БИОХИМИЯ, 2019, том 84, вып. 8, с. 1189 - 1204
УДК 576.385.5
ИССЛЕДОВАНИЕ in vitro РОЛИ TGF β1 И МУТАЦИЙ ГЕНА C Kit
В ПРОГРЕССИРОВАНИИ ГЕПАТОЦЕЛЛЮЛЯРНОЙ КАРЦИНОМЫ
У БОЛЬНЫХ, ИНФИЦИРОВАННЫХ ВИРУСОМ ГЕПАТИТА С
© 2019
M.E. El Houseini1, A. Ismail2, A.A. Abdelaal2, A.H. El Habashy3,
Z.F. Abdallah1, M.Z. Mohamed4, M. El Hadidi5,
W.C.S. Cho6, H. Ahmed7, T.A. Al Shafie1,8*
1 Department of Cancer Biology, National Cancer Institute, Cairo University,
Cairo 11796, Egypt; e mail: tamer.alshafie@pua.edu.eg
2 Department of Surgery, Faculty of Medicine, Ain Shams University, Cairo 11566, Egypt
3 Department of Pathology, National Cancer Institute, Cairo University, Cairo 11796, Egypt
4 Department of Medical Laboratory, Medical Center of Egyptian Railways, Cairo 11669, Egypt
5 Center of Informatics Science, Nile University, Giza 12525, Egypt
6 Department of Clinical Oncology, Queen Elizabeth Hospital, Kowloon, Hong Kong, China
7 GlycoMantra, Inc., Baltimore MD 21227, USA
8 Department of Pharmacology and Therapeutics, Faculty of Pharmacy and Drug Manufacturing,
Pharos University in Alexandria, Alexandria 21311, Egypt
Поступила в редакцию 19.03.2019
После доработки 19.04.2019
Принята к публикации 20.04.2019
Трансформирующий фактор роста β (TGF β - transforming growth factor β) является цитокином, который
подавляет рост опухоли в нормальных тканях и при доброкачественных опухолях. В то же время, при зло
качественной трансформации клеток TGF β может проявлять противоположные свойства, и опухолевые
клетки могут использовать это в своих целях. Установлено, что C Kit играет основную роль в активации
стволовых клеток и при регенерации печени после ее повреждения. Однако мало что известно о перекрест
ных связях между TGF β и C Kit и их роли в прогрессировании гепатоцеллюлярной карциномы (HCC -
hepatocellular carcinoma). В настоящей работе было изучено влияние возрастающих доз TGF β1 на стволо
вые клетки печени CD44+CD90+ (LSCs - liver stem cells) и экспрессию гена C Kit в образцах злокачествен
ных и прилегающих к ним тканях печени, полученных от 32 больных с гепатоцеллюлярной карциномой.
Собранные нами данные показали, что количество злокачественных стволовых клеток печени в два раза
превышало число клеток, не подвергающихся злокачественной трансформации. При обработке клеток пе
чени возрастающими количествами TGF β1, происходило последовательное подавление стволовых клеток,
как опухолевых, так и доброкачественных, однако низкие концентрации TGF β1 не подавляли злокачест
венные клетки. Кроме того, в отличие от злокачественных клеток печени, в клетках, не подвергающихся
злокачественной трансформации, в экзонах 9 и 11 не наблюдалось экспрессии гена C Kit. Мутационный
анализ показал, что мутации в экзоне 9, но не в экзоне 11, гена C Kit, в отдельных злокачественных клетках
печени приводят к изменению аминокислотной последовательности и нарушают структуру и функцию бел
ка. Следует особо отметить, что в злокачественных клетках печени выявлена ассоциация мутированного эк
зона 9 и высокого уровня вируса гепатита С (HCV - hepatitis C virus). При этом, экспрессия гена C Kit в этих
клетках не подавлялась при их обработке всеми использованными в работе дозами TGF β1. По нашим све
дениям, это первая работа, свидетельствующая о том, что у больных с гепатоцеллюлярной карциномой, у
которых в крови наблюдается высокая концентрация вируса гепатита С, в клетках печени происходит мута
ция гена C Kit. Результаты нашей работы подчеркивают необходимость изучения уровня TGF β1 и мутаций
гена C Kit у больных с хроническим поражением HCV, чтобы предотвратить развитие гепатоцеллюлярной
карциномы и улучшить качество лечения этих больных.
КЛЮЧЕВЫЕ СЛОВА: гепатоцеллюлярная карцинома, хроническое заболевание печени, TGF β1, стволо
вые клетки печени, мутации C Kit.
DOI: 10.1134/S0320972519080116
Принятые сокращения: HCC - гепатоцеллюлярная карцинома; TGF β1 - трансформирующий фактор роста бета 1;
LSC - стволовая клетки печени; CSC - стволовая опухолевая клетка; SCF - фактор стволовых клеток; C Kit - рецептор
фактора стволовых клеток; CLD - хронических заболеваниях печени; HCV - вирус гепатита С.
* Адресат для корреспонденции.
1189
1190
EL HOUSEINI и др.
Печень является основным эндокринным и
живаются при слабо дифференцированной ге
экзокринным органом, в котором происходит
патоцеллюлярной карциноме. Было показано,
интенсивный метаболизм веществ и который
что CD90 играет важную роль в метастазирова
осуществляет защиту организма. Печень обра
нии, воспалении и фиброзе ткани печени [8, 9].
зована несколькими типами клеток, преимуще
Клетки CD90+CD44+ представляют собой агрес
ственно гепатоцитами и холангиоцитами. Из
сивный фенотип, так как они способны, напри
других типов клеток следует отметить синусои
мер, образовывать метастатические поражения
дальные эндотелиальные клетки, Купферовские
в легких иммунодефицитных мышей [9]. Кроме
клетки и звездчатые клетки [1]. Среди случаев
того, предполагается, что положительность кле
первичного рака печени гепатоцеллюлярная
ток по двум маркерам: по CD44, а также по
карцинома (HCC) является основным гистоло
CD133 или CD90, обеспечивает более коррект
гическим типом рака и поражает ~80% больных
ную идентификацию опухолевых стволовых
с раком печени. Ее прогноз является неблаго
клеток печени [10].
приятным с общим пятилетним уровнем смерт
Показано, что TGF β, как плейотропный
ности >88%. Поэтому HCC является второй в
цитокин, в дополнение к его роли в развитии
мире по распространенности причиной смерти
фиброза печени через активацию звездчатых
от рака [2].
клеток и образование компонентов внеклеточ
Результаты работ, полученные в последние
ного матрикса (ECM - extracellular matrix), вов
годы, продолжают поддерживать гипотезу о том,
лечен в процессы регуляции дифференцировки
что опухолевые стволовые клетки (CSC - cancer
стволовых клеток в различные линии клеток
stem cell) являются корнем развития рака пече
[11, 12]. В недавних работах было показано, что
ни. Более 40% случаев HCC имеют клональное
есть корреляция между аберрантной TGF β
происхождение и образуются из клеток, подоб
опосредованной передачей сигнала и размноже
ных стволовым клеткам или CSC. [3]. Эти клет
нием CSC в процессе развития HCC при хрони
ки печени характеризуются поверхностными
ческих заболеваниях печени (CLD) [9]. Фактор
клеточными маркерами, такими как CD13,
стволовых клеток (SCF - stem cell factor) и его
CD133, CD90 и др. Кроме того, такие клетки ас
рецептор C Kit играют ключевую роль в регене
социируются с возникновением устойчивости к
рации печени после ее повреждения и частич
действию лекарств и неэффективностью прово
ной гепатэктомии [13]. Связывание SCF с этим
димого лечения [4]. Поэтому, необходимо сроч
рецептором приводит к запуску C Kit зависи
но расширить понимание механизмов и разра
мой передачи сигнала и последующей актива
ботать способы направленного воздействия на
ции PI3K/AKT , SRC и JAK/STAT зависимых
CSC при гепатоцеллюлярной карциноме.
сигнальных путей. Предполагается, что прог
Допускаются два пути происхождения ство
рессирование предраковых стволовых клеток
ловых клеток печени (LSC): эндогенный (внут
печени в раковые клетки ассоциировано с по
рипеченочные стволовые клетки) и экзогенный
вышенной экспрессией рецептора C Kit [14].
(внепеченочные стволовые клетки). В послед
Недавно появились сообщения о том, что взаи
нем случае они могут образовываться или из ге
мосвязь между TGF β и C Kit способствуют
матопоэтических стволовых клеток (HSC -
канцерогенезу в клетках HepG2 [15], однако не
hematopoietic stem cells) или из мезенхимных
обходимы дальнейшие исследования этих свя
стволовых клеток (MSC - mesenchymal stem
зей у больных с гепатоцеллюлярной карцино
cells) [5]. CD133 является специфическим мар
мой.
кером гематопоэтических стволовых клеток, в
В настоящей работе нами было предположе
то время как CD90 и CD44 являются специфи
но, что взаимодействия между трансформирую
ческими поверхностными маркерами мезен
щим фактором роста бета 1, стволовыми клетка
химных стволовых клеток. Таким образом, эти
ми печени и геном C Kit при CLD могут привес
поверхностные белки можно вполне уверенно
ти к развитию гепатоцеллюлярной карциномы.
использовать для характеризации LSC [6, 7].
Нами было предположено, что клетки печени,
Для идентификации LSC также используются
изолированные из злокачественной и доброка
различные белки, включая OV6, EpCAM и др. [8].
чественной частей опухолей, удаленных из пе
В частности, CD44 играет роль в ремодел
чени больных с HCC, могут имитировать клетки
линге ткани, адгезии клеток на внеклеточном
печени больных с CLD и гепатоцеллюлярной
матриксе и миграции клеток, т.е. в процессах,
карциномой соответственно. Поэтому, нами
которые вносят вклад в канцерогенез. CD90
была предложена модель ex vivo, чтобы изучить
также вовлечен во взаимодействия клеток и
эффект возрастающих доз TGF β1 на экспрес
внеклеточного матрикса и в межклеточные взаи
сию генов CD44+CD90+ и C Kit в стволовых
модействия, которые преимущественно обнару
клетках печени злокачественного и незлокачест
БИОХИМИЯ том 84 вып. 8 2019
РОЛЬ TGF β1 И МУТАЦИЙ ГЕНА C Kit ПРИ HCC
1191
венного происхождения. Мы считаем, что это
фугировали в течение 20 мин при 800 g, чтобы
исследование прольет свет на этиологическую
получить фракцию стволовых клеток печени,
основу развития гепатоцеллюлярной карцино
локализованную в среднем слое (рис. 1).
