БИОХИМИЯ, 2020, том 85, вып. 12, с. 1849 - 1857
УДК 577.121.7; 577.23
НЕФОСФОРИЛИРУЮЩЕЕ ОКИСЛЕНИЕ В МИТОХОНДРИЯХ
И СОПРЯЖЕННЫЕ С НИМ ПРОЦЕССЫ
Обзор
© 2020
Д.Б. Зоров1,2*, Н.В. Андрианова1, В.А. Бабенко1,2, Л.Е. Бакеева1,
С.Д. Зоров3, Л.Д. Зорова1,2, И.Б. Певзнер1,2, В.А. Попков1,2,
Е.Ю. Плотников1,2,4, Д.Н. Силачев1,2
1 НИИ физико химической биологии имени А.Н. Белозерского, Московский государственный университет
имени М.В. Ломоносова, 119991 Москва, Россия; электронная почта: zorov@belozersky.msu.ru
2 НМИЦ акушерства, гинекологии и перинатологии имени акад. В.И. Кулакова, 117997 Москва, Россия
3 Московский государственный университет имени М.В. Ломоносова,
факультет биоинженерии и биоинформатики, 119991 Москва, Россия
4 Институт молекулярной медицины, Первый Московский государственный
медицинский университет имени И.М. Сеченова, 119991 Москва, Россия
Поступила в редакцию 28.07.2020
После доработки 17.09.2020
Принята к публикации 17.09.2020
Механизмы окислительного фосфорилирования и его регуляции остаются одними из основных проблем
биоэнергетики. Эффективность энергизации митохондрий определяется взаимоотношением скорости ге"
нерации электрохимического потенциала ионов водорода и скоростями его расходования на синтез АТР и
использования АТР в эндергонических реакциях. Разобщение (частичное или полное), возникающее в про"
цессе неконтролируемой и контролируемой утечки ионов через внутреннюю митохондриальную мембрану,
с одной стороны, приводит к уменьшению относительного синтеза АТР, а с другой, подчиняясь закону сох"
ранения энергии, приводит к образованию тепла, генерация которого является насущной функцией орга"
низма. Кроме повышенного термогенеза, увеличение нефосфорилирующего окисления различных субстра"
тов сопровождается уменьшением трансмембранного потенциала и продукции активных форм кислорода и
активацией потребления кислорода, продукции воды и двуокиси углерода, повышением уровня внутрикле"
точного АDР и закислением цитозоля. В этом анализе каждый их этих факторов будет рассмотрен в отдель"
ности на предмет его роли в регуляции метаболизма.
КЛЮЧЕВЫЕ СЛОВА: митохондрии, вода, термогенез, набухание, биоэнергетика, разобщение.
DOI: 10.31857/S032097252012009X
ВВЕДЕНИЕ
ческую энергию макроэргической связи в моле"
куле АТР. Это означает, что при высокой степени
Окислительное фосфорилирование в совре"
сопряжения окисления с фосфорилированием
менной трактовке означает сопряжение окисле"
(то есть при максимально возможном превраще"
ния субстратов, сопровождающееся генерацией
нии энергии, освобождаемой при переносе
электрохимического потенциала ионов водоро"
электронов по дыхательной цепи митохондрий,
да, с его использованием АТР синтазой. Это по"
в энергию, запасенную в АТР) образуется много
нятие сопряжения априори предполагает, что
АТР (максимально - три молекулы при окисле"
химическая энергия, содержащаяся в субстратах
нии одной молекулы NАDН), а при низкой -
окисления, сначала трансформируется в элект"
меньше трех молекул. Практика показывает, что
рохимическую, а потом снова в химическую, со"
в любом случае, по крайней мере, ~1/3 энергии
держащуюся в макроэргической связи АТР. Сте"
окисления не сопряжено с синтезом АТР, а ос"
пень сопряжения между этими процессами оп"
вобождается в виде тепла [1]. При буквальном
ределяет коэффициент полезного действия ин"
рассмотрении в результате метаболизма субстра"
тегрального процесса, то есть насколько эффек"
тов окисления образуется АТР, вода, СО2 и выде"
тивно энергия окисления превращается в хими"
ляется тепло. Мы кратко рассмотрим в отдель"
ности все эти компоненты и некоторые способы
* Адресат для корреспонденции.
их регуляции, учитывая сопряженные элементы.
1849
1850
ЗОРОВ и др.
ТЕРМОГЕНЕЗ
тами. В целом, проблема соответствия доставки
энергии в отделы ткани и энергетических запро"
Считать тепло, освобождаемое в результате
сов ткани является одной из крайне важных в
митохондриального окисления, ненужной и
физиологическом отношении проблем, реше"
бессмысленной потерей неправильно, так как
ние которых очень значимо [6]. Возможно, ана"
это выделяемое тепло является основным спо"
логичное решение о необходимости высокой
собом поддержания температуры прежде всего у
васкуляризации природа нашла и для щитовид"
гомойотермных животных, к которым принад"
ной железы, производящей тиреоидные гормо"
лежит и человек. Более того, выделение тепла в
ны. Эти гормоны (с определенными оговорка"
результате митохондриального окисления надо
ми) некоторыми исследователями признаются
рассматривать не как императивную паразит"
как природные слабые разобщители, хотя пря"
ную утечку, а как тонко регулируемый физиоло"
мое разобщающее действие тиреоидных гормо"
гически необходимый процесс, который затра"
нов реализуется далеко не всегда, что указывает
гивает механизм сопряжения окислительного
на то, что если и происходит разобщение, то у
фосфорилирования.
этого процесса есть посредники [7-9].
Эта лишь внешне тривиальная мысль была
Разобщение окислительного фосфорилиро"
впервые высказана В.П. Скулачевым в 1960 г. на
вания, особенно в мягком режиме, допускаю"
основании измерений коэффициента Р/О мито"
щим умеренный синтез АТР митохондриями,
хондрий скелетных мышц голубя, подвергнутого
стало предметом множества исследований и бы"
охлаждению [2], и данные были представлены на
ло подробно проанализировано В.П. Скулаче"
5"ом Международном конгрессе биохимиков в
вым [10]. Основная мысль, высказанная им и
Москве [3]. В частности, было показано, что при
поддержанная мировым научным сообществом,
окислении пирувата с малатом коэффициент
заключается в том, что мягкое разобщение явля"
Р/О в митохондриях контрольных птиц был 2,07,
ется механизмом уменьшения ненужного и из"
а у птиц, подверженных охлаждению, 1,0. Надо
быточного эндогенного производства активных
отметить, что эти изменения сопровождались
форм кислорода. Этот вывод был сделан на ос"
активацией дыхания, т.е. скорость окисления
новании полученной экспоненциальной зави"
субстратов возрастала. Конечно же, возник воп"
симости генерации активных форм кислорода
рос о механизме такой регуляции. Немного поз"
(АФК), прежде всего перекиси водорода, от ве"
же первым кандидатом на роль такого регулято"
личин трансмембранного потенциала митохон"
ра, с полным правом претендующего называться
дрий, когда максимальная генерация АФК наб"
природным разобщителем окислительного фос"
людалась у гиперпотенциальных митохондрий
форилирования, стали свободные жирные кис"
[11]. Из этого не напрямую следует вывод, что
лоты [4], и именно они на долгие годы стали
генерация АФК митохондриями происходит не
предметом исследований команды Скулачева.
