БИОХИМИЯ, 2020, том 85, вып. 10, с. 1398 - 1410
УДК 577.24
ВИРУСНЫЕ ИНФЕКЦИИ: НЕГАТИВНЫЙ РЕГУЛЯТОР АПОПТОЗА
И ФАКТОР ОНКОГЕННОСТИ
Обзор
© 2020
А.В. Замараев1, Б. Животовский1,2, Г.С. Копеина1*
1 Московский государственный университет имени М.В. Ломоносова, факультет фундаментальной медицины,
119191 Москва, Россия; электронная почта: lirroster@gmail.com
2 Каролинский институт, Департамент медицины окружающей среды, SE*171 77 Стокгольм, Швеция
Поступила в редакцию 30.06.2020
После доработки 09.09.2020
Принята к публикации 09.09.2020
Нарушение процесса программируемой апоптотической гибели клеток тесно связано с этиологией различ
ных заболеваний, включая рак. Постоянные вирусные инфекции способны вызывать некоторые виды но
вообразований. Для этого онкогенные вирусы манипулируют как внешним путем запуска апоптоза, так и
внутренним, подавляя активность проапоптотических белков и сигнальных путей. Неадекватный иммун
ный надзор или подавление иммунного ответа способны индуцировать бесконтрольное размножение виру
са и пролиферацию клеток хозяина. В данном обзоре представлены современные данные о механизмах по
давления апоптотической гибели вирусами и их роль в онкогенезе.
КЛЮЧЕВЫЕ СЛОВА: онкогенные вирусы, апоптоз, онкологические заболевания.
DOI: 10.31857/S0320972520100073
ВВЕДЕНИЕ
Выделяют два основных пути передачи сиг
нала апоптоза: внешний путь (рецептор зависи
В многоклеточных организмах процесс апо
мый сигнальный путь с участием рецепторов
птотической гибели играет важную роль в под
смерти) и внутренний путь (митохондриаль
держании клеточного гомеостаза, морфогенеза,
ный). Внешний путь клеточной гибели начина
а также задействован на разных этапах развития
ется с взаимодействия специфических лигандов
и функционирования иммунной системы и уда
гибели с рецепторами смерти (CD95, TRAIL
лении поврежденных или инфицированных
R1/2, TNFR1) на поверхности клетки. Лиганды,
клеток [1]. Запуск апоптоза осуществляется пос
взаимодействующие с рецепторами, вызывают
редством внеклеточных или внутриклеточных
их олигомеризацию или конформационные из
факторов, таких как активация рецепторов
менения рецептора, что способствует присоеди
смерти, нарушение сигналов клеточного цикла,
нению белков адаптеров (FADD, TRADD) и не
накопление неправильно свернутых белков,
активных предшественников цистеиновых про
повреждение ДНК, метаболические нарушения,
теаз - инициаторных каспаз (прокаспаза 8,
10)
а также в ответ на различные бактериальные и
[2]. В составе такого высокомолекулярного
вирусные инфекции.
комплекса DISC происходит активация иници
Принятые сокращения: AIF - фактор, индуцирующий апоптоз; Akt - протеинкиназа В; Apaf 1 - фактор актива
ции апоптотической протеазы 1; Bcl 2 - семейство белков, регулирующих апоптоз; CD95/Fas/Apo 1 - рецептор смер
ти; cIAP- клеточный ингибитор белков апоптоза; DISC - сигнальный комплекс, индуцирующий клеточную гибель;
DREAM - белковый комплекс, ответственный за регулирование экспрессии генов клеточного цикла; EndoG - эндонук
леаза G; FADD - белок, взаимодействующий с доменом смерти CD95 рецептора; HBx - белок вируса гепатита Б;
HTLV 1 - ретровирус Т клеточного лейкоза человека 1 го типа; KSHV - герпесвирус, ассоциированный с саркомой Ка
поши; NF kB - транскрипционный ядерный фактор каппа Би; PI3K - фосфатидилинозитол 3 киназа; SARS CoV - од
ноцепочечный РНК вирус семейства короновирусов; TNF - фактор некроза опухолей; TNFR1 - рецептор фактора нек
роза опухоли 1; TRADD - белок, взаимодействующий с доменом смерти TNFR1 рецептора; TRAIL R1/2 - рецептор, от
носящийся к семейству TNF, инициирующий апоптоз; АФК - активные формы кислорода; ВИЧ 1 - вирус иммунодефи
цита человека; ВПЧ - вирус папилломы человека; ВЭБ - вирус Эпштейна-Барра; дАТФ - 2′ дезоксинуклеозид 5′ три
фосфат; ДНК - дезоксирибонуклеиновая кислота.
* Адресат для корреспонденции.
1398
ВИРУСЫ, АПОПТОЗ И КАНЦЕРОГЕНЕЗ
1399
аторных каспаз, которые вызывают, в свою оче
Некоторые вирусные инфекции для репли
редь, активацию эффекторных каспаз (каспазы
кации и выживания способны подавлять апоп
3,
6,
7) [3]. Данный каспазный каскад приво
тотическую гибель клеток. Многие вирусы, в
дит к гидролизу белков ядерной ламины, цитос
том числе и онкогенные, модулируют функции
келета, протеин киназ, белков репарации ДНК,
онкосупрессорного белка р53, активируют ан
а также инактивирует белки ингибиторы апоп
тиапоптотические сигнальные пути, ингибиру
тоза. В результате этих процессов происходят
ют рецептор зависимый и внутренний путь
несовместимые с поддержанием нормального
апоптоза, регулируя баланс про и антиапопто
состояния биохимические и морфологические
тических факторов. Подобные изменения апоп
изменения клетки.
тотических путей приводят к нарушению гомео
Внутриклеточные сигналы, такие как пов
статического баланса, что способствует прог
реждение ДНК клетки, окислительный стресс,
рессированию инфекции и вирус индуцирован
нарушение клеточного цикла или других сиг
ных патологий (таблица). Ниже представлены
нальных путей, приводят к апоптозу главным
различные механизмы вирусных инфекций,
образом через внутренний путь. Ключевым со
участвующих в подавлении апоптоза, и их роль в
бытием внутреннего пути является пермеабили
трансформации клеток.
зация митохондриальной мембраны, обуслов
ленное действием проапоптотических белков
семейства Bcl 2, таких как Bax и Bak. Образова
РЕГУЛЯЦИЯ ОНКОСУПРЕССОРНОГО
ние митохондриальных пор приводит к высво
БЕЛКА р53
бождению из митохондрий в цитоплазму проа
поптотических белков, таких как цитохром с,
Одним из центральных участников апопто
AIF, EndoG и др., через высокопроницаемые ка
тических сигнальных путей является онкосуп
налы на внешней поверхности митохондрий.
рессорный белок p53. Этот транскрипционный
Цитохром с, связываясь с белком Apaf 1 в при
фактор регулирует экспрессию ~500 целевых ге
сутствии дАТФ, вызывает его олигомеризацию,
нов, тем самым контролируя широкий спектр
обеспечивая рекрутирование прокаспазы 9. Та
клеточных процессов, включая остановку кле
ким образом, в цитоплазме клетки формируется
точного цикла, старение клеток, репарацию
белковый комплекс под названием апоптосома.
ДНК, метаболическую адаптацию и гибель кле
В составе этого комплекса происходит актива
ток [18]. Многие вирусные инфекции, чтобы до
ция инициаторной каспазы 9, которая, в свою
биться успешной репликации и распростране
очередь, активирует эффекторные каспазы,
ния, используют различные способы манипуля
приводя клетку к гибели [4].
ции функциями p53. Например, для экспансии
Виды опухоли, вызванные онкогенными вирусами [17]
Вирус
Вид опухоли
Ссылки
Вирус Эпштейна-Барра
40% лимфомы Ходжкина
[5, 6]
>95% эндемической лимфомы Беркитта
10% рак желудка
большинство носоглоточной карциномы (тип II и III)
саркома Капоши
другие лимфомы
Вирус гепатита В
53% гепатоцеллюлярной карциномы
[7]
Т лимфотропный вирус человека 1
> 99% взрослого Т клеточного лейкоза
[8, 9]
Вирус папилломы человека
> 95% рака шейки матки
[10, 11]
70% рака ротоглотки
другие аногенитальные карциномы
Вирус гепатита С
25% гепатоцеллюлярной карциномы
[12, 13]
неходжкинские В клеточные лимфомы
Герпесвирус, связанный с саркомой Капоши
> 99% саркомы Капоши
[14]
> 99% первичной выпотной лимфомы
Клеточный полиомавирус Меркеля
80% клеточной карциномы Меркеля
[15, 16]
БИОХИМИЯ том 85 вып. 10 2020
1400
ЗАМАРАЕВ и др.
некоторые вирусы вызывают p53 опосредован
зовать аутофагический путь для усиления своей
ную апоптотическую гибель клеток хозяина, од
репликации и подавления противовирусного
нако, некоторые вирусы могут стимулировать
интерферон зависимого пути.
пролиферацию клеток, ослабляя функцию p53,
Такие ДНК вирусы, как аденовирусы и SV40,
регулирующую клеточный цикл, и тем самым
используют механизм репликации ДНК клетки
могут провоцировать онкогенные процессы
хозяина для распространения вирусной инфек
[19].
