Суммированы данные, указывающие на неотению (продление молодости) у голого землекопа и человека.
В частности, у землекопа в настоящее время уже обнаружено 58 признаков неотении на уровне организма,
органов, тканей, клеток и метаболизма. Среди них можно найти признаки, продлевающие жизнь млекопи"
тающего. Такова, например, мягкая деполяризация митохондрий, предотвращающая генерацию этими ор"
ганеллами активных форм кислорода - мощного стимулятора старения. Деполяризация исчезает с возрас"
том у короткоживущих млекопитающих (мыши, Mus musculus) гораздо быстрее, чем у долгоживущих млеко"
питающих (голого землекопа и летучей мыши). Развитию неотеничной физиологии голых землекопов по"
служило их социальное устройство. Они живут в подземных колониях, где к размножению допускается
лишь царица и один или несколько самцов, защищенных многочисленными подчиненными от внешних уг"
роз и регулярно снабжаемых питанием. Человек, в свою очередь, добился постепенного увеличения про"
должительности жизни за счет неотении, а затем - технического прогресса, что прослеживается при срав"
нении кривых продолжительности жизни шимпанзе, охотников"собирателей - парагвайских аборигенов
племени Ache и жителей Швеции от XVII в. до наших дней.
КЛЮЧЕВЫЕ СЛОВА: старение, неотения, голый землекоп, человек, долгожительство, митохондрии.
DOI: 10.1134/S0320972519120078
В 2015 г. наша группа в Москве совместно с
явления признаков в онтогенезе неотеническо"
группой Т. Хильдебрандта в Берлине [1-3] и не"
го существа. Это связано с тем, что эволюция
зависимая объединенная группа Т. Парка в Чи"
вида может определяться не только, например,
каго, Стокгольме и Вене [4] обратили внимание
замедлением онтогенеза, но и какими"то совсем
на целый ряд признаков неотении (т.е. продле"
другими факторами, требующими не замедле"
ния жизни за счет замедления онтогенетических
ния, а наоборот, ускорения индивидуального
программ) грызунов (голых землекопов Hetero
развития. Именно очевидная мозаичность [6]
cephalus glaber, живущих, по крайней мере, в де"
затруднила принятие гипотезы Л. Болка о нео"
сять раз дольше мышей или крыс (см. также [5]).
тении H. sapiens [7, 8].
Особенно многочисленным оказался набор та"
Для выяснения подобных обстоятельств су"
ких признаков в статье из Москвы и Берлина,
щественными факторами являются: величина
опубликованной в Physiol. Rev. [3]. В ней был
разнообразия неотенических признаков и воз"
приведен список сорока трех явлений онтогене"
можная роль в контроле продолжительности
за, которые у землекопа либо не наблюдаются,
жизни представителей данного вида. В настоя"
либо возникают при индивидуальном развитии
щей статье дополнен список признаков неоте"
во много раз позже, чем у мышей, а также других
нии голого землекопа, описанных за последние
представителей семейства Bathyergidae, от кото"
два с половиной года, минувших после нашей
рого более 30 млн лет назад отделился голый
публикации 2017 г. в Physiol. Rev. [3]. Теперь этот
землекоп.
список вырос с 43 до 58 наименований и, что бо"
Отнесение неотении к причинам долголетия
лее важно, пополнился явлениями, определяю"
организма часто осложняется мозаичностью по" щими продолжительность жизни.
В 2018-2019 гг. в нашей группе М.Ю. Высо"
* Адресат для корреспонденции.
ких с сотр. был обнаружен феномен мягкой де"
1832
НЕОТЕНИЯ, ТЕХНИЧЕСКИЙ ПРОГРЕСС И ДОЛГОЖИТЕЛЬСТВО
1833
поляризации митохондрий, присущий почти
всем тканям млекопитающего. Этот регулятор"
ный механизм служит основным в системе пред"
отвращения генерации митохондриями актив"
ных форм кислорода (АФК). К концу жизни он
исчезает у обычных видов млекопитающих типа
мыши Mus musculus, но сохраняется десятилети"
ями у долгожителей - голого землекопа и лету"
чей мыши [9]. Наши результаты подтверждают
гипотезу о роли митохондриальных АФК как
ядов, которые образуются самим организмом и
постепенно отравляют его функционирование,
что, собственно, и служит механизмом старе"
ния [10].
