БИОХИМИЯ, 2019, том 84, вып. 8, с. 1139 - 1146
УДК 577.19
РЕСВЕРАТРОЛ СПОСОБСТВУЕТ ДИФФЕРЕНЦИРОВКЕ in vitro
ОСТЕОБЛАСТОВ MC3T3 E1 ПУТЕМ ПОТЕНЦИРОВАНИЯ
КАЛЬЦИНЕУРИН/NFATC1 ОПОСРЕДОВАНННОГО
СИГНАЛЬНОГО ПУТИ*
© 2019
Y. Huang1#, J. Huo2#, F.Q. Liu3, J. Liu3,
X.J. Zhang4, C.H. Guo4, L.H. Song4**
1 Department of Biochemistry, Changzhi Medical College, Changzhi 046000, Shanxi, China
2 Department of Biology, Changzhi Medical College, Changzhi 046000, Shanxi, China
3 Changzhi Medical College, Changzhi 046000, Shanxi, China
4 Department of Pharmacology, Changzhi Medical College, Changzhi 046000,
Shanxi, China; E mail: slh10282001@163.com
Поступила в редакцию 15.03.2019
После доработки 15.03.2019
Принята к публикации 08.04.2019
Ранее было показано, что ресвератрол (RSVL) может стимулировать in vitro дифференцировку остеобластов
MC3T3 E1. Однако механизмы, лежащие в основе анаболического влияния ресвератрола на остеобласты,
остаются в большей части невыясненными. Целью настоящей работы являлось изучение молекулярного
механизма действия ресвератрола на процесс дифференцировки клеток MC3T3 E1. Эти клетки на протяже
нии 8 дней подвергали обработке различными концентрациями ресвератрола (10-8 моль/л-10-6 моль/л) и
циклоспорина А (является специфическим ингибитором калцинеурин/NFAT опосредованного сигнально
го пути) в концентрации 10-6 моль/л. На основании результатов предварительных тестирований клеточной
пролиферации и определения активности щелочной фосфатазы, нами была определена концентрация рес
вератрола, равная 10-7 моль/л, которую затем использовали в последующих экспериментах. Определение
уровня экспрессии мРНК и белка, кодируемых генами, ассоциированными с процессом остеоза, а именно,
генами CaN, NFATc1 и Runx2 осуществляли с помощью ОТ ПЦР в реальном времени и вестерн блоттинга
соответственно. Было обнаружено, что ресвератрол вызывает повышение экспрессии генов CaN, NFATc1 и
Runx2 с образованием соответствующих мРНК и белков по сравнению с контрольной группой (p < 0,05).
Циклоспорин A, напротив, снижал стимулирующий эффект ресвератрола (p < 0,05). С помощью иммуно
гистохимических методов показано, что обработка клеток ресвератролом приводит к накоплению белка
NFATc1 в клеточных ядрах. Обработка клеток циклоспорином А вызывает ингибирование индукции ресве
ратролом накопления белка NFATc1. Ресвератрол оказывает свое действие на остеобласты при участии
CaN/NFATc1 опосредованного сигнального пути.
КЛЮЧЕВЫЕ СЛОВА: ресвератрол, циклоспорин A, кальцинеурин/NFATc1, дифференцировка
остеобластов.
DOI: 10.1134/S0320972519080062
Фитоэстрогены образуют группу существую
клеток млекопитающих. Ресвератрол (RSVL -
щих в природе химических соединений расти
resveratrol, 3,4’,5 тригидроксистильбен) являет
тельного происхождения, биологическая актив
ся пригодным в пищу полифенольным фитоэст
ность которых сходна с активностью эстрогенов
рогеном, который обнаруживается в кожуре
Принятые сокращения: ALP - alkaline phosphatase, щелочная фосфатаза; CaN - calcineurin, кальцинеурин; CsA -
cyclosporin A, циклоспорин А; NFATc1 - nuclear factor of activated T cells cytoplasmic 1, ядерный фактор активированных
Т клеток цитоплазматический 1; NF κB - nuclear factor kappa B, ядерный фактор каппа В; pNPP - p nitrophenyl phosphate,
п нитрофенилфосфат; PVDF - polyvinylidene fluoride, поливинилиденфторид; RIPA - radioimmunoprecipitation assay, метод
радиоиммунопреципитации; RSVL - resveratrol, ресвератрол; real time RT PCR - real time reverse transcription polymerase
chain reaction, ОТ ПЦР в реальном времени, полимеразная цепная реакция в реальном времени с обратной транскрипци
ей; Runx2 - Runt related transcription factor 2, родственный белку Runt фактор транскрипции 2; SDS - sodium dodecyl sulfate,
додецилсульфат натрия; α MEM - alpha mimumum essential medium, минимальная эссенциальная среда альфа; IHC -
immunohistochemistry, иммуногистохимия.
biokhimiya, в рубрике «Papers in Press», BM 19 077, 13.05.2019. Статья на английском языке опубликована в № 6 2019.
** Адресат для корреспонденции.
# Авторы внесли равный вклад в работу.
1139
1140
HUANG и др.
большинства сортов винограда, корнях арахиса
МЕТОДЫ ИССЛЕДОВАНИЯ
и в различных лекарственных растениях [1].
В различных исследованиях было показано, что
Клеточные культуры. Клетки MC3T3 E1 бы
ресвератрол является антиоксидантом, антинео
ли получены от Шанхайского института биохи
пластическим, антитромбоцитарным и проти
мии и биологии клеток (Китай). Культуральные
вовоспалительным средством [2]. Он также сни
среды и препарат MTT (3 (4,5 dimethyl 2 thia
жает потерю костной ткани у крыс с удаленным
zolyl) 2,5 diphenyl 2 H tetrazolium bromide;
3
яичником [3]. Тем не менее, механизмы, лежа
(4,5 диметил 2 тиазолил) 2,5 дифенил 2 H
щие в основе анаболического влияния ресверат
тетразолиум бромид) были приобретены в BD
рола на остеобласты, остаются в большей части
Company (США), а фетальная бычья сыворотка
невыясненными.