мы. Нами впервые показано, что умеренные и
Культивирование клеток печени и их изучение
высокие дозы TGF β1 способны подавлять рост
под микроскопом. Фракции паренхимных и не
популяции LSC и ингибировать экспрессию ге
паренхимных клеток печени, которые содержа
на C Kit с мутированным экзоном 9. Однако
ли LSC, культивировали в среде DMEM, содер
низкие концентрации TGF β1 не подавляли
жащей глутамин и неэссенциальные аминокис
рост стволовых клеток и не подавляли экспрес
лоты и дополненной по 20 нг/мл основного
сию гена C Kit. Кроме того, впервые нами были
фактора роста фибробластов (b FGF - basic
обнаружены различные замены в экзоне 9 гена
fibroblast growth factor) и эпидермального факто
C Kit, ассоциированные с высокими концентра
ра роста (EGF
- epidermal growth factor)
циями вируса гепатита С в злокачественных
(«Biowest Inc.», Франция). Культуры клеток вы
клетках печени, с которыми можно связать ус
держивали при 37 °C в увлажненном инкубаторе
тойчивость этих клеток к действию TGF β1.
в смеси из 95% воздуха и 5% CO2. Морфологию
Интересно, что уровень экспрессии гена C Kit в
клеток изучали на микроскопе с инвертирован
некоторых злокачественных клетках печени,
ной фазой Olympus® CKX41(«Olympus», Япо
полученных от больных, у которых в крови об
ния).
наруживается высокая концентрация HCV и му
Анализ мутаций гена C kit с помощью секве
тации в экзоне 9, оставался без изменений даже
нирования ДНК. Геномную ДНК из клеток пе
при их обработке средними и высокими дозами
чени получали с помощью набора QIAamp DNA
TGF β1.
Purification Kit («Qiagen», Германия). Определе
ние последовательностей ДНК проводили с по
мощью специфических праймеров следующим
МЕТОДЫ ИССЛЕДОВАНИЯ
образом: 1 я пара праймеров С Kit (расположе
на в экзоне 9): 5' TGACATGGTCAATGTTG
Дезагрегация и фракционирование клеток пе
GAA 3' (прямой) и 5' AGCCAGGGCTTTTGTT
чени. Данная работа проводилась в соответствии
TTCT 3' (обратный); 2 я пара праймеров С Kit
c протоколом экспертного совета национально
(расположена в экзоне 11): 5' CAGGTAAC
го онкологического института (Каир, Египет).
CATTTAT 3' (прямой) и 5' GTTATGTGTACC
В ней были обследованы 32 пациента, у которых
CAAAAAGG 3' (обратный), метод секвенирова
на основе лабораторных исследований, радио
ния ДНК на основе обрыва цепи с использова
логических анализов и изучения патологии был
нием красителей с помощью набора для секве
установлен диагноз гепатоцеллюлярная карци
нирования Big Dye Terminator v3.1 Cycle
нома и была проведена частичная гепатэктомия.
Sequencing kit («Applied Biosystems», США) и
Для получения клеток печени были обработаны
ДНК анализатора 3500 Genetic DNA Analyzers
злокачественные и доброкачественные ткани
(«Applied Biosystems», США). Для картирования
печени. Как было описано ранее [16, 17], были
полученных ридов относительно всех известных
разделены клетки основной паренхимы печени
генов из базы данных NCBI и определения ре
и гепатоциты, чтобы получить фракцию непа
ференсного гена C kit человека использовали
ренхимных клеток печени, которая содержит
программу BLAST. NG_007456.1 был выбран в
LSC. Вкратце, каждый образец злокачественной
качестве референсного гена благодаря наивыс
и доброкачественной ткани печени промывали
шему значению счета (score) при сравнении
гипотоническим солевым раствором и равным
нуклеотидных последовательностей [18]. Опре
объемом среды DMEM, дополненной 5% ным
деление однонуклеотидных полиморфизмов
раствором антибиотиков и противогрибковых
(SNPs) в мутированных генах проводили путем
средств. Затем каждый образец ткани измельча
их сравнения с идентифицированной референс
ли и подвергали перевариванию коллагеназой
ной последовательностью гена C kit с использо
III типа, чтобы получить по отдельности клеточ
ванием программ NovoSNP [19] и BioEdit [20].
ную суспензию злокачественных и доброкачест
Определение структуры мутантного белка
венных тканей печени. Каждую клеточную
C Kit. Сравнение структур референсного белка
суспензию пропускали через 100 мкм сито и
и мутированных последовательностей проводи
центрифугировали при 1000 g в течение 10 мин.
ли с помощью метода моделирования по гомо
Каждый ресуспендированный образец центри
логии с использованием программы Molecular
фугировали при 50 g в течение 1 мин 3 раза. За
Operating Environment tool [21]. Модели структу
тем, конечный супернатант наслаивали на смо
ры исходного белка и мутированного белка на
лу Ficoll Paque («Biowest», Франция) и центри
лагали друг на друга и показывали места амино
БИОХИМИЯ том 84 вып. 8 2019
1192
EL HOUSEINI и др.
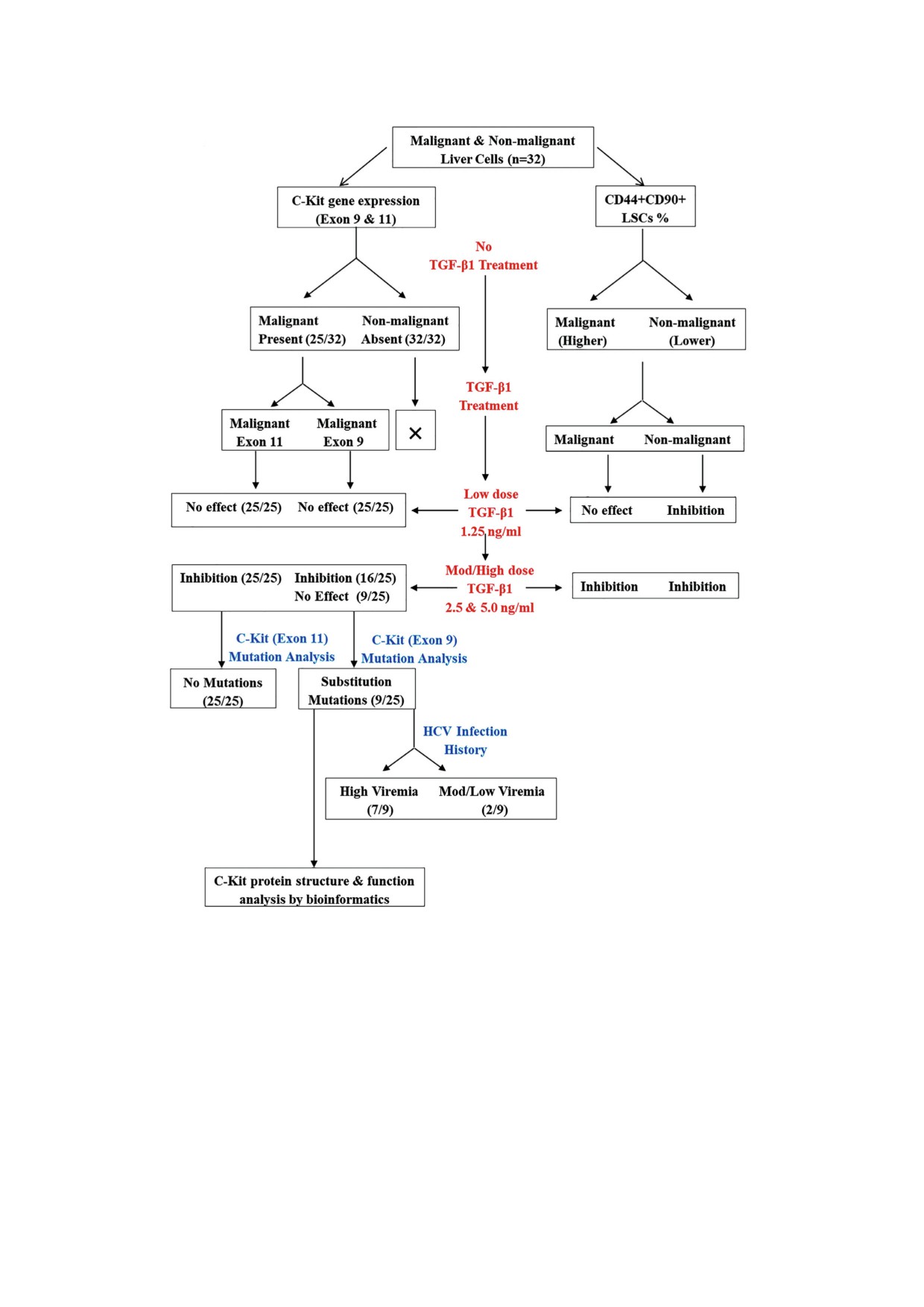
Рис. 1. План проведения исследования.
biokhsm/
кислотных замен. Новый мутантный аминокис
ный пакет SIFT [22]. Предсказания производи
лотный остаток представлен на модели из шари
ли на основе подсчета степени консервативнос
ков и палочек (ball and sticks representation) крас
ти аминокислотных остатков при сравнении
ным цветом, в то время как исходный амино
близкородственных аминокислотных последо
кислотный остаток показан синим цветом.
вательностей с помощью программы PSI
Определение функций мутантного белка C Kit.
BLAST (Position Specific Iterative Basic Local
Для предсказания эффекта аминокислотной заме
Alignment Search Tool). Значение отсечки (cut
ны на функцию белка, использовали программ
off) для вероятности произведения замены, ко
БИОХИМИЯ том 84 вып. 8 2019
РОЛЬ TGF β1 И МУТАЦИЙ ГЕНА C Kit ПРИ HCC
1193
торая была использована для обозначения влия
(обратный), ожидаемый размер продукта 300 п.н.