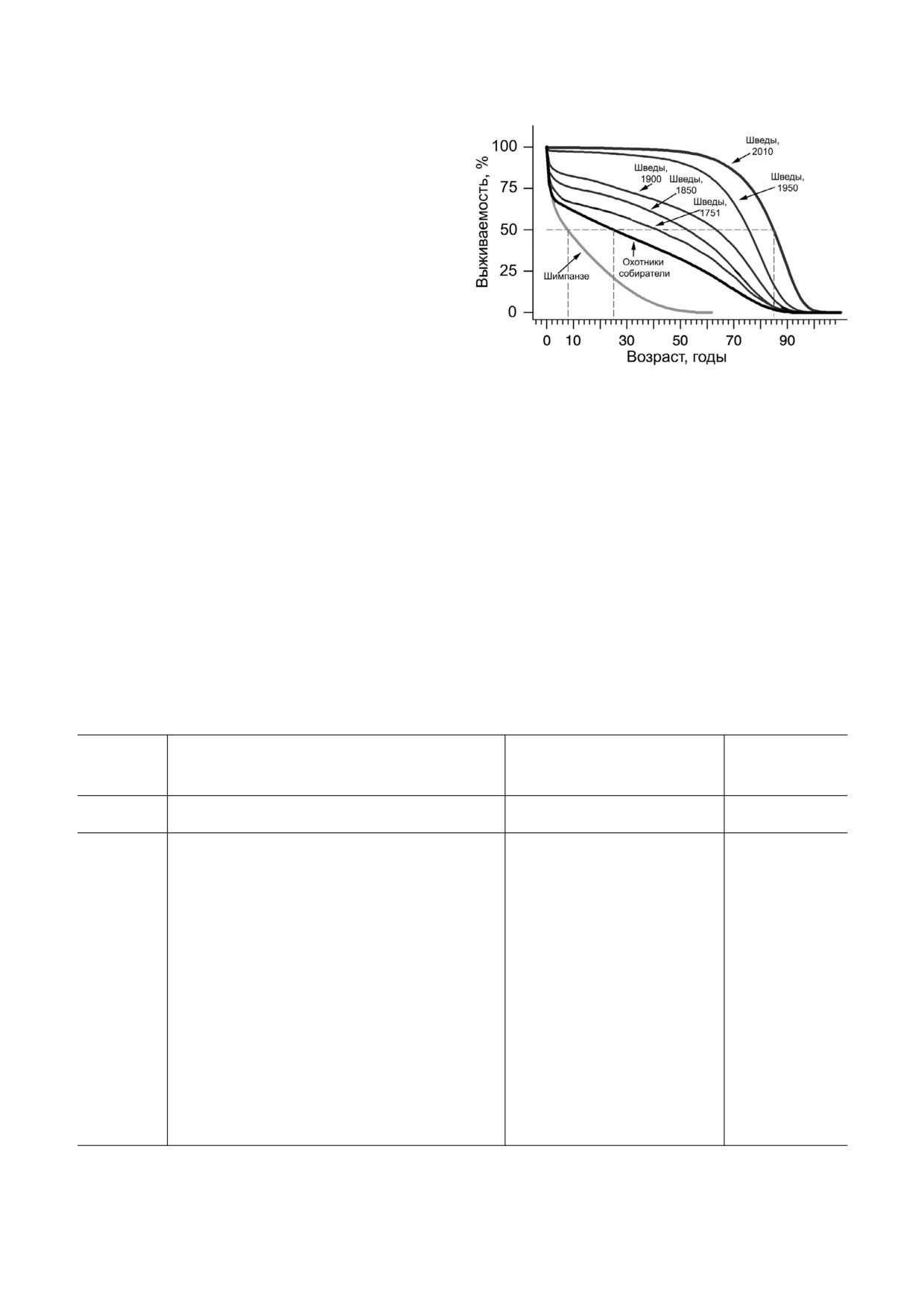
Большой интерес представляет сравнение
Возрастная зависимость выживания шведов (в 1751, 1850,
роли неотении в долгожительстве голых земле"
1900, 1950 и 2010 гг.), а также шимпанзе и охотников"соби"
копов с таковым у H. sapiens, который также от"
рателей, парагвайских дикарей племени Ache [11, 12]
носится к социальным организмам. По данным
Burger et al. [11, 12], приведенным на рисунке,
выживаемость современных цивилизованных
шимпанзе, развитием интеллекта. Что касается
людей близка к 100% вплоть до возраста ~ 60 лет,
дальнейшего уменьшения смертности человека,
после чего наступает все ускоряющийся спад,
оно обусловлено техническим прогрессом, темп
достигающий у шведов 50% ежегодной смерт"
которого несравнимо выше, чем скорость био"
ности к 90 годам. Парагвайские дикари имеют
логической эволюции.
50% выживаемость в возрасте 28 лет, а шимпан"
В таблице приведены списки признаков нео"
зе - в возрасте 9 лет. Трехкратное уменьшение
тении у голого землекопа и человека. Эти приз"
смертности дикарей по сравнению с шимпанзе,
наки наиболее заметны на организменном уров"
по"видимому, объясняется в основном коллек"
не организации. Но не только они отличают го"
тивной деятельностью по защите племени, ис"
лого землекопа и человека от других близких им
пользованием простейших орудий труда, строи"
видов, не использующих неотению (мышей и
тельством укрытий и высоким, в сравнении с
шимпанзе соответственно).
