БИОХИМИЯ, 2019, том 84, вып. 8, с. 1177 - 1188
УДК 577.6
ТЕРМОСТАБИЛЬНАЯ ЛИХЕНАЗА Clostridium thermocellum
ДОПУСКАЕТ ОДИНОЧНЫЕ И ТАНДЕМНЫЕ ИНСЕРЦИИ
БЕЛКОВЫХ ДОМЕНОВ*
© 2019
О.С. Павленко1**, O.A. Гра1, O.Н. Мустафаев2, К.В. Кабарбаева1,
Н.С. Садовская1, A.А. Тюрин1, В.С. Фадеев1, И.В. Голденкова4Павлова1**
1 Институт физиологии растений им. К.А. Тимирязева РАН, 127276 Москва, Россия;
электронная почта: Helliga.p@gmail.com и irengold58@gmail.com
2 Бакинский государственный университет, AZ 1148 Баку,
Азербайджан; электронная почта: orkhan@bioset.org
Поступила в редакцию 03.03.2019
После доработки 29.04.2019
Принята к публикации 30.04.2019
В этом исследовании лихеназа Clostridium thermocellum (эндо β 1,3;1,4 глюкан D гликозилгидролаза,
ЕС 3.2.1.73 (Р29716)) апробирована в роли белка, принимающего в двух областях инсерцию двух модельных
флуоресцентных белков (EGFP и TagRFP). Функциональная укладка всех доменов подтверждена сохране
нием ферментативной активности лихеназы и флуоресценции EGFP и TagRFP. Полученные результаты
убедительно свидетельствуют, что (i) два экспериментально апробированных петлевых региона лихеназы
могут служить областями для инсерции белков без нарушения ее фолдинга in vivo, (ii) лихеназа способна до
пускать не только одиночные, но и тандемные вставки крупных белковых доменов. Высокая активность и
термостабильность, эффективный рефолдинг in vitro лихеназы делает ее привлекательной в качестве ново
го принимающего «host» белка при использовании подхода инсерционного слияния доменов в инженерии
многофункциональных белков.
КЛЮЧЕВЫЕ СЛОВА: лихеназа, флуоресцентные белки, одиночные и тандемные инсерции белков.
DOI: 10.1134/S0320972519080104
Одно из ключевых направлений в инжене
который предназначен для разделения белков и
рии белков связано с дизайном и конструирова
поддержания благоприятного взаимодействия
нием слитых белков, биомолекул, состоящих из
между ними. Имеется немало успешных приме
двух или более белков и/или пептидов, которые
ров создания рекомбинантных белков с исполь
синтезируются как единая белковая конструк
зованием стратегии прямого «слияния» для раз
ция. Для инженерии таких белков используют
личного биотехнологического [2, 3, 5] и биофар
стратегию прямого «слияния» белков (или стра
мацевтического применения [6-8].
тегия «голова к хвосту») или стратегию инсер
Однако стратегия «голова к хвосту» может
ции доменов [1-3]. Стратегия «голова к хвосту»
терпеть неудачу вследствие того, что такие ре
достаточно проста и применяется в тех случаях,
комбинантные белки более часто подвержены
когда гибкие и неструктурированные N или
протеолитической деградации и структурно
С концевые области белка выступают в качестве
нестабильны [9]. В таких случаях более полез
«моста», чтобы обеспечить достаточное прост
ной может быть стратегия инсерции доменов
ранство между доменами белков для их пра
[10-12]. Эта стратегия основана на инсерции
вильной укладки [4]. Помимо этого, возможно
одного белкового домена («guest» домен) в дру
объединение белков через линкерный пептид,
гой белок (принимающий белок или «host» до
мен) [11]. Стоит отметить, что эта стратегия яв
ляется более сложной для дизайна, чем страте
Принятые сокращения: PGK - фосфоглицератки
наза, BLA - β лактамаза, СРБ - суммарный растворимый гия «прямого» слияния [12, 13]. Так, для успеш
белок.
ного создания белков путем инсерции домена
* Приложение к статье на английском языке опубликовано крайне важно предсказать потенциальные об
ласти интеграции в принимающем белке. Для
bio.msu.ru/biokhimiya/) и на сайте издательства Springer
поиска оптимальных сайтов для «врезания» в
2019.
белок предложено и успешно апробировано
** Адресат для корреспонденции.
несколько подходов: случайный и направлен
9
1177
1178
ПАВЛЕНКО и др.
ный мутагенез, в частности, комбинаторные
чувствительными методами как качественно,
подходы к вставке домена, которые используют
так и количественно [5, 13, 28]; (iii) компактная
DNAseI [14-16] или транспозоны [17, 18] для
структура белковой молекулы, согласно кото
введения случайных разрывов в сегмент ДНК
рой белок имеет несколько петлевых областей
мишени; циклические перестановки в белке,
[5, 13, 28, 29]; (iv) участки двух петлевых облас
которые дают представление о том, какие струк
тей (53 и 99 а.о.) каталитического домена тер
турные элементы важны для функционирова
мостабильной лихеназы допускают и введение
ния белка, а также позволяют найти сайты для
круговой перестановки, и выдерживают внут
вставки других белков [19, 20]; in silico алгорит
реннюю интеграцию небольших пептидов
мы для предсказания участков интеграции на
(6 а.о.), в том числе и тандемную, с сохранением
основе оценки низкой энергии нарушения
активности и термостабильности фермента [28].
структурно функциональных элементов белка
На основании ранее полученных нами ре
[21, 22] и ряд других.
зультатов о сохранении лихеназой активности и
Согласно текущему мнению, белки с инсер
термостабильности при внутренней интеграции
цией других белковых доменов стали уникаль
небольших пептидов высказано предположение
ными моделями для фундаментальных исследо
о толерантности этого белка к более крупным
ваний, в частности, (i) для изучения механизма
внутренним инсерциям в двух петлевых облас
кооперативной укладки белков и их функцио
тях (53 и 99 а.о.) [28]. Для того чтобы проверить
нирования [23]; (ii) для конструирования бел
эту гипотезу, необходимо оценить могут ли эти
ков переключателей [16]; (iii) для увеличения
области лихеназы выдержать внутреннюю ин
стабильности белковых молекул [13, 24] и
теграцию крупных доменов без драматической
(iv) для конструирования бифункциональных
потери ферментом активности и термотолерант
белков [1-3, 11, 13, 25].
ности.
Отметим, что стратегия инсерции доменов
использована на немногочисленном списке
белков, которые применены в качестве прини
МЕТОДЫ ИССЛЕДОВАНИЯ
мающего или «host» домена. Среди них есть как
умеренно стабильные [18, 23], так и термоста
Генно4инженерные методы. В работе исполь
бильные белки [13, 24-26]. Большинство «host»
зованы стандартные процедуры молекулярного
белков исследованы за счет вставки одного
клонирования [30], праймеры («Evrogen», Рос
«guest» домена, и проявляют к ней полную или
сия), ферменты и наборы реагентов («Promega»,
частичную толерантность, оцененную по сохра
США), («Fermentas», Литва), («QIAexpress»,
нению функциональной укладки обоих («host» и
США) и («Novagene», США). Плазмиды pQE
«guest») белков. Вставка одновременно двух
NC L 53 99 [28], pTagRFP C («Evrogen», Рос
«guest» доменов изучена для двух «host» бел
сия) и pQE egfp [31] использованы в качестве
ков - фосфоглицераткиназы (PGK) и β лакта
источников licB, rfp и egfp генов соответственно.
мазы (BLA): PGK допускает инсерцию двух раз
В последовательностях генов мишеней (RFP и
ных «guest» доменов в двух разных областях
EGFP) стартовый и стоп кодоны удалены.
