БИОХИМИЯ, 2020, том 85, вып. 12, с. 1939 - 1960
УДК 577.3;577.27
ОТ МЕТАБОЛИЗМА К ИММУНИТЕТУ: АФК И ДРУГИЕ СИГНАЛЫ
Обзор
© 2020
А.Ю. Андреев1*, Ю.Е. Кушнарева2, Н.Н. Старкова3, А.А. Старков4
1 The Scripps Research Institute, 10550 La Jolla, CA 92037, USA; E mail: alex_andreyev@mitoexperts.com
2 La Jolla Institute for Allergy and Immunology, 9420 La Jolla, CA 92037, USA
3 State University of New York, Maritime College, New York, NY 10465, USA
4 Brain and Mind Research Institute, Weill Medical College of Cornell University,
Weill Medical College of Cornell University, New York, NY 10065, USA
Поступила в редакцию 11.09.2020
После доработки 13.11.2020
Принята к публикации 17.11.2020
Функционирование иммунных клеток в значительной степени определяется их метаболизмом. Исследова
ние метаболизма иммунной системы по определению является мультидисциплинарным разделом иммуно
логии, который интегрирует данные о механизмах передачи энергии и биохимических путях. Одним из на
иболее важных понятий в области иммунологии является метаболическое переключение - вызванный ак
тивацией переход иммунных клеток к преимущественному использованию специфических катаболических
путей для получения энергии. Митохондрии участвуют в этом процессе и способствуют метаболической
адаптации через активацию различных механизмов, включая изменения продукции АТР в соответствии с
динамически меняющимися метаболическими потребностями, а также служат центрами управления для
внутриклеточной передачи сигналов. Последняя функция включает в себя один из наиболее изучаемых ми
тохондриальных процессов - генерацию активных форм кислорода (АФК). Роль митохондриальных АФК в
окислительном стрессе хорошо известна, в то время как их участие в иммунном ответе представляет собой
активно развивающееся направление. В этом обзоре мы обсуждаем роль сигнальных АФК и иммунометабо
лизма с точки зрения биоэнергетики. Мы также предлагаем критический взгляд на методологию оценки
АФК, описывая современные проблемы в этой области. В заключение, на основе анализа литературных
данных, мы полагаем что, в противоположность окислительному стрессу, продукция АФК с целью клеточ
ной регуляции в большей степени контролируется биогенезом митохондрий, чем метаболическим переклю
чением.
КЛЮЧЕВЫЕ СЛОВА: иммунометаболизм, активные формы кислорода, гликолитическое переключение,
макрофаги, Т клетки, биогенез митохондрий.
DOI: 10.31857/S0320972520120167
ВВЕДЕНИЕ
дриях (митоАФК), что подчеркивает важность
прооксидантов в исследованиях живых систем.
В последние десятилетия было опубликова
В 2002 г. мы предложили простую модель кине
но множество работ, посвященных исследова
тики генерации АФК как бимолекулярную ре
нию активных форм кислорода (АФК) в митохон
акцию между кислородом и митохондриальны
Принятые сокращения: АФК - активные формы кислорода; митоАФК - АФК, продуцируемые в митохондриях;
MPTP - пора, обуславливающая неспецифическую пермеабилизацию митохондрий (mitochondrial permeability transition
pore); ОЭТ - обратный транспорт электронов; СДГ - сукцинатдегидрогеназа; ЦТК - цикл трикарбоновых кислот;
ЭТЦ - электрон транспортная цепь; ARE - AU обогащенная последовательность DNA; ATF4 - транскрипционный
фактор ATF4, зависимый от циклического АМР; BMDM - макрофаги из костного мозга; CAD - декарбоксилаза цис ако
нитата; c Myc - протоонкоген MYC; DAMP - молекулярный паттерн, связанный с повреждениями; ERK - внеклеточ
ные сигнал регулируемые киназы; GAPDH - глицеральдегид 3 фосфатдегидрогеназа; G 3P - глицеральдегид 3 фосфат;
HIF1 - фактор 1, индуцируемый гипоксией; HIF1α - α субъединица HIF1; IFNγ - интерферон гамма; LDHA - лактат
дегидрогеназа A; LPS - липополисахарид; mTOR - мишень рапамицина млекопитающих; Nrf2 - ядерный фактор
(nuclear factor erythroid 2 related factor 2); NAC - N ацетилцистеин; NFAT - ядерный фактор активированных Т клеток;
NLRP3 - криопирин (nucleotide binding oligomerization domain, leucine rich repeat and pyrin domain containing 3 protein);
NO - оксид азота (II); NOX - NADРН оксидаза; PHD - пролилгидроксилаза; PPR - скорость генерации протонов;
SHP2 - тирозинфосфатаза 2 (Src homology region 2 domain containing phosphatase 2); TCR - рецептор Т клеток;
TLR4 - толл подобный рецептор 4; TNFα - фактор некроза опухоли; ΔΨ - мембранный потенциал; ρ0 клетки - клетки,
неспособные к дыханию из за дефекта митохондриальной DNA.
* Адресат для корреспонденции.
1939
1940
АНДРЕЕВ и др.
ми редокс центрами (АФК центрами), легко
В частности, основная АФК продуцирующая
подверженными автоокислению [1]. В соответ
активность ассоциирована с дигидролипоамид
ствии с законом действующих масс скорость
дегидрогеназой, являющейся компонентом α
этой реакции пропорциональна количествам
кетоглутаратного и пируватдегидрогеназного
(концентрациям) обоих реагентов. Данная мо
комплексов [12, 13].
дель также учитывает динамическую природу
Традиционно считается, что продукция АФК
АФК центров, которые постоянно циклически
происходит при окислительном стрессе, кото
переходят между восстановленной формой,
рый по определению вызывает разнообразные
способной к реакции с кислородом, и нереакци
повреждающие эффекты в клетках. Накоплен
онноспособной, окисленной формой. Таким
ные данные в пользу регуляторной функции
образом, продукция АФК пропорциональна од
АФК привели к сдвигу парадигмы к признанию
новременно общей концентрации АФК цент
«умеренных» количеств АФК как важных ком
ров и доле активной формы этих центров.
понентов передачи сигнала в клетке. Метаболи
Считается, что основным источником про
ческое репрограммирование активированных
дукции митоАФК в электрон транспортной це
иммунных клеток, особенно Т клеток и макро
пи (ЭТЦ) является комплекс I в сильно восста
фагов, требует значительных биоэнергетических
новленном состоянии, которое может быть дос
перестроек, которые должны влиять как на мас
тигнуто с помощью специфичного ингибитора
су митохондрий, так и на количество и редокс
комплекса I, ротенона, и других ингибиторов
состояние АФК центров, т.е. на оба компонента
ЭТЦ или путем создания определенных усло
предлагаемой модели продукции АФК [1]. Эти
вий (например, истощением цитохрома с). Нао
факторы следует учитывать при исследовании
борот, ротенон ингибирует продукцию АФК
АФК зависимых путей передачи сигнала.
при обратном электронном транспорте (ОЭТ) в
В этом обзоре мы представляем критический
гиперполяризованных митохондриях
[2-4].
взгляд на некоторые метаболические пути АФК
Этот процесс происходит в клетках, находя
зависимой передачи сигнала. Представив клю
щихся в патологических состояниях, способ
чевые иммунометаболические концепции, та
ствующих избыточному накоплению сукцината
кие как переключение на гликолиз и «размыка
и активации проксимальных сайтов продукции
ние» цикла трикарбоновых кислот (ЦТК), мы
АФК. Интересно, что окислительный стресс,
обсуждаем роль иммуномодулирующих метабо
индуцируемый ОЭТ, наблюдается при повреж
литов (иммунометаболитов). Такое обсуждение
дениях, вызванных ишемией/реперфузией
составляет базис для более углубленного анали
[5-7]. Тем не менее, как обсуждается далее,
за биоэнергетики иммунных клеток и их спо
вклад ОЭТ в АФК опосредованную передачу
собности генерировать митоАФК. Мы также
сигнала в нормальных физиологических усло
описываем возможные механизмы АФК опос
виях кажется маловероятным. Другим канони
редованной передачи сигнала в иммунном отве
ческим АФК центром является комплекс III,
те, в частности взаимосвязь между митоАФК и
описанный в новаторских исследованиях в
функционированием индуцируемого гипоксией
1972 году [8, 9]. Продукция АФК комплексом
фактора 1 (HIF1). Еще одной темой нашего об
III представляет собой устойчивый процесс, ко
зора являются митоАФК как молекулярные пат
торый, однако, можно наблюдать только в при
терны, связанные с повреждением (DAMP),
сутствии специфического ингибитора, антими
представляющие собой внутренние медиаторы
цина А, который искусственно увеличивает
воспалительного ответа. Мы также кратко рас
уровень нестабильного семихинона кофермен
сматриваем вопрос взаимодействия между ми
та Q (донора электронов) в части Q цикла со
тоАФК и внемитохондриальные АФК (в основ
стороны матрикса митохондрий. Неясно, моди
ном H2O2, продуцированной в NOX зависимых
фицируется ли комплекс III естественными эф
реакциях в макрофагах). Обзор завершается об
фекторами; следовательно, роль этого АФК
суждением экспериментальных методов, широ
центра в физиологических условиях остается
ко используемых в этой области, и возможных
под сомнением [2, 4, 10, 11]. Одним из аспектов
ошибок в интерпретации экспериментальных
продукции митоАФК, который в отличие от
результатов. Заинтересованный читатель может
ЭТЦ не получает заслуженного внимания, яв
найти более подробное описание затронутых
ляются источники АФК в митохондриальном
тем в специальных обзорах, процитированных в
матриксе. Основываясь на результатах ранних
соответствующих разделах.
исследований, было предположено, что дегид
Основными выводами из анализа концеп
рогеназы митохондриального матрикса вносят
ций, представленных в обзоре являются: 1) пе
вклад в продукцию АФК в нормальных митохо
редача регуляторных сигналов с участием мито
ндриях (в отсутствие ингибиторов ЭТЦ).
