БИОХИМИЯ, 2019, том 84, вып. 6, с. 849 - 858
УДК 577.22
ПРОТИВОПОЛОЖНЫЙ ЭФФЕКТ ИНГИБИТОРОВ
ROCK И Src КИНАЗЫ НА ЧУВСТВИТЕЛЬНОСТЬ КЛЕТОК
ЭУКАРИОТ К ИНВАЗИИ БАКТЕРИЙ Serratia grimesii
© 2019 Е.С. Божокина, О.А. Цаплина, С.Ю. Хайтлина*
Институт цитологии РАН, 194064 Санкт Петербург, Россия;
электронная почта: skhspb@gmail.com
Поступила в редакцию 20.09.2018
После доработки 22.11.2018
Принята к публикации 22.11.2018
Проникновение бактерий в клетки эукариот определяется сложным взаимодействием бактериальных и
клеточных факторов. Являясь частью окружающей среды, условно"патогенные бактерии разработали раз"
личные механизмы взаимодействия с рецепторами клеточной поверхности, такими как Е"кадгерин, интег"
рины или рецептор эпидермального фактора роста, что позволяет бактериям активировать компоненты
сигнальных путей эукариотических клеток и способствует проникновению бактерий в клетки, их выжива"
нию и внутриклеточному размножению. Исследуя механизмы проникновения условно"патогенных бакте"
рий Serratia grimesii в культивируемые эукариотические клетки, мы ранее установили, что обработка клеток
N"ацетилцистеином (NAC) увеличивает инвазию S. grimesii, и этот эффект коррелирует с увеличением
экспрессии Е"кадгерина. Так как NAC регулирует экспрессию ROCK киназы и Src киназы, целью данной
работы было выявление участия этих компонентов сигналинга в инвазии S. grimesii. Результаты ингибитор"
ного анализа показали, что чувствительность культивируемых клеток разного происхождения к инвазии
бактериями S. grimesii значительно увеличивается после обработки клеток специфическим ингибитором
протеинкиназы ROCK Y"27632. С другой стороны, введение в клетки Rho"ингибирующей siRNA или инги"
битора Src киназы Src"I1 уменьшало инвазию. Изменения чувствительности клеток к бактериям, вызван"
ные ингибиторами, коррелируют с соответствующими изменениями в экспрессии Е"кадгерина, увеличива"
ющейся при ингибировании ROCK киназы и уменьшающейся при ингибировании Src киназы. Эти резуль"
таты демонстрируют участие протеинкиназ ROCK и Src в инвазии бактерий S. grimesii в эукариотические
клетки и позволяют предположить, что в инвазию S. grimesii вовлечены компоненты других сигнальных пу"
тей, активность которых, по"видимому, увеличивается при Y"27632"зависимом ингибировании ROCK ки"
назы.
КЛЮЧЕВЫЕ СЛОВА: инвазия бактерий, Serratia grimesii, протеинкиназа ROCK, протеинкиназа Src, RhoA.
DOI: 10.1134/S0320972519060101
Система передачи сигнала эукариотических
системами секреции, которые позволяют достав"
клеток состоит из многочисленных сигнальных
лять факторы вирулентности непосредственно
путей, которые позволяют клеткам распозна"
в цитоплазму клетки"хозяина, чтобы управлять
вать изменения в окружающей среде, переда"
внутриклеточными процессами [6-9]. Напро"
вать внеклеточные сигналы в клетку и регулиро"
тив, условно"патогенные бактерии используют
вать поведение и взаимодействия клеток. Буду"
специфические поверхностные белки, которые
чи частью клеточного микроокружения, пато"
взаимодействуют с рецепторами клетки"хозяи"
генные и условно"патогенные внутриклеточные
на, такими как кадгерины [10, 11], интегрины
бактерии разработали различные механизмы
[12-14] или рецептор эпидермального фактора
для использования сигнальных путей эукарио"
роста [15, 16], что приводит к активации рецеп"
тических клеток для проникновения в клетку"
тор"опосредованных сигнальных каскадов и
хозяина, выживания и размножения в клетке
проникновению бактерий в клетки. Подобные
[1-5]. Для проникновения в эукариотические
механизмы могут быть использованы бактерия"
клетки многие патогенные бактерии обладают
ми, инвазивная активность которых скрыта до
тех пор, пока они не контактируют с поврежден"
Принятые сокращения: NAC - N"ацетилцистеин,
ными клетками или клетками пациента с ослаб"
КОЕ - колониеобразующие единицы.
ленным иммунитетом. Однако эти механизмы
* Адресат для корреспонденции.
еще недостаточно изучены.
849
850
БОЖОКИНА и др.
Грамотрицательные бактерии Serratia явля"
бриональной бычьей сыворотки («Thermo Fisher
ются условно"патогенными бактериями, спо"
Scientific Inc.», США) и 1% (v/v) NEAA (non"
собными вызывать внутрибольничные инфек"
essential amino acids) («Sigma"Aldrich», Герма"
ции или инфекции у пациентов с ослабленным
ния) без антибиотика при 37 °С в атмосфере 5%
иммунитетом [17, 18]. Ранее мы обнаружили,
CO2 до достижения 70-80% монослоя. Клетки
что бактерии S. grimesii, синтезирующие термо"
CaCo"2 и 3T3"SV40 культивировали в среде
лизиноподобную металлопротеиназу гримели"
DMEM («Биолот», Россия), а клетки SC5"MSC -
зин, специфически расщепляющую актин, мо"
в среде αМЕМ («Биолот», Россия), содержащих
гут проникать в эукариотические клетки. Оказа"
10% эмбриональной бычьей сыворотки без ан"
лось, также, что после трансформации плазми"
тибиотика, при 37 °С в атмосфере 5% CO2 до
дой, несущей ген гримелизина, неинвазивные
достижения 70-80% монослоя.
E. coli приобретают инвазивную активность [19,
Бактерии Serratia grimesii 30063 были получе"
20]. Мы также показали, что чувствительность
ны из «Немецкой коллекции микроорганизмов
клеток HeLa к инвазии S. grimesii и рекомбинан"
и клеточных культур» (DSMZ). Рекомбинан"
тными E. coli, трансформированными плазми"
тные E. coli SCS1, экспрессирующие гримели"
дой, несущей ген гримелизина, усиливалась при
зин, были получены, как описано ранее [25].
обработке клеток антиоксидантом N"ацетил"
Бактерии выращивали на питательном бульоне
цистеин (NAC) [20]. Этот эффект коррелировал
LB («Sigma"Aldrich», Германия), содержащем 1%"
с повышением экспрессии Е"кадгерина [20, 21],
ный пептон, 0,5% дрожжевого экстракта и 1%
трансмембранного белка, синтез которого уве"
NaCl (pH 7,0) при 37 °С с аэрацией в течение
личивается под воздействием N"ацетилцистеи"
времени, необходимого для появления в их
на [22]. С другой стороны, известно, что Е"кад"
экстрактах актин"гидролизующей активности.
герин участвует в интернализации бактерий
Затем суспензию бактерий в ростовой среде
Listeria monocytogenes [10]. Такая корреляция
осаждали на центрифуге MiniSpin («Eppendorf»,
указывает на то, что повышенная чувствитель"
Германия) при 13 000 об/мин в течение 10 мин.
ность клеток к S. grimesii может быть вызвана не"
Осадок ресуспендировали в среде DМЕМ («Би"
посредственным влиянием NAC на экспрессию
олот», Россия) без сыворотки.
гена Е"кадгерина. Однако известно, что NAC
Ингибиторный анализ. Чтобы ингибировать
также регулирует экспрессию других компонен"
сигнальный путь Rho/ROCK, исследуемые клет"
тов сигнальных путей, включая Rho/ROCK про"
ки в стандартной среде предварительно обраба"
теинкиназу [23] и Src протеинкиназу [22, 24],
тывали Y"27632 («Sigma"Aldrich», Германия) в
что, в свою очередь, может влиять на экспрес"
течение 30 мин при указанных концентрациях,
сию E"кадгерина. Поэтому в настоящей работе мы
используя в качестве исходного раствора 10 мМ
использовали специфические ингибиторы ROCK
раствор Y"27632 в диметисульфоксиде (ДМСО)
и Src протеинкиназ, чтобы выяснить, участвуют
(«Sigma"Aldrich», Германия). Для ингибирова"
ли эти протеинкиназы в проникновении S. gri
ния Src киназы клетки предварительно обраба"
mesii в клетки эукариот и сопровождается ли этот
тывали в течение 30 мин 10 мкМ Src"I1 (ингиби"
процесс регуляцией экспрессии Е"кадгерина.
