БИОХИМИЯ, 2019, том 84, вып. 8, с. 1167 - 1176
УДК 577.22, 577.32
СРАВНЕНИЕ РАЗМЕРОВ И СВОЙСТВ НАНОСФЕР
КОМПЛЕКСА ЦИТОХРОМА c С КАРДИОЛИПИНОМ
В ОСАДКЕ И В НЕПОЛЯРНОЙ СРЕДЕ
© 2019
Г.К. Владимиров2,3*, В.Е. Ременщиков2, А.М. Нестерова2,3,
В.В. Волков2,4, Ю.А. Владимиров1,2,3,5
1 Московский государственный университет им. М.В. Ломоносова,
факультет фундаментальной медицины, 119991 Москва, Россия;
электронная почта: ura'vladimirov@yandex.ru
2 Институт кристаллографии им. А. В. Шубникова
ФНИЦ «Кристаллография и фотоника» РАН, 119333 Москва, Россия
3 Институт регенеративной медицины, Первый московский государственный
медицинский университет им. И. М. Сеченова Министерства здравоохранения
Российской Федерации (Сеченовский университет), 119991 Москва, Россия
4 НИЦ Курчатовский институт, 123182 Москва, Россия
5 Российский национальный исследовательский медицинский
университет им. Н. И. Пирогова Министерства здравоохранения
Российской Федерации, 117997 Москва, Россия
Поступила в редакцию 20.02.2019
После доработки 28.03.2019
Принята к публикации 12.04.2019
Апоптоз, главный вид программируемой смерти клеток, играет важную роль в обновлении клеток нашего
организма и удалении неполноценных или переродившихся клеток, в том числе и раковых. Одно из самых
ранних событий апоптоза - это липидная пероксидация во внутренней митохондриальной мембране, кото#
рая катализируется комплексом цитохрома c (ЦитС) с митохондриальным фосфолипидом - кардиолипи#
ном (КЛ). Ранее было получено, что при смешивании растворов ЦитС и КЛ образуются наносферы комп#
лекса цитохрома с с кардиолипином (наносферы Цит#КЛ), имеющие диаметр 11-12 нм, которые состоят из
расплавленной молекулы белка и монослоя КЛ. Методами динамического светорассеяния раствора Цит#КЛ
в хлороформе и малоуглового рассеяния рентгеновских лучей в осадке Цит#КЛ показано, что в обоих слу#
чаях Цит#КЛ образует наносферы диаметром 8 и 11 нм, что соответствует ранее полученным соотношени#
ям липид/белок, соответственно 13-14 и 35-50. Эти результаты показали, что наносферы Цит#КЛ образу#
ются не только при кристаллизации комплекса, но и в гидрофобной среде. Цитохром с в комплексе имеет
структуру расплавленной глобулы, о чем свидетельствовало появление флуоресценции триптофанового и
тирозиновых остатков, отсутствующей в нативном белке из#за фёрстеровского резонансного переноса
энергии электронного возбуждения на гем. Известно также, что в комплексе Цит#КЛ разрывается коорди#
национная связь гемового железа с серой метионина#80 (исчезает полоса поглощения ~700 нм). Также про#
исходил разрыв железосерной связи в ЦитС при помещении его в водно#метанольный (1/1 по объему) раст#
вор. Эти изменения были обратимыми, что подтверждает вывод об изменении конформации белка в мета#
нольных растворах по типу расплавленной глобулы.
КЛЮЧЕВЫЕ СЛОВА: апоптоз, цитохром c, кардиолипин, комплекс Цит#КЛ, гидрофобная среда, динами#
ческое светорассеяние, малоугловое рентгеновское рассеяние, расплавленная глобула.
DOI: 10.1134/S0320972519080098
Апоптоз, главный вид программируемой
химических реакций, приводящих к аморазру#
смерти клеток, играет огромную роль в обновле#
шению клетки и ее захвату макрофагами, одно
нии клеток живого организма и удалении не#
из самых ранних событий - это липидная пе#
полноценных или переродившихся клеток, в
роксидация во внутренней митохондриальной
том числе раковых [1, 2]. В сложной цепи био#
мембране, которая катализируется комплексом
Принятые сокращения: ДСР - динамическое светорассеяние; КЛ - кардиолипин; МУР - малоугловое рассеяние
рентгеновских лучей; ТОКЛ - 1,1’,2,2’ тетраолеилкардиолипин; ЦитС - цитохром c; Цит#КЛ - комплекс цитохрома с
с кардиолипином; >Fe3+-S(Met80) - железосерные связи между железом гема и серой метионина в 80#м положении.
* Адресат для корреспонденции.
1167
1168
ВЛАДИМИРОВ и др.
гемопротеина цитохрома c со специфическим
мой, что и в гидрофобном (в сущности, углево#
митохондриальным фосфолипидом - кардио#
дородном) окружении, в котором комплекс ока#
липином (КЛ) [3-5].
жется внутри липидного бислоя мембран. Более
Первая гипотеза о структуре такого комп#
адекватной моделью может служить суспензия
лекса была высказана в 1977 г. Brown tr al. и за#
наносфер Цит#КЛ в углеводородном или ином
ключалась в том, что комплекс представляет со#
неполярном растворителе. В предыдущем ис#
бой молекулу цитохрома с (ЦитС) и прилегаю#
следовании такая суспензия Цит#КЛ в хлоро#
щий участок липидного бислоя мембраны
форме и гексане была приготовлена [21]. В на#
(мембранно#связанный цитохром с) [6]. Эту ги#
стоящей работе были усовершенствованы усло#
потезу в дальнейшем развивали и использовали
вия получения суспензии и измерены размеры
в ряде работ [7-9], см. также обзоры [10] и [11].
частиц Цит#КЛ методом динамического свето#
Поскольку при физиологических рН головка
рассеяния (ДСР) и состояние цитохрома c в этих
КЛ на поверхности липидного бислоя мембран
частицах методами спектрофотометрии и спект#
несет два отрицательных заряда ортофосфата, а
рофлуориметрии. Эти данные были сопоставле#
цитохром c имеет четыре избыточных положи#
ны с результатами изучения свойств наночастиц
тельных заряда, основной вклад взаимодей#
Цит#КЛ в осадке методом малоуглового рассея#
ствия ЦитС с мембраной вносит электростати#
ния рентгеновских лучей на лабораторном при#
ческое притяжение, но затем к нему добавляют#
боре и на синхротронной станции.
ся водородные связи и/или гидрофобные взаи#
модействия, что сопровождается изменением
конформации белка и появлением у цитохрома c
МЕТОДЫ ИССЛЕДОВАНИЯ
необычного свойства - он становится перокси#
дазой, которая способна окислять, в том числе и
В работе использовали следующие реактивы:
липиды мембран, в состав которых входят поли#
цитохром с из сердца лошади («Sigma#Aldrich»,
ненасыщенные жирные кислоты (см. обзоры
США); 1,1’,2,2’ тетраолеилкардиолипин (ТОКЛ,
[10], [11] и статьи [9, 12-15]).