ния замены на функцию, было равно 0,05. Ана
и 4 я пара праймеров C Kit (расположена в эк
лиз функций белка проводили с использовани
зоне 11): 5' AGCAAATCCATCCCCACACC 3'
ем InterPro веб портала, который классифици
(прямой) и 5' TGGAGCATGCCATTCACGAG 3'
рует белки в виде семейств и предсказывает до
(обратный), ожидаемый размер продукта
мены и значимые сайты [23, 24]. В нем находят
269 п.н.; и β актин: 5' AAAGGGTGTAACG
ся сигнатуры белков, собранные от различных
CAACTAA 3' (прямой) и 5' GGACCTGACT
баз данных, что может помочь в определении
GACTACCTCA 3' (обратный), ожидаемый раз
функций белков и изменений, возникающих в
мер продукта 612 п.н. Продукты ПЦР разделяли
белках в результате мутаций
с помощью гель электрофореза в агарозе и визу
Обработка клеток трансформирующим факто
ализировали с помощью транс иллюминатора в
ром роста бета1. Рекомбинантный TGF β1 чело
ультрафиолете и видимом свете.
века («Koma Biotech Inc.», Южная Корея) в ко
Обнаружение HCV с использованием ПЦР в
личестве 1,25, 2,5 и 5 нг/мл инкубировали с пер
реальном времени: титр вируса гепатита С опре
вичной культурой злокачественных и доброка
деляли у всех больных с помощью набора
чественных клеток печени в течение 24 ч. Затем
Anatolia Genework («Anatolia», Турция) и прибо
клетки печени собирали с использованием
ра DTprime Real Time Thermal cycler («ДНК тех
0,25% ного раствора трипсина в ЭДТА для про
нология», Россия).
ведения цитометрии клеток и ОТ ПЦР.
Статистическая обработка полученных ре
Проточная цитофлюориметрия клеток. Харак
зультатов. Чтобы определить % относительного
теристику LSC проводили с помощью проточ
изменения переменных величин, рассчитывали
ного цитофлуориметра Beckman Coulter Epics
относительную величину процента изменений:
XL Flow Cytometer («Beckman Coulter, Inc.»,
относительная величина процента изменений =
США) с использованием меченых фикоэритри
[(после обработки - до обработки)/до обработ
ном мышиных моноклональных антител против
ки] × 100. Непрерывно измеряемые значения
CD44 человека и меченых FITC мышиных мо
представлены в виде среднего значения ± стан
ноклональных антител против CD90 человека
дартное отклонение (SD). Сравнение данных
(«BD Biosciences», США) согласно инструкциям
между парными группами проводили при ис
производителя (смотри ниже).
пользовании повторяющихся измерений с по
Определение уровня экспрессии гена C Kit
мощью программы ANOVA. После этого, по
(экзоны 9 и 11) в клетках печени с помощью ПЦР
парные сравнения проводили с использованием
с обратной транскрипцией (reverse transcriptase
критерия наименьших достоверных различий
(RT) PCR). Препараты общей РНК получали
Фишера (Fisher’s Least Significant Difference).
путем экстракции РНК из злокачественных и
Значение величины p < 0,05 рассматривалось
доброкачественных клеток с использованием
как статистически достоверное. Все статисти
набора SV total RNA Isolation System
ческие анализы проводились при использова
(«Promega», США) в соответствии с инструкци
нии программного обеспечения SPSS version 15
ями производителя. Определение количества
(«SPSS Inc.», США.)
РНК проводили на спектрофотометре
Nanodrop 2000
(«Thermo Scientific», США).
кДНК первой цепи на основе РНК получали с
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
помощью набора High Capacity cDNA Reverse
Transcription Kit («Qiagen», Германия) в соответ
Данные о больных гепатоцеллюлярной карци
ствии с инструкциями производителя. Для про
номой. Лабораторные анализы показали повы
ведения ПЦР с обратной транскрипцией (RT
шенную функцию печени и содержание альфа
PCR, ОТ ПЦР), в качестве матриц использова
фетопротеина (AFP - α fetoprotein), в то время
ли кДНК первой цепи. Все процедуры ампли
как значения гематологических тестов были
фикации ПЦР выполняли на термоциклере
низкими (табл. 1 и 2). Из 32 больных с HCC,
Eppendorf Mastercycler («Eppendorf», Германия)
20 больных были в различной степени инфици
с использованием ДНК полимеразы Taq
рованы вирусом гепатита С. У 10 из них наблю
(«Promega», США) в буфере и растворе Mg2+,
далась виремия низкой степени, 3 больных бы
поставленных производителем. Отжиг произво
ли диагностированы с умеренной виремией и у
дили в диапазоне температур 58-61 °C. Были ис
7 больных была установлена высокая виремия.
пользованы следующие специфические прайме
Все принявшие участие в данном исследовании
ры: 3 я пара праймеров C Kit (расположена в
больные с HCC до проведения частичной гепа
экзоне 9): 5' CTGGAGCTTTCGGGAAGGTT 3'
тектомии не проходили химиотерапию. Только
(прямой) и 5' GGTGTGGGGATGGATTTGCT 3'
инфицированные вирусом гепатита С больные
10 БИОХИМИЯ том 84 вып. 8 2019
1194
EL HOUSEINI и др.
Таблица 1. Пол, возраст, диагноз, патологические, гематологические и радиологические данные больных с гепатоцеллю
лярной карциномой
Число случаев
n = 32
62,5% (n = 20)
Мужчины
37,5% (n = 12)
Женщины
58,8 ± 7,04
Возраст (лет)
гепатоцеллюлярная карцинома
Диагноз
Степень развития
I степень тяжести
25% (n = 8)
опухоли
II степень тяжести
75% (n = 24)
- цирротическая печень с очаговыми поражениями различных размеров;
US, CT & MRI
- большая масса с общим некрозом и агрессивные диспластические узлы;
- тромб главного PV;
- увеличенная селезенка;
− асциты.
US - УЗИ; CT - компьютерная томография; MRI - магнитно резонансная томография.
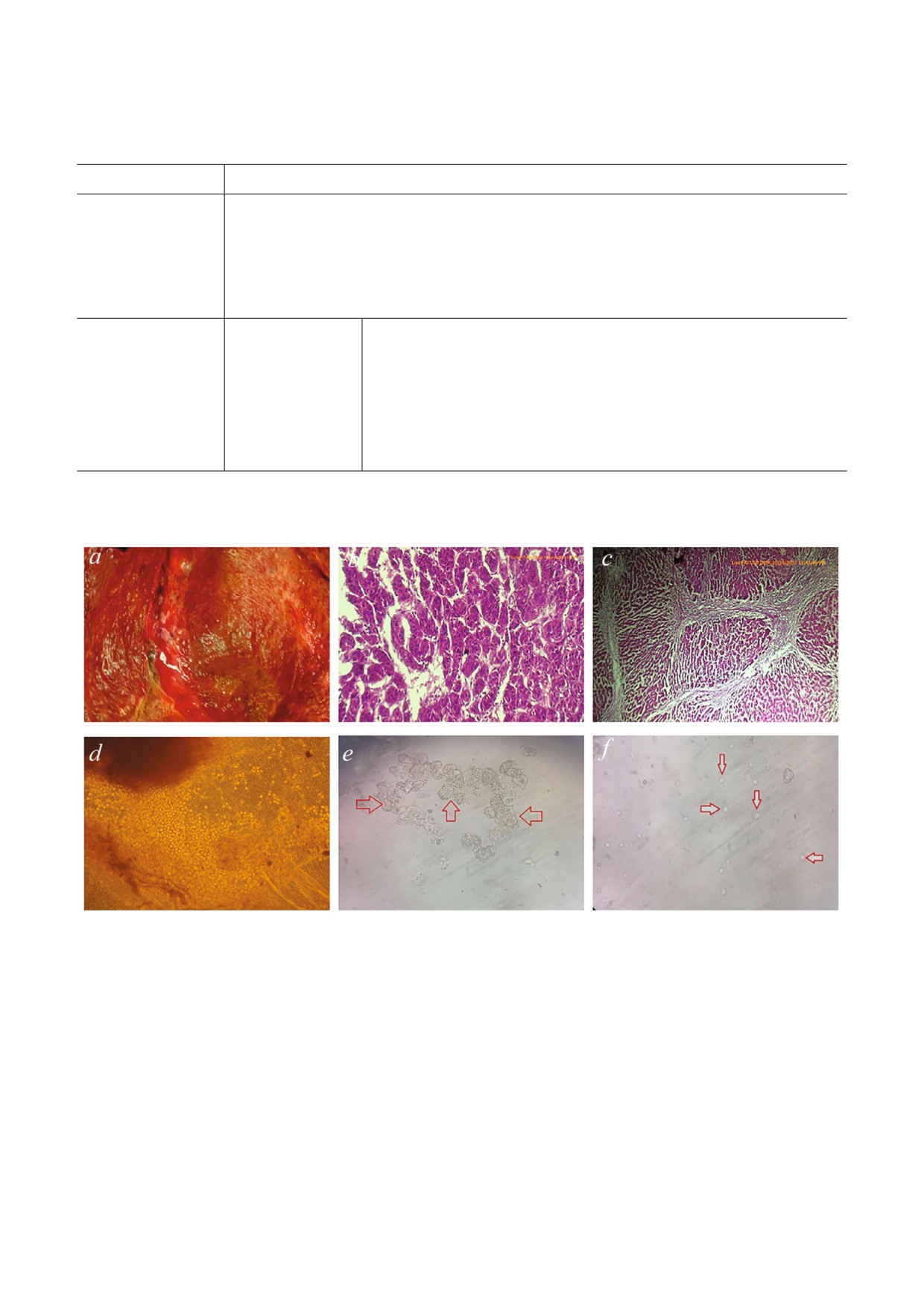
Рис. 2. a - Изображение целой, вырезанной опухоли печени, содержащей злокачественную и доброкачественную ткани.
