БИОХИМИЯ, 2020, том 85, вып. 1, с. 139 - 152
УДК 611.018.51
ЭТАНОЛЬНЫЙ ЭКСТРАКТ ОБОЛОЧКИ СЕМЯН ТАМАРИНДА
ЭФФЕКТИВНО ЗАЩИЩАЕТ ЭРИТРОЦИТЫ ОТ ЭРИПТОЗА,
ВЫЗВАННОГО ОКИСЛИТЕЛЬНЫМ СТРЕССОМ
© 2020
Дж. Кенгайя1, С.K.M. Нандиш1, Ч. Рамачандрайя1, Чандрама1, А. Шивайя1,
Дж.Дж. Вишалакши2, М. Поль2, С.М. Сантош3, Р.Л. Шанкар4, Д. Саннанингайя1*
1 Department of Studies and Research in Biochemistry and Centre for Bioscience and Innovation,
Tumkur University, 572103 Tumkur, India
2 Department of Studies in Biochemistry, University of Mysore, Manasagangothry, 570006 Mysore, India
3 Department of Medical Biochemistry and Microbiology (IMBM) Uppsala Biomedical Centre, 75237 Uppsala, Sweden
4 Department of Sericulture, Yuvaraja’s College, University of Mysore, 570006 Mysore, India;
E mail: sdevbiochem@gmail.com
Поступила в редакцию 01.04.2019
После доработки 04.10.2019
Принята к публикации 17.10.2019
Апоптоз или запрограммированная гибель эритроцитов - это ключевой момент в выявлении анемии при
различных патологических состояниях, включая сахарный диабет, хронические заболевания почек (ХБП),
рак, сепсис и др. Окислительный стресс, важный триггер в процессе ускоренной гибели эритроцитов при
эриптозе, является основным механизмом, лежащим в основе возникновения анемических состояний при
вышеуказанных патологиях. Поэтому выявление антиоксидантных и антиэриптотических средств для лече&
ния связанных со стрессом заболеваний сегодня является все более востребованным. В настоящей работе
продемонстрированы антиоксидантные и антиэриптотические свойства этанольного экстракта оболочки се&
мян тамаринда (TSCEE), действующие против индуцированного AAPH (2,2′ азобис (2&амидинопропан гид&
рохлорид)) окислительного стресса и эриптоза. Присутствие вероятных вторичных метаболитов в экстракте
TSCEE исследовали с помощью колоночной хроматографии RP&HPLC. Активные функциональные группы,
присутствующие в экстракте, были изучены с помощью инфракрасной спектроскопии с преобразованием
Фурье (FT&IR). Циклические вольтамперометрические исследования подтвердили антиоксидантный потен&
циал препарата. Кроме того, способность экстракта TSCEE защищать эритроциты была подтверждена при
оценке маркеров эриптоза, таких как активные формы кислорода (АФК), внутриклеточный уровень кальция
и экстернализация фосфатидилсерина. Также было показано, что экстракт TSCEE понижал уровень пере&
кисного окисления липидов, содержание карбонильных групп в белках, восстанавливал внутриклеточный
уровень глутатиона (GSH), способствовал активации антиоксидантных ферментов и ферментов, участвую&
щих в восстановлении пула GSH. В заключение следует отметить, что TSCEE имеет обширное терапевтичес&
кое применение, поэтому он может быть перспективным терапевтическим агентом для лечения эриптоза,
индуцированного окислительным стрессом, и следующей за этим анемии при различных патологиях.
КЛЮЧЕВЫЕ СЛОВА: этанольный экстракт оболочки семени тамаринда (TSCEE - Tamarind Seed Coat
Ethanol Extract), активные формы кислорода (АФК), эриптоз, (GSH/GSSG), G6PDH, антиоксидант, AAPH.
DOI: 10.31857/S032097252001011X
Эритроциты - это наиболее уникальные
дый день у здоровых взрослых людей появляют&
клеточные компоненты, присутствующие в кро&
ся ~200 миллионов эритроцитов, и их продол&
вотоке, они являются основным средством
жительность жизни составляет 120 дней. Цито&
снабжения кислородом и удаления углекислого
кин под названием эритропоэтин, образующий&
газа из тканей через газообмен в легких. Каж&
ся в почках, заставляет гемопоэтические стволо&
вые клетки из костного мозга проходить ряд ста&
Принятые сокращения: АФК - активные формы
дий дифференцировки, чтобы сформировать
кислорода; AAPH - 2,2′ азобис (2&амидинопропан гидро&
зрелые эритроциты, которые характеризуются
хлорид); GSH - восстановленный глутатион; GSH - окис& отсутствием митохондрий и ядра [1, 2]. Будучи
ленный глутатион; GRdx - глутатионредуктаза; GPx - глу& клетками, несущими кислород, эритроциты бо&
татионпероксидаза; GST
- глутатион&S&трансфераза;
лее подвержены окислительному повреждению.
MetHb - метгемоглобин; СОД - супероксиддисмутаза;
CAT - каталаза; ФС - фосфатидилсерин; ХБП - хрони&
Некоторые аспекты, такие как непрерывное
ческие болезни печени.
воздействие кислорода, высокое содержание
* Адресат для корреспонденции.
полиненасыщенных жирных кислот в их липид&
139
140
КЕНГАЙЯ и др.
ном бислое и автоокисление гемоглобина, при&
Несмотря на огромное терапевтическое приме&
водящее к деградации гема, делают циркулиру&
нение оболочки семян тамаринда его способ&
ющие эритроциты восприимчивыми к окисли&
ность защищать эритроциты не была исследова&
тельному повреждению [3]. В нормальных усло&
на. Таким образом, в настоящей работе показа&
виях эритроциты хорошо снабжены антиокси&
на эффективность защиты эритроцитов с по&
дантными молекулами и детоксицирующими
мощью TSCEE против AAPH&индуцированного
ферментами для борьбы с окислительным
окислительного стресса.
стрессом. Однако сообщалось, что при некото&
рых патологических состояниях, включая диа&
бет, рак и хронические заболевания почек
МЕТОДЫ ИССЛЕДОВАНИЯ
(ХБП), наблюдали повреждение эритроцитов в
результате окислительного стресса. В настоящее
Химические соединения и реактивы. 2,2′ азо&
время показано, что терапевтические препара&
бис (2&амидинопропан гидрохлорид) (AAPH);
ты, используемые для лечения этих патологи&
2′,7′&дихлородигидрофлуоресцеин (DCFDA);
ческих состояний, управляют окислительным
Fura&2AM, меченый аннексин V&FITC; HEPES
стрессом, вызывая повреждение эритроцитов.
[4&(2&гидроксиэтил)&1 пиперазин этансульфо&
Избыточный окислительный стресс вызывает
новая кислота]; тиобарбитуровая кислота
как лизис эритроцитов [4], так их старение и ги&
(TBA);
1,1′,3,3′&тетраметоксипропан (TMP);
бель от эриптоза. Эриптоз - это суицидальная
восстановленный глутатион (GSH); окислен&
гибель эритроцитов, которая характеризуется
ный глутатион (GSSG); O&фталевый альдегид;
сморщиванием клеток и скремблированием
1&хлор&2,4&динитробензол (CDNB) и кальцеин
рассматриваемыми как признаки апоптоза [5].
были приобретены у фирмы «Sigma», США. Все
Таким образом, устойчивый окислительный
остальные реактивы были аналитической чис&
стресс может изменить параметры крови за счет
тоты.
снижения количества эритроцитов и, наконец,
Растительный материал и получение экстракта
может привести к тяжелым анемическим состо&
оболочки семени тамаринда. Семена тамаринда
яниям. Анемия вызывает гипоксию в различных
были приобретены на местном рынке в городе
органах, а симптомы анемии включают рас&
Тумкур. Семена были тщательно помыты и высу&
стройство желудка, нарушение когнитивных
шены при комнатной температуре в течение 24 ч,
функций, депрессию, сердечно&сосудистые ослож&
после этого высушенные семена прогревали при
нения, головокружение и одышку при физичес&
температуре 50 °C в течение 15 мин, семенные
кой нагрузке. Кроме того, у пациентов, страда&
оболочки механически отделяли, гомогенизиро&
ющих раком, ХБП и сердечно&сосудистыми ос&
вали, перетирая пестиком в ступке, добавляя
ложнениями, окислительный стресс, вызван&
95% этанол. Полученный этанольный экстракт
ный анемией, является наиболее распростра&
фильтровали через фильтровальную бумагу Ват&
ненной проблемой [6]. Антиоксидантная тера&
ман № 1 и выпаривали спирт. Полученный
певтическая стратегия помогает в более эффек&
фильтрат полностью растворяли в воде и исполь&
тивном управлении вышеуказанной проблемой,
зовали для проведения дальнейшего анализа.
чтобы преодолеть эти вторичные побочные эф&
Анализ растительных компонентов экстракта
фекты при различных патологиях. Лекарствен&
TSCEE с помощью прибора HPLCNPDA. Расти&
ное растение тамаринд - источник богатый ан&
тельные компоненты экстракта TSCEE опреде&
тиоксидантными молекулами - является пред&
ляли при помощи HPLC&PDA («Waters Corpora&
почтительным выбором для терапии. Тамаринд
tion», США). Примерно 20 мкл образца TSCEE
индийский или индийский финик (Tamarindus
наносили на колонку C18 (150 мм × 4,60 мм, раз&
indica L) относится к семейству Fabaceae и под&
мер частиц 5 мкм). Колонку предварительно
семейству Caesalpiniodeae. Хотя родиной этого
уравновешивали 10%&м ацетонитрилом в воде
тропического дерева является Африка, оно ши&
для HPLC. Образец элюировали с использова&
роко представлено в мире. T. Indica является
нием линейного градиента воды для HPLC и
стручком и, подобно фруктам, имеет съедобную
ацетонитрила (растворители А и В, 9 : 1 v/v).
часть и семена. Съедобная часть используется в
Скорость потока составляла 1 мл/мин, и обра&
ресторанах и традиционной медицине по всему
зец сканировали при различных длинах волн в
миру. В то же время семенная оболочка
диапазоне 200-800 нм.
(TSCEE), содержит множество фитохимических
Инфракрасная спектрофотометрия с преобраN
веществ, главным образом полифенолов, алка&
зованием Фурье (FTNIR - Fourier Transform
лоидов, флавоноидов, тритерпенов и полисаха&
Infrared Spectrophotometer) и определение антиокN
ридов, используемых при лечении различных
сидантной активности экстракта TSCEE с поN
патофизиологических состояний человека [7].