в «Invitrogen» (США). Препарат циклоспорина
Белок кальцинеурин (CaN) является широко
А (CsA) был получен от «ALEXIS Laboratories»
распространенной серин треониновой фосфа
(США).
тазой. Ее субстратом внутри клеток выступает
Клетки MC3T3 E1 поддерживали в ростовой
ядерный фактор активированных Т клеток
среде (свободная от красного фенола мини
(NFAT). Кальцинеурин зависимое дефосфори
мальная эссенциальная среда, α MEM), допол
лирование факторов NFAT приводит к передаче
ненной 10% ной (в/в) фетальной бычьей сыво
сигналов о необходимости их локализации
роткой, 5 × 10-3 моль/л β глицерофосфата и
внутри ядра и транслокации этих факторов
25 г/л аскорбиновой кислоты, при 37 °C в 5% ной
внутрь ядра с последующей активацией опреде
CO2. Реагенты, которыми обрабатывали клетки,
ленных генов. Белки NFAT вместе образуют се
как показано ранее, добавляли в культуральную
мейство, состоящее из четырех факторов тран
среду через день. Затем культуральную среду ме
скрипции: NFAT1 (p, c2), NFAT2 (c, c1), NFAT3
няли каждые четыре дня.
(c4) и NFAT4 (x, c3). Показано, что в эмбрио
Определение выживаемости клеток с MTT.
нальных фибробластах NFATc1-/- происходит
Клетки инкубировали с различными концент
существенное нарушение процесса формирова
рациями ресвератрола (10-8 моль/л-10-6 моль/л)
ния костной ткани [4]. В настоящее время ос
в течение 8 дней. Затем клетки рассевали в 96
новное внимание уделяется изучению влияния
ячеечный планшет при плотности 3 × 103 клеток
активации гена NFATc1 на процесс остеокласто
на ячейку. После этого в каждую ячейку добав
генеза [5, 6]. В то же время до сих пор не выяс
ляли 20 мкл концентрированного раствора MTT
нена специфическая роль NFATc1 в процессе
в фосфатно солевом растворе (5 мг/мл) до дос
дифференцировки остеобластов. Кроме того, в
тижения конечной концентрации 0,5 мг/л MTT.
некоторых случаях полученные результаты про
Клетки инкубировали при 37°C в течение 4 ч.
тиворечат друг другу. Фактор транскрипции
Среду с MTT удаляли, образовавшиеся фиоле
Runx2 является основным регулятором процес
товые кристаллы формазана растворяли в
са дифференцировки остеобластов [7, 8]. Пока
150 мкл диметилсульфоксида и регистрировали
зано, что в его отсутствии дифференцировка ос
оптическую плотность (ОП) при 570 нм в каж
теобластов становится невозможной. В этих ус
дой ячейке с использованием ридера микро
ловиях происходит нарушение процесса форми
планшет. Контрольная группа выступала в каче
рования костей и снижение уровня транскрип
стве отрицательного контроля с 100% ой выжи
ции генов мишеней, на которые действует
ваемостью клеток. Пролиферацию клеток в об
Runx2 [9-11].
работанной группе, выраженную в процентах,
Основной целью настоящей работы было
рассчитывали следующим образом:
изучение молекулярного механизма действия
ОП обработанной группы/ОП контрольной
ресвератрола на процесс дифференцировки ос
группы × 100.
теобластов MC3T3 E1. Роль ресвератрола и воз
Определение активности щелочной фосфатазы.
можный механизм его действия оценивали пу
Клетки MC3T3 E1 при исходной плотности кле
тем анализа уровня экспрессии мРНК и белка у
ток, равной 2 × 104 клеток на ячейку, культивиро
генов, ассоциированных с остеозисом, таких
вали в среде α MEM в 24 ячеечном культураль
как CaN, NFATc1 и RunX2. Для проверки влия
ном планшете. После 8 дней культивирования
ния ресвератрола на процесс дифференцировки
клетки собирали и определяли активность ще
остеобластов, также был использован цикло
лочной фосфатазы (ALP) с помощью набора для
спорин А (CsA), специфический ингибитор
определения активности, содержащего п нитро
кальцинеурина. Его концентрация, равная
фенилфосфат (pNPP) («Sigma Chemical Co.»,
10-6 моль/л, была отобрана на основе результа
США) в соответствии с инструкциями произво
тов предварительных экспериментов и с учетом
дителя. Сначала клетки лизировали в лизисном
литературных данных [12].
буфере и инкубировали 30 мин при 37 °C. Затем
БИОХИМИЯ том 84 вып. 8 2019
ВЛИЯНИЕ РЕСВЕРАТРОЛА НА ДИФФЕРЕНЦИРОВКУ ОСТЕОБЛАСТОВ
1141
клеточные лизаты центрифугировали при 12 000 g
Таблица 1. Праймеры, использованные при проведении
ПЦР в реальном времени
при 4 °C в течение 10 мин. Осветленные лизаты
клеток переносили в новые центрифужные про
Праймер
Последовательность праймера (5′ 3′)
бирки объемом 1,5 мл и проводили определение
активности щелочной фосфатазы следующим
CaN F
CTCCCAGTTCAGCGTCAA
образом: 10 мкл осветленного клеточного лизата
CaN R
ATCGCCATCCTTATCCAG
помещали в 96 ячеечную прозрачную планшету.
NFATc1 F
GCGATCACTACTGGGATTTCT
Затем в каждую ячейку добавляли по 200 мкл
NFATc1 R
TCGAGGACGTGCATCTGTT
раствора п нитрофенилфосфата. Далее планше
RunX2 F
ATAGCAAAGGCCCTCACTAA
ту инкубировали в темноте в течение ~30 мин.