PGK [12], а BLA - тандемную инсерцию одного
Конструирование лихеназы со вставками до4
и того же белка, соединенную через короткий
менов (domain insertion). Плазмида pQE LicB/
гибкий линкер в одной области BLA [17, 27]. Та
53EGFP сконструирована в два этапа. Первона
ким образом, список принимающих белков, в
чально последовательность гена egfp клонирова
частности, белков, способных допускать не
на с помощью ПЦР с использованием плазмиды
только одиночные, но и тандемные вставки двух
pQE egfp в качестве матрицы и прямого EGF
«guest» доменов, крайне скромный.
A(F) и обратного EGF A(R) праймеров с вве
В этом исследовании мы применили подход
денными сайтами рестрикции BamHI и ApaI
инсерции домена к термостабильной лихеназе
(табл. 1). Ампликон клонирован в виде фраг
(эндо β 1,3;1,4 глюкан D гликозилгидролаза
мента BamHI - ApaI в плазмиду pQE NC L 53 99
ЕС 3.2.1.73 (Р29716)) Clostridium thermocellum,
(далее обозначали плазмиду как pQE LicB M)
как «host» белку, с использованием двух модель
[28], гидролизованную теми же эндонуклеазами
ных флуоресцентных белков. Привлекатель
рестрикции. Плазмида pQE LicB/99EGFP так
ность использования термостабильной лихена
же сконструирована в два этапа. Первоначально
зы для такого подхода базируется на следующем:
последовательность гена egfp получена с по
(i) белок сохраняет функциональность при N и
мощью ПЦР с использованием плазмиды pQE
C концевых слияниях [5, 28]; (ii) высокие тер
egfp в качестве матрицы и прямого EGF B(F) и
мостабильность и удельная активность лихена
обратного EGF B(R) праймеров с введенными
зы, которые могут быть оценены простыми и
сайтами рестрикции HindIII и SmaI (табл. 1).
БИОХИМИЯ том 84 вып. 8 2019
ЛИХЕНАЗА ДОПУСКАЕТ ТАНДЕМНЫЕ ИНСЕРЦИИ БЕЛКОВ
1179
Ампликон клонирован в виде фрагмента
Таблица 1. Последовательности праймеров, использован
ных в работе
HindIII - SmaI в плазмиду pQE LicB M [29],
гидролизованную теми же эндонуклеазами
Название
5'-3' последовательность праймеров
рестрикции. Плазмида pQE LicB/53EGFP/
праймера
99RFP сконструирована в два этапа. Первона
чально последовательность гена rfp получена с
EGF A(F)
GGATCCGTGAGCAAGGGCGA
помощью ПЦР с использованием плазмиды
EGF A(R)
GGGCCCCTTGTACAGCTCGT
pTagRFP C в качестве матрицы («Evrogen», Рос
EGF B(F)
AAGCTTGTGAGCAAGGGCGAG
сия) и прямого RFP(F), и обратного RFP(R)
праймеров с введенными сайтами рестрикции
EGF B(R)
CCCGGGCTTGTACAGCTCGTCC
HindIII и SmaI (табл. 1). Ампликон клонирован в
RFP(F)
AAGCTTGTGAGTAAAGGTGAAGA
виде фрагмента HindIII - SmaI в плазмиду pQE
RFP(R)
CCCGGGGTTCAATTTGTGACCTAG
LicB/53EGFP, гидролизованную теми же эндо
нуклеазами рестрикции. Рамки считывания ге
нов в плазмидах LicB/53EGFP, LicB/99EGFP и
LicB/53EGFP/99RFP проверены с помощью
только с негидролизованным лихенаном. Энзи
секвенирования («Evrogen», Россия).
мограммы получали окрашиванием геля после
Экспрессия и очистка белков. Штамм E. coli
разделения белков в 12% ном ДСН ПААГ в
XL1 Blue («Stratagene», США), несущий плаз
присутствии 0,1% лихенана, как описано ранее
миды pQE LicB M, pQE EGFP, pQE LicB/
[13, 28]. Кратко: лихенан (0,1%) добавляли в
53EGFP, pQE LicB/99EGFP и pQE LicB/
разделяющий гель до полимеризации. Прогре
53EGFP/99RFP, растили до OD600 0,6 при 37 °C в
тые белковые лизаты, содержащие равное коли
среде LB («Amresco», США) и затем индуциро
чество суммарного белка (определено по методу
вали 0,5 мM изопропил β D 1 тиогалактазидом
Бредфорда [32]), смешивали с буфером (Tris HCl,
(IPTG) при 37 °C в течение 16 ч. Клетки отделя
pH 6,8; 12,5% (v/v) глицерин, 0,5% SDS, 1,25%
ли от среды на центрифуге 5910R («Eppendorf»,
β меркаптоэтанол) и инкубировали при 70 °С
Германия), дважды промывали в буфере 50 мM
40 мин. После электрофореза гель инкубирова
Tris HCl, pH 8,0 и ресуспендировали в том же
ли при 70 °C 1 ч. Ферментативная активность
буфере. Разрушение клеток осуществляли
определена за счет окрашивания геля 0,5% ным
Френч прессом («Aminco», США) и очищали
раствором Конго красного с последующим от
центрифугированием. Полученный суммарный
мыванием в 1 М NaCl. При этом на месте актив
растворимый белок (СРБ) инкубировали при
ного белка обнаруживается просветленная об
65 °C 30 мин и очищали центрифугированием.
ласть, так как краситель связывается только с
Осадок высушивали, перерастворяли в 50 мM
негидролизованным лихенаном. Молекулярная
Tris HCl, pH 8,0 и использовали для очистки на
масса белков определена с помощью маркеров
колонке с Ni2+ NTA агарозой («Qiagen», Герма
Prestained Protein Ladder («Fermentas», Латвия).
ния), уравновешенной 50 мM Tris HCl, pH 8,0.
Активность лихеназы определена методом
Белки элюировали тем же буфером, содержа
восстанавливающих сахаров с использованием
щим линейный градиент (20-200 мM) имидазо
лихенана в качестве субстрата. Определение
ла. Элюированные белки подвергали диализу
восстанавливающих сахаров проводили с ис
против 50 мМ Tris HCl, pH 8,0, и очищенный
пользованием 3,5 динитросалициловой кисло
белок (ОБ) использовали в дальнейших экспе
ты (DNS) и контролировали калориметрически
риментах.
при 540 нм [13, 29] в устройстве CLARIO star
Аналитическая характеристика белков. Ча
(«BMGLABTECH», Германия). За единицу ак
шечный тест проводили, как описано ранее [13,
тивности принимали количество фермента, об
28]. Кратко: бактериальные колонии переноси
разующее 1 мкмоль восстанавливающих сахаров
ли на чашки Петри с агаризованной средой LB
за 1 мин в стандартных условиях. Стандартный
методом реплики и выращивали колонии при
анализ проводили в течение 10 мин в 50 мМ Tris
37 °С в течение ночи. Затем чашки заливали
HCl (рН 8,0) при 70 °С. Концентрацию белка
верхним слоем агара, содержащим 0,05% лихе
определяли по методу Бредфорда с бычьим сы
нана («Megazyme», Ирландия), и инкубировали
вороточным альбумином в качестве стандарта
1-16 ч. Активность лихеназы определяли окра
[32]. Термостабильность ферментов, инкубиро
шиванием гелей 0,5% ным раствором Конго
ванных в 50 мМ Tris HCl (pH 8,0) при 70 °C в те
красного («Sigma», США) с последующим от
чение 4 ч, оценивали по их активности, опреде
мыванием в 1М NaCl [13, 28]. При этом на мес
ленной стандартным анализом: оценивали от
те активного белка обнаруживается просветлен
носительную активность (%) каждого образца
ная область, так как краситель связывается
белка как (активность лихеназы в данной точке
БИОХИМИЯ том 84 вып. 8 2019
9*
1180
ПАВЛЕНКО и др.
инкубации при 70 °C)/(активность лихеназы без
меньших квадратов для случая нелинейной ре
инкубации) × 100. Обработка этанолом: СРБ об
грессии [34].