мощью циклической вольтамперометрии. Измере&
БИОХИМИЯ том 85 вып. 1 2020
ЭРИПТОЗ И ЗАЩИТНАЯ ЭФФЕКТИВНОСТЬ СЕМЯН ТАМАРИНДА
141
ния спектров FT&IR и циклическую вольтампе&
Определение уровня активных форм кислорода
рометрию проводили по методам Kareru et al. [8]
(АФК), внутриклеточного Сa2+ и экстернализации
и Arulpriya et al. [9]. ИК&спектры записывали,
фосфатидилсерина (ФС). Ключевые параметры
используя Frontier FT&IR/FIR system («Perkin
эриптоза, такие как повышение содержания
Элмера», США) в виде гранул КБР. Цикличес&
АФК, повышение уровня внутриклеточного Сa2+
кую вольтамперометрию (CV - сyclic voltamme&
и экстернализация фосфатидилсерина, оценива&
try) проводили на приборе CHI608E model
ли с помощью метода Driver et al. [12] с некото&
potential state в стандартных трехэлектродных
рыми незначительными изменениями. Для ин&
ячейках из стекловидного углерода с платино&
дукции эриптоза использовали AAPH (1 мМ), а
вой проволокой и электрода Ag/AgCl при ком&
для исследования ингибирования эритроциты
натной температуре.
(2%&ный гематокрит) предварительно инкубиро&
Забор крови человека. Забор крови у здоро&
вали с AAPH (1 мМ) и затем обрабатывали раз&
вых взрослых волонтеров проводили с исполь&
личными концентрациями экстракта TSCEE
зованием антикоагулянта, кислой цитратной
(0-200 мкг/мл). Конечный объем доводили до
декстрозы (ACD - Acid Citrate Dextrose; 85 мМ
200 мкл раствором Рингера, содержащим 125 мМ
цитрата натрия, 71 мМ лимонной кислоты и
NaCl, 5 мМ KCl, 3 мМ MgSO4, 5 мМ HEPES,
111 мМ декстрозы) в соотношении
6/1
=
2 мМ CaCl2, 10 мМ глюкозы (pH 7,4) и инкуби&
= кровь/ACD (v/v), после чего центрифугирова&
ровали в течение 30 мин при 37 °C. Далее в реак&
ли при 200 g в течение 10 мин при 37 °C. В ре&
ционную смесь добавляли флуоресцентные кра&
зультате была получена обогащенная тромбоци&
сители, такие как DCF&DA (10 мкМ), Fura&2AM
тами плазма (супернатант) и осадок, содержа&
(2 мкМ) и аннексин V&FITC (5 мкМ), и инкуби&
щий эритроциты. Осажденные эритроциты про&
ровали при 37 °C в течение 30 мин, чтобы опре&
мывали 2-3 раза 10 мМ фосатным солевым бу&
делить уровни АФК, внутриклеточного Ca2+ и
фером (PBS - Phosphate Buffer Saline, pH 7,4).
степень экстернализации ФС соответственно.
Далее получали 2%&ный гематокрит на основе
Полученную флуоресценцию измеряли с по&
осажденных эритроцитов с добавлением 1 мМ
мощью считывателя для микропланшет Varioskan
глюкозы в 10 мМ PBS, и использовали его для
multimode plate reader («Thermo Scientifics»,
дальнейшего анализа.
США) при возбуждении реакционной смеси при
Определение уровня индуцированного AAPH
488 нм с последующей эмиссией при 530 нм для
метгемоглобина (MetHb) и активности MetHbNреN
определения АФК. Для определения внутрикле&
дуктазы. Образование MetHb в эритроцитах оп&
точной концентрации ионов Ca2+ возбуждение
ределяли с помощью метода Benesch et al. [10] с
клеток проводили при 340 и 380 нм при постоян&
небольшими изменениями. Вкратце, по 1 мл
ной длине волны эмиссии 500 нм. Экстернализа&
эритроцитов (2%&ный гематокрит) помещали в
цию ФС проводили при длине волны возбужде&
пробирки, независимо обрабатывали различны&
ния 496 нм и длине волны эмиссии 530 нм.
ми концентрациями AAPH (0-500 мкМ) и ин&
Определение жизнеспособности эритроцитов.
кубировали в течение 24 ч при 37 °C. После за&
Анализ жизнеспособности эритроцитов прово&
вершения инкубации клетки лизировали в PBS,
дили по методу NaveenKumar et al. [13]. Сначала
содержащем 1%&ный Triton X&100 (1/1, v/v) и из&
эритроциты (2%&ный гематокрит) независимо
меряли поглощение при 630 нм. В качестве по&
обрабатывали или AAPH (1 мМ) или TSCEE
ложительного контроля эритроциты обрабаты&
(200 мкг/мл). При определении ингибирующей
вали 20 мМ NaNO2 (это означало, что образует&
роли клетки обрабатывали AAPH (1 мМ) в при&
ся 100% MetHb). Для определения защитного
сутствии возрастающих концентраций TSCEE
эффекта к предварительно обработанным
(0-200 мкг/мл) и инкубировали в течение 8 ч
AAPH (500 мкМ) эритроцитам добавляли
при 37 °C, затем их окрашивали кальцеином AM
различные концентрации экстракта TSCEE
(10 мкМ) и инкубировали в течение 30 мин при
(0-200 мкг/мл).
37 °C. Клетки дополнительно промывали и из&
Активность MethHb&редуктазы определяли
меряли флуоресценцию при длине волны воз&
по методу Sharma et al. [11]. Вкратце, к исход&
буждения 488 нм, а эмиссию регистрировали
ным AAPH (500 мкМ) и предварительно обра&
при 585 нм.
ботанным AAPH (500 мкМ) гемолизатам до&
Определение перекисного окисления липидов
бавляли различные концентрации TSCEE
(LPO - lipid peroxidation) и содержания карбоN
(0-200 мкг/мл) вместе с НАДН и 2,6 дихлорфе&
нильных групп в белках (PCC - protein carbonyl
нолиндофенолом в качестве акцептора электро&
content). Перекисное окисление липидов оцени&
нов, после чего измеряли поглощение образцов
вали с помощью метода Ohakawa et al. [14]. В су&
при 600 нм. Активность MethHb&редуктазы рас&
хие пробирки для определения размещали
считывали по скорости снижения поглощения.
~1,0-2,0 мг белка лизата эритроцитов, обрабо&
БИОХИМИЯ том 85 вып. 1 2020
142
КЕНГАЙЯ и др.
танных агонистом AAPH (1 мМ) и TSCEE
al. [7]. Вкратце, 2%&ный гематокрит в PBS обра&
(0-200 мкг/мл). Затем в каждую пробирку до&
батывали как контроль, а 1 мМ AAPH - как аго&
бавляли ~1,5 мл 20%&ной уксусной кислоты
нист соответственно. Для исследования инги&
(v/v) (pH 3,5), 0,2 мл 8%&го (w/v) SDS и 1,5 мл
бирующего влияния эритроциты, обработанные
0,8%&ной (w/v) тиобарбитуровой кислоты. Реак&
AAPH, инкубировали с TSCEE (0-200 мкг) в те&
ционную смесь нагревали при 45-60 °C в тече&
чение 2 ч. После завершения инкубации эритро&
ние 45 мин, затем центрифугировали при 450 g в
циты лизировали в дистиллированной воде и
течение 10 мин. Образующиеся аддукты экстра&
использовали для определения активности фер&
гировали 1&бутанолом (3 мл). Образующийся
ментов, таких как СОД и КАТ. Для определения
продукт TBARS (Thiobarbituric Acid&Reactive
активности СОД 0,1 мл лизата эритроцитов
Substance) регистрировали при 532 нм и рассчи&
(0,05 мг белка) добавляли в реакционную смесь
тывали его количество, используя в качестве
(1 мл), состоящую из фосфатного буфера
стандарта TMP (1,1,3,3&тетраметоксипропан).
(16 мМ, pH 7,8), смеси TEMED/ЭДТА (8 мМ/
Содержание карбонильных групп в белках из&
0,08 мМ) и 0,15% (w/v) кверцетина. Снижение
меряли с помощью динитрофенилгидразина
величины поглощения регистрировали в тече&
(DNPH) в соответствии с методом, описанным
ние 1 мин при 406 нм. За единицу принимали
Levine et al. [15]. К 1 мг/мл белка лизата эритро&
количество белка, которое ингибирует процесс
цитов, обработанных агонистом AAPH (1 мМ) и
автоокисления кверцетина на 50%. Для опреде&
TSCEE (0-200 мкг/мл), добавляли 5 мкл 10 мМ
ления активности КАТ в лизатах эритроцитов
DNPH в 2 N HCl и инкубировали в течение 1 ч
проводили измерение скорости гидролиза H2O2
при периодическом встряхивании при комнат&
при 240 нм. Образцы эритроцитов (0,05 мг бел&
ной температуре. Соответствующие контроль&
ка) добавляли в реакционную смесь (1мл), со&
ные образцы приготавливали путем добавления
держащую натрий&фосфатный буфер (100 мМ,
только 2N HCl. После завершения инкубации ре&
pH 7,4) и H2O2 (8,8 мМ). Регистрировали сниже&
акционную смесь осаждали с помощью 20%&ной
ние поглощения при 240 нм в течение 3 мин и
ТХУ и центрифугировали при 1200 g в течение
активность фермента выражали в виде количе&
15 мин. Полученный осадок дважды промывали
ства мМ разложившейся H2O2/мин/мг белка.
ацетоном, центрифугировали при 2500 g в тече&
Определение активности глутатионредукта
ние 15 мин и растворяли в 1 мл 20 мМ Tris&HCl
зы (GRdx), глутатионпероксидазы (GPx) и глута
буфере (pH 7,4), содержащем 0,14 M NaCl и 2%
тион S трансферазы (GST). Активность GRdx в
SDS. Поглощение супернатанта регистрировали
лизатах эритроцитов определяли по методу,
при 360 нм. Разницу в значении поглощения оп&
описанному Sundaram et al. [7]. Кратко, к 1 мл
ределяли и выражали в мкм карбонильных
100 мМ фосфатного буфера (pH 7,0), содержа&
групп/мг белка, используя коэффициент моляр&
щего 2 мМ ЭДТА, 20 мМ GSSG и 2 мМ НАДФН
ной экстинкции 22 мМ-1 см-1.
(в 0,1%&ном NaHCO3) добавляли 0,1 мл лизата
Определение содержания GSH и GSSG. Что&
эритроцитов (0,05 мг белка). Снижение величи&
бы выяснить редокс&статус эритроцитов, опре&
ны поглощения при 340 нм регистрировали в те&
деляли уровни GSH и GSSG, используя метод
чение 3 мин.