RunX2 R
AACTGGCTCTTCTGCTGATT
при комнатной температуре. После завершения
β актин F
AGACCTCTATGCCAACACAG
инкубации измеряли оптическую плотность об
β актин R
ACTCATCGTACTCCTGCTTG
разцов в ячейках при длине волны 405 нм на ри
дере многоячеечных планшет (США).
Полимеразная цепная реакция в реальном вре
мени с обратной транскрипцией (ОТ ПЦР в реаль
ный Тween 20. Затем мембраны инкубировали с
ном времени). Получали препарат общей РНК с
козлиными антикроличьими вторыми антитела
использованием набора MiniBEST Universal RNA
ми, коньюгированными с пероксидазой хрена
Extraction kit (Япония). Синтез кДНК осущест
(разведение 1 : 2000; A0208; «Beyotime Institute of
вляли с помощью набора PrimeScript RT Reagent
Biotechnology», Китай) при комнатной темпера
kit with gDNA Eraser (Япония). ОТ ПЦР прово
туре в течение 2 ч. В качестве контроля нагрузки
дили с использованием набора SYBR Green Mix
использовали β актин, который определяли с по
kit (Япония). Все образцы обрабатывали в трех
мощью мышиных поликлональных антител (раз
экземплярах в оптических пробирках в стрипах
ведение 1 : 5000; ab20272; «Abcam», Великобрита
по восемь штук (США). Последовательности
ния). Белки выявляли с помощью системы уси
праймеров представлены в табл.1. ОТ ПЦР в ре
ления хемолюминесценции («Amersham Biosci
альном времени проводили следующим образом:
ences», Германия) на приборе Image Quant LAS
исходное нагревание при 95 °C в течение 30 с и
4000 («GE Healthcare Life Sciences», Великобри
последующие 40 циклов по 5 с при 95 °C (денату
тания). Определение интенсивности белковых
рация) и при 60 °C (отжиг/элонгация) в течение
полос серого цвета осуществляли с помощью
30 с и затем диссоциация. Относительную вели
программного пакета Image J 1.37v software
чину экспрессии гена мишени рассчитывали с
(«National Institutes of Health», США).
использованием метода 2-ΔΔCt [13].
Иммуногистохимия. Клетки MT3T3 E1 в ко
Вестерн блоттинг. Клетки дважды промывали
личестве 1,0 × 105 клеток на ячейку рассевали на
ледяным натрий фосфатным буфером (PBS).
съемной крышке 6 ячеечной планшеты. Клетки
Общий белок клеток получали в лизисном буфе
культивировали в присутствии 10-7 моль/л рес
ре для проведения радиоиммунопреципитации
вератрола и 10-6 моль/л циклоспорина A в тече
RIP, содержащим 50 мМ Тris HCl, pH 8,0, 150 мМ
ние 8 дней. Затем клетки в течение 15 мин. фик
NaCl, 1% ный NP 40, 0,5% ный дезоксихолат
сировали в 4,0% ом параформальдегиде. Пер
натрия и 0,1% ный SDS, и определяли актив
меабилизацию клеток проводили в 0,5% ном
ность CaN, NFATc1 и RunX2. Содержание обще
Тriton X 100 в течение 10 мин. Далее клетки ин
го белка в лизатах клеток определяли с помощью
кубировали в блокирующем буфере. Затем до
наборов для определения концентрации белка,
бавляли мышиные моноклональные антитела
содержащих бицинхониновую кислоту (BCA)
против белка NFATc1 (разведение 1 : 200) и ин
(Китай). Белковые фракции разделяли с по
кубировали при 37 °C в течение 2 ч. Набор мы
мощью электрофореза в 10% ом полиакрил
шиных антител (PV 6002) использовали в соот
амидном геле c SDS, и разделенные белки пере
ветствии с инструкциями производителя
носили из геля на поливинилденфлюоридные
(«ZSGB BIO Co., Ltd.», Китай). Все изображе
(PVDF) мембраны. Затем среди разделенных бел
ния получали с помощью светового микроскопа
ков с помощью специфических антител выявля
Olympus BX51 («Olympus Corporation», Япония)
ли белки CaN (разведение 1 : 2000; ab137335;
при 400 кратном увеличении. На основании ин
«Abcam», Великобритания), NFATc1 (H 10; раз
тенсивности окрашивания, уровни экспрессии
ведение 1 : 500; «Santa Cruz Biotechnology», США)
NFATc1 оценивали следующим образом: - (без
и RunX2 (разведение 1 : 3000; ab23981; «Abcam»,
окрашивания), 1 (+), 2 (++) и 3 (+++). С учетом
Великобритания) в течение ночи при 4 °C.
доли NFATc1 положительных клеток, были ус
Мембраны PVDF трижды промывали натрий
тановлены следующие оценочные баллы:
Тris буферным раствором, содержащим 0,1%
- (0-1%; балл 0), + (1-24%; балл 1), ++ (25-49%;
БИОХИМИЯ том 84 вып. 8 2019
1142
HUANG и др.
балл 2) и +++ (50-100%; балл 3). Итоговые оце
ток, в этих клетках определяли активность ще
ночные баллы окрашивания NFATc1 были рас
лочной фосфатазы. Определение активности
считаны как суммы интенсивности окрашива
щелочной фосфатазы на единицу белка в клет
ния и процентный балл, которые выглядели
ках MC3T3 E1 проводили после обработки этих
следующим образом: -, нет экспрессии (общий
клеток в течение 8 дней различными концентра
балл 0); +, слабая экспрессия (общий балл 1-2);
циями ресвератрола. Максимальную активность
2+, умеренная экспрессия (общий балл 3-4) и
щелочной фосфатазы наблюдали в группе, об
3+, усиленная экспрессия (общий балл 5-6).