ции. Для этого вирусный Т антиген или белки
Одним из таких примеров является вирус ге
аденовируса Е1В связывают р53 и инактивируют
патита В. Исследования этого вируса показали,
его, позволяя клетке избегать остановки клеточ
что кодируемый им белок HBx (hepatitis B virus X
ного цикла и способствуя переходу в S фазу [25].
protein) способен негативно влиять на запуск
Более того, Т антиген вируса обладает свой
р53 индуцированного апоптоза [20]. На клеточ
ством «ДНК мимикрии» [25], имитируя заряд и
ных линиях гепатоцеллюлярной карциномы че
контур дуплекса ДНК. Данная особенность поз
ловека HepG2 и Hep3B была показана связь бел
воляет Т антигену связываться с р53 и тем са
ка HBx с цитоплазматическим р53, а также с
мым нарушать транскрипционную регуляцию
белком данного семейства p73. Данное взаимо
генов мишеней р53, что приводит к трансфор
действие предотвращало их ядерную транслока
мации клетки хозяина [26]. Связь между р53 и
цию и запуск проапоптотических факторов. В
SV40 может не только подавлять нормальные
дополнение к этому HBx усиливает экспрессию
функции р53, но и стимулировать «вспомога
N терминальной изоформы р73, которая явля
тельную» активность для развития SV40. N кон
ется трансдоминантным ингибитором р53 [21].
цевой домен p53 выступает в качестве основы
Также HBx способен усиливать экспрессию бел
для привлечения транскрипционных регулято
ка HURP (hepatoma upregulated protein), кото
ров, таких как p300/CBP и MDM2, тем самым
рый вызывает деградацию p53 и подавляет апоп
изменяя экспрессию клеточных генов, необхо
тоз, вызванный химиотерапевтическими ДНК
димых для распространения вируса [27].
повреждающими агентами [22]. Таким образом,
Вирус папилломы человека (ВПЧ), один из
реактивация вируса гепатита В, которая доволь
наиболее известных онкогенных вирусов, ассо
но часто наблюдается при лечении гематологи
циированный с доброкачественной гиперплази
ческих злокачественных новообразований, хи
ей ткани, а также с аногенитальными карцино
миотерапии рака молочной железы и других
мами, способен использовать особые свойства
злокачественных опухолей, способна приводить
своих вирусных белков Е6 и Е7 для ингибирова
к снижению ответа на лечение [23].
ния активности онкосупрессора р53. Вирусный
Наряду с исследованиями вируса гепатита В,
белок Е6 путем взаимодействия с Е3 убиквитин
многочисленные работы были проведены по
лигазой E6AP способен вызывать 26S протеа
изучению онкогенности вируса гепатита С. Бы
сомную деградацию р53, что приводит к сниже
ло показано, что повышенная экспрессия бел
нию уровня р53 в инфицированных клетках.
ков вируса NS2, NS3/4A и NS5A способна вли
Недавно было показано, что вирусный белок Е7
ять на р53 зависимый путь, индуцируя его пере
влияет на функционирование транскрипцион
мещение из ядра в цитоплазматическую/пери
ного репрессорного комплекса DREAM. Путь
нуклеарную область. В частности, неструктур
p53 p21 DREAM является одним из ключевых в
ный белок NS5A напрямую взаимодействует с
активации контрольных точек клеточного цик
р53 и подавляет p53 индуцированный апоптоз
ла с помощью p53. Запуская этот путь, р53 по
[24]. Высокий уровень белка NS5A также ассо
давляет ряд генов, контролируемых комплексом
циирован с появлением активных форм кисло
DREAM. Однако онкобелок Е7 способен нап
рода (АФК) в гепатоцитах, что может вызывать
рямую связываться с комплексом DREAM и
окислительный стресс и способствовать разви
усиливать экспрессию большинства генов кле
тию гепатоцеллюлярной карциномы. Наряду с
точного цикла, подавляя при этом функции р53
неструктурными компонентами белки капсида
[28]. Таким образом, белки Е6 и Е7 ВПЧ, влияя
вируса E1 и Е2 также являются причиной силь
на уровень р53 и его сигнальный путь, вызывают
ного окислительного стресса, что ускоряет раз
изменения в клеточном цикле хозяина, что при
витие опухоли. Интересно отметить, что
водит к нерегулируемому делению клеток и по
экспрессия белка NS3 NS5B достаточна для по
давлению апоптоза (рисунок).
явления двухмембранных везикул, которые на
Неонкогенные вирусы, такие как вирус ос
поминают аутофагосомы, а белок NS4B индуци
пы, имеющие большую ДНК [29], для обеспече
рует липидизацию LC3 и накопление аутофаго
ния ее синтеза уменьшают уровень и стабиль
сом [24]. Таким образом, вирус гепатита С, по
ность р53. Было обнаружено, что киназа B1R,
давляя апоптотическую гибель, может исполь
кодируемая ранним вирусным геном, гиперфос
БИОХИМИЯ том 85 вып. 10 2020
ВИРУСЫ, АПОПТОЗ И КАНЦЕРОГЕНЕЗ
1401
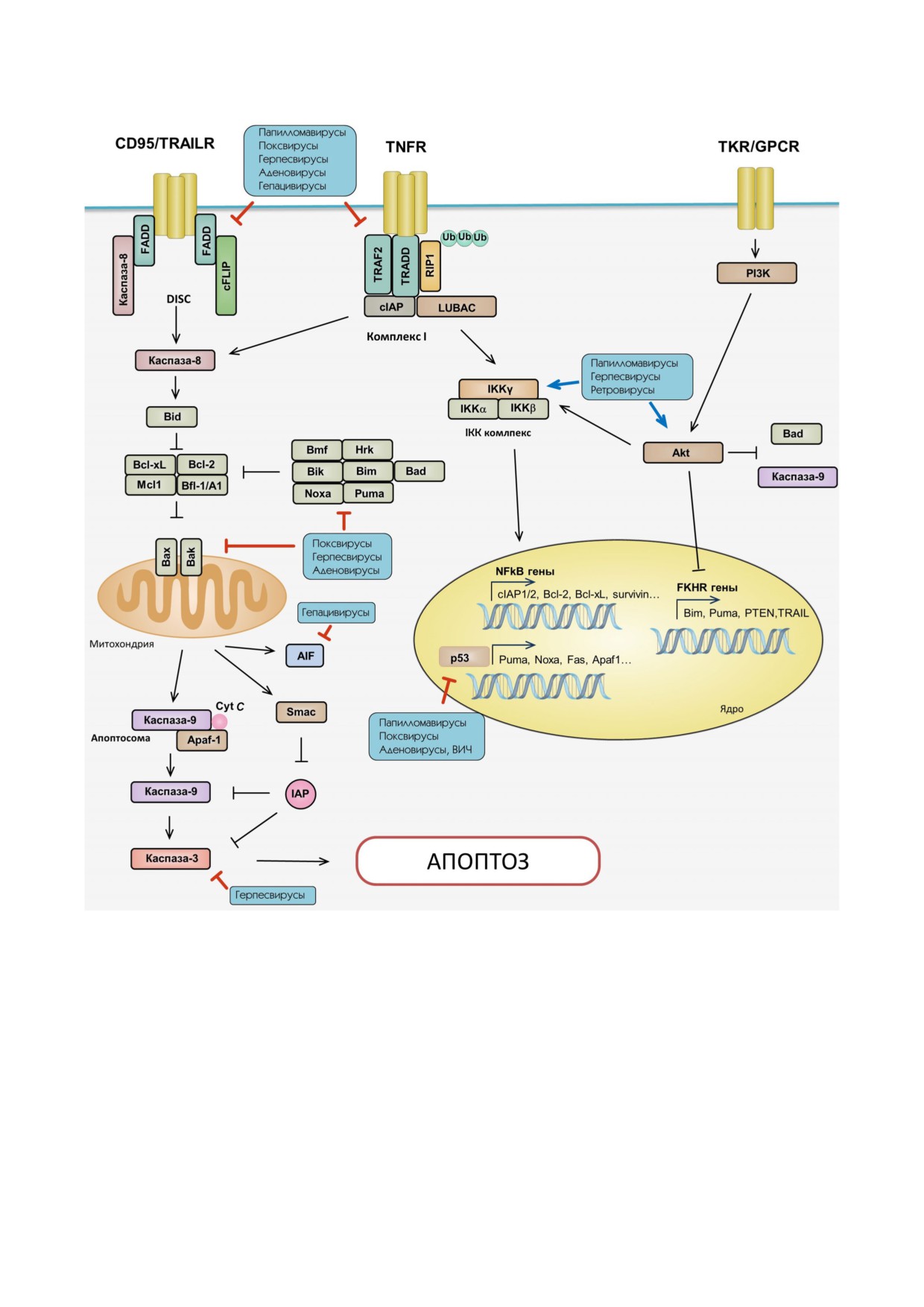
Подавление внешнего и внутреннего пути апоптоза вирусной инфекцией и модуляция клеточного сигналинга (обьясне
ния см. в тексте). Красные стрелки обозначают подавление вирусами соответствующей мишени, синие стрелки - актива
цию выбранного пути или белка. Красным курсивом отмечены гены, связанные с ингибированием апоптотической гибе
ли, синим курсивом - гены, участвующие в его запуске. (С цветным вариантом рисунка можно ознакомиться в электрон
форилирует р53 по остаткам Ser 15 и Thr 18, что
ком p53 в процессе инфицирования и в зависи
приводит к увеличению его убиквитинилирова
мости от стадии инфекции либо подавляет, либо
ния и деградации [30].
