БИОХИМИЯ, 2019, том 84, вып. 6, с. 827 - 835
УДК 577.355.3
АЛГОРИТМ ИЗВЛЕЧЕНИЯ КИНЕТИКИ СЛАБЫХ
ПОЛОС В ДИФФЕРЕНЦИАЛЬНЫХ СПЕКТРАХ
ПОГЛОЩЕНИЯ РЕАКЦИОННЫХ ЦЕНТРОВ
Rhodobacter sphaeroides*
© 2019
Р.А. Хатыпов1**, А.М. Христин1,
Л.Г. Васильева1, В.А. Шувалов1,2
1 Институт фундаментальных проблем биологии РАН, ФИЦ «Пущинский
научный центр биологических исследований РАН», 142290 Пущино,
Московская обл., Россия; электронная почта: rgreen1@rambler.ru
2 Институт химической физики им. Н.Н. Семенова РАН,
119991, Москва, Россия
Поступила в редакцию 15.02.2019
После доработки 21.03.2019
Принята к публикации 21.03.2019
Предложен способ построения кинетики ион радикальных полос на фоне интенсивного поглощения в
дифференциальных спектрах реакционных центров Rhodobacter sphaeroides при фемтосекундном возбужде
нии первичного донора электрона. Построенные предложенным методом кинетика полосы поглощения
при 1020 нм и кинетика выцветания полосы при 545 нм удовлетворительно описываются кинетическими
уравнениями для последовательного переноса электрона от возбужденного первичного донора на молекулу
мономерного бактериохлорофилла (BA) и затем - на молекулу бактериофеофитина, который служит акцеп
тором электрона (HA), c константами скоростей 3,5 ± 0,2 и 0,8 ± 0,1 пс соответственно. В кинетике полосы
поглощения бактериохлорофилла при 600 нм наблюдается как сверхбыстрое выцветание димера Р870, так и
более медленное выцветание мономера BA, обусловленное переходом его в анион радикальную форму.
Построенные кинетики ион радикальных полос согласуются с профилями концентраций состояний с раз
деленными зарядами, полученными нами методом глобального анализа экспериментальных данных в рам
ках модели последовательного переноса электрона в реакционных центрах.
КЛЮЧЕВЫЕ СЛОВА: фемтосекундная спектроскопия; бактериальные реакционные центры; перенос
электрона.
DOI: 10.1134/S0320972519060083
Фотосинтетические реакционные центры
дами. Кофакторами переноса электрона явля
Rhodobacter sphaeroides представляют собой
ются два бактериохлорофилла (БХл), образую
трансмембранные белковые комплексы, состо
щих димер первичного донора электрона (Р),
ящие из трех субъединиц L, M и H и десяти ко
два мономерных бактериохлорофилла (BA и BВ),
факторов переноса электрона, в которых проис
два бактерифеофитина (HA и HВ), два хинона
ходят начальные сверхбыстрые фотофизические
(QA и QВ), атом негемового железа и молекула
стадии процесса преобразования световой энер
каротина. Кофакторы организованы в две псев
гии в энергию состояний с разделенными заря
досимметричные цепи А и В, из которых актив
ной является только цепь А [1, 2].
Известно, что под действием света в реакци
Принятые сокращения: РЦ - реакционный центр;
Р - первичный донор электрона; БХл - бактериохлоро
онных центрах происходит трансмембранный
филл; ВА - мономерный БХл в активной цепи кофакторов;
перенос электрона от возбужденного донора Р*
НА - бактериофеофитин, который служит акцептором
электрона; QA - первичный хинонный акцептор; QB - вто
на молекулу хинона QA. В настоящее время об
ричный хинонный акцептор; фс - фемтосекунда; ΔA - из
щепринято, что трансмембранный перенос
менение поглощения; ΔGauss - изменение доли гауссовой электрона осуществляется через серию сверх
полосы поглощения.
быстрых реакций, включающих молекулы BA и HA
* Первоначально английский вариант рукописи опублико
активной цепи кофакторов в качестве промежу
точных акцепторов электрона. Существует зна
msu.ru/biokhimiya, в рубрике «Papers in Press», BM 19 039,
06.05.2019.
чительная экспериментальная сложность обна
** Адресат для корреспонденции.
ружения состояния P+B–, связанная с тем, что
827
6*
828
ХАТЫПОВ и др.
скорость переноса электрона на молекулу BA
оценки квантового выхода образования состоя
меньше скорости переноса электрона с B– на НА
ния P+Н– в мутантных РЦ Rba capsulatus. Другим
[3, 4]. Это приводит к малой заселенности со
подходом для решения проблемы перекрываю
стояния P+B–, которая не превышает нескольких
щихся полос было построение кинетики анион
десятков процентов. Другая сложность связана с
радикала B– при 1020 нм с использованием ап
тем, что спектр состояния P+B– замаскирован
проксимации спектральной подставки линией
спектрами других состояний, включая Р*. Обна
на широком интервале длин волн [13]. В данной
-
ружение минорной полосы анион радикала BA
работе мы предлагаем иной способ извлечения
при 1020 нм в дифференциальных спектрах по
кинетики отдельных, в частности, короткожи
глощения послужило основанием для обоснова
вущих, слабых полос поглощения и анализиру
ния четырехкомпонентной модели, в которой
ем эти кинетики в рамках модели последова
первичным акцептором электрона является мо
тельного переноса электрона.
лекула BA [5, 6]. Согласно этой модели, транс
мембранный перенос электрона осуществляется
от возбужденного донора Р* на молекулу BA с
МЕТОДЫ ИССЛЕДОВАНИЯ
константой скорости ~3 пс, затем на молекулу
HA с константой скорости ~1 пс и на молекулу
Реакционные центры Rba sphaeroides выде
QA с константой скорости ~200 пс.
