БИОХИМИЯ, 2019, том 84, вып. 3, с. 436 - 447
УДК 577.2
ЭКСПРЕССИЯ И ФУНКЦИОНАЛЬНЫЕ СВОЙСТВА
NMDA И GABAA РЕЦЕПТОРОВ ПРИ ДИФФЕРЕНЦИРОВКЕ
ИНДУЦИРОВАННЫХ ПЛЮРИПОТЕНТНЫХ СТВОЛОВЫХ КЛЕТОК
ЧЕЛОВЕКА В ВЕНТРАЛЬНЫЕ МЕЗЕНЦЕФАЛИЧЕСКИЕ НЕЙРОНЫ
© 2019 С.А. Антонов1*, Е.В. Новосадова1, А.Г. Кобылянский1,
С.Н. Иллариошкин2, В.З. Тарантул1, И.А. Гривенников1
1 Институт молекулярной генетики РАН, 123182 Москва,
Россия; электронная почта: vamore@inbox.ru
2 Научный центр неврологии, 125367 Москва, Россия
Поступила в редакцию 29.10.2018
После доработки 29.11.2018
Принята к публикации 29.11.2018
Ионотропные глутаматные и ГАМК рецепторы регулируют процесс дифференцировки и определяют функ
циональные свойства зрелых нейронов. Как недостаточная, так и избыточная активность этих систем ней
ротрансмиссии ассоциирована с различными заболеваниями нервной системы. В современной литературе
имеются ограниченные сведения относительно профиля экспрессии данных рецепторов и механизмов их
регуляции при дифференцировке специализированных типов нейронов человека. В настоящей работе с ис
пользованием ПЦР в реальном времени и иммуноцитохимического окрашивания был изучен профиль
экспрессии NMDA и GABAA рецепторов при дифференцировке in vitro вентральных мезенцефалических
нейронов, получаемых из индуцированных плюрипотентных стволовых клеток (ИПСК) человека. Иссле
дована связь между процессом созревания нейронов и динамикой экспрессии данных генов. Также посред
ством прижизненного наблюдения клеток с применением флуоресцентного кальциевого индикатора была
изучена функциональная активность рецепторов. С помощью трансдукции нейральных предшественников
репортерными генами TagGFP и TagRFP проанализирована роль NMDA и GABAA рецепторов в регуляции
роста нейритов и развитии спонтанной активности изучаемых клеток. Из полученных в работе результатов
следует, что агонисты изучаемых рецепторов могут быть использованы для оптимизации существующих
протоколов нейральной дифференцировки ИПСК, в частности для ускорения созревания нейронов.
КЛЮЧЕВЫЕ СЛОВА: NMDA рецепторы, GABAA рецепторы, индуцированные плюрипотентные стволо
вые клетки, мезенцефалические нейроны, дифференцировка.
DOI: 10.1134/S0320972519030138
Получение индуцированных плюрипотент
применении для моделирования конкретных
ных стволовых клеток (ИПСК) человека [1] и
физиологических и патологических процессов.
возможность их направленной дифференциров
Функционирование нейронов в роли актив
ки в специализированные типы нейронов от
ных элементов нервной системы зависит от
крывают уникальные возможности для изучения
экспрессии рецепторов нейротрансмиттеров,
патогенеза нейродегенеративных заболеваний
среди которых важную роль играют ГАМКА ре
in vitro [2]. При этом условия и временные сро
цепторы (GABAA R) и глутаматные NMDA ре
ки, в которых происходит дифференцировка
цепторы (NMDA R). Нарушения функций дан
нейронов при онтогенезе, не могут быть пол
ных рецепторов ассоциированы с различными
ностью воссозданы в культуре in vitro, что при
психическими и неврологическими заболевани
водит к фенотипическим отличиям у культиви
ями [4-7]. NMDA R и GABAA R играют важ
руемых нейронов [3] и создает ограничения в их
ную роль в развитии нервной системы, регули
Принятые сокращения: ГАМК - гамма аминомасляная кислота; NMDA - N метил D аспарагиновая кислота;
ИПСК - индуцированные плюрипотентные стволовые клетки; GABAA R - ГАМКА рецепторы; NMDA R - NMDA
рецепторы; ЭСК - эмбриональные стволовые клетки; ВМ нейроны - вентральные мезенцефалические нейроны; НП -
нейральные предшественники; кНП - коммитированные нейральные предшественники; рНП - ранние нейральные
предшественники; ОТ ПЦР - полимеразная цепная реакция с обратной транскрипцией; BDNF - нейротрофический
фактор мозга; GDNF - фактор роста нервов, выделенный из глии; ТД - терминальная дифференцировка.
* Адресат для корреспонденции.
436
NMDA И GABAA РЕЦЕПТОРЫ ПРИ ДИФФЕРЕНЦИРОВКЕ ИПСК ЧЕЛОВЕКА
437
руя пролиферацию, миграцию и дифференци
ген», Россия), следуя рекомендациям произво
ровку клеток [8, 9].
дителя.
В ряде работ было показано, что функцио
ОТ ПЦР в реальном времени. Образцы РНК
нальные NMDA R и GABAA R экспрессируют
выделяли с помощью набора РИБО золь А
ся в нейронах, получаемых из эмбриональных
(«Интерлабсервис», Россия), следуя инструкции
стволовых клеток (ЭСК) и ИПСК человека
производителя. ПЦР в реальном времени про
[10-12]. В то же время функции данных рецеп
водили с использованием амплификатора Mini
торов при дифференцировке ИПСК в специа
Opticon («Bio Rad», США), набора qPCRmix
лизированные типы нейронов, а также сходство
HS SYBR («Евроген», Россия) и праймеров, спе
профилей экспрессии данных генов in vitro и in
цифичных к указанным генам («ДНК Синтез»,
vivo остаются малоизученными.
Россия). Последовательности праймеров пред
Целью настоящей работы являлся анализ про
ставлены в таблице. Анализ полученных данных
филя экспрессии ряда субъединиц NMDA R и
осуществляли по методу 2-ΔΔCt [16].
GABAA R при дифференцировке ИПСК челове
Иммунофлуоресцентная детекция. Иммуно
ка в мезенцефалические нейроны и исследование
флуоресцентную детекцию проводили, как опи
функциональной активности данных рецепторов.
сано ранее [15], с использованием поликлональ
ных антител кролика к Glun2A («Sigma Aldrich»,
США) в разведении 1 : 300 и моноклональных
МЕТОДЫ ИССЛЕДОВАНИЯ
антител мыши к нестину («Millipore», США) в раз
ведении 1 : 750. Детекцию осуществляли с по
Культуры ИПСК и нейрональная дифференци
мощью поликлональных антител козы к имму
ровка. ИПСК линий IPSHD1.1S и IPSPDL2.15L
ноглобулинам мыши и кролика, конъюгирован
были получены путем репрограммирования
ных с красителями Alexa 546 и Alexa 488 соответ
фибробластов кожи человека с помощью ленти
ственно, в разведении 1 : 1000 («Thermo Fisher
вирусных векторов, несущих гены OCT4, SOX2,
Scientific», США).
Nanog и Klf4 [13]. Дифференцировку в вентраль
Метод TUNEL. Окрашивание апоптотичес
ные мезенцефалические нейроны (ВМ нейро
ких клеток проводили по методу, описанному в
ны) проводили в среде DMEM F12, содержав
работе Zhao и Darzynkiewicz [17], с использова
шей 2 мM L глутамин («ПанЭко», Россия) и 1×
нием Br dUTP («ДНК Синтез», Россия) и тер
добавку инсулин трансферрин селенит («ПанЭко»,
минальной нуклеотидилтрансферазы («NEB»,
Россия), по ранее опубликованному протоколу
Великобритания). Детекцию BrdU осуществля
[14], состоящему из трех стадий: 1) нейральной
ли с помощью моноклональных антител мыши
индукции с помощью двойного ингибирования
(1 : 200; «Santa Cruz», США). Для подтверждения
SMAD пути с использованием 10 мкМ SB431542
специфичности реакции окрашивание TUNEL+
(«Stemgent», США) и 80 нг/мл рекомбинантного
клеток сопоставляли с морфологией их ядер.
Noggin («Peprotech», США); 2) региональной спе
Визуализацию препаратов проводили с по
цификации с использованием 100 нг/мл Sonic
мощью флуоресцентного микроскопа Imager Z1
hedgehog (Shh) и 20 нг/мл FGF8 (фактор роста
(«Zeiss», Германия). Изображения анализиро
фибробластов 8; «Peprotech», США); 3) терми
вали с использованием программного пакета
нальной дифференцировки (ТД), которую про
ImageJ 1.49p (NCBI, США).
водили в монокультуре в среде с добавлением
Статистическую обработку данных проводи
20 нг/мл BDNF и 20 нг/мл GDNF («Peprotech»,
ли c помощью программы SigmaPlot 11 («Systat
США) либо при совместном культивировании с
Software Inc», США).
астроцитами мыши, полученными из мозга
В экспериментах по измерению внутрикле
19-20 дневных эмбрионов, в стандартной среде
точной концентрации ионов Ca2+, оценке вы
DMEM F12 без добавок (кроме 2 мM L глута
живаемости клеток и роста нейритов применяли
мина).