активируют его. На ранней стадии заражения
Вирус иммунодефицита человека 1 го типа
белки ВИЧ 1, такие как Nef и LTR, взаимодей
(ВИЧ 1), который вызывает снижение числа
ствуют непосредственно с р53 через его N кон
CD4 лимфоцитов и нарушение функций им
цевую часть и инактивируют этот белок, снижая
мунной системы, также взаимодействует с бел
его транскрипционную активность и как след
6 БИОХИМИЯ том 85 вып. 10 2020
1402
ЗАМАРАЕВ и др.
ствие апоптоз [31]. На более поздних стадиях
Помимо регуляции уровня рецепторов смер
Tat белки ВИЧ 1 способны индуцировать ак
ти на поверхности клетки и концентрации ли
тивность р53, стимулируя апоптоз Т клеток и
гандов, вирусы способны влиять на сборку глав
вызывая распространение ВИЧ 1 [32].
ных апоптотических комплексов внешнего пути
Недавние исследования вируса Эпш
апоптоза - DISC и комплекс I. Было показано,
тейн-Барра (ВЭБ) из семейства герпесвирусов
что многие вирусы из семейства герпесвирусов,
выявили вирусный онкоген EBNA1, который
а также поксвирусов кодируют белок vFLIP, го
влияет на путь p53 через свою мРНК. Кодируе
мологичный белку регулятору клеточной гибе
мый вирусом белок EBNA1 подавляет трансля
ли - cFLIP. Белок vFLIP содержит в своей
цию собственной мРНК, что приводит к актива
структуре два эффекторных домена смерти
ции PI3Kδ киназы и стабилизации р53 специ
(DED), благодаря которым он взаимодействует
фической убиквитинлигазы (MDM2), которая
с белком адаптером комплекса DISC - FADD,
вызывает моноубиквитинирование р53 и его
предотвращая активацию инициаторной каспа
деградацию под действием ядерных и цитоплаз
зы 8 и запуск апоптотического пути [40]. Вирус
матических протеасом [33]. Таким образом, раз
гепатита С, в свою очередь, способен регулиро
личные типы онкогенных вирусов, используя
вать уровень экспрессии эндогенного белка
собственные белки или РНК, способны влиять
cFLIP и подавлять активацию каспазы 8, тем
на эндогенный уровень p53 или ингибировать
самым смещая равновесие в сторону некропто
его транскрипционную активность, подавляя
тической гибели клеток [41, 42].
развитие апоптотического пути и стимулируя
Особо следует отметить роль ВПЧ в подавле
пролиферацию клеток хозяина.
нии TNF опосредованного апоптоза. ВПЧ ко
дирует два онкобелка, Е6 и Е7, которые непос
редственно ответственны за развитие ВПЧ ин
ИНГИБИРОВАНИЕ
дуцированного канцерогенеза. В ходе исследо
РЕЦЕПТОР-ЗАВИСИМОГО ПУТИ
ваний было показано, что онкобелок Е6 взаимо
АПОПТОТИЧЕСКОЙ ГИБЕЛИ
действует с доменом смерти TNFR1 и блокирует
взаимодействие TNFR1 с белком адаптером
Многие вирусы выработали различные стра
TRADD, подавляя запуск апоптоза [43]. Белок
тегии для вмешательства не только во внутрен
Е6 также может защищать клетки от TRAIL ин
ний клеточный путь, но и в сигнализацию всего
дуцированного апоптоза, способствуя деграда
внешнего пути апоптоза. Опосредованный ре
ции белка FADD и каспазы 8 [44]. Онкобелок
цепторами смерти апоптоз играет важную роль в
Е7 ВПЧ, в свою очередь, ингибирует TNF опос
патогенезе вирусной инфекции и противовирус
редованный апоптоз путем модуляции уровня
ной реакции хозяина. Многие вирусы приобре
E3 убиквитин лигазы сIAP2, которая участвует в
ли способность подавлять рецептор зависимый
деградации каспаз и белков комплекса
апоптоз и уклоняться от иммунного ответа хозя
DISC [45].
ина с помощью вирус кодируемых антиапопто
Были также отмечены случаи, когда вирус
тических факторов (рисунок) [34].
ные белки взаимодействовали напрямую с кас
Некоторые вирусы, такие как поксвирусы,
пазами и подавляли их активность. Представи
способны кодировать гомологичные рецепто
тели семейства герпесвирусов, вирус простого
рам смерти белки (T2 белок, CrmE), которые в
герпеса и цитомегаловирусы, кодируют белки
свободном виде способны нейтрализовать ли
ингибиторы - рибонуклеотидредуктазу R1 и
ганд TNF, необходимый для запуска внешнего
vICA соответственно. Эти белки способны по
пути апоптоза [35]. Вирусный белок гепатита В
давлять Fas опосредованный апоптоз путем свя
(HBc) выступает мощным ингибитором TRAIL
зывания с продоменом прокаспазы 8 через эф
индуцированного апоптоза в клетках гепатомы
фекторный домен смерти и предотвращать ее
человека. Устойчивость HBc экспрессирующих
активацию [46, 47]. Поксвирусы (вирус коровь
клеток к TRAIL индуцированному апоптозу
ей и кроличьей оспы, натуральной оспы и др.)
связана со значительным снижением экспрес
также кодируют консервативные ингибиторы
сии рецептора смерти 5 (DR5 или TRAILR2) и
сериновых протеаз - серпины, которые подав
наблюдалась у пациентов с хроническим гепа
ляют активность каспазы 8 и
10 при запуске
титом [36]. ВЭБ, напротив, предотвращает акти
внешнего пути апоптоза [48].
вацию TNF пути путем снижения экспрессии
Таким образом, многие вышеперечисленные
рецептора TNFR1 [37]. Следует отметить, что
онкогенные вирусы приобрели способность по
аденовирусы и вирус папилломы человека также
давлять внешний путь апоптоза и избегать им
способны подавлять экспрессию рецепторов
мунный ответ хозяина главным образом за счет
смерти Fas и TNFR1/2 [38, 39].
направленного подавления рецепторов смерти
БИОХИМИЯ том 85 вып. 10 2020
ВИРУСЫ, АПОПТОЗ И КАНЦЕРОГЕНЕЗ
1403
клеток хозяина и их лигандов, а также влияя на
инфекции vMIA, напротив, не имея гомологии с
ключевые компоненты рецепторного комплекса.
Bcl 2, по своей третичной структуре похож на
Bcl xL. vMIA способен связываться с проапоп
тотическими белками Bax и Bak, предотвращать
ИНГИБИРОВАНИЕ
их олигомеризацию и открытие митохондриаль
МИТОХОНДРИАЛЬНОГО
ных пор [58, 59].
ПУТИ АПОПТОЗА
Следует также отметить, что герпесвирус, ас
социируемый с саркомой Капоши, кодирует до
В процессе эволюции вирусов был выявлен
полнительный антиапоптотический белок, ко
еще один способ негативной регуляции апопто
торый является гомологом клеточного белка
за, а именно модуляция уровня белков семейства
survivin, члена семейства белков ингибиторов
Bcl 2. В настоящее время описано более 30 бел
апоптоза (IAP), блокирующих активность кас
ков млекопитающих этого семейства или ассо
паз и подавляющих гибель клеток. Этот белок
циированных с ним, которые условно разделены
называют K7 или вирусный ингибитор апоптоза
на анти и проапоптотические [49]. Антиапопто
(vIAP) [60]. Было показано, что vIAP связывает
тические белки семейства Bcl 2 включают Bcl 2,
ся с клеточным Bcl 2 белком, а также с активи
Bcl xL, Bcl W, Mcl 1 и Bfl 1/A1 и имеют общую
рованной каспазой 3, ингибируя ее протеолити
структурную гомологию. Эти антиапоптотичес
ческую активность и подавляя проапоптотичес
кие белки могут напрямую взаимодействовать с
кую сигнализацию в клетке [60].
проапоптотическими белками Bim, Puma, Bad,
Семейство поксвирусов, являясь крупным
Bid, Bik, Bmf, Hrk, Bax, Bak и Noxa и подавлять
ДНК вирусом, также содержит в своей структу
их активность. Активация апоптоза способна
ре ингибиторы апоптоза. Так, белок F1L вируса
смещать баланс в сторону проапоптотических
коровьей оспы, не имея сходств в своей после
белков Bcl 2 семейства, что приводит к сборке
довательности с белком Bcl 2, способен мимик
Bax Bak мультимерных пор на поверхности ми
рировать необычную топологическую укладку
тохондриальной мембраны, ее пермеабилизации
третичной структуры Bcl 2, что позволяет ему
и, как сказано выше, выходу цитохрома c и дру
связываться с такими проапоптотическими бел
гих факторов в цитозоль [50].
ками, как Bim [61] и Bak [62], предотвращая раз
Существует много крупных ДНК вирусов,
витие внутреннего пути апоптоза. Следует доба
которые имитируют белок Bcl 2 (vBcl 2), пред
вить, что многие представители семейства покс
отвращая накопление и олигомеризацию про
вирусов кодируют ингибиторы проапоптотичес
апоптотических белков Bax и Bak и запуск внут
ких BH3 содержащих белков, демонстрируют
реннего апоптотического пути. Остановка
высокую адаптивность к структуре Bcl 2 и спо
преждевременной гибели клеток хозяина на на
собность модулировать сигналинг митохонд
чальных стадиях вирусной инфекции имеет ре
рия зависимого апоптоза.
шающее значение для успешного инфицирова
Ряд штаммов семейства поксвирусов содер
ния [51] (рисунок).