в результате некой неконтролируемой утечки
Надо понимать, что активация дыхания кле"
электронов с компонентов дыхательной цепи, а
ток в пределах органа может приводить к воз"
является естественным, тонко регулируемым
можному дисбалансу между доставкой и потреб"
процессом, в ходе которого митохондрии, по"
лением кислорода и субстратов, что теоретичес"
мимо выполнения целого ряда насущных функ"
ки может приводить к ишемизации органа, яв"
ций [12], превращаются в систему, стабилизиру"
ляющейся нежелательной, поэтому максималь"
ющую уровень внутриклеточных АФК, тем са"
ная потеря сопряжения может нести в себе мно"
мым превращая ее в некотором роде в АФК"
жество патогенетических событий, заключаю"
стат. В этом плане митохондриальный термоге"
щихся не только в снижении рО2 в ткани, но и
нез является мощным регулятором уровня АФК
изменении в ней уровней АТР и АDР и внутри"
в митохондрии и клетке в целом. В частности,
клеточного рН, что будет рассмотрено далее.
повышение термогенеза позволит избежать не"
Исходя из этого, физиологически допустимой
желательную лавинную генерацию АФК
является умеренная потеря сопряжения, что
[13-15], имеющую колоссальное патогенети"
нашло отражение в применяемом сейчас терми"
ческое значение [16].
не «мягкое разобщение». Видимо, именно необ"
Несколько лет назад было выяснено, что
ходимостью предотвращения ишемизации и
осуществляемый жирными кислотами и други"
обусловлена высокая степень снабжения ткани
ми слабыми и сильными разобщителями цикли"
сосудами (васкуляризация) ткани бурого жира,
ческий процесс транспорта через бислойные
специализирующегося предпочтительно на тер"
мембраны протонированной и непротониро"
могенной функции за счет наличия в ткани ра"
ванной форм [17] является не единственным ме"
зобщающего белка UCP1 [5], обеспечивающего
ханизмом, обеспечивающим сброс мембранно"
термогенез, опосредованный жирными кисло"
го потенциала на внутренней митохондриаль"
БИОХИМИЯ том 85 вып. 12 2020
НЕФОСФОРИЛИРУЮЩЕЕ ОКИСЛЕНИЕ В МИТОХОНДРИЯХ
1851
ной мембране. Дополнительно к этому разоб"
конечной причиной гибели пациентов, пере"
щение окислительного фосфорилирования дос"
несших инсульт [31].
тигается наличием белков - переносчиков ра"
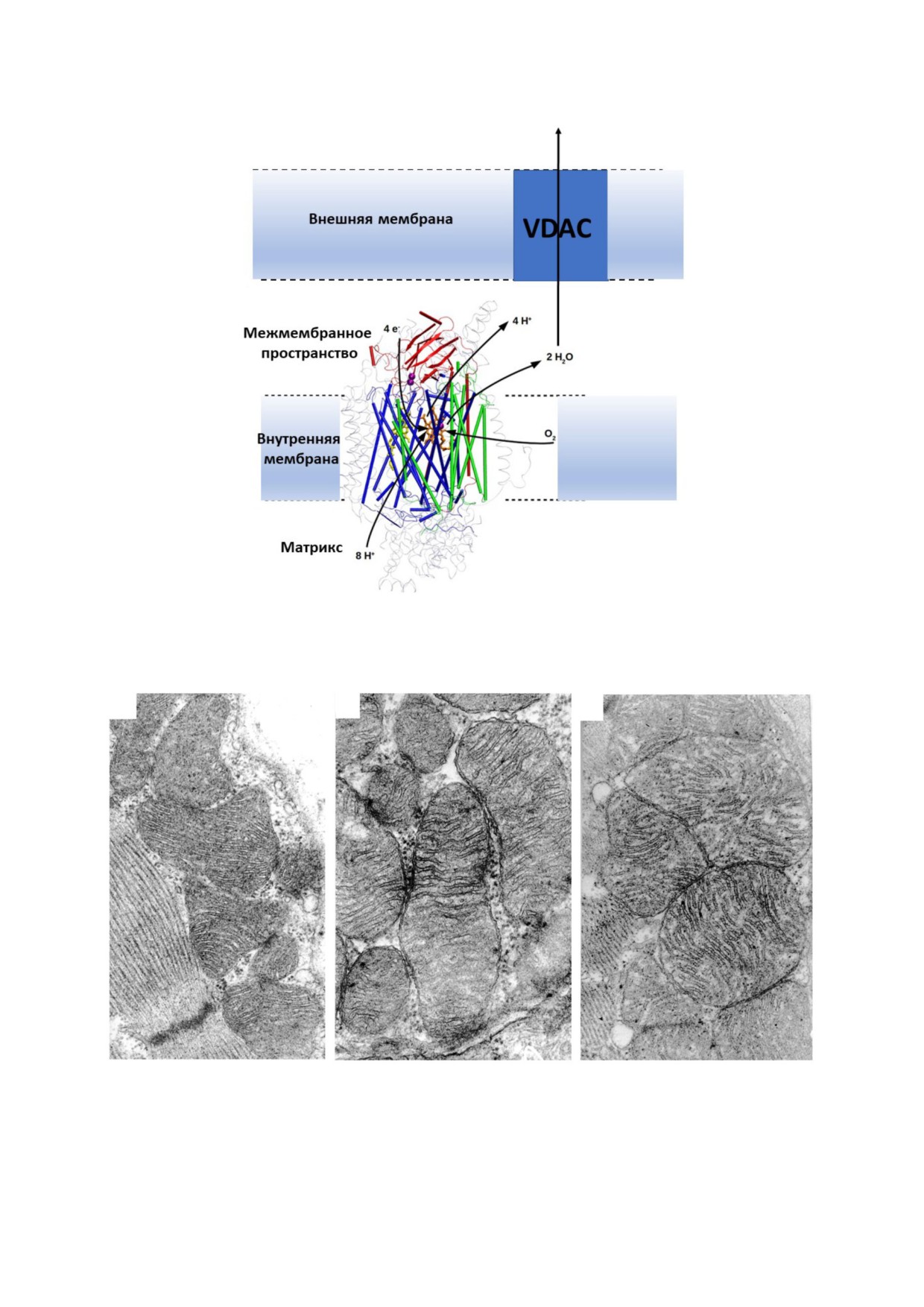
В митохондриях основное место образова"
зобщителей, среди которых не только UCP1, но
ния воды локализовано на цитохромокси"
и АDР/АТР антипортер [18], и дикарбоксилат"
дазе, освобождающей молекулы воды в межме"
ный переносчик во внутренней митохондриаль"
мбранное пространство (рис. 1) [32]. Дальней"
ной мембране [19, 20]. При всем этом в предпо"
шая судьба воды распределяется по двум нап"
лагаемом разобщении, опосредованном жирны"
равлениям. По одному сценарию практически
ми кислотами, остается много недоказанных
вся вода уходит в цитоплазму. Хотя признается,
предположений, но в общем виде можно гово"
что мембраны являются неплохими проводни"
рить о возможной естественной протонофорной
ками для воды, наиболее легким путем выхода
активности жирных кислот, наблюдаемой на
воды из митохондрий является ее вынос через
бислойных моделях и опосредованной протоно"
вольт"зависимый анионный канал (VDAС или,
форной активностью, вызванной взаимодей"
как его еще называют, пориновый канал [33]).