Sundaram et al. [16]. Вкратце, чтобы оценить
Активность GPx измеряли с помощью мето&
уровень GSH, к эритроцитам (2%&ный гематок&
да с сопряженными ферментами, как это было
рит), предварительно обработанным агонистом
описано Tung et al. [17]. В реакционную смесь
AAPH
(1 мМ) и ингибиторами TSCEE
(1 мл), содержащую 770 мкл 50 мМ натрий&фос&
(0-200 мкг/мл), добавляли o&фталевый альдегид
фатного буфера (pH 7,0), 100 мкл 10 мМ GSH,
(1 мг/мл) при комнатной температуре в течение
100 мкл 2 мМ НАДФН, 10 мкл 1,125 M азида нат&
15 мин. Далее чтобы определить соотношение
рия и 10 мкл 100 U/мл GPx, добавляли ~10 мкл
GSH/GSSG, до добавления o&фталевого альде&
лизата эритроцитов. После уравновешивания
гида образцы обрабатывали N&этилмалеимидом
смеси в течение 10 мин реакцию начинали, до&
(10 мкМ). Итоговую флуоресценцию измеряли
бавляя в реакционную смесь 50 мкл 5 мМ H2O2,
при 430 нм после возбуждения образцов при
и измеряли окисление НАДФН в течение 3 мин
365 нм. Концентрацию GSH и GSSG определя&
при 340 нм. Активность GPx рассчитывали, ис&
ли с использованием стандартных кривых.
пользуя коэффициент молярной экстинкции
Определение активности ферментов, участвуN
для НАДФН при 340 нм, равный 6,22 M-1см-1.
ющих в поддержании редоксNгомеостаза. Опреде
Активность GST измеряли в соответствии с
ление активности супероксиддисмутазы (СОД) и
методом Guthenberg et al. [18]. В реакционную
каталазы (КAT). Определение активности СОД
смесь (1 мл), содержащую 900 мкл 100 мМ PBS
и КAT в лизатах эритроцитов проводили с ис&
(pH 6,5,), 1 мМ ЭДТА, 50 мкл 20 мМ GSH и
пользованием метода, описанного Sundaram et
50 мкл CDNB (20 мМ раствор в этаноле), аго&
БИОХИМИЯ том 85 вып. 1 2020
ЭРИПТОЗ И ЗАЩИТНАЯ ЭФФЕКТИВНОСТЬ СЕМЯН ТАМАРИНДА
143
нист (AAPH, 1 мМ) и ингибитор (TSCEE,
ляются алкалоиды, флавоноиды и полифенолы
0-200 мкг/мл), добавляли предварительно обра&
(рис. 1, a). Далее спектры FTIR экстракта
ботанные эритроциты (2%&ный гематокрит).
TSCEE показали присутствие полосы поглоще&
Смесь инкубировали течение 3 ч, после чего регис&
ния при 3446 и 3429 см-1, предполагающие рас&
трировали увеличение поглощения при 340 нм в
тяжение связи O-H. Кроме того, в экстракте
течение 3 мин. За едицину активности глутатион&
TSCEE была обнаружена арильная полоса при
редуктазы, глутатиопероксидазы и глутатион&S&
590 см-1, а алкильные полосы появлялись при
трансферазы принимали количество фермента,
507 и 521 см-1. Это укрепляет предположение о
необходимое для получения 1,0 мкмоль соответ&
присутствии гидроксильных, арильных и ал&
ствующего продукта или потребление 1,0 мкмоль
кильных групп в экстракте TSCEE, что свиде&
соответствующего субстрата за 1 мин.
тельствует о наличии активных биохимических
Определение активности глюкозо 6 фосфат
соединений (рис. 1, b).
дегидрогеназы (G6PDH). Активность G6PDH оп&
Антиоксидантная активность экстракта
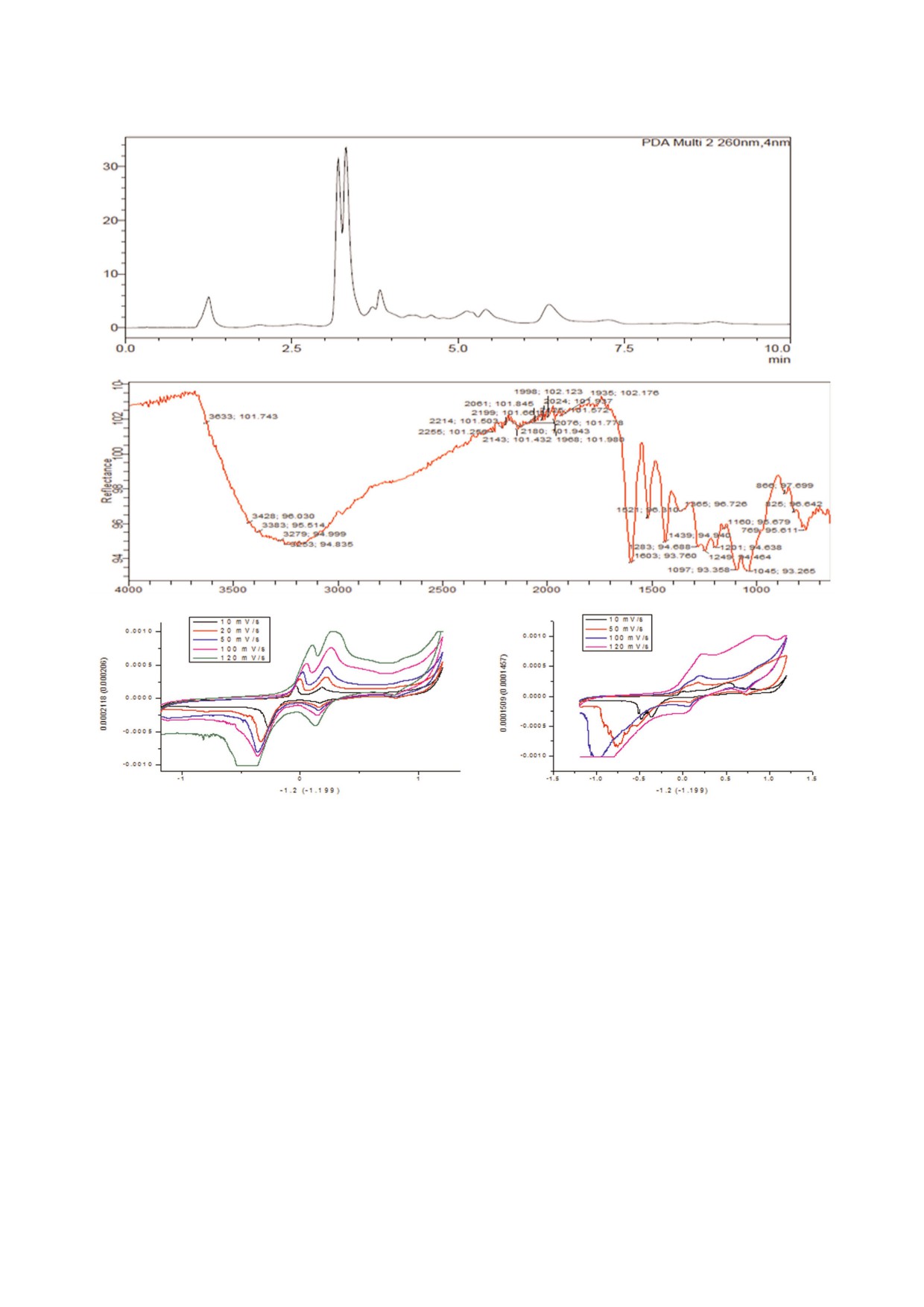
ределяли по увеличению величины поглощения
TSCEE. Для определения антиоксидантного по&
при 340 нм в течение 3 мин благодаря НАДФ&за&
тенциала экстракта TSCEE скорость окисли&
висимому превращению глюкозо&6&фосфата [16].
тельно&восстановительного сканирования из&
Вкратце, промытые эритроциты (2%&ный гема&
меряли, используя различные системы раство&
токрит в PBS) обрабатывали агонистом AAPH
рителей K4[Fe (CN)6] + K3[Fe(CN)6] + KCL
(1 мМ). При исследовании ингибирующего эф&
(5мМ) в диапазоне скорости сканирования
фекта обработанные AAPH эритроциты инкуби&
10-120 мВс-1 и KOH (0,3 мМ). Установлено, что
ровали с TSCEE (0-200 мкг) в течение 3 ч при
диапазон потенциалов окислительно&восстано&
37 °C. После завершения инкубации эритроциты
вительных пиков для электрода сравнения со&
суспендировали и лизировали в 1 мМ PBS буфе&
ставляет от
-1,2 до
+1,2 В относительно
ре. Лизаты использовали для определения ак&
Ag/AgCl (0,1 мМ KCL). Был приготовлен элект&
тивности G6PDH в реакционной смеси (1 мл)
род на основе углеродной пасты (CPE) путем из&
50 мМ Tris&HCl (pH 7,5), содержащей 3,8 мМ
мельчения порошка графита (70%) и силиконо&
НАДФ, 3,3 мМ глюкозо&6&фосфата и 6,3 мМ
вого масла (30%) при использовании небольшо&
MgCl2. За единицу активности глюкозо&6&фос&
го пестика и ступки при ручном перемешивании
фатдегирогеназы принимали количество фер&
в течение ~30 мин, что позволило получить го&
мента, способное производить
1,0 мкмоль
могенный чистый CPE. Он действует как рабо&
НАДФН в течение 1 мин.
чий электрод, платиновая проволока выступает
Определение концентрации белка. Концент&
в качестве противоположного электрода и
рацию белка в образцах определяли по методу
Аg/AgCl - в качестве электрода сравнения. Для
Lowry et al. [19], используя в качестве стандарта
этого экстракта нами были получены два пика
БСА.
окисления (Epc = -0,030 и 0,175) и два пика вос&
Статистическая обработка данных. Статисти&
становления (Epa = -0,270 и 0,164). Анодный
ческий анализ был выполнен при использова&
пиковый ток сдвигает Epa в анодном направле&
нии программы Graph Pad Prism. Все результаты
нии в присутствии TSCEE. Это указывало на то,
представляют среднее значение ± стандартная
что фитоконституенты TSCEE адсорбировались
ошибка отклонения трех независимых экспери&
на поверхности электрода. Наши наблюдения
ментов.
позволяют предположить, что процесс был диф&
фузионно контролируемым. Кроме того, было
обнаружено, что Epa смещается в анодном нап&
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
равлении с увеличением скорости развертки,
что указывает на квазиобратимый характер
Экстракт TSCEE содержит большое количестN
электродной реакции. Было также отмечено,
во алкалоидов, флавоноидов и полифенолов. Для
что анодный пиковый потенциал (Epa) и полу&
определения фитохимических составляющих в
пиковый потенциал (Epc) были смещены в сто&
экстракте TSCEE, образцы подвергали высоко&
рону более положительных значений, это свиде&
эффективной жидкостной хроматографии на
тельствует о том, что экстракт тушит свободные
колонке С18 с фотодиодным регистратором
радикалы кислорода. Рис. 1, c представляет со&
(RP&HPLC&Photo Diode Array detector). Мони&
бой электрохимические циклические вольтам&
торинг хроматографического разделения прово&
перограммы кривых экстракта TSCEE.