жит в структуре своего генома еще один анти
Одним из первых вирусов, в котором был об
апоптотический белок - vGAAP (viral Golgi anti
наружен гомолог Bcl 2 белка, был аденовирус.
apoptotic protein). vGAAP не является необходи
Белок Е1В 19К имеет гомологичную последова
мым компонентом для репликации вируса, од
тельность BH1 и BH2 доменам Bcl 2 белка. Бы
нако влияет на его вирулентность. Интересно,
ло показано, что E1B 19K способен взаимодей
что vGAAP демонстрирует очень высокую кон
ствовать с Bax , Bak и Bik белками клетки хо
сервативность с белком человека (hGAAP), ко
зяина и функционально взаимозаменять антиа
торый локализуется в комплексе Гольджи, обра
поптотический белок Bcl 2 при аденовирусной
зуя катионный канал, и регулирует потоки ио
инфекции, выступая мощным ингибитором
нов кальция (Ca2+) в клетке [63]. Повышенная
апоптоза [52, 53].
экспрессия vGAAP или hGAAP приводила к ус
Многие представители семейства вируса гер
тойчивости клеток к апоптотической гибели,
песа, такие как ВЭБ и герпесвирус, ассоциируе
что, вероятно, обусловлено снижением выхода
мый с саркомой Капоши, также кодируют гомо
Ca2+ из внутриклеточных запасов и уменьшени
логи Bcl 2 белка: BHRF1 и Ks Bcl 2 соответ
ем проникновения ионов Ca2+ в митохондрии
ственно [54, 55]. Исследования показали, что
[63]. Тем не менее детальный механизм, с по
BHRF1 и Ks Bcl 2 белки с большей аффин
мощью которого GAAP контролируют апоптоз,
ностью способны связываться с такими проа
до сих пор неизвестен.
поптотическими белками, как Bad, Bik, Bmf,
Почти все ингибирующие апоптоз вирусные
Hrk, Noxa, Bax, подавляя их и вызывая останов
белки vBcl 2 содержат трансмембранные якор
ку апоптоза [56, 57]. Белок цитомегаловирусной
ные домены, которые необходимы для их лока
БИОХИМИЯ том 85 вып. 10 2020
6*
1404
ЗАМАРАЕВ и др.
лизации на внешней митохондриальной мемб
рус ВПЧ, в котором каждый из вирусных онко
ране. По видимому, различные белки vBcl 2 ин
белков Е5, Е6 и Е7 прямо или косвенно направ
гибируют разные стадии активации Bax и транс
лен на PI3K Akt путь и способствует выжива
локацию на внешнюю митохондриальную
нию клеток и их делению, а также прогрессии
мембрану, что позволяет с высокой степенью ва
злокачественных образований [74].
риативности регулировать внутренний путь
Мембранный белок герпесвируса ВЭБ
апоптоза [64].
LMP2A также индуцирует фосфорилирование
Некоторые вирусы, такие как вирус гепатита
Akt и активирует путь PI3K Akt, что предотвра
В и С, способны влиять на события апоптоза,
щает удаление зараженных клеток и обеспечива
происходящие после открытия митохондриаль
ет селективное преимущество для LMP2A
ных пор. Вирус гепатита С, используя неструк
экспрессирующих клеток во время развития
турные белки NS5A/B, подавляет активацию
ВЭБ ассоциированных злокачественных ново
каспазы 3, предположительно, путем ингиби
образований [75]. LMP2A Опосредованная ак
рования каспазы 9 [65]. Вирусный белок НВx
тивация пути PI3K Akt также ингибирует диф
вируса гепатита В, связываясь с апоптотическим
ференцировку эпителиальных клеток в ВЭБ ин
фактором AIF, влияет на появление высокомо
фицированных клетках, тем самым способствуя
лекулярных фрагментов ДНК и конденсацию
прогрессированию ВЭБ связанных карцином и
хроматина и предотвращает развитие апоптоза
лимфом [76]. Вирус Капоши из семейства гер
[66]. Было также отмечено, что экспрессия HBx
песвирусов способен кодировать вирусный ре
способна влиять на локализацию и активность
цептор, сопряжённый с G белком (vGPCR),
белков Drp1 и Parkin, смещая равновесие кле
приводящий к фосфорилированию Akt и инду
точной гибели в сторону митофагии [67].
цирующий саркомогенез на аллографтной моде
Тем не менее основной мишенью внутренне
ли мышей [77]. Более того повышенная актива
го пути апоптоза для многих онкогенных виру
ция Akt наблюдалась и в биоптатах саркомы Ка
сов являются представители семейства Bcl 2.
поши человека, взятых у лиц с вирусом иммуно
Кодируемые вирусом белки демонстрируют вы
дефицита [77]. В В лимфоцитах экспрессия бел
сокую степень адаптивности к структуре Bcl 2
ка вируса Капоши К1 приводила к активации
белков и способность модулировать передачу
PI3P Akt пути, ингибированию фосфатазы
сигналов через различные механизмы.
PTEN и членов семейства транскрипционных
факторов Forkhead (FKHR), которые являются
ключевыми регуляторами клеточного цикла и
СИГНАЛЬНЫЙ ПУТЬ PI3K-Akt
апоптоза. Экспрессия вирусного белка К1 спо
КАК МИШЕНЬ ВИРУСНОЙ ИНФЕКЦИИ
собствует выживанию клеток и патогенезу виру
са, предотвращая преждевременный апоптоз ви
PI3K (фосфатидилинозитол 3 киназа) и Akt
рус инфицированных клеток (рисунок).
(протеинкиназа В) играют важную роль в регу
РНК содержащий ретровирус HTLV 1, в
ляции клеточного цикла и апоптоза [68]. Спо
свою очередь, модулирует Akt в CD4+ Т клетках,
собность Akt предотвращать апоптотическую
способствуя длительной латентной фазе [79].
гибель в клеточных линиях осуществляется пу
Было обнаружено, что онкобелок HTLV 1 (Tax)
тем фосфорилирования и ингибирования про
активирует путь Akt и индуцирует Akt зависи
апоптотических медиаторов, таких как Bad, Bax
мую инактивацию транскрипционного фактора
и каспаза 9 [69, 70], а также активацией тран
FOXO3 (Forkhead box O3), который вызывает
скрипционного фактора CREB, или IkB киназы
удаление CD4+ Т клеток путем индукции про
(IKK), являющейся положительным регулято
апоптотических и антипролиферативных генов
ром NF κB, для изменения экспрессии генов с
мишеней [79]. Таким образом, ингибирование
антиапоптотической активностью [71]. Поэтому
FOXO3 способствует выживанию и пролифера
во многих опухолевых тканях сигнальный путь
ции CD4+ Т клеток, которые сохраняют способ
PI3K и Akt гиперактивен [72].
ность распространять инфекционные частицы
Некоторые онкогенные вирусы, такие как
HTLV 1 [79].
ВПЧ, семейство герпесвирусов (ВЭБ, KSHV),
Т лимфотропный ретровирус человека, также
известный как ретровирус Т клеточного лей
CИГНАЛЬНЫЙ ПУТЬ NF-κB
коза человека 1 го типа (HTLV 1), развили ме
КАК АНТОГОНИСТ
ханизмы активации этого сигнального пути для
АПОПТОТИЧЕСКОЙ ГИБЕЛИ
ингибирования апоптоза или аутофагии, кото
рая может препятствовать вирусной репликации
Активация PI3K Akt пути может приводить
[73]. Наиболее широко изученным является ви
к запуску другого нисходящего сигнального пу
БИОХИМИЯ том 85 вып. 10 2020
ВИРУСЫ, АПОПТОЗ И КАНЦЕРОГЕНЕЗ
1405
ти NF κB, который также достаточно часто ак
АФК в клетках в связи с дисфункцией митохон
тивируется во многих видах раковых клеток и
дрий и реакцией на несвернутые белки. Такой
способствует развитию опухолевого процесса
вирус индуцированный окислительный стресс
[80]. Существует несколько механизмов, благо
способен запускать не только метаболические
даря которым NF κB антагонизирует гибель
изменения, но и активацию NF κB, что способ
клеток. Во первых, активация NF κB приводит
ствует онкогенезу в тканях печени и крови [89].
к повышению уровня антиапоптотических ге
Неонкогенные вирусы, такие как вирус гепатита
нов, таких как cIAP1/2, Bcl 2, Bcl xL, TRAF1/2,
дельта, коинфицирующий клетки хозяина сов
survivin, p21, а также индуцирует экспрессию не
местно с вирусом гепатита В, усиливают продук
которых проонкогенных генов и ряда провоспа
цию АФК и активируют путь NF κB и STAT3,
лительных цитокинов [81]. Кроме того, сигна
что может привести к ускорению развития пато
лизация NF κB способствует прогрессирова
логий печени и появлению гепатоцеллюлярной
нию опухоли, облегчая переход эпителиальных
карциномы [90].
клеток в мезенхимальные и метастазирование,
NF κB опосредованное воспаление играет
способствуя васкуляризации опухолей [82].
важную роль в функционировании правильного
Активация NF κB также является частью от
врожденного иммунного ответа на острую ин
вета на острую вирусную инфекцию, но некото
фекцию, тем не менее воздействие вирусных
рые вирусы могут использовать конститутивную
белков на ключевые мишени данного пути спо
активацию NF κB для своего распространения.