тор киназы Src"1), («Sigma"Aldrich», Германия),
из расчета 1 мкл 10 мМ раствора Src"I1 в ДМСО
на 1 мл культуральной среды. Чтобы исключить
МЕТОДЫ ИССЛЕДОВАНИЯ
влияние ДМСО, содержащегося в растворах ин"
гибиторов, к контрольным клеткам добавляли
Культуры клеток, штаммы бактерий и условия
эквивалентное количество ДМСО. Затем кле"
культивирования. Клетки эпителиоидной карци"
точную среду заменяли средой DMEM без сыво"
номы шейки матки человека M"HeLa клон 11,
ротки и проводили эксперименты по иммунофлуо"
клетки колоректальной аденокарциномы чело"
ресценции и количественный анализ инвазии.
века CaCo"2, эмбриональные фибробласты мыши,
Количественный анализ инвазии. Эффектив"
трансформированные вирусом SV40 (3T3"SV40)
ность инвазии S. grimesii и E. coli SCS1 оценива"
и мезенхимные стволовые клетки человека
ли с помощью гентамицинового метода (gen"
SC5"MSC были получены из «Коллекции куль"
tamicin protection assay) [26] с небольшими изме"
тур клеток позвоночных» (Институт цитологии
нениями [20]. Бактерии S. grimesii в среде
РАН, Санкт"Петербург, Россия). Клетки линии
DMEM без сыворотки добавляли к клеткам в
HeLa"RhoA, экспрессирующие RhoA, были лю"
количестве не менее 100 бактерий на клетку. Об"
безно предоставлена доктором Т. Адамом
щее количество бактерий определяли по интен"
(Charite, Берлин, Германия). Клетки M"HeLa и
сивности поглощения суспензии при длине вол"
HeLa"RhoA культивировали на среде αМЕМ
ны 600 нм. Клетки с бактериями инкубировали
(«Биолот», Россия), содержащей 10% (v/v) эм"
в атмосфере 5% СО2 при 37 °С в течение 2 ч. Для
БИОХИМИЯ том 84 вып. 6 2019
ВЛИЯНИЕ ИНГИБИТОРОВ ROCK И Src КИНАЗЫ НА ИНВАЗИЮ БАКТЕРИЙ
851
определения количества внутриклеточных бак"
3 мин, 45 циклов амплификации (денатурация 1 мин
терий (инвазии) клетки промывали 1 мл раство"
при 94 °C, отжиг 1 мин при 60 °C для GAPDH и
ра трипсин"ЭДТА («Sigma"Aldrich», Германия),
64 °С для Е"кадгерина, элонгация 1 мин при 72 °С),
добавляли среду DMEM, содержащую гентами"
и финальная достройка цепи 10 мин при 72 °C.
цин (50 мкг/мл), и инкубировали в течение 2 ч
Продукты амплификации разделяли в 1,5%"ном
при 37 °С с качанием, чтобы уничтожить вне"
геле и визуализировали с помощью окраски бро"
клеточные бактерии. Затем клетки лизировали
мистым этидием («Sigma"Aldrich», Германия). Для
дезоксихолатом натрия («Sigma"Aldrich», Герма"
определения молекулярного веса продуктов, по"
ния), для чего к 400 мкл суспензии клеток добав"
лученных с помощью ПЦР, использовали стан"
ляли 200 мкл 4,5%"ного раствора дезоксихолата
дартный маркер молекулярного веса DNA"Lad"
натрия. Лизат, объемом 100 мкл, быстро титро"
der 100 bp («Thermo Fisher Scientific Inc.», США).
вали холодной средой LB до нужной концентра"
Трансфекция клеток препаратами siRNA.
ции и высевали по 100 мкл на чашки с LB"агаром
siRNA, направленная против RhoA человека, и
для подсчета колониеобразующих единиц (КОЕ).
контрольная siRNA, содержащая случайную
Результаты представляют собой среднее значе"
нуклеотидную последовательность, были полу"
ние КОЕ из трех независимых экспериментов. Зна"
чены из фирмы Santa Cruz (США). Трансфек"
чения выражены как среднее ± S.D. Различия
цию клеток M"HeLa этими siRNA проводили с
считались значительными на уровне р < 0,05.
использованием реагента трансфекции siRNA
Флуоресцентная микроскопия. Контрольные
(«Santa Cruz», США), в соответствии с инструк"
и инфицированные клетки в PBS фиксировали
циями производителя. Эффективность РНК"ин"
3,7%"ным параформальдегидом («Sigma"Aldrich»,
терференции (RNAi) была проверена Вестерн"
Германия) в PBS в течение 10 мин при комнат"
блотт анализом. Клетки собирали и лизировали
ной температуре, три раза промывали PBS, об"
детергент"содержащим буфером для лизиса кле"
рабатывали 0,1%"ным раствором Тритона X"100
ток RIPA (25 мM Tris"HCl pH 7,6, 150 мM NaCl,
в PBS в течение 5 мин и промывали PBS. Для ви"
1% NP"40, 1% дезоксихолат натрия, 0,1% SDS)
зуализации Ф"актина клетки окрашивали рода"
(«Thermo Fisher Scientific Inc.», США). Концент"
мин"фаллоидином («Sigma"Aldrich», Германия)
рацию белка в лизате определяли с помощью ре"
в течение 15 мин при 37 °С в темноте и промы"
актива Брэдфорда («Bio"Rad», США). 10 мкг
вали PBS. Затем клетки окрашивали DAPI
белка наносили на дорожку SDS"электрофореза
(«Sigma"Aldrich», Германия), 1 : 2000, в течение
и после окончания электрофореза переносили
5 мин для визуализации клеточных ядер и бакте"
на PVDF"мембрану для выявления Е"кадгерина
рий, промывали PBS и заключали в монтажную
Вестерн"блоттингом. Контролем нагрузки слу"
среду. Образцы анализировали с использованием
жил β"актин и GAPDH.
конфокального инвертированного микроскопа
Вестерн@блоттинг. Клеточные экстракты фрак"
Leica SP5 TCS («Leica», Германия).
ционировали с помощью SDS"электрофореза и
ПЦР@анализ. Экспрессию гена Е"кадгерина
переносили на мембрану из поливинилиден"
анализировали с помощью полуколичественной
дифторида (PVDF) в соответствии с протокола"
ОТ"ПЦР. Общую РНК экстрагировали из клеточ"
ми производителя («Bio"Rad», США). После ин"
ных лизатов с использованием набора PowerLy"
кубации с 5%"ным обезжиренным молоком в
zer RNA Isolation («MO BIO», США). Концент"
TBST"буфере (10 мМ Tris"HCl, рН 8,0, 150 мМ
рацию РНК измеряли с помощью спектрофото"
NaCl, 0,5% (v/v) Твин 20) в течение 60 мин
мера. кДНК получали с помощью обратной
мембрану промывали один раз TBST"буфером и
транскрипции, выполненной с использованием
инкубировали с антителами против RhoA
набора First Strand cDNA Synthesis Kit («Thermo
(«Santa Cruz», США), (1 : 1000) или Е"кадгерина
Fisher Scientific Inc.», США). Реакцию проводи"
(«Santa Cruz», США), (1 : 1000) и β"актина
ли согласно инструкциям производителя с до"
(«Abcam», США), (1 : 5000) или GAPDH («Abcam»,
бавлением oligo(dT) праймеров («Евроген», Рос"
США), (1 : 5000) при 4 °C в течение 12 ч. Затем
сия). Ген"специфические пары праймеров были
мембраны промывали 3 раза по 10 мин и инку"
созданы с использованием программного обес"
бировали 2 ч со вторыми антителами, специ"
печения BLAST"primer. Праймеры для Е"кадге"
фичными к IgG мыши или IgG кролика, конью"
рина: прямой 5'"CTGAGAACGAGGANAACG"3'
гированными с пероксидазой хрена (1 : 3000).
и обратный 5'"TTCACATCCAGCACATCC"3'.
Мембраны трижды промывали TBST"буфером и
Праймеры для GAPDH: прямой 5'"TGCACCA"
проявляли с помощью системы реагентов для
CCAACTGCTTAGC"3', и обратный 5'"GGCAT"
выявления результатов Вестерн"блоттинга
GGACTGTGGTCATGAG"3'.
Pierce™ ECL Western Blotting Substrate («Thermo
ПЦР реакции были оптимизированы следу"
Fisher Scientific Inc.», США) в соответствии с
ющим образом: первичное плавление при 94 °C
протоколами производителя.
БИОХИМИЯ том 84 вып. 6 2019
852
БОЖОКИНА и др.
Статистический анализ. Статистический ана"
борку актинового цитоскелета с небольшими
лиз полученных данных проводили с помощью
островками Ф"актина, оставшимися в цито"
однофакторного дисперсионного анализа (ANOVA)
плазме, и остроконечными актин"содержащи"
с пакетом анализа данных Excel. Различия счи"
ми филоподии"подобными протрузиями, обра"
тались достоверными на уровне р < 0,05.