дили в диапазоне длин волн 200-800 нм в тече&
Экстракт TSCEE понижает индуцированное
ние 1 ч. Полученные хроматограммы свидетель&
AAPH образование АФК, уровень внутриклеточN
ствуют о том, что фитохимическими вещества&
ного Ca2+, смягчает экстернализацию ФС и пониN
ми, присутствующими в экстракте TSCEE, яв&
жает уровень гибели эритроцитов. Было изучено
БИОХИМИЯ том 85 вып. 1 2020
144
КЕНГАЙЯ и др.
a
b
c
Рис. 1. Анализ с использованием HPLC и антиоксидантная активность экстракта TSCEE. На рис. 1 представлены резуль&
таты проведенного анализа TSCEE с использованием RP&HPLC и FTIR и циклической вольтмаперометрии. a - TSCEE
анализировали с помощью метода RP&HPLC с использованием колонки C18 (150 × 4,60 мм; размер частиц 5 мкм).
Колонка, предварительно уравновешенная 10%&м ацетонитрилом в воде для HPLC. Образец элюировали с использова&
нием линейного градиента (9 : 1 v/v) растворителя A (вода для HPLC) и растворителя B (ацетонитрил) со скоростью по&
тока 1 мл/мин в диапазоне 200&800 нм. Белок элюировали при скорости потока 1 мл/мин при мониторинге в диапазоне
200-800 нм. b - Спектры FTIR (Инфракрасная спектроскопия с преобразованием Фурье) экстракта TSCEE показывают
вероятные функциональные группы в TSCEE. c - Результаты цикловольтамперометрического анализа, объясняющие ре&
докс&потенциал экстракта оболочки семян тамаринда.
biokhsm/
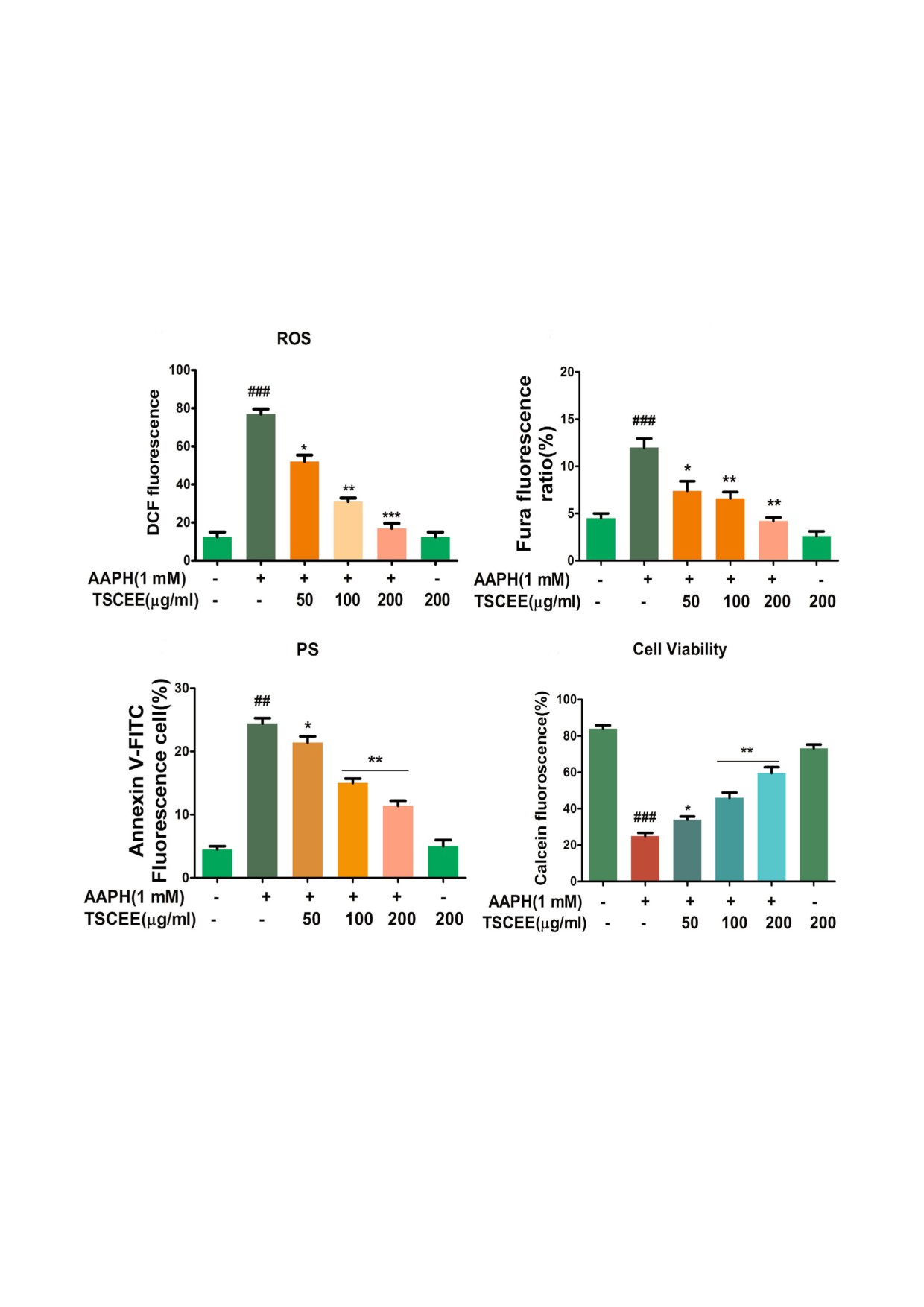
влияние экстракта TSCEE на запускаемую
TSCEE. Кроме того, экстракт TSCEE значитель&
AAPH генерацию АФК с помощью маркировки
но понижал уровень генерации АФК, вызванной
эритроцитов флуорофором DCF&DA. Значи&
AAPH, это явление зависело от дозы, и полное
тельное повышение уровня АФК наблюдали при
ингибирование наблюдали при концентрации
инкубации эритроцитов только с ААPH, в то
200 мкг (рис. 2, a). На рис. 2, b показано влияние
время как не было никаких изменений при ин&
TSCEE на индуцированное ААПГ повышение
кубации эритроцитов только с экстрактом
уровня Са2+ в эритроцитах. Для измерения уров&
БИОХИМИЯ том 85 вып. 1 2020
ЭРИПТОЗ И ЗАЩИТНАЯ ЭФФЕКТИВНОСТЬ СЕМЯН ТАМАРИНДА
145
ня содержания внутриклеточного Ca2+ исполь&
200 мкг. Устойчивое повышение внутриклеточ&
зовали флуоресцирующий индикатор их прису&
ного уровня Ca2+ является ключевым событием
тствия. Обработка отмытых эритроцитов с ис&
в инициировании скремблирования мембран и
пользованием АAPH вызывала повышение
экстернализации ФС, которое можно иденти&
уровня внутриклеточного Ca2+. Экстракт
фицировать с помощью окрашивания аннекси&
TSCEE повышал уровень внутриклеточного
ном V. Как видно на рис. 2, c AAPH усиливал
Ca2+ в результате дозозависимой индукции
связывание аннексина V, а экстракт TSCEE ин&
ААРН. Было обнаружено, что TSCEE осущес&
гибировал связывание аннексина в зависимос&
твляет значительную защиту при концентрации
ти от дозы, и значительное ингибирование на&
a
b
Ca2+
c
d
Рис. 2. Влияние TSCEE на AAPH&индуцированное образование АФК, внутриклеточный уровень ионов Ca2+, ФС и про&
цесс гибели эритроцитов. Влияние TSCEE на AAPH&индуцированное образование АФК и репрезентативная диаграмма,
на которой представлены средние значения изменений интенсивности флуоресценции DCF (a). b - Влияние TSCEE на
AAPH&индуцированные изменения уровня ионов Ca2+ внутри клеток. c - Влияние TSCEE на AAPH&индуцированную
экстернализацию ФС эритроцитов, и их репрезентативную гистограмму, показывающую медианные изменения интен&
сивности флуоресценции FITC&конъюгированного аннексина&V. d - Определение выживаемости клеток при использо&
вании регистрации флуоресценции кальцеина. Промытые эритроциты инкубировали с конечной реакционной смесью
2%&го гематокрита в растворе Рингера, дополненной AAPH в присутствии или в отсутствие TSCEE. Полученные данные
представлены в виде среднего значения ± SEM (n = 5), выраженных в виде процентов, на которые возросла флуоресцен&
ция DCF, Fura&2/AM и аннексина в сравнении с контролем. ## p < 0,01,### p < 0,001 достоверно по сравнению с контролем.
* p < 0,05, **p < 0,01, ***p < 0,001 достоверно по сравнению с AAPH.
biokhsm/
10 БИОХИМИЯ том 85 вып. 1 2020
146
КЕНГАЙЯ и др.
a
b
Рис. 3. Влияние TSCEE на AAPH&индуцированный окислительный стресс в эритроцитах и оценка маркером стресса. Со&
отношение GSH (a) и GSH/GSSG (b) в эритроцитах. Вкратце, промытые эритроциты инкубировали с конечной реакци&
онной смесью 2%&го гематокрита в PBS, дополненной AAPH (1 мМ) в присутствии или в отсутствие TSCEE. Для опреде&
ления соотношения GSH и GSH/GSSG были использованы лизаты отмытых эритроцитов, что достоверно отличалось от
контроля. Значения представлены в виде среднего ± SEM (n = 5); ### p < 0,001 достоверно по сравнению с контролем;
* p < 0,05, ** p < 0,01, *** p < 0,001 достоверно по сравнению с AAPH.