АФК, скорее всего, включает в себя умеренные
БИОХИМИЯ том 85 вып. 12 2020
АФК ЗАВИСИМАЯ ТРАНСДУКЦИЯ СИГНАЛОВ В ИММУНИТЕТЕ
1941
изменения, связанные с биогенезом митохонд
реакций биосинтеза, необходимых для роста
рий, а не резкие изменения в метаболическом
клеток [14, 23-25]. Парадоксально, что отличи
состоянии митохондрий, наблюдаемые при
тельной чертой активации иммунных клеток яв
окислительном стрессе; 2) метаболические из
ляется увеличенная продукция лактата, которая
менения в иммунной системе в целом способ
формально предотвращает утилизацию предс
ствуют увеличению преобразования энергии пу
тоящих метаболитов в синтезе макромоле
тем наиболее эффективного механизма - окис
кул [15]. Более того, переключение с эффектив
лительного фосфорилирования (а не переклю
ной продукции АТР митохондриями на менее
чения на гликолиз); 3) классически активиро
эффективный аэробный гликолиз выглядит не
ванные макрофаги являются исключением,
совместимым с энергетическими требованиями
т.к. обладают бактерицидной активностью, ко
клетки для осуществления анаболических реак
торая требует интенсивной продукции химичес
ций и эффекторных функций, а также пролифе
ки активных веществ (оксид азота (II)
рации.
(NO) и H2O2), способных повреждать митохонд
Действительно, метаболические фенотипы
рии.
активированных иммунных клеток и их функ
циональные подтипы различаются (особенно
это заметно для Т клеток и макрофагов). В то
ИММУНОМЕТАБОЛИЗМ:
время как гликолитическое переключение в ак
ОБЩИЕ АСПЕКТЫ
тивированных Т клетках происходит одновре
менно с быстрой активацией митохондриально
Беспрецедентный интерес к метаболической
го дыхания, стимуляция макрофагов приводит к
регуляции врожденного и приобретенного им
увеличению активности iNOS (индуцируемой
мунитета, а также развитие методов высокоточ
синтазы оксида азота) и NO опосредованному
ного «омик» профилирования клеток иммун
ингибированию ЭТЦ [26] (см. обсуждение этой
ной системы способствовали возникновению
проблемы ниже, в разделе, посвящённом биоэ
новой междисциплинарной области исследова
нергетической оценке гликолитического перек
ний: иммунометаболизма. Так, в серии недавних
лючения).
обзоров, представляющих последние достиже
Ферменты гликолиза контролируют эффек;
ния в этой области, основные метаболические
торные функции. Недавние данные об участии
пути рассматриваются в контексте специализи
гликолитических ферментов и метаболитов в
рованных функций иммунных клеток [14-19]. В
эпигенетической регуляции и пост транскрип
данной статье мы представляем некоторые не
ционном контроле эффекторных функций
давние концепции метаболизма иммунной сис
предполагают решающую роль гликолиза в им
темы, уделяя особое внимание гликолитическо
мунном ответе [27-30]. Например, было показа
му переключению и соответствующим измене
но, что экспрессия интерферона гамма (IFNγ) в
ниям в энергетическом метаболизме митохонд
Т клетках негативно регулируется связыванием
рий.
mRNA IFNγ глицеральдегид 3 фосфатдегидро
Гликолитическое переключение. Подобно ра
геназой (GAPDH); такое ингибирование транс
ковым клеткам, демонстрирующим классичес
ляции устраняется при индукции гликолитичес
кий эффект Варбурга, нормальные иммунные
кого переключения и вовлечении GAPDH в
клетки (в частности, Т лимфоциты и макрофа
производство энергии в активированных Т
ги), подвергнувшиеся действию антигенов, ци
клетках [27]. Обратная зависимость между дос
токинов и других стимулов, демонстрируют
тупностью GAPDH и уровнем трансляции цито
быстрый метаболический сдвиг от митохондри
кинов преполагает наличие дополнительной
ального окислительного фосфорилирования к
(неферментативной) функции GAPDH как
аэробному гликолизу. Хотя подобные метаболи
конститутивного ингибитора трансляции в по
ческие изменения хорошо известны (первые со
коящихся (неактивированных) клетках. Недав
общения об увеличении гликолитической ак
но было показано, что GAPDH участвует в сход
тивности в стимулированных лимфоцитах отно
ной метаболической регуляции трансляции
сятся к 1960-1970 гг. [20, 21]), необходимость
фактора некроза опухоли (TNF) в воспалитель
аэробного гликолиза для активации иммунных
ном ответе макрофагов [31]. Эти исследования
клеток и эффекторного ответа понятна не до
подтверждают данные более ранней работы, по
конца. Так как эффекторные функции иммун
казавшей способность GAPDH связывать AU
ных клеток, в частности продукция цитокинов
обогащенные последовательности (AU rich ele
лимфоцитами, связаны с увеличенной способ
ments, ARE) mRNA и влиять на экспрессию ге
ностью к пролиферации [22], активация глико
нов [32]. Оба субстрата GAPDH - глицеральде
лиза, возможно, требуется для осуществления
гид 3 фосфат (G 3P) и NAD+ - вытесняют ARE
БИОХИМИЯ том 85 вып. 12 2020
1942
АНДРЕЕВ и др.
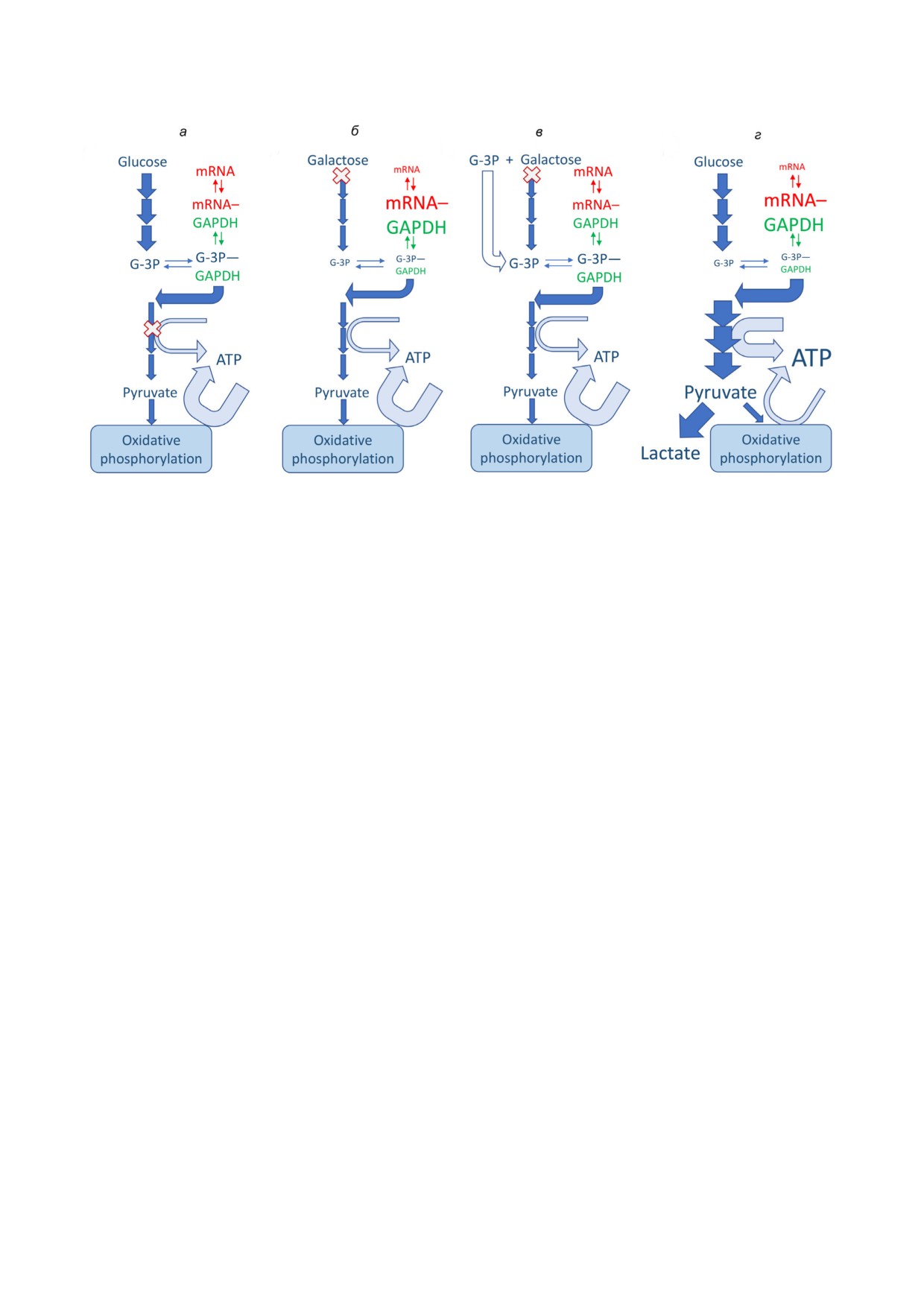
Рис. 1. Регуляция количества ARE-mRNA комплексов гликолитическим потоком. Активности ферментов регулируются
опосредованно путем изменений в концентрациях метаболитов. Соответственно, дополнительная mRNA связывающая
функция GAPDH определяется количеством субстратов (G 3P и пиридиновых нуклеотидов). GAPDH, mRNA и основ
ные реакции гликолитического пути показаны зеленым, красным и голубым цветами. Базовые реакции синтеза лактата
несущественны и не показаны на схеме (а, б). Ширина стрелки соответствует количеству метаболитов. X обозначает точ
ку ограничения метаболического пути. В нормальных условиях (а) точки ограничения представляют собой АТР продуци
рующие ферменты, частично ингибированные из за конкуренции с митохондриальной продукцией АТР. Галактоза вов
лечена в классический гликолитический путь, но поступает в него очень медленно (б, в). Это приводит к связыванию
ARE-mRNA (в том числе mRNA IFNγ) GAPDH и снижению ее концентрации (б), а также уменьшению концентрации
ARE-mRNA, включая IFNγ (б), что обратимо добавлением экзогенного G 3P (в) [27]. Гипотетическое гликолитическое
переключение (г) должно приводить к уменьшению уровня mRNA IFNγ (и белка), что противоречит наблюдаемой про
дукции IFNγ и нуждается в дополнительном исследовании. (С цветными вариантами рис. 1 и 2 можно ознакомиться в
из связывающего сайта [33, 34]. Соответствен
центрациях способны замещать ARE из GAPDH
но, G 3P усиливает продукцию IFNγ в гликоли
[27]. Так как внутриклеточные концентрации
тически недостаточных Т клетках, растущих на
пиридиновых и адениновых нуклеотидов дости
медленно метаболизируемой галактозе, но не в
гают миллимолярных значений, эти соединения
клетках, культивируемых на глюкозе, которые
должны эффективно конкурировать с АRЕ в
уже содержат значительные количества G 3P
любых условиях. Чтобы разрешить это противо
[27] (рис. 1, а-в). Однако следует отметить, что
речие, было предположено, что концентрации
при таком сценарии гликолитический поток ог
нуклеотидов в некоторых компартментах клет
раничен на стадия предшествующих реакции,
ки (ядре или цитоплазме) могут быть значитель
катализируемой GAPDH. Если гликолитичес
но ниже, чем во всей клетке [27], но это допуще
кий поток ограничен на стадиях, следующих за
ние нуждается в проверке.