натриевая соль, «Avanti Polar Lipids», США);
Концепция мембранно#связанного цитохро#
KH2PO4 («Sigma#Aldrich») для приготовления
ма c оказалась, однако, не единственной. В 2011 г.
калий#фосфатного буферного раствора. В каче#
на основании данных, полученных методом ма#
стве растворителей использовали: метанол
лоуглового рентгеновского рассеяния (МУР)
(«Sigma#Aldrich»), хлороформ («Sigma#Aldrich»).
осадка, состоящего из комплекса цитохрома с
Подготовка проб для малоуглового рентгенов7
с кардиолипином, была предложена совершенно
ского рассеяния. Исходные растворы ЦитС гото#
другая схема строения комплекса, согласно ко#
вили в 0,05 М калий#фосфатном буферном раст#
торой молекула цитохрома с находится в центре
воре в концентрации 10-30 мг/мл; тетраолеил#
наносферы и окружена монослоем из молекул
кардиолипина - в растворе метанола в 30#крат#
кардиолипина [16]. В таком комплексе, назван#
ном избытке по отношению к ЦитС
ном Цит#КЛ, молекула цитохрома c находилась
(моль/моль), затем или добавляли в раствор
в «набухшем» состоянии, что проявлялось в уве#
ЦитС (для получения комплекса), или разбав#
личении ее диаметра (5,6 нм вместо 4,0 нм) [16]
ляли калий#фосфатным буферным раствором
и появлении флуоресценции тирозиновых и
(контрольный раствор кардиолипина). Общий
триптофанового аминокислотных остатков
объем пробы составлял 100 мкл.
[17-19], которая в нативном ЦитС потушена ге#
Исследование интенсивности малоуглового
мом, расстояние которого до этих остатков в на#
рентгеновского рассеяния. Измерения интенсив#
тивном белке лежит в пределах радиуса Фёрсте#
ности малоуглового рассеяния проводили на
ра. Эта гипотеза объясняет большинство опи#
двух установках - автоматическом малоугловом
санных в литературе свойств комплекса в мо#
рентгеновском дифрактометре АМУР#К и стан#
дельных системах, включая соотношение ли#
ции BioSAXS BM29 (7-15 кэВ) синхротрона
пид/белок [11, 20], пероксидазную и липоксиге#
ESRF («European Synchrotron Radiation Facility»,
назную активность комплекса Цит#КЛ.
Франция, 2D детектор PILATUS#1M («DECTRIS»,
Предложенная структура наносферы Цит#КЛ
Швейцария) интенсивность излучения 1012 фо#
предполагает, что, имея неполярную поверх#
тонов/с). Для установки АМУР#К измерения
ность, она будет в условиях живой клетки распо#
проводили при фиксированной длине волны
лагаться не в водной среде, а внутри гидрофоб#
излучения λ, равной 0,154 нм (CuKα линия ост#
ной фазы биологических мембран, как это было
рофокусной трубки ISO#DEBYEFLEX 3003 с
сформулировано в работе [11]. Однако нет уве#
медным анодом), и коллимационной системой
ренности в том, что структура Цит#КЛ в составе
Кратки, энергетический спектр излучения фор#
микрокристаллического осадка будет той же са#
мировали с помощью монохроматора из пиро#
БИОХИМИЯ том 84 вып. 8 2019
НАНОСФЕРЫ КОМПЛЕКСА ЦИТОХРОМА c С КАРДИОЛИПИНОМ
1169
литического графита, число угловых отсчетов
Предполагается, что смесь хлороформ#мета#
составляло 3300, сечение рентгеновского пучка
нол выделяет все липиды [23]. Таким образом,
составляло 0,2 × 8 мм при диапазоне углов рас#
можно предположить, что в верхней фазе нет
сеяния, соответствующем значениям модуля
комплекса ЦитС с ТОКЛ, но между двумя фаза#
волнового вектора 0,1 < s < 12 нм-1 (s = 4πsinθ/λ,
ми появлялся осадок, к которому должно отно#
2θ — угол рассеяния). Для установки станции
ситься 40% от исходного количества комплекса
BM29 измерения проводили при фиксирован#
ЦитС с ТОКЛ, так как в нижней фазе мы обна#
ной длине волны излучения λ, равной 0,123 нм,
ружили 60% от исходного количества комплекса
в интервале волновых векторов 0,07 < s < 5,5 нм-1.
ЦитС с ТОКЛ (измерено по поглощению).
Образцы помещали в 1,0 мм стеклянные ка#
Спектрофотометрический анализ изменений
пилляры со стенками толщиной 0,01 мм, каме#
конформации (развертывания) цитохрома c в ме7
ры образца вакуумировали с помощью форваку#
таноле и комплексе Цит7КЛ проводили на
умного масляного насоса. Время измерений од#
Specord 200 («Analytik Jena», Германия) с ис#
ного образца раствора составляло 3 ч.
пользованием кварцевых кювет с длиной опти#
Полученные экспериментальные данные
ческого пути 1,00 см. Спектры регистрировали
интенсивности малоуглового рассеяния были
в пробе объемом 3000 мкл в диапазоне 650-
нормированы на интенсивность падающего
750 нм.
пучка, после чего вычитали рассеяние от раст#
Измеряли спектры поглощения ЦитС в вод#
ворителя или фоновое рассеяние.
ных и водно#метанольных растворах с различ#
Обработку и анализ данных проводили с ис#
ной объемной долей метанола. Характерный
пользованием программ SASPLOT и PEAK, вхо#
пик железосерных связей в геме ЦитС наблюда#
дящих в пакеты ATSAS, разработанные в Ин#
ли при длине волны 699 нм. Фоновый спектр
ституте кристаллографии им. А. В. Шубникова
вычитали из спектра ЦитС в соответствии со
ФНИЦ «Кристаллография и фотоника» РАН
следующим уравнением:
[22]. Основной результат - кривая малоуглового
рассеяния в координатах: вектор рассеяния
AFe-S = A699 - (A670 + A728)/2.
(абсцисса)/логарифм интенсивности рассеяния
(ордината). Вручную выделяли интересующий
Уравнение основано на том, что длина вол#
участок кривой в области выраженного острого
ны максимума поглощения железосерных свя#
пика и для него находили основные параметры,
зей (699 нм) находится точно посередине интер#
среди которых наиболее интересные для нас -
вала между длинами волн 670 нм и 728 нм.
средний размер микрокристаллических струк#
Статистический анализ данных. Статистиче#
тур в нанометрах (Long Range Order) и межплос#
скую обработку данных проводили с использо#
костное расстояние (d) в микрокристалле, соот#
ванием компьютерной программы GraphPad
ветствующее данному пику рассеяния (Bragg
Prism 6 («GraphPad Software»). Все данные при#
spacing).