работанной ресвератролом в концентрации
Средние значения получали на основе значений
10-7 моль/л, для которой было показано значи
от десяти случайно выбранных полей зрения.
тельное увеличение активности в сравнении с
Статистическая обработка полученных ре
контрольной группой (рис. 1, b).
зультатов. Полученные данные представлены в
Влияние обработки ресвератролом на уровень
виде среднего значения ± стандартное откло
мРНК генов CaN, NFATc1 и Runx2, ассоциирован
нения. В соответствии с планом проведения
ных с остеозисом. На рис. 2 представлена карти
экспериментов, для сравнения результатов, по
на влияния ресвератрола на экспрессию мРНК
лученных в различных группах, использовали
генов CaN, NFATc1 и RunX2, полученная с по
метод одновариантного анализа с последую
мощью ОТ ПЦР в реальном времени.
щим определением значительных различий с
Значительное повышение уровня экспрес
помощью t теста. Значения величины p менее
сии мРНК CaN наблюдали в группе, обработан
0,05 рассматривали как статистически досто
ной ресвератролом, по сравнению с контроль
верные.
ной группой (p < 0,05). В то же время в группе,
обработанной ресвератролом и циклоспорином А,
было выявлено значительное снижение уровня
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
экспрессии мРНК CaN по сравнению с группой,
обработанной только ресвератролом (p < 0,05).
Влияние ресвератрола на процесс пролифера
Сходные результаты были получены при опре
ции клеток. Оптимальную концентрацию ресве
делении уровня экспрессии мРНК генов NFATc1
ратрола определяли с помощью метода оценки
и RunX2 (все значения p < 0,05).
выживаемости клеток с использованием MTT.
5RunX2. Чтобы оценить эффект действия
В соответствии с полученными результатами,
ресвератрола на клетки MC3T3 E1, были опре
для проведения дальнейших экспериментов бы
делены уровни белков, кодируемых генами CaN,
ла выбрана концентрация ресвератрола, равная
NFATc1 и RunX2. Как показано на рис. 3, ресве
10-7 моль/л (рис. 1, а).