GAPDH, устранение ограничений будет иметь
Альтернативный механизм координации
противоположный эффект, и накопление ин
гликолитического переключения и биосинтеза
термедиатов, включая G 3P, должно прекра
цитокинов основан на участии лактатдегидроге
титься. Гипотетический сценарий «метаболи
назы А (LDHA), которая катализирует превра
ческого переключения» с окислительного фос
щение пирувата в лактат. Основываясь на ре
форилирования (в котором гликолитические
зультатах генетических экспериментов по сни
АТР продуцирующие ферменты после GAPDH
жению уровня LDHA, было показано, что при
частично ингибированы из за конкуренции с
аэробном гликолизе экспрессия IFNγ контро
митохондриями) на гликолитическую продук
лируется LDHA через ацетил КоА зависимый
цию АТР показан на рис. 1, г.
эпигенетический механизм (ацетилирование
В рамках этой модели, однако, не находит
гистонов), а не GAPDH опосредованной регу
места факт, что продукт реакции NADН и его
ляцией транскрипции [28]. Проявление обоих
структурный аналог АТР в микромолярных кон
механизмов также могет зависеть от каноничес
БИОХИМИЯ том 85 вып. 12 2020
АФК ЗАВИСИМАЯ ТРАНСДУКЦИЯ СИГНАЛОВ В ИММУНИТЕТЕ
1943
ких функций этих белков «домашнего хозяй
митохондриями и способствует его транспорту
ства» (housekeeping proteins), в особенности в
из этих органелл, таким образом повышая уров
парадигме потери активности. Было предсказа
ни синтеза ацетил КоА из цитрата, селективно
но, что GAPDH является ещё одним фермен
го ацетилирования гистонов и транскрипции
том, определяющим скорость гликолиза (в до
IFNγ [28]. Отметим, что ацетил КоА использу
полнение к классическим ферментам, регулиру
ется и в других клеточных процессах, таких как
ющим скорость гликолиза, таким как гексоки
биосинтез жирных кислот и холестерина, а так
наза, фосфофруктокиназа и пируваткиназа)
же в ацетилировании гистонов и негистоновых
[35], в то время как LDHA определяет конечную
белков [41, 42].
реакцию аэробного гликолиза. Таким образом,
Что касается метаболизма цитрата, одним из
GAPDH и LDHA являются важными детерми
потенциально важных регуляторов продукции
нантами в установлении концентраций глико
цитокинов (особенно в макрофагах) и
литических компонентов в устойчивом состоя
NADP+/NADPН баланса в клетке является ан
нии реакции, редокс равновесии NAD+/NADН
типортер цитрата/малата (SLC25A1) [43, 44].
и уровне выживания клеток. Исследования при
Эти и другие недавние исследования сформиро
менения ингибирования LDHA для подавления
вали новый раздел в изучении метаболизма им
эффекта Варбурга в терапии раковых заболева
мунных клеток, сфокусированном на роли мета
ний показали, что такое ингибирование нару
болитов ЦТК (см. ниже, а также об
шает биосинтез серина и аспарагиновой кисло
зор Williams и O’Neill [44]).
ты. Однако в раковых клетках этому эффекту
Концепция «разомкнутого цикла». Основыва
противостоит опосредованная ATF4 (тран
ясь на метаболическом перепрограммировании
скрипционным фактором из семейства цАМР
макрофагов и дендритных клеток, было предпо
зависимых белков) активация экспрессии ге
ложено, что в условиях метаболического перек
нов, ответственных за выживание клеток [36].
лючения и/или нарушения митохондриального
Аналогично пролиферация и эффекторные
дыхания (например, из за продукции NO акти
функции активированных Th1 клеток, включа
вированными макрофагами) происходит рас
ют в себя ATF4 зависимый транскрипционный
согласование ЦТК и ЭТЦ, которое приводит к
ответ, в особенности в условиях дефицита ами
накоплению иммуномодуляторных интермеди
нокислот [37]. Кроме того, лактат per se напря
атов, наиболее заметными из которых являются
мую модулирует функционирование иммунных
сукцинат и цитрат [44, 45]. Однако следует ожи
клеток [38-40] и в высоких концентрациях спо
дать, что подавление транспорта электронов в
собствует снижению продукции цитокинов. Не
ЭТЦ посредством механизма дыхательного
давние исследования обнаружили новый лак
контроля или путем прямого ингибирования
тат зависимый эпигенетический механизм (мо
вызовет почти полное восстановление NAD+ до
дификация лактатом остатков лизина в гисто
NADН. Это приведет к остановке ЦТК из за де
нах), который связан с метаболическим репрог
фицита NAD+, который является субстратом в
раммированием макрофагов после контакта с
четырех реакциях цикла, включая пируватде
бактериальными патогенами. Митохондриаль
гидрогеназную реакцию на входе в цикл. В дан
ный ингибитор ротенон увеличивал уровень
ном случае образования сукцината и цитрата не
данной модификации, что свидетельствует о
будет (рис. 2, а-в). Для обеспечения прохожде
взаимосвязи между активностью окислительно
ния реакций цикла необходимо удаление избыт
го фосфорилирования в митохондриях и регуля
ка восстановительных эквивалентов, который
цией транскрипции. Действительно, количество
должен быть транспортирован из митохондрий
лактата обратно зависит от утилизации пирувата
переносчиком малата/сукцината (рис. 2, б) или
в митохондриях, которая прекращается, когда
утилизирован каким либо еще образом [46].
пул пиридиновых нуклеотидов полностью вос
Вопрос, достаточно ли этих активностей для пе
станавливается в присутствии ротенона. [29].
резапуска ЦТК должен быть решен экспери
LDHA зависимая эпигенетическая модуляция
ментально.
экспрессии цитокинов напрямую связана с ме
Иммунометаболиты: «строительные» блоки
таболизмом митохондрий, особенно с биосин
анаболизма или регуляторные молекулы? Метабо
тезом цитрата в ЦТК. Стимулированные CD4+
литы могут играть роль предшественников стро
T клетки с дефицитом активности LDHA харак
ительных блоков (аминокислот или жирных
теризуются повышенными митохондриальным
кислот) при пролиферации клеток и биосинтезе
дыханием, активностью ЦТК и пониженным
белков и/или регуляторных молекул. При рас
уровнем ацетил КоА [28]. Это предполагает, что
смотрении анаболических процессов необходи
в клетках дикого типа гликолитическое перек
мо принимать во внимание два соображения.
лючение предотвращает утилизацию цитрата
Во первых, маловероятно, что клетки пожерт
БИОХИМИЯ том 85 вып. 12 2020
1944
АНДРЕЕВ и др.
вуют энергетической эффективностью ради
сценариях метаболического репрограммирова
преимуществ анаболизма как «большего блага»,
ния, может служить в лучшем случае для уме
как предполагает концепция гликолитического
ренной коррекции пищевого имбаланса (за иск
переключения. С точки зрения биохимии, им
лючением печени и жировой ткани, специали
мунный ответ является исключительно энерге
зирующихся в липогенезе [47]). Это также озна
тически затратным механизмом, в котором
чает, что в экспериментах in vitro стандартная
энергия расходуется на синтез белка (пролифе
среда для культивации клеток должна
рация и дифференцировка клеток, продукция
пополняться определёнными питательными ве
цитокинов и антител), работы моторных белков
ществами, если они становятся компонентами,
цитоскелета (морфологические изменения кле
лимитирующими рост клеток.