ведены в формате среднее значение ± стандарт#
Приготовление комплекса Цит7КЛ в неполяр7
ная ошибка среднего. Статистические различия
ном окружении было проведено на основе метода
данных анализировали с использованием непа#
Фолча для экстракции липидов [23] в соответ#
раметрического U#критерия Манна-Уитни.
ствии с протоколом, описанным в работе [24] с
Значение p < 0,05 считали статистически значи#
небольшими изменениями: 260 мкМ водно#ме#
мым.
танольного раствора (4/1 об/об) ЦитС смешива#
ли с 13,5 мМ метанольным раствором ТОКЛ,
чтобы получить водно#метанольный (1/1 об/об)
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
раствор комплекса ЦитС с ТОКЛ с молярным от#
ношением 1/30. При этом отношении ТОКЛ на#
Исследование структуры комплекса Цит7КЛ
ходилось в избытке, что облегчило переход комп#
методом малоуглового рассеяния. Ранее в нашей
лекса из водно#метанольного раствора в хлоро#
лаборатории был получены данные, свидетель#
формную фазу. Затем хлороформ добавляли к
ствующие о наличии упорядоченной структуры
водно#метанольному раствору ЦитС с ТОКЛ для
в растворе, содержащем комплекс Цит#КЛ при
достижения объемного соотношения хлоро#
молярном отношении 1/35 [16]. При воспроиз#
форм/вода/метанол, равного 2/1/1. Смесь интен#
ведении этих результатов методом МУР для
сивно перемешивали в течение 2 мин. После
межплоскостного расстояния получены наряду
спонтанного разделения фаз нижнюю (хлоро#
с пиками первого семейства, которые были
форм#метанольную) фракцию, содержащую
слегка сдвинуты (10,95 и 5,42 нм) по отношению
экстрагированный комплекс Цит#КЛ, осторож#
к литературным данным (11,25 и 5,47 нм соглас#
но отбирали стеклянной пипеткой Пастера.
но [16]), также и пики второго семейства (8,26 и
БИОХИМИЯ том 84 вып. 8 2019
1170
ВЛАДИМИРОВ и др.
Рис. 1. Кривые интенсивности малоуглового рассеяния
Рис. 2. Дифракционные пики от микрокристалла комп#
комплекса Цит#КЛ в 0,05 М калий#фосфатном буферном
лекса Цит#КЛ в 0,05 М калий#фосфатном буферном раст#
растворе (рН 3,6) при концентрации цитохрома с 1,85 мМ,
воре (рН 7,2), при концентрации цитохрома с 0,1 мМ, со#
соотношение ЦитС/ КЛ - 1/30. Цифры в максимумах рас#
отношение ЦитС/ КЛ - 1/30; где 1 - узкий пик первого
сеяния соответствуют межплоскостным расстояниям в
порядка (Long range order = 715 нм, d = 8 нм); 2 - узкий
микрокристаллах (все значения даны в нм). Пунктирными
пик второго порядка (Long range order = 586 нм, d = 4 нм).
линиями указаны значения, рассчитанные из данных по
Данные получены на установке BM29 синхротрона ESRF
пикам первого порядка (10,95/2 = 5,47, 8,26/2 = 4,13). Дан#
и обработаны с использованием программ PEAK и
ные получены на приборе АМУР#К и обработаны с ис#
SASPLOT
пользованием программы SASPLOT
4,01 нм). Видно, что различия между рассчитан#
Для увеличения интенсивности дифракци#
ными и данными, полученными из эксперимен#
онных пиков была уменьшена концентрации
тальных кривых (5,42 и 4,01 нм), составляют
ЦитС в исследуемых растворах с 10 до 1 мг/мл.
0,05 и 0,12 нм (рис. 1).
Такая возможность появилась после того, как
Следует отметить, что пики второго семей#
стало доступным проведение измерений на
ства воспроизводились не всегда. Было проведе#
станции излучения BM29 синхротрона ESRF.
но варьирование концентрации ЦитС, pH сис#
Кроме того, существенным явился переход рН
темы, и были найдены условия воспроизводи#
исследуемых растворов из нейтральной в кис#
мости пиков второго семейства. Установлено,
лую. Еще одним весомым дополнением стала
что наиболее отчетливо они проявлялись при
модификация раствора кардиолипина в мета#
низких рН и высоких концентрациях ЦитС (при
ноле путем добавки определенного количества
pH 3,6 и концентрации ЦитС 23 мг/мл пик вто#
исходного калий#фосфатного буферного раст#
рого семейства был более отчетлив, чем при
вора.
рН 6,5 и более низких концентрациях ЦитС).
Таким образом, на станции BM29 синхро#
Обобщение всех полученных данных по величи#
трона ESRF удалось измерить узкие дифракци#
нам межплоскостных расстояний при рН 3,6 и
онные пики, а при рН 7,2 наблюдали не один, а
6,3 дано в табл. 1. Таким образом, согласно полу#
два узких пика, соответствующие первому и вто#
ченным результатам, диаметр комплекса Цит#КЛ
рому порядкам отражения от плоскостей с d =
может составлять либо 11 нм, либо 8 нм с неко#
= 8,07 и 4,00 нм (рис. 2). При pH 6,5, 7,2 и 7,4 на#
торым разбросом данных.
блюдали пики одной группы - 8,07 ± 0,03 (n = 5),
Таблица 1. Сводные данные по межплоскостным расстояниям в опытах, проведенных на установке АМУР#К и обработан#
ных программой SASPLOT. Межплоскостные расстояния приведены в виде «среднее значение ± стандартная ошибка
среднего, нм», для каждого значения pH n = 3
pH
Пики 1#го семейства,
Пики 1#го семейства,
Пики 2#го семейства,
Пики 2#го семейства,
1#го порядка, нм
2#го порядка, нм
1#го порядка, нм
2#го порядка, нм
6,3
10,91 ± 0,06
5,41 ± 0,01
8,01 ± 0,01
3,99 ± 0,01
3,6
10,95 ± 0,00
5,49 ± 0,09
8,26 ± 0,00
4,00 ± 0,01
БИОХИМИЯ том 84 вып. 8 2019
НАНОСФЕРЫ КОМПЛЕКСА ЦИТОХРОМА c С КАРДИОЛИПИНОМ
1171
при pH 7,4 - также пики второй группы -
11,35 ± 0,21 (n = 5).
Изучение размеров молекул цитохрома c и
комплекса Цит7КЛ в растворе методом динами7
ческого светорассеяния. Прежде всего, была оце#
нена возможность определения методом ДСР
размера мелких частиц, по форме близких к
сферическим. Обычно измеряются частицы раз#
мером более 10 нм, что связано с тем, что длина
волны света, рассеяние которого изучается, сос#
тавляет 633 нм. В работах [25, 26], тем не менее
определялись размеры небольших белков. С по#
мощью метода ДСР мы определили диаметр
Рис. 3. Флуктуации размера частиц в растворах, измерен#
ные методом динамического рассеяния: а - средние значе#
частиц ЦитС в водном растворе (рис. 3, а), кото#
ния диаметров частиц в водном растворе ЦитС, получен#
рый составил 3,6 ± 0,1 нм, что по значению
ные при последовательных измерениях в одной пробе; б -
близко к размерам ЦитС в белковых кристаллах
средние значения диаметров частиц Цит#КЛ, растворен#
или водных растворах, измеренным с помощью
ных в хлороформной фазе (см. «Методы исследования»)
методов дифракции рентгеновского излучения
и малоуглового рассеяния [16]. Как и ожида#
лось, при последовательных измерениях в од#
последовательных измерениях в одной пробе
ной пробе, измеряемый диаметр частиц изме#
средние размеры частиц варьировали в доста#
нялся незначительно, что говорит о том, что
точно широком диапазоне значений (рис. 3, б).