biokhsm/
a
b
c
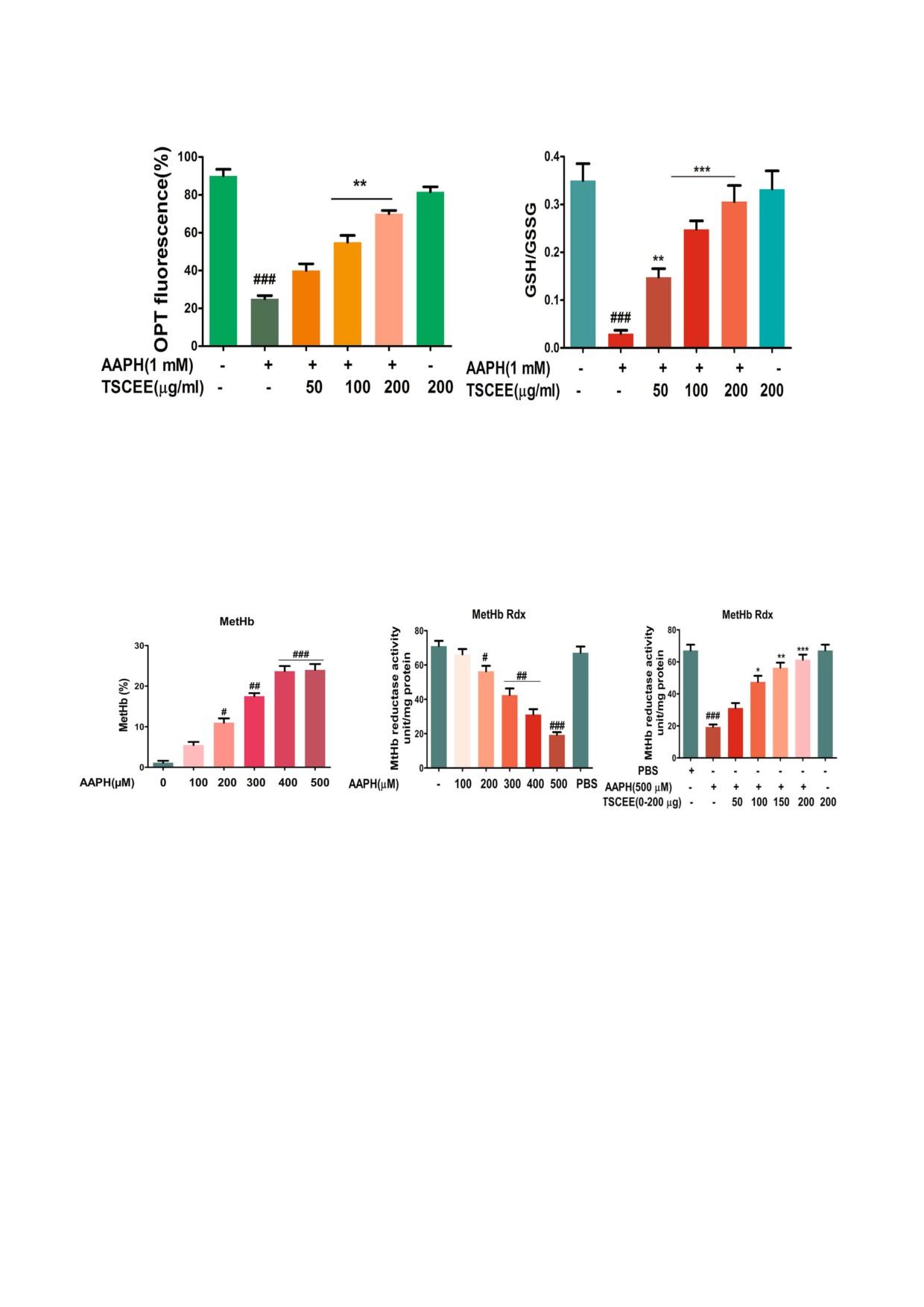
Рис. 4. Влияние AAPH на образование MetHb определяли по активности MetHb&редуктазы. a - AAPH дозозависимо ин&
дуцировал образование MetHb при концентрации 100, 200, 300, 400, 500 мкМ в течение 24 ч при 37 °C. b - активность
MetHb&редуктазы в лизате эритроцитов, обработанном AAPH (0&500 мкМ). c - Ингибирование AAPH&индуцированного
образования MetHb с помощью преинкубации с TSCEE. Все данные представлены в виде среднего ± SEM (n = 5);
# p < 0,05, ## p < 0,01, ### p < 0,001 по сравнению с контролем PBS; * p < 0,05, ** p < 0,01, *** p < 0,001 по сравнению толь&
ко с AAPH.
biokhsm/
блюдали при концентрации 200 мкг. Для опре&
морфологии клеток в концен&трации 200 мкг,
деления эффективности защиты экстракта
которая была сопоставима только с положи&
TSCEE для эритроцитов, обработанных AAPH,
тельными контрольными эри&троцитами. Поэ&
проводили анализ жизнеспособности клеток с
тому можно предположить, что TSCEE защи&
использованием кальцеинового красителя (рис.
щает эритроциты от эриптоза, индуцированно&
2, d). Примечательно, что TSCEE показал зна&
го окислительным стрессом.
чительную защитную эффективность на пов&
TSCEE восстанавливает уровень GSH в эрипN
режденных клетках эри&троцитов, поскольку
тотических эритроцитах. GSH играет жизненно
он способствовал сохранению нормальной
важную роль в смягчении окислительного
БИОХИМИЯ том 85 вып. 1 2020
ЭРИПТОЗ И ЗАЩИТНАЯ ЭФФЕКТИВНОСТЬ СЕМЯН ТАМАРИНДА
147
стресса, так как он участвует в детоксикации че&
эритроцитов, обработанном AAPH, дозозависи&
рез конъюгацию. Снижение уровня GSH оказы&
мым образом по сравнению с необработанным
вает негативное влияние на клеточную и моле&
лизатом эритроцитов (рис. 4, с).
кулярную функцию. Поэтому нами было изуче&
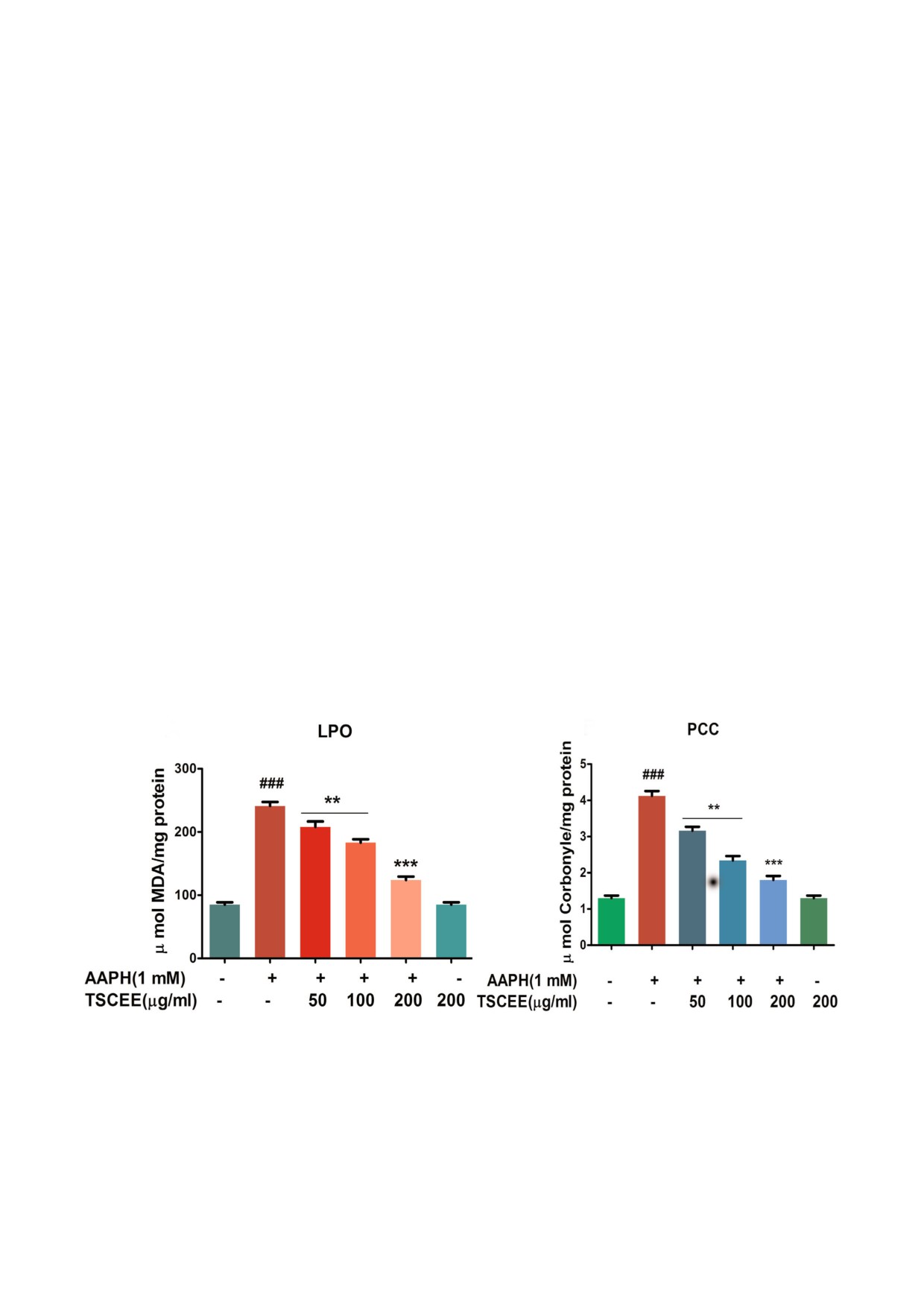
TSCEE уменьшает вызванные действием
но влияние экстракта TSCEE на уровень GSH в
AAPH перекисное окисление липидов и карбониN
эритроцитах, обработанных AAPH. В нормаль&
лирование белков. Перекисное окисление липи&
ных эритроцитах был выявлен обычный уровень
дов - ключевое событие, которое демонстриру&
GSH. С другой стороны, в эритроцитах, обрабо&
ет последствия окислительного стресса. На
танных AAPH, уровень GSH был понижен в
рис. 5, a представлены результаты изучения вли&
сравнении с контролем. Следует подчеркнуть,
яния экстракта TSCEE на индуцированное
что 200 мкг экстракта TSCEE способствовали
AAPH перекисное окисление липидов. Эритро&
восстановлению нормального уровня GSH в
циты, которые подверглись воздействию AAPH,
эритроцитах, обработанных AAPH (рис. 3, a).
демонстрировали значительное повышение
Кроме того, обработка клеток экстрактом
уровня перекисного окисления липидов, что
TSCEE эффективно восстанавливала соотноше&
можно видеть по существенному увеличению
ние GSH/GSSG, нарушенное применением
содержания малонового альдегида (МДА). Вы&
ААРН (рис. 3, b).
зывает интерес тот факт, что TSCEE понижал
TSCEE восстанавливает уровень редуктазы
уровень МДА в дозозависимом режиме, и пол&
MetHb в эриптотических эритроцитах. В обрабо&
ное ингибирование наступало при концентра&
танных AAPH лизатах РБК наблюдался повы&
ции 200 мкг/мл. Далее исследовали защитное
шенный процент образования MetHb по срав&
действие TSCEE на степень окисления белков в
нению с необработанным лизатом эритроцитов
эритроцитах человека. Экспериментальные ре&
(рис. 4, а). Исследовали активность редуктазы
зультаты показали значительное повышение
MetHb, ответственной за реверсию MetHb в ге&
уровня карбонильных групп в эритроцитах об&
моглобин. В поврежденных эритроцитах уро&
работанных AAPH по сравнению с контролем.
вень редуктазы MetHb был резко снижен по
Более того, экстракт TSCEE в концентрации
сравнению с лизатом эритроцитов, обработан&
200 мкг/мл вызывал значительное снижение
ным PBS (рис. 4, b). Интересно, что TSCEE нор&
уровня карбонильных групп в эритроцитах,
мализовал уровень редуктазы MetHb в лизате
подвергшихся воздействию AAPH (рис. 5, b).
a
b
Рис. 5. Влияние TSCEE на AAPH&индуцированный окислительный стресс в эитроцитах и оценка маркеров стресса.