ток), а также функционирование ионных насо
Регуляторные механизмы представляются
сов (ионотропная передача сигнала). Большин
более значимыми, чем анаболические процес
ство клеток, включая иммунные, обладают из
сы. Как отмечено выше касательно их регуля
быточной дыхательной ёмкостью, т.е. превыше
торных функций, сукцинат и цитрат участвуют в
нием емкости ЦТК над общими энергетически
пост трансляционной и пост транскрипцион
ми потребностями, что может быть использова
ной регуляции. Изменения их уровней (в осо
но для биосинтеза без необходимости подавле
бенности, в модели «разомкнутого» ЦТК) зна
ния биоэнергетических процессов. Во вторых,
чительны и достаточны, чтобы вызвать значи
клетки организма, как правило, получают боль
мый эффект. Количество цитрата в макрофагах
шую часть строительных блоков (аминокислот,
костного мозга (bone marrow derived
жирных кислот, холестерина) с пищей. Возмож
macrophages, BMDM) стимулированных толл
ность организма добавить что либо к этому ис
подобным рецептором 4 (TLR 4) увеличивается
точнику весьма ограничена. Из двадцати амино
в 2 раза [48], что может приводить к сходному
кислот десять являются незаменимыми и еще
возрастанию количества ацетил КоА в цитозоле
две синтезируются из незаменимых аминокис
и повышению скорости реакции ацетилирова
лот. Все полиненасыщенные жирные кислоты
ния гистонов. Кроме того, ацетил КоА может
являются незаменимыми или синтезируются из
быть конвертирован в малонил КоА за счет ак
таковых. Способность синтезировать пальмитат
тивности ацетил КоА карбоксилазы. Было по
из цитрата или серина и глицина (из 3 фосфо
казано, что содержание малонил КоА действи
глицерата), которая часто рассматривается в
тельно увеличивается примерно в 2,5 раза, что
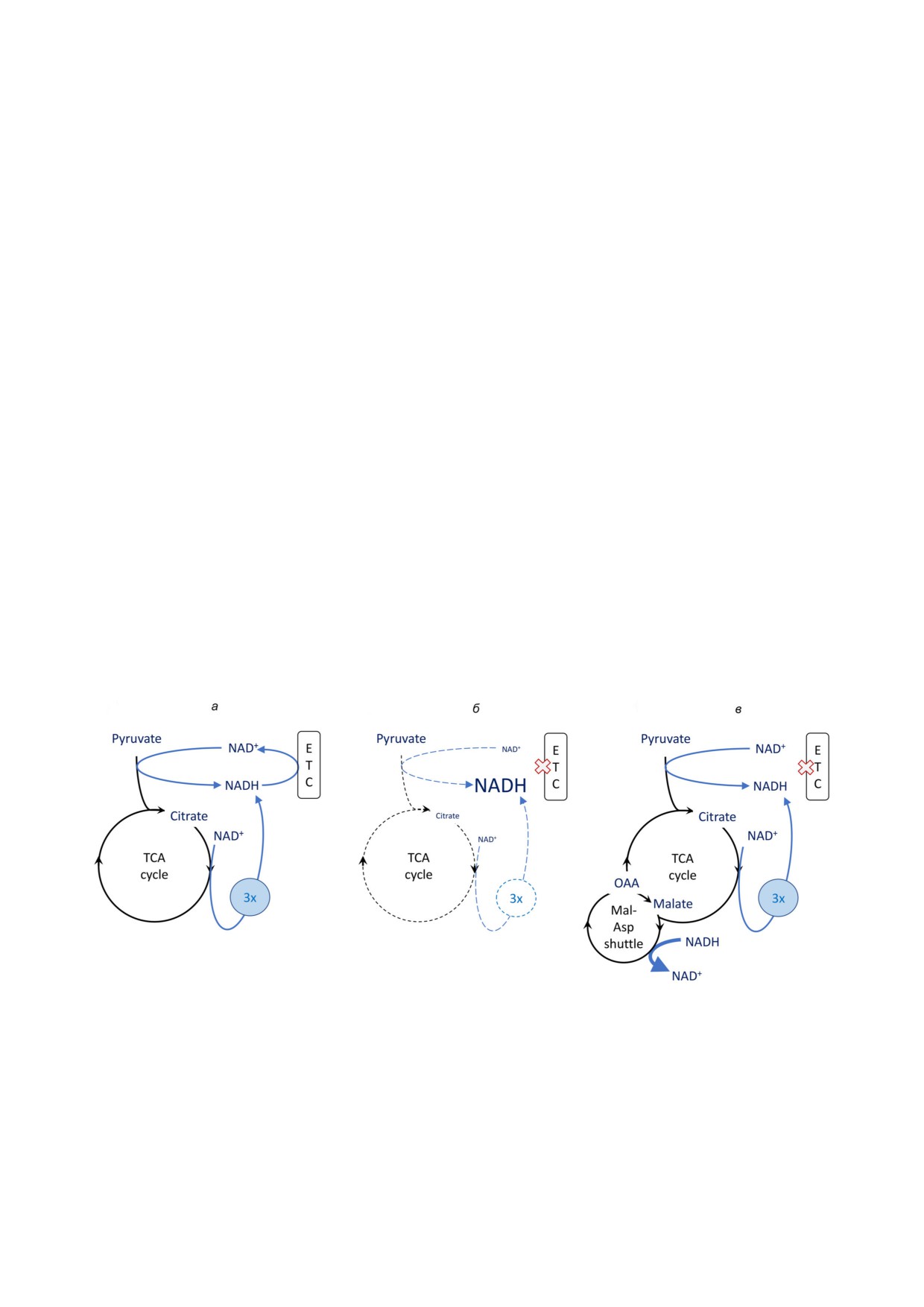
Рис. 2. Подавление и восстановление биосинтетической активности ЦТК доступностью NAD+. Биосинтетическая актив
ность показана на примере синтеза цитрата, необходимого для биосинтеза липидов. Реакции пиридиновых нуклеотидов
показаны голубыми стрелками; «3x» обозначает 3 молекулы NADH, образованных в одном обороте ЦТК. Ширина стрел
ки соответствует величине потока; размер шрифта соответствует количеству метаболита; X обозначает подавление актив
ности ЭТЦ. а - Нормальная продукция цитрата при окислительном фосфорилировании. б - Низкая продукция цитрата
в условиях подавления активности ЭТЦ, в том числе путем дыхательного контроля в условиях гликолитического перек
лючения. в - Гипотетическое восстановление синтеза цитрата через обратный малат аспартатный (Mal Asp) обмен; окис
ление NADH оксалоацетатом (OAA). Стрелка, соответствующая превращению NADH в NAD+, ассоциирована с симво
лом Mal Asp обмена (в противоположность конверсии OAA в малат) для простоты схемы. Символы NADH (и/или NAD+)
представляют общий пул NADH (и/или NAD+)
БИОХИМИЯ том 85 вып. 12 2020
АФК ЗАВИСИМАЯ ТРАНСДУКЦИЯ СИГНАЛОВ В ИММУНИТЕТЕ
1945
приводит к малонилированию различных бел
возрастает до миллимолярных концентраций в
ков, в том числе и GAPDH [49]. Малонилирова
течение нескольких часов после активации кле
ние лизина 213 (или его замена на глутаминовую
ток TLR4, что превращает итаконат в наиболее
кислоту, симулирующая эту модификацию) уве
представленный дикарбоксилат в клетке
личивает ферментативную активность прибли
[51, 54]. Такая кинетика сходна с кинетикой дру
зительно в 3 раза и полностью высвобождает
гих важных регуляторных метаболитов, таких
связанный транскрипт TNFα [49]. Таким обра
как простагландины [55] и оксид азота [56]. Она
зом, одна и та же модификация, индуцируемая
отражает кинетику цис аконитатдекарбоксила
повышением содержания цитрата, одновремен
зы (CAD; также известной как иммуноподавля
но усиливает энергетические и эффекторные
ющий белок гена 1, IRG1), фермента, количест
функции.
во которого сильно (почти в 200 раз) возрастает
Содержание сукцината в TLR 4 стимулиро
при индукции и который превращает интерме
ванных BMDM также повышено в 2 раза [48].
диат ЦТК аконитат в итаконат [54]. У млекопи
Сукцинат ингибирует пролилгидроксилазу
тающих эта реакция происходит почти исклю
(PHD) с IC50 ~ 0,5 мМ, что находится в пределах
чительно в макрофагах [57]. Хотя остается неяс
его внеклеточной концентрации [50]. Из за
ным, что заставляет аконит, связанный в актив
структурной схожести ингибирование имеет
ном центре аконитазы, диссоциировать и всту
конкурентный характер и может быть предотв
пать в реакцию, катализируемую CAD.
ращено с помощью α кетоглутарата, который
Роль итаконата остается предметом для дис
является субстратом фермента [48]. Соответ
куссий: является ли его действие провоспали
ственно, активность PHD должна зависеть от
тельным благодаря ингибированию СДГ и на
соотношения между этими метаболитами в
коплению сукцината [51] или противовоспали
большей степени, чем от содержания каждого из
тельным [50, 58]? Последнее предположение
них. Количество α кетоглутарата в TLR 4 сти
подтверждается фактом, что содержание цито
мулированных макрофагах RAW 264.7 остается
кинов IL 1β, IL 6, IL 18 и IL 12 (но не TNFα) в
неизменным, в то время как содержание сукци
клетках с нокаутированной CAD слегка повы
ната увеличивается в 3 раза, что сдвигает соот
шено (на 20-30%), предполагая существование
ношение в пользу ингибирования [51]. Ингиби
иммунной супрессии итаконатом в клетках ди
рование PHD предотвращает дестабилизацию
кого типа. Однако уровень транскрипта hif 1α в
HIF1α (см. раздел «Митохондриальные АФК и
клетках с нокаутом CAD увеличивается в 10 раз,
регуляция HIF1») и позволяет транскрипцию
приводя к увеличению количества белка HIF1α
многих генов, включая ген провоспалительного
[52], что само по себе может объяснить умерен
цитокина IL 1β [48].
ный провоспалительный эффект. Благодаря
Общий пул метаболитов ЦТК (в том числе
присутствию реактивной двойной связи, сопря
сукцината и цитрата) в активированных макро
женной с карбоксигруппой, итаконовая кислота
фагах увеличивается, что предполагает актива
может участвовать в реакциях присоединения
цию анаплероза [48]. Исследования с использо
Михаэля с тиолами белков. Было предположе
ванием стабильных изотопов показали, что наи
но, что итаконат оказывает свое действие через
более значительный вклад в анаплероз вносит
пост трансляционные модификации Kelch по
превращение глутамина в α кетоглутарат через
добного белка 1 (KEAP1) с образованием аддук
глутамат [48]. Накопление сукцината может
тов Михаэля и последующей активацией проти
осуществляться в результате ингибирования
вовоспалительного ядерного фактора Nrf2 [58].
сукцинатдегидрогеназы (СДГ), например ита
В целом, однако, учитывая мощный ответ ита
конатом (см. ниже) [51, 52]. Однако классичес
коната на стимуляцию, вполне вероятно, что мы
кого пункта перекреста на уровне СДГ нет, т.к.
пока слабо преставляем себе полный потенциал
количество интермедиатов пост СДГ, фумарата
этого метаболита.
и малата, не только не уменьшается, но даже мо
Несколько замечаний о передаче сигнала. Зна
жет увеличиваться [48], что свидетельствует о
чительный интерес в исследовании иммуноме
незначительной роли СДГ в аккумуляции сук
таболизма сосредоточен на динамических пост
цината. Эксперименты по изучению метаболи
активационных изменениях в метаболизме ли
ческих путей с использованием стабильных изо
пидов, в частности на зависимом от гликолиза
топов исключили возможность биосинтеза сук
de novo синтезе жирных кислот (СЖК). Метабо
цината непосредственно из итаконата [51].
лический сдвиг, сопровождаемый переходом от
Итаконат вполне можно рассматривать как
окисления жирных кислот (ОЖК) к СЖК, акти
архетипичный метаболит иммунной системы
вируется сигнальными каскадами, ассоцииро
[53]. Хотя итаконат отсутствует в покоящихся
ванными с белком мишенью рапамицина мле
макрофагах, его количество экспоненциально
копитающих, mTOR (mammalian target of
БИОХИМИЯ том 85 вып. 12 2020
1946
АНДРЕЕВ и др.
rapamycin), действию которого противостоит
[69] и стимулированных липополисахаридами
активность
AMP активируемой киназы
(LPS) макрофагов [70]. В этом разделе обзора
(AMPK), которая, в свою очередь, активирует
мы попытались оценить эти сценарии с точки
ОЖК [15, 59]. Сериновая/треониновая киназа
зрения биохимика. Наш анализ построен преж
mTOR связывает доступность питательных ве
де всего на данных, полученных в обстоятель
ществ и кислорода с соответствующими тран
ном исследовании гликолитического переклю
скрипционными программами. Эта регулятор
чения в TLR4 активированных макрофагах [70],
ная сеть контролирует различные аспекты врож
которое позволило расшифровать системы ме
денного и приобретенного иммунитета, вклю
таболических изменений, происходящих при
чая поляризацию макрофагов, гликолитическое
провоспалительной поляризации макрофагов,
репрограммирование Т клеток, клеточную миг
приводящей в итоге к стабилизации HIF1α и
рацию, функционирование регуляторных Т
продукции цитокинов.