система содержит одинаковые частицы и что
Это указывает на то, что в пробе содержались
точность измерений достаточно высока. Комп#
частицы различных размеров. Скорее всего, это
лекс Цит#КЛ в водно#хлороформ#метанольном
объясняется тем, что Цит#КЛ содержал различ#
растворе был получен из ЦитС и ТОКЛ (в мо#
ное количество молекул кардиолипина, при#
лярном соотношении 1/30) и затем переведен в
крепленных к ЦитС.
хлороформ#метанольную фазу, после чего был
Были выявлены три наиболее часто встречаю#
измерен размер частиц в пробе. Средний диа#
щиеся диапазона (табл. 2), в которые попадали
метр частиц в пробе составил 10,9 ± 0,9 нм, что
измеренные средние диаметры частиц: 4,0-5,5
практически точно совпадает со значением в
(группа 1); 7,0-8,5 (группа 2) и 11,5-13,0 нм
11 ± 1 нм, полученным для кристаллов Цит#КЛ
(группа 3). Средние диаметры частиц в группах
[16]. Однако в отличие от размеров частиц, по#
достоверно различались и составили: группа 1 -
лученных при последовательных измерениях в
4,69 ± 0,21, группа 2 - 7,79 ± 0,28, группа 3 -
водном растворе ЦитС, в данном случае при
12,1 ± 0,17 нм.
Изучение влияния метанола на комплекс
Цит7КЛ методом спектрофотометрии. Ранее бы#
ло показано, что наличие метанола необходимо
Таблица 2. Результаты обработки данных, полученных ме#
тодом динамического светорассеяния при последователь#
для того, чтобы комплекс Цит#КЛ был успешно
ных измерениях пробы, содержащей хлороформ#мета#
экстрагирован в неполярную фазу [24]. Поэтому
нольный раствор Цит#КЛ
представлялось интересным изучить влияние
Диаметр
Число
Средний размер частиц, ср. знач. ±
непосредственно метанола на свойства ЦитС,
частиц, нм
измерений
стандартная ошибка среднего, нм
чтобы понять, какие изменения происходят с
цитохромом с в процессе приготовления ком#
0,0-4,0
1
плекса с кардиолипином еще до добавления в
4,0-5,5
4
4,69 ± 0,21
систему последнего. Для этого в нашей работе
5,5-7,0
2
мы использовали спектрофотометрический ме#
7,0-8,5
5
7,79 ± 0,28
тод и изучали поглощение в области 699 нм, где
8,5-10,0
1
поглощают железосерные связи между железом
10,0-11,5
4
10,51 ± 0,18
гема и серой метионина в 80#м положении (см.
11,5-13,0
8
12,1 ± 0,17
«Методы исследования»).
13,0-14,5
3
13,36 ± 0,20
Показано, что при изменении концентрации
14,5-16,0
2
метанола от
0 до
10% поглощение связи
16,0-17,5
2
>Fe3+-S(Met80) при 699 нм снижается на 10%, а
17,5-19,0
1
при концентрации 50% происходит почти пол#
20,5-22,0
1
ное исчезновение полосы поглощения, что на#
блюдали и ранее в работе нашей группы [24].
БИОХИМИЯ том 84 вып. 8 2019
1172
ВЛАДИМИРОВ и др.
гидрофобном растворителе (табл. 2). Были вы#
явлены частицы с диаметрами 12,1 ± 0,17 нм и
7,79 ± 0,28 нм, количество которых в растворе
значительно превышало количество частиц дру#
гих размеров.
Таким образом, нами были получены дан#
ные, которые указывают на существование оди#
наковых частиц в составе осадка Цит#КЛ и в
растворе Цит#КЛ в гидрофобном растворителе.
В обоих случаях частицы могут иметь диаметр
~11 или 8 нм. Различия в диаметре этих двух ви#
дов наносфер соответствуют разнице в соотно#
шении липид/белок (как описано в работе [21]),
которое при нейтральных рН составляет, по раз#
ным данным, от 35/1 [16] до 59/1, а в кислой
среде (pH < 5) - от 13/1 до 14/1 [27].
Естественно, возникает вопрос, соответствует
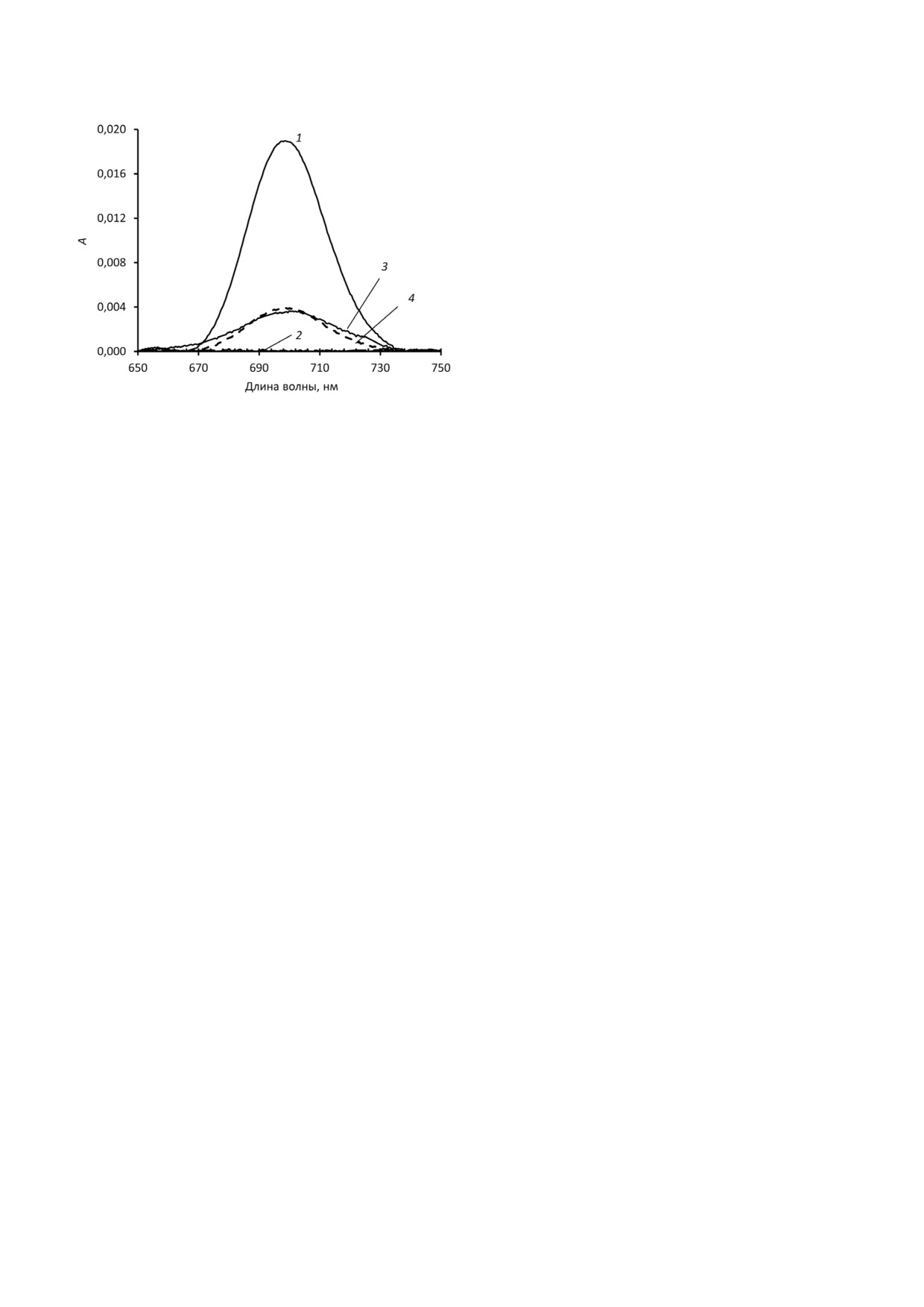
Рис. 4. Спектры поглощения в растворах ЦитС, указываю#
щие на обратимость действия метанола на состояние
ли полученное в данной работе соотношение ли#
>Fe3+-S связей (699 нм). Раствор 1 - 50 мкМ ЦитС (0% ме#
пид/белок соотношению в комплексах Цит#КЛ в
танола); раствор 2 - 50 мкМ раствор ЦитС (50% метанола
мембранах митохондрий или других клеточных
(об./об.)); раствор 3 - раствор 1, разбавленный в пять раз
водой (Cкон (ЦитС) = 10 мкМ, 10% метанола (об./об.));
мембранах? К сожалению, существующие мето#
раствор 4 (пунктирная линия) - 10 мкМ раствор ЦитС (0%
ды не позволяют отделить эти комплексы от ос#
метанола)
новной массы цитохрома с и кардиолипина в
митохондриальных мембранах. Но маловероят#
но, что молекулы цитохрома с и кардиолипина,
Далее было изучено, является ли разрушение
которые в водной и неполярной средах образу#
железосерной связи метанолом обратимым или
ют одну и ту же структуру - наносфера Цит#КЛ,
необратимым процессом. Полученные данные
с одинаковым размером и соотношением ли#
говорят о том, что в случае раствора ЦитС с объ#
пид/белок, по какой#то причине не образуют
емной долей метанола, равной 50%, этот про#
эту структуру в живой клетке, а образуют нечто
цесс является обратимым - при 5#кратном раз#
совершенно иное. При этом как в модельных
бавлении такого раствора дистиллированной
системах, так, вероятно, и в клеточных мембра#
водой (что сопровождается снижением объем#