рабатывали тремя объемами 96% ного этанола
Статистический анализ. Все процедуры про
при 4 °С в течение 8-10 ч и последующим цент
водили как минимум в трех независимых экспе
рифугированием при 12 °С и 100 g («Eppendorf»,
риментах, и результаты выражали в виде средне
Германия). Осадок высушивали при комнатной
го значения ± стандартное отклонение, если не
температуре и растворяли в 50 мМ Tris HCl,
указано особо.
pH 8,0, и определяли активность лихеназы, как
указано выше.
Флуоресценцию ЕGFP в бактериальных
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
трансформантах определяли визуально при осве
щении УФ светом. Спектры флуоресценции
Дизайн термостабильной лихеназы с инсерцией
ЕGFP регистрировали на спектрофлуориметре
доменов флуоресцентных белков. В этом исследо
Саrу Eclipse («Varian B.V.», Австралия). Измере
вании термостабильная лихеназа C. thermocellum
ния получены при длине волн возбуждения и
использована как потенциальный принимаю
эмиссии 488 и 507 нм соответственно. Флуорес
щий «host» белок. Лихеназа - мономерный бе
ценцию RFP у бактерий регистрировали с ис
лок, состоящий из 215 а.о., он имеет компактную
пользованием микроскопа Axio Imager Z2
структуру белковой молекулы и характеризуется
(«Zeiss», Германия), оснащенного цифровой ка
повышенной термостабильностью и высокой
мерой AxioCam MR и фильтром № 43. Модуль
удельной активностью. Для инсерции доменов
ApoTome («Zeiss», Германия), позволяющий
использован модифицированный вариант тер
уменьшить очаговую флуоресценцию, исполь
мостабильной лихеназы NC L 53 99 (в этой ра
зовался для регистрации флуоресценции.
боте обозначен как LicB M) с областью вставки
Кинетика процесса рефолдинга. Образцы
небольших пептидов между 53 и 99 а.о. в катали
белков (50 мкМ) нагревали до 95 °С в денатури
тическом домене лихеназы (рис. 1). В этом вари
рующем растворе (8 М мочевина, 1 мМ дитио
анте гена термостабильной лихеназы кодоны для
треитол) в течение 4 мин [33]. Реакции рефол
интегрированных пептидов формируют уни
динга начинали после 100× разведения в том же
кальные сайты рестрикции на уровне нуклео
буфере без мочевины. Кинетика рефолдинга
тидных последовательностей BamHI, ApaI,
лихеназы оценена по восстановлению ее актив
HindIII и SmaI [28]. Такой дизайн последователь
ности с использованием алгоритма Левенберга-
ности позволяет легкое применение подхода ин
Марквардта, который реализует метод наи
серции доменов для лихеназы как «host» белка.
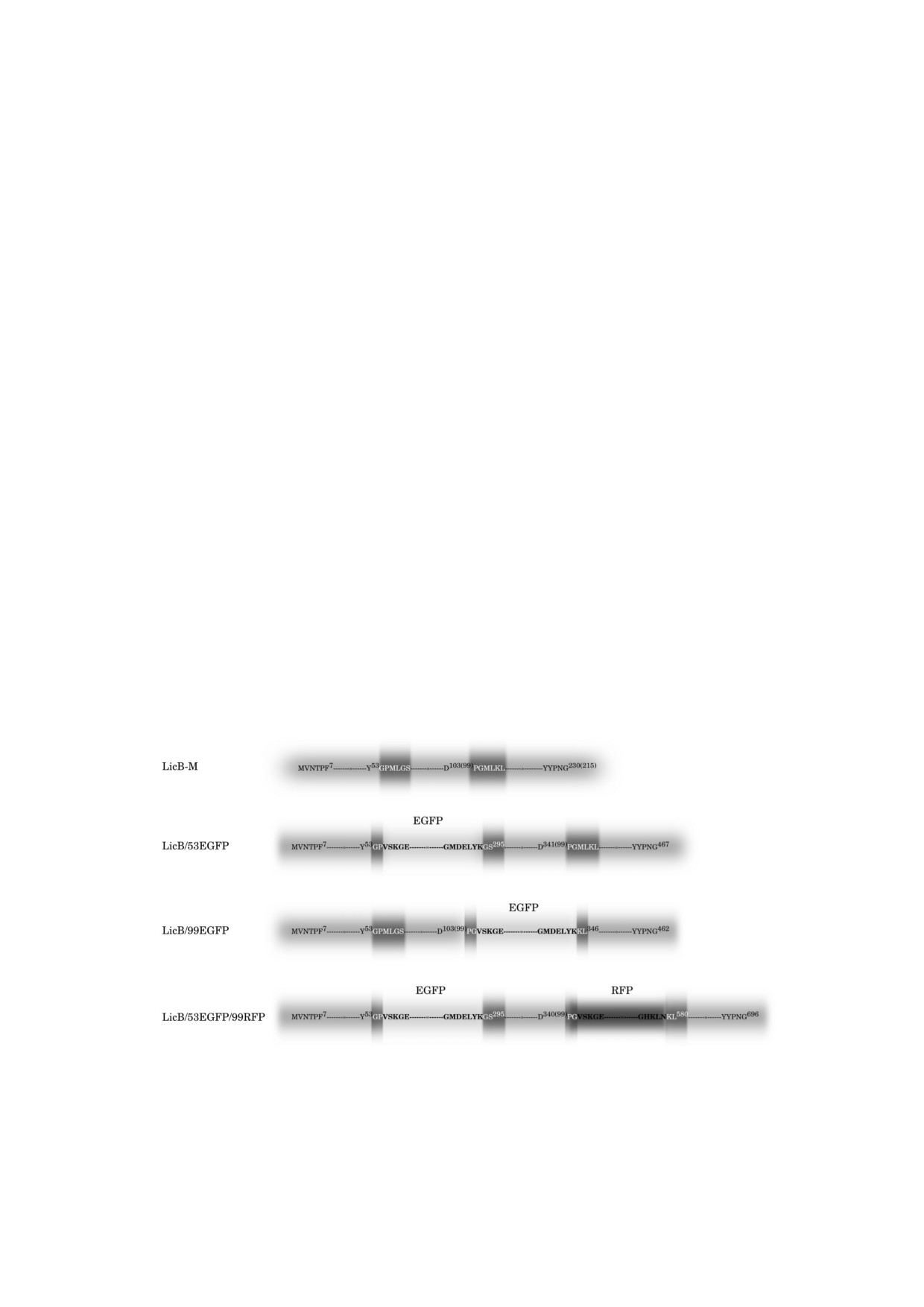
Рис. 1. Последовательности LicB M (исходный вариант лихеназы), LicB/53EGFP и LicB/99EGFP (варианты лихеназы с
инсерцией EGFP в область 53 или 99 а.о. соответственно) и LicB/53EGFP/99RFP (вариант лихеназы с инсерциями EGFP
и TagRFP в область 53 и 99 а.о. соответственно). Светло серые блоки - последовательности лихеназы; темно серые бло
ки - интегрированные малые пептиды или их части; EGFP- зеленый флуоресцентный белок; TagRFP - красный флуо
ресцентный белок. Цифрами у аминокислотных остатков обозначены порядковые номера а.о. в белке, цифрами в скоб
ках обозначены порядковые номера а.о. каталитического домена термостабильной лихеназы
БИОХИМИЯ том 84 вып. 8 2019
ЛИХЕНАЗА ДОПУСКАЕТ ТАНДЕМНЫЕ ИНСЕРЦИИ БЕЛКОВ
1181
Интегрированные домены - EGFP (PDBID:
in vivo. В пользу этого предположения свиде
4KAG) [35] и TagRFP (AMY56669.1) [36] - моно
тельствуют и результаты анализа суммарного
мерные белки, состоящие из 238 и 236 а.о. соот
растворимого белка методом ДСН ПААГ
ветственно, с относительной близостью N и
электрофореза (рис. 2, в), который демонстри
C концевых областей, функционально активная
рует сравнимый уровень экспрессии в E. coli как
укладка которых может быть оценена простой ви
нативных белков LicB M и EGFP, так и вариан
зуализацией за счет их флуоресценции [33, 37, 38].