собно вызвать трансформацию клеток. Актив
Например, трансмембранный белок LMP1 ВЭБ
ность NF κB не только способствует пролифе
стимулирует развитие лимфомы, активируя за
рации опухолевых клеток, подавляет апоптоз,
пуск NF κB пути [83]. LMP1 способен взаимо
но и индуцирует эпителиально мезенхималь
действовать с адаптерными молекулами TRAF,
ный переход и метастазирование [91]. Таким об
как клеточные рецепторы TNFR семейства, что
разом, подавление NF κB пути в инфицирован
обуславливает его участие в передаче внутрикле
ных клетках делает данный путь перспективной
точных сигналов. Таким образом, LMP1 инду
терапевтической мишенью.
цированная активация NF κB способствует
пролиферации и выживанию инфицированных
клеток [84].
ЗАКЛЮЧЕНИЕ И ПЕРСПЕКТИВЫ
NF κB также конститутивно активируется в
большинстве клеток первичной выпотной лим
Когда нарушаются нормальные механизмы
фомы (PEL), индуцированной вирусом Капоши
контроля роста клеток и ее гибели, некоторые
[85]. В этих клетках вирусный белок vFLIP акти
клетки могут проявлять неконтролируемую
вирует путь NF κB, связываясь непосредствен
пролиферацию и перестают выполнять свои
но с регуляторной субъединицей комплекса
тканеспецифические функции, что приводит к
IKK - NEMO (также известна как IKK gamma),
развитию рака. Считается, что заражение онко
что приводит к активации этого комплекса и
генными вирусами вызывает ~15-20% всех опу
высвобождению компонентов ДНК связываю
холевых заболеваний человека [92]. В настоящее
щих транскрипционных факторов [86]. У транс
время наиболее известны ВЭБ, вирус гепатита В
генных мышей, экспрессирующих вирусный бе
и С, Т лимфотропный вирус человека 1 (HTLV
лок vFLIP, активация NF κB пути приводит к
1), папилломавирусы человека, а также герпес
усиленной пролиферации лимфоцитов и увели
вирус, ассоциированный с саркомой Капоши
чению частоты встречаемости лимфомы [87].
(KSHV), и клеточный полиомавирус Меркеля
Ретровирус HTLV 1 способен похожим обра
(MCPyV) (таблица).
зом влиять на активацию NF κB пути. Вирус
Следует также отметить еще один тип виру
ный онкобелок Tax модулирует клеточные сиг
сов, наиболее исследуемый и актуальный в пос
нальные пути для усиления пролиферации Т
леднее время - SARS CoV 2 семейства короно
клеток и выживания. Новые исследования по
вирусов. Данный вирус принадлежит к семей
казали, что белок Tax, влияя на активацию убик
ству неретровирусных РНК вирусов с одноце
витин зависимых киназ, способен подвергаться
почечным РНК геномом. Многие исследования
К63 зависимому полиубиквитинированию. Эта
продемонстрировали способность вируса запус
модификация имеет ключевое значение для его
кать как внешний, так и внутренний путь апоп
взаимодействия с NEMO и активации NF κB
тотической гибель клеток хозяина, что облегча
(рисунок) [88].
ет распространение вируса и отягчает клиничес
Следует отметить, что некоторые онкоген
кую картину [93, 94]. Однако некоторые авторы
ные вирусы, такие как вирус гепатита В и С,
предполагают, что SARS CoV 2 может способ
ВЭБ, способны вызывать увеличение уровня
ствовать постоянной инфекции внутри хозяина,
БИОХИМИЯ том 85 вып. 10 2020
1406
ЗАМАРАЕВ и др.
фиброзу легких, что через какое то время, нап
Логичным подходом к профилактике или ле
ример, через несколько лет может привести к
чению раковых заболеваний вирусной этиоло
всплеску новообразований [95]. Один из меха
гии является направленное подавление вируса.
низмов, который был предложен авторами на
Этот принцип был подтвержден успехами в кли
основании гомологии белков с SARS CoV 1, это
нической практике, которые резко снизили ко
разрушение белка онкосупрессора р53. Другим
личество вирус ассоциированных опухолевых
механизмом, который может привести к канце
заболеваний [97]. Появление противовирусной
рогенезу, является цитокиновый шторм и окис
терапии вируса гепатита С позволило значи
лительный стресс. Последний может выступить
тельно улучшить результативность лечения по
как инициатором, так и промотором канцероге
давляющего большинства пациентов и остается
неза благодаря прямому мутагенному действию
эффективным способом профилактики гепато
АФК на ДНК, а также способствовать пролифе
целлюлярной карциномы [98]. В настоящее вре
рации и инвазии клеток. Однако эксперимен
мя применение вакцины против ВПЧ и вируса
тальные данные, подтверждающие эти гипоте
гепатита В в развитых и некоторых развиваю
зы, еще не выявлены.
щихся странах позволило существенно снизить
Онкогенные вирусы человека имеют разно
заболеваемость раком шейки матки, печени и
образные геномы, клеточные тропизмы, онко
других вирус ассоциированных заболеваний.
логические патологии, однако у них есть много
Также сейчас успешно проводятся испытания
общих свойств, которые могут привести к онко
вакцины и иммунотерапии против ВЭБ [99].
генезу.
Профилактика или лечение ВЭБ может снизить
Так, большинство онкогенных вирусов пере
частоту возникновения лимфопролифератив
даются между людьми и вызывают хронические
ных заболеваний, а также некоторых лимфом и
инфекции, которые длятся годами без явных
карциномы носоглотки [100].
симптомов. На протяжении этого времени он
Таким образом, разработка противовирус
когенные вирусы адаптируются к клетке хозяи
ных препаратов, а также иммунологическая те
на, меняя клеточные процессы и нарушая им
рапия, направленная на антигены опухоли, яв
мунное распознавание. Вирусные онкобелки
ляются приоритетными целями, которые долж
способны манипулировать как внешним путем
ны быть достигнуты в современной борьбе с он
запуска апоптоза, так и внутренним, влияя на
кологическими заболеваниями.
экспрессию рецепторов смерти, сборку апопто
тических комплексов, подавляя активность кас
Финансирование. Работа выполнена при под
паз и проапоптотических белков. Однако подав
держке гранта РНФ (проект 19 15 00125). Рабо
ление иммунной системы и неадекватный им
та в лабораториях авторов также поддержана
мунный надзор способны индуцировать бескон
грантами РФФИ (18 29 09005, 20 015 00157),
трольное размножение вируса и активную
Шведским (190345) и Стокгольмским онколо
экспрессию вирусных белков, которые наруша
гическими фондами.
ют регуляцию пролиферации клеток хозяина и
Конфликт интересов. Авторы заявляют об от
стимулируют образование опухолей [96]. Хотя
сутствии конфликта интересов.
злокачественное перерождение является объе
Соблюдение этических норм. Настоящая ра
диняющим патологическим признаком для он
бота не содержит описания выполненных авто
когенных вирусов, тем не менее он не является
рами исследований с участием людей или ис
эволюционно выгодным для вируса и необходи
пользованием животных в качестве объектов.
мым для его распространения.
СПИСОК ЛИТЕРАТУРЫ
1.
Jin, Z., and El Deiry, W. S. (2005) Overview of cell death
their potential therapeutic role, Cell. Mol. Life Sci., 72,
signaling pathways, Cancer Biol. Ther., 4, 139163,
505 517, doi: 10.1007/s00018 014 1757 2.
doi: 10.4161/cbt.4.2.1508.
5.
Raab Traub, N. (2012) Novel mechanisms of EBV
2.
Schleich, K., Warnken, U., Fricker, N., Öztürk, S.,
induced oncogenesis, Curr. Opin. Virol., 2, 453458,
Richter, P., et al. (2012) Stoichiometry of the CD95 death
doi: 10.1016/j.coviro.2012.07.001.
inducing signaling complex: experimental and modeling
6.
Young, L. S., and Rickinson, A. B. (2004) Epstein-Barr virus: 40
evidence for a death effector domain chain model, Mol.
years on, Nat. Rev. Cancer, 4, 757 768, doi: 10.1038/nrc1452.
Cell, 47, 306 319, doi: 10.1016/j.molcel.2012.05.006.
7.
Levrero, M., and Zucman Rossi, J. (2016) Mechanisms of
3.
Ashkenazi, A., and Dixit, V. M. (1998) Death receptors:
HBV induced hepatocellular carcinoma, J. Hepatol., 64,
signaling and modulation, Science, 281, 13051308,
84 101, doi: 10.1016/j.jhep.2016.02.021.
doi: 10.1126/science.281.5381.1305.
8.
Gessain, A., and Cassar, O. (2012) Epidemiological aspects
4.
Zamaraev, A. V., Kopeina, G. S., Zhivotovsky, B., and
and world distribution of HTLV 1 infection, Front.
Lavrik, I. N. (2014) Cell death controlling complexes and
Microbiol., 3, doi: 10.3389/fmicb.2012.00388.
БИОХИМИЯ том 85 вып. 10 2020
ВИРУСЫ, АПОПТОЗ И КАНЦЕРОГЕНЕЗ
1407
9.
Matsuoka, M., and Jeang, K. T. (2007) Human T cell
ciency of Simian virus 40 large T antigen induced cellular
leukaemia virus type 1 (HTLV 1) infectivity and cellular
transformation, J. Virol., 83, 10106 10118, doi: 10.1128/
transformation, Nat. Rev. Cancer,
7,
270280,
jvi.00174 09.
doi: 10.1038/nrc2111.
28.
Engeland, K. (2018) Cell cycle arrest through indirect
10.