нах, соотношение липид/белок зависит от окру#
ной доли метанола до 10%) при 699 нм наблюда#
жающих условий, в том числе, от pH. Получен#
лось поглощение, составляющее 92% от погло#
ные размеры частиц соответствуют структуре
щения водного раствора ЦитС аналогичной
наносферы, в которой глобула частично рас#
концентрации (рис. 4).
плавленного цитохрома c (диаметр 5,6 нм) окру#
жена жидким гидрофобным слоем углеводород#
ных цепей молекул кардиолипина. В пользу то#
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
го, что ЦитС в комплексе Цит#КЛ, помещенном
в гидрофобную среду, имеет структуру расплав#
В предыдущих работах нами было показано,
ленной глобулы, говорят также появление флу#
что при смешивании ЦитС, находящегося в вод#
оресценции триптофанового и тирозиновых ос#
ном растворе, с раствором митохондриального
татков, отсутствующей в нативном белке из#за
фосфолипида кардиолипина образуется ком#
фёрстеровского резонансного переноса энергии
плекс Цит#КЛ с совершенно определенным со#
электронного возбуждения на гем, и разрыв ко#
отношением липид/белок [11]. При изучении
ординационной связи гемового железа с серой
микрокристаллического осадка, выпадающего
метионина#80 (исчезновение полосы поглоще#
из концентрированных водных растворов ком#
ния ~700 нм) [21]. Согласно работе [28], в кото#
плекса Цит#КЛ, были обнаружены частицы с
рой изучали мутантные формы ЦитС, нативная
диаметром ~11 нм [16]. В данной работе при
структура железосерной связи поддерживается
низких значениях рН в микрокристаллическом
аминокислотными остатками Lys72, Lys73 и
осадке наряду с частицами диаметром 11,35 ±
Lys79. При этом Lys72 и Lys79 влияют на связы#
± 0,21 также обнаружены и частицы с диамет#
вание ЦитС с КЛ, а наличие Lys73 необходимо
ром 8,07 ± 0,03 нм. Кроме того, с помощью ме#
для проявления комплексом Цит#КЛ перокси#
тода ДСР впервые определили размер частиц в
дазной активности [28]. Судя по всему, разрыв
растворе комплекса цитохром с/кардиолипин в
железосерной связи между гемом и Met80 затра#
БИОХИМИЯ том 84 вып. 8 2019
НАНОСФЕРЫ КОМПЛЕКСА ЦИТОХРОМА c С КАРДИОЛИПИНОМ
1173
Рис. 5. Схема строения частиц комплекса Цит#КЛ и изменения конформации ЦитС при их формировании. Размеры час#
тиц приведены в масштабе. Частицы: верхний ряд - схема строения глобулы ЦитС, находящегося в нативном состоянии
(а - расположение остатка Tyr и гема (черный прямоугольник), б - глобула до образования частицы Цит#КЛ, в - распо#
ложение атома гемового железа >Fe относительно атома серы Met80, координационная железосерная связь); средний
ряд - строение расплавленной глобулы ЦитС (г - полученной после воздействия метанолом, в результате чего диаметр
глобулы увеличился, а остатки Tyr удалились от гема, д - в составе частицы Цит#КЛ, полученной после добавления КЛ к
ЦитС, е - полученной после воздействия метанолом, в результате чего диаметр глобулы увеличился, а атом серы Met80
удалился от гема и координационная железосерная связь была разрушена); нижний ряд - частицы Цит#КЛ, выпавшие в
осадок из водной среды (ж), экстрагированные в неполярный растворитель (з) и образовавшиеся в мембранах митохон#
дрий (и) идентичны по строению тем, что образовывались в водном растворе (д). Стрелки: 1 - отдаление остатков Tyr и
Trp от гема, приводящее к появлению характерной флуоресценции в метанольном растворе ЦитС; 2 - образование час#
тиц Цит#КЛ (д) при добавлении к водному раствору ЦитС метанольного раствора КЛ (светло#серый круг - глобула ЦитС
в нативном состоянии, темно#серый - расплавленная глобула ЦитС, пунктирный контур - диаметр частицы Цит#КЛ с
соотношением липид/белок, равном приблизительно 13/1, сплошной черный контур - диаметр частицы Цит#КЛ с соот#
ношением липид/белок, равном приблизительно 60/1); 3 - отдаление остатка Met80 от гема и разрыв координационной
связи между атомом гемового железа >Fe и атомом серы Met80, приводящий к уменьшению поглощения при 699 нм в ме#
танольном растворе ЦитС, 3* - возвращение глобулы ЦитС из расплавленного состояния в исходное, как следует из дан#
ных на рис. 4; 4, 5 и 6 - переход частиц Цит#КЛ из водного раствора в кристаллический осадок, неполярный раствори#
тель и мембрану митохондрий, соответственно
гивает и эти аминокислотные остатки, тем са#
ном растворителе подтверждается эксперимен#
мым влияя на образование комплекса Цит#КЛ и
тальными данными, а структура Цит#КЛ, вклю#
на проявление им свойств пероксидазы. В рабо#
ченного в липидный слой мембраны митохонд#
те [29] взаимодействие ЦитС с КЛ приводило к
рий, в настоящее время остаётся предположи#
повышению проницаемости липидных везикул,
тельной. На схеме видно, что рассмотренные в
авторы также отмечают разрыв железосерных
данной статье частицы относятся к трем типам
связей.