клеток (Treg) и продукцию цитокинов различны
Понятие гликолитического переключения
ми типами иммунных клеток [15, 59-62]. Одним
предполагает активацию гликолиза, так что уро
из плейотропных эффектов mTOR является уси
вень продукции АТР в гликолизе начинает пре
ление биогенеза митохондрий и окислительного
вышать уровень его продукции при окислитель
метаболизма [62]. Помимо сенсора кислорода
ном фосфорилировании, что вызывает ингиби
HIF 1α (см. ниже), к необходимым транскрип
рование окислительного фосфорилирования за
ционным факторам, контролирующим метабо
счет дефицита субстрата (АDP). Соответствен
лическое репрограммирование, относятся
но, соотношение ATP к АDP (AТP/АDP) после
Ca2+/кальцинейрин зависимый ядерный фак
перехода клеток на гликолиз должно увеличи
тор активированных Т клеток (NFAT) [63, 64] и
ваться. Однако в TLR4 стимулированных
c Myc (MYC протоонкоген), один из основных
BMDM соотношение ATP/АDP снижается при
регуляторов экспрессии генов, ответственных за
мерно на 40%, что соответствует увеличению
выживание клеток [23, 45, 60, 65]. Недавно было
уровня АDP с 5 до 8% от общего пула нуклеоти
показано, что ядерный фактор kappa B (NF kB),
дов. Хотя такой подъем в уровне АDP должен
являющийся транскрипционным активатором в
вызывать активацию дыхания, дыхание клеток
воспалительном ответе, играет роль негативного
снижается примерно на 40%, что подразумевает
регулятора при избыточном воспалении [66].
совершенно иной механизм: активное подавле
Данный механизм включает в себя сложное вза
ние окислительного фосфорилирования, а не
имодействие между избирательной элиминаци
дыхательный контроль (контроль уровней
ей поврежденных митохондрий (митофагией) и
АDP). С наибольшей вероятностью окислитель
активацией NLRP3 инфламмасомы [67]. Эти
ное фосфорилирование подавляется оксидом
компоненты сигнальной сети являются объек
азота, NO [71]. С 1989 года было известно, что
том активных исследований и были подробно
активированные макрофаги производят NO,
описаны в обзорах, посвящённых передаче сиг
который вызывает подавление дыхания в клет
нала в клетках [62, 66, 68].
ках мишенях [72]. NO обратимо ингибирует ок
сидазу цитохрома c (дыхательный комплекс IV),
конкурируя с кислородом. NO продуцируется
БИОЭНЕРГЕТИЧЕСКАЯ ОЦЕНКА
iNOS, экспрессируемой при активации клеток.
ГЛИКОЛИТИЧЕСКОГО ПЕРЕКЛЮЧЕНИЯ
Так, культивируемые макрофаги способны про
изводить NO в количествах до 1 мкМ, что доста
Часто повторяющейся темой в исследовани
точно для подавления их дыхания [73]. Длитель
ях иммунометаболизма является концепция ме
ное воздействие NO может вызвать необратимое
таболического переключения, т.е. обязательного
ингибирование дыхательного комплекса I [74].
перехода клеток к использованию специализи
NO подавлет активность аконитазы 2 и пируват
рованных путей метаболизма для осуществле
дегидрогеназы, т.е. стадий, предшествующих
ния иммунных функций. В частном случае гли
дыхательной цепи, что может служить причи
колитического переключения эта концепция
ной наблюдаемого снижения уровня потребле
включает в себя набор представлений и поня
ния кислорода дыхательной цепью [75]. Неста
тий, таких как переход к аэробному гликолизу,
бильность NO может объяснить существенные
сигнальная активность АФК и/или иммуноме
различия в степени ингибирования дыхания,
таболитов, которые в целом или по отдельности
наблюдаемой в разных работах.
прилагаются к исследованиям различных кле
Использование платформ для анализа мета
ток иммунной системы. Во всей полноте такой
болических потоков, разработанных для однов
механизм был предложен, например для моно
ременной оценки потребления кислорода (OCR,
цитов, инфицированным вирусом SARS CoV 2
oxygen consumption rate) и скорости закисления
БИОХИМИЯ том 85 вып. 12 2020
АФК ЗАВИСИМАЯ ТРАНСДУКЦИЯ СИГНАЛОВ В ИММУНИТЕТЕ
1947
внеклеточной среды (ECAR, extracellular acidifi
фагов (продукцию цитокинов) и значительно
cation rate) с развитым анализом данных [76],
снижает смертность животных в моделях бакте
позволяет количественно оценить относитель
риальной инфекции и/или гипервоспаления
ные вклады окислительного фосфорилирования
[48, 80, 81]. Эти результаты подчеркивают суще
и гликолиза в общую биоэнергетическую актив
ствование точной настройки энергетических
ность клеток иммунной системы. В работе
требований в иммунном ответе, которая сохра
Mils et al. [70] представлены подробные данные
няется, несмотря на существующий энергети
для такого анализа, которые включают достаточ
ческий дефицит, но при этом существенно зави
но редко измеряемую скорость генерации прото
сит от остаточного обеспечения энергией за счет
нов (proton production rate, PPR) (в данном слу
метаболизма глюкозы. Последний, очевидно, не
чае измерения ECAR было недостаточно). Наш
ограничен гликолизом и включает в себя окис
анализ этих данных основан на суммарных сте
лительный метаболизм гликолитически проду
хиометриях окислительного фосфорилирования
цированного пирувата в митохондриях. Остает
и гликолитического пути. В соответствии с пос
ся вопрос, насколько другие митохондриальные
ледними оценками выход ATP в окислительном
субстраты могут замещать пируват.
фосфорилировании составляет 30-32 молекулы
Следует заметить, что, несмотря на актива
АТР на молекулу глюкозы (как следует, напри
цию гликолиза, подавление окислительного
мер из соотношения АТР/О, приводимых
фосфорилирования в альтернативно активиро
Nichalls и Ferguson [77]) вместо канонических 36
ванных макрофагах (например, в ответ на IL 4)
молекул ATP [78]. Соответственно, мы принима
отсутствует. Наоборот, в таких клетках происхо
ем, что АТР/O2 = 5,45 [79]. Общепринятая стехи
дит опосредованный транскрипционным акти
ометрия гликолиза - продукция одной молеку
ватором PGC 1β биогенез митохондрий, кото
лы АТР на одну молекулу лактата (АТР/Н+ = 1)
рый приводит к увеличению их способности к
[77]. Однако расчёт продукции АТР в гликолизе,
окислительному метаболизму [82, 83], соответ
основанный на определении PPR, предоставляет
ствующему повышенным энергетическим тре
оценку только верхнего предела (максимальных
бованиям клеток [71, 84, 85]. Сходные измене
значений) из за вклада CO2, производимого ми
ния происходят в биоэнергетике Т клеток в от
тохондриями. Мы конвертировали опублико
вет на стимуляцию рецепторов Т клеток, когда
ванные значения PPR и OCR в скорость синтеза
активируются одновременно гликолиз и окис
АТР. В состоянии покоя 2 ⋅ 105 BMDM продуци
лительное фосфорилирование [27, 86]. Таким
руют ~1500 пмоль ATP в мин; стимуляция TLR4
образом, даже если усиление гликолиза являет
снижает уровень синтеза АТР клетками пример
ся характерным ответом иммунных клетк на
но на 40% (до ~900 пмоль/мин). Вклад гликоли
различные стимулы, гликолитическое переклю
за увеличивается с ~3% (покоящиеся клетки) до
чение имеет ограниченное значение.
~17% (активированные клетки). Такие измене
Обратный электронный транспорт и АФК. До
ния метаболизма не достигают уровня, позволя
полнительного обсуждения в данном контексте
ющего говорить о «переключении», восприни
заслуживает роль продукции АФК, опосредо
маемом как переход к преимущественной опоре
ванной обратным электронным транспортом.
на гликолиз для удовлетворения энергетических
Вполне вероятно, что гликолитическое перек
нужд клеток.
лючение могло бы приводить к активации ОЭТ
Парадоксально, что в TLR4 стимулирован
при соблюдении определенных условий. Экспе
ных макрофагах митохондриальная функция
рименты на изолированных митохондриях по
ингибирована, в то время как вклад гликолиза
казали, что данный механизм исключительно
является малосущественным и не обеспечиваю
чувствителен к любой деполяризации митохон
щим энергетических потребностей клетки. Это
дриальной мембраны, поскольку он движим
приводит к снижению синтеза АТР и соотноше
мембранным потенциалом (ΔΨ)
«в горку»,
ния АТР/ADР, неизбежно вызывая подавление
т.е. против редокс потенциала переносчиков
АТР потребляющих процессов, которые в по
[87, 88]. Для работы этого механизма митохонд
коящихся клетках представляются собой, по
рии должны быть в, по существу, нефосфорили
большей части, процессы «домашнего хозяй
рующем состоянии (состояние 4, гиперполяри
ства», абсолютно необходимые для выживания.
зация). Этого не происходит в митохондриях,
Однако в более широком биологическом смыс
стимулированных BMDM, которые сохраняют
ле для провоспалительных макрофагов задача
более чем половину нормальной АТР синтети
выживания вполне может не стоять как сущест
ческой активности (см. выше). Прямые измере
венная.
ния выявили, что, хотя сигнал ΔΨ чувствитель
Введение ингибитора гликолиза, 2 дезок
ного зонда ТМРМ увеличивается при стимуля
сиглюкозы, подавляет иммунный ответ макро
ции клеток, он не достигает уровня, наблюдае
БИОХИМИЯ том 85 вып. 12 2020
1948
АНДРЕЕВ и др.
мого в состоянии 4, индуцированном олигоми
чение сукцината из алкилированного производ
цином [70]. Кроме того, такое изменение про
ного (например, диэтилсукцината) зависит от
порционально увеличению флуоресценции ΔΨ
его расщепления неспецифическими внутрик
нечувствительного митохондриального зонда
леточными эстеразами неизвестной природы.