ратрол значительно повышал уровень белков
Влияние ресвератрола на активность щелоч
CaN, NFATc1 и RunX2 (p < 0,05). В то же время
ной фосфатазы. Чтобы оценить влияние ресве
уровни этих белков в группе, обработанной рес
ратрола на дифференцировку остеогенных кле
вератролом и циклоспорином А, были значи
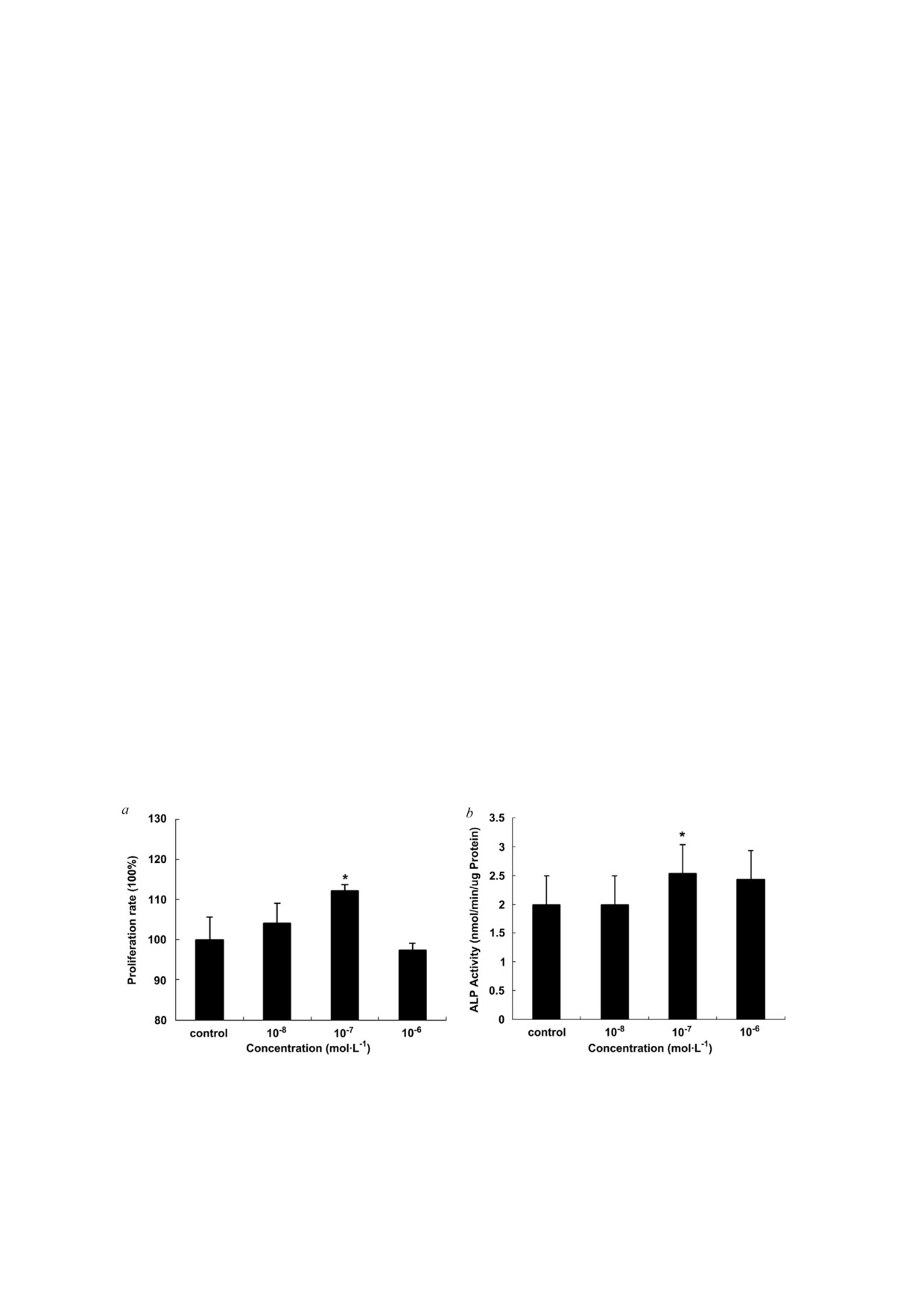
Рис. 1. Оценка пролиферации клеток MC3T3 E1 после их обработки ресвератролом (n = 6). Для проведения сравнитель
ной оценки пролиферации клеток MC3T3 E1 после их обработки ресвератролом использовали тест на выживаемость кле
ток с использованием MTT. При обработке клеток ресвератролом в концентрации 10-7 моль/л наблюдалась наибольшая
пролиферация в сравнении с контрольной группой (a). Дифференцировка клеток оценивалась по активности щелочной
фосфатазы (b); * p < 0,05 в сравнении с контрольной группой
БИОХИМИЯ том 84 вып. 8 2019
ВЛИЯНИЕ РЕСВЕРАТРОЛА НА ДИФФЕРЕНЦИРОВКУ ОСТЕОБЛАСТОВ
1143
Рис. 2. Экспрессия мРНК генов, ассоциированных с остеозисом. Клетки MC3T3 E1 подвергали различным обработкам в
течение 8 дней. Экспрессию мРНК генов CaN (a), NFATc1 (b) и RunX2 (c) определяли с помощью ОТ ПЦР в реальном вре
мени. Данные представлены в виде среднего значения ± стандартное отклонение (n = 3); * p < 0,05 против контрольной
группы. # p < 0,05 между группой клеток, обработанной только ресвератролом, и группой клеток, обработанных циклос
порином А и ресвератролом
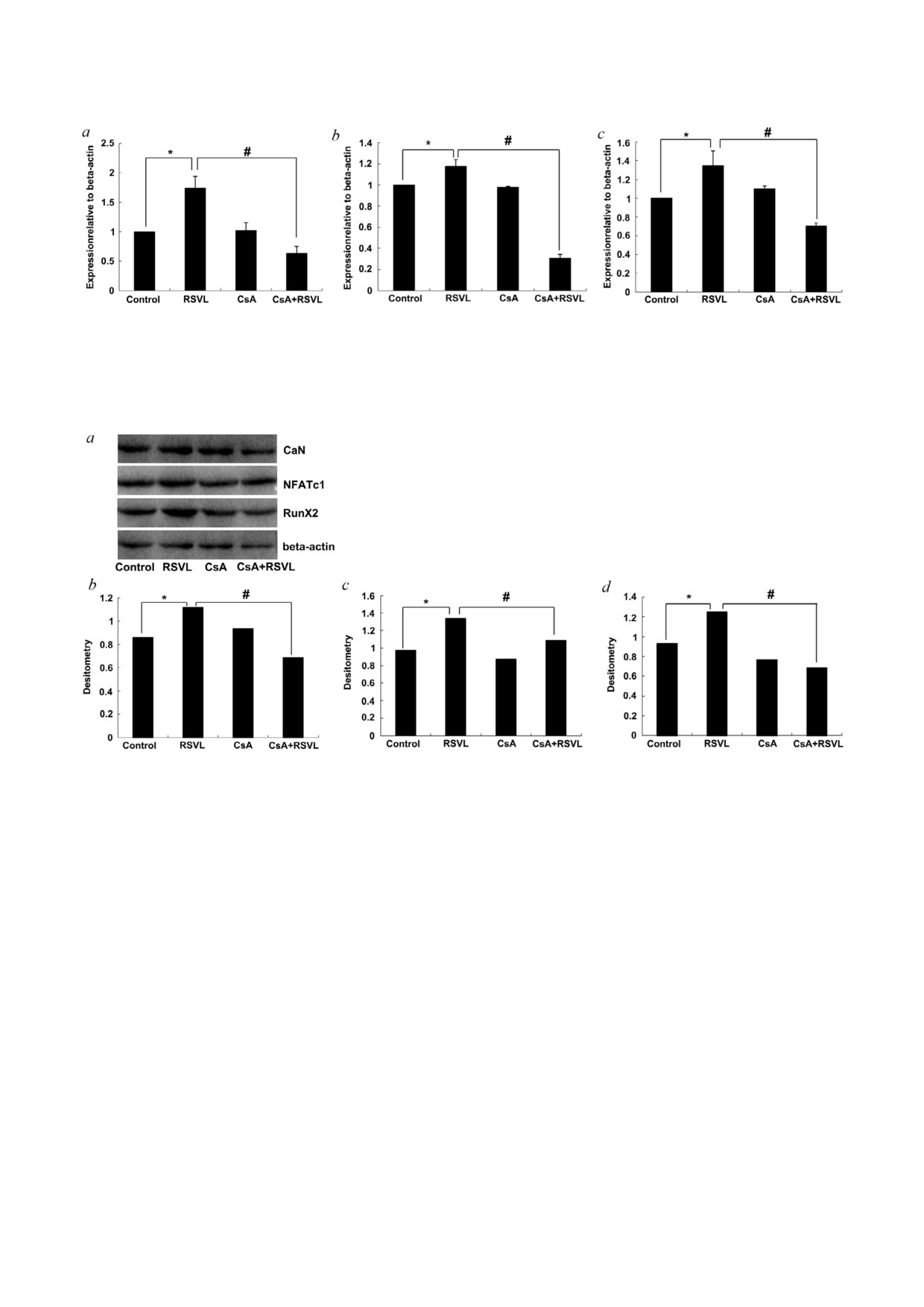
Рис. 3. Уровни белков, ассоциированных с остеопорозом, после различных обработок клеток. Уровни белков CaN,
NFATc1 и RunX2 (a). Денситометрический анализ белков CaN (b), NFATc1 (c) и RunX2 (d). В качестве контроля белковой
нагрузки использовали β актин; * p < 0,05 против контрольной группы, # p < 0,05 между группой, обработанной ресверат
ролом, и группой, обработанной ресвератролом и циклоспорином А
тельно снижены, по сравнению с группой, обра
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
ботанной только ресвератролом.