Schiffman, M., Clifford, G., and Buonaguro, F. M. (2009)
transcriptional repression by p53: I have a DREAM, Cell
Classification of weakly carcinogenic human papillo
Death Differ., 25, 114 132, doi: 10.1038/cdd.2017.172.
mavirus types: addressing the limits of epidemiology at the
29.
Moss, B. (1990) Regulation of vaccinia virus transcription,
borderline, Infect. Agent. Cancer, 4, doi: 10.1186/1750
Annu. Rev. Biochem., 59, 661 688, doi: 10.1146/annurev.bi.
9378 4 8.
59.070190.003305.
11.
Harper, D. M., and DeMars, L. R. (2017) HPV vaccines -
30.
Santos, C. R., Vega, F. M., Blanco, S., Barcia, R., and
a review of the first decade, Gynecol. Oncol., 146, 196 204,
Lazo, P. A. (2004) The vaccinia virus B1R kinase induces
doi: 10.1016/j.ygyno.2017.04.004.
p53 downregulation by an Mdm2 dependent mechanism,
12.
Mitchell, J. K., Lemon, S. M., and McGivern, D. R.
Virology, 328, 254 265, doi: 10.1016/j.virol.2004.08.013.
(2015) How do persistent infections with hepatitis C virus
31.
Greenway, A. L., McPhee, D. A., Allen, K., Johnstone, R.,
cause liver cancer? Curr. Opin. Virol., 14, 101108,
et al. (2002) Human immunodeficiency virus type 1 Nef
doi: 10.1016/j.coviro.2015.09.003.
binds to tumor suppressor p53 and protects cells against
13.
Goossens, N., and Hoshida, Y. (2015) Hepatitis C virus
p53 mediated apoptosis, J. Virol.,
76,
26922702,
induced hepatocellular carcinoma, Clin. Mol. Hepatol., 21,
doi: 10.1128/jvi.76.6.2692 2702.2002.
105 114, doi: 10.3350/cmh.2015.21.2.105.
32.
Thakur, B. K., Chandra, A., Dittrich, T., Welte, K., and
14.
Schulz, T. F., and Cesarman, E. (2015) Kaposi Sarcoma
Chandra, P. (2012) Inhibition of SIRT1 by HIV 1 viral
associated Herpesvirus: mechanisms of oncogenesis, Curr.
protein Tat results in activation of p53 pathway, Biochem.
Opin. Virol., 14, 116 128, doi: 10.1016/j.coviro.2015.08.016.
Biophys. Res. Commun., 424, 245250, doi: 10.1016/
15.
Wendzicki, J. A., Moore, P. S., and Chang, Y. (2015) Large
j.bbrc.2012.06.084.
T and small T antigens of Merkel cell polyomavirus, Curr.
33.
Gnanasundram, S., Malbert Colas, L., Chen, S., Fusée, L.,
Opin. Virol., 11, 38 43, doi: 10.1016/j.coviro.2015.01.009.
Daskalogianni, C., et al. (2020) MDM2’s dual mRNA
16.
Liu, W., MacDonald, M., and You, J. (2016) Merkel cell
binding domains co ordinate its oncogenic and tumour
polyomavirus infection and Merkel cell carcinoma, Curr.
suppressor activities, Nucleic Acids Res., 48, 6775 6787,
Opin. Virol., 20, 20 27, doi: 10.1016/j.coviro.2016.07.011.
doi: 10.1093/nar/gkaa431.
17.
Krump, N. A., and You, J. (2018) Molecular mechanisms
34.
Benedict, C. A., Norris, P. S., and Ware, C. F. (2002) To
of viral oncogenesis in humans, Nat. Rev. Microbiol., 16,
kill or be killed: viral evasion of apoptosis, Nat. Immunol.,
684 698, doi: 10.1038/s41579 018 0064 6.
3, 1013 1018, doi: 10.1038/ni1102 1013.
18.
Vousden, K. H., and Lane, D. P. (2007) p53 in health and
35.
Reading, P. C., Khanna, A., and Smith, G. L. (2002)
disease, Nat. Rev. Mol. Cell Biol., 8, 275 283, doi: 10.1038/
Vaccinia virus CrmE encodes a soluble and cell surface
nrm2147.
tumor necrosis factor receptor that contributes to virus vir
19.
Kaminskyy, V., and Zhivotovsky, B. (2010) To kill or be
ulence, Virology, 292, 285 298, doi: 10.1006/viro.2001.
killed: how viruses interact with the cell death machinery:
1236.
symposium, J. Int. Med., 267, 473 482, doi: 10.1111/
36.
Du, J., Liang, X., Liu, Y., Qu, Z., Gao, L., et al. (2009)
j.1365 2796.2010.02222.x.
Hepatitis B virus core protein inhibits TRAIL induced
20.
Wang, X. W., Gibson, M. K., Yeh, H., Forrester, K.,
apoptosis of hepatocytes by blocking DR5 expression, Cell
Harris, C. C., et al. (1995) Abrogation of p53 induced
Death Differ., 16, 219 229, doi: 10.1038/cdd.2008.144.
apoptosis by the Hepatitis B virus X gene, Cancer Res., 55,
37.
Morrison, T. E., Mauser, A., Klingelhutz, A., and Kenney,
6012 6016, doi: 10.1385/1 59259 079 9:57.
S. C. (2004) Epstein-Barr virus immediate early protein
21.
Knoll, S., Fürst, K., Thomas, S., Baselga, S. V., Stoll, A.,
BZLF1 inhibits tumor necrosis factor alpha induced sig
Schaefer, S., and Pützer, B. M. (2011) Dissection of cell
naling and apoptosis by downregulating tumor necrosis
context dependent interactions between HBx and p53
factor receptor 1, J. Virol., 78, 544 549, doi: 10.1128/
family members in regulation of apoptosis: A role for HBV
jvi.78.1.544 549.2004.
induced HCC, Cell Cycle, 10, 3554 3565, doi: 10.4161/
38.
Benedict, C. A., Norris, P. S., Prigozy, T. I., Bodmer, J. L.,
cc.10.20.17856.
Mahr, J. A., et al. (2001) Three adenovirus E3 proteins
22.
Chao, C. C. K. (2016) Inhibition of apoptosis by onco
cooperate to evade apoptosis by tumor necrosis factor relat
genic hepatitis B virus X protein: Implications for the treat
ed apoptosis inducing ligand receptor 1 and
2, J. Biol.
ment of hepatocellular carcinoma, World J. Hepatol., 8,
Chem., 276, 3270 3278, doi: 10.1074/jbc.m008218200.
1061 1066, doi: 10.4254/wjh.v8.i25.1061.
39.
Kabsch, K., and Alonso, A.
(2002) The Human
23.
Voican, C. S., Mir, O., Loulergue, P., Dhooge, M.,
Papillomavirus type 16 E5 protein impairs TRAIL and
Brezault, C., et al. (2016) Hepatitis B virus reactivation in
FasL mediated apoptosis in HaCaT cells by different
patients with solid tumors receiving systemic anticancer
mechanisms, J. Virol., 76, 12162 12172, doi: 10.1128/jvi.
treatment, Ann. Oncol., 27, 2172 2184, doi: 10.1093/
76.23.12162 12172.2002.
annonc/mdw414.
40.
Thome, M., Schneider, P., Hofmann, K., Fickenscher, H.,
24.
Vescovo, T., Refolo, G., Vitagliano, G., Fimia, G. M., and
Meinl, E., et al. (1997) Viral FLICE inhibitory proteins
Piacentini, M. (2016) Molecular mechanisms of hepatitis
(FLIPs) prevent apoptosis induced by death receptors,
C virus induced hepatocellular carcinoma, Clin. Microbiol.
Nature, 386, 517 521, doi: 10.1038/386517a0.
Infect., 22, 853 861, doi: 10.1016/j.cmi.2016.07.019.
41.
Kim, H., and Ray, R. (2014) Evasion of TNF α mediated
25.
Levine, A. J., and Oren, M. (2009) The first 30 years of
apoptosis by hepatitis C virus, Methods Mol. Biol., 1155,
p53: Growing ever more complex, Nat. Rev. Cancer, 9,
125 132, doi: 10.1007/978 1 4939 0669 7_11.
749 758, doi: 10.1038/nrc2723.
42.
Nailwal, H., and Chan, F. K. M. (2019) Necroptosis in
26.
Liu, X., and Marmorstein, R. (2006) When viral oncopro
anti viral inflammation, Cell Death Differ., 26, 4 13,
tein meets tumor suppressor: a structural view, Genes Dev.,
doi: 10.1038/s41418 018 0172 x.
20, 2332 2337, doi: 10.1101/gad.1471706.
43.
Filippova, M., Filippov, V. A., Kagoda, M., Garnett, T.,
27.
Hermannstadter, A., Ziegler, C., Kuhl, M., Deppert, W.,
Fodor, N., and Duerksen Hughes, P. J. (2009) Complexes
and Tolstonog, G. V. (2009) Wild type p53 enhances effi
of Human Papillomavirus type 16 E6 proteins form pseu
БИОХИМИЯ том 85 вып. 10 2020
1408
ЗАМАРАЕВ и др.
do death inducing signaling complex structures during
58.
Karbowski, M., Norris, K. L., Cleland, M. M., Jeong, S. Y.,
tumor necrosis factor mediated apoptosis, J. Virol., 83,
and Youle, R. J. (2006) Role of Bax and Bak in mitochon
210 227, doi: 10.1128/jvi.01365 08.
drial morphogenesis, Nature, 443, 658 662, doi: 10.1038/
44.