ствием жирных кислот с белками внутренней
Что касается второй опции, то в принципе вода
мембраны митохондрий, при этом важным фак"
может поступать в матрикс митохондрий, где
тором, участвующим в эффективности разобще"
особенно высокое осмотическое + онкотичес"
ния, является значение трансмембранного по"
кое давление, и в норме ортодоксальная конфи"
тенциала [21]. Примечательным стало подтвер"
гурация матрикса митохондрий есть один из по"
ждение транспорта протона через транслокатор
казателей митохондриального гомеостаза. В
адениновых нуклеотидов, опосредованного
этом плане наличие во внутренней митохондри"
жирными кислотами, очень схожего по характе"
альной мембране аквапоринов [34, 35] внешне
ру с механизмом разобщающего действия жир"
выглядит нелогичным, ибо их присутствие уско"
ных кислот при взаимодействии с разобщаю"
рит поступление воды в матрикс митохондрий.
щим белком UCP1, в результате чего подтверж"
Хотя, справедливости ради, надо отметить, что
дается также и возможная термогенная функция
быстрое уравновешивание объема митохондрий
АDР/АТР антипортера [22]. Есть данные, что
после осмотического воздействия [36] может
сами митохондрии достаточно устойчивы к зна"
свидетельствовать о том, что пассивного транс"
чительному повышению температуры [23], так
порта воды и без наличия аквапоринов вполне
что калоригенное действие митохондриальных
может хватить для обеспечения осмотической
переносчиков может не отражаться на функцио"
регуляции, и специфическая функция аквапо"
нальной активности митохондрий. Пока остает"
ринов не сводится к транспорту воды, а заклю"
ся загадочным процесс ингибирования альфа"
чается в переносе некоторых других веществ,
кетохолестанолом действия даже самых мощ"
осуществляя, например, транспорт ионов аммо"
ных разобщителей [24], что может предполагать
ния, сопряженный с транспортом воды, что
участие липидов в процессе разобщения.
предполагается для аквапорина 8 [37].
Отметим, что набухание межмембранного
пространства и высокоамплитудное набухание
ВОДА
митохондрий в целом, наблюдаемые на ультра"
структурном уровне, являются признаками па"
Подавляющее большинство исследователей
тогенного стрессорного воздействия. При этом
отчетливо признает кислород"потребляющую
небольшое набухание митохондрий является не
функцию митохондрий, однако результирую"
только не патологическим, а наоборот, регуля"
щее образование воды в митохондриях уходит из
торным, приводящим к активации митохондри"
общего внимания. Надо понимать, что на одну
ального метаболизма, как адаптивный ответ на
молекулу потребленного кислорода образуется
умеренный стресс. В кардиомиоцитах вызван"
две молекулы воды. Но при этом, если проблеме
ное окислительным стрессом увеличение объе"
доставки кислорода в ткань и в саму митохонд"
ма митохондрий всего на несколько процентов
рию посвящено громадное число публикаций,
приводит к активации дыхания на одну треть с
то создается впечатление, что проблемы оттока
пропорциональным увеличением синтеза АТР,
воды от митохондрий и целой клетки не сущест"
генерации активных форм кислорода (АФК),
вует. При этом в патологической клетке можно
обеспечивающих мобилизацию активных за"
наблюдать локальное или глобальное (высокоа"
щитных сигнальных систем [38] и приводящих к
мплитудное) набухание митохондрий (т.е. на
соответствию энергетических поступлений и их
микроуровне [25-28]) или отек всей клетки или
затрат [6]. Это небольшое увеличение объема
органа (т.е. на макроуровне) [29, 30]. Заметим,
митохондрий наблюдалось при действии гормо"
что именно фатальный отек мозга становится
нов, в частности, глюкагона, также обеспечива"
БИОХИМИЯ том 85 вып. 12 2020
1852
ЗОРОВ и др.
Рис. 1. Функционирование цитохромоксидазы с предполагаемым транспортом воды. (С цветным вариантом рисунка
а
б
в
Рис. 2. Ультраструктура митохондрий в диафрагме крысы. Электронная микроскопия, выполненная после стандартной
фиксации и подготовки образца. Просмотр осуществляли на микроскопе Hitachi HU"12. а - Диафрагма контрольной
крысы, содержащейся при 28°С; б - диафрагма крысы, подвергнутой холодовому стрессу (2°С, 15 мин); в препарат диа"
фрагмы контрольной крысы после добавления 40 μМ 2,4"динитрофенола in vitro при 28°C
БИОХИМИЯ том 85 вып. 12 2020
НЕФОСФОРИЛИРУЮЩЕЕ ОКИСЛЕНИЕ В МИТОХОНДРИЯХ
1853
ющего активацию метаболизма [39]. В начале
белками перемещение митохондрий является
1970"х гг. мы наблюдали активацию тканевого
строго энергозависимым, требующим АТР (воз"
дыхания, сопровождающуюся небольшим уве"
можно и/или мембранного потенциала [43]), в
личением in situ межмембранного пространства
то время как предполагаемый water"jet механизм
и просветлением матрикса митохондрии диаф"
в нефосфорилирующих условиях (при деэнерги"
рагмальной ткани крыс, подвергнутых холодо"
зации митохондрий, соответствующей состоя"
вому стрессу (рис. 2). Таким образом, набухание
нию 3u или близкому к нему) за счет активации
митохондрий также подчиняется общей законо"
дыхания, сопровождающейся усиленной генера"
мерности «multet nocem» (много вредно), доми"
цией и выходом из митохондрий воды, будет бо"
нирующей в биологическом мире [15] и которой
лее активным, чем в покоящемся состоянии
нужно придерживаться при использовании фар"
(состояние 4 или близкое к нему). Мы предпола"
макологических препаратов. При этом неболь"
гаем, что если такой механизм митохондриаль"
шое набухание митохондрий является не только
ной подвижности существует, то по определе"
физиологически допустимым, но и необходи"
нию будет прямая связь между уменьшением
мым для мобилизации метаболизма и его соот"
степени сопряжения митохондрий и повышени"
ветствия возросшим потребностям, в то время
ем скорости образования Н2О цитохромоксида"
как дальнейшее набухание митохондрий может
зой и, соответственно, созданием реактивного
приводить к гибели клетки. Можно лишь пред"
импульса, способного обеспечить перемещение
положить, что это малое изменение объема ми"
митохондрий.