Ресвератрол индуцирует накопление NFATc1 в
Для лечения остеолитических костных забо
ядрах клеток. Для определения in situ уровня
леваний применяются различные лекарства,
NFATc1 в клетках MC3T3 E1 было проведено
например, бисфосфонаты, эстрогены и ралок
иммуногистохимическое окрашивание клеток.
сифен [14]. Однако, несмотря на эффективность
Результаты представлены на рис. 4. В табл. 2 по
действия этих лекарств, их применение ограни
казаны уровни NFATc1 в ядрах MC3T3 E1, опре
чено из за характерных для них побочных эф
деленные с использованием антител против бел
фектов, среди которых можно отметить тромбо
ка NFATc1. Иммуногистохимически было обна
эмболию и раздражение пищевода [15]. Конеч
ружено, что ресвератрол индуцировал накопле
ной целью разработки лекарств против остеопо
ние NFATc1 в ядрах клеток и способствовал зна
роза является нахождение агента, который мог
чительному усилению окрашивания NFATc1. Об
бы улучшить формирование кости через стиму
работка циклоспорином А приводила к предот
ляцию остеобластов и одновременное ингиби
вращению способности ресвератрола индуциро
рование остекластов без возникновения неже
вать экспрессию NFATc1 в клетках MC3T3 E1.
лательных побочных явлений. Для достижения
БИОХИМИЯ том 84 вып. 8 2019
1144
HUANG и др.
Рис. 4. Иммуногистохимическое окрашивание белка NFATc1 в клетках MC3T3 E1.
biokhsm/
этой цели были изучены природные соединения
активацией ERK1/2 [17]. Ресвератрол предот
и фитоэстрогены в качестве кандидатов на анти
вращает потерю костной ткани путем стимули
остеопоротические агенты. В предыдущих рабо
рования про остеогенного фактора BMP2[18]. В
тах было показано, что ресвератрол может сти
то же время до сих пор не было сообщений о су
мулировать пролиферацию и дифференцировку
ществовании взаимосвязи между RSVL инду
остеобластов [16-18]. Также было проведено
цированной дифференцировкой остеобластов и
изучение механизма действия ресвератрола.
CaN/NFATc1 зависимым сигнальным путем.
Например, показано, что ресвератрол может
Действительно, в нашей предыдущей работе
предотвращать апоптоз остеобластов путем уси
было показано, что ресвератрол может индуци
ления экспрессии β катенина [16]. Ресвератрол
ровать повышение уровня мРНК, продукта гена
стимулирует пролиферацию и дифференциров
Runx2. Белок Runx2, также известный как связы
ку остеобластов мезенхимных стволовых клеток
вающий фактор ядра альфа 1, является факто
костного мозга человека через эстрогеновый ре
ром транскрипции, жизненно необходимым для
цептор зависимый механизм и сопряжение с
дифференцировки остеобластов и образования
Таблица 2. Уровни NFATc1 в клетках MC3T3 E1, определенные с помощью иммуногистохимии
Всего
Группа
n
Значение p
-
1+
2+
3+
Контроль
6
1 (16,7%)
5 (83,3%)
0
0
p < 0,01
RSVL
6
0
0
2 (33,3%)
4 (66,7%)
p < 0,01
CsA+RSVL
6
2 (33,3%)
3 (50%)
1 (16,7%)
0
CsA
6
2 (33,3%)
4 (66,7%)
0
0
БИОХИМИЯ том 84 вып. 8 2019
ВЛИЯНИЕ РЕСВЕРАТРОЛА НА ДИФФЕРЕНЦИРОВКУ ОСТЕОБЛАСТОВ
1145
костной ткани. Нарушение экспрессии Runx2
жет быть задействован в процессе дифференци
приводило к полному нарушению формирова
ровки остеобластов, индуцированном ресверат
ния костей у зародышей мышей [19]. Эти резуль
ролом. Это наблюдение соответствует результа
таты позволили предположить, что существует
там, полученным Sun et al. [23], которые сообщи
важная связь между действием ресвератрола и
ли о том, что повышенная экспрессия CaN уси
транскрипцией в остеобластах, специфичной для
ливала остеобластогенез, а делеция гена CaN или
костных клеток. Более того, полученные резуль
его ингибирование снижали формирование
таты указывали на необходимость поиска новых
костной ткани и дифференцировку остеоблас
внутриклеточных молекул, на которые воздей
тов. Кроме того, ранее было показано, что ресве
ствует ресвератрол, и ассоциированных с ними
ратрол предотвращал ингибирование циклоспо
сигнальных путей, которые управляют, в конеч
рином А пролиферации и дифференцировки ме
ном итоге, дифференцировкой костных клеток.
зенхимных стволовых клеток костного мозга
Производя поиск сигнального пути, который мог
(BMSCs - bone marrow stem cells) в остеобласты
бы свести вместе индукцию ресвератролом и ак
[12]. Следовательно, в настоящей работе было
тивацию гена Runx2 в клетках MC3T3 E1, мы об
показано, что ресвератрол индуцировал диффе
ратили наше внимание на CaN/NFATc1 зависи
ренцировку остеобластов in vitro через потенци
мый сигнальный путь, поскольку он является
рование CaN/NFATc1 опосредованной оси пе
жизненно необходимым для сигнального каска
редачи сигнала. Эффект действия ресвератрола
да, задействованного в контроле дифференци
может быть обусловлен его участием в регуляции
ровки, апоптоза и адаптации широкого круга ти
транскрипции генов, участвующих в дифферен
пов клеток и тканей [20-22]. Поэтому было целе
цировке остеобластов. Тем не менее, необходимо
сообразно изучить возможность активации сиг
в дальнейшем изучить возможность наблюдения
нальной оси CaN/NFATc1 под действием ресве
подобных изменений in vivo.