MK 801, мусцимол и глутамат натрия («Sigma
Измерение внутриклеточной концентрации ио
Aldrich», США).
нов кальция. Измерение проводили с использо
ванием флуоресцентного индикатора Fluo4 («Ther
mo Fisher Scientific», США) c помощью конфо
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
кального микроскопа LSM 510 («Zeiss», Герма
ния) [15]. При аппликации L глутамата («Sigma
Экспрессия генов, кодирующих субъединицы
Aldrich», США) одновременно в среду вводили
NMDA R, GABAA R и хлорные переносчики
15 мкМ глицина (коагониста NMDA R).
NKCC1 и KCC2, при дифференцировке ИПСК че
Трансфекция ИПСК. Трансфекцию осущест
ловека в ВМ нейроны. Для изучения субъеди
вляли с помощью вектора Lv Tag GFP2 («Евро
ничного состава NMDA R и GABAA R в ВМ ней
БИОХИМИЯ том 84 вып. 3 2019
438
АНТОНОВ и др.
Праймеры, используемые для проведения ОТ ПЦР в реальном времени
Ген
Прямой праймер, 5' → 3'
Обратный праймер, 5' → 3'
gapdh
AGCCACATCGCTCAGACACC
GTACTCAGCGGCCAGCATCG
grin1
AGGATGGGGACCGGAAGTTCG
AGGGCTCCTGGTGGATCGTCA
grin2a
GAAGTAATGGCACCGTCTCACC
GATTCTGGACAGGCACGGAGTT
grin2b
TCATCCCTGAGCCCAAAAGCAG
CCTCCAGGGTCACAATGCTCAG
grin2c
CATCAGTGTGATGGTGGCTCGCA
CCCGGACTTCTTGCCTCTGGTGA
grin2d
CGCTCGTGCTCACGCCCAA
GTAGGACAGGAAGGCCCGGTGG
grin3a
AAGGGGAGGGGATGTGGTAAGCG
GGCCACAACCTTGCTTTGCCTT
grin3b
CCCGGGGCGCTTCTTGGCA
AAGTCCAGCTGGCCGTCCCG
gabra1
ATGCCCATGCTTGCCCACTA
TGTTGAGCCAGAAGGAGACTTGTG
gabra2
AAAGAGGATGGGCTTGGGATGGG
GGCTTCTTGTTGGGTTCTGGCGT
gabra3
GCTGCGACCTGGGCTTGGAGAT
TGGAAGAAGGTGTCCGGTGTCCA
gabrb1
ATTCGCTTGCGGCCGGACT
GCAGCTGTGGTTGTGATTCGGA
gabrb2
ACGATGCTTCAGCTGCAAGGGTG
GGGCCCCCTCCCAAAGAAGATGT
gabrg1
GAGCTAGTGCAGCACACGTA
ACCCAGGTTTTGTTCACCGT
gabrg2
GAGCTAGTGCAGCACACGTA
CACGTGGATAGCCATAACTGGA
slc12a2
TTCCAGAAGTTTAGGGCCCGA
CGTGGCCCAAAGTTTTCTGCAA
slc12a5
CCTCAGTCACAGGGATCATGGC
CACAATGCCATCCCTCGAGATG
ронах человека была осуществлена направлен
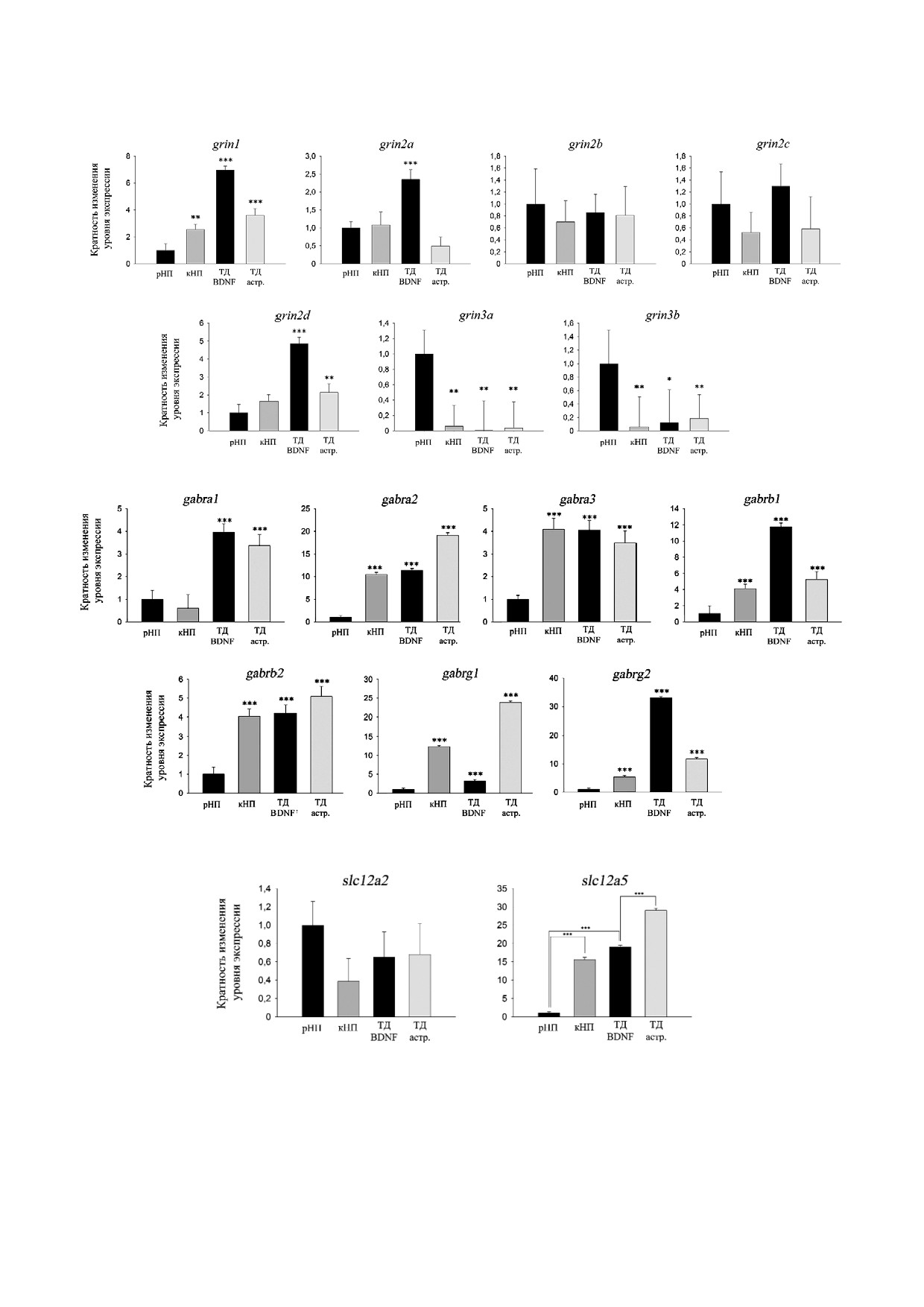
Исходя из значений ΔCT, соотношение уров
ная дифференцировка ИПСК, как описано ра
ней транскрипции генов, кодирующих субъеди
нее [14], и получены популяции клеток с содер
ницы NMDA R, было следующим (от большего
жанием значительной доли (>35%) дофаминер
к меньшему): grin1 >> grin2a = grin2b > grin2c >
гических нейронов, иммунопозитивных к тиро
grin3a > grin2d > grin3b.
зингидроксилазе, а также ГАМК+ нейронов
Уровни транскрипции генов grin1, grin2a и
(5-10%).
grin2d прогрессивно увеличивались на последо
С помощью ОТ ПЦР в реальном времени в
вательных стадиях дифференцировки. Эти из
этих культурах были проанализированы уровни
менения имели сходный характер в обеих лини
транскрипции генов, кодирующих субъедини
ях ИПСК, поэтому далее приводятся репрезен
цы NMDA R и GABAA R, а также генов slc12a2
тативные данные только по линии IPSHD1.1S
и slc12a5 на трех последовательных стадиях диф
(рис. 1, а). Уровень транскрипции указанных ге
ференцировки: ранних мультипотентных ней
нов был значительно выше при ТД клеток с ре
ральных предшественниках (рНП), коммитиро
комбинантными BDNF/GDNF по сравнению с
ванных ВМ предшественниках (кНП) и нейро
ТД в совместной культуре с астроцитами мыши.
нах при терминальной дифференцировке (ТД).
Уровень транскрипции гена grin2b оставался
Последнюю стадию дифференцировки осущест
стабильным на всех изученных стадиях и не за
вляли в двух вариантах условий: в среде с реком
висел от условий ТД.
бинантными нейротрофическими факторами
Из семи изученных субъединиц GABAA R
BDNF и GDNF (ТД BDNF) или в стандартной
наиболее выраженные изменения в транскрип
среде при совместном культивировании изучае
ции наблюдались для генов gabra2, gabrb1, gabrg1
мых клеток с астроцитами мыши (ТД астр.).