Неотеничные черты голого землекопа и человека на различных уровнях организации
Уровни
Общие признаки
организации
Голый землекоп
Человек
человека и голого
землекопа
1
2
3
4
Организмен"
гораздо меньшая масса тела по сравнению
«плоское лицо», ортогнатия [8];
отсутствие или
ный
с 20 родственными видами - членами семейства
уменьшение или отсутствие во"
уменьшение ко"
Bathyergidae [3, 13];
лос на теле [8];
личества волос;
отсутствие волос (уникально для взрослых грызу"
центральное положение заты"
длительный срок
нов) [3, 14, 26];
лочного отверстия (у большин"
жизни;
гораздо более длительный период беременности,
ства других приматов оно миг"
длительный
чем ожидается от животных с сопоставимой мас"
рирует назад во время онтоге"
период роста;
сой тела [13, 15-17];
неза) [8];
длительный срок
гораздо более длительный период роста и созрева"
сохранение черепных структур
беременности;
ния новорожденных, по сравнению с другими гры"
до преклонного возраста [8];
длительный
зунами [3, 17];
структура рук и ног [8];
период младенче"
увеличение репродуктивной функции с возрастом
форма таза [8];
ства;
(а не ее угнетение, наблюдаемое у других млекопи"
вентрально направленная пози"
длительный пе"
тающих) [3, 18];
ция полового канала у женщин
риод полового
ограниченная способность поддерживать постоян"
[8];
созревания.
ную температуру тела, как у новорожденных жи"
определенные вариации строе"
вотных [3, 19-21];
ния зубов и черепных швов [8];
возрастные заболевания, такие как рак, диабет,
отсутствие гребня черепа [8];
сердечно"сосудистые заболевания, патологии
отсутствие гребня бровей [8];
5 БИОХИМИЯ том 84 вып. 12 2019
1834
ПОПОВ, СКУЛАЧЕВ
Продолжение таблицы
1
2
3
4
головного мозга, печени и некоторые заболевания
тонкость костей черепа [8];
почек, а также многие инфекции очень редки или
положение глазниц под череп"
отсутствуют как причины смерти голых землекопов
ной полостью [8];
(как в случае с молодыми млекопитающими) [3, 22,
брахицефалия [8];
23];
мелкие зубы [8];
задержка в развитии скелетной системы [3, 24];
позднее прорезывание зубов [8];
высокая максимальная продолжительность жизни;
невозможность полного враще"
по сравнению с другими грызунами очень низкий
ния большого пальца [8];
уровень смертности, который увеличивается незна"
длительный период инфан"
чительно, по крайней мере, в течение первых 24
тильной зависимости [8];
лет [3, 19, 23];
длительный период роста [8];
отсутствие связанных с возрастом снижений ког"
высокая продолжительность
нитивных функций [3, 23].
жизни [8];
короткие конечности по срав"
нению с размером тела [8];
ноги длиннее рук [3];
вертикальное положение тела [3];
более длительное время бере"
менности по сравнению с
шимпанзе [3].
Тканевый
отсутствие у взрослых особей ушных раковин, как
форма наружного уха [8];
очень недораз"
у новорожденных животных [3, 26];
эпикантическая («монгольс"
витый вомеро"
отсутствие у взрослых особей мошонки, как у но"
кая») складка в углах глаз [8];
назальный ор"
ворожденных [3, 16, 27];
относительно большая масса
ган, как у ново"
распределение кальбиндина по неонатальному ти"
мозга [8];
рожденных [3,
пу в области ca3 гиппокампа [3, 28];
половые губы у женщин [8];
23, 49];
низкая чувствительность к экзогенному аденозину
отсутствие бакулюма (кости
повышенная си"
в гиппокампе [3, 28, 29];
пениса) [3];
наптическая ак"
очень длительный период развития мозга по срав"
небольшой нос [3];
тивность и плас"
нению с другими грызунами [4];
люди сохраняют более прими"
тичность;
недоразвитие многочисленных морфологических
тивную форму скелета и мышц,
неполная мие"
признаков в легких [3, 30, 31];
тогда как у обезьян развивалась
линизация нейт"
мозг новорожденных голых землекопов в два раза
более сложная и специализи"
ральных воло"
больше мозга новорожденных мышей. При рожде"
рованная анатомия этих орга"
кон в течение
нии масса мозга мыши составляет ~ 17% массы
нов [3];
нескольких лет
мозга взрослой мыши, но уже через 2 недели жизни
мозг человека намного больше,
после рождения;
достигает 90% массы мозга взрослого животного,
чем у шимпанзе [3];
взрывное фор"
тогда как мозг голых землекопов составляет при
скорость роста человеческого
мирование си"
рождении ~ 41% размера взрослого органа и дости"
мозга увеличивается до самого
напсов («избы"
гает 90% массы только к 3"месячному возрасту [5];
рождения, тогда как у шимпан"
точный синап"
голые землекопы рождаются с четко определенным
зе данный параметр начинает
тогенез») на
молекулярным слоем зубчатой извилины в мозге и
уменьшаться задолго до этого
ранних стадиях
со значительно меньшим количеством клеток,
[3];
развития голов"
имеющих фактор транскрипции Sox2, тогда как
нейроны у взрослых людей
ного мозга.