зовавшимися на периферии клетки. Многочис"
ленные бактерии были расположены вдоль
протрузий и между протрузиями, а также интер"
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
нализованы в клетках, что указывает на то, что
инкубация клеток с Y"27632, предшествующая
Ингибирование активности ROCK киназы уве@
заражению клеток бактериями, существенно
личивает инвазию S. grimesii. Чтобы выяснить,
повышает чувствительность клеток М"HeLa к
участвует ли Rho/ROCK киназа в проникнове"
инвазии. Чтобы оценить этот эффект количе"
нии S. grimesii в эукариотические клетки, мы
ственно, инвазия бактерий в клетки М"HeLa, об"
сначала использовали конфокальную флуорес"
работанные ингибитором ROCK киназы Y"27632
центную микроскопию для визуализации внут"
в диапазоне 1-20 мкМ, была определена с по"
риклеточных бактерий в клетках М"HeLa, пред"
мощью гентамицинового метода [26].
варительно обработанных специфическим ин"
Как показано на рис. 2, а, 5-20 мкМ Y"27632
гибитором ROCK киназы Y"27632. Контроль"
увеличивали восприимчивость клеток M"HeLa
ные клетки M"HeLa (рис. 1, а) и клетки, обрабо"
к проникновению S. grimesii дозо"зависимым
танные 10 мкМ Y"27632 (рис. 1, б) в течение
образом, с максимальным увеличением инвазии
30 мин, инфицировали бактериями S. grimesii,
после инкубации клеток с 10 мкМ Y"27632.
как описано в разделе «Методы исследования».
Дальнейшее увеличение концентрации ингиби"
Сравнение контрольных клеток (рис. 1, а) и кле"
тора приводило к уменьшению чувствительнос"
ток, обработанных Y"27632 (рис. 1, б) показыва"
ти клеток к бактериям, что может быть связано с
ет, что ингибитор вызывал значительную раз"
дозо"зависимым влиянием Y"27632 на форми"
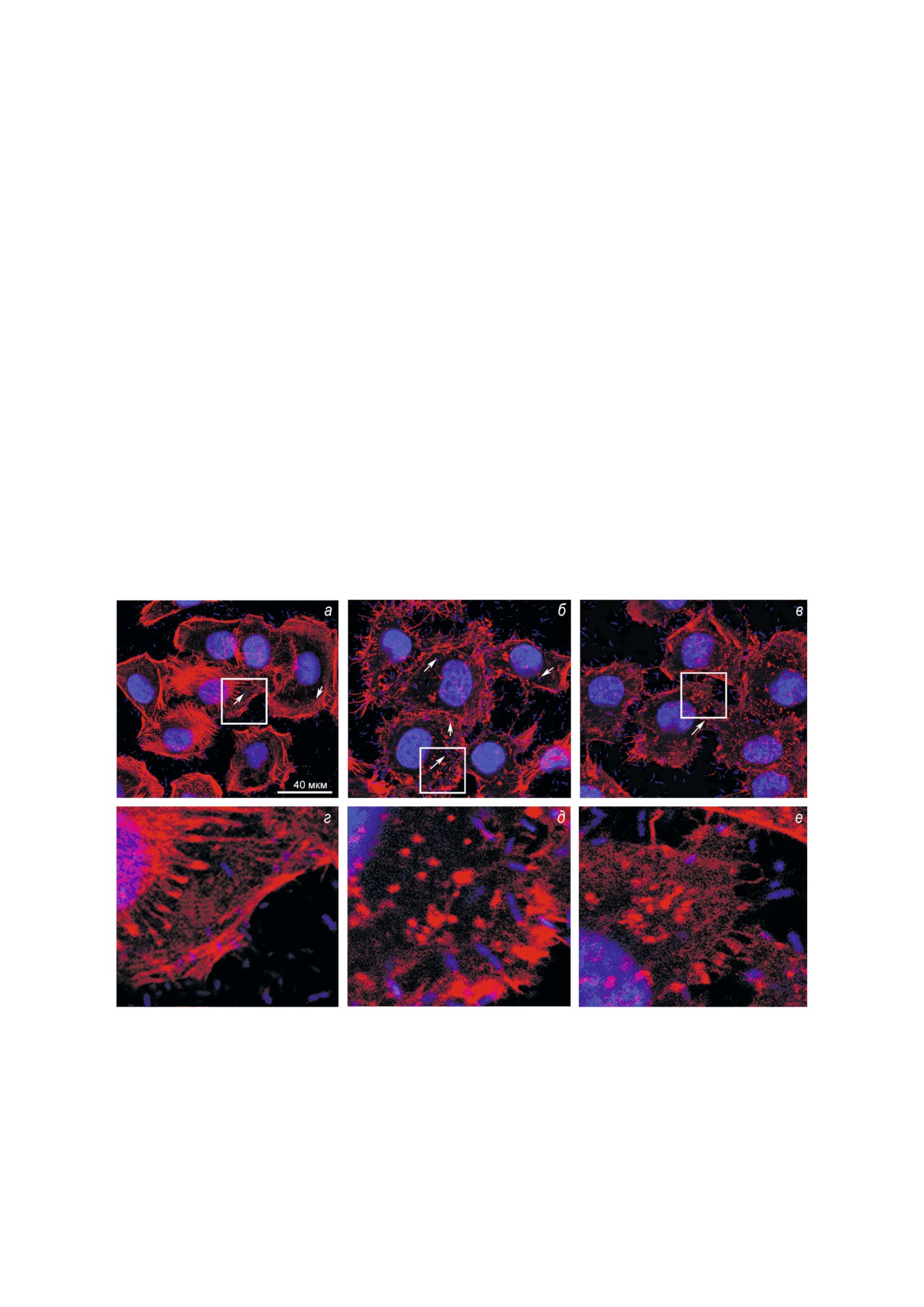
Рис. 1. Влияние ингибирования ROCK киназы и Src киназы на чувствительность клеток M"HeLa к инвазии бактериями
S. grimesii, по данным флуоресцентной конфокальной микроскопии. Клетки M"HeLa, выращенные на покровных стеклах (а),
предварительно инкубировали с 10 мкМ Y"27632 (б) или 10 мкМ Src"I1 (в) и инфицировали бактериями S. grimesii, как
описано в Материалах и Методах. Ф"актин окрашивали родамин"фаллоидином (красный); ядра и бактерии окрашивали
DAPI (синий). Внутриклеточные бактерии отмечены стрелами. В нижнем ряду рисунка (г, д, е) показаны увеличенные
изображения, соответствующие выделенным участкам изображений верхнего ряда рисунка (а, б, в).
biokhsm/
БИОХИМИЯ том 84 вып. 6 2019
ВЛИЯНИЕ ИНГИБИТОРОВ ROCK И Src КИНАЗЫ НА ИНВАЗИЮ БАКТЕРИЙ
853
рование клеточных выростов и перестройки ци"
Инкубация клеток с 10 мкМ Y"27632 приво"
тоскелета [27, 28], которые играют существен"
дила также к 2-3"кратному увеличению инва"
ную роль в бактериальной инвазии. В частнос"
зии в клетки CaCo"2 и 3T3"SV40, а также в клет"
ти, было показано, что повышение концентра"
ки линии HeLa"RhoA, содержащей ~20% RhoA"
ции Y"27632 над оптимальной приводит к раз"
трансфицированных клеток HeLa (рис. 2, б).
борке кортикального цитоскелета [27]. Когда
Кроме того, подобный эффект наблюдался, ког"
клетки вступают в контакт с бактериями, этот
да эти клетки были инфицированы рекомби"
процесс может уменьшить инвазию бактерий.