b - Гистопатологическое изображение злокачественной опухоли печени. Опухоль состоит из трабекул и нескольких гроз
девидных образований злокачественных гепатоцитов, для которых характерно центральное гиперхроматичное ядро и
обильная эозинофильная цитоплазма. Эти трабекулы отделены друг от друга тонкой фиброваскулярной стромой. c - Гис
топатологическое изображение цирротической ткани печени с диффузными нарушениями нормальной структуры пече
ни (утрата нормальных взаимоотношений центра и частей). Есть соединяющая волокнистая перемычка с образованием
паренхиматозных узлов из регенерирующих гепатоцитов. d - Изображение, полученное с помощью инвертированного
микроскопа, демонстрирующее первичный эксплантат печени, из которого высвобождаются различные печеночные и
непеченочные клетки. e - Изображение, полученное с помощью инвертированного микроскопа, демонстрирующее гепа
тоцит (клетка паренхимы печени) в виде многоугольной клетки большого размера в 1 ый день культивирования; увели
чение ×1000. f - Изображение, полученное с помощью инвертированного микроскопа, демонстрирующее непаренхима
тозные клетки печени округлой и овальной формы маленького размера в 1 ый день культивирования; увеличение ×1000.
biokhsm/
БИОХИМИЯ том 84 вып. 8 2019
РОЛЬ TGF β1 И МУТАЦИЙ ГЕНА C Kit ПРИ HCC
1195
получали комбинированную противовирусную
клетки (NPCs - non parencymal cells), в число ко
терапию в виде пегилированного интерферо
торых входят также LSC, - меньше и округлой
на α и рибавирина и в разной степени реагиро
формы (рис. 2, f).
вали на прием этих препаратов, прежде чем у
Более высокое содержание популяции
них была диагностирована HCC (табл. 3).
CD44+90+ LSC среди злокачественных клеток пе
Культура клеток печени. Все вырезанные части
чени по сравнению с доброкачественными клетка
печени (рис. 2, а) содержали кусочки злокачест
ми печени. Принято считать, что прогрессирова
венной и доброкачественной ткани, что было
ние гепатоцеллюлярной карциномы обусловле
также подтверждено гистопатологическими ме
но CSC, которые способны к самообновлению и
тодами (рис. 2, b и с). Клетки печени выделяли из
бесконечному размножению [25]. В настоящей
злокачественной и доброкачественной тканей
работе с помощью проточного цитометра нами
печени и далее их культивировали (рис. 2, d). Ге
была определена доля LSC среди клеток печени,
патоциты имели форму больших многоугольни
полученных в результате дезагрегации доброка
ков (рис. 2, e), в то время как непаренхимные
чественной и злокачественной тканей. Для про
Таблица 2. Функции печени, гематологические тесты, опухолевый маркер, вирусные маркеры больных с гепатоцеллюляр
ной карциномой
Лабораторные исследования
Показатели
Среднее значение ± SD
Контрольные значения
Тесты определения функций
аланинаминотрансфераза (ед./л)
35,25 ± 13,59
До 40 ед./л
печени
аспартатаминотрансфераза ед./л)
54,91 ± 12,99
До 40 ед./л
общий билирубин (мг/дл)
1,91 ± 1,49
До 1,0 мг/дл
прямой билирубин (мг/дл)
0,78 ± 0,55
До 0,2 мг/дл
альбумин (г/дл)
3,25 ± 0,44
3,5 5,5 г/дл
щелочная фосфатаза (ед./л)
116 ± 50,68
До 180 ед./л
ГГТ (ед./л)
75,3 ± 53,6
12 32 ед./л
Гематологические тесты
гемоглобин (г/дл)
10,5 ± 2,0
12,5 14,5 г/дл
лейкоциты (103/мкл)
4,1 ± 2,1
4 11 × 103
тромбоциты (103/мкл)
98 ± 43
150 450 × 103
Опухолевый маркер
АФП (нг/мл)
177 ± 130
10 нг/мл
Таблица 3. Степень виремии при вирусном гепатите и режим приема противовирусных препаратов больных гепатоцеллю
лярной карциномой*
Тип вирусного гепатита
Степень виремии
(Среднее значение ± SD)
Контрольные
Противовирусный
IU/мл
значения
режим*
Non HBV/Non HCV (n = 12)
нет
-
не определены
none
HBV/HCV (n = 0)
нет
-
не определены
none
HBV положительный (n = 0)
нет
-
не определены
none
HCV положительный (n = 20)
низкая § (n = 10)
70,000 ± 110
не определены
респондер
умеренная † (n = 3)
450,000 ± 100,000
респондер
высокая # (n = 7)
1.5,000,000 ± 190,000
нереспондер
* Все обследованные больные с гепатоцеллюлярной карциномой (n = 32) не проходили химиотерапию до проведения час
тичной гепатэктомии. Все HCV положительные больные (n = 20) получали пэгилированный интерферон и рибавирин до
того момента, когда у них диагностировали гепатоцеллюлярную карциному.
§ Низкая степень виремии: 16-200,000 IU/мл.
† Умеренная виремия: 200,000-1,000,000 IU/мл.
# Высокая виремия: >1,000,000 IU/мл.
БИОХИМИЯ том 84 вып. 8 2019
10*
1196
EL HOUSEINI и др.
a
b
d
1
2
c
3
4
e
Рис. 3 (см. подпись на стр. 1197).
БИОХИМИЯ том 84 вып. 8 2019
РОЛЬ TGF β1 И МУТАЦИЙ ГЕНА C Kit ПРИ HCC
1197
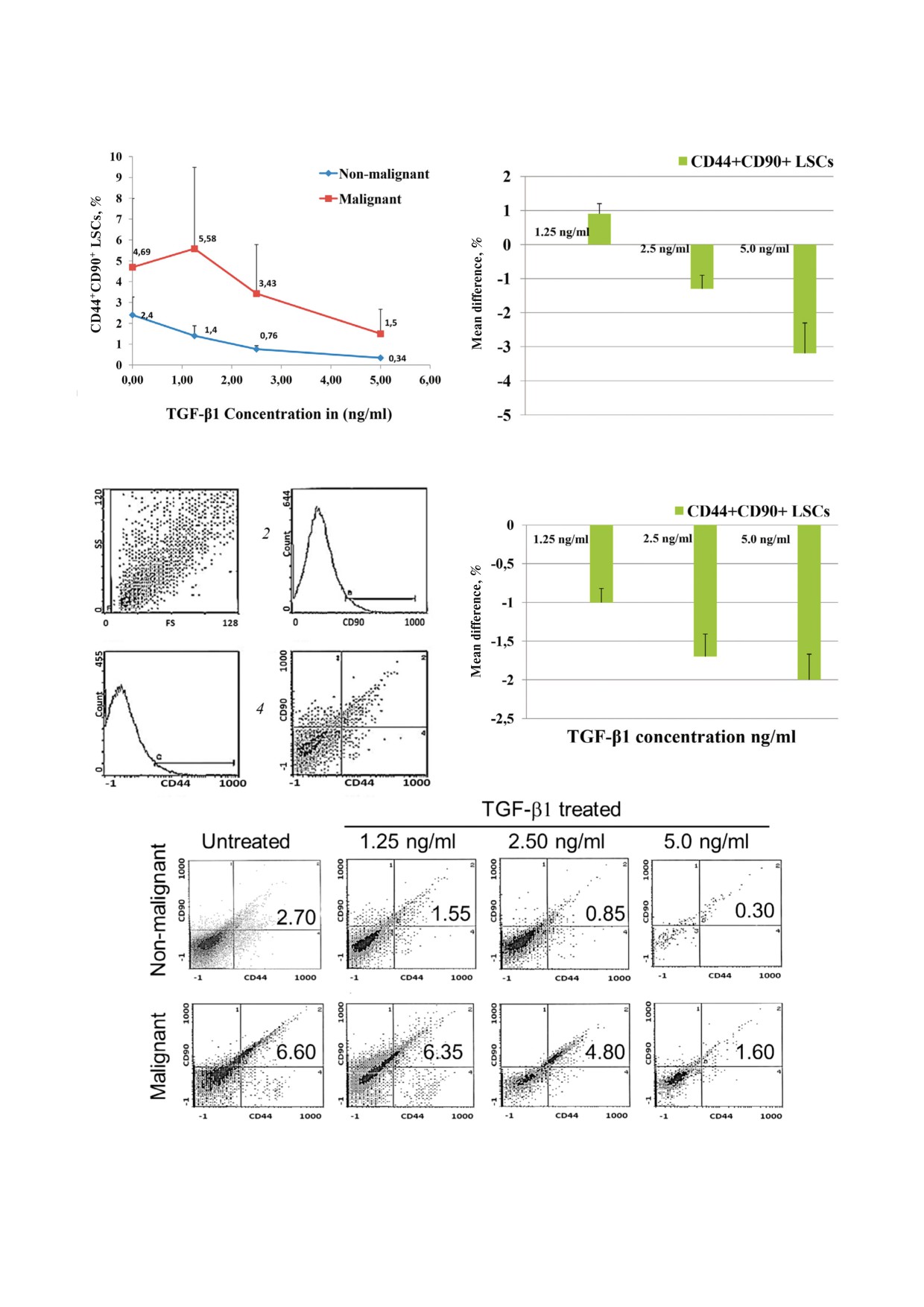
Таблица 4. Сравнение с помощью метода проточной цитометрии экспрессии печеночных стволовых клеток CD44+CD90+
(LSCs) до и после их обработки in vitro повышающимися дозами TGF β1 в клетках печени, происходящих из доброкаче
ственной и злокачественной тканей
Клетки печени, обработанные TGF β1
Источник ткани
Клетки необработанной
печени
1,25 нг/мл
2,50 нг/мл
5,0 нг/мл
Значение p
CD44+CD90+ LSCs из
2,4 ± 0,9
1,4 ± 0,5
0,76 ± 0,16
0,34 ± 0,06
0,001*
доброкачественной ткани (%)
CD44+CD90+ LSCs из
4,69 ± 3,3
5,58 ± 4
3,43 ± 2,4
1,5 ± 1,2
0,019*
злокачественной ткани (%)
* Различия статистически достоверны при p < 0,05.