Вкратце, промытые эритроциты инкубировали с конечной реакционной смесью 2%&го гематокрита в PBS, дополненной
AAPH (1 мМ) в присутствии или в отсутствие TSCEE. Влияние TSCEE на индуцированное AAPH (a) перекисное окисле&
ние липидов (LPO), b - содержание карбонильных групп в белках (PCC), LPO и PCC определяли с использованием об&
разцов мембран эритроцитов (1 мг/мл). Полученные данные представлены в виде среднего значения ± SEM (n = 5).
### p < 0,001 достоверно по сравнению с контролем; ** p < 0,01, *** p < 0,001 достоверно по сравнению с AAPH.
biokhsm/
БИОХИМИЯ том 85 вып. 1 2020
10*
148
КЕНГАЙЯ и др.
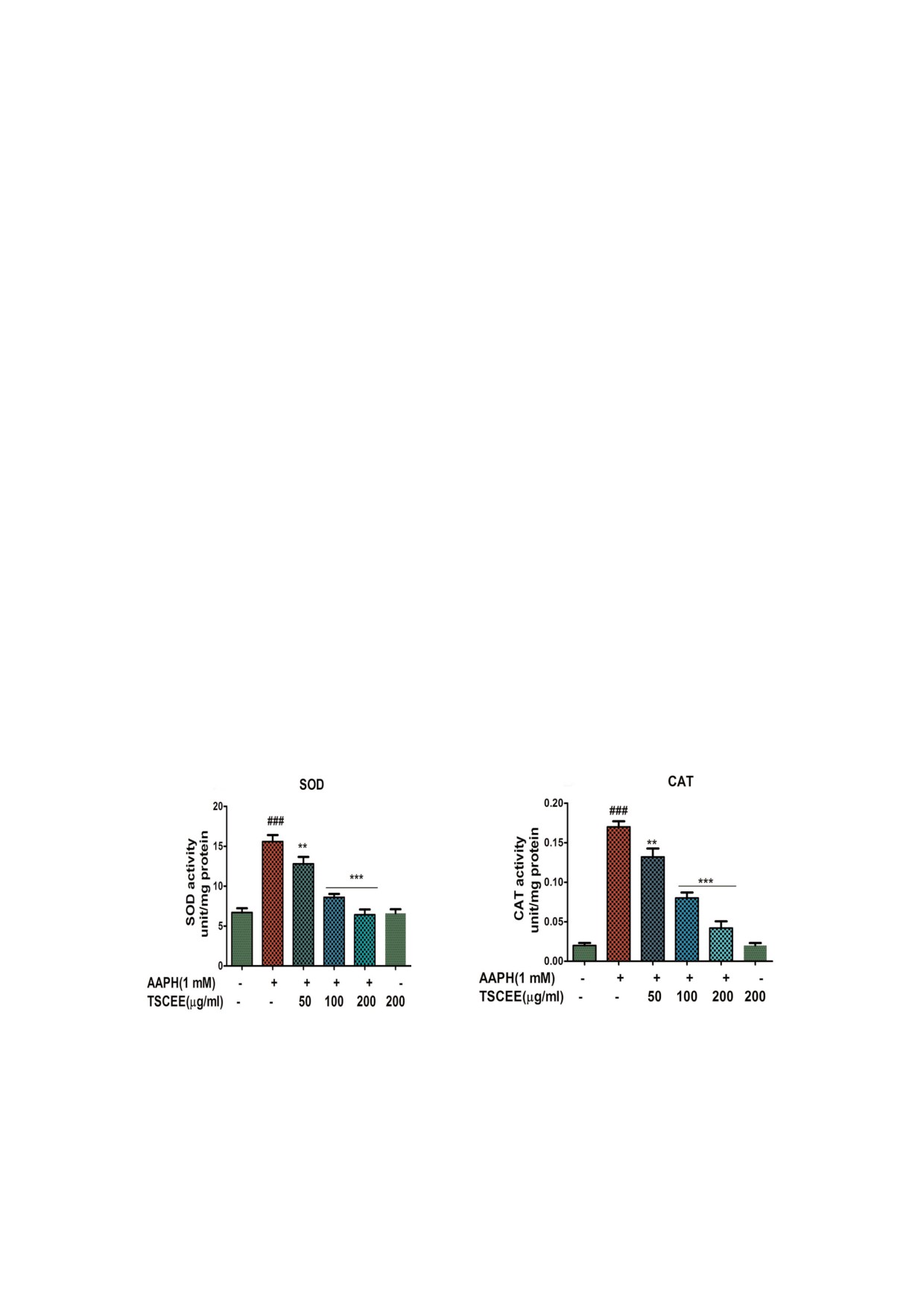
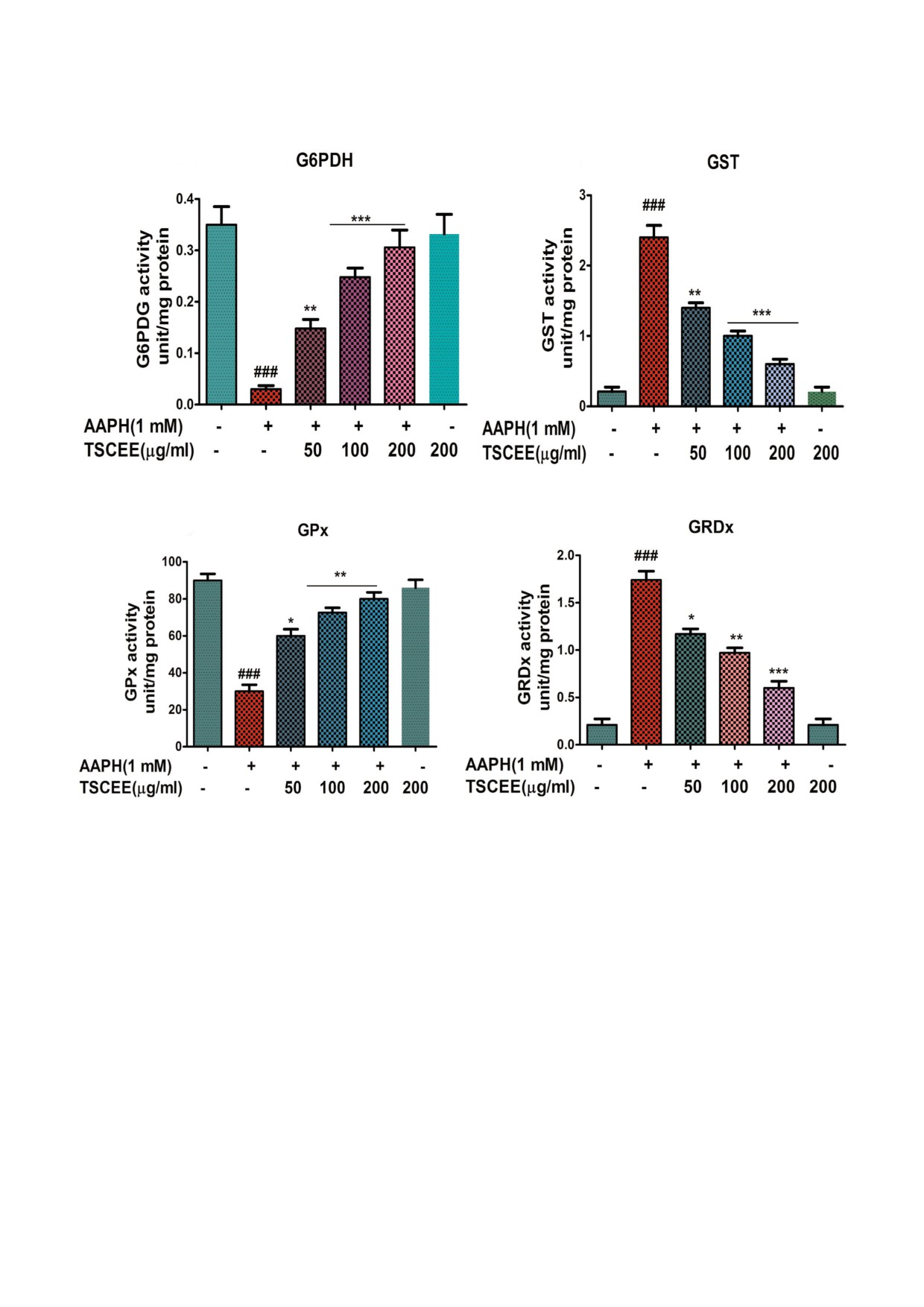
TSCEE модулирует АAPHNиндуцированное
существенному восстановлению активности
окислительное повреждение и восстанавливает
этих ферментов. Кроме того, активность фер&
активность ферментов, участвующих в окислиN
ментов, задействованных в кругообороте глута&
тельноNвосстановительном гомеостазе. Постоян&
тиона, таких как GRdx (рис. 7, c) и GPx (рис. 7, d),
ный окислительный стресс является первичным
снижалась после обработки клеток AAPH. В то
фактором, который истощает антиоксидантную
же время обработка эритроцитов экстрактом
систему клеток. Эндогенные антиоксидантные
TSCEE (200 мкг) приводила к существенному
ферменты, такие как СОД и КАТ, играют ключе&
восстановлению активности GRdx и GPx в
вую роль в восстановлении исходного редокс&
эритроцитах, обработанных AAPH.
состояния клеток. Поэтому в настоящей работе
было оценено защитное действие экстракта
TSCEE на уровни антиоксидантных ферментов.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Значительное повышение активности антиок&
сидантных ферментов (СОД и КАТ) было отме&
Эритроциты являются основными клеточ&
чено в эритроцитах, обработанных AAPH
ными компонентами, которые в кровотоке час&
(1 мМ) (рис. 6, a и b), и резкое снижение актив&
то подвергаются воздействию различных хими&
ности СОД и КАТ наблюдали в эритроцитах, об&
ческих соединений способных быстро разру&
работанных TSCEE (0-200 мкг). В дополнение к
шить мембрану эритроцитов и воздействовать
изменениям активности антиоксидантных фер&
на их нормальное функционирование. Эритро&
ментов еще одним ключевым событием в борьбе
циты являются основными несущими кислород
с окислительным стрессом является повышение
клетками, поэтому они наиболее склонны к
уровня эндогенного GSH. Поэтому в настоящей
окислительному повреждению, которое может в
работе были определены уровни ферментов, ко&
дальнейшем закончиться гемолизом или выз&
торые восстанавливают исходный уровень GSH,
вать самоубийственную смерть, называемую
включая GST, GR, GPx и G6PDH, в эритроци&
эриптозом [20]. Подобно апоптозу, эриптоз ха&
тах, обработанных AAPH (1 мМ) в присутствии
рактеризуется увеличением внутриклеточного
или в отсутствие TSCEE (рис. 7, a и b). Было по&
Ca2+, сморщиванием клеток, обесцвечиванием
казано, что в эритроцитах, обработанных AAPH
мембран и экспонированием фосфатидилсери&
(1 мМ), происходит значительное увеличение
на на клеточной мембране. Сообщалось, что
активности всех антиоксидантых ферментов.