почти все клетки зубчатой извилины в мозге ново"
сохраняют определенные «не"
рожденных мышей являются Sox2"положительны"
совершеннолетние» характе"
ми. Оба вида рождаются с сопоставимыми уровня"
ристики, такие как повышен"
ми клеток, экспрессирующих Sox2 в желудочковой
ная синаптическая активность
и субвентрикулярной зонах, но этот уровень резко
и пластичность, а также непол"
падает у мышей в течение первой постнатальной
ная миелинизация в течение
недели, в то время как у голых землекопов он дос"
первых двух десятилетий жиз"
товерно не изменяется до 3"х лет [5];
ни [48];
очень высокая устойчивость нейронов головного
веретенообразные нейроны
мозга к аноксии и последующему окислительному
намного крупнее и многочис"
стрессу при реоксигенации (как у новорожденных
леннее у людей, чем у челове"
животных) [3, 29, 32];
кообразных обезьян [48].
отсутствие релаксации синаптических парных им"
пульсов в отличие от других взрослых млекопитаю"
щих [3, 28, 32];
поддержание способности к регенерации и удлине"
нию нейронов в зрелом возрасте [3, 23, 28, 33];
БИОХИМИЯ том 84 вып. 12 2019
НЕОТЕНИЯ, ТЕХНИЧЕСКИЙ ПРОГРЕСС И ДОЛГОЖИТЕЛЬСТВО
1835
Продолжение таблицы
1
2
3
4
легочные нейроэпителиальные тельца, число кото"
рых обычно уменьшается в течение первой постна"
тальной недели, представлены в больших количе"
ствах у голых землекопов в возрасте более двух не"
дель [3, 34];
отсутствие снижения эластичности кровеносных
сосудов с возрастом [3, 18, 25];
отсутствие снижения площади коры головного
мозга с возрастом [3, 18];
отсутствие возрастного снижения минеральной
плотности костей [3, 18, 25];
отсутствие какого"либо ухудшения состояния сус"
тавных хрящей с возрастом [3, 18];
чувствительность гладкой мускулатуры голых зем"
лекопов к NO не уменьшается с возрастом в отли"
чие от изменений, наблюдаемых у других грызунов
[3, 35, 36];
у мышей корковый слой 1 не становится четко оп"
ределенным до 2"недельного возраста и теряет
экспрессию DCX к 3 неделям. Голые землекопы
рождаются с более организованным расположени"
ем коры и более развитой корой головного мозга,
тем не менее период созревания их коры дольше,
чем у мышей [5];
созревание нейронов в гиппокампе у мышей дос"
тигает своего максимального значения к 6 меся"
цам, тогда как у голых землекопов этот уровень
медленно увеличивается в течение первого десяти"
летия жизни [3, 4].
Клеточный
отсутствие перегрузки Ca2+ при длительном воз"
оба вида имеют
и молеку"
буждении нейронов [3, 32, 37];
наибольшее сре"
лярный
сильная задержка развития единой митохондриаль"
ди млекопитаю"
ной системы в скелетных мышцах в постнатальном
щих количество
периоде [3, 30];
мотивов PXXP в
увеличение, а не уменьшение количества митохон"
богатом проли"
дрий и их активности с возрастом [2, 3, 23, 38];
ном домене p53
отсутствие у землекопов, в отличие от мышей, воз"
(5 для человека
растного снижения коэффициента дыхательного
и 4 для голых
контроля митохондрий [9];
землекопов);
отсутствие возрастного общего снижения метабо"
у людей и голых
лизма [3, 18, 25];
землекопов уро"
отсутствие какого"либо увеличения индекса пере"
вень тирозин"
кисного окисления липидов и их окислительного
гидроксилазы
повреждения с возрастом, которое наблюдается у
возрастает до
других млекопитающих при старении [3, 39];
2"3"летнего воз"
нет увеличения продукции активных форм кисло"
раста);
рода (АФК) с возрастом в отличие от других млеко"
стойкая
питающих [3, 9, 23];
экспрессия 3R
состав аксонов голых землекопов более сходен с
тау"белка.