и gabrg2 (рис. 1, б). При ТД с рекомбинантными
Анализ транскрипции генов при ТД проводили
факторами был обнаружен более высокий уро
на 15 й день культивирования. Такие условия
вень транскрипции генов gabrb1 и gabrg2, а при
широко используются в современных протоко
ТД в совместной культуре с астроцитами мыши -
лах получения специализированных типов ней
более высокий уровень транскрипции генов
ронов из ИПСК [12, 18]. Изменения уровня
gabra2, gabrb2 и gabrg1.
экспрессии генов рассчитывали относительно
Дифференцировка изучаемых клеток сопро
уровня экспрессии в рНП, принимаемого за еди
вождалась значительным увеличением уровня
ницу.
транскрипции гена slc12a5, что в наибольшей
БИОХИМИЯ том 84 вып. 3 2019
NMDA И GABAA РЕЦЕПТОРЫ ПРИ ДИФФЕРЕНЦИРОВКЕ ИПСК ЧЕЛОВЕКА
439
а
б
в
Рис. 1. Динамика транскрипции генов при нейрональной дифференцировке ИПСК (линия IPSHD1.1S). рНП - ранние
мультипотентные предшественники, кНП - коммитированные ВМ предшественники, ТД BDNF - терминальная диф
ференцировка в среде с рекомбинантным нейротрофическим фактором, ТД астр. - терминальная дифференцировка в
совместной культуре с астроцитами мыши. а - Субъединицы NMDA R; б - субъединицы GABAA R; в - хлорные пере
носчики Slc12a2 (NKCC1) и Slc12a5 (KCC2). n = 4; * p < 0,05; ** p < 0,01; *** p < 0,001 по t критерию Стьюдента
БИОХИМИЯ том 84 вып. 3 2019
440
АНТОНОВ и др.
степени проявлялось при ТД в совместных куль
лы, активация которых в незрелых нейронах вы
турах с астроцитами мыши. Уровень экспрессии
зывает деполяризацию клеточной мембраны и
гена slc12a2 при этом оставался стабильным
открытие потенциал зависимых кальциевых ка
(рис. 1, в).
налов [22]. Таким образом, активность NMDA R
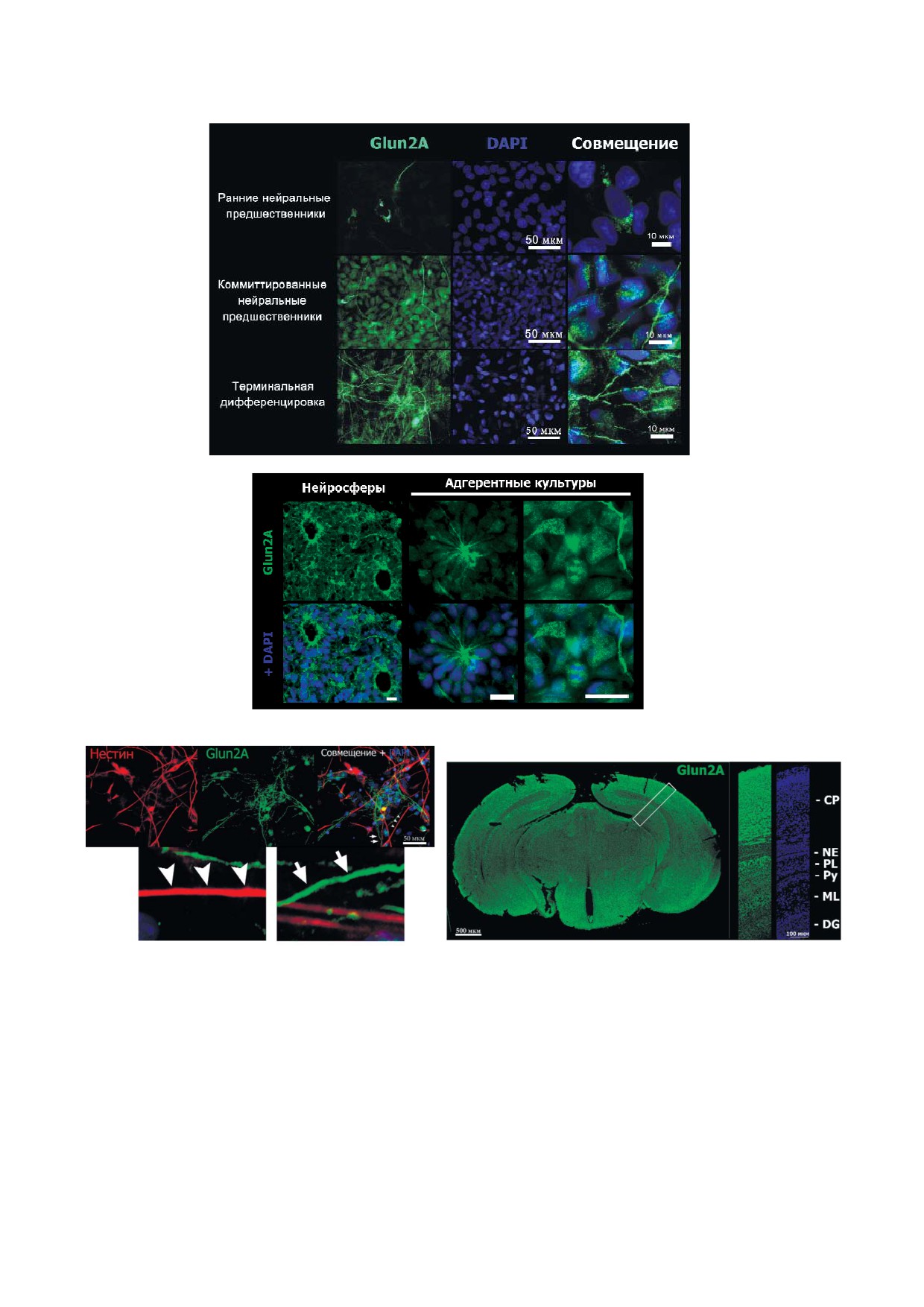
Glun2A субъединица NMDA R экспрессирует
и GABAA R ассоциирована с изменениями
ся в нейральных предшественниках и незрелых
внутриклеточной концентрации ионов кальция
нейронах человека. Известно, что у мыши
([Ca2+]i).
Glun2A субъединица NMDA R, кодируемая ге
Для установления корреляции между экспрес
ном grin2a, специфически локализуется в зре
сией генов, кодирующих субъединицы NMDA R
лых нейронах [19]. С помощью иммунофлуорес
и GABAA R, и формированием в изучаемых
центного окрашивания была проанализирована
клетках функциональных рецепторов были про
экспрессия Glun2A на последовательных стади
ведены измерения [Ca2+]i в клетках на стадии ТД
ях нейрональной дифференцировки ИПСК че
при аппликации агонистов глутаматных рецеп
ловека (рис. 2, а). Окрашивание Glun2A отсут
торов и GABAA R, с использованием для этой
ствовало в недифференцированных ИПСК че
цели кальциевого индикатора Fluo4.