ведения более точной идентификации стволо
изучить влияние TGF β1 на LSCs, клетки пече
вых клеток печени нами были выбраны клетки
ни получали из злокачественных и доброкачест
печени, экспрессировавшие два общих маркера
венных участков и обрабатывали возрастающи
стволовых клеток, CD44 и CD90, которые обыч
ми количествами TGF β1. Было показано, что
но экспрессируются почти во всех LSC, но в
эти стволовые клетки по разному отвечают на
различной степени в зависимости от здоровой
действие TGF β1. Доля стволовых клеток
или поврежденной печени [26]. Стволовые
CD44+CD90+ из доброкачественной ткани пече
клетки печени пролиферируют в ответ на раз
ни последовательно снижалась в ответ на обра
личные повреждения печени в процессе регене
ботку различными дозами TGF β1 (1,25, 2,5 и
рации, который зависит от типа повреждения
5,0 нг/мл) (рис. 3, a, табл. 4). В то же время про
печени и тяжести прогрессирования болезни
цент CD44+CD90+ среди злокачественных кле
[27]. Нами было показано, что процент
ток печени снижался только при обработке кле
CD44+CD90+ LSC среди злокачественных кле
ток умеренными и высокими дозами TGF β1
ток печени была в 2 раза выше, чем в доброкачест
(2,5 и 5,0 нг/мл) (рис. 3, a, табл. 5). Показаны ре
венных клетках печени (рис. 3, f-c, табл. 4 и 5).
зультаты попарного сравнения CD44+CD90+
Таким образом, повышение количества стволо
LSC, полученных из доброкачественной и зло
вых клеток в злокачественных тканях печени
качественной тканей, обработанных возрастаю
может быть ассоциировано с образованием опу
щими количествами TGF β, и необработанных
холевой ткани.
клеток (рис. 3, b и c, табл. 4 и 5). Полученные
Различные ответы популяций CD44+90+ LSC
данные были проанализированы с помощью
из доброкачественных и злокачественных тканей
программ SPSS v.20 и Prism. Результаты репре
на обработку TGF β1. Выявлена ассоциация
зентативного анализа CD44+CD90+ LSC в доб
между аберрантным супрессорным эффектом
рокачественной и злокачественной тканях с по
TGF β1 и размножением стволовых клеток пе
мощью метода проточной цитометрии показа
чени при возникновении гепатоцеллюлярной
ны на рис. 3, d и e. Нами было установлено, что
карциномы у больных с CLD [12, 13]. Чтобы
в отличие от стволовых клеток, полученных из
Рис. 3 (см. стр. 1196). a - Влияние обработки in vitro возрастающими количествами (1,25, 2,5 и 5,0 нг/мл) трансформиру
ющего фактора роста TGF β1 на стволовые клетки печени CD44+CD90+, выделенные из образцов злокачественной и доб
рокачественной тканей печени. Средние значения различий влияния возрастающих концентраций TGF β на стволовые
клетки CD44+CD90+, происходящие из доброкачественной (b) и злокачественной ткани (c) в сравнении с необработан
ными клетками. d - Этапы характеризации стволовых клеток печени с помощью метода проточной цитометрии с исполь
зованием клеточных маркеров CD90 и CD44: 1 - мертвые клетки и обломки клеток удаляли с помощью метода исключе
ния прямого рассеивания и бокового рассеяния (FSc/SSc - Forward scatter/Side scatter) точечных участков образцов на ос
новании размера и зернистости клеток, 2 - гистограмма CD44 PE, использованная для идентификации печеночных ство
ловых клеток CD44+, 3 - гистограмма CD90 ФИТЦ, использованная для идентификации печеночных стволовых клеток
CD90+ и 4 - контурная диаграмма CD90 ФИТЦ и CD44 ФЭ для идентификации печеночных стволовых клеток
CD44+CD90+. Верхний правый квадрант (область D2) считался содержащим популяцию CD44+CD90+ LSC. e - Репре
зентативные цитограммы, показывающие стволовые клетки CD44+CD90+ из образцов доброкачественной и злокачест
венной ткани в отсутствии или в присутствии увеличивающихся концентраций TGF β1.
biokhsm/
БИОХИМИЯ том 84 вып. 8 2019
1198
EL HOUSEINI и др.
доброкачественной ткани печени, низкие дозы
Таблица 5. Попарные сравнения CD44+CD90+ LSCs из доб
рокачественных и злокачественных тканей, обработанных TGF β (1,25 нг/мл) не вызывали ингибирование
возрастающими концентрациями TGF β1, в сравнении с CD44+CD90+ LSC, полученных из злокачест
необработанными клетками
венной ткани. С другой стороны, умеренные и
высокие дозы (2,5 и 5,0 нг/мл) этого фактора
Клетки печени, обработанные TGF β1
роста вызывали ингибирование CD44+CD90+
1,25 нг/мл
2,50 нг/мл
5,0 нг/мл
LSC, как подвергающихся, так и не подвергаю
щихся злокачественной трансформации. Этот
результат согласуется с тем, что низкий уровень
CD44+CD90+
1,0 (0,5; 1,4)
1,7 (0,9; 2,4)
2,0 (1,2; 2,9)
TGF β1, который часто наблюдается при хрони
LSCs из добро
p = 0,003*
p = 0,002*
p = 0,001*
качественной
ческих заболеваниях печени, не приводит к по
ткани (%)
давлению стволовых клеток, что, в конечном
итоге, повышает риск развития гепатоцеллю
CD44+CD90+
-0,9 (-1,7; -0,06)
1,3 (0,18; 2,3)
3,2 (0,8; 5,6)
лярной карциномы [13].
LSCs из зло
p = 0,04*
p = 0,03*
p = 0,02*
Экспрессия гена C Kit (экзоны 9 и 11) в добро
качественной
ткани (%)
качественных клетках печени, не подвергающихся
злокачественной трансформации, отличается от
† Средние значения различий с 95% CI в круглых скобках.
экспрессии этого гена в злокачественных клетках.
Статистически достоверные значения p при α < 0,05.
Известно, что рецептор C Kit и его лиганд, SCF,
a
b
c
Рис. 4. a - Репрезентативная полуколичественная ПЦР гена C Kit (экзоны 11 и 9 соответственно). Оба экзона гена C Kit
(экзон 11 и 9) экспрессируются в злокачественных клетках печени. В то же время в доброкачественных клетках их экспрес
сия не наблюдается. b - Экспрессия гена C Kit (экзон 11, 9 соответственно) после обработки возрастающими количества
ми TGF β1 (1,25, 2,5 и 5,0 нг/мл. TGF β1 ингибировал экспрессию экзона 11 гена C Kit в, в то время как экспрессия экзо
на 9 не изменялась. c - График, демонстрирующий изменения экспрессии гена C Kit (экзон 11, 9) в ответ на обработку зло
качественных клеток печени трансформирующим фактором роста TGF β1. Экспрессия не подвергшегося мутациям гена
C Kit (экзоны 9 и 11) ингибировалась умеренными и высокими дозами и не ингибировалась низкими дозами, в то время
как экспрессия мутантного по экзону 9 гена C Kit не изменялась при изученных в данной работе дозах TGF β1.
biokhsm/
БИОХИМИЯ том 84 вып. 8 2019
РОЛЬ TGF β1 И МУТАЦИЙ ГЕНА C Kit ПРИ HCC
1199
Таблица 6. Типы мутаций гена C Kit, наблюдаемые в экзо
но экспрессии C Kit (экзон 9 и 11) с использо
нах 11 и 9
ванием обеих пар праймеров (рис. 4).
Обнаружение мутаций в гене C Kit (экзон 9) в
Ген С kit
Мутация
отдельных злокачественных клетках печени. Му
тации в гене C Kit наблюдаются при различных
Экзон 11
нет мутаций
n = 25 из 25
типах рака человека. Считается, что они могут
способствовать росту и прогрессированию опу
Экзон 9
нет мутаций
холей, и они выявляются при самых различных
n = 16 из 25
случаях рака человека. Поэтому есть мнение,
что они могут способствовать росту и прогрес
мутации, приводящие к замене
n = 9 из 25
сии опухолевых клеток [30]. Эти мутации могут
затрагивать самые различные экзоны, но часто
мутации происходят в экзонах 11 и 9 [31]. В свя
зи с тем, что в клетках из доброкачественной
участвуют в пути передачи сигнала, ассоцииро
ткани печени не наблюдалась экспрессия C Kit,
ванного с выживанием, пролиферацией и диф
мы изучили мутации в гене C Kit, в экзонах 9 и
ференцировкой стволовых клеток, играя роль в
11 в клетках печени, выделенных из злокачест
развитии HCC [28]. Однако в то же время было
венных тканей. Во всех злокачественных клет
показано, что гепатоциты, находящиеся в пред
ках не было обнаружено ни одной мутации в эк
раковом состоянии (т.е., гепатоциты, находя
зоне 11 (данные не приведены). С другой сторо
щиеся в хронических узлах) экспрессируют ген
ны, 9 различных мутаций, приводящих к изме
C kit, и эта аномалия может быть обусловлена
нению аминокислотной последовательности
гетерогенностью различных видов гепатоцел
белка (substitution mutations) были обнаружены
люлярной карциномы [29]. Нами была изучена
в экзоне 9, в 9 из 25 популяций клеток печени,
экспрессия гена C Kit (экзоны 9 и 11) в клетках
изолированных из злокачественных тканей
печени, полученных из доброкачественной или
(табл. 6 и 7; рис. 5).
злокачественной тканей. С использованием
Мутированный ген C Kit кодировал белок с
1 ой пары праймеров, локализованной в экзоне
нарушенной структурой и функцией. Принято
9, было показано, что экспрессия гена C Kit (эк
считать, что мутации в гене C Kit способствуют
зон 9) была положительной в 28 случаях из 32
канцерогенезу [32, 33]. В частности, было пока
(87,5%) в доброкачественных клетках и в 25 из
зано, что мутации, приводящие к аминокислот
32 случаев (78.1%) в клетках, изолированных из
ным заменам, вызывают изменения в структуре
злокачественной ткани. Аналогичные результа
белка. Кроме того, при многих типах рака, таких
ты были получены в случае гена C Kit (экзон 11)
как рак кишечника [34] и рак простаты [35, 36],
с использованием 2 ой пары праймеров, лока
мутации в гене C Kit приводят к образованию
лизованной в экзоне 11. С другой стороны, в
обрезанных форм белка C Kit. Поэтому, необхо
доброкачественных клетках не было обнаруже
димо рассмотреть взаимосвязь между мутация
Таблица 7. Влияние аминокислотных замен, ассоциированных с экзоном 9, на структуру и функции белка
Положение нуклеотида
Влияние мутации на структуру и функцию белка
No.