человеческим, в результате чего голые землекопы
сохраняют экспрессию 3R тау даже после заверше"
ния роста мозга, в то время как у мышей наблюда"
ется резкое снижение 3R тау уже на 8 день после
рождения [5];
выключение мягкой деполяризации митохондрий у
голых землекопов не происходит вплоть до 30"лет"
него возраста, в то время как у мышей этот меха"
низм защиты от митохондриальных АФК прекра"
щает работать уже через 1 год после рождения [9];
исчезновение мягкой деполяризации митохондрий
печени: у мышей " сразу после родов, у голых
БИОХИМИЯ том 84 вып. 12 2019
5*
1836
ПОПОВ, СКУЛАЧЕВ
Окончание таблицы
1
2
3
4
землекопов он сохраняется, по крайней мере до 10 лет [9];
инактивация гена, кодирующего FAS"активируемую серин/ треонин"
киназу (FASTK), антиапоптотический и провоспалительный фермент
[3, 15];
отсутствие снижения уровней супероксиддисмутаз (Sod) 1 и 2 и ката"
лазы с возрастом [3, 9, 40, 41];
снижение уровня адениннуклеотидов в митохондриях взрослых голых
землекопов и более высокий уровень дыхания после истощения до"
бавленного ADP (как у эмбрионов крыс, мышей и у новорожденных)
[3];
необычно высокая протеасомная активность у взрослых животных [3,
14, 23, 42, 43];
изменения в последовательности В"цепи инсулина, приводящие к
снижению гормональной активности до эмбрионального уровня [3,
15, 44];
отсутствие снижения уровня IGF2 у взрослых голых землекопов [3,
15];
снижение экспрессии IGF"1, инсулин"индуцированного гена 2
(INSIG2) и увеличение экспрессии генов IGF1R и резистина (RETN) у
взрослых голых землекопов [3, 15];
отсутствие способности ощущать боль, вызванную капсаицином или
кислотой из"за недостатка компонента Р [3, 21];
постоянное присутствие субъединицы CluN2D рецептора NMDA
(глутамата) во взрослом возрасте, которое присуще другим млекопи"
тающим только у новорожденных [9, 45];
отсутствие снижения толерантности к глюкозе у старых животных [3,
18, 20];
отсутствие повышения при старении уровня гликированного гемо"
глобина [3, 18, 46];
постоянное присутствие высокомолекулярного гиалуронана в меж"
клеточных пространствах [3, 47];
у мышей уровень нейрофиламентов легкой цепи (NF"L) быстро уве"
личивается после рождения и достигает уровня взрослого в течение
первой постнатальной недели, в то время как у голых землекопов уве"
личение NF"L на 33% происходит между рождением и 2-4 неделями,
а к 4 месяцам NF"L на 168% больше, чем у новорожденных [5];
экспрессия DCX (ассоциированного с микротрубочками белка -
двойного кортина) у мышей выше, чем у голых землекопов, но быст"
ро снижается как в зубчатой извилине, так и в субвентикулярной зо"
не, тогда как у голых землекопов наблюдается постепенное снижение
экспрессии [5];
уровень изоформы Map2D (белок, ассоциированный с микротрубоч"
ками) у голых землекопов остается стабильным до 6 месяца жизни, в
то время как у крыс экспрессия этого белка полностью падает к 3 не"
деле жизни [5];
уровень синаптофизина у голых землекопов резко возрастает в пери"
од с 2-4 месяцев до 9 месяцев после рождения, в то время как у мы"
шей эти события происходят в первые постнатальные дни 10-35 [5];
экспрессия тирозингидроксилазы в мозге голых землекопов прогрес"
сирующе увеличивается между рождением и 3"летним возрастом, тог"
да как у крыс эти события происходят более быстро - за 4-5 недель
развития [5];
голый землекоп, в отличие от мыши, экспрессирует связанный с мие"
лином гликопротеин (MAG) при рождении [5];
появление количественного доминирования гексокиназы II над более
активной гексокиназой I в митохондриях мышц: у мышей до 3 меся"
цев, у голых землекопов - через 11 лет [9].
Конфликт интересов. Авторы заявляют об от"
Соблюдение этических норм. В данном обзоре
сутствии между ними конфликта интересов.
отсутствуют эксперименты, где в качестве моде"
лей использованы люди или животные.
БИОХИМИЯ том 84 вып. 12 2019
НЕОТЕНИЯ, ТЕХНИЧЕСКИЙ ПРОГРЕСС И ДОЛГОЖИТЕЛЬСТВО
1837
СПИСОК ЛИТЕРАТУРЫ
1.
Skulachev, V.P. (2015) Moscow news: two more representa
17.
Sherman, P.W., Jarvis, J.U.M., and Alexander, R.D. (1991)
tives of sodium motive force generators (Na+ cbb3 oxidase
The Biology of the naked mole rat, Princeton University
and Na+ bacteriorhodopsin); natural delay of the aging pro
Press, Princeton, N.J.
gram (neoteny) in mammals, namely in naked mole rat and
18.
Buffenstein, R. (2008) Negligible senescence in the longest
«naked ape» (human). Conf. MIP 2015, Lučn Bouda, Pec
living rodent, the naked mole"rat: insights from a success"
pod Snezkou, Czech Republic.
fully aging species, J.Comp. Phys. B Bioch. Syst. Environ.
2.
Holtze, S., Eldarov, C.M., Vays, V.B., Vangeli, I.M.,
Phys., 178, 439-445, doi: 10.1007/s00360"007"0237"5.
Vysokikh, M.Y., Bakeeva, L.E., Skulachev, V.P., and
19.
Buffenstein, R. (2005) The naked mole"rat? A new long"
Hildebrandt, T.B. (2016) Study of age"dependent structural and
living model for human aging research, J. Gerontol. A Biol.
functional changes of mitochondria in skeletal muscles and
Sci. Med. Sci., 60, 1369-1377.
heart of naked mole rats (Heterocephalus glaber), Biochemistry
20.
Buffenstein, R., Kang, J., and Biney, A. (2007) Glucose
(Moscow), 81, 1429-1437, doi: 10.1134/S000629791612004X.
tolerance and insulin sensitivity in an extremely long"living
3.
Skulachev, V.P., Holtze, S., Vyssokikh, M.Y., Bakeeva, L.E.,
rodent, the naked mole"rat., FASEB J., 21, A1423-A1423.