структур: 1) глобула ЦитС в нативном состоянии
Вся совокупность полученных в работе ре#
(верхний ряд), 2) ЦитС в состоянии «расплав#
зультатов, а также данные литературы позволили
ленной глобулы» (структуры г и е) и 3) наносфе#
предложить гипотетическую схему образования
ра Цит#КЛ (структуры д, ж, з, и). При этом ЦитС
и строения частиц Цит#КЛ, находящихся в раз#
в составе наносферы Цит#КЛ находится, судя по
личных условиях (рис.5). При этом структура
всему, в том же состоянии «расплавленной гло#
Цит#КЛ в кристаллическом осадке и неполяр#
булы». Наносферы Цит#КЛ образуются в двух
БИОХИМИЯ том 84 вып. 8 2019
1174
ВЛАДИМИРОВ и др.
формах с различными размерами - с диаметром
комплекса цитохром с/кардиолипин в хлоро#
~8 нм и ~11 нм. Это связано с различием в тол#
форме необходимо присутствие метанола в вод#
щине липидного слоя, окружающего расплав#
ной фазе не менее 50% [24].
ленную глобулу ЦитС, которая, в свою очередь,
Таким образом, показано, что разрушение
зависит от содержания липида в составе нано#
железосерных связей в ЦитС, помещенном в
сферы (~13 молекул КЛ при диаметре наносфе#
среду, содержащую метанол, является обрати#
ры, равном 8 нм, и ~60 - при диаметре 11 нм).
мым, что говорит об изменении конформации
Образовавшаяся в водном растворе частица (д)
цитохрома с по типу расплавленной глобулы.
может переходить в осадок (стрелка 4) или быть
По#видимому, интактный ЦитС в водном раст#
переведена в неполярный растворитель (стрелка 5).
воре неспособен «вытягивать» молекулы кар#
При этом данные о частицах в микрокристалли#
диолипина из липидного бислоя, например, из
ческом осадке Цит#КЛ и о частицах Цит#КЛ в
сформировавшихся липосом, но при плавле#
неполярных растворителях указывают на то, что
нии глобулы гемопротеина он приобретает
в обоих случаях частицы имеют одинаковые раз#
большее сродство к кардиолипину и легче об#
меры, а также, вероятно, и одинаковое строение.
разует с ним комплекс. Выяснение механизма
В пользу последнего предположения говорят
процессов формирования комплекса Цит#КЛ в
данные измерения флуоресценции тирозиновых
митохондриях нуждается в дальнейших иссле#
и триптофановых остатков цитохрома и погло#
дованиях.
щения цитохрома при длине волны 699 нм [21].
Ранее нами было сделано предположение, что
Цит#КЛ, будучи гидрофобной наносферой,
Финансирование. Работа выполнена при фи#
встраивается в липидный бислой внутренней
нансовой поддержке Российского научного
мембраны митохондрий и там осуществляет ка#
фонда (грант 17#74#10248), а также с использо#
тализ пероксидации липидов, находясь в гидро#
ванием оборудования Центра коллективного
фобном окружении (стрелка 6) [11].
пользования ФНИЦ Кристаллография и фото#
Следует отметить, что влияние рН на свой#
ника РАН и при частичной поддержке Мини#
ства комплекса Цит#КЛ отмечали и другие ав#
стерства науки и высшего образования в рамках
торы. Например, в работе [30] показано, что
выполнения работ по Государственному зада#
только при значениях pH выше 6 (что, по на#
нию ФНИЦ Кристаллография и фотоника РАН
шим данным, полученным с помощью МУР, со#
в части программного обеспечения анализа
ответствует наносферам с диаметром 11,35 ±
данных малоуглового рассеяния, а также проек#
0,21, но не 8,07 ± 0,03 нм) наблюдалось образо#
та ESRF BAG MX#2079 в части измерений ма#
вание синглетного кислорода при работе ком#
лоуглового рассеяния на станции BioSAXS
плекса Цит#КЛ, окисляющего кардиолипин,
BM29.
что свидетельствует о различиях в фермента#
Конфликт интересов. Авторы заявляют об от#
тивных свойствах Цит#КЛ в зависимости от со#
сутствии конфликта интересов.
отношения липид/белок, которое, в свою оче#
Соблюдение этических норм. Настоящая
редь, зависит от pH.
статья не содержит описания каких#либо иссле#
Согласно полученным в нашей лаборатории
дований с использованием людей или животных
результатам, для успешного получения раствора
в качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Ouyang, L., Shi, Z., Zhao, S., Wang, F.T., Zhou, T.T., Liu, B.,
catalytic interactions of cytochrome c with cardiolipin and
and Bao, J.K. (2012) Programmed cell death pathways in
phosphatidylserine, Free Radic. Biol. Med.,
37,
cancer: a review of apoptosis, autophagy and programmed
1963-1985, doi: 10.1016/j.freeradbiomed.2004.08.016.
necrosis, Cell Prolif., 45, 487-498, doi: 10.1111/ j.1365#
4.
Kagan, V.E., Tyurin, V.A., Jiang, J., Tyurina, Y.Y., Ritov, V.B.,
2184.2012.00845.x.
Amoscato, A.A., Osipov, A.N., Belikova, N.A., Kapralov, A.A.,
2.