MitoTracker Green, что, скорее всего, предпола
Эти активности достаточны для захвата и удер
гает ответ на увеличение объема матрикса мито
жания в клетках различных зондов и/или инги
хондрий, а не на изменение ΔΨ. Также, в исход
биторов. Однако способность расщеплять доста
ном определении ОЭТ является транспортом
точное количество молекул метаболитов ЦТК
электронов с сукцината на NAD+ и проявляется
для поддержания нормального метаболического
в высоко восстановленном состоянии
потока пока не была доказана. В частности, из
NAD+/NADН. Однако в активированных
упомянутой выше работы Mils et al. [70] очевид
BMDM 90% пиридиновых нуклеотидов окисле
но, что диэтилсукцинат (называемый в этой ра
ны. В целом, по нашему мнению, стимулиро
боте просто сукцинатом) не способен увеличить
ванные BMDM не обладают необходимыми ус
внутриклеточный пул сукцината. Тем не менее
ловиями для ОЭТ и ОЭТ опосредованной про
он вызывает характерный ряд биологических
дукции АФК. Также при изучении ингибитор
эффектов, таких как стимуляция TLR4 зависи
ных свойств АФК митохондриальные ингибито
мой провоспалительной передачи сигнала (про
ры, способные эффективно блокировать ОЭТ,
дукции IL 1β) и подавление противовоспали
добавлялись за 3 часа до стимуляции клеток
тельного сигнала (IL 10 и IL 1RA), не влияя при
[70], что оставляло достаточно времени для про
этом на TNFα. Удивительно, что диэтиловый
явления множественных альтернативных эф
эфир бутилмалоната (ингибитор переносчика
фектов. Хотя вполне возможно, что определяю
дикарбоксилата) проявляет сходные биологи
щим механизмом все же является АФК зависи
ческие свойства, хотя известно, что он подавля
мая передача сигнала, осуществляемая через
ет окисление сукцината в изолированных мито
стабилизацию HIF1α, происхождение АФК в
хондриях путем предотвращения его доступа к
данном случае остаётся непонятным. Упомяну
СДГ. Это несоответствие предполагает, что оба
тое выше увеличение сигнала MitoTracker Green
диетиловых эфира могут оказывать сходный по
[70] согласуется с предположением об увеличе
бочный эффект. Кроме того, диэтилсукцинат
нии массы митохондрий, что также поддержи
per se вызывает очень незначительную продук
вается синтезом дополнительных молекул мито
цию АФК (~40% при концентрации 5 мМ) по
хондриальной DNA в активированных макро
сравнению с 4 кратным увеличением генерации
фагах [89]. Это предполагает биогенез с однов
АФК лигандом TLR4, LPS. К сожалению, отве
ременным увеличением количества митохонд
ты АФК на комбинацию этих двух эффекторов,
риальных редокс переносчиков, в том числе
вызывающих упомянутый выше характерный
АФК продуцирующих центров. Само по себе
ряд биологических эффектов, не известны. В от
это может быть достаточным для увеличения об
личие от диэтилсукцината, эффект LPS на повы
щей продукции АФК, которая к тому же может
шение клеточного уровня сукцината (в отсут
дополнительно усиливаться в слегка восстанов
ствии экзогенных источников) в 3 раза превы
ленной среде (изменение уровня восстановле
шает эффект ингибитора СДГ, малоната. Это
ния NADН с ~8 до ~13% [70]), предположитель
предполагает, что TLR4 зависимые метаболи
но за счет частичного блокирования дыхатель
ческие изменения не заканчиваются перерасп
ной цепи.
ределением в общем пуле интермедиатов ЦТК и
Сукцинат и продукция АФК. Классические
могут увеличивать этот пул примерно до 10 раз
эксперименты на изолированных митохондриях
(анаплероз). Такое увеличение полностью сов
используют окисление экзогенного сукцината
местимо с предполагаемым ростом митохондри
как источник ОЭТ зависимой генерации АФК,
альной массы (см. выше).
равно как и продукции АФК комплексом III.
Клетки ρ0 как модель гликолитического перек;
Однако сукцинат неизбежно присутствует в ин
лючения. Клетки с индуцируемым дыхательным
тактных клетках как метаболит, поэтому потреб
дефицитом (ρ0), полученные путем экспрессии
ность в экзогенном сукцинате трудно объяснить,
доминатно негативной DNA полимеразы γ и
если только не происходит значительного исто
описанные нами ранее [90], использовались для
щения метаболитов ЦТК. В последнем же случае
моделирования «метаболического переключе
любой анаплеротический интермедиат способен
ния» и исследования влияния внутриклеточных
заменить сукцинат. Так, гораздо легче обеспе
сигналов, продуцируемых митохондриями
чить доставку глутамина (предшественника α
[91, 92]. В митохондриях этих клеток отсутству
кетоглутарата), чем сукцината, который не про
ет дыхательная цепь, что нарушает их способ
никает через плазматическую мембрану. Полу
ность окислять образованный в ЦТК NADН и
БИОХИМИЯ том 85 вып. 12 2020
АФК ЗАВИСИМАЯ ТРАНСДУКЦИЯ СИГНАЛОВ В ИММУНИТЕТЕ
1949
поддерживать функционирование ЦТК. Это
тохондриальных АФК, отличающейся от их ши
приводит к снижению биосинтеза цитрата и его
роко известных проапоптотических функций и
продукта, ацетил КоА, и, как следствие, к
других повреждающих клетку активностей, ин
уменьшению ацетилирования гистонов. Нес
дуцируемых при нарушении работы митохонд
колько специфичных сайтов теряют ацетилиро
рий в различных патологических состояниях.
вание гистонов в ρ0 клетках, что подразумевает
МитоАФК, функционирующие как сигнальные
потенциальное подавление экспрессии генов
молекулы, вовлечены в автофагию, митофагию,
[91]. Было показано, что деацетилирование гис
биогенез митохондрий, клеточный цикл и дру
тонов обратимо за счет экспрессии альтернатив
гие клеточные процессы, контролируемые ре
ной дыхательной цепи, состоящей из оксидоре
докс зависимой передачей сигнала и тран
дуктазы NDI1 и митохондриальной оксидазы
скрипционными факторами. Белки, функцио
AOX, что было интерпретировано как реактива
нирование которых регулируется АФК, пере
ция ЦТК путем окисления NADН. Однако со
числены во многих обзорах. Список белков ми
отношение NAD+/NADН в клетках с феноти
шеней АФК включает ATF4, ядерный фактор
пом ρ0 осталось неизменным [91], предполагая
kappa B (NF kB), митоген активируемые кина
существование иных механизмов деацетилиро
зы (MAPK), внеклеточные сигнал регулируе
вания гистонов. Совместное действие NDI1 и
мые киназы (ERKs), p38 MAPK, c Jun N терми
AOX не способно приводить к генерации АТР,
нальные киназы (JNKs) [93]; тирозинфосфатазы
следовательно, экспрессия этих белков не вос
и p53 [10, 94-103]. Тем не менее механизм обра
станавливает пролиферцию ρ0 клеток [91], что
зования АФК и АФК зависимой передачи сиг
указывает на исключительную важность биоэ
нала в контексте иммунометаболического регу
нергетических (а не биосинтетических) функ
лирования требует дальнейших исследований и
ций. Способность ρ0 клеток к пролиферации
вероятного переосмысления существующих
может быть незначительно (на 10-15%) восста
концепций.
новлена нормализацией ΔΨ. Эти результаты,
Т;клетки: основные моменты. Множествен
параллельно с данными об ответе индикатора
ные данные позволяют предположить, что ми
CellROX, позволили предположить механизм
тоАФК являются потенциально незаменимыми
АФК зависимой передачи сигнала по HIF1
факторами для активации Т клеток, их эффек
опосредованному пути [91]. Однако регуляция
торных функций и дифференциации [42, 49, 96,
образования АФК посредством ΔΨ осуществля
102, 104-108]. Раннее исследование TCR акти
ется через гипервосстановленное состояние ми
вированных клеток описывает немедленное
тохондрий, которое может быть детектировано
действие TCR зависимой продукции АФК на
по гипервосстановленности пары NAD+/
сигнальные пути, регулирующие пролифера
NADН, но не наблюдается в ρ0 клетках [91]. Как
цию и выживание Т клеток [109]. Удивительно,
альтернативное объяснение можно предполо
что разные АФК оказывали раличное действие
жить, что ΔΨ необходим для импорта белков в
на клетки: перекись водорода активировала пе
митохондрии, в том числе компонентов биосин
редачу пролиферационного сигнала в ERK за
тетических систем, критически необходимых
висимом каскаде, в то время как супероксид
для пролиферации клеток и функционирования
анион индуцировал клеточную смерть (через
основного АФК продуцирующего фермента,
экспрессию лигандов FAS). При этом ряд эф
дигидролипоамиддегидрогеназы [12, 13]. В це
фектов, включая активацию NFAT и экспрес
лом модель, использующая ρ0 клетки, подчер
сию IL 2, не зависели от АФК [109]. Хотя авто
кивает важность биоэнергетики для функцио
ры исследования не указали место генерации
нирования клеток, в то время как роль гликоли
АФК, представляется вероятным участие мито
тического переключения представляется спор
АФК в описанных эффектах. На основе резуль
ной. Данная модель не объясняет механизм об
татов химической и генетической инактивации
разования АФК, и, принимая во внимание на
компонентов ЭТЦ другие авторы связали Т
личие поврежденной дыхательной цепи, значе
клеточные ответы с сигнальной функцией ми
ние этой модели остается неясным.
тоАФК, образованных в ЭТЦ [104, 110]. Так,
CD4+ T клетки с дефицитом железо серного
белка Rieske (функционально обеспечивающего
МИТОХОНДРИАЛЬНЫЕ АФК
классический АФК генерирующий центр в
КАК СИГНАЛЬНЫЕ КОМПОНЕНТЫ
комплексе III) демонстрировали нарушения в
ИММУННОГО ОТВЕТА
Ca2+ зависимом NFAT-IL 2 сигнальном каска
де, в то время как остальные редокс зависимые
В последнее время увеличивается количест
пути (например, активность ERK) не были зат
во статей, посвященных регуляторной роли ми
ронуты дефицитом белка Rieske. Данные экспе
БИОХИМИЯ том 85 вып. 12 2020
1950
АНДРЕЕВ и др.
риментов по обращению этого эффекта с ис
кад и реакции, опосредованные интерфероном
пользованием экзогенной H2O2 поддерживают
типа I [117-119]. Это раскрывает еще одну важ
роль АФК зависимого фактора в индукции IL
ную роль митохондрий в иммунитете, которая
2. Интересно, что в этих условиях гликолиз не
дополняет их функцию по обеспечению клетки
был необходим для активации T клеток и про
АТР и классическую передачу сигнала в апопто
дукции IL 2 [104]. В другой работе было обнару
зе через высвобождение цитохрома с [120, 121].