тохондрий не сопровождается разрывом их
внешней мембраны, в то время как большее на"
бухание, вплоть до т.н. высокоамплитудного,
АТР
приводит к увеличивающимся разрывам внеш"
ней мембраны, которое сопровождается выхо"
Мы отдаем себе отчет, что глобальное обсуж"
дом из межмембранного пространства сигналь"
дение роли АТР в клетке просто невозможно в
ных белков, индуцирующих клеточную гибель.
рамках такого обзора, поэтому позволяем себе
Отметим, что высокоамплитудное набухание
сосредоточиться лишь на тех аспектах, которые
митохондрий, наблюдаемое при открытии в ми"
мало обсуждаются в научном мире.
тохондриях мегаканала (индукции неспецифи"
Чем выше вклад нефосфорилирующего
ческой проницаемости), сопровождается, с од"
окисления в интегральное потребление кисло"
ной стороны, полной деэнергизацией, а с дру"
рода митохондриями, тем больший эффект это
гой стороны, неспособностью обеспечить нор"
произведет на уровни внутриклеточного АТР,
мальную (ортодоксальную) конформацию мат"
даже при сохранении энергетических затрат, не
рикса митохондрий, определенно указывающей
говоря уже о физиологических ситуациях, кото"
на взаимосвязь этих двух процессов/состояний.
рые сопряжены с активацией метаболизма. Но
Рассматривая первый вариант судьбы воды,
прежде всего нужно понимать, что изменения
образованной в митохондриях, а именно ее вы"
уровня АТР неотвратимо вызовут изменения в
ход через VDAC, мы позволим себе предполо"
содержании его партнеров по простой схеме
жить, что этот огромный массив воды будет спо"
гидролиза"синтеза, которая очень схематично
собен генерировать реактивный импульс, доста"
выглядит как АТР = АDР + Рi + Н+, т.е. сущест"
точный для обеспечения перемещения отдель"
вует полное сопряжение освобождения и связы"
ных митохондрий в клетках с мало структуриро"
вания в системе протона с уровнем АТР или
ванной цитоплазмой (например, в фибробластах
АDР (здесь мы опускаем роль ионов Mg, считая
или астроглиальных клетках). Неравномерное
хелирующие способности АТР и АDР близки"
распределение VDAC по поверхности внешней
ми). В любом клеточном компартменте увеличе"
митохондриальной мембраны было продемон"
ние содержания АТР сопутствует некоторому
стрировано раньше [40], и оно теоретически мо"
защелачиванию компартмента, а его уменьше"
жет обеспечивать реактивное движение целых
ние сопровождается закислением. Именно это
органелл или их частей, которое мы наблюдали.
свойство и определяет возникновение ацидоза в
Такое реактивное движение может быть рас"
ткани, испытывающей проблемы с доставкой
смотрено как еще один механизм подвижности
энергических субстратов и кислорода (напри"
митохондрий, дополняющий уже известный ме"
мер, при ишемии), приводящей к падению
ханизм перемещения митохондрий, идущий с
уровня АТР в ишемизированной области. По"
участием адаптерных моторных белков, ассоци"
нятно, что внутриклеточные буферы в значи"
ированных с цитоскелетом [41, 42]. Эти типы
тельной степени гасят кислотный «пожар», од"
движений, различающиеся по механизмам, пре"
нако и у этих буферов есть пределы, в результате
дусматривают, что опосредованное моторными
чего в ткани возникают градиенты кислотности,
БИОХИМИЯ том 85 вып. 12 2020
1854
ЗОРОВ и др.
при этом в областях с низкими значениями
тельного фосфорилирования, поток через цикл
внутриклеточных рН начинают активироваться
Кребса пропорциональным образом увеличива"
деградационные процессы [44-46]. Повторим,
ется, генерация СО2 также возрастает, внося до"
что именно АТРазная активность, необходимая
полнительный вклад в общее закисление.
для выполнения внутриклеточных функций,
Казалось бы, умеренное закисление обладает
определяет степень ацидоза, а не результат пе"
защитными свойствами, что было показано на
реключения на гликолитический путь, непра"
примере ишемии [55], в частности за счет акти"
вильно названный «лактатным ацидозом», ибо
вации связывания белкового ингибитора (IF1) c
путь превращения глюкозы в молочную кислоту
АТР синтазным комплексом [56], что препят"
не сопряжен с суммарной генерацией протона
ствует нежелательное в условиях энергетическо"
(см. подробное объяснение в [47]).
го кризиса расходование АТР на поддержание
Кроме этого, надо понимать, что повышение
мембранного потенциала митохондрий в усло"
концентрации АDР, сопряженное с гидролизом
виях гипоксии/ишемии [43, 57]. Однако было
АТР, может привести к смещению равновесия
обнаружено, что простым эффектом на измене"
аденилаткиназной реакции в направлении по"
ние рН роль СО2 не ограничивается. Стало изве"
вышения уровня АМР, имеющего очень важное
стным, что возрастающие уровни двуокиси угле"
значение, в частности, через регуляцию АМР"
рода и сопряженного с ней бикарбоната, напри"
киназной активности [48].