Skulachev, M.V., Markov, A.V., Hildebrandt, T.B., and
21.
Park, T.J., Comer, C., Carol, A., Lu, Y., Hong, H.S., and
Sadovnichii, V.A. (2017) Neoteny, prolongation of youth:
Rice, F.L. (2003) Somatosensory organization and behav"
from naked mole rats to «naked apes» (Humans), Physiol.
ior in naked mole"rats: II. Peripheral structures, innerva"
Rev., 97, 699-720, doi: 10.1152/physrev.00040.2015.
tion, and selective lack of neuropeptides associated with
4.
Penz, O.K., Fuzik, J., Kurek, A.B., Romanov, R., Larson, J.,
thermoregulation and pain, J. Comp. Neurol., 465,
Park, T.J., Harkany, T., and Keimpema, E.
(2015)
104-120, doi: 10.1002/cne.10824.
Protracted brain development in a rodent model of extreme
22.
Delaney, M.A., Nagy, L., Kinsel, M.J., and Treuting, P.M.
longevity, Sci. Rep., 5, 11592, doi: 10.1038/srep11592.
(2013) Spontaneous histologic lesions of the adult naked
5.
Orr, M.E., Garbarino, V.R., Salinas, A., and Buffenstein, R.
mole rat (Heterocephalus glaber): a retrospective survey of
(2016) Extended postnatal brain development in the
lesions in a zoo population, Vet. Pathol., 50, 607-621,
longest"lived rodent: prolonged maintenance of neotenous
doi: 10.1177/0300985812471543.
traits in the naked mole"rat brain, Front Neurosci., 10, 504,
23.
Edrey, Y.H., Hanes, M., Pinto, M., Mele, J., and
doi: 10.3389/fnins.2016.00504.
Buffenstein, R. (2011) Successful aging and sustained good
6.
Bufill, E., Agusti, J., and Blesa, R. (2011) Human neoteny
health in the naked mole rat: a long"lived mammalian
revisited: the case of synaptic plasticity, Am. J. Hum. Biol.,
model for biogerontology and biomedical research, ILAR
23, 729-739, doi: 10.1002/ajhb.21225.
J., 52, 41-53.
7.
Bolk, L. (1926) The problem of human development, Gustav
24.
Henry, E.C., Dengler"Crish, C.M., and Catania, K. C.
Fischer, Jena.
(2007) Growing out of a caste - reproduction and the
8.
Bolk, L. (1927) On the origin of human races, Proc.
making of the queen mole"rat, J. Exper.Biol., 210,
Koninklijke Akadem Wetenschappen Te Amsterdam, 30, 320-328.
261-268, doi: 10.1242/jeb.02631.
9.
Vyssokikh, M.Y., Holtze, S., Averina, O.A., Lyamzaev, K.G.,
25.
O’Connor, T.P., Lee, A., Jarvis, J.U.M., and Buffenstein,
Severin, F.F., Skulachev, M.V., Hildebrandt, T.B., and
R. (2002) Prolonged longevity in naked mole"rats: age"
Skulachev, V.P. (2019) Mild depolarization of the inner
related changes in metabolism, body composition and gas"
mitochondrial membrane, crucial component of anti"
trointestinal function, Comp. Biochem. Physiol. A Mol.
aging program, Proc. Natl. Acad. Sci. USA, in press.
Integr. Physiol., 133, 835-842, doi: 10.1016/S1095"
10.
Skulachev, V.P. (2012) What is «phenoptosis» and how to
6433(02)00198"8.
fight it? Biochemistry (Mosc.) Phenoptosis, 77, 689-706,
26.
Rüppell, E. (1845) Heterocephalus glaber (Rüppell). in
doi: 10.1134/S0006297912070012.
Abliantllungren aus dem Gebiete der beschreibenden
11.
Burger, O., Baudisch, A., and Vaupel, J.W. (2012) Human mor"
Naturgeschichte, pp. 99-101.
tality improvement in evolutionary context, Proc. Natl. Acad.
27.
Klonisch, T., Fowler, P.A., and Hombach"Klonisch, S.
Sci. USA, 109, 18210-18214, doi: 10.1073/pnas.1215627109.
(2004) Molecular and genetic regulation of testis descent
12.
Burger, O. (2017) Evolutionary demography of the human
and external genitalia development, Dev. Biol., 270, 1-18,
mortality profile in The Evolution of Senescence in the Tree of
doi: 10.1016/j.ydbio.2004.02.018.
Life, Cambridge Univ Press, Cambridge.
28.
Amrein, I., Becker, A.S., Engler, S., Huang, S.H., Muller, J.,
13.
Mcnab, B.K. (1979) Influence of body size on the energet"
Slomianka, L., and Oosthuizen, M.K. (2014) Adult neuro"
ics and distribution of fossorial and burrowing mammals,
genesis and its anatomical context in the hippocampus of
Ecology, 60, 1010-1021, doi: 10.2307/1936869.
three mole"rat species, Front. Neuroanat.,
8,
39,
14.