нантными E. coli SCS1, трансформированными
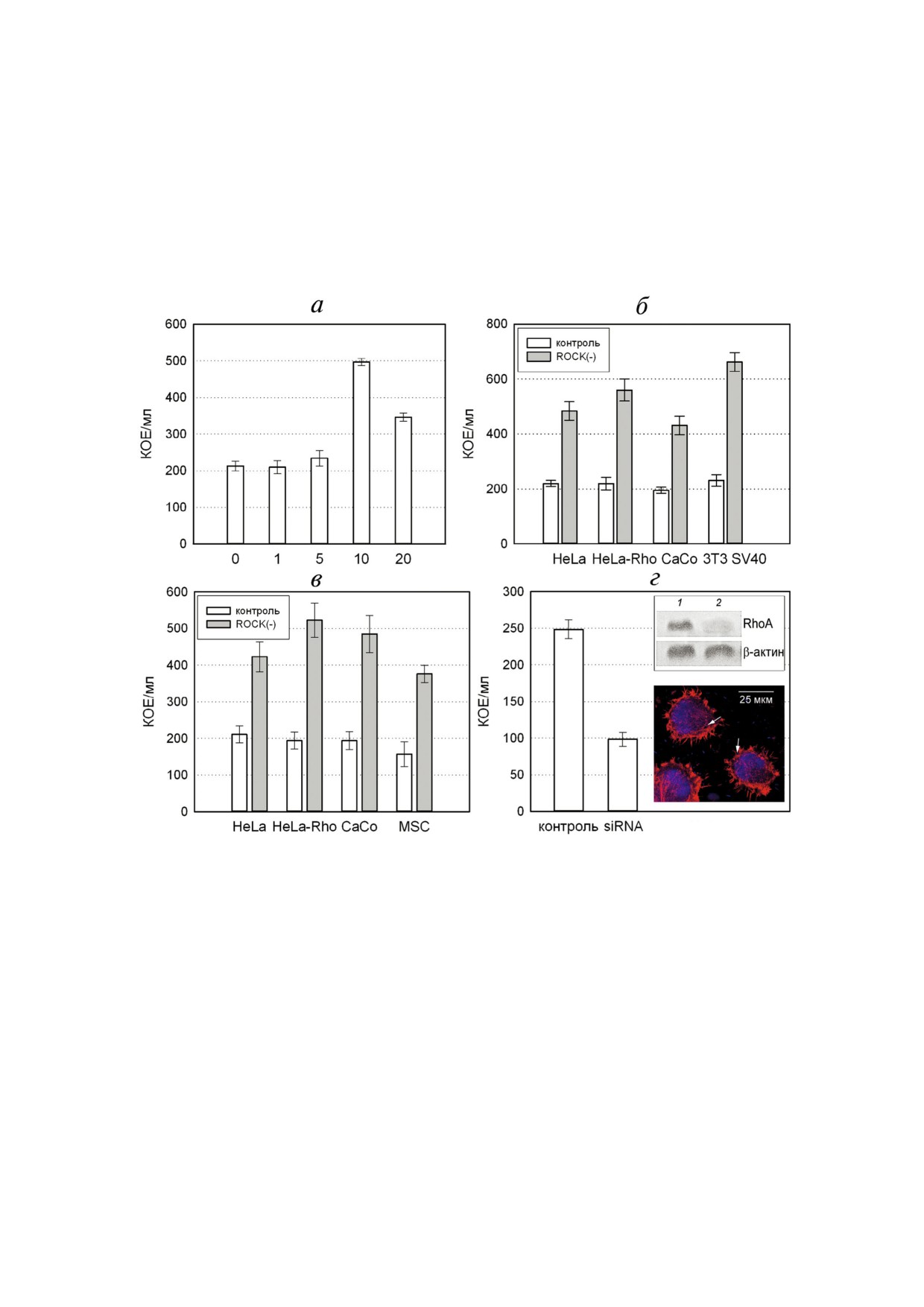
Рис. 2. Количественная оценка влияния ингибирования ROCK киназы на чувствительность клеток к инвазии бактерия"
ми S. grimesii. а - Влияние концентрации ингибитора Y"27632 на инвазию бактерий S. grimesii в клетки M"HeLa. б - Вли"
яние 10 мкМ Y"27632 на инвазию S. grimesii в клетки M"HeLa, HeLa"RhoA, CaCo"2 и 3T3"SV40. в - Влияние 10 мкМ
Y"27632 на инвазию бактерий E. coli SCS1, трансформированных геном гримелизина, в клетки M"HeLa, HeLa"RhoA,
CaCo"2 и SC5"MSC. Контроль - необработанные клетки, ROCK(") - клетки, предварительно инкубированные с 10 мкМ
Y"27632, как описано в «Методах исследования». Результаты представляют собой среднее значение КОЕ из трех незави"
симых экспериментов. Значения выражены как среднее ± S.D. Различия считались значительными на уровне р < 0,05.
г - Влияние ингибирования RhoA на чувствительность клеток M"HeLa к инвазии бактериями S. grimesii. Клетки M"HeLa,
выращенные на покровных стеклах, инкубировали с siRNA, направленной против RhoA, до того, как они были инфици"
рованы S. grimesii. Контроль - клетки трансфицированы siRNA, содержащей случайную нуклеотидную последователь"
ность. Представлены результаты количественной оценки влияния ингибирования siRNA на чувствительность клеток
M"HeLa к бактериальной инвазии. Значения выражены как среднее ± S.D. Различия считались значительными на уровне
р < 0,05. На врезках показаны результаты ингибирования RhoA по данным Вестерн"блот анализа и иммунофлуоресцен"
ции. Вестерн"блот анализ (верхняя врезка) - клетки M"HeLa, необработанные (дорожка 1) или трансфицированные
siRNA, направленной против RhoA (дорожка 2). β"актин служил контролем нагрузки. Иммунофлуоресцентный анализ
(нижняя врезка) - клетки M"HeLa, выращенные на покровных стеклах, инкубировали с siRNA, направленной против
RhoA, и инфицировали бактериями S. grimesii. Внутриклеточные бактерии S. grimesii показаны стрелками. Ф"актин окра"
шивали родамин"фаллоидином (красный); ядра и бактерии окрашивали DAPI (синий).
biokhsm/
БИОХИМИЯ том 84 вып. 6 2019
854
БОЖОКИНА и др.
плазмидой, несущей ген гримелизина, фактора
сколько меньше, чем действие Src"I1. Эти ре"
вирулентности S. grimesii [25] (рис. 2, в). Так как
зультаты указывают на то, что при ингибирова"
влияние ингибитора Y"27632 на инвазию бакте"
нии Srс киназы Y"27632"зависимое ингибирова"
рий во все исследованные клетки оказалось
ние ROCK практически не увеличивает уровень
одинаковым, в дальнейших экспериментах бы"
инвазии. Эти данные могут быть полезны в экс"
ли использованы только клетки линии M"HeLa.
периментах, направленных на понимание меха"
Поскольку активность ROCK киназы зави"
низма активации инвазии бактерий S. grimesii
сит от RhoA [29], другим способом ингибирова"
ингибитором Y"27632.
ния ROCK киназы может быть ингибирование
Ингибирование активности ROCK и Src по@раз@
активности RhoA. Чтобы проверить это предпо"
ному влияет на экспрессию и накопление E@кадге@
ложение, мы ингибировали RhoA путем предва"
рина. Увеличение инвазии S. grimesii в клетки
рительной инкубации клеток M"HeLa с siRNA,
направленной против RhoA. Контролем служи"
ли клетки M"HeLa, трансфицированные конт"
рольной siRNA, содержащей случайную нуклео"
тидную последовательность. По данным флуо"
ресцентной микроскопии, изменения морфоло"
гии клеток M"HeLa и цитоскелета, вызванные
введением siRNA, ингибирующей экспрессию
RhoA, были сходны с изменениями, вызванны"
ми действием специфического ингибитора
ROCK киназы Y"27632. Введение siRNA приво"
дило к округлению клеток, разборке актинового
цитоскелета в цитоплазме и образованию прот"
рузий на периферии клеток (рис. 2, г). Однако в
отличие от результатов действия Y"27632, в
клетках, обработанных анти"RhoA"siRNA, ко"
личество внутриклеточных бактерий было низ"
ким (рис. 2, г). Количественная оценка этого
эффекта показала, что ингибирование RhoA
уменьшило инвазию бактерий S. grimesii в клет"
ки M"HeLa в 2 раза (рис. 2, г), что указывает на
независимое от RhoA влияние ROCK киназы на
чувствительность клеток к инвазии бактериями.
Ингибирование активности Src киназы умень@
шает инвазию S. grimesii. Аналогично эффекту,
полученному в результате действия Y"27632, об"
работка клеток HeLa ингибитором Src киназы
Src"I1 (рис. 1, в) вызывала разборку актинового
цитоскелета в центральной части клеток и пере"
стройку актиновых структур на периферии.