мер в условиях реперфузии после ишемии, нап"
Другой момент, хотя и мало связанный с
рямую действуют на процесс аутофагии, ингиби"
энергетикой, состоит в том, что АТР, при нахож"
руя его [58] и, тем самым, препятствуют устране"
дении его вне клетки, может рассматриваться
нию нефункциональных митохондрий, что чре"
как индуктор нативного воспалительного про"
вато дальнейшим повреждением биологической
цесса из"за принадлежности АТР к группе
системы [59]. Но, кроме роли в рН"регуляции,
DAMPs (damage"associated molecular patterns)
СО2 и бикарбонат несут значительную сигналь"
[49], и его выход из клетки возможен, в частнос"
ную функцию, в частности, участвуя в ряде важ"
ти, при массивном или локальном повреждении
ных редокс"реакций [60-62], при этом возраста"
ткани или как результат некротической гибели
ющий уровень бикарбоната провоцирует окис"
одиночных клеток. Кроме АТР, мощными вне"
ление белков [63]. Очень слабая проницаемость
клеточными регуляторными функциями обла"
мембран для бикарбоната нивелируется наличи"
дает аденозин, образующийся из АМР [50]. На"
ем специфических переносчиков для него
до отметить возможную связь между внутри" и
[64-66], дефект которых приводит к патологиям,
внеклеточными уровнями адениновых нуклео"
включая системный ацидоз, дисфункцию мозга
тидов, учитывая наличие в клеточной мембране
и почек и гипертонию (суммировано в [63]).
паннексинов [51, 52].
В пределах физиологических значений рН
СО2 в чистом виде почти не существует, в основ"
ном превращаясь в бикарбонат, который регу"
ДВУОКИСЬ УГЛЕРОДА
лирует множество ферментов. Среди них адени"
латциклаза [67], сукцинатдегидрогеназа [68],
Одним из основных рН"буферов в организме
митохондриальная АТРаза [69, 70] и АТР синта"
является химически равновесная система бикар"
за [71], что позволяет отнести СО2/бикарбонат"
бонат/двуокись углерода (HCO3-/CO2) [53, 54],
ную систему к достаточно мощному фактору,
отсюда ясна важная роль стационарного уровня
регулирующему окислительное фосфорилиро"
СО2 и ее продукции в кислотном гомеостазе в
вание. Недавняя находка сопрягающего
митохондрии, цитоплазме, внеклеточном прост"
действия бикарбоната [72] подтверждает эту
ранстве и крови. По определению, увеличение
точку зрения, указывая на еще одну точку уп"
уровня СО2 приведет к ацидозу, а его уменьше"
равления механизмом разобщения окислитель"
ние - к алкалозу в биологическом компартмен"
ного фосфорилирования.
те. В митохондриях СО2 образуется в цикле три"
карбоновых кислот в двух последовательных ре"
акциях из изоцитрата (с катализом на изоцитрат"
Финансирование. Работа выполнена при фи"
дегидрогеназе) и альфа"кетоглутарата (с катали"
нансовой поддержке Российского научного
зом на альфа"кетоглутаратдегидрогеназе), при
фонда (грант 19"14"00173).
этом, чем более активно функционирует цикл
Конфликт интересов. Авторы заявляют об от"
Кребса, тем больше образуется СО2 и тем больше
сутствии конфликта интересов.
вероятность закисления матрикса митохондрий.
Соблюдение этических норм. В данной работе
Учитывая, что при активации дыхания, в част"
нет исследований, в которых были бы исполь"
ности, при уменьшении сопряжения окисли"
зованы в качестве объектов люди или животные.
БИОХИМИЯ том 85 вып. 12 2020
НЕФОСФОРИЛИРУЮЩЕЕ ОКИСЛЕНИЕ В МИТОХОНДРИЯХ
1855
СПИСОК ЛИТЕРАТУРЫ
1.
Brand, M. D., Chien, L. F., Ainscow, E. K., Rolfe, D. F.,
18.
Andreyev, A. Yu., Bondareva, T. O., Dedukhova, V. I.,
and Porter, R. K. (1994) The causes and functions of mito"
Mokhova, E. N., Skulachev, V. P., Tsofina, L. M., Volkov,
chondrial proton leak, Biochim. Biophys. Acta, 1187, 132"
N. I., and Vygodina, T. V. (1989) The ATP/ADP"
139, doi: 10.1016/0005"2728(94)90099"x.
antiporter is involved in the uncoupling effect of fatty acids
2.
Скулачев В. П., Маслов С. П. (1960) Роль окисления в
on mitochondria, Eur. J. Biochem.,
182,
585"592,
терморегуляции, Биохимия, 25, 1058"1064.
doi: 10.1111/j.1432"1033.1989.tb14867.x.
3.
Skulachev, V. P. (1961) Regulation of the coupling of oxi"
19.
Wieckowski, M. R., and Wojtczak, L. (1997) Involvement
dation and phosphorylation, 5th Internat. Cong. of Biochem.
of the dicarboxylate carrier in the protonophoric action of
(Moscow), 5, 367"373.
long"chain fatty acids in mitochondria, Biochem. Biophys.
4.
Скулачев В. П., Маслов С. П., Сивкова В. Г. Калини"
Res. Commun., 232, 414"417, doi: 10.1006/bbrc.1997.6298.
ченко Л. П., Маслова Г. М. (1963) Холодовое разобще"
20.
Samartsev, V. N., Smirnov, A. V., Zeldi, I. P., Markova, O. V.,
ние окисления и фосфорилирования в мышцах белых
Mokhova, E. N., and Skulachev, V. P. (1997) Involvement
мышей, Биохимия, 28, 70"79.
of aspartate/glutamate antiporter in fatty acid"induced
5.
Nedergaard, J, and Cannon, B. (2018) Brown adipose tis"
uncoupling of liver mitochondria, Biochim. Biophys. Acta,
sue as a heat"producing thermoeffector, Handb. Clin.
1319, 251"257, doi: 10.1016/s0005"2728(96)00166"1.
Neurol., 156, 137"152, doi: 10.1016/B978"0"444"63912"
21.
Rupprecht, A., Sokolenko, E. A., Beck, V., Ninnemann, O.,
7.00009"6.
Jaburek, M., Trimbuch, T., Klishin, S. S., Jezek, P.,
6.
Yaniv, Y., Juhaszova, M., Nuss, H. B., Wang, S., Zorov, D. B.,
Skulachev, V. P., and Pohl, E. E. (2010) Role of the trans"
Lakatta, E. G., and Sollott, S. J. (2010) Matching ATP
membrane potential in the membrane protein leak,
supply and demand in mammalian heart: in vivo, in vitro,
Biophys. J., 98, 1503"1511, doi: 10.1016/j.bpj.2009.12.4301.
and in silico perspectives, Ann. N. Y. Acad. Sci., 1188, 133"
22.
Bertholet, A. M., Chouchani, E. T., Kazak, L.,
142, doi: 10.1111/j.1749"6632.2009.05093.x.
Angelin, A., Fedorenko, A., et al. (2019) H+ transport is an
7.