окислительный стресс вызывает эриптоз путем
Более того, наблюдали значительное пониже&
активации чувствительного к ионам Ca2+ про&
ние активности G6PDH и GST, а обработка кле&
ницаемого неизбирательного катионного кана&
ток экстрактом TSCEE (200 мкг) приводила к
ла. Проникновение внутрь клетки значительных
a
b
Рис. 6. TSCEE модулирует AAPH&индуцированные окислительные повреждения и восстанавливает активность фермен&
тов, участвующих в окислительно&восстановительном гомеостазе. Активность супероксиддисмутазы (a) и каталазы (b) в
эритроцитах. Вкратце, промытые эритроциты инкубировали с конечной реакционной смесью 2%&го гематокрита в PBS,
дополненной AAPH (1 мМ) в присутствии или в отсутствие TSCEE. Полученные данные представлены в виде среднего
значения ± SEM (n = 5); ### p < 0,001 достоверно по сравнению с контролем; ** p < 0,01, *** p < 0,001 достоверно по срав&
нению с AAPH.
biokhsm/
БИОХИМИЯ том 85 вып. 1 2020
ЭРИПТОЗ И ЗАЩИТНАЯ ЭФФЕКТИВНОСТЬ СЕМЯН ТАМАРИНДА
149
a
b
c
d
Рис. 7. Влияние TSCEE на активность антиоксидантных ферментов в эритроцитах, обработанных AAPH. Вкратце, про&
мытые эритроциты инкубировали с конечной реакционной смесью 2%&го гематокрита в PBS, дополненной AAPH (1 мМ)
в присутствии или в отсутствие TSCEE (a). b - Активность G6PDH и GST. c - Активность GPx. d - Активность GRdx.
Значения представлены в виде среднего ± SEM (n = 5); ### p < 0,001 достоверно по сравнению с контролем; * p < 0,05,
** p < 0,01, *** p < 0,001 достоверно по сравнению с AAPH.
biokhsm/
количеств ионов Ca2+ может привести к возник&
того, окислительный стресс может привести к
новению последующих событий эриптоза, на&
активации каспаз, тем самым запуская эриптоз
пример, сморщивание клеток путем активации
[21].
каналов, чувствительных к ионам Ca2+ и K+, с
При некоторых случаях окислительного
последующей гиперполяризацией мембраны.
стресса, ассоциированного с патологическими
Выход ионов Ca2+ вызывает сморщивание кле&
состояниями, такими как рак, диабет, ХБП и
ток из&за потери KCl вместе с водой в результате
др., наблюдается повышенная гибель эритроци&
осмоса. Повышение концентрации ионов Ca2+
тов в результате эриптоза и последующая тяже&
также ассоциируется со скремблингом мембран
лая анемия. Кроме того, было показано, что
и ФС, основными признаками эриптоза. Кроме
несколько противоопухолевых препаратов так&
БИОХИМИЯ том 85 вып. 1 2020
10*
150
КЕНГАЙЯ и др.
же индуцируют эриптоз [3, 21]. Таким образом,
подготавливая их к эриптотической гибели кле&
регулирование окислительного стресса может
ток [21]. Таким образом, способность экстракта
стать ключом к борьбе с анемией при этих кли&
TSCEE противостоять эриптозу может быть
нических заболеваниях. Было показано, что ряд
связана с его врожденным антиоксидантным
антиоксидантных молекул растительного про&
потенциалом, благодаря которому он может ту&
исхождения обеспечивают защиту от индуциро&
шить АФК и таким образом снижать уровень
ванного окислительным стрессом эриптоза [22].
окислительного стресса. Более того, TSCEE в
Лекарственное использование различных час&
значительной степени понижал уровень переки&
тей тамариндового дерева хорошо документиро&
сей липидов и содержание карбонильных групп
вано в традиционной медицине [23]. В настоя&
в белках, которые являются маркерами окисли&
щем исследовании оценивали защитную эф&
тельного стресса. Перекисное окисление липи&
фективность TSCEE против индуцированного
дов является одним из основных индикаторов
окислительным стрессом эриптоза. Хроматог&
окислительных повреждений, вызванных АФК.
рамма экстракта TSCEE, полученная при RP&
Эритроциты богаты ненасыщенными липида&
HPLC анализе, показала присутствие различ&
ми, и их мембраны более склонны к окисли&
ные биологически активные компоненты таких
тельному повреждению. Известно, что большое
как флавоноиды, каротиноиды и полифенолы.
количество перекисей липидов вызывает нару&
Более того, данные анализа FTIR указывают на
шение нормального функционирования клетки,
присутствие в растительных компонентах
снижение текучести мембран, инактивацию
экстракта TSCEE карбонильных, гидроксиль&
мембраносвязанных рецепторов и ферментов и
ных и кетоновых групп. Гидроксильные группы
повышение неспецифической проницаемости
могут тушить образовавшиеся во время окисли&
ионов, что в итоге приводит к повреждению
тельного стрессa свободные радикалы, и это
клетки [27-29]. Кроме того, карбонилирование
обстоятельство может быть отнесено на счет эф&
белков может вызвать изменения структуры и
фективности защиты экстракта TSCEE. Более
функций клетки [30]. Содержание продуктов
того, с помощью циклических вольтамперомет&
перекисного окисления липидов и карбониль&
рических исследований также была подтвержде&
ных групп в белках также в значительной степе&
на антиоксидантная активность экстракта
ни снижалось в результате действия TSCEE, это
TSCEE.
свидетельствовало о том, что TSCEE может ин&
AAPH запускает окислительный стресс, ин&
гибировать вызванные воздействием ААРН на&
дуцирующий образование метгемоглобина с по&
рушения в эритроцитах.
ниженной редуктазной активностью. Превра&
Выживание эритроцитов абсолютно зависит
щение гемоглобина в MetHb может отменить
от их способности поддерживать клеточный
способности связывать молекулярный кисло&
окислительно&восстановительный гомеостаз.
род, превращая его в нефункциональный белок.
Как ферментативные, так и неферментативные
Это может привести к возникновению состоя&
механизмы работают совместно для борьбы с
ния, называемого метгемоглобинемией [24-26].
окислительным стрессом. GSH - основной эн&
Метгемоглобинредуктаза является ключевым
догенный антиоксидант, который помогает клет&
ферментом, который превращает MetHb в ге&
кам избавляться от повышенных уровней АФК
моглобин и восстанавливает его функциональ&
[31, 32]. Было обнаружено, что TSCEE восполня&
ную активность. Возможно, благодаря сниже&
ет количество глутатиона в эритроцитах, кото&
нию уровня окислительного стресса и восста&
рые были предварительно обработаны индукто&
новлению активности метгемоглобинредуктазы
ром свободных радикалов, AAPH. Кроме того,
TSCEE в значительной степени снижает образо&
экстракт TSCEE также способствовал восстанов&
вание MetHb. На основе предыдущих исследо&
лению активности антиоксидантных ферментов,
ваний можно предполагать, что окислительный
таких как СОД и КАТ. Кроме того, TSCEE вос&
стресс является главным фактором, индуцирую&
становил активность ферментов (GST, GR GPx и
щим образования MetHb. Любые такие измене&
G6PDH), участвующих в пополнении GSH. В
ния делают эритроциты более восприимчивыми
процессе регенерации GSH молекула НАДФН
к старению и гибели клеток. Поврежденные
является одним из ключевых факторов, которые
эритроциты удаляются из кровотока в результа&
образуются в эритроцитах с помощью фермента
те особого процесса гибели клеток, который на&
глюкозо&6&фосфатдегидрогеназы (G6PDH) в хо&
зывается эриптозом. Повышение концентрации
де гексозомонофосфатного пути (HMP shunt
АФК и последующее проникновение ионов Ca2+
pathway) [33]. Поэтому защитная эффективность
в эритроциты, в свою очередь, может активиро&
TSCEE может быть обусловлена ее способ&
вать каспазы и способствовать транслокации
ностью восстанавливать клеточный окислитель&
ФС на поверхность эритроцитов, тем самым
но&восстановительный гомеостаз.
БИОХИМИЯ том 85 вып. 1 2020
ЭРИПТОЗ И ЗАЩИТНАЯ ЭФФЕКТИВНОСТЬ СЕМЯН ТАМАРИНДА
151
В заключение, можно резюмировать, что
Дели за представленную стажировку. DS благода&
TSCEE способен эффективно защищать эритро&
рит проф. K Кемпарайю, университет Мисора,
циты от индуцированных ААРН окислительных
проф. Гириш K.С., доктора П. Рагхавендра Ку&
повреждений и эриптоза. Это может быть связа&
мар, Бхагялаксми M., университет Тумкура за их
но с наличием активных фитохимических ве&
помощь при выполнении данной работы. Авторы
ществ, таких как алкалоиды, флавоноиды и по&
также выражают благодарность Маникара M. и
лифенольные соединения. Благодаря способнос&
Светакумар Б., университет Мисора за их любез&
ти тушить свободные радикалы терапевтические
ную помощь при выполнении работы.
средства на основе лекарственных растений мо&
Конфликт интересов. Авторы заявляют об от&
гут стать эффективным средством в борьбе с
сутствии конфликта интересов.
окислительным стрессом и возникающим вслед&
Соблюдение этических норм. Все процедуры,
ствие его эриптозом и противодействовать ане&
выполненные в исследовании с участием людей,
мии при различных патологических состояниях.
соответствуют этическим стандартам институ&
ционального и/или национального комитета по
исследовательской этике и Хельсинкской де&
Финансирование. Работа была выполнена при
кларации 1964 года и ее последующим измене&
финансовой поддержке Правительства Индии,
ниям или сопоставимым нормам этики. От каж&
Нью Дели и VGST и Правительства Каранатака.
дого из включенных в исследование участников
Благодарности. Джайанна выражает благодар&
было получено информированное доброволь&
ность UGC&RGNF, Правительству Индии, Нью ное согласие.
СПИСОК ЛИТЕРАТУРЫ
1.
Palis, J. (2014) Primitive and definitive erythropoiesis in
Hemshekhar, M., Girish, K.S., and Kemparaju, K.
mammals, Front. Physiol., 5, 3, doi: 10.3389/fphys.2014.
(2015) Oxidative stress&induced methemoglobinemia
00003.
is the silent killer during snakebite: a novel and strategic
2.
Lang, K.S., Lang, P.A., and Bauer, C. (2005) Mechanisms
neutralization by melatonin, J. Pineal Res., 59, 240-254.
of suicidal erythrocyte death, Cell. Physiol. Biochem., 15,
12. Driver, A.S., Kodavanti, P.R., and Mundy, W.R. (2000)
195-202.
Age related changes in reactive oxygen species production
3.