Кроме того, наблюдалась локализация бактерий
на поверхности протрузий. Однако, в отличие от
эффекта, вызываемого Y"27632, предваритель"
ная обработка клеток ингибитором Src"I1 при"
водила к значительному уменьшению количе"
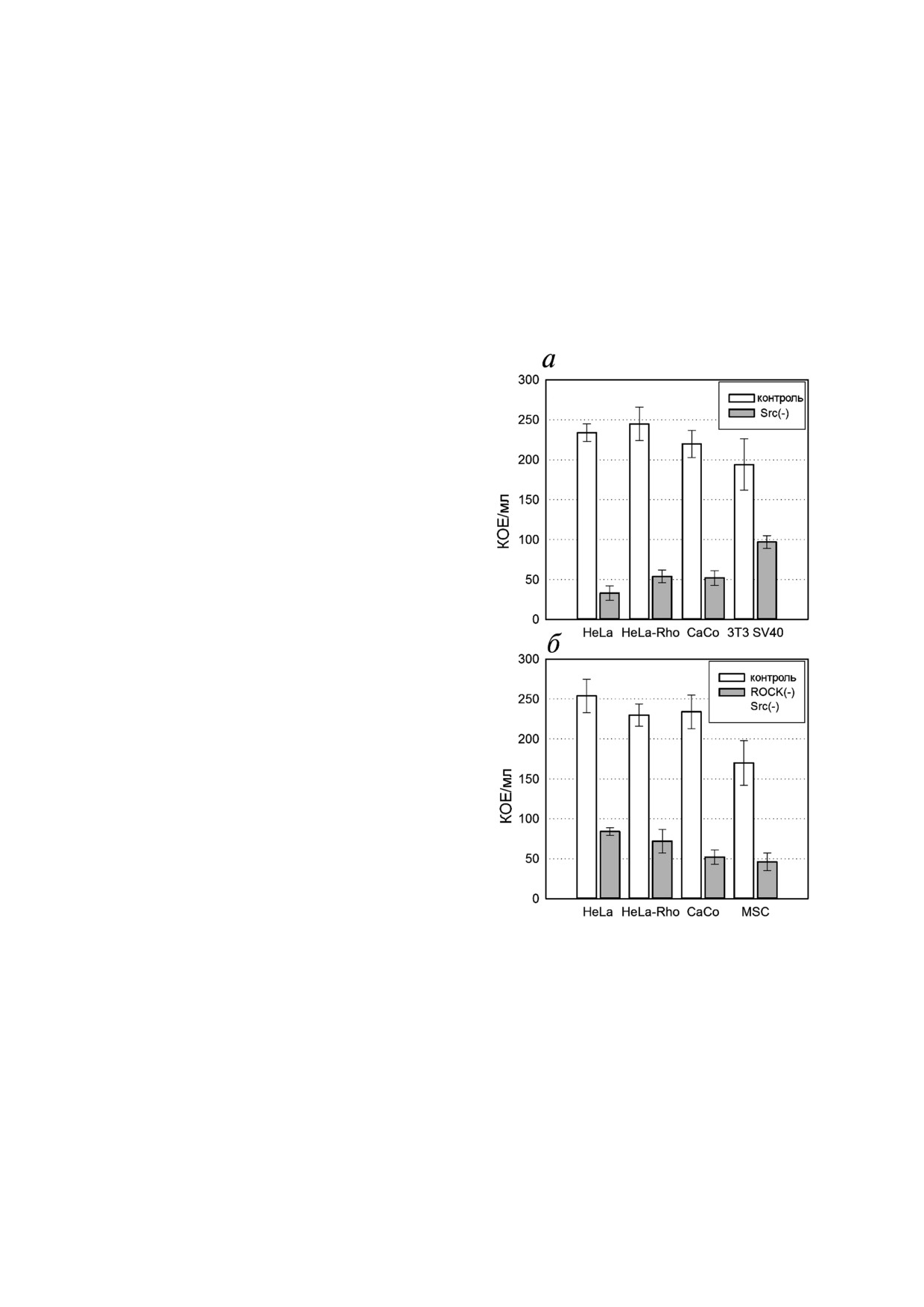
Рис. 3. Количественная оценка влияния ингибирования
ства внутриклеточных бактерий (рис. 1, в). Как
Src киназы и Src/ROCK киназ на чувствительность клеток
видно на рис. 3, а, инкубация клеток с ингиби"
к инвазии бактериями S. grimesii. а - Клетки инкубировали
тором Src"I1 уменьшала эффективность инва"
с 10 мкМ Src"I1 до того, как они были инфицированы S. gri
mesii. Контроль - необработанные клетки, Src(") - клетки,
зии бактерий S. grimesii в клетки M"HeLa, HeLa"
инкубированные с 10 мкМ Src"I1 перед заражением. б - Клет"
RhoA и CaCo"2 в 5-6 раз, а в клетки 3T3"SV40 -
ки инкубировали с Y"27632 и Src(") ингибитором до того,
примерно в 2 раза.
как они были инфицированы S. grimesii. Контроль - необ"
Ингибирование инвазии наблюдалось и тог"
работанные клетки, ROCK("), Src(") - клетки, инкубиро"
ванные со смесью 10 мкМ Src"I1 и 10 мкМ Y"27632 до за"
да, когда клетки предварительно инкубировали
ражения. Результаты представляют собой среднее значе"
со смесью ингибиторов Y"27632 и Src"I1, а затем
ние КОЕ из трех независимых экспериментов. Значения
инфицировали бактериями (рис. 3, б). Совмест"
выражены как среднее ± S.D. Различия считались значи"
ное действие двух ингибиторов было лишь не"
тельными на уровне р < 0,05
БИОХИМИЯ том 84 вып. 6 2019
ВЛИЯНИЕ ИНГИБИТОРОВ ROCK И Src КИНАЗЫ НА ИНВАЗИЮ БАКТЕРИЙ
855
HeLa и CaCo"2 коррелирует с повышенной
как известно, что это влияние зависит от мно"
экспрессией Е"кадгерина [21, 22]. С другой сто"
жества клеточных факторов, в том числе, типа
роны, недавние исследования показали, что как
клеток и состояния цитоскелета [35-37].
активация ROCK [30, 31], так и ее ингибирова"
ние [32] могут вызывать значительные измене"
ния в экспрессии генов. Показано также, что
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
повышенная экспрессия и/или повышенная ак"
тивность Src [33], а также ингибирование Src [34]
Инвазия эукариотических клеток условно"
вызывают изменения в экспрессии многочис"
патогенными бактериями активируется контак"
ленных генов. Поэтому мы использовали полу"
том поверхностных белков бактерий с рецепто"
количественный ОТ"ПЦР и Вестерн"блоттинг,
рами клетки"хозяина, такими как E"кадгерин,
чтобы выяснить, может ли обработка клеток
интегрин или рецептор эпидермального факто"
M"HeLa ингибиторами Y"27632 или Src"I1 вли"
ра роста [10, 12, 13, 16]. Изучая инвазию услов"
ять на экспрессию E"кадгерина. Изменение экс"
но"патогенных бактерий S. grimesii в эукариоти"
прессии Е"кадгерина наблюдали через 4 ч после
ческие клетки, мы показали, что индуцирован"
обработки клеток ингибиторами. На рис. 4, а
ное N"ацетилцистеином увеличение чувстви"
показано, что инкубация клеток M"HeLa с 10 мкМ
тельности этих клеток к инвазии коррелирует с
Y"27632 в течение 30 мин приводила к ~5"крат"
увеличением экспрессии E"кадгерина [20, 21].
ному увеличению экспрессии гена Е"кадгерина.