Тип мутации в экзоне 9
Контроль
Исследуемый
Структура белка
Функция белка
образец
1
92
176
G > T
G31 > V
вредная
2
232
36
C > A
молчащая мутация
(нет влияния)
3
167
123
A > C
N56 > T
вредная
4
210
80
A > T
укорочение белка
вредная
5
211
79
G > A
укорочение белка
вредная
6
171
126
T > A
D57 > E
(допустимая)
7
91
201
G > T
G31 > L
вредная
8
92
200
G > T
G31 > L
вредная
9
171
121
T > C
молчащая мутация
(нет эффекта)
БИОХИМИЯ том 84 вып. 8 2019
1200
EL HOUSEINI и др.
a
b
c
d
Рис. 5. Типы мутаций в гене C Kit (экзон 9) и ассоциированные аминокислотные замены, и нарушение структуры белка.
Мутация 1: наблюдалась в положении 92 референсного гена, где гуанин (G) в положении 176 был заменен на тимин (T)
(счет 8) (A1). Эта мутация оказывала отрицательное воздействие на структуру и функцию белка из за замены остатка гли
цина (G) в положении 31 на остаток валина (V) (A3, A4). Мутация 2: наблюдалась в положении 232 референсного гена, в
котором цитозин (C) был заменен на аденин (A) в положении 36 в образце (счет 15) (A2). Это молчащая мутация, которая
не вызывает изменений структуры и функции белка. Мутация 3: наблюдалась в положении 167 референсного гена, в ко
тором аденин (A) был заменен на цитозин (C) в положении 123 в образце (счет 8) (B1). Эта мутация оказывала отрица
тельное воздействие на структуру и функцию белка из за замены остатка аспарагина (N) в положении 56 на остаток тре
онина (T) (B3, B4). Мутация 4: наблюдалась в положении 210 референсного гена, в котором аденин (A) был заменен на
тимин (T) в положении 80 в образце (счет 15) (B2). Эта мутация приводила к укорочению белка. Эта мутация оказывала
отрицательное воздействие на структуру и функцию белка (B4, B5). Мутация 5: наблюдалась в положении 211 референс
ного гена, в котором гуанин (G) был заменен на аденин (A) в положении 79 в образце (счет 8) (B3). Эта мутация приво
дила к укорочению белка. Эта мутация оказывала отрицательное воздействие на структуру и функцию белка (B4, B5). Му
тация 6: наблюдалась в положении 171 референсного гена, в котором тимин (T) был заменен на аденин (A) в положении
126 в образце (счет 9) (C1). Эта мутация оказывала отрицательное воздействие на структуру и функцию белка из за заме
ны остатка аспарагиновой кислоты (D) в положении 57 на остаток глутаминовой кислоты (E) (C2, C3). Мутация 7: на
блюдалась в положении 91 референсного гена, в котором гуанин (G) был заменен на тимин (T) в положении 201 в образ
це (счет 14) (D1). Эта мутация оказывала отрицательное воздействие на структуру и функцию белка из за замены остатка
глицина (G) в положении 31 на остаток лейцина (L) (D4, D5). Мутация 8: наблюдалась в положении 92 референсного ге
на, в котором гуанин (G) был заменен на тимин (T) в положении 200 в образце (счет 8) (D2). Эта мутация оказывала от
рицательное воздействие на структуру и функцию белка из за замены остатка глицина (G) в положении 31 на остаток лей
цина L) (D4, D5). Мутация 9: наблюдалась в положении 171 референсного гена, в котором тимин (T) был заменен на ци
тозин (C) в положении 121 в образце (счет 9) (D3). Это молчащая мутация, которая не вызывает изменений структуры и
функции белка.
biokhsm/
БИОХИМИЯ том 84 вып. 8 2019
РОЛЬ TGF β1 И МУТАЦИЙ ГЕНА C Kit ПРИ HCC
1201
ми и экспрессией этого гена [37]. В нашей рабо
сии C Kit (экзоны 9 и 11) во всех злокачествен
те было показано, что большинство мутаций,
ных клетках (25 из 25) остается без изменений
обнаруженных в гене C Kit, приводят к измене
при обработке клеток низкими дозами TGF β1
нию аминокислотной последовательности и на
(1,25 нг/мл). Интересно, что высокие и умерен
рушению нативной структуры и функций белка.
ные дозы TGF β1 (2,5 нг/мл и 5 нг/мл) были
Впервые нами с помощью программного пакета
способны подавлять экспрессию экзонов 9 и 11
SIFT было показано, что из 9 мутаций в экзоне
гена C Kit во всех злокачественных клетках пе
9, обнаруженных в 9 образцах злокачественной
чени, но не в клетках с мутантным экзоном 9,
ткани печени, 2 мутации приводили к укороче
которые оставались без изменений. Эти резуль
нию аминокислотной последовательности, а
таты, во первых, подтверждают неэффектив
5 мутаций вызывали появление аминокислот
ность низких концентраций TGF β1. Во вто
ных замен. Кроме того, были обнаружены
рых, они предполагают, что мутации в гене C Kit
2 молчащие мутации и 1 мутация с заменой ос
(экзон 9) могут быть вероятной причиной отсут
татка аспарагиновой кислоты на остаток глута
ствия чувствительности этих клеток к действию
миновой кислоты (D57>E). Эти мутации не
даже высоких доз TGF β1 (рис. 4, b и c).
приводили к заметным структурным изменени
Корреляция между мутациями в гене C Kit, тя
ям и вызывали только положительный эффект в
жестью инфицирования вирусом гепатита С и ус
отношении функционирования белка. Все вы
тойчивостью к противовирусной терапии больных
явленные мутации наблюдали на поверхности
с HCC. Показано, что у больных, инфицирован
белковой молекулы, на концах функциональ
ных вирусом гепатита С, в стволовых клетках
ных участков («рук») белка, и их расположение
печени наблюдается повышенная экспрессия
критично для нормального функционирования
гена C Kit. В особенности, это касается боль
белка, вызывая нежелательные последствия
ных, у которых развивается гепатоцеллюлярная
(табл. 7 и рис. 5).
карцинома [40, 41]. В то же время, мало данных
Кроме того, ранее во многих работах было
о том, есть ли у таких больных корреляция меж
показано, что мутации, приводящие к усилению
ду мутациями в гене C Kit и тяжестью инфици
функций (gain of function mutations) гена C Kit,
рования вирусом гепатита С. Из 32 больных
широко распространены в природе и имеют от
HCC, которые приняли участие в этом исследо
ношение к прогрессированию рака [35]. Прове
вании, 20 человек были в различной степени тя
денный нами функциональный анализ с ис
жести инфицированы вирусом гепатита С: 10 с
пользованием программы InterProScan показал,
низкой виремией, 3 с умеренной и 7 больных с
что все изученные последовательности гена
высокой виремией.
C Kit образуют семейство гомологичных генов с
Следует отметить, что в нашей работе было
белковой сигнатурой, сходной с фолдом, харак
показано, что в 7 из 9 случаев HCC с наблюдае
терным для иммуноглобулин подобных белков.
мыми мутациями в гене C Kit больные являются
Удивительно, но при этом мутированные после
положительными по вирусу гепатита С, и у них
довательности не совпадают с рецептором тиро
наблюдается виремия высокой степени. В то же
зиновых киназ и рецептором фактора роста туч
время в образцах, взятых у больных, также ин
ных/стволовых клеток. Необходимые дальней
фицированных вирусом гепатита С, но с низким
шие исследования, чтобы выяснить являются ли
или умеренным содержанием вируса в крови,
такие мутантные белки C Kit с аномальной
мутации в гене C Kit выявлены не были. Также, у
функцией результатом или же драйвером гепа
7 таких больных была выявлена устойчивость к
тоцеллюлярной карциномы на поздних стадиях
действию противовирусной терапии с пэгилиро
заболеваний печени.
ванным интерфероном α и рибавирином. Полу
Различные ответы экспрессии мутантного и
ченные нами результаты согласуются с результа
немутантного экзона 9 C Kit на воздействие TGF
тами Du et al. и Li et al., которые показали, что
β1. Перекрестные связи между TGF β и wnt/
длительное применение некоторых типов про
β катенин , WNK1 зависимыми сигнальными
тивовирусных лекарств против HBV и HCV мо
путями и др., оказывающими влияние на биоло
жет способствовать возникновению мутаций,
гию стволовых клеток, хорошо известны [38,
повышающих устойчивость к действию лекарств
39]. Однако мало что известно о перекрестных
и способствовать канцерогенезу [42, 43]. По на
связях между TGF β и C Kit при развитии гепа
шим сведениям, это первая работа, в которой
тоцеллюлярной карциномы. Поэтому, нами был
показана корреляция между мутациями в гене
изучен эффект экспрессии гена C Kit (экзоны 9
C Kit и высокой виремией у больных с HCC, ин
и 11) в злокачественных клетках печени при ле
фицированных HСV, с одной стороны, и устой
чении возрастающими количествами TGF β1.
чивостью к лечению с помощью пэглированного
Нами было установлено, что уровень экспрес
интерферона α и рибавирина, с другой стороны.
БИОХИМИЯ том 84 вып. 8 2019
1202
EL HOUSEINI и др.
C Kit представляет собой потенциальную
CA203420 National Institute of Health, предостав
терапевтическую мишень у больных с раком
ленным HA.