Rodriguez, K.A., Edrey, Y.H., Osmulski, P., Gaczynska, M.,
doi: 10.3389/fnana.2014.00039.
and Buffenstein, R. (2012) Altered composition of liver
29.
Larson, J., Drew, K.L., Folkow, L.P., Milton, S.L., and
proteasome assemblies contributes to enhanced proteasome
Park, T.J. (2014) No oxygen? No problem! Intrinsic brain
activity in the exceptionally long"lived naked mole"rat,
tolerance to hypoxia in vertebrates, J. Exp. Biol., 217,
PLoS One, 7, e35890, doi: 10.1371/journal.pone.0035890.
1024-1039, doi: 10.1242/Jeb.085381.
15.
Fand, X., Seim, I., Huang, Z., Gerashchenko, M.V.,
30.
Maina, J.N., Gebreegziabher, Y., Woodley, R., and
Xiong, Z., Turanov, A.A., Zhu, Y., Lobanov, A.V., Fan, D.,
Buffenstein, R. (2001) Effects of change in environmental
Yim, S.H., Yao, X., Ma, S., Yang, L., Lee, S.G., Kim, E.B.,
temperature and natural shifts in carbon dioxide and oxy"
Bronson, R.T., Sumbera, R., Buffenstein, R., Zhou, X.,
gen concentrations on the lungs of captive naked mole"rats
Krogh, A., Park, T.J., Zhang, G., Wang, J., and Gladyshev, V.N.
(Heterocephalus glaber): a morphological and morphomet"
(2014) Adaptations to a subterranean environment and
ric study, J. Zool.,
253,
371-382, doi:
10.1017/
longevity revealed by the analysis of mole rat genomes, Cell
S0952836901000346.
Rep., 8, 1354-1364, doi: 10.1016/j.celrep.2014.07.030.
31.
Maina, J.N., Maloiy, G.M.O., and Makanya, A.N. (1992)
16.
Jarvis, J.U.M. (1991) Reproduction of naked mole rats. in
Morphology and morphometry of the lungs of 2 East"
The Biology of the Naked Mole Rat: Monographs in Behavior
African mole rats, tachyoryctes"splendens and
and Ecology (Sherman, P.W., Jarvis, J.U.M., and Alexander,
Heterocephalus"Glaber (Mammalia, rodentia), Zoomor
R.D. eds.), Princeton Univ. Press, Oxford. pp. 384-425.
phology, 112, 167-179, doi: 10.1007/Bf01633107.
БИОХИМИЯ том 84 вып. 12 2019
1838
ПОПОВ, СКУЛАЧЕВ
32.
Larson, J., and Park, T.J. (2009) Extreme hypoxia tole"
longevity differences among the longest"living rodents and
rance of naked mole"rat brain, Neuroreport,
20,
similarly"sized mice, J. Gerontol. Series A Biol. Sci. Med.
1634-1637, doi: 10.1097/Wnr.0b013e32833370cf.
Sci., 61, 1009-1018, doi: 10.1093/gerona/61.10.1009.
33.
Park, K.K., Luo, X., Mooney, S.J., Yungher, B.J., Belin, S.,
42.
Rodriguez, K.A., Osmulski, P.A., Pierce, A., Weintraub, S.T.,
Wang, C., Holmes, M.M., and He, Z. (2017) Retinal gan"
Gaczynska, M., and Buffenstein, R. (2014) A cytosolic
glion cell survival and axon regeneration after optic nerve
protein factor from the naked mole"rat activates protea"
injury in naked mole"rats, J. Comp. Neurol., 525, 380-388,
somes of other species and protects these from inhibition,
doi: 10.1002/cne.24070.
Biochim. Biophys. Acta, 1842, 2060-2072, doi: 10.1016/
34.
Pan, J., Park, T.J., Cutz, E., and Yeger, H.
(2014)
j.bbadis.2014.07.005.
Immunohistochemical characterization of the chemosen"
43.
Triplett, J.C., Tramutola, A., Swomley, A., Kirk, J.,
sory pulmonary neuroepithelial bodies in the naked mole"
Grimes, K., Lewis, K., Orr, M., Rodriguez, K., Cai, J.,
rat reveals a unique adaptive phenotype, PLoS One, 9,
Klein, J.B., Perluigi, M., Buffenstein, R., and Butterfield, D.A.
e112623, doi: 10.1371/journal.pone.0112623.
(2015) Age"related changes in the proteostasis network in
35.
Csiszar, A., Ahmad, M., Smith, K.E., Labinskyy, N., Gao, O.,
the brain of the naked mole"rat: Implications promoting
Kaley, G., Edwards, J.G., Wolin, M.S., and Ungvari, Z.
healthy longevity, Biochim. Biophys. Acta,
1852,
(2006) Bone morphogenetic protein"2 induces proinflam"
2213-2224, doi: 10.1016/j.bbadis.2015.08.002.
matory endothelial phenotype, Amer.J. Pathol., 168,
44.
Perez, V.I., Buffenstein, R., Masamsetti, V., Leonard, S.,
629-638, doi: 10.2353/ajpath.2006.050284.