ловека (данные не приведены). В культурах рНП
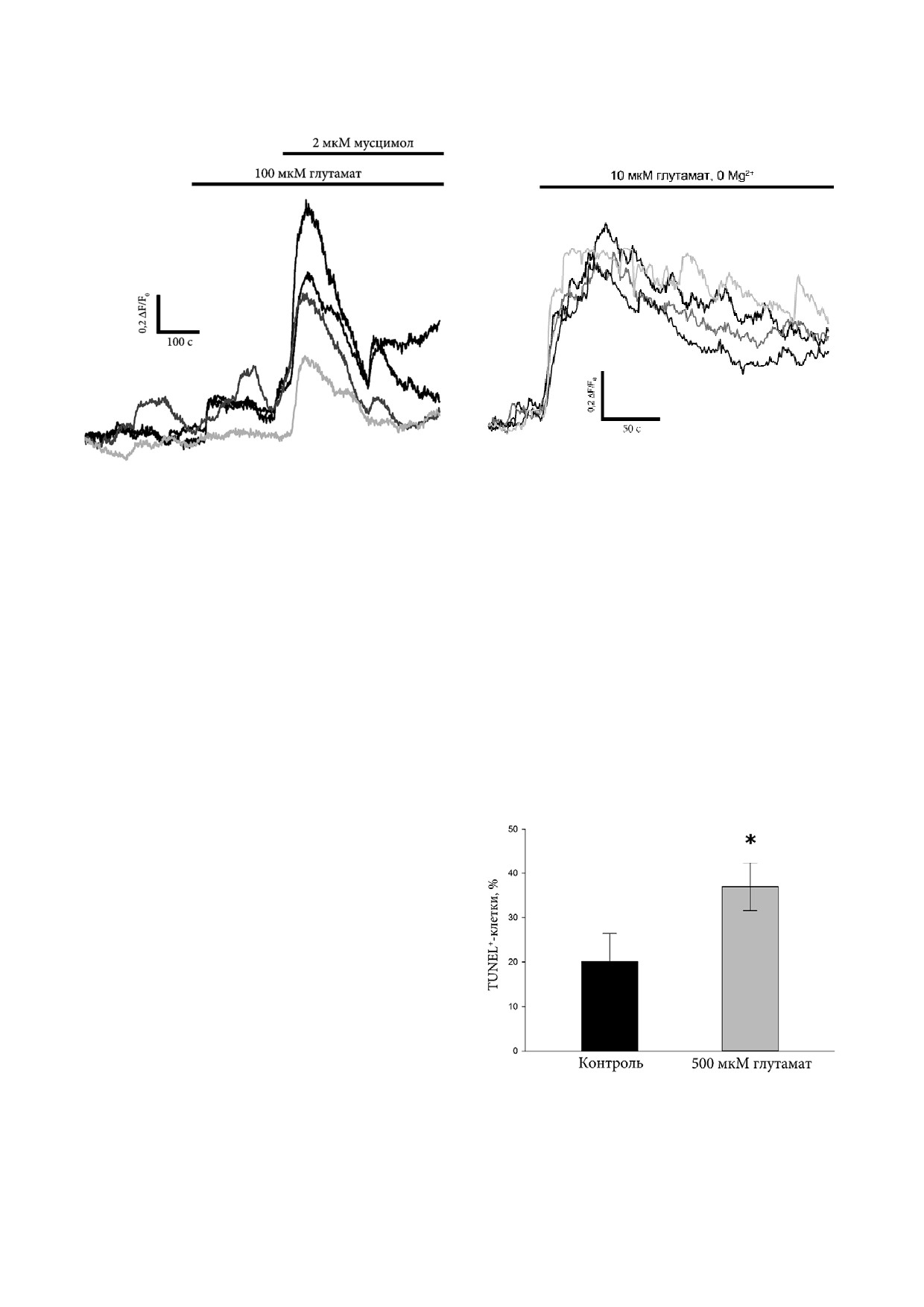
Ранее нами было продемонстрировано, что с
встречались отдельные клетки, экспрессирую
8 го по 30 й дни в ИПСК, находящихся на ста
щие Glun2A, в то время как все клетки в культу
дии ТД, активация GABAA R оказывает в изуча
рах кНП являлись Glun2A положительными.
емых нейронах возбуждающее действие, вызы
Иммунореактивность Glun2A наблюдалась так
вая увеличение [Ca2+]i [15]. В настоящей работе
же во всех клетках при ТД, при этом наиболее
аналогичное возбуждающее действие ГАМК и
интенсивно окрашивались нейриты.
селективного агониста GABAA R мусцимола
Было обнаружено, что в формирующих ро
наблюдалось на 15 й день ТД нейронов при их
зетки НП человека, являющихся специализиро
совместном культивировании с астроцитами
ванным типом мультипотентных НП [20], как в
мыши. В тех же клетках аппликация глутамата
адгерентной культуре, так и в суспензионных
(100 мкМ) в среде, содержавшей 0,7 мM Mg2+,
нейросферах иммунореактивность Glun2A пре
не приводила к изменению [Ca2+]i (рис. 3, а). В от
обладает в апикальной области клеток (рис. 2, б).
сутствие ионов Mg2+ в значительной доле клеток
Для сравнения профилей экспрессии Glun2A
при аппликации 10 мкМ глутамата наблюдалось
при нейрональной дифференцировке у человека
выраженное увеличение [Ca2+]i (рис. 3, б). По
и мыши было проведено иммунофлуоресцент
скольку известно, что ионы Mg2+ вызывают по
ное окрашивание первичных культур НП мыши
тенциал зависимое ингибирование NMDA R,
с помощью антител к Glun2A и маркеру НП нес
не влияя при этом на функционирование других
тину (рис. 2, в). В данных культурах наблюда
типов ионотропных глутаматных рецепторов
лась Glun2A иммунореактивность в нейритах,
[21], данный результат указывает на наличие в
однако солокализация с нестином не была об
изучаемых клетках на данном этапе дифферен
наружена.
цировки функциональных NMDA R, но не дру
Для сопоставления экспрессии Glun2A в НП
гих типов (AMPA или каинатных) ионотропных
человека in vitro и мыши in vivo было проведено
глутаматных рецепторов.
иммунофлуоресцентное окрашивание срезов
Согласно литературным данным, преоблада
эмбрионального мозга мыши. Иммунореактив
ющим типом глутаматных рецепторов в разви
ность различной интенсивности присутствовала
вающихся нейронах является NMDA R [23], в то
в кортикальной пластинке, полиморфном и мо
время как в зрелых нейронах - AMPA R [24]. По
лекулярном слоях гиппокампа - зонах, в кото
результатам настоящего исследования, анало
рых происходит созревание нейронов после их
гичное преобладание NMDA R наблюдается и
миграции (рис. 2, г). В то же время окрашивание
при дифференцировке ВМ нейронов человека.
Glun2A отсутствовало в неокортикальной субвент
Глутамат проявляет умеренную цитотоксич
рикулярной зоне, гиппокампальном нейроэпите
ность в культурах ВМ нейронов, полученных из
лии и зубчатой извилине гиппокампа, которые
ИПСК человека. Ранее Gupta et al. [25] продемон
являются прогениторными зонами, содержащи
стрировали эксайтотоксическое действие глута
ми популяции делящихся НП (рис. 2, г, врезка).
мата в культурах нейронов неустановленного
Экспрессия субъединиц NMDA R и GABAA R
типа, получаемых из ЭСК человека. Восприим
в ВМ нейронах человека in vitro ассоциирована с
чивость к глутамату отсутствовала у 2 недель
образованием в этих клетках функциональных ре
ных нейронов и увеличивалась в ходе диффе
цепторов. NMDA R являются катионными ка
ренцировки, достигая максимума в 8 недельных
налами, проницаемыми для ионов Ca2+ [21].
культурах [25]. Сведения о токсичности глута
GABAA R представляют собой анионные кана
мата в культурах ВМ нейронов, получаемых из
БИОХИМИЯ том 84 вып. 3 2019
NMDA И GABAA РЕЦЕПТОРЫ ПРИ ДИФФЕРЕНЦИРОВКЕ ИПСК ЧЕЛОВЕКА
441
а
б
в
г
Рис. 2. а - Иммунофлуоресцентное окрашивание Glun2A при нейрональной дифференцировке ИПСК. Левый столбец -
Glun2A, средний столбец - окрашивание ядер клеток DAPI, правый столбец - увеличенные фрагменты совмещенных
изображений; б - локализация Glun2A в апикальной области формирующих розетки НП человека. Как в суспензионных
нейросферах (слева), так и в адгерентной культуре (центральная колонка) интенсивность окрашивания Glun2A преобла
дает в апикальной зоне клеток. В других типах НП человека (справа) Glun2A не проявляет асимметричного распределе
ния. Масштабный отрезок - 20 мкм; в - двойное иммунофлуоресцентное окрашивание клеток НП мыши с помощью ан
тител к Glun2A (зеленый цвет) и нестину (красный цвет). Иммунореактивность Glun2A+ (длинные стрелки) не солокали
зуется с нестин положительными отростками (короткие стрелки). В нижнем ряду представлены увеличенные фрагменты
изображений; г - иммунофлуоресцентное окрашивание Glun2A (зеленый цвет) в мозге 19 дневного эмбриона мыши. Вы
деленный рамкой участок изображения приведен в увеличенном виде справа. CP - кортикальная пластинка, NE - гип
покампальный нейроэпителиальный слой, PL - полиморфный слой гиппокампа, Py - слой гиппокампальных пирами
дальных нейронов, ML - молекулярный слой гиппокампа, DG - зубчатая извилина гиппокампа. Ядра окрашены DAPI
(синий цвет)
БИОХИМИЯ том 84 вып. 3 2019
442
АНТОНОВ и др.
а
б
Рис. 3. Динамика изменений [Ca2+]i в клетках на 15 й день ТД в совместной культуре с астроцитами мыши: а - при воз
действии глутамата и мусцимола в стандартной среде (раствор Хенкса). Представлены графики изменения интенсивнос
ти флуоресценции Ca2+ индикатора Fluo4 от времени в четырех репрезентативных клетках; б - при воздействии глутама
та в среде, не содержавшей Mg2+ (раствор Хенкса без добавления солей Mg2+). Представлены графики изменения интен
сивности флуоресценции Ca2+ индикатора Fluo4 от времени в четырех репрезентативных клетках
ИПСК человека, в современной литературе от
Поскольку базовая среда DMEM F12, ис
сутствуют.
пользуемая для культивирования клеток в дан
В данной работе для выявления апоптоти
ной работе, содержит 50 мкМ L глутамат, для
ческих клеток в культурах ВМ нейронов челове
изучения роли NMDA R в созревании нейронов
ка был использован метод TUNEL. В наших
был применен MK 801 (селективный антаго
экспериментах глутамат вводился в культураль
нист NMDA R) в концентрации 10 мкМ. ТД про
ную среду в концентрации 500 мкМ на 14-15 й
водили в присутствии MK 801, за 24 ч до регист
день ТД. Было установлено, что в изучаемых
рации спонтанной активности ингибитор отмы
культурах исходно существует относительно
вали. Характеристики спонтанной активности
высокий базальный уровень апоптотической ги
определяли с помощью измерения динамики
бели клеток: доля TUNEL+ ядер составляла
изменений [Ca2+]i.