Garnett, T. O., Filippova, M., and Duerksen Hughes, P. J.
nature05111.
(2006) Accelerated degradation of FADD and procaspase 8
59.
Norris, K. L., and Youle, R. J. (2008) Cytomegalovirus
in cells expressing human papilloma virus 16 E6 impairs
proteins vMIA and m38.5 link mitochondrial morphogen
TRAIL mediated apoptosis, Cell Death Differ., 13, 1915
esis to Bcl 2 family proteins, J. Virol., 82, 6232 6243,
1926, doi: 10.1038/sj.cdd.4401886.
doi: 10.1128/jvi.02710 07.
45.
Yuan, H., Fu, F., Zhuo, J., Wang, W., Nishitani, J., An, D. S.,
60.
Wang, H. W., Sharp, T. V., Koumi, A., Koentges, G., and
Chen, I. S. Y., and Liu, X. (2005) Human papillomavirus
Boshoff, C. (2002) Characterization of an anti apoptotic
type 16 E6 and E7 oncoproteins upregulate c IAP2 gene
glycoprotein encoded by Kaposi’s sarcoma associated her
expression and confer resistance to apoptosis, Oncogene,
pesvirus which resembles a spliced variant of human sur
24, 5069 5078, doi: 10.1038/sj.onc.1208691.
vivin, EMBO J., 21, 2602 2615, doi: 10.1093/emboj/
46.
Dufour, F., Sasseville, A. M. J., Chabaud, S., Massie, B.,
21.11.2602.
Siegel, R. M., and Langelier, Y. (2011) The ribonucleotide
61.
Taylor, J. M., Quilty, D., Banadyga, L., and Barry, M.
reductase R1 subunits of herpes simplex virus types 1 and 2
(2006) The vaccinia virus protein F1L interacts with Bim
protect cells against TNFα and FasL induced apoptosis
and inhibits activation of the pro apoptotic protein Bax, J.
by interacting with caspase 8, Apoptosis, 16, 256 271,
Biol. Chem.,
281,
3972839739, doi:
10.1074/jbc.
doi: 10.1007/s10495 010 0560 2.
M607465200.
47.
McCormick, A. L., Skaletskaya, A., Barry, P. A., Mocarski,
62.
Postigo, A., Cross, J. R., Downward, J., and Way, M.
E. S., and Goldmacher, V. S. (2003) Differential function
(2006) Interaction of F1L with the BH3 domain of Bak is
and expression of the viral inhibitor of caspase 8 induced
responsible for inhibiting vaccinia induced apoptosis, Cell
apoptosis (vICA) and the viral mitochondria localized
Death Differ.,
13,
16511662, doi:
10.1038/sj.cdd.
inhibitor of apoptosis (vMIA) cell death suppressors con
4401853.
served in primate and rodent cytomegaloviruses, Virology,
63.
Carrara, G., Parsons, M., Saraiva, N., and Smith, G. L.
316, 221 233, doi: 10.1016/j.virol.2003.07.003.
(2017) Golgi anti apoptotic protein: a tale of camels, cal
48.
Veyer, D. L., Carrara, G., Maluquer de Motes, C., and
cium, channels and cancer, Open Biol., 7, 170045,
Smith, G. L. (2017) Vaccinia virus evasion of regulated cell
doi: 10.1098/rsob.170045.
death, Immunol. Lett., 186, 68 80, doi: 10.1016/j.imlet.
64.
Cross, J. R., Postigo, A., Blight, K., and Downward, J.
2017.03.015.
(2008) Viral pro survival proteins block separate stages in
49.
Chipuk, J. E., Moldoveanu, T., Llambi, F., Parsons, M. J.,
Bax activation but changes in mitochondrial ultrastructure
and Green, D. R. (2010) The BCL 2 family reunion, Mol.
still occur, Cell Death Differ., 15, 997 1008, doi: 10.1038/
Cell, 37, 299 310, doi: 10.1016/j.molcel.2010.01.025.
cdd.2008.14.
50.
Danial, N. N., and Korsmeyer, S. J. (2004) Cell death:
65.
Masalova, O., Lesnova, E., Solyev, P., Zakirova, N.,
critical control points, Cell, 116, 205 219, doi: 10.1016/
Prassolov, V., et al. (2017) Modulation of cell death path
s0092 8674(04)00046 7.
ways by Hepatitis C virus proteins in Huh7.5 hepatoma
51.
Altmann, M., and Hammerschmidt, W.
(2005)
cells, Int. J. Mol. Sci.,
18,
2346, doi:
10.3390/
Epstein-Barr virus provides a new paradigm: a require
ijms18112346.
ment for the immediate inhibition of apoptosis, PLoS Biol.,
66.
Liu, H., Yuan, Y., Guo, H., Mitchelson, K., Zhang, K., et
3, 1 10, doi: 10.1371/journal.pbio.0030404.
al. (2012) Hepatitis B virus encoded X protein suppresses
52.
Han, J., Wallen, H. D., Nuñez, G., and White, E. (1998) E1B
apoptosis by inhibition of the caspase independent path
19,000 molecular weight protein interacts with and inhibits
way, J. Proteome Res., 11, 48034813, doi: 10.1021/
CED 4 dependent, FLICE mediated apoptosis, Mol. Cell.
pr2012297.
Biol., 18, 6052 6062, doi: 10.1128/mcb.18.10.6052.
67.
Kim, S. J., Khan, M., Quan, J., Till, A., Subramani, S.,
53.
Farrow, S. N., White, J. H. M., Martinou, I., Raven, T.,
and Siddiqui, A. (2013) Hepatitis B virus disrupts mito
Pun, K. T., Grinham, C. J., Martinou, J. C., and Brown, R.
chondrial dynamics: induces fission and mitophagy to
(1995) Cloning of a bcl 2 homologue by interaction with
attenuate apoptosis, PLoS Pathog., 9, 1 12, doi: 10.1371/
adenovirus E1B 19K, Nature, 374, 731 733, doi: 10.1038/
journal.ppat.1003722.
374731a0.
68.
Brazil, D. P., Yang, Z. Z., and Hemmings, B. A. (2004)
54.
Sarid, R., Sato, T., Bohenzky, R. A., Russo, J. J., and
Advances in protein kinase B signalling: AKTion on multi
Chang, Y. (1997) Kaposi’s sarcoma associated herpesvirus
ple fronts, Trends Biochem. Sci.,
29,
233242,
encodes a functional Bcl 2 homologue, Nat. Med., 3, 293
doi: 10.1016/j.tibs.2004.03.006.
298, doi: 10.1038/nm0397 293.
69.
Datta, S. R., Brunet, A., and Greenberg, M. E. (1999)
55.
Henderson, S., Huen, D., Rowe, M., Dawson, C.,
Cellular survival: a play in three akts, Genes Dev., 13, 2905
Johnson, G., and Rickinson, A. (1993) Epstein-Barr
2927, doi: 10.1101/gad.13.22.2905.
virus coded BHRF1 protein, a viral homologue of Bcl 2,
70.
Takino, J. I., Sato, T., Nagamine, K., and Hori, T. (2019)
protects human B cells from programmed cell death, Proc.
The inhibition of Bax activation induced apoptosis by
Natl. Acad. Sci. USA, 90, 84798483, doi: 10.1073/
RasGRP2 via R Ras PI3K Akt signaling pathway in the
pnas.90.18.8479.
endothelial cells, Sci. Rep., 9, 16717, doi: 10.1038/s41598
56.
Kvansakul, M., Yang, H., Fairlie, W. D., Czabotar, P. E.,
019 53419 4.
Fischer, S. F., et al. (2008) Vaccinia virus anti apoptotic
71.
Fresno Vara, J. Á., Casado, E., de Castro, J., Cejas, P.,
F1L is a novel Bcl 2 like domain swapped dimer that
Belda Iniesta, C., and González Barón, M.
(2004)
binds a highly selective subset of BH3 containing death lig
P13K/Akt signalling pathway and cancer, Cancer Treat.
ands, Cell Death Differ., 15, 1564 1571, doi: 10.1038/
Rev., 30, 193 204, doi: 10.1016/j.ctrv.2003.07.007.
cdd.2008.83.
72.
Zhao, H. F., Wang, J., Shao, W., Wu, C. P., Chen, Z. P.,
57.
Flanagan, A. M., and Letai, A. (2008) BH3 domains define
To, S. T., and Li, W. P. (2017) Recent advances in the use
selective inhibitory interactions with BHRF 1 and KSHV
of PI3K inhibitors for glioblastoma multiforme: current
BCL 2, Cell Death Differ., 15, 580 588, doi: 10.1038/
preclinical and clinical development, Mol. Cancer, 16, 100,
sj.cdd.4402292.
doi: 10.1186/s12943 017 0670 3.
БИОХИМИЯ том 85 вып. 10 2020
ВИРУСЫ, АПОПТОЗ И КАНЦЕРОГЕНЕЗ
1409
73.
Surviladze, Z., Sterk, R. T., DeHaro, S. A., and Ozbun, M. A.
the resistance to apoptosis associated with Kaposi’s sarco
(2013) Cellular entry of Human Papillomavirus type 16
ma associated Herpesvirus infection, J. Virol., 91, e01170
involves activation of the phosphatidylinositol
3
17, doi: 10.1128/jvi.01170 17.
kinase/Akt/mTOR pathway and inhibition of autophagy,
87. Chugh, P., Matta, H., Schamus, S., Zachariah, S., Kumar, A.,
J. Virol., 87, 2508 2517, doi: 10.1128/jvi.02319 12.
Richardson, J. A., Smith, A. L., and Chaudhary, P. M.