тов лихеназы с инсерцией домена в LicB/
Методами молекулярного клонирования
53EGFP и LicB/99EGFP.
сконструированы последовательности: LicB/
Используя очищенный белок, оценены
53EGFP и LicB/99EGFP, в которых ген egfp ин
свойства принимающего белка (активность
тегрирован в последовательность гена лихеназы
LicB M) и вставленного белка (спектр флуорес
в область 53 или 99 а.о. соответственно (рис. 1),
ценции EGFP). Первоначально мы провели
т.е. последовательности с одной инсерцией
анализ очищенных белков методом энзимо
EGFP в разных областях лихеназы. Дополни
грамм, на которых детектируются одиночные
тельно сконструирована последовательность
просветленные полосы активности. Это может
LicB/53EGFP/99RFP, которая несет вставки
свидетельствовать о том, что белки LicB/
EGFP и TagRFP в регионах 53 и 99 а.о. каталити
53EGFP и LicB/99EGFP в образцах представле
ческого домена лихеназы соответственно (рис. 1),
ны в форме мономеров (рис. 2, г), а также о том,
т.е. последовательность лихеназы с инсерцией
что молекулярные массы этих белков соответ
двух флуоресцентных белков в двух разных об
ствуют теоретически рассчитанным (~52 кДа).
ластях лихеназы.
Сравнительный анализ удельной активности
Экспрессия и функциональные свойства тер4
очищенных белков показал, что белки LicB/
мостабильной лихеназы с инсерцией одного доме4
53EGFP и LicB/99EGFP, в целом, не демон
на в две разные петлевые области. Первоначаль
стрируют драматического снижения удельной
но мы попытались выяснить: (i) может ли тер
активности по сравнению с исходным вариан
мостабильная лихеназа в принципе быть ис
том LicB M. Так, вариант LicB/53EGFP имеет
пользована в качестве принимающего белка, т.е.
удельную активность 314 ± 10 ед/мг, что сходно
сможет ли лихеназа образовать стабильную и
с таковой для исходного варианта LicB M - 385 ±
функциональную укладку при вставке крупных
11 ед/мг, а активность варианта LicB/99EGFP
доменов; (ii) сохранит ли вставленный домен
составляет 147 + 14 ед/мг, т.е. проявляет лишь
(в нашем случае флуоресцентный белок) спо
~40% активности по сравнению с LicB M. Та
собность к функциональной укладке; (iii) есть
ким образом, у обоих вариантов белков (LicB/
ли преференция у одного из выбранных сайтов
53EGFP и LicB/99EGFP) с инсерцией одного до
интеграции в термостабильной лихеназе, как
мена активность лихеназы, в целом, сохранена.
принимающем домене?
Функциональность EGFP оценена в образ
Для того, чтобы ответить на эти вопросы, ис
цах СРБ: EGFP, LicB/53EGFP и LicB/99EGFP.
пользовали сконструированные последователь
Показано, что LicB/53EGFP и LicB/99EGFP
ности LicB/53EGFP и LicB/99EGFP (рис. 1), а
имеют сходные с нативным EGFP спектры воз
также исходные последовательности LicB M и
буждения и эмиссии белков (рис. 2, д). Эти дан
EGFP, которые перенесены в экспрессионные
ные служат убедительным подтверждением то
вектора на основе системы векторов pQE. Полу
го, что вставленный домен EGFP сохраняет
ченные трансформанты бактерий обозначены
способность к функциональной укладке в при
нами в соответствии с наименованием сконстру
нимающем белке.
ированных последовательностей: LicB/53EGFP,
Экспрессия и функциональные свойства тер4
LicB/99EGFP, LicB M и EGFP.
мостабильной лихеназы с тандемной инсерцией
Анализ бактериальных трансформантов ме
доменов. Далее мы попытались совместить два
тодом чашечного теста показал, что присущую
мономерных домена в двух положениях (53 и
лихеназе активность проявляют как LicB M, так
99 а.о.) термостабильной лихеназы (LicB M) -
и трансформанты LicB/53EGFP и LicB/99EGFP
зеленый (EGFP) и красный (TagRFP) флуорес
(просветленные пятна вокруг колоний) (рис. 2, а).
центные белки, и выяснить: (i) может ли лихена
При этом у трансформантов LicB/ 53EGFP и
за образовывать функциональную укладку при
LicB/99EGFP выявлена и флуоресценция EGFP
вставке одновременно двух доменов; и (ii) сохра
(рис. 2, б). Это с большой вероятностью свиде
нят ли при этом вставленные флуоресцентные
тельствует о том, что в этих белках оба домена
белки способность к функциональной укладке.
принимают функционально активную уклад
Для этого использовали последовательности
ку - «host» (LicB M) и «guest» (EGFP), и эти
LicB/53EGFP/99RFP (рис. 1) и TagRFP, которые
белки, по крайней мере умеренно, стабильны
перенесены в экспрессионные вектора на осно
БИОХИМИЯ том 84 вып. 8 2019
1182
ПАВЛЕНКО и др.
а
б
д
в
г
Рис. 2. Характеристика исходного варианта лихеназы (LicB M) и вариантов лихеназы с инсерцией EGFP (LicB/53EGFP
и LicB/99EGFP). а - Чашечный тест бактериальных трансформантов (об активности лихеназы свидетельствуют просвет
ленные пятна вокруг колоний). б - Чашечный тест бактериальных трансформантов (EFGP флуоресценция). Для рисун
ков а и б - бактериальные трансформанты LicB M (1), EGFP (2), LicB/53EGFP (3) и LicB/99EGFP (4) соответственно.
в - Электрофореграмма сырых белковых лизатов EGFP (1), LicB/53EGFP (2), LicB/99EGFP (3), LicB M (4), сырого бел
кового лизата штамма XL1 Blue, как контроль (5). г - Энзимограмма очищенных белков EGFP (1), LicB/53EGFP (2),
LicB/99EGFP (3), LicB M (4). М - маркер молекулярных масс. д - Спектры возбуждения и эмиссии флуоресценции
EGFP. 1 - Нативный EGFP, 2 - LicB/99EGFP. Спектры LicB/53EGFP идентичны LicB/99EGFP (б).
biokhsm/
ве системы векторов pET, и получены трансфор
53EGFP/99RFP с тандемной вставкой доменов
манты бактерий, обозначенные нами в соответ
(рис. 3, а). Этот результат является свидетель
ствии с наименованием сконструированных
ством того, что белок с двойной вставкой доме
последовательностей: LicB/53EGFP/99RFP и
нов может свернуться в естественных условиях и
TagRFP.
проявляет стабильность. Далее часть СРБ была
Чашечный тест бактериальных трансфор
очищена, и оценены свойства «host» белка (ак
мантов показал, что активность лихеназы про
тивность LicB M) и «guest» белков (флуорес
являют LicB M и LicB/53EGFP/99RFP, и флуо
ценция EGFP и TagRFP). После разделения
ресценция EGFP выявлена как у трансформан
белков в ДСН ПААГ методом энзимограмм бы
та EGFP, так и у LicB/53EGFP/99RFP (данные в
ла охарактеризована способность LicB M и ва
статье не приводятся). Это является убедитель
рианта LicB/53EGFP/99RFP гидролизовать
ным аргументом в пользу того, что
субстрат (просветленные полосы активности)
LicB/53EGFP/99RFP и принимающий белок
(рис. 3, б). На зимограмме наблюдалась един
LicB M, и по крайней мере один из «guest» до
ственная полоса активности с молекулярной
менов (EGFP) функционально активны.