Maley, G. F., and Lardi, H. A. (1953) Metabolic effects of
integral function of the mitochondrial ADP/ATP carrier,
thyroid hormones in vitro. II. Influence of thyroxine and
Nature, 571, 515"520, doi: 10.1038/s41586"019"1400"3.
triiodothyronine on oxidative phosphorylation, J. Biol.
23.
Chrétien, D., Bénit, P., Ha, H. H., Keipert, S., El"Khoury, R.,
Chem., 204, 435"444, PMID: 13084614.
et al. (2018) Mitochondria are physiologically maintained
8.
Hoch, F. L., and Lipmann, F. (1954) Proc. Natl. Acad. Sci.
at close to 50°C, PLoS Biol., 16, e2003992, doi: 10.1371/
USA, 40, 909"921, doi: 10.1073/pnas.40.10.909.
journal.pbio.2003992.
9.
Dickens, F., and Salmony, D. (1956) Effects of thyroid
24.
Starkov, A. A., Dedukhova, V. I., and Skulachev, V. P.
hormones in vitro on tissue respiration, oxidative phospho"
(1994) 6"Ketocholestanol abolishes the effect of the most
rylation and the swelling of mitochondria, Biochem. J., 64,
potent uncouplers of oxidative phosphorylation in mito"
645"651, doi: 10.1042/bj0640645.
chondria, FEBS Lett., 355, 305"308, doi: 10.1016/0014"
10.
Skulachev, V. P. (1998) Uncoupling: new approaches to an
5793(94)01211"3.
old problem of bioenergetics, Biochim. Biophys. Acta, 1363,
25.
Lehninger, A. L. (1962) Water uptake and extrusion by
100"124, doi: 10.1016/s0005"2728(97)00091"1.
mitochondria in relation to oxidative phosphorylation,
11.
Korshunov, S. S., Skulachev, V. P., and Starkov, A. A. (1997)
Physiol. Rev., 42, 467"517, doi: 10.1152/physrev.1962.42.
High protonic potential actuates a mechanism of produc"
3.467.
tion of reactive oxygen species in mitochondria, FEBS Lett.,
26.
Solenski, N. J., diPierro, C. G., Trimmer, P. A., Kwan, A. L.,
416, 15"18, doi: 10.1016/s0014"5793(97)01159"9.
and Helm, G. A. (2002) Ultrastructural changes of neu"
12.
Zorov, D. B., Krasnikov, B. F., Kuzminova, A. E.,
ronal mitochondria after transient and permanent cerebral
Vysokikh, M. Yu., and Zorova, L. D. (1997) Mitochondria
ischemia, Stroke, 33, 816"824, doi: 10.1161/hs0302.
revisited. Alternative functions of mitochondria, Biosci.
104541.
Rep., 17, 507"520, doi: 10.1023/a:1027304122259.
27.
Kaasik, A., Kuum, M., Joubert, F., Wilding, J., Ventura"
13.
Zorov, D. B., Filburn, C. R., Klotz, L. O., Zweier, J. L.,
Clapier, R., and Veksler, V. (2010) Mitochondria as a
and Sollott, S. J. (2000) Reactive oxygen species (ROS)"
source of mechanical signals in cardiomyocytes,
induced ROS release: a new phenomenon accompanying
Cardiovasc. Res., 87, 83"91, doi: 10.1093/cvr/cvq039.
induction of the mitochondrial permeability transition in
28.
Zorov, D. B., Vorobjev, I. A., Popkov. V. A., Babenko, V. A.,
cardiac myocytes, J. Exp. Med.,
192,
1001"1014,
Zorova, L. D., et al. (2019) Lessons from the discovery of
doi: 10.1084/jem.192.7.1001.
mitochondrial fragmentation (fission): a review and
14.
Zorov, D. B., Juhaszova, M, and Sollott, S. J. (2006)
update, Cells, 8, 175, doi: 10.3390/cells8020175.
Mitochondrial ROS"induced ROS release: an update and
29.
Ware, A. J., D’Agostino, A. N., and Combes, B. (1971)
review, Biochim. Biophys. Acta,
1757,
509"517,
Cerebral edema: a major complication of massive hepatic
doi: 10.1016/j.bbabio.2006.04.029.
necrosis, Gastroenterology, 61, 877"884, PMID: 125688.
15.
Zorov, D. B., Juhaszova, M., and Sollott, S. J. (2014)
30.
Durward, Q. J., Del Maestro, R. F., Amacher, A. L., and
Mitochondrial reactive oxygen species (ROS) and ROS"
Farrar, J. K. (1983) The influence of systemic arterial pres"
induced ROS release, Physiol. Rev.,
94,
909"950,
sure and intracranial pressure on the development of cere"
doi: 10.1152/physrev.00026.2013.
bral vasogenic edema, J. Neurosurg.,
59,
803"809,
16.
Зоров Д. Б., Банникова С. Ю., Белоусов В. В., Высо"
doi: 10.3171/jns.1983.59.5.0803.
ких М. Ю., Зорова Л. Д., Исаев Н. К., Красников
31.
Preston, E., and Webster J. (2004) A two"hour window for
Б. Ф., Плотников Е. Ю. (2005) Активные формы кис"
hypothermic modulation of early events that impact
лорода и азота: друзья или враги, Биохимия, 70, 215"
delayed opening of the rat blood"brain barrier after
221.
ischemia, Acta Neuropathol., 108, 406"412, doi: 10.1007/
17.
Liberman, E. A., Topaly, V. P., Tsofina, L. M., Jasaitis, A. A.,
s00401"004"0905"4.
and Skulachev, V. P. (1969) Mechanism of coupling of
32.
Schmidt, B., McCracken, J., and Ferguson"Miller, S.
oxidative phosphorylation and the membrane potential of
(2003) A discrete water exit pathway in the membrane pro"
mitochondria, Nature, 222, 1076"1078, doi: 10.1038/
tein cytochrome c oxidase, Proc. Natl. Acad. Sci. USA, 100,
2221076a0.
15539"15542, doi: 10.1073/pnas.2633243100.
БИОХИМИЯ том 85 вып. 12 2020
1856
ЗОРОВ и др.
33.
Zimmerberg, J., and Parsegian, V. A. (1987) Water move"
50.
Borea, P. A., Gessi, S., Merighi, S., Vincenzi, F., and
ment during channel opening and closing, J. Bioenerg.
Varani, K. (2015) Pharmacology of adenosine receptors:
Biomembr., 19, 351"358, doi: 10.1007/BF00768538.
the state of the art, Physiol. Rev., 98, 1591"1625,
34.
Gena, P., Fanelli, E., Brenner, C., Svelto, M., and
doi: 10.1152/physrev.00049.2017.
Calamita, G. (2009) News and views on mitochondrial
51.