Кроме того, была обнаружена колокализация
Напротив, аналогичная инкубация клеток M"HeLA
S. grimesii с Е"кадгерином клеток 3T3 и 3T3"SV40,
с 10 мкМ Src"I1 уменьшала экспрессию гена
что указывает на возможное участие Е"кадгери"
Е"кадгерина примерно в 5 раз (рис. 4, а). В соот"
на в инвазии S. grimesii в эукариотические клет"
ветствии с этими результатами, вестерн"блоттинг
ки [38]. Результаты настоящей работы подтверж"
показал увеличение и уменьшение количества
дают это предположение, показывая, что инги"
E"кадгерина, вызванное ингибиторами Y"27632
бирование ROCK" или Src"ассоциированных
и Src"I1 соответственно (рис. 4, б). Эти результа"
сигнальных путей сильно влияет на чувствитель"
ты хорошо коррелируют с приведенными выше
ность клетки"хозяина к проникновению S. grimesii,
данными о влиянии ингибирования Y"27632 и
и эти эффекты коррелируют с соответствующи"
Src"I1 на инвазию S. grimesii. Для того, чтобы оп"
ми изменениями экспрессии Е"кадгерина. При
ределить влияние введения анти"RhoA"siRNA на
этом ингибирование Src подавляло как инвазию,
уровень экспрессии и количество Е"кадгерина,
так и экспрессию Е"кадгерина, тогда как инги"
необходимы дополнительные эксперименты, так
бирование ROCK усиливало как инвазию, так и
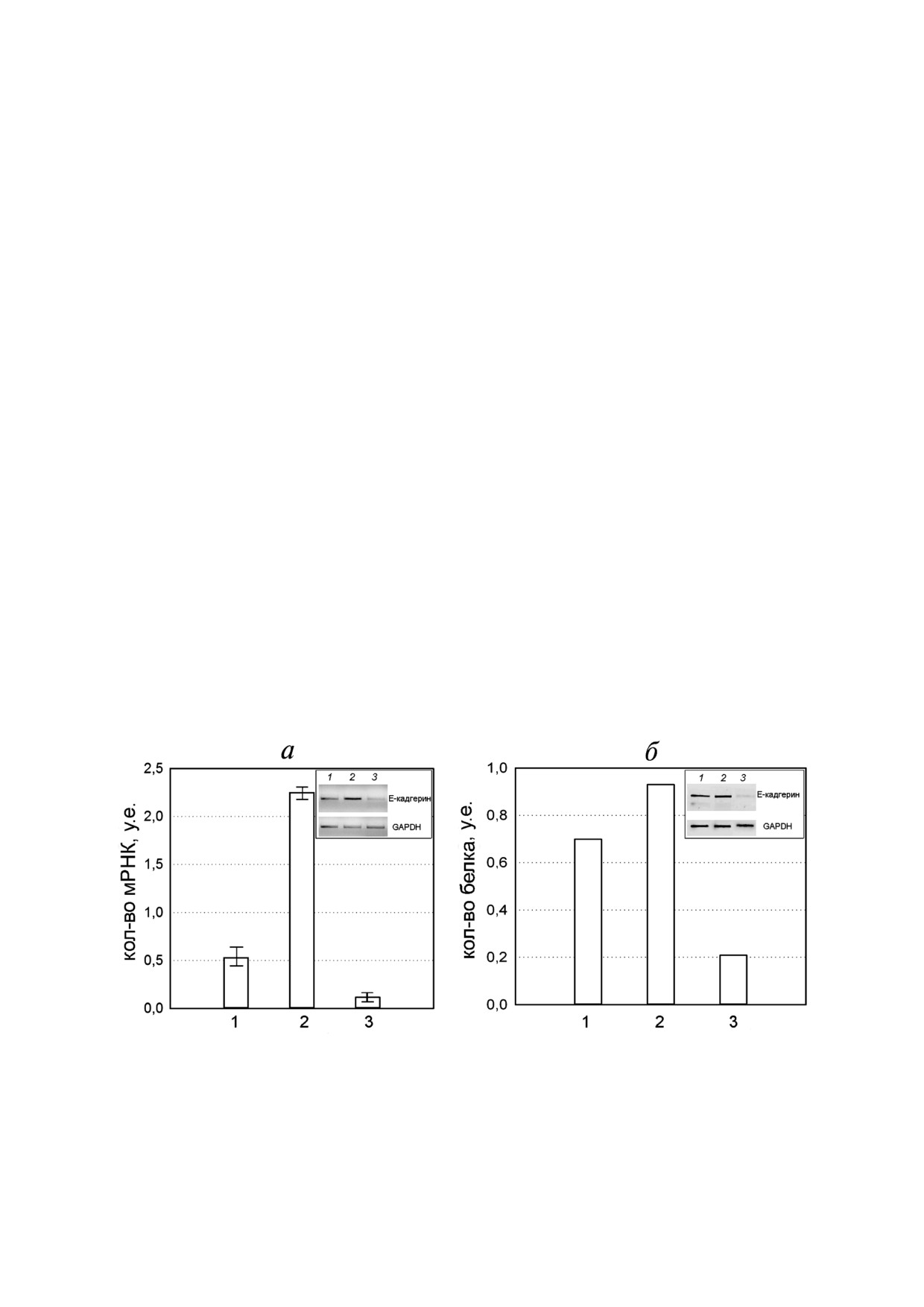
Рис. 4. Влияние ингибирования ROCK киназы и Src киназы на экспрессию и накопление E"кадгерина. Уровень экспрес"
сии E"кадгерина определяли полуколичественной ОТ"ПЦР (а) и тестировали с помощью Вестерн"блот анализа (б). 1 - Конт"
роль (необработанные клетки M"HeLa), 2 - клетки M"HeLa обрабатывали 10 мкМ Y"27632, 3 - клетки M"HeLa обраба"
тывали 10 мкМ Src"I1. Изменение экспрессии Е"кадгерина наблюдали через 4 ч, после обработки клеток ингибиторами.
Результаты ПЦР"анализа представляют собой среднее значение из трех независимых экспериментов. Значения выраже"
ны как среднее ± S.D. Различия считались значительными на уровне р < 0,05. На врезках представлены данные ОТ"
ПЦР электрофореза (а) и Вестерн"блот анализа (б), GAPDH служил в качестве внутреннего контроля
БИОХИМИЯ том 84 вып. 6 2019
856
БОЖОКИНА и др.
экспрессию Е"кадгерина. Эти изменения были
бирование активности ROCK с помощью Y"27632
одинаковыми для всех исследованных клеток, а
увеличивало вирулентность Listeria monocyto
также для двух видов бактерий, S. grimesii и ре"
genes при инвазии этих бактерий в мышиные
комбинантных E. coli SCS1, использованных в
фибробласты и гепатоциты [23] и усиление ин"
наших экспериментах.
вазии Yersinia enterocolitica в клетки HeLa [47].
Контакт бактерий с рецептором клеточной
При этом Y"27632"зависимое ингибирование
поверхности инициирует процесс интернализа"
ROCK увеличивало никотин"зависимую инва"
ции бактерий, включающий активацию сиг"
зию E. coli К1 в клетки микрососудистого эндо"
нальных каскадов и перестройки цитоскелета.
телия мозга человека, по"видимому, в результате
Src"киназа является одним из ключевых регуля"
перестроек актинового цитоскелета и морфоло"
торов этого процесса. Показано, что при интер"
гических изменений, индуцированных никотин"
нализации Listeria monocytogenes, индуцирован"
зависимым активированием PI3 киназы [45].
ной взаимодействием интерналина InlA c E"кад"
Предполагается, что ингибирование ROCK уве"
герином, Src киназа фосфорилирует кортактин,
личивало интернализацию Yersinia, способствуя
который необходим для активации комплекса
повышению активности Rac1 [47]. Было также
Arp2/3 и полимеризации актина [39]. Интерна"
показано влияние ингибирования ROCK на уве"
лизация листерии включает также Src"опосре"
личение экспрессии кадгерина и некоторых
дованное фосфорилирование тирозиновых ос"
специфических компонентов адгезии и подвиж"
татков E"кадгерина [40]. Показано также, что
ности нормальных и раковых клеток [31, 48, 49].
протеинкиназы Src семейства участвуют в опос"
Поэтому необходимы дальнейшие исследова"
редованной интегрином интернализации Staphy
ния, чтобы выяснить, участвуют ли эти или дру"
lococcus aureus [12], Yersinia pseudotuberculosis [13]
гие компоненты сигнальных путей в увеличении
и уропатогенной Escherichia coli [41]. Активация
эффективности инвазии S. grimesii в ответ на ин"
Src протеинкиназы была также выявлена при
гибирование ROCK киназы с помощью Y"27632.
инвазии Escherichia coli K1 в клетки микрососу"
Таким образом, проведенный нами ингиби"
дистого эндотелия мозга человека, которая со"
торный анализ выявил участие протеинкиназ
провождалась перестройками цитоскелета. Как
Src и ROCK в проникновении условно"патоген"
инвазия E. coli, так и индуцированная E. coli пе"
ных бактерий Serratia grimesii в эукариотические
рестройка актиновых филаментов блокирова"
клетки. Результаты нашей работы показали, что
лись ингибиторами Src [42]. Исходя из этих дан"
ингибирование Src киназы с помощью Src"I1 при"
ных, мы предполагаем, что Src киназа не только
водит к уменьшению инвазии бактерий и экспрес"
влияет на экспрессию гена E"кадгерина, но и
сии E"кадгерина. Инвазия бактерий уменьша"
участвует в регуляции перестроек цитоскелета,
лась также при ингибировании синтеза RhoA,
необходимых для интернализации S. grimesii. Та"
активатора ROCK киназы, с помощью анти"
ким же образом ингибирование инвазии S. grime
RhoA"siRNA. Напротив, ингибирование ROCK
sii, вызванное введением в клетки анти"RhoA"
киназы с помощью Y"27632 усиливало инвазию
siRNA, может быть связано с отсутствием RhoA"
и экспрессию E"кадгерина. Полученные резуль"
зависимых перестроек цитоскелета. Поскольку
таты указывают на то, что в инвазии S. grimesii
RhoA является активатором ROCK киназы [29],
наряду с Src" и ROCK"опосредованными сиг"
уменьшение инвазии бактерий S. grimesii после
нальными путями участвуют компоненты дру"
введения в клетки анти"RhoA"siRNA, можно
гих сигнальных путей, активность которых, по"
также объяснить тем, что при ингибировании
видимому, увеличивается при Y"27632"зависи"
синтеза RhoA ROCK киназа не активируется.
мом ингибировании ROCK киназы.
С другой стороны, увеличение инвазии S. gri
mesii в результате обработки клеток ингибито"
Финансирование. Работа выполнена при
ром ROCK Y27632 оказалось весьма неожидан"
поддержке Российского фонда фундаменталь"
ным. ROCK играет важную роль в регуляции пе"
ных исследований (грант № 17"04"00558).
рестроек актомиозинового цитоскелета, активи"
Конфликт интересов. Авторы заявляют об от"
рованных RhoA, таких как образование стресс"
сутствии конфликта интересов.