массой, соответствующей теоретически рассчи
Методом ДСН ПААГ электрофореза прове
танной массе LicB/53EGFP/99RFP (~88 кДа)
ден анализ СРБ, результаты которого свидетель
(рис. 3, б). Удельная активность очищенного
ствуют о сходном уровне экспрессии в E. coli как
LicB/53EGFP/99RFP белка оказалась сравни
для LicB M, так и для варианта LicB/
мой с таковой у варианта LicB/99EGFP (с инсер
БИОХИМИЯ том 84 вып. 8 2019
ЛИХЕНАЗА ДОПУСКАЕТ ТАНДЕМНЫЕ ИНСЕРЦИИ БЕЛКОВ
1183
цией EGFP в область 99 а.о.) и составила 155 ±
ность практически на первоначальном уровне, а
15 ед/мг. Флуоресцентная микроскопия бакте
вариант LicB/53EGFP/99RFP проявляет схо
риальных трансформантов подтвердила флуо
жий с LicB M показатель остаточной удельной
ресценцию TagRFP у варианта LicB/53EGFP/
активности.
99RFP (рис. 3, в). Дополнительно установлено,
Рефолдинг лихеназы in vitro. Последствием
что очищенный белок LicB/ 53EGFP/99RFP об
инсерций доменов может быть полная или час
ладает флуоресценцией с максимумами возбуж
тичная потеря способности принимающего бел
дения/эмиссии, характерными как для EGFP
ка к рефолдингу in vitro [18]. Основным методо
(488 и 507 нм), так и для TagRFP (555 и 584 нм)
логическим приемом изучения укладки и ста
(данные в статье не приводятся). Полученные
бильности белков является исследование их раз
результаты указывают на то, что оба вставлен
ворота (unfolding) под воздействием денатури
ных белка (TagRFP и EGFP) способны к функ
рующих агентов и последующего рефолдинга in
циональной укладке при инсерции в лихеназу.
vitro. Мы исследовали способность лихеназы в
Термостабильность очищенных белков оце
исходном варианте белка LicB M и в белках с
нена в процессе инкубации при 70 °С в отсут
инсерцией доменов LicB/53EGFP, LicB/
ствии субстрата, и остаточная удельная актив
99EGFP и LicB/53EGFP/99RFP к повторной
ность лихеназы рассчитана после 4 ч инкубации.
укладке in vitro после денатурации их мочевиной
Отмечено снижение удельной активности лихе
и этанолом. Для оценки способности лихеназы
назы только для варианта LicB/53EGFP. Вари
к рефолдингу in vitro использована фермента
ант LicB/99EGFP сохраняет удельную актив
тивная активность, как единственный показа
а
б
в
Рис. 3. Характеристика исходного варианта лихеназы (LicB M) и варианта лихеназы с инсерциями EGFP и TagRFP
(LicB/53EGFP/99RFP). а - Электрофореграмма сырых белковых лизатов LicB M (1), LicB/53EGFP/99RFP (2). б - Энзимо
грамма очищенных белков LicB M (1), LicB/53EGFP/99RFP (2). М - маркер молекулярных масс. в - Флуоресцентная мик
роскопия бактериальных трансформантов. Бактериальные трансформанты, экспрессирующие нативный TagRFP (1), бакте
риальные трансформанты, экспрессирующие LicB/53EGFP/99RFP (2), бактериальные трансформанты, с пустым pQE век
тором (3). I - бактериальные клетки в проходящем свете; II - визуализация флуоресценции RFP в бактериальных клетках.
biokhsm/
БИОХИМИЯ том 84 вып. 8 2019
1184
ПАВЛЕНКО и др.
инсерцией доменов (LicB/53EGFP, LicB/
99EGFP и LicB/53EGFP/99RFP) составляет
80-85% от начальной активности лихеназы у
этих белков (рис. 4). Рефолдинг вариантов лихе
назы с инсерцией доменов измерен и по восста
новлению флуоресценции EGFP и TagRFP в ко
нечной точке - 160 мин. Оказалось, что белки
LicB/53EGFP, LicB/99EGFP и LicB/53EGFP/
99RFP проявляли флуоресценцию с максимума
ми возбуждения / эмиссии, характерными для
EGFP (488 и 507 нм) (рис. S1 в Приложении), а
вариант LicB/53EGFP/99RFP - для TagRFP
(555 и 584 нм).
Дополнительно оценена и возможность ре
фолдинга in vitro лихеназы после осаждения эта
нолом всех вариантов белков с инсерциями до
менов. Показано, что как для исходного вариан
Рис. 4. Способность лихеназы к рефолдингу in vitro. Кине
та (LicB M), так и для всех вариантов белков с
тика рефолдинга in vitro лихеназы LicB M и белков с ин
инсерцией домена, ферментативная активность
серцией доменов LicB/53EGFP, LicB/99EGFP и LicB/
лихеназы не только сохраняется, но достоверно
53EGFP/99RFP после денатурации мочевиной.
увеличивается (≥10×) (табл. 2). Такое увеличе
С цветным вариантом рис. 4 можно ознакомиться в элект
ние уровня ферментативной активности может
journal/biokhsm/
быть объяснено значительным уменьшением
доли прочих суммарных растворимых белков (от
10 до 20 раз) в образцах белков после осаждения
тель, который позволяет однозначную дискри
этанолом (табл. 2). При этом вставленные доме
минацию функциональной укладки этого белка.
ны (EGFP и TagRFP) у LicB/53EGFP, LicB/
Для определения кинетики рефолдинга in
99EGFP и LicB/53EGFP/99RFP также способ
vitro лихеназы LicB M и белков с инсерцией до
ны к рефолдингу in vitro после денатурации эта
менов LicB/53EGFP, LicB/99EGFP и LicB/
нолом (данные не показаны).
53EGFP/99RFP использовали концентрацию
мочевины и условия инкубации, ранее указан
ные для EGFP [33]. Способность этих белков
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
складываться in vitro первоначально исследова
на за счет определения степени восстановления
В этом исследовании мы определили влияние
активности лихеназы. Так, исходный вариант
инсерции домена(ов) размером более чем 200 а.о.
лихеназы (LicB M) повторно сворачивается in
в две области термостабильной лихеназы на спо
vitro с практически полной обратимостью, вос
собность к укладке в функциональную конфор
становленная активность после проведения
мацию лихеназы и модельных флуоресцентных
цикла рефолдинга in vitro для вариантов белков с
белков, использованных как «guest» домены.
Таблица 2. Активность лихеназы и уровень суммарных растворимых белков до и после осаждения этанолом
Активность лихеназы, ед/мкгСРБ
Уровень СРБ, мкг/мкл
Название белка
До осаждения
После осаждения
До осаждения
После осаждения
этанолом
этанолом
этанолом
этанолом
NC L 53 99
25,46 ± 2,11
267,76 ± 1,58
0,75 ± 0,05
0,03 ± 0,01
EGFP
n/d
n/d
n/d
n/d
RFP
n/d
n/d
n/d
n/d
NC EGFP 53
21,25 ± 1,23
211,48 ± 1,42
0,55 ± 0,05
0,06 ± 0,01
NC EGFP 99
11,35 ± 1,31
115,31 ± 1,85
0,65 ± 0,05
0,04 ± 0,01
NC EGFP 53 RFP 99
12,45 ± 1,51
106,55 ± 1,43
0,35 ± 0,05
0,03 ± 0,01
СРБ - суммарный растворимый белок.
БИОХИМИЯ том 84 вып. 8 2019
ЛИХЕНАЗА ДОПУСКАЕТ ТАНДЕМНЫЕ ИНСЕРЦИИ БЕЛКОВ
1185
Положения вставок в лихеназе, в целом, бы
существует только в нативном состоянии белка
ли выбраны с опорой на общий критерий: они
[37]. Для мониторинга функциональной уклад
расположены на поверхности петель и не при
ки флуоресцентных белков мы применили ме
нимают непосредственное участие в формиро
тоды, которые подтвердили, что флуоресцент
вании каталитического сайта. Согласно экспе
ные белки, как «guest» домены, способны к об
риментальным данным, только правильно вы
разованию функциональной конформации при
бранные петлевые участки могут выдерживать
одиночной и тандемной инсерции в термоста
вставки без драматической потери функцио
бильную лихеназу (рис. 2, б и д; 3, в и рис. S1 в
нальной укладки [39-41]. Например, для PGK
Приложении).