ляли согласно описанной ранее методике [14].
Затем РЦ были суспендированы в буфере, со
3 пс
1 пс
200 пс
держащем 20 мМ Tris HCl (pH 8,0) и 0,1% (v/v)
P* —→ BA —→ HA ——→ QA
Тритон X 100, и сконцентрированы до поглоще
ния 0,5 при 870 нм (длина оптического пути 1 мм).
В последнее время исследователи первичных
Спектры поглощения реакционных центров из
процессов разделения зарядов в реакционном
меряли на спектрофотометре UV1800 («Shimadzu»,
центре (РЦ) фотосинтеза все чаще используют
Япония).
глобальный анализ массива дифференциальных
Подробное описание фемтосекундного ла
спектров поглощения, измеренных с фемтосе
зерного спектрометра приведено ранее [15].
кундным (фс) временным разрешением [7], ко
Фемтосекундные импульсы света (~35 фс, ~3 мДж)
торый позволяет описать весь набор спектраль
получали с помощью фемтосекундного лазера
ных данных с использованием математической
MaiTai SP с регенеративным усилителем Spitfire
и кинетической модели.
Ace («Spectra Physics», США). Для возбуждения
Этот метод позволяет аппроксимировать
первичного донора электрона при 870 нм из
экспериментальные данные с помощью разных
континуума, полученного на сапфире, свето
моделей: 1) модели последовательного переноса
фильтром RG850 («Newport», США) выделяли
электрона [8-11]; 2) модели переноса электрона
длины волн >850 нм. Измерительный луч дели
с учетом обратимости реакций разделения заря
ли на зондирующий и опорный. После вращаю
дов [8-10]; 3) модели разделения зарядов по
щейся кюветы с образцом (толщина 1 мм) зон
двум каналам дезактивации в гетерогенном бел
дирующий и опорный импульсы направляли
ковом окружении кофакторов в РЦ [10]. Следу
сначала в спектрограф Spectra Pro 2300i, а затем
ет отметить, что разделение короткоживущих
на CCD, Pixis 400BR («Princeton Instruments»,
состояний методом глобального анализа затруд
США) для регистрации спектров в диапазонах
нено, если длительность возбуждающего им
450-720 или 750-1100 нм. Относительная поля
пульса достаточно мала и наблюдаются когерент
ризация импульса возбуждения была параллель
ные эффекты. Таким образом, использование
на измерительному импульсу. Частота вращения
глобального анализа требует от исследователя
кюветы и частота возбуждения (10 Гц) подбира
привлечения дополнительной информации о
лась таким образом, чтобы каждый возбуждаю
процессах разделения зарядов для предложения
щий импульс освещал новую часть РЦ. Интен
адекватной физической модели.
сивность возбуждающих импульсов ослабляли
В ряде случаев по прежнему остается акту
так, чтобы возбуждались менее 20% реакцион
альным анализ кинетики изменения поглоще
ных центров в образце. Все измерения проводи
ния ΔA на заданной длине волны. Однако такой
ли при комнатной температуре.
анализ бывает осложнен вкладом различных пе
Глобальный анализ массива разрешенных во
рекрывающихся спектральных компонентов. В ка
времени дифференциальных спектров поглощения
честве возможного подхода для решения этой
проводили с помощью программы Glotaran [16].
проблемы в работе Carter et al. [12] был проведен
Алгоритм извлечения кинетики спектральных
экспоненциальный анализ сложной кинетики
полос. При фемтосекундном возбуждении пер
изменения поглощения ΔA при 600 нм для
вичного донора электрона в дифференциальных