жено, что продукция IL 2 и IL 4 активирован
Для того, чтобы эмитировать такие «SOS» сиг
ными Т клетками существенно снижалась ан
налы, митохондрии должны быть избирательно
тиоксидантом N ацетилцистеином (NAC). Про
пермеабилизованы или в какой то степени пов
дукция интерлейкинов в клетках, дефицитных
реждены, хотя точные пути для высвобождения
по комплексу I дыхательной цепи, также была
DAMP пока не известны. Как кандидаты в
снижена. Авторы предположили, что именно
структуры, участвующие в высвобождении DNA
комплекс I является источником регуляторных
и АФК и последующей активации NLRP3, были
АФК [110]. Индуцированная TCR экспрессия
предложены компоненты митохондриальной
цитокинов и продукция H2O2 ингибировалась
апоптотической поры. Пермеабилизация мито
ротеноном (в концентрациях, значительно пре
хондрий при апоптозе включает два хорошо изу
вышающих специфические для ингибирования
ченных механизма: формирование неселектив
комплекса I), что вызывает вопрос - действи
ной поры во внутренней мембране митохонд
тельно ли сигнал АФК приходит из комплекса I.
рий (mitochondrial permeability transition pore,
Менее инвазивный метод (по отношению к
MPTP) и формирование Bax/Bak зависимой
функционированию дыхательной цепи) заклю
поры во внешней мембране митохондрий (mito
чается в использовании классических антиокси
chondrial outer membrane permeabilization поры,
дантов и антиоксидантов, специфичных для ми
MOMP) [122]. Последний механизм не наруша
тохондрий. Хотя такой подход и недостаточен
ет целостность внутренней митохондриальной
для идентификации молекулярных механизмов
мембраны. Считается, что он не участвует в им
и участников АФК опосредованной передачи
мунном ответе, хотя было показано, что актива
сигнала (таких как сайт генерации АФК и бел
ция эффекторных киназ после индукции
ков мишеней), он обеспечивает необходимые
MOMP белками Bax/Bak способствует NLRP3
для подтверждения концепции результаты, до
зависимой продукции IL 1β [123]. Неселектив
казывающие участие АФК в разнообразных сиг
ная MPTP может пропускать DNA [124, 125].
нальных путях. В разделе «Инструменты» мы
Так, недавно было предположено, что TDP 43
обсудим преимущества и недостатки наиболее
зависимое воспаление опосредовано транслока
часто применяемых систем для защиты от АФК.
цией митохондриальной DNA через MPTP. Од
АФК как «SOS» сигналы. Mолекулярныe
нако это предположение было сделано на осно
фрагменты, ассоциированныe с патогенами
ве наблюдаемых эффектов ингибитора MPTP,
(pathogen associated molecular patterns, PAMPs)
циклоспорина А, примененного в дозе пример
или повреждениями (DAMPs), узнаются рецеп
но в 10 раз превышающей необходимую для спе
торами иммунных клеток как сигналы опаснос
цифической активности [126]. Не было исклю
ти и запускают острый иммунный ответ, приво
чено, что в данном случае наблюдаемые эффек
дящий к элиминации патогена и регенерации
ты были результатом действия на альтернатив
поврежденной ткани. Иммунный ответ на
ную мишень - кальцинейрин. Кроме того, фор
DAMP часто называют стерильным воспалени
мирование MPTP обычно приводит к полной
ем, т.к. его причиной являются внутриклеточ
потере функции митохондрий, и понадобился
ные (а не внешние) агенты [111]. Изучение рас
бы какой то дополнительный механизм для
познавания сигналов опасности макрофагами
предотвращения этого. В недавнем исследова
позволило идентифицировать сигнальные пути,
нии был описан регуляторный механизм, кото
ведущие к активации NLRP3 инфламмасомы,
рый включает в себя взаимодействие между ти
которая служит молекулярной платформой для
розинфосфатазой 2 (SHP2) и транслоказой аде
интеграции прокаспазы I и процессинга про
ниновых нуклеотидов 1 (ANT1), одним из пред
воспалительных цитокинов при стрессе [67, 89,
полагаемых компонентов МPTP. Было предпо
112-115]. Активирующие инфламмасому
ложено, что SHP2 опосредованное ингибирова
DAMP включают в себя молекулы, образован
ние ANT1 ограничивает высвобождение DAMP
ные в митохондриях, такие как АФК, кардиоли
и предотвращает избыточную активацию ин
пин и митохондриальная DNA [67, 89, 113, 116].
фламмасомы [127]. Так как преобладающим
Высвобожденная митохондриальная DNA так
АФК является H2O2, которая свободно проника
же участвует в антивирусном врожденном имму
ет через мембраны, логично преположить, что
нитете, запуская cGAS STING сигнальный кас
сигнал митоАФК, вызывающий активацию
БИОХИМИЯ том 85 вып. 12 2020
АФК ЗАВИСИМАЯ ТРАНСДУКЦИЯ СИГНАЛОВ В ИММУНИТЕТЕ
1951
инфламмасомы, связан с незначительными из
ми кислорода, т.к. значения Км только слегка
менениями в митохондриях и не требует катаст
превышают концентрацию кислорода в водной
рофического увеличения проницаемости мито
среде (230-250 мкМ) [133]. Следовательно, ак
хондрий, вызываемого открытием МРТР. В этом
тивность этих белков должна почти линейно за
отношении обратимое состояние MPTP и сос
висеть от парциального давления кислорода в
тояние с низкой проводимостью [10, 128, 129]
условиях гипоксии. Существование любого дру
заслуживают особое внимание, т.к. могут быть
гого механизма для оценки количества кислоро
связаны с АФК зависимой передачей сигнала от
да предполагает, что фундаментальный конт
функционально интактных митохондрий.
роль в соответствии с законом действия масс ка
В активации инфламмасомы NLRP3 участ
ким либо образом утерян, и активность фер
вуют митоАФК, производимые комплексами I и
ментов остается высокой, несмотря на практи
III (но, ожидаемо, не комплексом II) [113]. Ин
ческое отсутствие одного из субстратов (кисло
тересно, что продукция АФК и последующий
рода). Разработка и доказательство существова
воспалительный иммунный ответ ингибирова
ния подобного сценария может стать благород
лись подавлением активности потенциал зави
ной задачей для будущих энзимологов.
симых анионных каналов (voltage dependent
Аргументом в пользу митоАФК опосредо
anion channels, VDACs), что может связывать
ванного механизма является повышенный ответ
АФК зависимую передачу сигнала и транспорт
АФК чувствительных зондов при гипоксии [92,
метаболитов [113]. МитоАФК также передает
131, 134]. Эти результаты следует интерпретиро
сигнал о повреждениях через окисление мито
вать с осторожностью, особенно в отсутствии
хондриальной DNA, т.к. только окисленная
кинетических данных [92, 134]. Например, наб
DNA связывается с NLRP3 и активирует ее
людаемые обратимые изменения в флуоресцен
[89, 114]. В другом исследовании была выявлена
ции дихлорофлуоресцеина [131] трудно объяс
ключевая роль паркин опосредованной мито
нить. Ответ сайтов продукции АФК на измене
фагии в NF kB зависимом контроле избыточ
ния парциального давления кислорода был ис
ной активации NLRP3 инфламмасомы [67]. Это
следован в такой простой системе, как изолиро
и другие исследования [130] подчеркивают важ
ванные митохондрии. Результаты этих исследо
ность систем контроля качества в митохондри
ваний варьировали, но ни в одной из работ не
ях, интегрирующих митофагию, динамику ми
была обнаружена повышенная продукция АФК
тохондрий и биогенез этих органелл в развитии
в условиях гипоксии [135-137]. Было сделано
врожденного иммунного ответа и воспаления.
заключение, что ответы АФК на гипоксию в
клетках не являются внутренне присущим свой
ством митохондрий [136], и их происхождение
МИТОАФК И РЕГУЛЯЦИЯ HIF1
остается неизвестным [95]. Экзогенная пере
кись водорода, возможно, стабилизирует HIF1α
Хотя транскрипционный фактор HIF1 ши
чувствительным к каталазе способом [92], что в
роко вовлечен в ряд адаптивных процессов,
некоторой степени поддерживает идею о том,
включая иммунный ответ, идея об участии АФК
что в данный процесс могут быть вовлечены не
в регуляции его критически важной субъедини
митохондриальные АФК. Тем не менее базовый
цы HIF1α возникла при изучении адаптации
вопрос о механизме подавления гидроксилиро
клеток к физиологической гипоксии (0,3-3%
вания HIF1α АФК остается открытым.
кислорода). Было предположено, что в таких ус
ловиях стабилизация HIF1α критически зави
сит от сигнальных митоАФК, таким образом
НЕМИТОХОНДРИАЛЬНЫЕ АФК
приписывая митохондриям роль сенсоров кис
В ИММУНИТЕТЕ
лорода [131]. Субъединица HIF1α нестабильна
при нормальном содержании кислорода в клет
Основной темой этого обзора являются ми
ке из за быстрой деградации, инициируемой
тоАФК, однако обсуждение АФК зависимой
белком PDH из семейства α кетоглутарат зави
передачи сигнала было бы неполным без упоми
симых оксигеназ. Гидроксилирование повышает
нания других (иногда более значительных) ис
в 1000 раз аффинность HIF1α к убиквитин ли
точников АФК. В ответ на вторжение патогена,
газному комплексу, содержащему опухолевый
фагоцитирующие клетки секретируют огром
супрессор pVHL (von Hippel Lindau tumor sup
ные количества АФК. Этот процесс называется
pressor) как распознающий компонент. HIF1α
дыхательным взрывом и катализируется фер
подвергается быстрому убиквитинированию и
ментами из семейства NADРН оксидаз (NOX)
направляется для деградации протеасомой
(см. обзор [138], описывающий семь известных
[132]. Белки PDH являются хорошими сенсора
белков этого семейства). Основная NADРН ок
БИОХИМИЯ том 85 вып. 12 2020
1952
АНДРЕЕВ и др.
сидаза иммунного ответа, NOX2, представляет
цированный NOX, должен как либо отличаться
собой мультисубъединичный белковый комп
от сигнала, возникшего в митохондриях, т.к. на
лекс, который формируется на плазматической
иболее вероятно, что в обоих случаях такой
мембране клеток в строго регулируемом процес
АФК будет относительно стабильная перекись
се в ответ на внеклеточные сигналы. Фермент
водорода, H2O2, способная свободно проникать
использует внутриклеточные восстановитель
через мембрану. Относительный вклад этих двух
ные эквиваленты в виде NADРН для восстанов
источников был недавно оценен в нефагоцити
ления кислорода до супероксид аниона (кото
рующих клетках (миобластах). Хотя нет причи
рый, в свою очередь, дисмутирует в перекись во
ны ожидать, что эти клетки обладают повышен
дорода) с выбросом АФК во внеклеточную сре
ной активностью NOX, вклад этого фермента в
ду [138, 139]. В фагоцитирующих клетках (мак
продукцию АФК был значителен (~40%) [145].