показано, что только две петлевые области из
Таким образом, сохранение ферментативной
четырех апробированных допускают вставку
активности лихеназы и флуоресценции EGFP и
двух разных «guest» доменов в двух разных об
TagRFP у вариантов лихеназы с инсерцией до
ластях [12]. Нами ранее продемонстрировано,
менов позволяет предположить, что: (i) расстоя
что только две петлевые области 53 и 99 а.о. из
ния между структурными элементами могут
трех экспериментально апробированных допус
быть не критичными для формирования функ
кали введение круговых перестановок и толерант
ционально активного домена лихеназы типа
ны к интеграции небольших пептидов [28].
«jelly roll»; (ii) EGFP и TagRFP могут быть встав
Сохранение ферментативной активности у
лены в выбранные области лихеназы и иметь не
вариантов лихеназы со вставками крупных до
зависимые единицы укладки и функционирова
менов (рис. 1-3) демонстрирует, что крупные
ния, в том числе и за счет близости их N и
структурированные вставки могут быть введены
C концевых областей.
в лихеназу в двух экспериментально выбранных
Продемонстрировано, что как лихеназа, так
областях. Отметим, что каталитический домен
и флуоресцентные белки, которые выступают в
лихеназы имеет классическую структуру домена
роли «host» и «guest» белков соответственно, не
типа β «jelly rolle», характерную для семейства
теряют способность к повторной укладке in vitro
гликозид гидролаз 16 (GH16), которая удиви
после воздействия денатурирующих факторов.
тельно устойчива к изменениям аминокислот
Прежде всего мы отметили, что у всех вариантов
ной последовательности и топологии цепи [5,
лихеназы сохраняется способность к ренатура
42], что, вероятно, способствует образованию
ции до активной формы фермента в геле после
стабильной и функциональной укладки лихена
электрофореза в денатурирующих условиях
зы при инсерции крупных доменов в двух экспе
(рис. 2, г и 3, б). Полученные результаты позво
риментально подобранных областях.
ляют судить о том, что повторная укладка этих
Нами отмечено и отсутствие значимых раз
белков возможна после денатурации, но не поз
личий в продукции нативных белков и белков с
воляют сделать заключение о том, насколько
инсерцией домена(ов) (рис. 2, в и 3, а). Извест
она эффективна. Другими словами, на основа
но, что в случае, когда N или C концевые об
нии анализа энзимограмм невозможно оценить
ласти «guest» домена достаточно далеки друг от
относительную скорость процесса рефолдинга
друга, это сокращает число степеней свободы в
лихеназы и пропорцию фракции белка в натив
динамике белков, и может вызвать нежелатель
ном и денатурированном состояниях на пути
ные последствия, такие как неправильный фол
укладки белков с инсерцией доменов при ре
динг белков, низкий уровень продукции белков
фолдинге.
и нарушение биологической активности за счет
Сравнение кривых кинетики рефолдинга у
стерических затруднений [12]. EGFP и TagRFP,
исходного варианта лихеназы и всех вариантов с
использованные нами в качестве «guest» доме
инсерцией доменов демонстрирует относитель
нов, характеризуются относительной близостью
но высокую эффективность повторной укладки
N и C концевых областей [36-39], что, вероят
белков (рис. 4). Известно, что денатурация флуо
но, сопутствовало успеху конструирования ста
ресцентного белка, как правило, нарушает флуо
бильных и функционально активных вариантов
ресценцию [43], и изменения в структуре флуо
лихеназы с инсерцией доменов. Таким образом,
ресцентного белка могут быть оценены по от
нами показана толерантность белковой структу
клонениям спектра его возбуждения [38]. Отсут
ры лихеназы для больших встроек (рис. 2 и 3),
ствие отклонений в спектрах возбуждения сви
которые не вызывают стерических затруднений
детельствует о том, что флуоресцентные белки,
для функциональной укладки лихеназы [16].
как «guest» домены, при их инсерции в лихеназу
Флуоресцентные белки имеют уникальный
также не теряют способность к повторной ук
хромофор, и характеристики флуоресценции
ладке in vitro (рис. S1 в Приложении). Учитывая,
чувствительны к структуре белка, а перенос
что полупериод рефолдинга ЕGFP на порядок
энергии к хромофору в нейтральном состоянии
выше, чем таковые у вариантов лихеназы с ин
БИОХИМИЯ том 84 вып. 8 2019
1186
ПАВЛЕНКО и др.
серцией доменов, можно предположить потен
ности, при длительном хранении белковых пре
циально возможный путь укладки этих белков -
паратов в контейнере закрытой системы, кото
это путь укладки в начале принимающего белка,
рый подвержен изменениям температуры, что,
а затем белка вставки. Это подтверждает гипоте
как следствие, часто приводит к агрегации и де
зу о том, что если сложенное состояние белка
градации белкового препарата [5, 13]. В пятых,
стабильно, то процесс укладки не сильно сдер
слитые белки, сконструированные на основе
живается и не требует строгого упорядочения
термостабильной лихеназы, могут быть нарабо
или координации [18, 24, 42].
таны не только в бактериальной системе, но и
Следует отметить несколько примечатель
на основе иных экспрессионных платформ -
ных свойств термостабильной лихеназы при ис
дрожжи, растения, клетки млекопитающих, как
пользовании ее в качестве принимающего бел
это было продемонстрировано в ряде работ
ка. Во первых, белок сохраняет функциональ
[31, 44].
ную укладку при значительных размерах встраи
Таким образом, наши результаты убедитель
ваемых доменов (рис. 1, 2, 3), хотя, как правило,
но свидетельствуют о том, что термостабильная
при использовании подхода инсерции встраива
лихеназа C. thermocellum может быть включена в
емый домен короче, чем принимающий белок
список «host» белков, способных допускать не
[12]. Во вторых, метод энзимограмм позволяет
только одиночные, но и тандемные вставки до
точно определять молекулярные массы белков,
менов, который ранее включал фосфоглицерат
сконструированных с использованием лихеназы
киназу (PGK) [12] и β лактамазу (BLA) [27], и
(рис. 2, г и 3, б), и характеризуется высокой
имеет преимущества, связанные с ее примеча
чувствительностью [13, 28]. Дополнительно ме
тельными свойствами. Эти свойства делают тер
тод энзимограмм позволяет быстро оценить
мостабильную лихеназу крайне привлекатель
белки на способность к ренатурации in vitro
ной в качестве нового принимающего «host»
(рис. 2, г и 3, б). В третьих, высокий уровень
белка при использовании подхода инсерцион
термостабильности лихеназы, а также сохране
ного слияния доменов для инженерии много
ние лихеназой функциональной укладки после
функциональных белков. Тем не менее в буду
осаждения нецелевых белков этанолом могут
щем требуется проверка сохранения функцио
быть использованы для быстрой и экономичной
нальной укладки лихеназы с другими вставлен
очистки белковых лизатов (табл. 2). Так, ранее
ными белками.
показано, что за счет термообработки при 65 °С
удаляется до 50% загрязняющих белков, остав
ляя слитые с лихеназой белки сохранными [13].
Финансирование. Работа поддержана Рос
В четвертых, термостабильная лихеназа может
сийским фондом фундаментальных исследова
быть использована для «guest» белков, которые
ний (проект № 16 34 00249; AAT).
стабильны только в узком диапазоне рН и тем
Конфликт интересов. Авторы заявляют об от
ператур, поскольку сама функционирует в ши
сутствии конфликта интересов.
роком диапазоне температур и рН. Это свой
Соблюдение этических норм. Настоящая
ство, в целом, может оказаться полезным и для
статья не содержит описания каких либо иссле
увеличения стабильности белков, сконструиро
дований с участием людей и животных в качест
ванных с использованием лихеназы, и, в част
ве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Cota, J., Oliveira, L.C., Damasio, A.R., Citadini, A.P.,
4.