Saleem, M., Asif, J., Asif, M., and Saleem, U. (2018)
and Kini, V. (2005) Cytochrome c acts as a cardiolipin oxy#
Amygdalin, from apricot kernels, induces apoptosis and
genase required for release of proapoptotic factors, Nature
causes cell cycle arrest in cancer cells: an updated review,
Chem. Biol., 1, 223-232, doi: 10.1038/nchembio727.
Anticancer Agents Med. Chem., 18, 1650-1655, doi: 10.2174/
5.
Kagan, V.E., Bayir, A., Bayir, H., Stoyanovsky, D.,
1871520618666180105161136.
Borisenko, G.G., Tyurina, Y.Y., Wipf, P., Atkinson, J.,
3.
Kagan, V.E., Borisenko, G.G., Tyurina, Y.Y., Tyurin, V.A.,
Greenberger, J.S., Chapkin, R.S., and Belikova, N.A.
Jiang, J., Potapovich, A.I., Kini, V., Amoscato, A.A., and
(2009) Mitochondria#targeted disruptors and inhibitors of
Fujii, Y. (2004) Oxidative lipidomics of apoptosis: redox
cytochrome c/cardiolipin peroxidase complexes: a new
БИОХИМИЯ том 84 вып. 8 2019
НАНОСФЕРЫ КОМПЛЕКСА ЦИТОХРОМА c С КАРДИОЛИПИНОМ
1175
strategy in anti#apoptotic drug discovery, Mol. Nutr. Food
Biophys. Acta, 1808, 2147-2155, doi: 10.1016/j.bbamem.
Res., 53, 104-114, doi: 10.1002/mnfr.200700402.
2011.04.009.
6.
Brown, L.R., and Wuthrich, K. (1977) NMR and ESR
18.
Belikova, N.A., Vladimirov, Y.A., Osipov, A.N., Kapralov, A.A.,
studies of the interactions of cytochrome c with mixed car#
Tyurin, V.A., Potapovich, M.V., Basova, L.V., Peterson, J.,
diolipin#phosphatidylcholine vesicles, Biochim. Biophys.
Kurnikov, I.V., and Kagan, V.E. (2006) Peroxidase activity
Acta, 468, 389-410, doi: 10.1016/0005#2736(77)90290#5.
and structural transitions of cytochrome c bound to cardio#
7.
Sinibaldi, F., Howes, B.D., Piro, M.C., Polticelli, F.,
lipin#containing
membranes, Biochemistry,
45,
Bombelli, C., Ferri, T., Coletta, M., Smulevich, G., and
4998-5009, doi: 10.1021/bi0525573.
Santucci, R. (2010) Extended cardiolipin anchorage to
19.
Kapralov, A.A., Kurnikov, I.V., Vlasova, I.I., Belikova, N.A.,
cytochrome c: a model for protein#mitochondrial mem#
Tyurin, V.A., Basova, L.V., Zhao, Q., Tyurina, Y.Y., Jiang, J.,
brane binding, J. Biol. Inorg. Chem., 15, 689-700,
Bayir, H., Vladimirov, Y.A., and Kagan, V.E. (2007) The
doi: 10.1007/s00775#010#0636#z.
hierarchy of structural transitions induced in cytochrome c
8.
Mandal, A., Hoop, C.L., DeLucia, M., Kodali, R.,
by anionic phospholipids determines its peroxidase activa#
Kagan, V.E., Ahn, J., and van der Wel, P.C. (2015)
tion and selective peroxidation during apoptosis in cells,
Structural changes and proapoptotic peroxidase activity of
Biochemistry, 46, 14232-14244, doi: 10.1021/bi701237b.
cardiolipin#bound mitochondrial cytochrome c, Biophys. J.,
20.
Proskurnina, E.V., Alekseev, A.V., Demin, E.M.,
109, 1873-1884, doi: 10.1016/j.bpj.2015.09.016.
Izmailov, D.Y., and Vladimirov, Y.A. (2013) Cyt#CL com#
9.
Hanske, J., Toffey, JR., Morenz, A.M., Bonilla, A.J.,
plex: Peroxidase activity and role in lipid peroxidation,
Schiavoni, K.H., and Pletneva, E.V. (2012) Conformation#
FEBS J., 280, 264.
al properties of cardiolipin#bound cytochrome c, Proc.
21.
Vladimirov, G.K., Vikulina, A.S., Volodkin, D., and
Natl. Acad. Sci. USA, 109, 125-130, doi: 10.1073/pnas.
Vladimirov, Y.A. (2018) Structure of the complex of
1112312108.
cytochrome c with cardiolipin in non#polar environment,
10.
Kagan, V.E., Bayir, H.A., Belikova, N.A., Kapralov, O.,
Chem. Phys. Lipids, 214, 35-45, doi: 10.1016/j.chemphyslip.
Tyurina, Y.Y., Tyurin, V.A., Jiang, J., Stoyanovsky, D.A.,
2018.05.007.
Wipf, P., Kochanek, P.M., Greenberger, J.S., Pitt, B.,
22.
Konarev, P.V., Volkov, V.V., Sokolova, A.V., Koch, M.H.J., and
Shvedova, A.A., and Borisenko, G. (2009) Cytochrome c/
Svergun, D.I. (2003) PRIMUS: a Windows PC#based system
cardiolipin relations in mitochondria: a kiss of death, Free
for small#angle scattering data analysis, J. Appl. Crystallogr., 36,
Radic. Biol. Med.,
46,
1439-1453, doi:
10.1016/
1277-1282, doi: 10.1107/S002188980301277923.
j.freeradbiomed.2009.03.004.
23.
Folch, J., Lees, M., and Sloane Stanley, G.H. (1957)
11.
Vladimirov, Y.A., Proskurnina, E.V., and Alekseev, A.V.
A simple method for the isolation and purification of total
(2013) Molecular mechanisms of apoptosis. Structure of
lipides from animal tissues, J. Biol. Chem., 226, 497-509.
cytochrome c-cardiolipin complex, Biochemistry (Moscow),
24.
Vikulina, A.S., Alekseev, A.V., Proskurnina, E.V., and
78, 1086-1097, doi: 10.1134/S0006297913100027.
Vladimirov, Y.A. (2015) The complex of cytochrome c with
12.
Jemmerson, R., Liu, J., Hausauer, D., Lam, K.P.,
cardiolipin in non#polar environment, Biochemistry
Mondino, A., and Nelson, R.D. (1999) A conformational
(Moscow), 80, 1298-1302, doi: 10.1134/S0006297915100107.
change in cytochrome c of apoptotic and necrotic cells is
25.
Ali, S., Farooqi, H., Prasad, R., Naime, M., Routray, I.,
detected by monoclonal antibody binding and mimicked
Yadav, S., and Ahmad, F. (2010) Boron stabilizes peroxide
by association of the native antigen with synthetic phos#
mediated changes in the structure of heme proteins, Int. J.
pholipid vesicles, Biochemistry,
38,
3599-3609,
Biol. Macromol., 47, 109-115, doi: 10.1016/j.ijbiomac.
doi: 10.1021/bi9809268.