БИОХИМИЯ том 84 вып. 6 2019
ПЕРВИЧНЫЕ ПРОЦЕССЫ РАЗДЕЛЕНИЯ ЗАРЯДОВ В РЦ
829
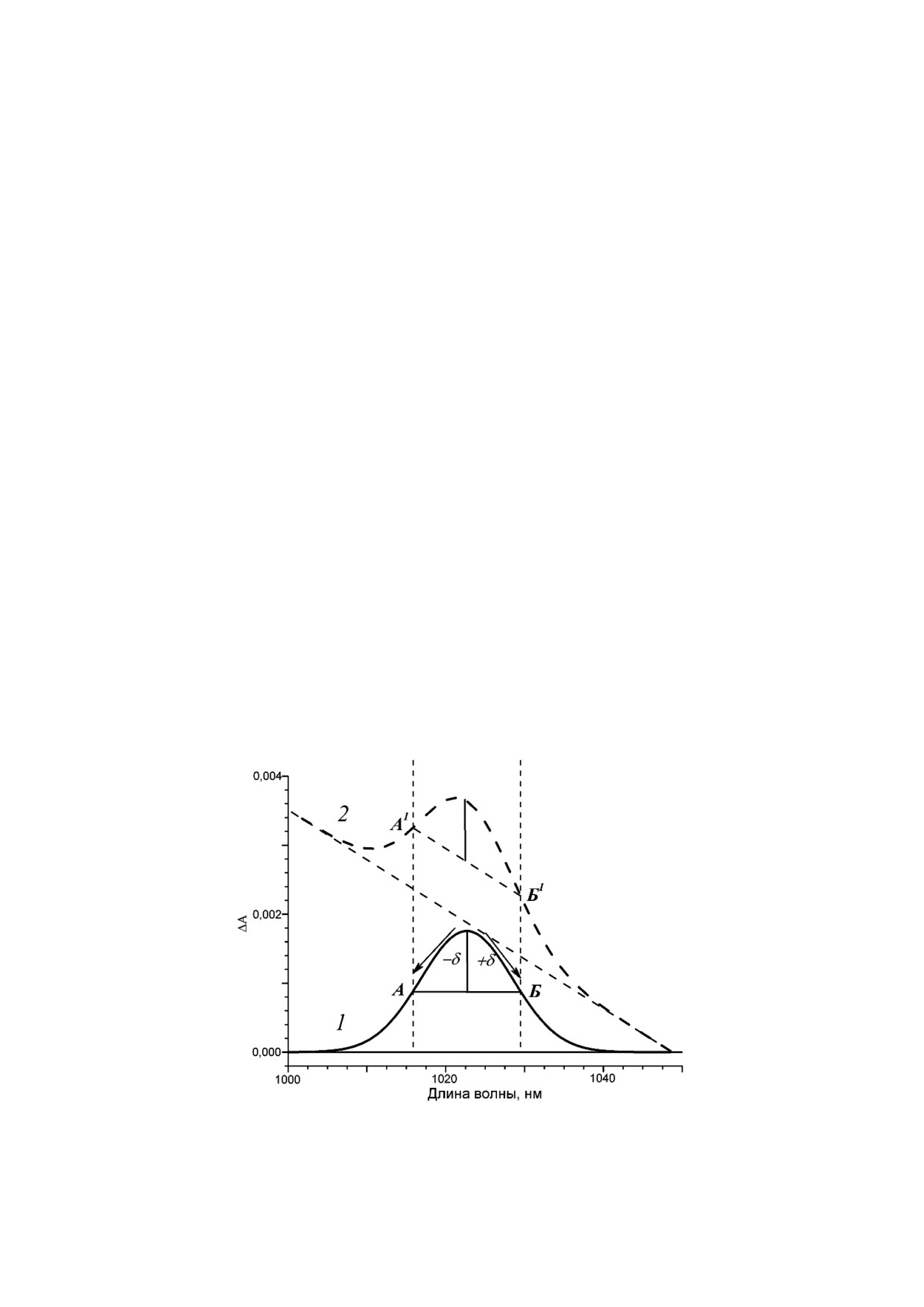
спектрах на разных задержках времени можно
тральная подставка линейна. Если на некоторой
наблюдать появление полос поглощения ком
произвольной задержке времени, обозначив
понентов электронного транспорта. Пусть по
подставку прямой линией, провести прямую,
глощение компонента проявляется в виде сим
параллельную подставке, то она пересечет гаус
метричной гауссовой полосы. Эволюцию этого
сову полосу поглощения в точках А1 и Б1, равно
спектрального компонента во времени отражает
отстоящих от максимума полосы поглощения.
кинетика изменений поглощения ΔA в макси
Легко увидеть, что на разных задержках времени
муме полосы поглощения. С другой стороны, в
отрезок А1Б1 всегда остается параллельным
окрестности максимума поглощения всегда
подставке, а когда полоса отсутствует, он лежит
можно найти точки А и Б, равноудаленные от
на подставке (рис. 1, кривая 2). Если из кинети
максимума на интервал длин волн δ (рис. 1, кри
ки ΔΑ в максимуме полосы поглощения вычесть
вая 1). Очевидно, что в силу симметричности
векторную полусумму кинетики ΔA в точках А1
полосы среднее значение длин волн в точках А
и Б1, то в итоговой векторной разности мы полу
и Б есть длина волны максимума поглощения,
чим одну и ту же долю спектральной полосы на
а среднее значение поглощения в точках А и Б
каждой задержке времени. В этом случае кине
есть середина отрезка АБ. Разность оптической
тика ΔGauss, построенная предложенным выше
плотности в максимуме поглощения и в середи
способом, является кинетикой доли спектраль
не отрезка АБ есть доля амплитуды гауссовой
ной полосы, независимо от того с какой кон
полосы, кинетика изменений которой ΔGauss
стантой скорости изменяются амплитуда и угол
такая же, как кинетика изменений поглощения
наклона линейной подставки.
ΔA в максимуме полосы, но кратно меньшая по
Если на некотором интервале длин волн
амплитуде.
спектральная подставка имеет кривизну, не ме
В области любой полосы поглощения всегда
няющуюся во времени, то эта кривизна не вли
вносят вклад другие спектральные компоненты,
яет на кинетику ΔGauss, а только смещает ее на
которые образуют меняющуюся во времени
шкале оптической плотности. И только в том
спектральную подставку. Поэтому на любой за
случае, когда кривизна подставки меняется во
держке времени оптическая плотность в макси
времени, она вносит искажения в кинетику
муме полосы поглощения есть сумма оптичес
ΔGauss, но искажения существенно меньшие,
кой плотности полосы поглощения и ее под
чем вклад всей спектральной подставки в целом.
ставки. Вследствие этого кинетика изменений
Модель последовательного переноса электро<
поглощения ΔA в максимуме полосы поглоще
на в реакционных центрах. В рамках последова
ния включает как кинетику самой полосы, так и
тельной модели разделения зарядов в комплек
кинетику изменения спектральной подставки.
сах реакционных центров пурпурных бактерий
Пусть на некотором интервале длин волн спек
под действием света на пикосекундной шкале
Рис. 1. Модель симметричной гауссовой полосы в дифференциальном спектре поглощения: 1 - без подставки, 2 - на фо
не линейной спектральной подставки
БИОХИМИЯ том 84 вып. 6 2019
830
ХАТЫПОВ и др.