ратрола. В настоящей работе клетки MC3T3 E1
Таким образом, в данной работе были полу
обрабатывали 10-7 моль/л ресвератрола в течение
чены доказательства влияния ресвератрола на
8 дней. После обработки была изучена пролифе
процесс дифференцировки остеобластов и учас
рация клеток, экспрессия мРНК и белка у генов,
тия CaN/NFATc1 зависимого сигнального пути
которые ассоциированы с остеозисом (CaN,
в индуцированной ресвератролом стимуляции
NFATc1 и Runx2), и маркеры дифференцировки
остеобластов. Эти данные способствуют раскры
при остеогенезе (активность щелочной фосфата
тию молекулярных механизмов, лежащих в ос
зы) (рис. 1, b). Показано, что обработка ресверат
нове стимуляции остеобластов ресвератролом.
ролом в концентрации 10-7 моль/л RSVL вызыва
ла повышение экспрессии соответствующих
мРНК и увеличивала содержание белков CaN,
Финансирование. Выполнение работы про
NFATc1 и RunX2 (рис. 2, рис. 3). С помощью им
водили при поддержке грантов программ
муногистохимии также было показано, что обра
Changzhi Medical College Science and Technology
ботка клеток ресвератролом вызывала накопле
Innovation Team (грант CX201413) и Shanxi
ние белка NFATc1 в клеточных ядрах (рис. 4).
Province Health and Family Planning Commission
Чтобы проверить результаты наших наблюдений,
research topic (грант 201602024).
клетки MC3T3 E1 были обработаны циклоспо
Конфликт интересов. Авторы заявляют об от
рином А, который действует как ингибитор, спе
сутствии между ними конфликта интересов.
цифичный для CaN/NFATc1 зависимого пути. В
Соблюдение этических норм. Настоящая
результате действия циклоспорина А, было отме
статья не содержит описания каких либо иссле
нено стимулирующее действие ресвератрола на
дований с использованием людей или животных
экспрессию генов CaN, NFATc1 и RunX2 как на
в качестве объектов.
уровне мРНК, так и на уровне белка (рис. 2 и рис.
Благодарности. Авторы выражают благодар
3), что свидетельствовало о том, что
ность проф. Сун (Song) за ее тщательное руковод
CaN/NFATc1 зависимый сигнальный путь мо
ство данной работой.
СПИСОК ЛИТЕРАТУРЫ
1.
Chen, R.S., Wu, P.L., and Chiou, R.Y. (2002) Peanut roots as
and bone loss in stroke prone spontaneously hypertensive
a source of resveratrol, J. Agric. Food Chem., 50, 1665 1667.
rats, J. Nutr. Sci. Vitaminol. (Tokyo), 46, 78 83.
2.
Fremont, L. (2000) Biological effects of resveratrol, Life
4.
Koga, T., Matsui, Y., Asagiri, M., Kodama, T.,
Sci., 66, 663 673.
Crombrugghe, B., Nakashima, K., and Takayanagi, H.
3.
Mizutani, K., Ikeda, K., Kawai, Y., and Yamori, Y. (2000)
(2005) NFAT and Osterix cooperatively regulate bone for
Resveratrol attenuates ovariectomy induced hypertension
mation, Nat. Med., 11, 880 885.
7 БИОХИМИЯ том 84 вып. 8 2019
1146
HUANG и др.
5.
Penolazzi, L., Zennaro, M., Lambertini, E., Tavanti, E.,
14. Jakob, F., Genest, F., Baron, G., Stumpf, U., Rudert, M.,
Torreggiani, E., Gambari, R., and Piva, R.
(2007)
and Seefried, L. (2015) Regulation of bone metabolism in
Induction of estrogen receptor alpha expression with decoy
osteoporosis: novel drugs for osteoporosis in development,
oligonucleotide targeted to NFATc1 binding sites in
Unfallchirurg, 118, 925 932.
osteoblasts, Mol. Pharmacol., 71, 1457 1462.
15. Rachner, T.D., Khosla, S., and Hofbauer, L.C. (2011)
6.
Zhou, F., Shen, Y., Liu, B., Chen, X., Wan, L., and Peng, D.
Osteoporosis: now and the future, Lancet, 377, 1276 1287.
(2017) Gastrodin inhibits osteoclastogenesis via down regulat
16. Li, P., Wang, Y., Liu, X., Zhou, Z., Wang, J., Zhou, H.,
ing the NFATc1 signaling pathway and stimulates osseointe
Zheng, L., and Yang, L. (2019) Atypical antipsychotics
gration in vitro, Biochem. Biophys. Res. Commun., 484, 820 826.
induce human osteoblasts apoptosis via Wnt/β catenin sig
7.
Ziros, P.G., Gil, A.P., Georgakopoulos, T., Habeos, I.,
naling, BMC Pharmacol. Toxicol., 20, 10.
Kletsas, D., Basdra, E.K., and Papavassiliou, A.G. (2002)
17. Dai, Z., Li, Y., Quarles, L.D., Song, T., Pan, W., Zhou, H.,
The bone specific transcriptional regulator Cbfa1 is a tar
and Xiao, Z. (2006) Resveratrol enhances proliferation and
get of mechanical signals in osteoblastic cells, J. Biol.
osteoblastic differentiation in human mesenchymal stem
Chem., 277, 23934 23941.
cells via ER dependent ERK1/2 activation, Phytomedicine,
8.