(20,1 ± 6,3)% (рис. 4). В присутствии 500 мкМ
глутамата доля TUNEL+ ядер возрастала до
(36,9 ± 5,4)%. Этот результат свидетельствует о
существовании в изучаемых культурах субпопу
ляции нейронов, уязвимых к токсическому
действию глутамата.
Участие NMDA R в регуляции спонтанной ак
тивности ВМ нейронов. Спонтанная активность
критически важна для созревания различных
типов нейронов [26]. Возникновение спонтан
ной активности рассматривалось в ряде работ в
качестве критерия оценки функциональной
зрелости нейронов, получаемых из ИПСК чело
века [12, 27, 28]. Поскольку известно, что
NMDA R вовлечены в регуляцию созревания
многих типов нейронов [29, 30], было проана
лизировано участие этого рецептора в форми
Рис. 4. Оценка токсичности глутамата в культурах ВМ ней
ровании спонтанной возбудимости клеток по
ронов человека с помощью метода TUNEL. Общее число кле
средством оценки максимальной амплитуды
ток определяли по числу ядер, окрашенных DAPI. * p < 0,001;
Ca2+ спайков.
n = 3
БИОХИМИЯ том 84 вып. 3 2019
NMDA И GABAA РЕЦЕПТОРЫ ПРИ ДИФФЕРЕНЦИРОВКЕ ИПСК ЧЕЛОВЕКА
443
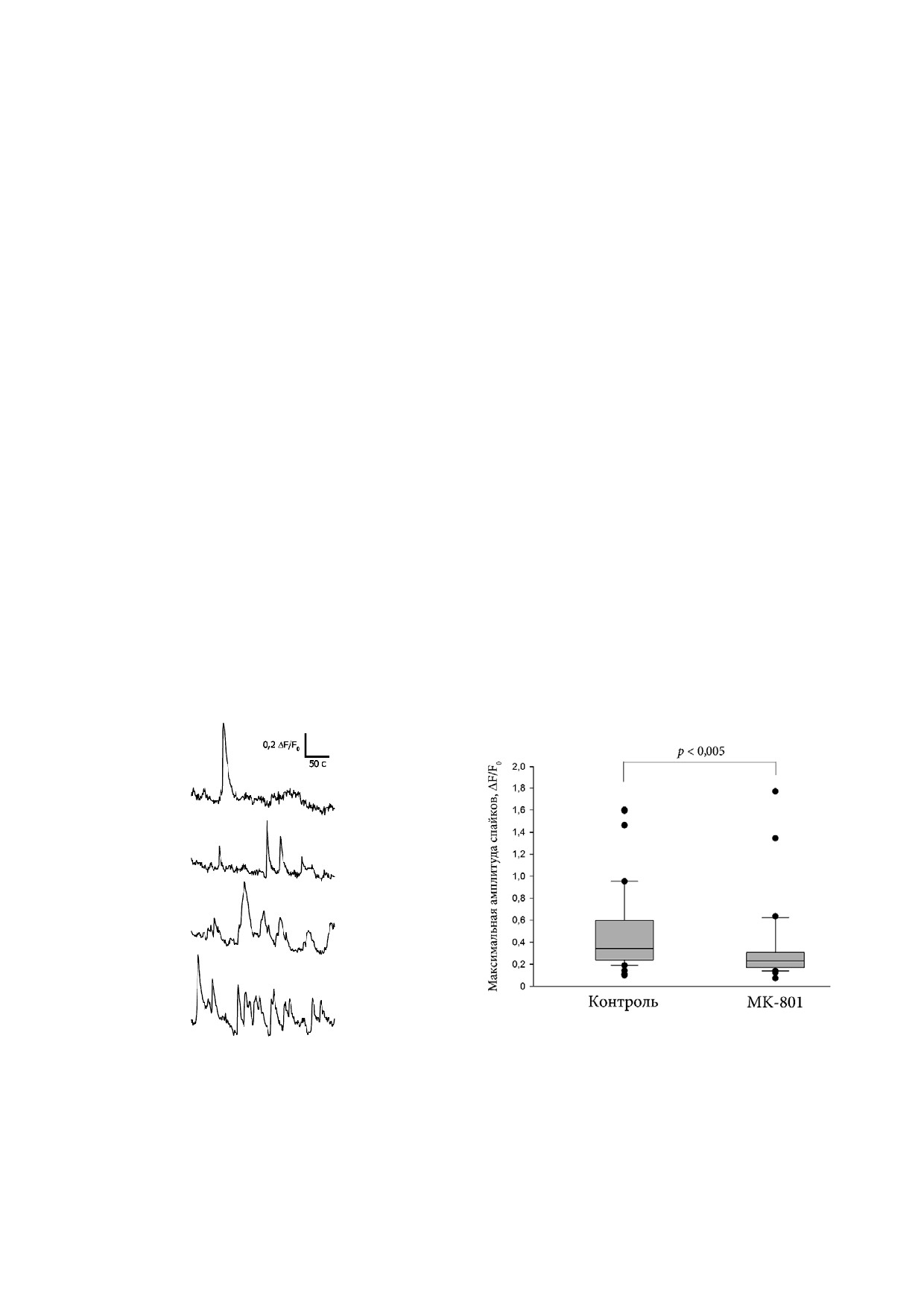
Появление спонтанной активности наблюда
довательных временных точках дифференци
лось в нейронах, получаемых из ИПСК на 10-15 й
ровки (4, 7 и 11 й дни ТД) свидетельствуют об
дни ТД. Репрезентативные записи спонтанной
эффективности данного подхода для изучения
активности индивидуальных клеток представле
факторов, влияющих на нейритогенез (данные
ны на рис. 5, а. Было установлено, что амплитуда
не приведены).
спонтанных Ca2+ спайков была значительно ни
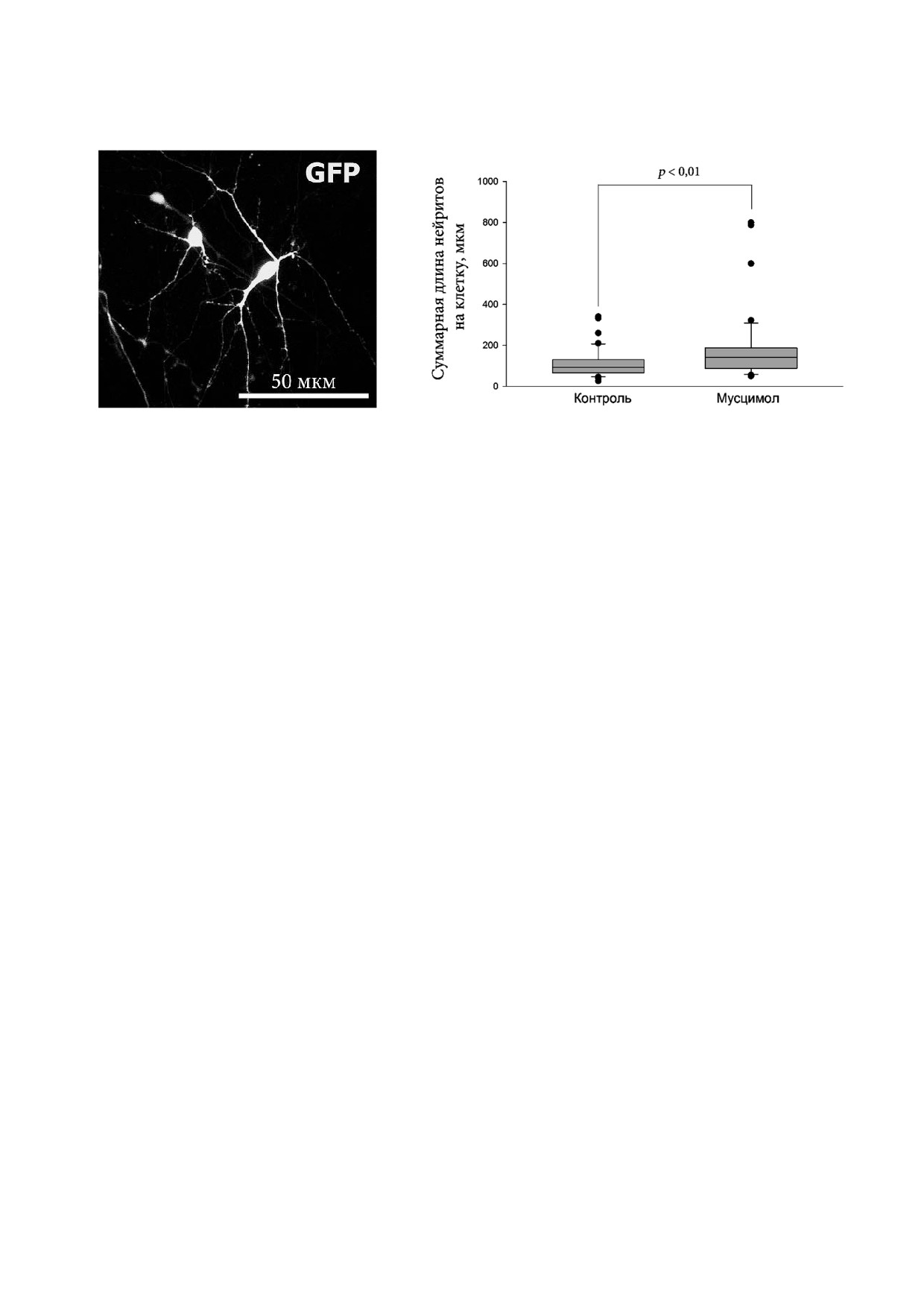
Дифференцировка в присутствии мусцимо
же в клетках, дифференцировавшихся в среде с
ла, селективного агониста GABAA R, в концент
добавлением MK 801, по сравнению с контролем
рации 2 мкМ приводила к достоверному увели
(рис. 5, б). Полученные результаты указывают на
чению длины нейритов индивидуальных клеток
участие NMDA R в регуляции функционального
при ТД (рис. 6, б). Этот результат согласуется с
созревания ВМ нейронов человека.