времени происходит перенос электрона от воз
Из приведенных уравнений видно, что при
бужденного первичного донора P* на молекулу
последовательном переносе электрона концент
BA с константой скорости k1, а затем от молеку
рация P* с течением времени экспоненциально
лы BA на бактериофеофитин HA с константой
убывает, концентрация B– сначала нарастает до
скорости k2:
максимума, а затем убывает до нуля, а концент
рация H– нарастает до значения а.
k1
k2
P* —→ BA —→ HA.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В начальный момент времени [P*] = a, [B–] = 0,
[H–] = 0. Поскольку в элементарном акте разде
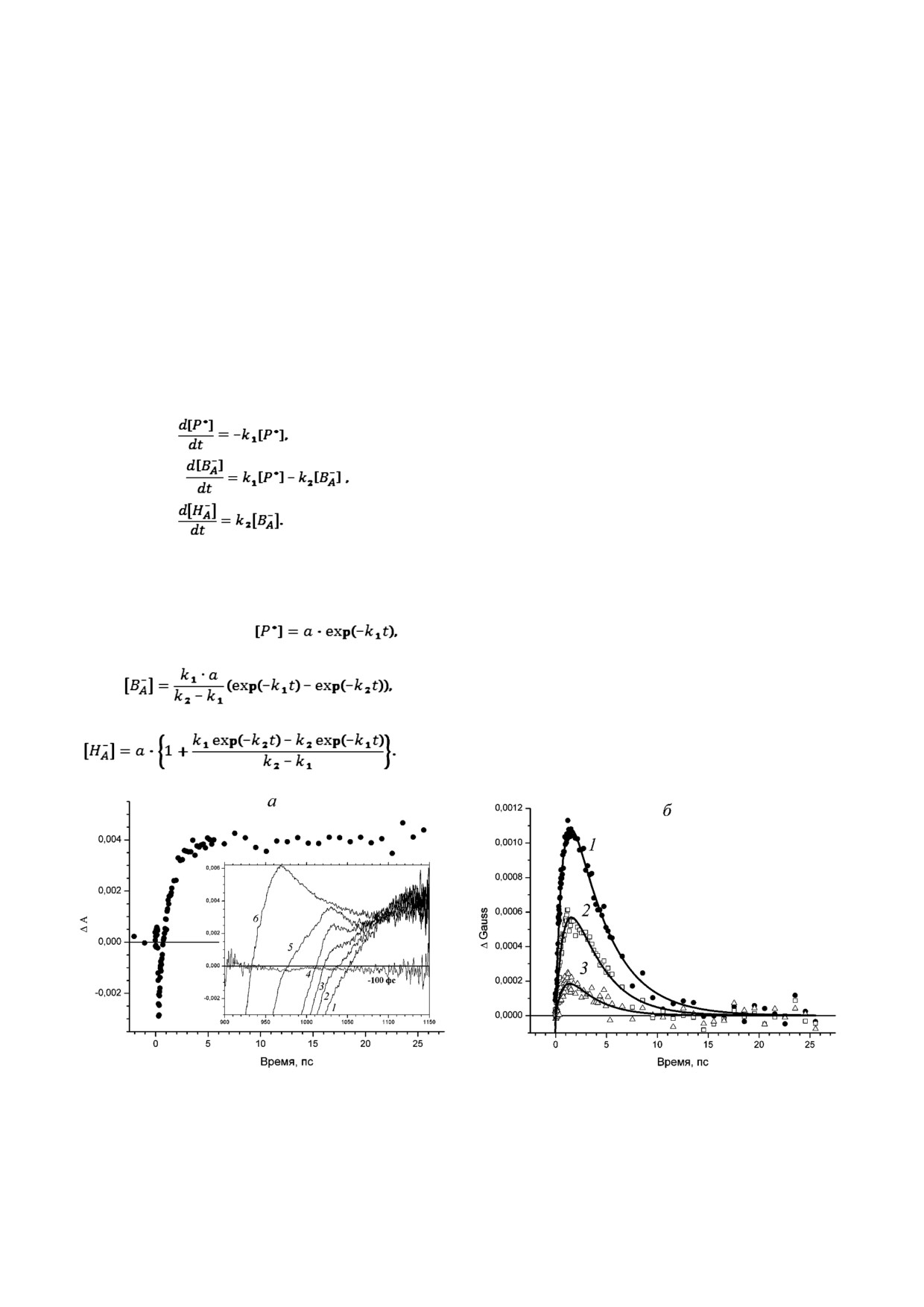
На рис. 2 представлены кинетика изменений
ления зарядов происходит перенос одного элек
поглощения ΔA и кинетика изменений доли
трона, то уравнение материального баланса:
амплитуды полосы ΔGauss в области поглоще
[P*] + [B–] + [H–] = a.
ния анион радикала B– в РЦ Rba. sphaeroides.
Система дифференциальных уравнений для
Кинетику изменений поглощения ΔA1020 (рис. 2, а)
компонентов переноса электрона имеет вид:
трудно приписать промежуточному акцептору
электрона, поскольку она не затухает на шкале
времени до нескольких наносекунд (данные не
приведены) [4]. Это связано с тем, что в области
1020 нм существенный вклад вносят другие
спектральные компоненты, которые образуют
подставку [17]. Действительно, на задержках
времени менее одной пикосекунды в этой кине
тике наблюдаются отрицательные изменения
Решение этой системы уравнений позволяет
поглощения, обусловленные стимулированным
получить уравнения кинетических кривых для
излучением при 935 нм, а на задержках времени
концентраций P*, B– и H–:
в десятки пикосекунд - положительные изме
нения, обусловленные поглощением катион
(1)
радикала первичного донора электрона.
Эволюция полосы поглощения при 1020 нм
происходит на фоне меняющейся во времени
(2)
спектральной подставки (рис. 2, а, вставка). На
задержках времени до десятков фемтосекунд эта
подставка обусловлена положительным погло
щением при 1100 нм и отрицательным поглоще
(3)
нием стимулированного излучения при 935 нм.