кишечника и некоторых больных с гепатоцел
Благодарности. Эта работа была поддержана
люлярной карциномой [44, 45], и эти пациен
национальным онкологическим институтом
ты могут отвечать на действие TGF β1. В на
Каирского университета, Египет. Авторы хотели
шей работе было показано, что экспрессия
бы выразить благодарность всем сотрудникам
C Kit в клетках печени, полученных от боль
отдела биологии рака за проявленное внимание
ных с HCC и высокой степенью виремии, ко
и оказанную помощь.
торые также не поддаются лечению пэгилиро
Вклад авторов. Все авторы принимали учас
ванным интерфероном α и рибавирином, ос
тие в подготовке этой статьи. MZ собрал полу
тается без изменений в присутствии умерен
ченные данные. TS, HA, WC прочли текст
ных и высоких доз TGF β1. Поэтому, эти боль
статьи. AI и AA запланировали и выполнили ра
ные могут не реагировать на действие TGF β1,
боту. MH, ZF и MH обобщили полученные ре
что делает возможными развитие у них рака
зультаты и доработали статью. AH внес значи
печени. В целом, чтобы улучшить лечение
тельный вклад в разработку плана исследова
больных с хроническими заболеваниями пече
тельской работы, ее проведение и доработал со
ни и предотвратить развитие HCC, необходи
держание этой статьи. Все авторы прочли и
мо изучать мутации гена C Kit, особенно, у
одобрили конечный вариант этой статьи.
больных гепатоцеллюлярной карциномой, ас
Конфликт интересов. Авторы заявляют об от
социированной с HCV, которые не поддаются
сутствии конфликта интересов.
лечению противовирусными препаратами и в
Соблюдение этических норм. В соответствии
крови у которых наблюдается высокая конце
с этическими нормами, для проведения иссле
нтрация этого вируса.
дований было получено разрешение от эксперт
ного совета (IRB - Institution Review Board) на
По нашим сведениям, настоящая работа яв
ционального онкологического института Ка
ляется первым сообщением о том, что больные,
ирского университета (#IRB00004025). Выпол
инфицированные HCV c высокой степенью ви
нение работы осуществлялось в соответствии с
ремии, которые невосприимчивы к лечению
местными и международными положениями и
противовирусными препаратами, наиболее ве
руководствами для взятия образцов и проведе
роятно имеют мутации в гене C Kit и предрас
ния исследований. Это исследование подтвер
положены к развитию гепатоцеллюлярной кар
дило положения Хельсинкской декларации.
циномы. Наша работа подчеркивает необходи
Информированное согласие. Все больные бы
мость изучения мутаций в гене C Kit у хрониче
ли информированы об исследовательском ха
ских больных, инфицированных HCV c устой
рактере этой работы, и все они предоставили
чивостью к противовирусной терапии для пре
свое информированной согласие в письменном
дотвращения развития гепатоцеллюлярной кар
виде. Все больные, которые приняли участие в
циномы и улучшения качества лечения этих
выполнении данной работы, предоставили ин
больных.
формированное согласие.
Дополнительная информация. Полученные в
ходе проведения настоящей работы данные и
Финансирование. Выполнение данной рабо
результаты их анализа доступны на сайте
ты было частично поддержано грантом
СПИСОК ЛИТЕРАТУРЫ
1.
Friedman, S.L. (2008) Mechanisms of hepatic fibrogene
cancer stem cells: current trends and future challenges,
sis, Gastroenterology, 134, 1655-1669, doi: 10.1053/
World J. Stem Cells, 7, 1185-1201, doi: 10.4252/wjsc.v7.
j.gastro.2008.03.003.
i9.1185.
2.
Jemal, A., Bray, F., Center, M.M., Ferlay, J., Ward, E., and
5.
Xu, L.B., and Liu, C. (2014) Role of liver stem cells in
Forman, D. (2011) Global cancer statistics, CA Cancer J.
hepatocarcinogenesis, World J. Stem Cells, 6, 579-590,
Clin., 61, 69-90, doi: 10.3322/caac.20107.
doi: 10.4252/wjsc.v6.i5.579.
3.
Rao, S., Zaidi, S., Banerjee, J., Jogunoori, W., Sebastian, R.,
6.
Qiu, L., Li, H., Fu, S., Chen, X., and Lu, L. (2018)
Mishra, B., Nguyen, B.N., Wu, R.C., White, J., Deng, C.,
Surface markers of liver cancer stem cells and innovative
Amdur, R., Li, S., and Mishra, L. (2017) Transforming
targeted therapy strategies for HCC, Oncol. Lett., 15,
growth factor β in liver cancer stem cells and regeneration,
2039-2048.7, doi: 10.3892/ol.2017.7568.
Hepatol. Commun., 1, 477-493, doi: 10.1002/hep4.1062.
7.
Yamashita, T., Honda, M., Nakamoto, Y., Baba, M., Nio, K.,
4.
Dragu, D.L., Necula, L.G., Bleotu, C., Diaconu, C.C.,
Hara, Y., Zeng, S.S., Hayashi, T., Kondo, M., Takatori, H.,
and Chivu Economescu, M. (2015). Therapies targeting
Yamashita, T., Mizukoshi, E., Ikeda, H., Zen, Y.,
БИОХИМИЯ том 84 вып. 8 2019
РОЛЬ TGF β1 И МУТАЦИЙ ГЕНА C Kit ПРИ HCC
1203
Takamura, H., Wang, X.W., and Kaneko, S. (2013)
Group, Montreal, QC, Canada, 2010, URL: http://
Discrete nature of EpCAM+ and CD90+ cancer stem cells
in hepatocellular carcinoma, Hepatology, 57, 1484-1497,
22.
Kumar, P., Henikoff, S., and Ng, P.C. (2009) Predicting
doi: 10.1002/hep.26168.
the effects of coding non synonymous variants on protein
8.
Yamashita, T., Ji, J., Budhu, A., Forgues, M., Yang, W.,
function using the SIFT algorithm, Nat. Protoc., 4,
Wang, H.Y., Jia, H., Ye, Q., Qin, L.X., Wauthier, E.,
1073-1081, doi: 10.1038/nprot.2009.86.
Reid, L.M., Minato, H., Honda, M., Kaneko, S.,
23.
Jones, P., Binns, D., Chang, H.Y., Fraser, M., Li, W.,
Tang, Z.Y., and Wang, X.W. (2009) EpCAM positive hepa
McAnulla, C., Maslen, J., Mitchell, A., Nuka, G., Pesseat, S.,
tocellular carcinoma cells are tumor initiating cells with
Quinn, A.F., Sangrador Vegas, A., Scheremetjew, M.,
stem/progenitor cell features, Gastroenterology,
136,
Yong, S.Y., Lopez, R., and Hunter, S. (2014) InterProScan 5:
1012-1024, doi: 10.1053/j.gastro.2008.
genome scale
protein
function
classification,
9.
Ding, W., Mouzaki, M., You, H., Laird, J.C., Mato, J.,
Bioinformatics, 30, 1236-1240, doi: 10.1093/bioinformatics/
Lu, S.C., and Rountree, C.B. (2009) CD133+ liver cancer
btu031.
stem cells from methionine adenosyl transferase 1A defi
24.
Finn, R.D., Attwood, T.K., Babbitt, P.C., Bateman, A.,
cient mice demonstrate resistance to transforming growth
Bork, P., et al. (2017) InterPro in 2017 beyond protein
factor (TGF) beta induced apoptosis, Hepatology, 49,
family and domain annotations, Nucleic Acids Res., 45,
1277-1286, doi: 10.1002/hep.22743.
D190-D199, doi: 10.1093/nar/gkw1107.
10.
Xiang, Y., Yang, T., Pang, B.Y., Zhu, Y., and Liu, Y.N.
25.
Oishi, N., and Wang, X.W. (2011) Novel therapeutic strate
(2016) The progress and prospects of putative biomarkers
gies for targeting liver cancer stem cells, Int. J. Biol. Sci., 7,
for liver cancer stem cells in hepatocellular carcinoma,
517-535.
Stem Cells Int., 2016:7614971, doi: 10.1155/2016/7614971.
26.
Tanaka, M., Itoh, T., Tanimizu, N., and Miyajima, A.
11.
Amin, R., and Mishra, L. (2008) Liver stem cells and
(2011) Liver stem/progenitor cells: their characteristics
TGF β in hepatic carcinogenesis, Gastrointestinal Cancer
and regulatory mechanisms, J. Biochem., 149, 231-239,
Res., 2, S27-S30.
doi: 10.1093/jb/mvr001.
12.
Majumdar, A., Curley, S.A., Wu, X., Brown, P., Hwang, J.P.,
27.
Irfan, A., and Ahmed, I. (2015) Could stem cell therapy be
Shetty, K., Yao, Z.X., He, A.R., Li, S., Katz, L., Farci, P.,
the cure in liver cirrhosis? J. Clin. Exp. Hepatol., 5,
and Mishra, L. (2012) Hepatic stem cells and transforming
142-146, doi: 10.1016/j.jceh.2014.03.042.
growth factor β in hepatocellular carcinoma, Nat. Rev.
28.
Yashpal, N.K., Li, J., Wang, R. (2004) Characterization of
Gastroenterol. Hepatol., 9, 530-538, doi: 10.1038/nrgastro.
C Kit and nestin expression during Islet cell development
2012.114.
in the prenatal and postnatal rat pancreas, Dev. Dyn., 229,
13.
Ren, X., Hu, B., and Colletti, L. (2008) Stem cell factor
813-825.
and its receptor, c kit, are important for hepatocyte prolif
29.
Mansuroglu, T., Baumhoer, D., Dudas, J., Haller, F.,
eration in wild type and tumor necrosis factor receptor 1
Cameron, S., Lorf, T., Fuzesi, L., and Ramadori, G. (2009)
knockout mice after 70% hepatectomy, Surgery, 143,
Expression of stem cell factor receptor C Kit in human nontu
790-802, doi: 10.1016/j.surg.2008.03.021.
moral and tumoral hepatic cells, Eur. J. Gastroenterol. Hepatol.,
14.
Chen, L., Shen, R., Ye, Y., Pu, X.A., Liu, X., Duan, W.,
21, 1206-1211, doi: 10.1097/MEG.0b013e328317f4ef.