74.
Zhang, L., Wu, J., Ling, M. T., Zhao, L., and Zhao, K. N.
(2005) Constitutive NF κB activation, normal Fas
(2015) The role of the PI3K/Akt/mTOR signalling path
induced apoptosis, and increased incidence of lymphoma
way in human cancers induced by infection with human
in human herpes virus 8 K13 transgenic mice, Proc. Natl.
papillomaviruses, Mol. Cancer, 14, doi: 10.1186/s12943
Acad. Sci. USA, 102, 12885 12890, doi: 10.1073/pnas.
015 0361 x.
0408577102.
75.
Fukuda, M., and Longnecker, R. (2004) Latent membrane
88. Lavorgna, A., and Harhaj, E. W. (2014) Regulation of
protein 2A inhibits transforming growth factor 1 induced
HTLV 1 tax stability, cellular trafficking and NF κB acti
apoptosis through the phosphatidylinositol 3 kinase/Akt
vation by the ubiquitin proteasome pathway, Viruses, 6,
pathway, J. Virol., 78, 1697 1705, doi: 10.1128/jvi.78.4.
3925 3943, doi: 10.3390/v6103925.
1697 1705.2004.
89. Kgatle, M. M., Spearman, C. W., Kalla, A. A., and
76.
Scholle, F., Bendt, K. M., and Raab Traub, N. (2000)
Hairwadzi, H. N. (2017) DNA oncogenic virus induced
Epstein-Barr virus LMP2A transforms epithelial cells,
oxidative stress, genomic damage, and aberrant epigenetic
inhibits cell differentiation, and activates Akt, J. Virol., 74,
alterations, Oxid. Med. Cell. Longev.,
2017,
1 16,
10681 10689, doi: 10.1128/jvi.74.22.10681 10689.2000.
doi: 10.1155/2017/3179421.
77.
Sodhi, A., Montaner, S., Patel, V., Gómez Román, J. J.,
90. Williams, V., Brichler, S., Khan, E., Chami, M., Dény, P.,
Li, Y., Sausville, E. A., Sawait, E. T., and Gutkind, J. S.
Kremsdorf, D., and Gordien, E. (2012) Large hepatitis
(2004) Akt plays a central role in sarcomagenesis induced
delta antigen activates STAT 3 and NF κB via oxidative
by Kaposi’s sarcoma herpesvirus encoded G protein cou
stress, J. Viral Hepat., 19, 744 753, doi: 10.1111/j.1365
pled receptor, Proc. Natl. Acad. Sci. USA, 101, 4821 4826,
2893.2012.01597.x.
doi: 10.1073/pnas.0400835101.
91. Xia, Y., Shen, S., and Verma, I. M. (2014) NF κB, an
78.
Tomlinson, C. C., and Damania, B. (2004) The K1 protein
active player in human cancers, Cancer Immunol. Res., 2,
of Kaposi’s sarcoma associated Herpesvirus activates the
823 830, doi: 10.1158/2326 6066.cir 14 0112.
Akt signaling pathway, J. Virol.,
78,
19181927,
92. Zur Hausen, H., and de Villiers, E. M. (2014) Cancer
doi: 10.1128/jvi.78.4.1918 1927.2004.
“causation” by infections - Individual contributions and
79.
Olagnier, D., Sze, A., Bel Hadj, S., Chiang, C., Steel, C.,
synergistic networks, Semin. Oncol.,
41,
860875,
Han, X., Routy, J. P., Lin, R., Hiscott, J., and van
doi: 10.1053/j.seminoncol.2014.10.003.
Grevenynghe, J. (2014) HTLV 1 Tax mediated inhibition
93. Ren, Y., Shu, T., Wu, D., Mu, J., Wang, C., et al. (2020)
of FOXO3a activity is critical for the persistence of termi
The ORF3a protein of SARS CoV 2 induces apoptosis in
nally differentiated CD4+ T cells, PLoS Pathog., 10,
cells, Cell. Mol. Immunol., 17, 881 883, doi: 10.1038/
e1004575, doi: 10.1371/journal.ppat.1004575.
s41423 020 0485 9.
80.
Bai, D., Ueno, L., and Vogt, P. K. (2009) Akt mediated
94. Varga, Z., Flammer, A. J., Steiger, P., Haberecker, M.,
regulation of NF κB and the essentialness of NF κB for
Andermatt, R., et al. (2020) Endothelial cell infection and
the oncogenicity of PI3K and Akt, Int. J. Cancer, 125,
endotheliitis in COVID 19, Lancet, 395, 14171418,
2863 2870, doi: 10.1002/ijc.24748.
doi: 10.1016/S0140 6736(20)30937 5.
81.
Feng, C., Wu, B., Fan, H., Li, C., and Meng, S. (2014)
95. Alpalhão, M., Ferreira, J. A., and Filipe, P.
(2020)
NF kappaB induced gp96 up regulation promotes hepa
Persistent SARS CoV 2 infection and the risk for cancer,
tocyte growth, cell cycle progression and transition, Acta
Med. Hypotheses, 143, 109882, doi: 10.1016/j.mehy.
Microbiol. Sinica, 54, 1212 1220.
2020.109882.
82.
Huber, M. A., Azoitei, N., Baumann, B., Grünert, S.,
96. Mesri, E. A., Feitelson, M. A., and Munger, K. (2014)
Sommer, A., et al. (2004) NF κB is essential for epithelial
Human viral oncogenesis: a cancer hallmarks analysis, Cell
mesenchymal transition and metastasis in a model of breast
Host Microbe, 15, 266 282, doi: 10.1016/j.chom.2014.
cancer progression, J. Clin. Invest.,
114,
569581,
02.011.
doi: 10.1172/jci21358.
97. Van Kriekinge, G., Castellsagué, X., Cibula, D., and
83.
Kulwichit, W., Edwards, R. H., Davenport, E. M., Baskar,
Demarteau, N. (2014) Estimation of the potential overall
J. F., Godfrey, V., and Raab Traub, N. (1998) Expression
impact of human papillomavirus vaccination on cervical
of the Epstein-Barr virus latent membrane protein
1
cancer cases and deaths, Vaccine,
32,
733739,
induces B cell lymphoma in transgenic mice, Proc. Natl.
doi: 10.1016/j.vaccine.2013.11.049.
Acad. Sci. USA, 95, 11963 11968, doi: 10.1073/pnas.95.20.
98. McQuaid, T., Savini, C., and Seyedkazemi, S. (2015)
11963.
Sofosbuvir, a significant paradigm change in HCV treat
84.
Wang, L. W., Jiang, S., and Gewurz, B. E.
(2017)
ment, J. Clin. Transl. Hepatol., 3, 27 35, doi: 10.14218/
Epstein-Barr virus LMP1 mediated oncogenicity, J.
jcth.2014.00041.
Virol., 91, e01718 16, doi: 10.1128/jvi.01718 16.
99. Schiller, J. T., and Lowy, D. R. (2010) Vaccines to prevent
85.
Gopalakrishnan, R., Matta, H., and Chaudhary, P. M.
infections by oncoviruses, Annu. Rev. Microbiol., 64, 23 41,
(2013) A purine scaffold HSP90 inhibitor BIIB021 has
doi: 10.1146/annurev.micro.112408.134019.
selective activity against KSHV associated primary effu
100. Bu, W., Joyce, M. G., Nguyen, H., Banh, D. V., Aguilar, F.,
sion lymphoma and blocks vFLIP k13 induced NF κB,
et al. (2019) Immunization with components of the viral
Clin. Cancer Res., 19, 5016 5026, doi: 10.1158/1078
fusion apparatus elicits antibodies that neutralize
0432.ccr 12 3510.
Epstein-Barr Virus in B cells and epithelial cells,
86.
Briggs, L. C., Chan, A. W. E., Davis, C. A., Whitelock, N.,
Immunity, 50, 1305 1316, doi: 10.1016/j.immuni.2019.
Hotiana, H. A., et al. (2017) IKKγ mimetic peptides block
03.010.
БИОХИМИЯ том 85 вып. 10 2020
1410
ЗАМАРАЕВ и др.
VIRAL INFECTIONS: NEGATIVE REGULATOR
OF APOPTOSIS AND FACTOR OF ONCOGENICITY
Review
A. V. Zamaraev1, B. Zhivotovsky1,2, and G. S. Kopeina1*
1 Lomonosov Moscow State University, Faculty of Basic Medicine, 119191 Moscow, Russia; E*mail: lirroster@gmail.com
2 Institute of Environmental Medicine, Karolinska Institute, SE*171 77 Stockholm, Sweden
Received June 30, 2020
Revised September 9, 2020
Accepted September 9, 2020
The disruption of apoptotic cell death process is closely associated with the etiology of various diseases, including can
cer. Permanent viral infections can cause different types of cancers. Oncogenic viruses manipulate both external and
internal apoptosis pathways, and inhibit the activity of proapoptotic proteins and signaling pathways, which facilitates
carcinogenesis. Ineffective immune surveillance or immune response suppression can induce uncontrolled virus
propagation and host cell proliferation. In this review, we discuss current data that provide insights into mechanisms
of apoptotic death suppression by viruses and their role in oncogenesis.
Keywords: oncogenic viruses, apoptosis, carcinogenesis
БИОХИМИЯ том 85 вып. 10 2020