Рис. 2. а - Кинетика изменений поглощения ΔA при 1020 нм. Вставка: дифференциальные спектры поглощения, изме
ренные на задержках времени 0,3 пс (1), 0,53 пс (2), 0,85 пс (3), 1,5 пс (4), 3,7 пс (5) и 10,6 пс (6). б - Кинетика доли спект
ральной полосы ΔGauss при 1020 нм, построенная на интервалах длин волн δ = ± 15 нм (кривая 1), δ = ± 10 нм (кривая 2),
δ = ± 5 нм (кривая 3). Сплошными линиями показана аппроксимация кинетики уравнением 2
БИОХИМИЯ том 84 вып. 6 2019
ПЕРВИЧНЫЕ ПРОЦЕССЫ РАЗДЕЛЕНИЯ ЗАРЯДОВ В РЦ
831
На небольшом интервале длин волн в окрест
последовательного переноса электрона [8, 11].
ности 1020 нм сумму положительной и отрица
На рис. 3 сравниваются кинетика изменений
тельной гауссовых функций, образующих под
поглощения ΔA и кинетика изменения полосы
ставку, можно, в первом приближении, предста
ΔGauss в области QX перехода БХл при 600 нм.
вить, как прямую линию. На задержках времени
В кинетике изменений поглощения ΔA600 при
до десятков пикосекунд амплитуды этих спект
фемтосекундном возбуждении Р870 происходит
ральных полос уменьшаются, что приводит к
быстрое увеличение поглощения, обусловлен
изменению угла наклона подставки, и появляет
ное поглощением возбужденного первичного
ся поглощение катион радикала первичного до
донора электрона, которое, по видимому, пре
нора электрона, которое увеличивает амплитуду
вышает по амплитуде выцветающую полосу
подставки. При таком рассмотрении эволюция
поглощения при 600 нм (рис. 3, а). Далее на пи
полосы поглощения анион радикала B– проис
косекундной шкале времени в кинетике ΔA на
ходит на линейной подставке, угол наклона и
блюдается экспоненциальный спад со временем
амплитуда которой существенно изменяются во
жизни ~3,5 пс, обусловленный уменьшением
времени. На рисунке 2, б представлены кинети
концентрации возбужденного первичного доно
ки изменения доли спектральной полосы погло
ра электрона P*870. В то же время на вставке к
щения (ΔGauss) при 1020 нм, построенные для
рис. 3, а видно, что в дифференциальных спект
интервалов длин волн δ = ± 15 нм (кривая 1),
рах поглощения полоса при 600 нм полностью
δ = ± 10 нм (кривая 2) и δ = ± 5 нм (кривая 3).
выцветает за время ~0,1 пс. Таким образом, в
Уменьшение δ приводит к уменьшению соотно
кинетике изменений поглощения ΔA в области
шения сигнал-шум. Показано (рис 2, б), что вид
QX перехода БХл при 600 нм преобладают изме
построенной кинетики ΔGauss не зависит от
нения поглощения, обусловленные эволюцией
уровня экспериментального шума, неизбежного
возбужденного первичного донора электрона.
при измерении дифференциальных спектров
Кинетика изменения полосы ΔGauss при 600 нм,
поглощения. При всех значениях δ кинетика
построенная для интервала длин волн δ = ± 10 нм,
ΔGauss представляет собой кинетику, характер
отражает эволюцию спектральной полосы при
ную для промежуточного акцептора электрона.
600 нм без вклада возбужденного первичного
Однако уменьшение соотношения сигнал-шум
донора электрона (рис. 3, б). В кинетике наблю
приводит к увеличению погрешности в опреде
дается быстрое (~60 фс) выцветание QX полосы
лении временных констант. Эта кинетика удов
поглощения димера Р870, связанное с переходом
летворительно аппроксимируется уравнением (2)
первичного донора в возбужденное состояние.
для промежуточного акцептора электрона, ко
Далее на пикосекундной шкале времени и до
торый принимает электрон с константой ско
2,5 нс (рис. 3, б, вставка) амплитуда выцветания
рости 3,5 ± 0,2 пс и отдает электрон с константой
ΔGauss не изменяется, поскольку возбужден
скорости 0,8 ± 0,1 пс, что хорошо согласуется с
ный первичный донор электрона отдает элект
константами скоростей для молекулы BA в модели
рон в активную цепь кофакторов с квантовым
Рис. 3. а - Кинетика ΔA при 600 нм, измеренная от -2 до 27,5 пс. Сплошной линией показана экспоненциальная аппрок
симация кинетики. Вставка: дифференциальные спектры поглощения, измеренные на задержках времени -0,5 пс (1),
-0,15 пс (2), -0,05 пс (3), 0,05 пс (4) и 0,1 пс (5). б - Кинетика изменения доли спектральной полосы ΔGauss при 600 нм
(интервал длин волн δ ± 10 нм), построенная от -2 до 27,5 пс. На вставке показана кинетика ΔGauss при 600 нм, постро
енная от -20 до 2,5 нс
БИОХИМИЯ том 84 вып. 6 2019
832
ХАТЫПОВ и др.
выходом, близким к единице, и переходит в ка
блюдается быстрое увеличение поглощения,
тион радикальную форму P+870.
обусловленное поглощением возбужденного
Обращает на себя внимание, что на ранних
первичного донора электрона. На пикосекунд
задержках времени в кинетике ΔGauss наблю
ной шкале времени происходит уменьшение
дается дополнительный всплеск выцветания
поглощения, связанное как с уменьшением
(рис. 3, б, вставка). Поскольку уровень выцвета
концентрации возбужденного Р*870, так и с вы
ния полосы при 600 нм на наносекундной шка
цветанием полосы поглощения бактериофео
ле времени обусловлен поглощением двух моле
фитина HA (рис. 4, а, вставка) [18].