Bissinger, R., Modicano, P., Alzoubi, K., Honisch, S.,
in rat brain homogenates, Neurotoxicol. Teratol., 22,
Faggio, C., Abed, M., and Lang, F. (2014) Effect of
175-181.
saponins on erythrocytes, Int. J. Hematol., 100, 51-59.
13. NaveenKumar, S.K., Thushara, R.M., Sundaram, M.S.,
4.
Lang, E., Bissinger, R., Qadri, S.M., and Lang, F. (2017)
Hemshekhar, M., Paul, M., Thirunavukkarasu, C.,
Suicidal death of erythrocytes in cancer and its chemother&
Basappa, Nagaraju, G., Raghavan, S.C., Girish, K.S.,
apy: a potential target in the treatment of tumor&associated
Kemparaju, K., and Rangappa, K.S. (2015) Unconjugated
anaemia, Int. J. Cancer, 141, 1522-1528.
bilirubin exerts pro&apoptotic effect on platelets via p38&
5.
Lang, F., Lang, K.S., Lang, P.A., Huber, S.M., and Wieder, T.
MAPK activation, Sci. Rep., 5, 15045.
(2006) Mechanisms and significance of eryptosis, Antioxid.
14. Ohkawa, H., Ohishi, N., and Yagi, K. (1979) Assay for lipid
Redox Signal., 8, 1183-92.
peroxides in animal tissues by thiobarbituric acid reaction,
6.
Girish, K.S., Paul, M., Thushara, R.M., Hemshekhar, M.,
Anal. Biochem., 95, 351-358.
Shanmuga, S.M., Rangappa, K.S., and Kemparaju, K.
15. Levine, R.L., Garland, D., and Oliver, C.N.
(1990)
(2013) Melatonin elevates apoptosis in human platelets via
Determination of carbonyl content in oxidatively modified
ROS mediated mitochondrial damage, Biochem. Biophys.
proteins, Methods Enzymol., 186, 464-478.
Res. Commun., 438, 198-204.
16. Sundaram, M.S., Hemshekhar, M., Thushara, R.M.,
7.
Sundaram, M.S., Hemshekhar, M., Santhosh, M.S., Paul,
Santhosh, M.S., Kumar, S.K., Paul, M., Devaraja, S.,
M., Sunitha, K., Thushara, R.M., Naveen Kumar, S.K.,
Kemparaju, K., Rangappa, K.S., and Girish, K.S. (2014)
Devaraja, S., Rangappa, K.S., Kemparaju, K., and Girish, K.S.
Tamarind seed extract mitigates the liver oxidative stress in
(2015) Tamarind Seed (Tamarindus indica) Extract amelio&
arthritic rats, Food Funct., 5, 587-597.
rates adjuvant&induced arthritis via regulating the media&
17. Tung, B.T., Hai, N.T., and Thu, D.K. (2017) Antioxidant
tors of cartilage/bone degeneration, inflammation and
and acetylcholinesterase inhibitory activities in vitro of
oxidative stress, Sci. Rep., 5, 11117.
different fraction of Huperzia squarrosa (Forst.) Trevis
8.
Kareru, P.G., Keriko, J.M., Gachanja, A.N., and Kenji, G.M.
extract and attenuation of scopolamine&induced cognitive
(2008) Direct detection of triterpenoid saponins in medic&
impairment in mice, J. Ethnopharmacol., 198, 24-32.
inal plants, Afr. J. Tradit. Complement. Altern. Med., 5,
18. Guthenberg, C., Alin, P., and Mannervik, B.
(1985)
56-60.
Glutathione transferase from rat testis, Methods Enzymol.,
9.
Arulpriya, P., Lalitha, P., and Hemalatha, S. (2010) Cyclic
113, 507-510.
voltametric assessment of the antioxidant activity of petro&
19. Lowry, O.H., Rosebrough, N.J., Farr, A.L., and Randall, R.J.
leum ether extract of Samanea saman (Jacq) Merr, Adv.
(1951) Protein measurement with the folin phenol reagent,
Appl. Sci. Res., 1, 24-35.
J. Biol. Chem., 193, 265-275.
10.
Benesch, R.E., Benesch, R., and Yung, S.
(1973)
20. Föller, M., Huber, S.M., and Lang, F. (2008) Erythrocyte
Equations for the spectrophotometric analysis of hemoglo&
programmed cell death, IUBMB Life, 60, 661-668.
bin mixtures, Anal. Biochem., 55, 245-248.
21. Lang, F., Abed, M., Lang, E., and Föller, M. (2014)
11.
Sharma, R.D., Katkar, G.D., Sundaram, M.S.,
Oxidative stress and suicidal erythrocyte death, Antioxid.
Paul, M., NaveenKumar, S.K., Swethakumar, B.,
Redox Signal, 21, 138-153.
БИОХИМИЯ том 85 вып. 1 2020
152
КЕНГАЙЯ и др.
22.
Jagadisha, S., Hemshekhara, M., NaveenKumara, S.K.,
28. Shaik, N., Lupescu, A., and Lang, F. (2013) Inhibition of
Sharath Kumar, K.S., Sundaram, M.S., Basappa., Girisha, K.S.,
suicidal erythrocyte death by probucol, J. Cardiovasc.
and Rangappa, K.S. (2017) Novel oxolane derivative
Pharmacol., 61, 120-126.
DMTD mitigates high glucose induced erythrocytes apo&
29. Zierle, J., Bissinger, R., and Lang, F. (2016) Inhibition by
ptosis by regulating oxidative stress, Toxicol. Appl.
teriflunomide of erythrocyte cell membrane scrambling
Pharmacol., 334,167-179.
following energy depletion, oxidative stress and ionomycin,
23.
Bhadoriya, S.S., Ganeshpurkar, A., Narwaria, J., Rai, G.,
Cell. Physiol. Biochem., 39, 1877-1890.
and Jain, A.P. (2011) Tamarindus indica: extent of explored
30. Sebastin Santhosh, M., Hemshekhar, M., Thushara, R.M.,
potential, Pharmacogn. Rev., 5, 73-71.
Devaraja, S., Kemparaju, K., and Girish, K.S. (2013)
24.
Dong, X., Kiss, L., Petrikovics, I., and Thompson, D.E.
Vipera russelli venom induced oxidative stress and hemato&
(2017) Reaction of dimethyl trisulfide with haemoglobin,
logical alterations: amelioration by crocin a dietary col&
Chem. Res. Toxicol., 30, 1661-1663.
orant, Cell. Biochem. Funct., 31, 41-50.
25.
Ding, Y., Liu, H., Tekwani, B.L., Nanayakkara, N.P.D.,
31. Vitak, T.Y., Wasser, S.P., Nevo, E., and Sybirna, N.O.
Khan, I.A., Walker, L.A., and Doerksen, R.J.
(2016)
(2017) Enzymatic system of antioxidant protection of
Methemoglobinemia hemotoxicity of some antimalarial 8&
erythrocytes in diabetic rats treated with medicinal mush&
aminoquinoline analogues and their hydroxylated deriva&
rooms Agaricus brasiliensis and Ganoderma lucidum
tives: density functional theory computation of ionization
(Agaricomycetes), Int. J. Med. Mushrooms, 19, 697-708.
potentials, Chem. Res. Toxicol., 29, 1132-1141.
32. Jha, R., and Rizvi, S.I. (2011) Carbonyl formation in
26.
Puchulu&Campanella, E., Chu, H., Anstee, D.J., Galan,
erythrocyte membrane proteins during aging in humans,
J.A., Tao, W.A., and Low, P.S. (2013) Identification of the
Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech.
components of a glycolytic enzyme metabolon on the
Repub., 155, 39-42.
human red blood cell membrane, J. Biol. Chem., 288,
33. Vishalakshi, G.J., Hemshekhar, M., Kemparaju, K., and
848-858.
Girish, K.S. (2018) Para&tertiary butyl catechol induces
27.
Qadri, S.M., Mahmud, H., Föller, M., and Lang, F. (2009)
eryptosis in vitro via oxidative stress and hemoglobin leak&
Inhibition of suicidal erythrocyte death by xanthohumol,
age in human erythrocytes, Toxicol. In Vitro,
52,
J. Agric. Food Chem., 57, 7591-7595.
286-296.
PROTECTIVE EFFICACY OF TAMARIND SEED COAT ETHANOLIC
EXTRACT ON OXIDATIVE STRESS INDUCED ERYPTOSIS
J. Kengaiah1, S. K. M. Nandish1, Ch. Ramachandraiah1, Chandramma1, A. Shivaiah1,
G. J. Vishalakshi2, M. Paul2, S. M. Santhosh3, R. L. Shankar4, and D. Sannaningaiah1*
1 Department of Studies and Research in Biochemistry and Centre for Bioscience and Innovation,
Tumkur University, 572103 Tumkur, India
2 Department of Studies in Biochemistry, University of Mysore, Manasagangothry, 570006 Mysore, India
3 Department of Medical Biochemistry and Microbiology (IMBM), Uppsala Biomedical Centre, 75237 Uppsala, Sweden
4 Department of Sericulture, Yuvaraja’s College, University of Mysore, 570006 Mysore, India; E mail: sdevbiochem@gmail.com
Received April 1, 2019
Revised October 4, 2019
Accepted October 17, 2019
Suicidal erythrocyte death or eryptosis is the key event in eliciting anaemia in multitude of pathological conditions
including diabetes, chronic kidney diseases, cancer, sepsis, etc. Oxidative stress is an important trigger in accelerating
erythrocyte loss by eryptosis and has been shown to be the underlying mechanism for anaemic conditions in above
said pathologies. Thus, the identification of antioxidant and anti&eryptotic agents for management of the oxidative
stress&related ailments is in increasing demand today. The current work demonstrates antioxidant and anti&eryptotic
assets of tamarind seed coat ethanol extract (TSCEE) against AAPH (2,2′&azobis(2&amidinopropane)hydrochloride)&
induced oxidative stress and eryptosis. The presence of probable secondary metabolites in the TSCEE extract was
investigated by RP&HPLC column chromatography. The active groups present in the TSCEE were studied using the
Fourier&transform infrared spectroscopy. Cyclic voltammetric studies confirmed the antioxidant potential of TSCEE.
Further, the red blood cells protection property of TSCEE was confirmed by assessing various eryptotic markers such
as reactive oxygen species, cellular calcium level, and phosphatidyl serine exposure. In addition, TSCEE reduced lipid
peroxidation and protein carbonyl content, restored cellular glutathione level, antioxidant enzymes and enzymes
involved in glutathione replenishment. Thus, TSCEE may be a promising therapeutic agent for curing oxidative
stress&induced eryptosis and subsequent anaemia in various pathologies.
Keywords: tamarind seed coat ethanol extract (TSCEE), reactive oxygen species (ROS), eryptosis, GSH/GSSG,
G6PDH, antioxidant, AAPH
БИОХИМИЯ том 85 вып. 1 2020