Rizk, M., Antranikian, G., and Elleuche, S. (2012) End to
Hoffmam, Z.B., Alvarez, T.M., Codima, C.A., Leite, V.B.,
end gene fusions and their impact on the production of mul
Pastore, G., de Oliveira Neto, M., Murakami, M.T.,
tifunctional biomass degrading enzymes, Biochem. Biophys.
Ruller, R., and Squina, F.M. (2013) Assembling a
Res. Commun., 428, 1-5, doi: 10.1016/j.bbrc.2012.09.142.
xylanase lichenase chimera through all atom molecular
5.
Goldenkova Pavlova, I.V., Tyurin Acapital A.C., and
dynamics simulations, Biochim. Biophys. Acta, 1834,
Mustafaev, O.N. (2018) The features that distinguish
1492-1500, doi: 10.1016/j.bbapap.2013.02.030.
lichenases from other polysaccharide hydrolyzing enzymes
2.
Furtado, G.P., Ribeiro, L.F., Lourenzoni, M.R., and
and the relevance of lichenases for biotechnological appli
Ward, R.J. (2013) A designed bifunctional laccase/beta
cations, Appl. Microbiol. Biotechnol., 102, 3951-3965,
1,3 1,4 glucanase enzyme shows synergistic sugar release
doi: 10.1007/s00253 018 8904 x.
from milled sugarcane bagasse, Protein Eng. Des. Sel., 26,
6.
Faria, A.R., de Castro Veloso, L., Coura Vital, W., Reis, A.B.,
15-23, doi: 10.1093/protein/gzs057.
Damasceno, L.M., Gazzinelli, R.T., and Andrade, H.M.
3.
Liu, W.C., Lin, Y.S., Jeng, W.Y., Chen, J.H., Wang, A.H.,
(2015) Novel recombinant multiepitope proteins for the
and Shyur, L.F. (2012) Engineering of dual functional
diagnosis of asymptomatic leishmania infantum infected
hybrid glucanases, Protein Eng. Des. Sel., 25, 771-780,
dogs, PLoS Negl. Trop. Dis., 9, e3429, doi: 10.1371/journal.
doi: 10.1093/protein/gzs083.
pntd.0003429.
БИОХИМИЯ том 84 вып. 8 2019
ЛИХЕНАЗА ДОПУСКАЕТ ТАНДЕМНЫЕ ИНСЕРЦИИ БЕЛКОВ
1187
7.
Jacquet, N., Navarre, C., Desmecht, D., and Boutry, M.
23.
Peng, Q., and Li, H. (2009) Direct observation of tug of
(2014) Hydrophobin fusion of an influenza virus hemag
war during the folding of a mutually exclusive protein,
glutinin allows high transient expression in Nicotiana ben
J. Am. Chem. Soc., 131, 13347-13354, doi: 10.1021/
thamiana, easy purification and immune response with
ja903480j.
neutralizing activity, PLoS One, 9, e115944, doi: 10.1371/
24.
Pierre, B., Xiong, T., Hayles, L., Guntaka, V.R., and Kim, J.R.
journal.pone.0115944.
(2011) Stability of a guest protein depends on stability of a
8.
Schmidt, S. R. (2013) Fusion protein technologies for biophar*
host protein in insertional fusion, Biotechnol. Bioeng., 108,
maceuticals: applications and challenges, John Wiley & Sons.
1011-1020, doi: 10.1002/bit.23039.
9.
Schoeffler, A.J., May, A.P., and Berger, J.M.
(2010)
25.
Ribeiro, L.F., Furtado, G.P., Lourenzoni, M.R., Costa
A domain insertion in Escherichia coli GyrB adopts a novel
Filho, A.J., Santos, C.R., Nogueira, S.C., Betini, J.A.,
fold that plays a critical role in gyrase function, Nucleic
Polizeli Mde, L., Murakami, M.T., and Ward, R.J. (2011)
Acids Res., 38, 7830-7844, doi: 10.1093/nar/gkq665.
Engineering bifunctional laccase xylanase chimeras for
10.
Betton, J.M., Jacob, J.P., Hofnung, M., and Broome
improved catalytic performance, J. Biol. Chem., 286,
Smith, J.K. (1997) Creating a bifunctional protein by
43026-43038, doi: 10.1074/jbc.M111.253419.
insertion of beta lactamase into the maltodextrin binding
26.
Elleuche, S. (2015) Bringing functions together with fusion
protein, Nat. Biotechnol., 15, 1276-1279, doi: 10.1038/
enzymes - from nature’s inventions to biotechnological
nbt1197 1276.
applications, Appl. Microbiol. Biotechnol., 99, 1545-1556,
11.
Ay, J., Gotz, F., Borriss, R., and Heinemann, U. (1998)
doi: 10.1007/s00253 014 6315 1.
Structure and function of the Bacillus hybrid enzyme
27.
Edwards, W.R., Williams, A.J., Morris, J.L., Baldwin, A.J.,
GluXyn 1: native like jellyroll fold preserved after insertion
Allemann, R.K., and Jones, D.D. (2010) Regulation of
of autonomous globular domain, Proc. Natl. Acad. Sci.
beta lactamase activity by remote binding of heme: func
USA, 95, 6613-6618, doi: 10.1073/pnas.95.12.6613.
tional coupling of unrelated proteins through domain
12.
Yu, K., Liu, C., Kim, B.G., and Lee, D.Y. (2015) Synthetic
insertion, Biochemistry, 49, 6541-6549, doi: 10.1021/
fusion protein design and applications, Biotechnol. Adv.,
bi100793y.
33, 155-164, doi: 10.1016/j.biotechadv.2014.11.005.
28.
Tyurin, A.A., Sadovskaya, N.S., Nikiforova, K.R.,
13.
Tyurin, A.A., Kabardaeva, K.V., Mustafaev, O.N.,
Mustafaev, O.N., Komakhin, R.A., Fadeev, V.S., and
Pavlenko, O.S., Sadovskaya, N.S., Fadeev, V.S., Zvonova, E.A.,
Goldenkova Pavlova, I.V. (2015) Clostridium thermocel
and Goldenkova Pavlova, I.V. (2018) Expression of soluble
lum thermostable lichenase with circular permutations and
active interferon alphaA in Escherichia coli periplasm by
modifications in the N terminal region retains its activity
fusion with thermostable lichenase using the domain inser
and thermostability, Biochim. Biophys. Acta, 1854, 10-19,
tion approach, Biochemistry (Moscow), 83, 259-269,
doi: 10.1016/j.bbapap.2014.10.012.
doi: 10.1134/S0006297918030069.
29.
Schimming, S., Schwarz, W.H., and Staudenbauer, W.L.
14.
Biondi, R.M., Veron, M., Baehler, P.J., and Reymond, C.D.
(1992) Structure of the Clostridium thermocellum gene
(1998) Random insertion of GFP into the cAMP depen
licB and the encoded β 1,3 1,4 glucanase, Eur. J.
dent protein kinase regulatory subunit from Dictyostelium
Biochem., 204, 13-19, doi: 10.1111/j.1432 1033.1992.
discoideum, Nucleic Acids Res.,
26,
4946-4952,
tb16600.x.
doi: 10.1093/nar/26.21.4946.
30.
Sambrook, J., Fritsch, E.F., and Maniatis, T.
(1989)
15.
Guntas, G., Mitchell, S.F., and Ostermeier, M. (2004)
Molecular cloning: a laboratory manual, Cold Spring
A molecular switch created by in vitro recombination of
Harbor Laboratory Press, New York.
nonhomologous genes, Chem. Biol., 11, 1483-1487,
31.
Piruzian, E.S., Goldenkova, I.V., Musiychuk, K.A.,
doi: 10.1016/j.chembiol.2004.08.020.
Kobets, N.S., Arman, I.P., Bobrysheva, I.V., Chekhuta, I.A.,
16.