кул бактериохлорофилла специальной пары, то
В кинетике изменения полосы ΔGauss при
этот всплеск, по видимому, принадлежит треть
545 нм никаких быстрых процессов не наблюда
ей молекуле БХл. Если допустить, что диполь
ется (рис. 4, б). На пикосекундной шкале време
ные силы QX переходов четырех молекул бакте
ни происходит выцветание полосы при 545 нм,
риохлорофилла в РЦ соизмеримы, можно оце
обусловленное тем, что молекула HA принимает
нить, что эта третья молекула БХл выцветает
электрон и переходит в анион радикальную
только на ~20% в отличие от первых двух моле
форму H–. На ранних задержках времени в ки
кул. Всплеск выцветания достигает максимума
нетике выцветания полосы при 545 нм наблюда
на задержке времени 1,6 пс и затем спадает в те
ется слабый перегиб, который отличает ее от
чение ~10 пс (рис. 3, б). Кинетика этого допол
экспоненциальной кинетики. Действительно, эта
нительного компонента воспроизводит кинети
кинетика хорошо описывается уравнением (3)
ку спектральной полосы при 1020 нм (рис. 2, б)
для вторичного электронного акцептора с кон
и, по видимому, обусловлена выцветанием QX по
стантами скоростей 3,4 ± 0,2 и 0,7 ± 0,1 пс. Все
лосы поглощения молекулы BA при переходе ее
это позволяет заключить, что молекула HA про
в анион радикальную форму. Сравнение рис. 3, а
являет свойства не первичного, а вторичного ак
и 3, б позволяет заключить, что в кинетике ΔA
цептора электрона. Обращает на себя внимание
при 600 нм фемтосекундное выцветание полосы
то, что на задержках времени >15 пс можно на
поглощения первичного донора электрона и пи
блюдать появление слабой релаксации, связан
косекундное выцветание молекулы BA, связан
ной с переносом электрона на следующий хи
ное с переносом на нее электрона, скрыто изме
нонный акцептор QA.
нениями поглощения возбужденного первично
На вставке к рис. 4, б представлена кинетика
го донора электрона, а в кинетике ΔGauss эти
ΔGauss при 545 нм, построенная на шкале вре
изменения наблюдаются в явном виде.
мени до 2,5 нс. На этой шкале времени видно,
Кинетика изменений поглощения ΔA в об
что молекула HA также является промежуточ
ласти QX перехода бактериофеофитина при 545 нм
ным акцептором электрона от возбужденного
представлена на рис. 4, а. В этой кинетике на
донора Р* на хинонный акцептор QA. Кинетика
Рис. 4. а - Кинетика ΔA при 545 нм, измеренная от -2 до 27,5 пс. Вставка: дифференциальные спектры поглощения, из
меренные на задержках времени 0,1 пс (1), 1,0 пс (2), 2,0 пс (3), 3,0 пс (4) и 8,0 пс (5). б - Кинетика изменения доли спект
ральной полосы ΔGauss при 545 нм (интервал длин волн δ ± 10 нм), построенная от -2 до 27,5 пс. Сплошной линией по
казана аппроксимация кинетики уравнением 3. На вставке показана кинетика ΔGauss при 545 нм, построенная от -20 до
2,5 нс
БИОХИМИЯ том 84 вып. 6 2019
ПЕРВИЧНЫЕ ПРОЦЕССЫ РАЗДЕЛЕНИЯ ЗАРЯДОВ В РЦ
833
на вставке к рис. 4, б удовлетворительно описы
ках этих моделей мы построили модельные про
вается уравнением (2) с константами скоростей
фили концентраций состояний Р*, P+B– и P+H–.
3,5 ± 0,4 и 210 ± 10 пс.
Обнаружили, что в случае переноса электрона
Полученные в работе результаты позволяют
с последовательно убывающими константами
построить зависимость концентраций состоя
скоростей профиль концентрации состояния
ний P*, P+B– и P+H– от времени, то есть полу
P+B– в максимуме достигает не 20, а ~60%. Для
чить экспериментально измеренные кинетичес
модели переноса электрона с учетом обратных
кие кривые состояний с разделенными заряда
реакций заселенность состояния P+B– длитель
ми (рис. 5, а). Кинетика изменений поглощения
ное время сохраняется, и это состояние можно
ΔA при 930 нм (рис. 5, а, кривая 1), обусловлен
наблюдать на шкале времени до нескольких со
ная затуханием стимулированного излучения,
тен пикосекунд. И только для последовательной
отражает эволюцию возбужденного состояния
модели переноса электрона с константами 3, 1 и
первичного донора электрона. Кинетика спект
200 пс без учета обратных реакций получаемые
ральной полосы ΔGauss при 545 нм (рис. 5, а,
профили изменения концентраций состояний с
кривая 3) отражает эволюцию состояния P+H–.
разделенными зарядами имеют большое сход
Мы нормировали эти кинетические кривые на
ство с измеренной нами кинетикой слабых по
единицу, поскольку квантовый выход разделе
лос поглощения.
ния зарядов P+H– в РЦ близок к единице. Кине
Сравнение рис. 5, а и 5, б показывает, что ки
тика спектральной полосы ΔGauss при 1020 нм
нетика слабых полос поглощения ΔGauss при
(рис. 5, а, кривая 2) отражает эволюцию состоя
1020 и 545 нм и кинетика ΔA при 930 нм, по
ния P+B–. Эту кинетику мы нормировали на 0,2,
строенные без предварительного привлечения
так как QX полоса поглощения молекулы BA при
каких либо моделей, с хорошей точностью вос
600 нм выцветает только на ~20%.