описанными ранее эффектами агонистов
Роль GABAА R в регуляции роста нейритов
GABAA R для других типов нейронов [31] и ука
ВМ нейронов человека. Для изучения роста ней
зывает на важную регуляторную роль GABAA R
ритов индивидуальных клеток была проведена
в определении фенотипа дифференцирующихся
трансдукция лентивирусными векторами, несу
ВМ нейронов человека. Поскольку увеличение
щими репортерные гены Tag%GFP2 и Tag%RFP.
длины нейритов является одним из характерных
Иммунофлуоресцентное окрашивание цитоске
изменений, сопровождающих созревание ней
летных белков βIII тубулина и MAP 2 не позво
ронов [22], агонисты GABAA R могут рассмат
ляло нам установить принадлежность нейритов
риваться в качестве стимуляторов созревания
к той или иной отдельной клетке в связи с тяго
нейронов человека. Данный вывод согласуется с
тением данных культур к росту в относительно
результатами Rushton et al. [32], обнаруживши
высокой плотности и формированием много
ми способность ГАМК стимулировать созрева
слойных структур и плотных сплетений нейри
ние нейронов неустановленного регионального
тов. Клетки, меченые Tag GFP2 либо Tag RFP,
фенотипа, получаемых из ИПСК человека.
смешивали с нетрансдуцированными клетками
исходной линии в отношении 1 : 10, что обеспе
чивало легкость идентификации и отслежива
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
ния нейритов отдельных меченых клеток при
сохранении оптимальной для роста плотности
Известно, что дифференцировка и созревание
культуры (рис. 6, а). Полученные статистически
нейронов сопровождаются реорганизацией субъ
значимые различия длины нейритов на после
единичного состава NMDA R и GABAA R [9, 33].
а
б
Рис. 5. Спонтанные изменения [Ca2+]i в культурах ВМ нейронов человека на 15 й день ТД. а - На репрезентативных гра
фиках представлены изменения флуоресценции Ca2+ индикатора Fluo4 в индивидуальных клетках в зависимости от вре
мени. Спонтанная активность проявлялась в виде отдельных Ca2+ спайков, очередей спайков или пейсмейкерной актив
ности; б - снижение максимальной амплитуды спонтанных Ca2+ спайков в культурах нейронов после 10 дней дифферен
цировки в среде с MK 801. Медианные значения вычисляли для 70 и более нейронов для каждой повторности, n = 4. Дос
товерность различий оценивали с помощью U критерия Манна-Уитни
БИОХИМИЯ том 84 вып. 3 2019
444
АНТОНОВ и др.
а
б
Рис. 6. Влияние мусцимола на длину нейритов индивидуальных клеток. В сумме было проанализировано >300 нейронов
в трех независимых экспериментах. а - Репрезентативное изображение нейронов, экспрессирующих Tag GFP; б - ре
зультаты анализа общей длины нейритов на клетку на 4 й день ТД
Несмотря на значительное число работ, посвя
жается уровень транскрипции гена gabra2, коди
щенных направленной дифференцировке ИПСК
рующего преобладающую эмбриональную α субъ
в ВМ нейроны, функции рецепторов нейротранс
единицу GABAA R. Параллельно усиливается
миттеров в получаемых клетках детально не
экспрессия gabra1, доминантной α субъедини
изучались [34, 35]. В данной работе впервые был
цы в различных регионах центральной нервной
проанализирован субъединичный состав и функ
системы взрослого организма [37]. Было пока
циональная активность NMDA R и GABAA R в
зано, что в ходе дифференцировки ВМ нейро
ВМ нейронах человека, получаемых из ИПСК.
нов человека in vitro уровень экспрессии gabra2
Гены grin2a, gabra1 и slc12a5 являются потенци
значительно увеличивается на стадии ТД, пре
альными маркерами зрелых нейронов. Было по
вышая уровень экспрессии gabra1. Таким обра
казано, что в изучаемых культурах профиль
зом, в изучаемых клетках, независимо от усло
экспрессии этих генов может значительно варь
вий ТД, преобладали GABAA R эмбрионального
ировать в зависимости от условий ТД и не пред
типа. Наблюдаемая нами динамика экcпрессии
определяется региональным фенотипом нейро
генов grin1, gabra2 и gabrb1 согласуется с резуль
нов. Это наблюдение позволяет предположить,
татами полнотранскриптомного анализа куль
что созревание нейронов не является единым
тур ВМ нейронов, получаемых из ИПСК чело
процессом - обретение отдельных зрелых фено
века [38]. Аналогично нашим результатам, Mom
типических характеристик может иметь незави
cilovic et al. было показано, что уровень транс
симую регуляцию.
крипции гена gabra2 превышает уровень транс
Согласно нашим данным, Glun2A экспрес
крипции гена gabra1 при ТД [38]. В то же время
сируется во всех кНП и постмитотических ВМ
транскриптомный анализ не выявил прогрес
нейронах человека. Сходный профиль экспрес
сивного роста экспрессии grin2a и grin2d на рас
сии наблюдается в переднем мозге эмбриона че
сматриваемых в настоящей работе стадиях диф
ловека, где Glun2A экспрессируется совместно с
ференцировки (рНП, кНП и ТД) [38]. Возмож
маркерами различных типов НП и молодых
но, данные противоречия объясняются разли
постмитотических нейронов [36]. Кроме того,
чиями в используемых протоколах дифферен
экспрессия Glun2A была продемонстрирована в
цировки.
незрелых нейронах неустановленного регио
Было установлено, что в ходе дифференци
нального фенотипа, получаемых из ИПСК че
ровки усиливается экспрессия гена slc12a5, что
ловека [11]. Наши результаты анализа экспрес
также наблюдалось в работе Momcilovic et al.
сии Glun2A у мыши in vitro и in vivo согласуются
[38]. Это явление характерно для созревания
с литературными данными [33], из чего следует,
большинства типов нейронов в ЦНС [22], кроме
что Glun2A может рассматриваться как маркер
дофаминергических мезенцефалических нейро
зрелых нейронов мыши, но не человека.
нов крысы [39]. Можно предположить, что про
Известно, что в ходе созревания различных
филь экспрессии slc12a5 различается в данном
типов нейронов in vivo, в т.ч. ВМ нейронов, сни
типе нейронов у человека и грызунов. Сообща
БИОХИМИЯ том 84 вып. 3 2019
NMDA И GABAA РЕЦЕПТОРЫ ПРИ ДИФФЕРЕНЦИРОВКЕ ИПСК ЧЕЛОВЕКА
445
лось, что в зрелых нейронах BDNF индуцирует
14-15 й дни ТД аппликация высоких концент
снижение уровня экспрессии slc12a5 [40], что
раций глутамата вызывает частичную гибель кле
может объяснять наблюдаемые нами различия
ток (рис. 4), но процент уязвимых клеток значи
уровня экспрессии данного гена в зависимости
тельно ниже, чем в первичных культурах ВМ ней
от условий ТД.
ронов грызунов [43]. Вероятно, эти различия
С помощью измерения динамики [Ca2+]i бы
объясняются гетерогенностью изучаемых куль
ло показано, что уже на 15 й день ТД в изучае
тур и асинхронностью созревания нейронов.
мых клетках происходит образование функцио
В настоящей работе впервые было показано,
нальных NMDA R и GABAA R (рис. 3). Эти дан
что рост нейритов и спонтанная активность кле
ные указывают на возможность изучения эф
ток, т.е. характеристики нейронов, тесно свя
фектов ГАМКергических и глутаматергических
занные с их созреванием, способны регулиро
препаратов в культурах ВМ нейронов человека
ваться в культурах ВМ нейронов человека при
на данной стадии дифференцировки без необ
участии NMDA R и GABAA R и их лигандов
ходимости продолжительного культивирования
(рис. 5, б; рис. 6, б). Таким образом, агонисты
клеток с целью получения зрелых нейронов [27].