Wen, J., Zimmerer, J., Wang, Y., Liu, Y., Lasky, L.C.,
30.
Abbaspour Babaei, M., Kamalidehghan, B., Saleem, M.,
Heerema, N.A., Perrotti, D., Ozato, K., Kuramochi
Huri, H.Z., and Ahmadipour, F. (2016) Receptor tyrosine
Miyagawa, S., Nakano, T., Yates, A.J., Carson, W.E. 3rd,
kinase (C Kit) inhibitors: a potential therapeutic target in
Lin, H., Barsky, S.H., and Gao, J.X. (2007) Precancerous
cancer cells, Drug Des. Devel. Ther., 10, 2443-2459,
stem cells have the potential for both benign and malignant
doi: 10.2147/DDDT.S89114.
differentiation, PLoS One, 2, e293.
31.
Hussain, S.R., Naqvi, H., Ahmed, F., Babu, S.G., Bansal, С.,
15.
Rojas, A., Zhang, P., Wang, Y., Foo, W.C., Munoz, N.M.,
and Mahdi, F. (2012) Identification of the C Kit gene
Xiao, L., Wang, J., Gores, G.J., Hung, M.C., and
mutations in biopsy tissues of mammary gland carcinoma
Blechacz, B. (2016) A positive TGF β/C KIT feedback loop
tumor, J. Egypt. Natl. Cancer Institute, 24, 97-103.
drives tumor progression in advanced primary liver cancer,
32.
McDonell, L.M., Kernohan, K.D., Boycott, K.M., and
Neoplasia, 18, 371-386, doi: 10.1016/j.neo.2016.04.002.
Sawyer, S.L. (2015) Receptor tyrosine kinase mutations in
16.
Wang, M.K., Sun, H.Q., Xiang, Y.C., Jiang, F., Su, Y.P.,
developmental syndromes and cancer: two sides of the
and Zou, Z.M. (2012) Different roles of TGF β in the
same coin, Hum. Mol. Genet., 24, R60-R66, doi: 10.1093/
multi lineage differentiation of stem cells, World J. Stem
hmg/ddv254.
Cells, 4, 28-34.
33.
Lennartsson, J., and Ronnstrand, L. (2006) The stem cell
17.
Anzano, M.A., Roberts, A.B., Smith, J.M., Sporn, M.B.,
factor receptor/C Kit as a drug target in cancer, Curr.
and De Larco, J.E. (1983) Sarcoma growth factor from
Cancer Drug Targets, 6, 561-571.
conditioned medium of virally transformed cells is com
34.
Mostafa, M., El Serafi, Abeer A., Bahnassy, M.D., Nasr, M.,
posed of both type alpha and type beta transforming growth
Ali, Salem M., Eid, Mahmoud M., Kamel, Nayera A.,
factors, Proc. Natl. Acad. Sci. USA, 80, 6264-6268.
Abdel Hamid, Abdel Rahman, N., and Zekri, Ph.D.
18.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., and
(2010) the prognostic value of C Kit, K ras codon 12, and
Lipman, D.J. (1990) Basic local alignment search tool,
p53 codon 72 mutations in Egyptian patients with stage II
J. Mol. Biol., 215, 403-410.
colorectal cancer, Cancer, 116, 4954-4964.
19.
Weckx, S., Del Favero, J., Rademakers, R., Claes, L.,
35.
Cardoso, H.J., Figueira, M.I., and Socorro, S. (2017) The
Cruts, M., De Jonghe, P., Van Broeckhoven, C., and De
stem cell factor (SCF)/c kit signaling in testis and prostate
Rijk, P. (2005) novoSNP, a novel computational tool for
cancer, J. Cell Commun. Signal., 11, 297-307.
sequence variation discovery, Genome Res., 15, 436-442.
36.
Paronetto, M.P., Farini, D., Sammarco, I., Maturo, G.,
20.
Hall, T.A. (1999) BioEdit: a user friendly biological
Vespasiani, G., Geremia, R., Rossi, P., and Sette, C.
sequence alignment editor and analysis program for
(2004) Expression of a truncated form of the C Kit tyro
Windows 95/98/NT, in Nucleic Acids Symposium Series,
sine kinase receptor and activation of Src kinase in human
Oxford University Press, No 41, pp. 95-98.
prostatic cancer, Am. J. Pathol., 164, 1243-1251.
21.
Chemical Computing Group: the molecular operating
37.
Bai, C.G., Liu, X.H., Xie, Q., Feng, F., and Ma, D.L.
environment (MOE), v. 2010. 10. Chemical Computing
(2005) A novel gain of function mutant in C Kit gene and
БИОХИМИЯ том 84 вып. 8 2019
1204
EL HOUSEINI и др.
its tumorigenesis in nude mice, World J. Gastroenterol., 11,
42. Du, Y., Su, T., Ding, Y., and Cao, G. (2012) Effects
7104-7108.
of antiviral therapy on the recurrence of hepatocellular
38. Sun, Q., Guo, S., Wang, C.C., Sun, X., Wang, D., Xu, N.,
carcinoma after curative resection or liver transplanta
Jin, S.F., and Li, K.Z. (2015) Cross talk between TGF β/
tion, Hepat. Mon., 12, e6031, doi: 10.5812/hepatmon.
Smad pathway and Wnt/β catenin pathway in pathological
6031.
scar formation, In.t J. Clin. Exp. Pathol., 8, 7631-7639.
43. Li, L., Liu, W., Chen, Y.H., Fan, C.L., Dong, P.L.,
39. Lee, B.H., Chen, W., Stippec, S., and Cobb, M.H. (2007)
Wei, F.L., Li, B., Chen, D.X., and Ding, H.G. (2013)
Biological cross talk between WNK1 and the transforming
Antiviral drug resistance increases hepatocellular carcino
growth factor Smad signaling pathway, J. Biol. Chem., 282,
ma: a prospective decompensated cirrhosis cohort study,
17985-17996.
World J. Gastroenterol., 19, 8373-81.
40. Tekin Koruk, S., Ozardali, I., Dincoglu, D., Guldur, M.,
44. Shah, Y.M., and van den Brink, G.R. (2015) c Kit as a
and Calisir, C. (2012) Can the presence of C Kit positive
novel potential therapeutic target in colorectal cancer,
hepatic progenitor cells in chronic hepatitis C have a role in
Gastroenterology, 149, 534-537, doi: 10.1053/j.gastro.
the follow up of the disease? Erciyes Med. J., 34, 44-49.
2015.07.027.
41. Kwon, Y.C., Bose S.K., Steele, R., Meyer, K.,
45. Chung, C.Y., Yeh, K.T., Hsu, N.C., Chang, J.H., Lin, J.T.,
Di Bisceglie, A.M., Ray, R.B., and Ray, R.
(2015)
Horng, H.C., and Chang, C.S. (2005) Expression of C Kit
Promotion of cancer stem like cell properties in hepatitis C
protooncogene in human hepatocellular carcinoma,
virus infected hepatocytes, J. Virol., 89, 11549-11556.
Cancer Lett., 217, 231-236.
ROLE OF TGF β1 AND C Kit MUTATIONS
IN HEPATOCELLULAR CARCINOMA PROGRESSION
IN PATIENTS WITH HEPATITIS C VIRUS: in vitro STUDY
M. E. El Houseini1, A. Ismail2, A. A. Abdelaal2, A. H. El Habashy3,
Z. F. Abdallah1, M. Z. Mohamed4, M. El Hadidi5,
W. C. S. Cho6, H. Ahmed7, and T. A. Al Shafie1,8*
1 Department of Cancer Biology, National Cancer Institute, Cairo University,
Cairo 11796, Egypt; e mail: tamer.alshafie@pua.edu.eg
2 Department of Surgery, Faculty of Medicine, Ain Shams University, Cairo 11566, Egypt
3 Department of Pathology, National Cancer Institute, Cairo University, Cairo 11796, Egypt
4 Department of Medical Laboratory, Medical Center of Egyptian Railways, Cairo 11669, Egypt
5 Center of Informatics Science, Nile University, Giza 12525, Egypt
6 Department of Clinical Oncology, Queen Elizabeth Hospital, Kowloon, Hong Kong, China
7 GlycoMantra, Inc., Baltimore MD 21227, USA
8 Department of Pharmacology and Therapeutics, Faculty of Pharmacy and Drug Manufacturing,
Pharos University in Alexandria, Alexandria 21311, Egypt
Received March 19, 2019
Revised April 19, 2019
Accepted April 20, 2019
Transforming growth factor β (TGF β) is a tumor suppressing cytokine in normal and non malignant conditions.
Yet, in malignancy, TGF β can exert opposite effects that cancer cells may exploit for their benefits. C Kit plays a
prominent role in stem cell activation and liver regeneration after injury. However, little is known about cross talk
between TGF β and C Kit and its role in hepatocellular carcinoma (HCC) progression. Here, we studied the effect
of increasing doses of TGF β1 on CD44+CD90+ liver stem cells (LSCs) and C Kit gene expression using malignant
and adjacent non malignant liver tissues excised from 32 HCC patients. Results showed that malignant LSCs per
centage was 2.0 fold higher compared to their non malignant counterparts. When treated with increasing doses of
TGF β1, malignant and non malignant LSCs were progressively suppressed, but low TGF β1 dose failed to suppress
malignant LSCs. Moreover, C Kit exons 9 and 11 were not expressed in non malignant LSCs, in contrast to their
malignant counterparts. Mutation analysis of C Kit detected mutations in exon 9, but not in exon 11, in some malig
nant liver cells resulting in changes of amino acid sequence and in the deregulated protein structure and function.
Interestingly, in malignant liver cells, mutated exon 9 was shown to be associated with high level hepatitis C virus
(HCV) viremia, and its expression was not suppressed by TGF β1 treatment at all doses. To our knowledge, this is the
first report that the C Kit gene is mutated in patients with HCC associated with high level HCV viremia. Our study
underscores the need to investigate TGF β1 level and C Kit mutations in patients with chronic HCV for HCC pre
vention and better therapeutic management.
Keywords: HCC, chronic liver disease, TGF β1, liver stem cells, C Kit mutation
БИОХИМИЯ том 84 вып. 8 2019