производят профили концентраций состояний с
В то же время мы провели глобальный ана
разделенными зарядами для последовательной
лиз экспериментального массива данных в рам
четырехкомпонентной модели переноса элект
ках нескольких кинетических моделей: 1) моде
рона в РЦ.
ли переноса электрона с последовательно убы
Это позволяет утверждать, что кинетические
вающими константами скоростей k1 > k2 > k3 [9];
кривые, построенные предложенным в работе
2) модели переноса электрона с константами
алгоритмом, относительно мало искажены вкла
скоростей прямых реакций k12 < k23 > k34 с уче
дом сопутствующих спектральных компонентов
том реакций рекомбинации зарядов состояния
и достаточно точно отражают эволюцию кон
P+H– через молекулу BA с константами скоростей
центраций состояний с разделенными зарядами.
рекомбинации k21 = 1/12 пс и k32 = 1/21 пс [8, 10]
Полученные данные подтверждают результаты,
и 3) для последовательной модели переноса
полученные нами ранее [6, 13] и недавней рабо
электрона с константами скоростей реакций
ты Dominguez et al. [11], согласно которым в РЦ
k1 < k2 > k3 без учета обратных реакций [11]. В рам
происходит именно последовательный гомо
Рис. 5. а - Кинетика ΔA при 928 нм (кривая 1) и кинетики ΔGauss при 1020 нм (кривая 2) и 545 нм (кривая 3), построен
ные от -2 до 27,5 пс. б - Профили концентраций состояний P* (кривая 1), P+B– (кривая 2), P+H– (кривая 3), полученные
методом глобального анализа экспериментальных данных
БИОХИМИЯ том 84 вып. 6 2019
834
ХАТЫПОВ и др.
генный перенос электрона, включающий моле
Финансирование. Работа выполнена в рамках
кулу ВА в качестве первичного акцептора [19].
госзадания № АААА А17030110140 5 и при час
Это согласуется и с тем фактом, что и кинетика
тичной поддержке Российского фонда фунда
полосы при 1020 нм, и профиль концентрации
ментальных исследований (гранты № 17 00 00207
состояния P+B– в этой модели затухают на ин
и № 17 00 00209 КОМФИ).
тервале времени до ~10 пс. В то время, как в мо
Благодарности. Авторы выражают благодар
дели обратимого переноса электрона, заселен
ность А.Я. Шкуропатову и А.А. Забелину за по
ность состояния P+B– сохраняется на шкале
мощь в работе и обсуждение результатов.
времени до нескольких сотен пикосекунд [8, 10].
Конфликт интересов. Авторы заявляют об от
Полученные данные свидетельствуют также о
сутствии конфликта интересов.
том, что при возбуждении донора электрона им
Соблюдение этических норм. Настоящая
пульсами света длительностью ~30 фс в РЦ при
статья не содержит описания каких либо иссле
комнатной температуре происходит разделение
дований с использованием людей или животных
зарядов по не адиабатическому механизму.
в качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Kirmaier, C., and Holten, D. (1987) Primary photoche
sphaeroides, Biophys. J., 104, 2493-2502, doi: 10.1016/j.
mistry of reaction centers from the photosynthetic purple
bpj.2013.04.026.
bacteria, Photosynth. Res., 13, 225-260, doi: 10.1007/
11.
Dominguez, P., Himmelstoss, M., Michelmann, J.,
BF00029401.
Lehner, F., Gardiner, A.T, Cogdell, R.J., and Zinth, W.
2.
Woodbury, N.W., and Allen, J.P. (2004) In Anoxygenic pho>
(2014) Primary reactions in photosynthetic reaction cen
tosynthetic bacteria (Blankenship, R.E., Madigan, M.T.,
ters of Rhodobacter sphaeroides - time constants of the ini
and Bauer, C.E., eds) Kluwer Academic Publishers, New
tial electron transfer, Chem. Phys. Lett., 601, 103-109,
York, 527-557, doi: 10.1007/0 306 47954 0_24.
doi: 10.1016/j.cplett.2014.03.085.
3.
Holzapfel, W., Finkele, U., Kaiser, W., Oesterhelt, D.,
12.
Carter, B., Boxer, S.B., Holten, D., and Kirmaier, C.
Scheer, H., Stilz, H.U., and Zinth, W. (1989) Observation
(2012) Photochemistry of a bacterial photosynthetic reac
of a bacteriochlorophyll anion radical during the primary
tion center missing the initial bacteriochlorophyll electron
charge separation in a reaction center, Chem. Phys. Lett.,
acceptor, J. Phys. Chem. B, 116, 9971?9982, doi: 10.1021/
160, 1-7, doi: 10.1016/0009 2614(89)87543 8.
jp305276m.
4.
Arlt, T., Schmidt, S., Kaiser, W., Lauterwasser, C., Meyer, M.,
13.
Yakovlev, A.G., Shkuropatov, A.Y., and Shuvalov, V.A.
-
Scheer, H., and Zinth, W. (1993) The accessory bacterio
(2002) Nuclear wavepacket motion between P* and P+BA
chlorophyll: a real electron carrier in primary photosynthesis,
potential surfaces with subsequent electron transfer to HA
Proc. Natl. Acad. Sci. USA, 90, 11757-11761, doi: 10.1073/
in bacterial reaction centers. 1. Room temperature,
pnas.90.24.11757.
Biochemistry, 41, 2667-2674, doi: 10.1021/bi0101244.
5.
Kennis, J.T., Shkuropatov, A.Y., van Stokkum, I.H.M.,
14.