Salmon, A.B., Mele, J., Andziak, B., Yang, T., Edrey, Y.,
36.
Csiszar, A., Pacher, P., Kaley, G., and Ungvari, Z. (2005)
Friguet, B., Ward, W., Richardson, A., and Chaudhuri, A.
Role of oxidative and nitrosative stress, longevity genes and
(2009) Protein stability and resistance to oxidative stress
poly(ADP"ribose) polymerase in cardiovascular dysfunction
are determinants of longevity in the longest"living rodent,
associated with aging, Curr. Vasc. Pharmacol., 3, 285-291.
the naked mole"rat, Proc. Natl. Acad. Sci. USA, 106,
37.
Peterson, B.L., Park, T.J., and Larson, J. (2012) Adult
3059-3064, doi: 10.1073/pnas.0809620106.
naked mole"rat brain retains the NMDA receptor subunit
45.
Peterson, B.L., Larson, J., Buffenstein, R., Park, T.J., and
GluN2D associated with hypoxia tolerance in neonatal
Fall, C.P. (2012) Blunted neuronal calcium response to
mammals, Neurosci. Lett., 506, 342-345, doi: 10.1016/
hypoxia in naked mole"rat hippocampus, PLoS One, 7,
j.neulet.2011.11.042.
e31568, doi: 10.1371/journal.pone.0031568.
38.
Csiszar, A., Labinskyy, N., Orosz, Z., Zhao, X.M.,
46.
Yang, T., Buffenstein, R., and O’Connor, T.P.
(2002)
Buffenstein, R., and Ungvari, Z. (2007) Vascular aging in
Disparate age effects on gastrointestinal enzymes in naked
the longest"living rodent, the naked mole rat, Am. J.
mole rats, Int. Comp. Biol., 42, 1340-1341.
Physiol. Heart Circ. Physiol., 293, 919-927, doi: 10.1152/
47.
Tian, X., Azpurua, J., Hine, C., Vaidya, A., Myakishev"
ajpheart.01287.2006.
Rempel, M., Ablaeva, J., Mao, Z.Y., Nevo, E., Gorbunova, V.,
39.
Andziak, B., and Buffenstein, R. (2006) Disparate patterns
and Seluanov, A. (2013) High"molecular"mass hyaluronan
of age"related changes in lipid peroxidation in long"lived
mediates the cancer resistance of the naked mole rat,
naked mole"rats and shorter"lived mice, Aging Cell, 5,
Nature, 499, 346-349, doi: 10.1038/Nature12234.
525-532, doi: 10.1111/j.1474"9726.2006.00246.x.
48.
Buffenstein, R., and Yahav, S. (1991) Is the naked mole"rat
40.
Andziak, B., O’Connor, T.P., and Buffenstein, R. (2005)
Heterocephalus glaber an endothermic yet poikilothermic
Antioxidants do not explain the disparate longevity
mammal? J. Therm. Biol., 16, 227-232, doi: 10.1016/
between mice and the longest"living rodent, the naked
0306"4565(91)90030"6.
mole"rat, Mech. Ageing Dev.,
126,
1206-1212,
49.
Smith, T.D., Bhatnagar, K.P., Dennis, J.C., Morrison, E.E.,
doi: 10.1016/j.mad.2005.06.009.
and Park, T.J. (2007) Growth"deficient vomeronasal organs
41.
Hulbert, A.J., Faulks, S.C., and Buffenstein, R. (2006)
in the naked mole"rat (Heterocephalus glaber), Brain Res.,
Oxidation"resistant membrane phospholipids can explain
1132, 78-83, doi: 10.1016/j.brainres.2006.11.021.
NEOTENIC TRAITS OF Heterocephalus glaber AND Homo sapiens
Mini review
N. A. Popov and V. P. Skulachev*
Belozersky Institute of Physico Chemical Biology, Lomonosov Moscow State University,
119991 Moscow, Russia; E mail: skulach@belozersky.msu.ru
Received July 31, 2019
Revised September 24, 2019
Accepted September 24, 2019
Some data indicating neoteny (prolongation of youth and retardation of aging) in naked mole rat (Heterocephalus
glaber) and Homo sapiens are summarized. In the case of naked mole rat, 58 neotenic traits are described when organ"
ismal, tissue, cellular and metabolic aspects are considered. Among them, one may find mild depolarization of mito"
chondria, which prevents mitochondrial generation of reactive oxygen species, potent stimulators of aging. Effect of
mild depolarization inhibits an aging program of naked mole rat and prolongs life of this rodent. Mild depolarization
disappears with age in short"lived mammals (mouse Mus musculus) much sooner than in long"lived mammals (naked
mole rat and bat). Development of neoteny in naked mole rats is stimulated by social organization of their colonies
where sexual reproduction is monopolized by a queen and one or a few males who are protected and served by numer"
ous subordinates. Humans also developed neoteny which now is substituted by technical progress. Quite different rates
of neoteny and technical progress allow to discriminate between contributions of these two longevity mechanisms.
Keywords: aging, neoteny, naked mole rats, humans, longevity, mitochondria
БИОХИМИЯ том 84 вып. 12 2019