2010.05.013.
13.
Tuominen, E.K., Zhu, K., Wallace, C.J., Clark#Lewis, I.,
26.
Kobayashi, H., Nagao, S., and Hirota, S.
(2016)
Craig, D.B., Rytomaa, M., and Kinnunen, P.K. (2001)
Characterization of the cytochrome c membrane#binding
ATP induces a conformational change in lipid#bound
site using cardiolipin#containing bicelles with NMR,
cytochrome c, J. Biol. Chem., 276, 19356-19362, doi:
Angewandte Chem. Intern. Edition, 55, 14019-14022,
10.1074/jbc.M100853200.
doi: 10.1002/anie.201607419.
14.
Balakrishnan, G., Hu, Y., Oyerinde, O.F., Su, J., Groves, J.T.,
27.
Проскурнина Е.В., Проскурнин М.А., Алексеев А.В.,
and Spiro, T.G. (2007) A conformational switch to beta#
Галимова В.Р., Владимиров Ю.А. (2018) Определение
sheet structure in cytochrome c leads to heme exposure.
состава комплекса цитохрома c с кардиолипином при
Implications for cardiolipin peroxidation and apoptosis,
помощи спектрофотометрии и термолинзовой спект#
J. Am. Chem. Soc., 129, 504-505, doi: 10.1021/ja0678727.
рометрии, Технологии живых систем, 15, 27-33.
15.
Hong, Y., Muenzner, J., Grimm, S.K., and Pletneva, E.V.
28.
Sinibaldi, F., Howes, B.D., Droghetti, E., Polticelli, F.,
(2012) Origin of the conformational heterogeneity of car#
Piro, M.C., Di Pierro, D., Fiorucci, L., Coletta, M.,
diolipin#bound cytochrome c, J. Am. Chem. Soc., 134,
Smulevich, G., and Santucci, R. (2013) Role of lysines in
18713-18723, doi: 10.1021/ja307426k.
cytochrome c#cardiolipin interaction, Biochemistry, 52,
16.
Vladimirov, Y.A., Nol’, Y.T., and Volkov, V.V.
(2011)
4578-4588, doi: 10.1021/bi400324c.
Protein-lipid nanoparticles that determine whether cells
29.
Kitt, J.P., Bryce, D.A., Minteer, S.D., and Harris, J.M.
will live or die, Crystallogr. Rep.,
56,
553-559,
(2017) Raman spectroscopy reveals selective interactions
doi: 10.1134/S1063774511040250.
of cytochrome c with cardiolipin that correlate with mem#
17.
Kapralov, A.A., Yanamala, N., Tyurina, Y.Y., Castro, L.,
brane permeability, J. Am. Chem. Soc., 139, 3851-3860,
Samhan#Arias, A., Vladimirov, Y.A., Maeda, A.,
doi: 10.1021/jacs.7b00238.
Weitz, A.A., Peterson, J., Mylnikov, D., Demicheli, V.,
30.
Miyamoto, S., Nantes, I.L., Faria, P.A., Cunha, D.,
Tortora, V., Klein#Seetharaman, J., Radi, R., and Kagan, V.E.
Ronsein, G.E., Medeiros, M.H., and Di Mascio, P. (2012)
(2011) Topography of tyrosine residues and their involve#
Cytochrome c#promoted cardiolipin oxidation generates
ment in peroxidation of polyunsaturated cardiolipin in
singlet molecular oxygen, Photochem. Photobiol. Sci., 11,
cytochrome c/cardiolipin peroxidase complexes, Biochim.
1536-1546, doi: 10.1039/c2pp25119a.
БИОХИМИЯ том 84 вып. 8 2019
1176
ВЛАДИМИРОВ и др.
COMPARISON OF DIMENSIONS AND PROPERTIES OF NANOSPHERES
OF THE CYTOCHROME c COMPLEX WITH CARDIOLIPIN
IN SEDIMENT AND IN NON7 POLAR MEDIUM
G. K. Vladimirov1,2,3*, V. E. Remenshchikov2, A. M. Nesterova2,3,
V. V. Volkov2,4, and Yu. A. Vladimirov1,2,3,5
1 Lomonosov Moscow State University, Faculty of Fundamental Medicine,
119991 Moscow, Russia; E'mail: ura'vladimirov@yandex.ru
2 Shubnikov Institute of Crystallography of Federal Scientific Research Centre “Crystallography and Photonics”,
Russian Academy of Sciences, 119333 Moscow, Russia
3 Institute for Regenerative Medicine, Sechenov First Moscow State Medical University (Sechenov University),
Ministry of Public Health of Russian Federation, 119991 Moscow, Russia
4 National Research Center “Kurchatov Institute”, 123182 Moscow, Russia
5 Pirogov Russian National Research Medical University,
Ministry of Public Health of Russian Federation, 117997 Moscow, Russia
Received February 20, 2019
Revised March 28, 2019
Accepted April 12, 2019
Apoptosis, the major type of programmable cell death, plays a critical role in the renewal of cells in our body and in
the elimination of defective or degenerated cells, including cancer cells. One of the earliest events of apoptosis involves
lipid peroxidation in the inner mitochondrial membrane, which is catalyzed by a complex of cytochrome c (CytC)
with mitochondrial phospholipid - cardiolipin (CL). According to our data obtained earlier, when mixing solutions
of CytC and CL, nanospheres of the CytC complex with CL (Cyt-CL) with a diameter of 11#12 nm are formed,
which consist of a molten protein molecule and a CL monolayer. In this study, using the dynamic light scattering of
the Cyt-CL solution in chloroform and the small#angle X#ray scattering in the Cyt-СL sediment, it was shown that
in both cases the Cyt-CL forms nanospheres with 8# and 11#nm diameters, which corresponds to the previously
obtained lipid/protein ratio, respectively, 13 : 14 and 35 : 50. These data show that the Cyt-CL nanospheres are
formed not only during the crystallization of the complex but also in the hydrophobic environment. CytC in the com#
plex has a molten globule structure, as evidenced by the appearance of the tryptophan and tyrosine fluorescence,
which is absent in the native protein due to the Förster resonance transfer of the electron excitation energy to heme.
Under these conditions, the coordination bond between the heme iron and sulfur of methionine#80 is broken in the
Cyt-CL complex (the absorption band about 700 nm vanishes). When methanol is added to the aqueous CytC solu#
tion (water-methanol, 1 : 1 v/v), this iron-sulfur bond is broken also. The latter changes were reversible, which sup#
ports the conclusion that changes of CytC conformation in methanol#containing solutions correspond to the molten
globule type.
Keywords: apoptosis, cytochrome c, cardiolipin, Cyt-CL complex, hydrophobic environment, dynamic light scatter#
ing, small#angle X#ray scattering, molten globule
БИОХИМИЯ том 84 вып. 8 2019