Shuvalov, V.A., Shkuropatov, A.Ya., Kulakova, S.M.,
Gast, P., Hoff, A.J., Shuvalov, V.A., and Aartsma, T.J.
Ismailov, M.A., and Shkuropatova, V.A. (1986) Photo
(1997) Formation of a long lived P+B– state in plant pheo
reactions of bacteriopheophytins and bacteriochlorophylls
phytin exchanged reaction centers of Rhodobacter
in reaction centers of Rhodopseudomonas sphaeroides and
sphaeroides R26 at low temperature, Biochemistry, 36,
Chloroflexus aurantiacus, Biochim. Biophys. Acta, 849,
16231-16238, doi: 10.1021/bi9712605.
337-346, doi: 10.1016/0005 2728(86)90145 3.
6.
Yakovlev, A.G., Shkuropatov, A.Y., and Shuvalov, V.A.
15.
Khatypov, R.A., Khristin, A.M., Fufina, T.Yu., and
(2000) Nuclear wavepacket motion producing a reversible
Shuvalov, V.A. (2017) An alternative pathway of light
charge separation in bacterial reaction centers, FEBS Lett.,
induced transmembrane electron transfer in photosynthetic
466, 209-212, doi: 10.1016/S0014 5793(00)01081 4.
reaction centers of Rhodobacter sphaeroides, Biochemistry
7.
Van Stokkum, I., Larsen, D., van Grondelle, R. (2004)
(Moscow), 82, 692-697, doi: 10.1134/S0006297917
Global and target analysis of time resolved spectra,
060050.
Biochim. Biophys. Acta, 1657, 82-104, doi: 10.1016/j.
16.
Snellenburg, J.J., Laptenok, S.P., Seger, R., Mullen, K.M.,
bbabio.2004.04.011.
and van Stokkum, I.H.M. (2012) Glotaran: a Java based
8.
Holzwarth, A.R., and Muller, M.G. (1996) Energetics and
graphical user interface for the R package TIMP, J. Stat.
kinetics of radical pairs in reaction centers from
Soft., 49, 1-22, doi: 10.18637/jss.v049.i03.
Rhodobacter sphaeroides: a femtosecond transient absorp
17.
Sporlein, S., Zinth, W., and Wachtveilt, J.
(1998)
tion study, Biochemistry, 35, 11820-11831, doi: 10.1021/
Vibrational coherence in photosynthetic reaction centers
bi9607012.
observed in the bacteriochlorophyll anion band, J. Phys.
9.
Kakitani, Y., Hou, A., Miyasako, Y., Koyama, Y., and
Chem. B, 102, 7492-7496, doi: 10.1021/jp9817473.
Nagae, H. (2010) Rates of the initial two steps of electron
18.
Heller, B., Holten, D., and Kirmaier, C. (1996) Effects of
transfer in reaction centers from Rhodobacter sphaeroides
Asp residues near the L side pigments in bacterial reaction
as determined by singular value decomposition followed by
centers, Biochemistry, 35, 15418-15427, doi: 10.1021/
global fitting, Chem. Phys. Lett., 492, 142-149, doi: 10.
bi961362f.
1016/j.cplett.2010.03.071.
19.
Shuvalov, V.A., and Duysens L.N.M. (1986) Primary elec
10.
Zhu, J., van Stokkum, I.H.M., Paparelli, L., Jones, M.R.,
tron transfer reactions in modified reaction centers from
and Groot, M.L. (2013) Early bacteriopheophytin reduc
Rhodopseudomonas sphaeroides, Proc. Natl. Acad. Sci. USA,
tion in charge separation in reaction centers of Rhodobacter
83, 1690-1694, doi: 10.1073/pnas.83.6.1690.
БИОХИМИЯ том 84 вып. 6 2019
ПЕРВИЧНЫЕ ПРОЦЕССЫ РАЗДЕЛЕНИЯ ЗАРЯДОВ В РЦ
835
THE METHOD OF EXTRACTING THE KINETICS
OF WEAK BANDS IN THE TRANSIENT ABSORPTION
SPECTRA OF Rhodobacter sphaeroides
REACTION CENTERS
R. A. Khatypov1*, A. M. Khristin1, L. G. Vasilyeva1,
and V. A. Shuvalov1,2
1 Institute of Basic Biological Problems, Russian Academy of Sciences,
Pushchino Scientific Center for Biological Research of the Russian
Academy of Sciences, 142290 Pushchino, Moscow Region, Russia;
E>mail: rgreen1@rambler.ru
2 Semenov Institute of Chemical Physics, Russian Academy of Sciences,
119991 Moscow, Russia
Received February 15, 2019
Revised March 21, 2019
Accepted March 21, 2019
An algorithm to plot kinetics of ion radical bands on the strong absorption background in transient absorption spec
tra of Rhodobacter sphaeroides reaction centers upon femtosecond excitation of primary electron donor is proposed.
The rising kinetics of transient absorption band at 1020 nm and the band bleaching kinetics at 545 nm built by the
proposed method are fitted adequately by kinetic equations for sequential electron transfer from an excited primary
donor to BA molecule of bacteriochlorophyll monomer and then to bacteriopheophytin HA molecule with time con
stants of 3.5 ± 0.2 and 0.8 ± 0.1 ps, respectively. In the kinetics of bacteriochlorophyll absorption band at 600 nm,
both an ultrafast bleaching of P870 dimer and slower bleaching of the BA monomer due to its transition to anion radi
cal are observed. The plotted kinetics of ion radical bands are consistent with the concentrations’ profiles of the
charge separated states fitted by the global target analysis of experimental data with use of the model of sequential
electron transfer in reaction centers.
Keywords: femtosecond spectroscopy, bacterial reaction centers, electron transfer
БИОХИМИЯ том 84 вып. 6 2019