изучаемых рецепторов потенциально могут
Активация GABAA R в изучаемых клетках
быть использованы для оптимизации существу
приводит к увеличению [Ca2+]i, что характерно
ющих протоколов нейральной дифференциров
для незрелых нейронов [22]. Данный характер
ки ИПСК, в частности для ускорения созрева
ответов сохраняется до 30 дней ТД. Глутамат
ния нейронов.
индуцированный ток ионов Ca2+, ингибируе
мый ионами Mg2+, свидетельствует о наличии
функциональных NMDA R в изучаемых клет
Финансирование
ках (рис. 3, а). На 10-15 й дни ТД при апплика
ции глутамата в непроточной системе увеличе
Работа выполнена при поддержке РФФИ
ние [Ca2+]i в клетках имеет транзиентный харак
(грант № 17 04 01661; Антонов С.А., Новосадо
тер. На более поздних сроках дифференцировки
ва Е.В., Кобылянский А.Г., Гривенников И.А.;
повышенный уровень [Ca2+]i сохраняется на
эксперименты по кальциевому имиджингу,
протяжении длительного периода времени. Та
трансфекции, ОТ ПЦР и иммунофлуоресцен
ким образом, в ходе созревания ВМ нейронов
ции) и РНФ (грант № 14 15 01047 П; Новосадо
человека снижается склонность NMDA R к де
ва Е.В., Иллариошкин С.Н., Гривенников И.А.;
сенситизации.
работы по культивированию, дифференцировке
Эксайтотоксичность рассматривается как
и исследованию апоптоза нейронов человека).
одна из основных причин гибели нейронов при
ишемическом инсульте и нейродегенеративных
Конфликт интересов
заболеваниях [4-6]. Моделирование этого про
цесса in vitro является важным для изучения па
Авторы заявляют об отсутствии конфликта
тогенеза этих заболеваний и поиска новых лека
интересов.
рственных средств. Эксайтотоксичность опос
редована через гиперактивацию глутаматных ре
Соблюдение этических норм
цепторов и необратимое увеличение [Ca2+]i [41].
Токсичность глутамата также может реализовы
Все применимые международные, националь
ваться через инверсию работы цистин глутамат
ные и/или институциональные принципы ухода
ного антипортера [42]. Было показано, что на
и использования животных были соблюдены.
СПИСОК ЛИТЕРАТУРЫ
1.
Takahashi, K., Tanabe, K., Ohnuki, M., Narita, M.,
turing strategy improves functional neuronal development
Ichisaka, T., Tomoda, K., and Yamanaka, S. (2007) In
of human neural progenitor cells, J. Neurochem., 109,
duction of pluripotent stem cells from adult human fibrob
238-247, doi: 10.1111/j.1471 4159.2009.05954.x.
lasts by defined factors, Cell, 131, 861-872, doi: 10.1016/
4.
Zhou, Q., and Sheng, M. (2013) NMDA receptors in nervous
j.cell.2007.11.019.
system diseases, Neuropharmacology, 74, 69-75, doi: 10.1016/
2.
Новосадова Е.В., Гривенников И.А. (2014) Индуциро
j.neuropharm.2013.03.030.
ванные плюрипотентные стволовые клетки: от полу
5.
Koutsilieri, E., and Riederer, P. (2007) Excitotoxicity and
чения до применения в биохимических и биомедици
new antiglutamatergic strategies in Parkinson’s disease and
нских исследованиях, Успехи биологической химии, 54,
Alzheimer’s disease, Parkinsonism Relat. Disord., 13,
3-38.
S329-S331, doi: 10.1016/s1353 8020(08)70025 7.
3.
Schaarschmidt, G., Schewtschik, S., Kraft, R., Wegner, F.,
6.
Dong, X.X., Wang, Y., and Qin, Z.H. (2009) Molecular
Eilers, J., Schwarz, J., and Schmidt, H. (2009) A new cul
mechanisms of excitotoxicity and their relevance to patho
БИОХИМИЯ том 84 вып. 3 2019
446
АНТОНОВ и др.
genesis of neurodegenerative diseases, Acta Pharmacol.
J. Neurosci., 24, 8885-8895, doi: 10.1523/jneurosci.2476
Sin., 30, 379-387, doi: 10.1038/aps.2009.24.
04.2004.
7.
Rowley, N.M., Madsen, K.K., Schousboe, A., and Steve
20.
Elkabetz, Y., Panagiotakos, G., Al Shamy, G., Socci, N.D.,
White, H. (2012) Glutamate and GABA synthesis, release,
Tabar, V., and Studer, L. (2008) Human ES cell derived
transport and metabolism as targets for seizure control,
neural rosettes reveal a functionally distinct early neural stem cell
Neurochem. Int., 61, 546-558, doi: 10.1016/j.neuint.
stage, Genes Dev., 22, 152-165, doi: 10.1101/gad.1616208.
2012.02.013.
21.
Dingledine, R., Borges, K., Bowie, D., and Traynelis, S.F.
8.
Nacher, J., and McEwen, B.S. (2006) The role of N me
(1999) The glutamate receptor ion channels, Pharmacol.
thyl D asparate receptors in neurogenesis, Hippocampus,
Rev., 51, 7-61.
16, 267-270, doi: 10.1002/hipo.20160.
22.
Ben Ari, Y. (2002) Excitatory actions of gaba during deve
9.
Galanopoulou, A.S. (2008) GABAA receptors in normal
lopment: the nature of the nurture, Nat. Rev. Neurosci., 3,
development and seizures: friends or foes? Curr. Neuro%
728-739, doi: 10.1038/nrn920.
pharmacol., 6, 1-20, doi: 10.2174/157015908783769653.
23.
Herlenius, E., and Lagercrantz, H. (2010) Neurotrans
10.
Young, A., Machacek, D.W., Dhara, S.K., Macleish, P.R.,
mitters and neuromodulators during brain development, in
Benveniste, M., Dodla, M.C., Sturkie, C.D., and Stice, S.L.
The newborn brain: neuroscience and clinical applications
(2011) Ion channels and ionotropic receptors in human
(Lagercrantz, H., ed.), 2nd еdn, Cambridge University
embryonic stem cell derived neural progenitors, Neuro%
Press, pp. 99-119.
science, 192, 793-805, doi: 10.1016/j.neuroscience.2011.
24.
Rogawski, M.A. (2011) Revisiting AMPA receptors as an anti
04.039.
epileptic drug target, Epilepsy Curr., 11, 56-63, doi: 10.5698/
11.
Zhang, W.B., Ross, P.J., Tu, Y., Wang, Y., Beggs, S.,
1535 7511 11.2.56.
Sengar, A.S., Ellis, J., and Salter, M.W. (2016) Fyn kinase
25.
Gupta, K., Hardingham, G.E., and Chandran, S. (2013)
regulates GluN2B subunit dominant NMDA receptors in
NMDA receptor dependent glutamate excitotoxicity in
human induced pluripotent stem cell derived neurons, Sci.
human embryonic stem cell derived neurons, Neurosci.
Rep., 6, 23837, doi: 10.1038/srep23837.
Lett., 543, 95-100, doi: 10.1016/j.neulet.2013.03.010.
12.
Ishii, M.N., Yamamoto, K., Shoji, M., Asami, A., and
26.
Bergey, G.K., Fitzgerald, S.C., Schrier, B.K., and Nelson, P.G.
Kawamata, Y. (2017) Human induced pluripotent stem cell
(1981) Neuronal maturation in mammalian cell culture is
(hiPSC) derived neurons respond to convulsant drugs
dependent on spontaneous electrical activity, Brain Res.,
when co cultured with hiPSC derived astrocytes, Toxi%
207, 49-58, doi: 10.1016/0006 8993(81)90678 8
cology, 389, 130-138, doi: 10.1016/j.tox.2017.06.010.
27.
Lam, R.S., Topfer, F.M., Wood, P.G., Busskamp, V., and
13.
Некрасов Е.Д., Лебедева О.С., Честков И.В., Сюсина М.А.,
Bamberg, E. (2017) Functional maturation of human stem
Федотова Е.Ю., Лагарькова М.А., Киселев С.Л., Гри
cell derived neurons in long term cultures, PLoS One, 12,
венников И.А., Иллариошкин С.Н. (2011) Получение
e0169506, doi: 10.1371/journal.pone.0169506.
и характеристика индуцированных плюрипотентных
28.
Gunhanlar, N., Shpak, G., van der Kroeg, M., Gouty
стволовых клеток человека из фибробластов кожи па
Colomer, L.A., Munshi, S.T., Lendemeijer, B., Ghazvini, M.,
циентов с нейроденеративными заболеваниями, Клеточ%
Dupont, C., Hoogendijk, W.J.G., Gribnau, J., de Vrij, F.M.S.,
ная трансплантология и тканевая инженерия, 6, 1-7.
and Kushner, S.A. (2017) A simplified protocol for differen
14.
Лебедева О.С., Новосадова Е.В., Мануилова Е.С., Ар
tiation of electrophysiologically mature neuronal networks
сеньева Е.Л., Киселев С.Л., Лагарькова М.А., Хаспе
from human induced pluripotent stem cells, Mol.
ков Л.Г., Иллариошкин С.Н., Гривенников И.А. (2014)
Psychiatry, 23, 1336-1344, doi: 10.1038/mp.2017.56.