Kanwar, M., Wright, R.C., Date, A., Tullman, J., and
and Glazkova, D. (2002) A reporter system for prokaryotic
Ostermeier, M. (2013) Protein switch engineering by
and eukaryotic cells based on the thermostable lichenase
domain insertion, Methods Enzymol., 523, 369-388,
from Clostridium thermocellum, Mol. Genet. Genomics,
doi: 10.1016/B978 0 12 394292 0.00017 5.
266, 778-786, doi: 10.1007/s00438 001 0595 8.
17.
Simm, A.M., Baldwin, A.J., Busse, K., and Jones, D.D.
32.
Bradford, M.M. (1976) A rapid and sensitive method for
(2007) Investigating protein structural plasticity by survey
the quantitation of microgram quantities of protein utiliz
ing the consequence of an amino acid deletion from TEM 1
ing the principle of protein dye binding, Anal. Biochem.,
beta lactamase, FEBS Lett.,
581,
3904-3908,
72,
doi: 10.1016/j.febslet.2007.07.018.
2697(76)90527 3.
18.
Edwards, W.R., Busse, K., Allemann, R.K., and Jones, D.D.
33.
Evdokimov, A.G., Pokross, M.E., Egorov, N.S., Zaraisky, A.G.,
(2008) Linking the functions of unrelated proteins using a
Yampolsky, I.V., Merzlyak, E.M., Shkoporov, A.N.,
novel directed evolution domain insertion method, Nucleic
Sander, I., Lukyanov, K.A., and Chudakov, D.M. (2006)
Acids Res., 36, e78, doi: 10.1093/nar/gkn363.
Structural basis for the fast maturation of Arthropoda green
19.
Bliven, S., and Prlic, A. (2012) Circular permutation in
fluorescent protein, EMBO Rep.,
7,
1006-1012,
proteins, PLoS Comput. Biol., 8, e1002445, doi: 10.1371/
doi: 10.1038/sj.embor.7400787.
journal.pcbi.1002445.
34.
Kanzow, C., Yamashita, N., and Fukushima, M. (2004)
20.
Jones, A.M., Mehta, M.M., Thomas, E.E., Atkinson, J.T.,
Levenberg-Marquardt methods with strong local conver
Segall Shapiro, T.H., Liu, S., and Silberg, J.J. (2016) The
gence properties for solving nonlinear equations with con
structure of a thermophilic kinase shapes fitness upon ran
vex constraints, J. Comput. Appl. Math., 172, 375-397,
dom circular permutation, ACS Synth. Biol., 5, 415-425,
doi: 10.1016/j.cam.2004.02.013.
doi: 10.1021/acssynbio.5b00305.
35.
Arpino, J.A., Rizkallah, P.J., and Jones, D.D.
(2014)
21.
Selvam, R.A., and Sasidharan, R. (2004) DomIns: a web
Structural and dynamic changes associated with beneficial
resource for domain insertions in known protein structures,
engineered single amino acid deletion mutations in
Nucleic Acids Res., 32, D193-D195, doi: 10.1093/
enhanced green fluorescent protein, Acta Crystallogr. D, 70,
nar/gkh047.
2152-2162, doi: 10.1107/S139900471401267X.
22.
Kufner, K., and Lipps, G. (2013) Construction of a
36.
Merzlyak, E.M., Goedhart, J., Shcherbo, D., Bulina, M.E.,
chimeric thermoacidophilic beta endoglucanase, BMC
Shcheglov, A.S., Fradkov, A.F., Gaintzeva, A., Lukyanov, K.A.,
Biochem., 14, 11, doi: 10.1186/1471 2091 14 11.
Lukyanov, S., Gadella, T.W., and Chudakov, D.M. (2007)
БИОХИМИЯ том 84 вып. 8 2019
1188
ПАВЛЕНКО и др.
Bright monomeric red fluorescent protein with an extend
40. Zverlov, V.V., and Schwarz, W.H. (2008) Bacterial cellulose
ed fluorescence lifetime, Nat. Methods, 4, 555-557, doi:
hydrolysis in anaerobic environmental subsystems -
10.1038/nmeth1062.
Clostridium thermocellum and Clostridium stercorarium,
37. Subach, O.M., Malashkevich, V.N., Zencheck, W.D.,
thermophilic plant fiber degraders, Ann. N. Y. Acad. Sci.,
Morozova, K.S., Piatkevich, K.D., Almo, S.C., and
1125, 298-307, doi: 10.1196/annals.1419.008.
Verkhusha, V.V. (2010) Structural characterization of
41. Viguera, A. R., and Serrano, L. (1997) Loop length,
acylimine containing blue and red chromophores in
intramolecular diffusion and protein folding, Nature Struct.
mTagBFP and TagRFP fluorescent proteins, Chem. Biol.,
Biol., 4, 939-946, doi: 10.1038/nsb1197 939.
17, 333-341, doi: 10.1016/j.chembiol.2010.03.005.
42. Bloom, J.D., Labthavikul, S.T., Otey, C.R., and Arnold, F.H.
38. Stepanenko, O.V., Stepanenko, O.V., Kuznetsova, I.M.,
(2006) Protein stability promotes evolvability, Proc. Natl.
Verkhusha, V.V., and Turoverov, K.K. (2013) Beta barrel
Acad. Sci. USA, 103, 5869-5874, doi: 10.1073/pnas.
scaffold of fluorescent proteins: folding, stability and
0510098103.
role in chromophore formation, Int. Rev. Cell. Mol.
43. Ward, W.W. (2006) Biochemical and physical properties of
Biol., 302, 221-278, doi: 10.1016/B978 0 12 407699
green fluorescent protein, Methods Biochem. Anal., 47,
0.00004 2.
39-66, doi: doi. 10.1002/0471739499.ch3.
39. Zoldak, G., Carstensen, L., Scholz, C., and Schmid, F.X.
44. Gerasymenko, I., Sheludko, Y., Frabel, S., Staniek, A., and
(2009) Consequences of domain insertion on the stability
Warzecha, H. (2019) Combinatorial biosynthesis of small
and folding mechanism of a protein, J. Mol. Biol., 386,
molecules in plants: engineering strategies and tools, Methods
1138-1152, doi: 10.1016/j.jmb.2008.12.052.
Enzymol., 617, 413-442, doi: 10.1016/bs.mie.2018.12.005.
THE THERMOSTABLE LICHENASE FROM Clostridium thermocellum
AS A HOST PROTEIN IN THE DOMAIN INSERTION APPROACH
O. S. Pavlenko1*, O. A. Gra1, K. V. Kabarbaeva1, O. N. Mustafaev2, N. S. Sadovskaya1,
V. S. Fadeev1, A. А. Tyurin1, and I. V. Goldenkova4Pavlova1*
1 K.А. Timiryazev Institute of Plant Physiology, Russian Academy of Sciences,
127276 Moscow, Russia; E*mail: Helliga.p@gmail.com, irengold58@gmail.com
2 Baku State University, Department of Biophysics and Molecular Biology,
Baku, AZ 1148, Azerbaijan; E*mail: orkhan@bioset.org
Received March 3, 2019
Revised April 29, 2019
Accepted April 30, 2019
The Clostridium thermocellum lichenase (endo β 1,3;1,4 glucan D glycosyl hydrolase, EC 3.2.1.73 (P29716)) has
been tested as a protein hosting insertion of two model fluorescent proteins (EGFP and TagRFP) in two regions.
Functional folding of all domains is confirmed by preserved lichenase activity and fluorescence of EGFP and TagRFP.
The results demonstrate that (i) these two experimentally tested loop regions of the lichenase permit protein insertions
without damaging its folding in vivo, and (ii) the lichenase permits not only single but also tandem insertions of large
protein domains. High activity, thermostability, and efficient in vitro refolding of lichenase make it attractive as a new
host protein in the domain insertion approach.
Keywords: lichenase, fluorescent proteins, single and tandem protein insertions
БИОХИМИЯ том 84 вып. 8 2019