Ziros, P.G., Basdra, E.K., and Papavassiliou, A.G. (2008)
14, 806 814.
Runx2: of bone and stretch, Int. J. Biochem. Cell Biol., 40,
18. Zhao, M., Ko, S.Y., Garrett, I.R., Mundy, G.R.,
1659 1663.
Gutierrez, G.E., and Edwards, J.R. (2018) The polyphenol
9.
Vimalraj, S., and Selvamurugan, N. (2013) MicroRNAs:
resveratrol promotes skeletal growth in mice through a sir
synthesis, gene regulation and osteoblast differentiation,
tuin 1-bone morphogenic protein 2 longevity axis, Br. J.
Curr. Issues Mol. Biol., 15, 7 18.
Pharmacol., 175, 4183 4192.
10.
Kang, J.S., Alliston, T., Delston, R., and Derynck, R.
19. Maeno, T., Moriishi, T., Yoshida, C.A., Komori, H.,
(2005) Repression of Runx2 function by TGF beta
Kanatani, N., Izumi, S., Takaoka, K., and Komori, T. (2011)
through recruitment of class II histone deacetylases by
Early onset of Runx2 expression caused craniosynostosis,
Smad3, EMBO J., 24, 2543 2555.
ectopic bone formation, and limb defects, Bone, 49, 673 682.
11.
Jeon, E.J., Lee, K.Y., Choi, N.S., Lee, M.H., Kim, H.N.,
20. Rao, A., Luo, C., and Hogan, P.G. (1997) Transcription
Jin, Y.H., Ryoo, H.M., Choi, J.Y., Yoshida, M., Nishino, N.,
factors of the NFAT family: regulation and function, Annu.
Oh, B.C., Lee, K.S., Lee, Y.H., and Bae, S.C. (2006) Bone
Rev. Immunol., 15, 707 747.
morphogenetic protein 2 stimulates Runx2 acetylation,
21. Flanagan, W.M., Corthesy, B., Bram, R.J., and Crabtree, G.R.
J. Biol. Chem., 281, 16502 16511.
(1991) Nuclear association of a T cell transcription factor
12.
Song, L.H., Pan, W., Yu, Y.H., Quarles, L.D., Zhou, H.H.,
blocked by FK 506 and cyclosporine A, Nature, 352, 803 807.
and Xiao, Z.S. (2006) Resveratrol prevents CsA inhibition
22. Zayzafoon, M. (2006) Calcium/calmodulin signaling con
of proliferation and osteoblastic differentiation of mouse
trols osteoblast growth and differentiation, J. Cell.
bone marrow derived mesenchymal stem cells through an
Biochem., 97, 56 70.
ER/NO/cGMP pathway, Toxicol. In Vitro, 20, 915 922.
23. Sun, L., Blair, H. C., Peng, Y., Zaidi, N., Adebanjo, O. A.,
13.
Schmittgen, T.D., and Livak, K.J. (2008) Analyzing real
Wu, X. B., Wu, X. Y., Iqbal, J., Epstein, S., and Abe, E.
time PCR data by the comparative CT method, Nat.
(2005) Calcineurin regulates bone formation by the
Protoc., 3, 1101 1108.
osteoblast, Proc. Natl. Acad. Sci. USA, 102, 17130 17135.
RESVERATROL PROMOTES in vitro DIFFERENTIATION
OF OSTEOBLASTIC MC3T3 E1 CELLS VIA POTENTIATION
OF THE CALCINEURIN/NFATc1 SIGNALING PATHWAY
Y. Huang1#, J. Huo2#, F. Q. Liu3, J. Liu3, X. J. Zhang4, C. H. Guo4, and L. H. Song4*
1 Department of Biochemistry, Changzhi Medical College, Changzhi 046000, Shanxi, China
2 Department of Biology, Changzhi Medical College, Changzhi 046000, Shanxi, China
3 Changzhi Medical College, Changzhi 046000, Shanxi, China
4 Department of Pharmacology, Changzhi Medical College, Changzhi 046000,
Shanxi, China; E mail: slh10282001@163.com
Received March 15, 2019
Revised April 8, 2019
Accepted April 8, 2019
Resveratrol has been shown to stimulate differentiation of osteoblastic MC3T3 E1 cells in vitro; however, the mecha
nisms underlying the anabolic effect of resveratrol on osteoblasts remain largely unknown. Our study was aimed to inves
tigate the molecular mechanism of resveratrol induced differentiation of MC3T3 E1 cells. MC3T3 E1 cells were treat
ed for 8 days with different concentrations of resveratrol (10-8 10-6 M) and 10-6 M cyclosporine A (CsA), a specific
inhibitor of the calcineurin/NFAT pathway. According to the results of pilot studies of cell proliferation and alkaline
phosphatase activity, 10-7 M concentration of resveratrol was used in subsequent experiments. The levels of mRNA
expression of the osteosis related genes CaN, NFATc1, and Runx2 were analyzed by real time RT PCR; the levels of the
corresponding proteins were estimated by Western blot analysis. Resveratrol upregulated expression of the CaN, NFATc1,
and Runx2 genes at both mRNA and protein levels compared to the control group (p < 0.05), while CsA reduced the
effects of resveratrol (p < 0.05). Using immunohistochemical staining, we showed that resveratrol induced NFATc1 accu
mulation in the cell nuclei, and treatment with CsA inhibited resveratrol mediated induction of NFATc1, suggesting that
the calcineurin/NFATc1 signaling pathway plays an important role in the regulatory effect of resveratrol on osteoblasts.
Keywords: resveratrol, cyclosporine A, calcineurin/NFATc1, osteoblast differentiation
БИОХИМИЯ том 84 вып. 8 2019