фибрилл, подвижность клеток, сокращение и
Соблюдение этических норм. Настоящая
адгезия [43, 44], и ингибирование ROCK часто
статья не содержит описания каких"либо иссле"
приводит к ингибированию бактериальной ин"
дований с использованием животных или учас"
вазии [45, 46]. Вместе с тем, показано, что инги"
тием людей в качестве объектов.
БИОХИМИЯ том 84 вып. 6 2019
ВЛИЯНИЕ ИНГИБИТОРОВ ROCK И Src КИНАЗЫ НА ИНВАЗИЮ БАКТЕРИЙ
857
СПИСОК ЛИТЕРАТУРЫ
1.
Ham, H., Sreelatha, A., and Orth, K. (2011) Manipulation
experiments to current practice, Clin. Microbiol. Rev., 24,
of host membranes by bacterial effectors, Nat. Rev.
755-791, doi: 10.1128/CMR.00017"11.
Microbiol., 9, 635-646, doi: 10.1038/nrmicro2602.
19.
Bozhokina, E.S., Tsaplina, O.A., Efremova, T.N., Kever, L.V.,
2.
Bhavsar, P.A., Guttman, J.A., and Finlay, B.B. (2007)
Demidyuk, I.V., Kostrov, S.V., Adam, T., Komissarchik, Y.Y.,
Manipulation of host-cell pathways by bacterial patho"
and Khaitlina, S.Y. (2011) Bacterial invasion of eukaryotic
gens, Nature, 449, 827-834, doi: 10.1038/nature06247.
cells can be mediated by actin"hydrolysing metalloproteases
3.
Carabeo, R. (2011) Bacterial subversion of host actin
grimelysin and protealysin, Cell Biol. Int., 35, 111-118,
dynamics at the plasma membrane, Cell. Microbiol., 13,
doi: 10.1042/CBI20100314.
1460-1469, doi: 10.1111/j.1462"5822.2011.01651.x.
20.
Bozhokina, E., Vakhromova, E., Gamaley, I., and
4.
Personic, N., Barlocher, K., Finsel, I., and Hilbi, H.
Khaitlina, S. (2013) N"acetylcysteine increases suscepti"
(2016) Subversion of retrograde trafficking by translocated
bility of HeLa cells to bacterial invasion, J. Cell. Biochem.,
pathogen effectors, Trends Microbiol., 24, 450-462, doi: 10.
114, 1568-1574, doi: 10.1002/jcb.24498.
1016/j.tim.2016.02.003.
21.
Bozhokina, E., Khaitlina, S., and Gamaley, I. (2015)
5.
Weber, M.M., and Faris, R. (2018) Subversion of the endo"
Dihydrolipoic but not alpha"lipoic acid affects suscepti"
cytic and secretory pathways by bacterial effector proteins,
bility of eukaryotic cells to bacterial invasion, Biochem.
Front. Cell Dev. Biol., 6, 1-17, doi: 10.3389/fcell.2018.
Biophys. Res. Commun., 460, 697-702, doi: 10.1016/j.bbrc.
00001.
2015.03.092.
6.
Rosenshine, I., and Finlay, B.B. (1993) Exploitation of
22.
Parasassi, T., Brunelli, R., Bracci"Laudiero, L., Greco, G.,
host signal transduction pathways and cytoskeletal func"
Gustafsson, A.C., Krasnowska, E.K., Lundeberg, J.,
tions by invasive bacteria, BioEssays, 15, 17-24, doi: 10.
Lundeberg, T., Pittaluga, E., Romano, M.C., and
1002/bies.950150104.
Serafino, A. (2005) Differentiation of normal and cancer
7.
Cossart, P., and Sansonetti, P.J. (2004) Bacterial invasion:
cells induced by sulfhydryl reduction: biochemical and
the paradigms of enteroinvasive pathogens, Science, 304,
molecular mechanisms, Cell Death Differ., 12, 1285-1296,
242-248, doi: 10.1126/science.1090124.
doi: 10.1038/sj.cdd.4401663.
8.
Boumart, Z., Velge, P., and Wiedemann, A.
(2014)
23.
Kirchner, M., and Higgins, D.E. (2008) Inhibition of
Multiple invasion mechanisms and different intracellular
ROCK activity allows InlF"mediated invasion and
behaviors: a new vision of Salmonella-host cell interac"
increased virulence of Listeria monocytogenes, Mol.
tion, FEMS Microbiol. Lett., 361, 1-7, doi: 10.1111/1574"
Microbiol., 68, 749-767, doi: 10.1111/j.1365"2958.2008.
6968.12614.
06188.x.
9.
Mattock, E., and Blocker, A.J. (2017) How do the viru"
24.
Krasnowska, E.K., Pittaluga, E., Brunati, A.M., Brunelli, R.,
lence factors of Shigella work together to cause disease?
Costa, G., De Spirito, M., Serafino, A., Ursini, F., and
Front. Cell Infect. Microbiol., 64, doi: 10.3389/fcimb.
Parasassi, T. (2008) N"acetyl"l"cysteine fosters inactivation
2017.00064.
and transfer to endolysosomes of c"Src, Free Radic. Biol.
10.
Bonazzi, M., Lecuit, M., and Cossart, P. (2009) Listeria
Med., 45, 1566-1572, doi: 10.1016/j.freeradbiomed.2008.
monocytogenes internalin and E"cadherin: from structure to
09.012.
pathogenesis, Cell. Microbiol., 11, 693-702, doi: 10.1111/
25.
Bozhokina, E., Khaitlina, S., and Adam, T.
(2008)
j.1462"5822.2009.01293.x.
Grimelysin, a novel metalloprotease from Serratia grimesii,
11.
Ribet, D., and Cossart, P. (2015) How bacterial pathogens
is similar to ECP32, Biochem. Biophys. Res. Commun., 367,
colonize their hosts and invade deeper tissues, Microb.
888-892, doi: 10.1016/j.bbrc.2008.01.003.
Infect., 17, 173-183, doi: 10.1016/j.micinf.2015.01.004.
26.
Prouty, A.M., and Gunn, J.S. (2000) Salmonella enterica
12.
Agerer, F., Antje, M., Ohlsen, K., and Hauck, C.R. (2003)
serovar typhimurium invasion is repressed in the presence
Integrin"mediated Invasion of Staphylococcus aureus into
of bile, Infect. Immun., 68, 6763-6769.
human cells requires Src family protein"tyrosine kinases,
27.
Svoboda, K.K.H., Moessner, P., Field, T., and Acevedo, J.
J. Biol. Chem., 278, 42524-42531, doi: 10.1074/jbc.
(2004) ROCK inhibitor (Y27632) increases apoptosis and
M302096200.
disrupts the actin cortical mat in embryonic avian corneal
13.
Eitel, J., Heise, T., Thiesen, U., and Dersch, P. (2005) Cell
epithelium, Dev. Dyn., 229, 579-590, doi: 10.1002/dvdy.
invasion and IL"8 production pathways initiated by YadA
20008.
of Yersinia pseudotuberculosis require common signalling
28.
Heng, B.C. (2009) Effect of Rho"associated kinase
molecules (FAK, c"Src, Ras) and distinct cell factors, Cell.
(ROCK) inhibitor Y"27632 on the post"thaw viability of
Microbiol., 7, 63-77, doi: 10.1111/j.1462"5822.2004.
cryopreserved human bone marrow"derived mesenchymal
00434.x.
stem cells, Tissue Cell, 41, 376-380, doi: 10.1016/j.tice.
14.
Scibelli, A., Roperto, S., Manna, L., Pavone, L.M., Tafuri, S.,
2009.01.004.
Della Morte, R., and Staiano, N. (2007) Engagement of
29.
Riento, K., and Ridley, A.J. (2003) ROCKs: multifunc"
integrins as a cellular route of invasion by bacterial
tional kinases in cell behaviour, Nat. Rev. Mol. Cell Biol., 4,
pathogens, Veter. J., 173, 482-491, doi: 10.1016/j.tvjl.
446-456, doi: 10.1038/nrm1128.
2006.01.010.
30.
Rajakyla, E.K., and Vartiainen, M.K. (2014) Rho, nuclear
15.
Wiedemann, A., Mijouin, L., Ayoub, M.A., Barilleau, E.,
actin, and actin"binding proteins in the regulation of tran"
Canepa, S., Teixeira"Gomes, A.P., Le Vern, Y., Rosselin, M.,
scription and gene expression, Small GTPases, 5, e27539,
Reiter, E., and Velge, P. (2016) Identification of the epider"
doi: 10.4161/sgtp.27539.
mal growth factor receptor as the receptor for Salmonella
31.
Rath, N., Morton, J.P., Julian, L., Helbig, L., Kadir, S.,
Rck"dependent invasion, FASEB J., 304, 4180-4191, doi: 10.
McGhee, E.J., Anderson, K.I., Kalna, G., Mullin, M.,
1096/fj.201600701R.
Pinho, A.V., Rooman, I., Samuel, M.S., and Olson, M.F.
16.