Kang, J., Kang, N., Lovatt, D., Torres, A., Zhao, Z., Lin, J.,
water transport, Front. Biosci. (Landmark Ed), 14, 4189"
and Nedergaard, M. (2008) Connexin 43 hemichannels are
4419, doi: 10.2741/3522.
permeable to ATP, J. Neurosci.,
28,
4702"4711,
35.
Lee, W. K., and Thévenod, F. (2006) A role for mitochon"
doi: 10.1523/JNEUROSCI.5048"07.2008.
drial aquaporins in cellular life"and"death decisions?
52.
Chekeni, F. B., Elliott, M. R., Sandilos, J. K., Walk, S. F.,
Am. J. Physiol. Cell Physiol., 291, 195"202, doi: 10.1152/
Kinchen, J. M., et al. (2010) Pannexin 1 channels mediate
ajpcell.00641.2005.
“find"me” signal release and membrane permeability dur"
36.
Calamita, G., Gena, P., Meleleo, D., Ferri, D., and Svelto,
ing apoptosis, Nature, 467, 863"867, doi: 10.1038/
M. (2006) Water permeability of rat liver mitochondria: a
nature09413.
biophysical study, Biochim. Biophys. Acta, 1758, 1018"
53.
Roos, A., and Boron, W. F. (1981) Intracellular pH,
1024, doi: 10.1016/j.bbamem.2006.07.008.
Physiol. Rev.,
61,
296"434, doi:
10.1152/phys"
37.
Saparov, S. M., Liu, K., Agre, P., and Pohl, P. (2007)
rev.1981.61.2.296.
In vitro analysis and modification of aquaporin pore selec"
54.
Aickin, C. C. (1986) Intracellular pH regulation by verte"
tivity, J. Biol. Chem., 282, 5296"5301, doi: 10.1074/jbc.
brate muscle, Annu. Rev. Physiol.,
48,
349"361,
M609343200.
doi: 10.1146/annurev.ph.48.030186.002025.
38.
Juhaszova, M., Zorov, D. B., Kim, S. H., Pepe, S., Fu, Q.,
55.
Inserte, J., Barba, I., Hernando, V., Abellán, A., Ruiz
et al. (2004) Glycogen synthase kinase"3beta mediates
Meana, M., Rodr guez Sinovas, A., and Garcia"Dorado, D.
convergence of protection signaling to inhibit the mito"
(2008) Effect of acidic reperfusion on prolongation of
chondrial permeability transition pore, J. Clin. Invest., 113,
intracellular acidosis and myocardial salvage, Cardiovasc.
1535"1549, doi: 10.1172/JCI19906.
Res., 77, 782"790, doi: 10.1093/cvr/cvm082.
39.
Halestrap, A. P. (1989) The regulation of the matrix volume of
56.
Pullman, M. E., and Monroe, G. C. (1963) A naturally
mammalian mitochondria in vivo and in vitro and its role in
occurring inhibitor of mitochondrial adenosine triphos"
the control of mitochondrial metabolism, Biochim. Biophys.
phatase, J. Biol. Chem., 238, 3762"3769, PMID: 14109217.
Acta, 973, 355"382, doi: 10.1016/s0005"2728(89)80378"0.
57.
Di Lisa, F., Blank, P. S., Colonna, R., Gambassi, G.,
40.
Konstantinova, S. A., Mannella, C. A., Skulachev, V. P.,
Silverman, H. S., Stern, M. D., and Hansford, R. G.
and Zorov, D. B. (1995) Immunoelectron microscopic
(1995) Mitochondrial membrane potential in single living
study of the distribution of porin on outer membranes of rat
adult rat cardiac myocytes exposed to anoxia or metabolic
heart mitochondria, J. Bioenerg. Biomembr., 27, 93"99,
inhibition, J. Physiol., 486, 1"13, doi: 10.1113/jphysiol.
doi: 10.1007/BF02110336.
1995.sp020786.
41.
Maeder, C. I., Shen, K., and Hoogenraad, C. C., (2014)
58.
Queliconi, B. B., Kowaltowski, A. J., and Gottlieb, R. A.
Axon and dendritic trafficking, Curr. Opin. Neurobiol.,
(2016) Bicarbonate increases ischemia"reperfusion dam"
2014, 27, 165"170, doi: 10.1016/j.conb.2014.03.015.
age by inhibiting mitophagy, PLoS One, 11, e0167678,
42.
Melkov, A., and Abdu, U. (2018) Regulation of long"dis"
doi: 10.1371/journal.pone.0167678.
tance transport of mitochondria along microtubules, Cell.
59.
Zorov, D. B., Popkov, V. A., Zorova, L. D., Vorobjev, I. A.,
Mol. Life Sci., 75, 163"176, doi: 10.1007/s00018"017"
Pevzner, I. B., et al. (2017) Mitochondrial aging: is there a
2590"1.
mitochondrial clock? J. Gerontol. A Biol. Sci. Med. Sci., 72,
43.
Zorova, L. D., Popkov, V. A., Plotnikov, E. Y., Silachev, D.
1171"1179, doi: 10.1093/gerona/glw184.
N., Pevzner, I. B., et al. (2018) Mitochondrial membrane
60.
Liochev, S. I., and Fridovich, I. (2002) Copper, zinc super"
potential, Anal. Biochem., 552, 50"59, doi: 10.1016/
oxide dismutase and H2O2. Effects of bicarbonate on inac"
j.ab.2017.07.009.
tivation and oxidations of NADPH and urate, and on con"
44.
Rizack, M. A. (1964) An epinephrine"sensitive lipolytic
sumption of H2O2, J. Biol. Chem., 277, 34674"34678,
activity in adipose tissue, J. Biol. Chem., 236, 657"662,
doi: 10.1074/jbc.M204726200.
PMID: 14169136.
61.
Medinas, D. B., Cerchiaro, G., Trindade, D. F., and
45.
Eastman, A. (1994) Deoxyribonuclease II in apoptosis and
Augusto, O. (2007) The carbonate radical and related oxi"
the significance of intracellular acidification, Cell Death
dants derived from bicarbonate buffer, IUBMB Life, 59,
Differ., 1, 7"9, PMID: 17180001
255"262, doi: 10.1080/15216540701230511.
46.
Gottlieb, R. A., Giesing, H. A., Zhu, J. Y., Engler, R. L.,
62.
Queiroz, R. F., Paviani, V., Coelho, F. R., Marques, E. F.,
and Babior, B. M. (1995) Cell acidification in apoptosis:
Di Mascio, P., and Augusto, O. (2013) The carbonylation
granulocyte colony"stimulating factor delays programmed
and covalent dimerization of human superoxide dismutase
cell death in neutrophils by up"regulating the vacuolar
1 caused by its bicarbonate"dependent peroxidase activity
H(+)"ATPase, Proc. Natl. Acad. Sci. USA, 92, 5965"5968,
is inhibited by the radical scavenger tempol, Biochem. J.,
doi: 10.1073/pnas.92.13.5965.