Получение и характеристика клеточной модели бо
29.
Hirasawa, T., Wada, H., Kohsaka, S., and Uchino, S.
лезни Паркинсона на основе индуцированных плю
(2003) Inhibition of NMDA receptors induces delayed
рипотентных стволовых клеток, в сб. Стволовые клет%
neuronal maturation and sustained proliferation of progeni
ки и регенеративная медицина (под ред. Ткачука В.А.),
tor cells during neocortical development, J. Neurosci. Res.,
Изд во Московского Университета, Москва, с. 154-168.
74, 676-687, doi: 10.1002/jnr.10795.
15.
Antonov, S.A., Novosadova, E.V., Arsenyeva, E.L.,
30.
Yamasaki, M., Okada, R., Takasaki, C., Toki, S., Fukaya, M.,
Grefenstein, M.A., Zykova, A.A., Kobylyansky, A.G.,
Natsume, R., Sakimura, K., Mishina, M., Shirakawa, T.,
Manuilova, E.S., Grivennikov, I.A., Illarioshkin, S.N., and
and Watanabe, M. (2014) Opposing role of NMDA recep
Myasoedov, N.F. (2016) Investigation of the effects of GABA
tor GluN2B and GluN2D in somatosensory development
receptor agonists in the differentiation of human induced
and maturation, J. Neurosci., 34, 11534-11548, doi: 10.1523/
pluripotent stem cells into dopaminergic neurons, Dokl.
jneurosci.1811 14.2014.
Biol. Sci., 470, 244-246, doi: 10.1134/s0012496616050045.
31.
Sernagor, E., Chabrol, F., Bony, G., and Cancedda, L.
16.
Livak, K.J., and Schmittgen, T.D. (2001) Analysis of rela
(2010) GABAergic control of neurite outgrowth and
tive gene expression data using real time quantitative PCR
remodeling during development and adult neurogenesis:
and the 2-ΔΔCT method, Methods, 25, 402-408, doi: 10.1006/
general rules and differences in diverse systems, Front. Cell
meth.2001.1262.
Neurosci., 4, 11, doi: 10.3389/fncel.2010.00011.
17.
Zhao, H., and Darzynkiewicz, Z. (2017) Rapid detection
32.
Rushton, D.J., Mattis, V.B., Svendsen, C.N., Allen, N.D.,
of DNA strand breaks in apoptotic cells by flow and
and Kemp, P.J. (2013) Stimulation of GABA induced Ca2+
image cytometry, Methods Mol. Biol., 1644, 139-149,
influx enhances maturation of human induced pluripotent
doi: 10.1007/978 1 4939 7187 9_12.
stem cell derived neurons, PLoS One, 8, e81031, doi: 10.1371/
18.
Verpelli, C., Carlessi, L., Bechi, G., Fusar Poli, E.,
journal.pone.0081031.
Orellana, D., Heise, C., Franceschetti, S., Mantegazza, R.,
33.
Paoletti, P., Bellone, C., and Zhou, Q. (2013) NMDA
Mantegazza, M., Delia, D., and Sala, C. (2013) Compara
receptor subunit diversity: impact on receptor properties,
tive neuronal differentiation of self renewing neural prog
synaptic plasticity and disease, Nat. Rev. Neurosci., 14,
enitor cell lines obtained from human induced pluripotent
383-400, doi: 10.1038/nrn3504.
stem cells, Front. Cell. Neurosci., 7, 1-14, doi: 10.3389/
34.
Hartfield, E.M., Yamasaki Mann, M., Ribeiro Fernandes, H.J.,
fncel.2013.00175.
Vowles, J., James, W.S., Cowley, S.A., and Wade Martins, R.
19.
Liu, X.B., Murray, K.D., and Jones, E.G. (2004) Switching
(2014) Physiological characterisation of human iPS
of NMDA receptor 2A and 2B subunits at thalamic and
derived dopaminergic neurons, PLoS One, 9, e87388, doi:
cortical synapses during early postnatal development,
10.1371/journal.pone.0087388.
БИОХИМИЯ том 84 вып. 3 2019
NMDA И GABAA РЕЦЕПТОРЫ ПРИ ДИФФЕРЕНЦИРОВКЕ ИПСК ЧЕЛОВЕКА
447
35. Stanslowsky, N., Haase, A., Martin, U., Naujock, M.,
GABAA receptor mediated inhibition in rat substantia
Leffler, A., Dengler, R., and Wegner, F. (2014) Functional
nigra, J. Neurosci., 23, 8237-8246, doi: 10.1523/jneu
differentiation of midbrain neurons from human cord
rosci.23 23 08237.2003
blood derived induced pluripotent stem cells, Stem Cell
40. Kaila, K., Price, T.J., Payne, J.A., Puskarjov, M., and
Res. Ther., 5, 35, doi: 10.1186/scrt423.
Voipio, J. (2014) Cation chloride cotransporters in neu
36. Bagasrawala, I., Memi, F., Radonjic, N., and Zecevic, N.
ronal development, plasticity and disease, Nat. Rev.
(2017) N Methyl D aspartate receptor expression patterns
Neurosci., 15, 637-654, doi: 10.1038/nrn3819.
in the human fetal cerebral cortex, Cereb. Cortex, 27,
41. Wang, C., Liu, F., Patterson, T.A., Paule, M.G., and
5041-5053, doi: 10.1093/cercor/bhw289.
Slikker, W., Jr. (2017) Relationship between ketamine
37. Skilbeck, K.J., Johnston, G.A., and Hinton, T. (2010)
induced developmental neurotoxicity and NMDA recep
Stress and GABA receptors, J. Neurochem.,
112,
tor mediated calcium influx in neural stem cell derived
1115-1130, doi: 10.1111/j.1471 4159.2009.06539.x.
neurons, Neurotoxicology, 60, 254-259, doi: 10.1016/
38. Momcilovic, O., Liu, Q., Swistowski, A., Russo Tait, T.,
j.neuro.2016.04.015.
Zhao, Y., Rao, M.S., and Zeng, X. (2014) Genome wide
42. Kritis, A.A., Stamoula, E.G., Paniskaki, K.A., and Vavilis, T.D.
profiling of dopaminergic neurons derived from human
(2015) Researching glutamate induced cytotoxicity in dif
embryonic and induced pluripotent stem cells, Stem Cells
ferent cell lines: a comparative/collective analysis/study, Front.
Dev., 23, 406-420, doi: 10.1089/scd.2013.0412.
Cell Neurosci., 9, 1-18, doi: 10.3389/fncel. 2015.00091.
39. Gulacsi, A., Lee, C.R., Sik, A., Viitanen, T., Kaila, K.,
43. Kikuchi, S., and Kim, S.U. (1993) Glutamate neurotoxicity
Tepper, J.M., and Freund, T.F. (2003) Cell type specific
in mesencephalic dopaminergic neurons in culture, J. Neuro%
differences in chloride regulatory mechanisms and
sci. Res., 36, 558-569, doi: 10.1002/jnr.490360508.
EXPRESSION AND FUNCTIONAL PROPERTIES
OF NMDA AND GABAA RECEPTORS DURING
THE DIFFERENTIATION OF HUMAN INDUCED
PLURIPOTENT STEM CELLS INTO VENTRAL
MESENCEPHALIC NEURONS
S. A. Antonov1*, E. V. Novosadova1, A. G. Kobylyansky1,
S. N. Illarioshkin2, V. Z. Tarantul1, and I. A. Grivennikov1
1 Institute of Molecular Genetics, Russian Academy of Sciences,
123182 Moscow, Russia; E%mail: vamore@inbox.ru
2 Research Center of Neurology, 125367 Moscow, Russia
Received October 29, 2018
Revised November 29, 2018
Accepted November 29, 2018
Ionotropic glutamate and GABA receptors regulate differentiation and determine functional properties of mature
neurons. Both insufficient and excessive activity of these neurotransmission systems is associated with various nervous
system diseases. Our knowledge regarding the expression profiles of these receptors and the mechanisms of their regu
lation during differentiation of specialized human neuron subtypes is limited. Here, using real time PCR and
immunocytochemical staining, we have investigated expression profile of the NMDA and GABAA receptor subunits
during in vitro differentiation of human induced pluripotent stem cells (iPSCs) into ventral mesencephalic neurons.
The most prominent changes were observed in expression levels of grin1, grin2a, grin2d, gabra2, gabrg1 and gabrg2
genes. Furthermore, differentiation was accompanied with a significant increase in potassium chloride KCC2 sym
porter expression, that is indicative of neuronal maturation. Using calcium imaging, it was shown that the expression
of these genes is associated with the formation of functional receptors. We have found that NMDA and GABAA recep
tors are involved in the regulation of neurite outgrowth and the development of spontaneous activity in the studied
cells. The obtained results indicate a possibility of using ligands of the NMDA and GABAA receptors for regulating
the phenotype of human neurons during their differentiation in vitro.
Keywords: NMDA receptors, GABAA receptors, induced pluripotent stem, mesencephalic neurons, differentiation
БИОХИМИЯ том 84 вып. 3 2019