Mambu, J., Virlogeux"Payant, I., Holbert, S., Grepinet, O.,
(2017) ROCK signaling promotes collagen remodeling to
Velge, P., and Wiedemann, A. (2017) An updated view on
facilitate invasive pancreatic ductal adenocarcinoma tumor
the Rck invasin of Salmonella: still much to discover, Front.
cell growth, EMBO Mol. Med., 9, 198-218, doi: 10.15252/
Cell. Infect. Microbiol., 7, 500, doi 10.3389/fcimb.2017.
emmm.201606743.
00500.
32.
Piltti, J., Bygdell, J., Fernandez"Echevarria, C., and
17.
Grimont, F., and Grimont, P.A.D. (2006) The genus
Marcellino, D. (2017) Rho"kinase inhibitor Y"27632 and
Serratia, Prokaryotes, 6, 219-244, doi: 10.1007/0"387"
hypoxia synergistically enhance chondrocytic phenotype
30746"x_11.
and modify S100 protein profiles in human chondrosarcoma
18.
Mahlen, S.D. (2011) Serratia infections: from military
cells, Sci. Rep., 7, 3708, doi: 10.1038/s41598"017"03958"5.
8 БИОХИМИЯ том 84 вып. 6 2019
858
БОЖОКИНА и др.
33. Yeatman, T.J. (2010) A renaissance for Src, Nat. Rev.
ted uropathogenic Escherichia coli, PLoS Pathog., 3, e100,
Cancer, 4, 470-480, doi: 10.1038/nrc1366.
doi: 10.1371/journal.ppat.0030100.
34. Kruewel, T., Schenone, S., Radi, M., Maga, G.,
42. Liu, W., Zhao, W.D., Yan, J.C., Ren, Z.Y., Fang, W.G.,
Rohrbeck, A., Botta, M., and Borlak, J. (2010) Molecular
Zhu, L., Shang, D.S., and Chen, Y.H. (2010) Involvement
characterization of c"Abl/c"Src kinase inhibitors targeted
of Src tyrosine kinase in Escherichia coli invasion of human
against murine tumour progenitor cells that express stem
brain microvascular endothelial cells, FEBS Lett., 584,
cell markers, PLoS One, 5, e14143, doi: 10.1371/journal.
27-32, doi: 10.1016/j.febslet.2009.10.090.
pone.0014143.
43. Amano, M., Nakayama, M., and Kaibuchi, K. (2010) Rho"
35. Chang, Y.W., Marlin, J.W., Chance, T.W., and Jakobi R.
Kinase/ROCK: a key regulator of the cytoskeleton and cell
(2006) RhoA mediates cyclooxygenase"2 signaling to dis"
polarity, Cytoskeleton, 67, 545-554, doi: 10.1002/cm.20472.
rupt the formation of adherens junctions and increase cell
44. Julian, L., and Olson, M.F. (2014) Rho"associated coiled"
motility, Cancer Res., 66, 11700-11708, doi: 10.1158/
coil containing kinases (ROCK), Small GTPases, 5,
0008"5472.CAN"06"1818.
e29846, doi: 10.4161/sgtp.29846.
36. Shankar, J., and Nabi, I.R. (2015) Actin cytoskeleton reg"
45. Chen, Y.H., Chen, S.H., Jong, A., Zhou, Z.Y., Li, W.,
ulation of epithelial mesenchymal transition in metastatic
Suzuki, K., and Huang, S.H. (2002) Enhanced Escherichia
cancer cells, PLoS One, 10, e0119954, doi: 10.1371/jour"
coli invasion of human brain microvascular endothelial
nal.pone.0119954.
cells is associated with alternations in cytoskeleton induced
37. Lee, G., Kim H.J., and Kim, H."M. (2016) RhoA"JNK
by nicotine, Cell. Microbiol., 4, 503-514, doi: 10.1016/j.
regulates the E"cadherin junctions of human gingival
febslet.2009.10.090.
epithelial cells, J. Dent. Res., 95, 284-291, doi: 10.1177/
46. Soong, G., Martin, F.J., Chun, J., Cohen, T., Ahn, D.S.,
0022034515619375.
and Prince, A. (2011) Staphylococcus aureus protein A
38. Ivlev, A.P., Efremova, T.N., Khaitlina, S.Yu., and
mediates invasion across airway epithelial cells through
Bozhokina, E.S. (2018) Difference in susceptibility of 3T3
activation of RhoA GTPase signaling and proteolytic activity,
and 3T3"SV40 cells to invasion by opportunistic pathogens
J. Biol. Chem., 286, 35891-35898, doi: 10.1074/jbc.M111.
Serratia grimesii, Cell Tissue Biol., 12, 33-40.
295386.
39. Sousa, S., Cabanes, D., Bougneres, L., Lecuit, M.,
47. Wolke, S., Ackermann, N., and Heesemann, J. (2011) The
Sansonetti, P., Tran"Van"Nhieu, G., and Cossart, P. (2007)
Yersinia enterocolitica type 3 secretion system (T3SS) as
Src, cortactin and Arp2/3 complex are required for E"cad"
toolbox for studying the cell biological effects of bacterial
herin"mediated internalization of Listeria into cells, Cell.
Rho GTPase modulating T3SS effector proteins, Cell. Micro
Microbiol., 9, 2629-2643, doi: 10.1111/j.1462"5822.2007.
biol., 13, 1339-1357, doi: 10.1111/j.1462"5822.2011.01623.x.
00984.x.
48. Wang, L., Xue, L., Yan, H., Li, J., and Lu, Y. (2010) Effects
40. Bonazzi, M., Veiga, E., Pizarro"Cerda, J., and Cossart, P.
of ROCK inhibitor, Y"27632, on adhesion and mobility in
(2008) Successive post"translational modifications of E"
esophageal squamous cell cancer cells, Mol. Biol. Rep., 37,
cadherin are required for InlA"mediated internalization of
1971-1977, doi: 10.1007/s11033"009"9645"9.
Listeria monocytogenes, Cell. Microbiol., 10, 2208-2222,
49. Fu, P., Tang, R., Yu, Z., Li, C., Chen, X., Xie, M., Wang, W.,
doi: 10.1111/j.1462"5822.2008.01200.x.
and Luo, X. (2016) Rho"associated kinase inhibitors pro"
41. Eto, D.S., Jones, T.A., Sundsbak, J.L., and Mulvey, M.A.
mote microglial uptake via the ERK signaling pathway,
(2007) Integrin"mediated host cell invasion by type 1"pilia"
Neurosci. Bull., 32, 83-91, doi: 10.1007/s12264"016"0013"1.
THE OPPOSITE EFFECTS OF ROCK AND Src KINASE INHIBITORS
ON SUSCEPTIBILITY OF EUKARYOTIC CELLS TO INVASION
BY BACTERIA Serratia grimesii
E. S. Bozhokina, O. A. Tsaplina, and S. Yu. Khaitlina*
Institute of Cytology, Russian Academy of Sciences, 194064 St. Petersburg, Russia;
E mail: skhspb@gmail.com
Received September 20, 2018
Revised November 22, 2018
Accepted November 22, 2018
Internalization of bacteria into eukaryotic cells is determined by a sophisticated interaction of bacterial and host cell fac"
tors. Being a part of cell environment, opportunistic intracellular bacteria developed various mechanisms to interact with
cell surface receptors such as E"cadherin, integrins or the epidermal growth factor receptor, to activate components of sig"
naling pathways of eukaryotic cells and promote their own uptake, survival, and intracellular replication. Studying the
mechanisms that promote penetration of opportunistic bacteria Serratia grimesii into cultured eukaryotic cells, we have
previously shown that pre"treatment of cells with N"acetylcysteine (NAC) increases Serratia invasion, and this effect cor"
relates with an increased E"cadherin expression. As NAC is known to regulate both ROCK kinase and Src kinase expres"
sion, the aim of our work was to reveal participation of these signaling components in S. grimesii invasion. Here we demon"
strate that Y"27632, a specific inhibitor of Rho"associated protein kinase (ROCK), significantly enhanced invasion of cul"
tured eukaryotic cells by opportunistic bacteria S. grimesii. On the other hand, invasion of the same cells by S. grimesii was
inhibited with siRNA directed against RhoA or with the Src kinase inhibitor Src"I1. The inhibitor effects correlated with
the corresponding changes in expression of E"cadherin gene, up"regulated by the ROCK inhibition and down"regulated
by the Src inhibition. These results demonstrate participation of Src and ROCK protein kinases in invasion of eukaryotic
cells by opportunistic pathogen S. grimesii and suggest that additional signaling pathways are involved in the S. grimesii
uptake. Activity of these pathways appears to be increasing with the Y"27632"dependent inhibition of ROCK kinase.
Keywords: bacterial invasion, ROCK kinase, Src kinase, RhoA, Serratia grimesii
БИОХИМИЯ том 84 вып. 6 2019