455, 37"46, doi: 10.1042/BJ20130180.
47.
Silachev, D. N., Gulyaev, M. V., Zorova, L. D., Khailova,
63.
Queliconi, B. B., Marazzi, T. B., Vaz, S. M., Brookes, P. S.,
L. S., Gubsky, L. V., et al. (2015) Magnetic resonance spec"
Nehrke, K., Augusto, O., and Kowaltowski, A. J. (2013)
troscopy of the ischemic brain under lithium treatment.
Bicarbonate modulates oxidative and functional damage in
Link to mitochondrial disorders under stroke, Chem. Biol.
ischemia"reperfusion, Free Radic. Biol. Med., 55, 46"53,
Interact., 237, 175"182, doi: 10.1016/j.cbi.2015.06.012.
doi: 10.1016/j.freeradbiomed.2012.11.007.
48.
Steinberg, G. R., and Kemp, B. E. (2009) AMPK in health
64.
Kumar, S., Flacke, J., Kostin, S., Appukuttan, A., Reusch,
and disease, Physiol. Rev., 89, 1025"1078, doi: 10.1152/
H. P., and Ladilov, Y. (2011) SLC4A7 sodium bicarbonate
physrev.00011.2008.
cotransporter controls mitochondrial apoptosis in
49.
Zhang, Q., Raoof, M., Chen, Y., Sumi, Y., Sursal, T.,
ischaemic coronary endothelial cells, Cardiovasc. Res., 89,
Junger, W., Brohi, K., Itagaki, K., and Hauser, C. J. (2010)
392"400, doi: 10.1093/cvr/cvq330.
Circulating mitochondrial DAMPs cause inflammatory
65.
Alka, K., and Casey, J. R. (2014) Bicarbonate transport in
responses tom injury, Nature, 464, 104"107, doi: 10.1038/
health and disease, IUBMB Life,
66,
596"615,
nature08780.
doi: 10.1002/iub.1315.
БИОХИМИЯ том 85 вып. 12 2020
НЕФОСФОРИЛИРУЮЩЕЕ ОКИСЛЕНИЕ В МИТОХОНДРИЯХ
1857
66.
Nozik"Grayck, E., Huang, Y."C. T., Carraway, M. S., and
the binding change mechanism, J. Bioenerg. Biomembr.,
Piantadosi, C. (2003) Bicarbonate"dependent superoxide
16, 407"419, doi: 10.1007/BF00743235.
release and pulmonary artery tone, Am. J. Physiol. Heart
70. Roveri, O. A., and Calcaterra, N. B. (1985) Steady"state
Circ. Physiol., 285, 2327"2335, doi: 10.1152/ajpheart.
kinetic of F1"ATPase. Mechanism of anion activation,
00507.2003.
FEBS Lett.,
192
123"127, doi:
10.1016/0014"
67.
Acin"Perez, R. Salazar, E., Kamenetsky, M., Buck, J.,
5793(85)80056"9.
Levin, L. R., and Manfredi, G. (2009) Cyclic AMP pro"
71. Lodeyro, A. F. Calcaterra, N. B., and Roveri, O. A. (2001)
duced inside mitochondria regulates oxidative phosphory"
Inhibition of steady"state mitochondrial ATP synthesis by
lation, Cell Metabol., 9, 265"276, doi: 10.1016/j.cmet.
bicarbonate, an activating anion of ATP hydrolysis,
2009.01.012.
Biochim. Biophys. Acta, 1506, 236"243, doi: 10.1016/
68.
Zeylemaker, W. P., Klaasse, A. D. Slater, E. C., and
s0005"2728(01)00221"3.
Veeger, C. (1970) Studies on succinate dehydrogenase. VI.
72. Khailova, L. S., Vygodina, T. V., Lomakina, G. Y., Kotova.
Inhibition by monocarboxylic acids, Biochim. Biophys.
E. A., and Antonenko, Y. N. (2020) Bicarbonate suppress"
Acta, 198, 415"422, doi: 10.1016/0005"2744(70)90120"8.
es mitochondrial membrane depolarization induced by
69.
Kasho, V. N., and Boyer, P. D. (1984) Relationships of ino"
conventional uncouplers, Biochem. Biophys. Res. Commun.,
sine triphosphate and bicarbonate effects on F1"ATPase to
530, 29"34, doi: 10.1016/j.bbrc.2020.06.131.
NONPHOSPHORYLATING OXIDATION IN MITOCHONDRIA
AND RELATED PROCESSES
Review
D. B. Zorov1,2*, N. V. Andrianova1, V. A. Babenko1,2, L.E. Bakeeva1, S. D. Zorov3,
L. D. Zorova1,2, I. B. Pevzner1,2, V. A. Popkov1,2, E. Y. Plotnikov1,2,4, and D. N. Silachev1,2
1 Belozersky Institute of Physico Chemical Biology, Lomonosov Moscow State University,
119991 Moscow, Russia; E mail: zorov@belozersky.msu.ru
2 Kulakov National Medical Research Center of Obstetrics, Gynecology and Perinatology, 117997 Moscow, Russia
3 Lomonosov Moscow State University, Faculty of Bioengineering and Bioinformatics, 119991 Moscow, Russia
4 Sechenov First Moscow State Medical University, Institute of Molecular Medicine, 119991 Moscow, Russia
Received July 28, 2020
Revised September 17, 2020
Accepted September 17, 2020
The mechanism of oxidative phosphorylation and its regulation remain one of the main problems of bioenergetics.
Efficiency of the mitochondrial energization is determined by the relationship between the rate of generation of elec"
trochemical potential of hydrogen ions and the rate of its expenditure on the synthesis of ATP and the use of ATP in
endergonic reactions. Uncoupling (partial or complete), which occurs in the process of uncontrolled and controlled
leakage of ions through the inner mitochondrial membrane, on the one hand leads to the decrease in the relative syn"
thesis of ATP, and on the other, being consistent with the law of conservation of energy, leads to the formation of heat,
generation of which is an essential function of the organism. In addition to increased thermogenesis, the increase of
non"phosphorylating oxidation of various substrates is accompanied by the decrease in transmembrane potential, pro"
duction of reactive oxygen species, and activation of oxygen consumption, water and carbon dioxide production,
increase in the level of intracellular ADP and acidification of the cytosol. In this review, each of these factors will be
considered separately for its role in regulating metabolism.
Keywords: mitochondria, water, thermogenesis, swelling, bioenergetics, uncoupling
9 БИОХИМИЯ том 85 вып. 12 2020