рофагах и нейтрофилах) NOX2 продуцируемые
Сходный анализ источников АФК в различных
АФК играют основную роль в уничтожении ин
типах иммунных клеток позволит существенно
вазивных патогенов. Эти же АФК могут участво
расширить наши знания о роли АФК зависимой
вать в сигналах обратной связи. К примеру, они
передачи сигнала в иммунитете.
подавляют ответ Т клеток на активацию TCR в
И наоборот, митохондрии могут вносить
смешанных культурах нейтрофилов и Т клеток
вклад в окислительный взрыв и бактерицидную
[140]. В отсутствии фагоцитов внутриклеточные
активность, как было показано для макрофагов
NOX генерируемые АФК могут играть роль в
[146] и нейтрофилов [147]. Важно отметить, что
индуцированной активацией смерти Т клеток
при этом митохондрии специально привлека
[106]. Вдобавок к этим иммуносупрессорным
ются к сайтам бактерицидной активности в
механизмам АФК могут участвовать в актива
NOX2 содержащих фагосомах [146]. Возможно
ции Т клеток. Активация TCR вызывает внек
также существование положительной обратной
леточную генерацию супероксида, скорее всего,
связи между митохондриальной и NOX зависи
при участии NOX2. Удивительно, что этот про
мой продукцией АФК, что может приводить к
цесс не требуется для активации Т клеток, их
усилению бактерицидного действия [147-149].
пролиферации и продукции цитокинов [141].
На основе этих и других данных было сделано
заключение, что NOX2 необходим для правиль
ИНСТРУМЕНТЫ
ной дифференцировки в Th2 клетки и ухода от
Th17 фенотипа, но не для активации Т клеток.
Любые результаты хороши настолько, нас
Анализ действия антиоксидантов привел к ги
колько хороши использованные в исследовани
потезе, что реально действующими молекулами
ях методы. Ниже мы приводим примеры неко
при активации Т клеток являются растворимые
торых ключевых методов, применяемых в изуче
в липидах продукты перекисного окисления или
нии АФК и метаболических сигнальных процес
их ферментативно образованные аналоги (лей
сов.
котриены), но не АФК как таковые [106].
АФК зонды. Некоторые из методов оценки
АФК, образующиеся при функционирова
скорости продукции и конечных уровней АФК,
нии NOX, также могут участвовать в активации
их недостатки и потенциальные ошибки уже об
инфламмасомы NLRP3 в макрофагах [142], что
суждались в нашем предыдущем обзоре, посвя
было подтверждено несколькими линиями до
щенном АФК и использованию дихлорофлуо
казательств. Даже если специфичность ингиби
ресцеина и его эфирного аналога (H2 DCF DA
торов NOX можно подвергнуть сомнению [143],
или DCF DA) [11]. Однако в последние годы
сложность NOX опосредованных ответов,
широкое признание получило положительно за
включающих транслокацию и сборку множест
ряженное производное дигидроэтидия, полу
венных субъединиц, предоставляет свидетель
ченное его конъюгацией с трифенилфосфонием
ства в пользу предложенной модели, как было
(MitoSOX), которое используются как специфи
показано в генетических экспериментах и ис
ческий сенсор супероксида в митохондриях.
следовании по локализации белковых компо
Следует отметить, что такие зонды подвержены
нентов [142]. Активация инфламмасомы в ответ
таким же ограничениям, как и остальные ΔΨ
на частицы также соответствует модели фагоци
зависимые флуоресцентные индикаторы
тоз индуцируемой активации NOX и продукции
[150, 151]. Флуоресценция MitoSOX зависит не
АФК, сходной с дыхательным взрывом [144]. Та
только от активности АФК, но и от обратимой
ким образом, NOX представляется вполне веро
компартментализации в митохондриальном
ятной альтернативой митохондриям в качестве
матриксе, цитозоле и внеклеточной среде, кото
источника АФК в клетках. Не существует оче
рая определяется ΔΨ на митохондриальной и
видной причины, почему сигнал АФК, проду
плазматической мембранах. Любые изменения в
БИОХИМИЯ том 85 вып. 12 2020
АФК ЗАВИСИМАЯ ТРАНСДУКЦИЯ СИГНАЛОВ В ИММУНИТЕТЕ
1953
ΔΨ приведут к изменениям в концентрации и
перекисью водорода, которая легко проникает
флуоресценции зонда в матриксе и цитозоле.
через мембраны, факт защиты клеток с по
Другим фактором может быть разное связыва
мощью этих антиоксидантов является весьма
ние зонда с ядерной и митохондриальной DNA
спорным указателем происхождения АФК. Оба
[151] (связывание MitoSOX с DNA многократно
соединения были разработаны как каталитичес
увеличивает его флуоресценцию). Кроме того,
кие антиоксиданты [159, 160], т.к. являются
зонд может разобщать митохондрии и ингиби
структурными аналогами природного редокс
ровать оксидазу цитохрома с (комплекс IV) в
переносчика кофермента Q, который подверга
очень низких концентрациях [152], что делает
ется циклическому окислению/восстановлению
необходимым контроль и учет этих параметров
в дыхательной цепи. MitoQ больше похож на ко
при интерпретации экспериментальных дан
фермент Q, в то время как структура SkQ была
ных. Существование более чем одного флуорес
оптимизирована для повышения реактивности и
центного продукта превращает количественную
способности проникать через мембрану
оценку АФК с использованием зонда в качест
[160, 161]. Способность SkQ восстанавливаться
венную, если только продукты реакции не ана
комплексами I и II и таким образом регенериро
лизируются методом высокоэффективной жид
вать была показана в статье Skulachev [160]. Нуж
костной хроматографии [153]. Хотя абсолютно
но отметить, что как MitoQ, так и SkQ в микро
надежных зондов для измерения АФК не суще
молярных концентрациях являются сильными
ствует, MitoSOX представляется одним из наи
прооксидантами [160], поэтому необходимо
более сложных в использовании зондов.
тщательно подбирать условия, чтобы избежать
Антиоксиданты. N ацетилцистеин. Хотя
перегрузки митохондрий этими соединениями.
NAC широко применяется как общий антиок
В субмикромолярных концентрациях MitoQ и
сидант, его, скорее всего, следует считать вос
SkQ проявляют антиоксидантные свойства, и те
становителем и умеренным протектором сульф
рапевтический диапазон концентраций SkQ ши
гидрильных групп. NAC также участвует в по
ре такового для MitoQ [160]. В целом SkQ предс
полнении пула глутатиона, но только в условиях
тавляется более перспективным соединением,
его значительного истощения [154, 155]. Меха
чем MitoQ, и, по нашему мнению, ему следует
низм нейтрализации АФК этим антиоксидан
отдавать предпочтение при выборе зондов для
том находится под вопросом, учитывая низкую
изучения окислительного стресса и АФК зави
константу скорости реакции с супероксидом
симой передачи сигнала. С другой стороны, лю
[156-158]. NAC слабо реагирует с H2O2 и супе
бые выводы об источнике происхождения АФК,
роксидом [157]. Взаимодействие NAC с супе
сделанные на основе действия этих оксидантов,
роксидом представляет собой цепную реакция,
будут, в лучшем случае, спорными.
в которой супероксид регенерируется. Реакция
Эфиры метаболитов. Так как многие иммуно
завершается с превращением всего супероксида
метаболиты не способны проникать через плаз
в другую разновидность АФК, H2O2 [157]. Мета
матическую мембрану, в исследованиях метабо
болические реакции, приводящие к образова
лизма иммунной системы обычно используются
нию более восстанавливающих низкомолеку
их алкилированые производные [70, 163]. Одна
лярных соединений серы описаны в работе
ко не следует забывать, что эти производные яв
Ezerina et al. [158], но их значение для детокси
ляются самостоятельными соединениями с уни
кации АФК нуждается в подтверждении.
кальными химическими свойствами, а не прос
SkQ и MitoQ. Хинон MitoQ и в меньшей сте
то предшественниками исследуемых метаболи
пени его близкий структурный аналог, SkQ
тов. Например, диметилитаконат и диметилфу
[159, 160], способные специфически аккумули
марат гораздо легче формируют аддукты Михаэ
роваться митохондриями, широко используются
ля с белками, чем итаконат и фумарат соответ
как специфические антиоксиданты для внутри
ственно [58]. С другой стороны, диметилитако
митохондриальных АФК [159]. Следует отме
нат не конвертируется в итаконат при попада
тить, что пространственная селективность этих
нии в клетку, так что его использование теряют
компонентов постулируется на основании сооб
смысл [164]. Было предложено использовать 4
ражений их близкого расположения к источни
октилитаконат [58], но это соединение оказыва
кам митохондриальных АФК и компартментали
ет совершенно другое действие на индуцируе
зации [161]. С другой стороны, SkQ and MitoQ
мый частицами иммунный ответ макрофагов
являются эффективными нейтрализаторами
костного мозга, чем итаконат. Авторы отметили
(scavengers) свободных радикалов в мембранах,
существенную разницу между действием эндо
включая те, которые генерируются из экзоген
генного итаконата и экзогенного 4 октил эфира
ного пероксида [162]. Принимая во внимание,
[163]. Добавление диметилсукцината к BMDM
что большинство клеточных АФК представлено
не вызывало увеличения количества сукцината
15 БИОХИМИЯ том 85 вып. 12 2020
1954
АНДРЕЕВ и др.
в этих клетках [70]. Отметим, что использование
чить обсуждение биоэнергетических аспектов
эфиров в случае макрофагов может быть не нуж
метаболизма иммунной системы в предлагае
ным, т.к. концентрация итаконата в этих клет
мый обзор об АФК зависимой передаче сигна
ках может быть повышена до миллимолярных
ла. Представляется, что наиболее безопасным
значений путем их длительной инкубации с не
способом АФК зависимой передачи сигнала,
модифицированным соединением [51]. Кроме
предотвращающим развитие окислительного
того, благодаря фагоцитарным свойствам мак
стресса, является фоновая генерация АФК. При
рофагов возможна эффективная доставка в
стимуляции она усиливается за счет биогенеза
клетку таких канонических непроникающих
митохондрий, а не за счет быстрого метаболи
молекул, как кофермент A и карбоксиатракти
ческого переключения. Соответственно, биоэ
лозид [84]. Очевидно, работа с Т клетками и
негетические процессы в клетке поддерживают
другими иммунными клетками может потребо
ся на том же уровне или активируются для осу
вать использования иных стратегий. В целом
ществления энергозатратных функций, напри
следовало бы следить за изменениями в уровнях
мер таких, как пролиферация. Тем не менее,
требуемых метаболитов при использовании их
когда макрофаги (клетки «часовые») обнаружи
производных и контролировать эффекты от
вают патоген «нарушитель», производство ци
щепляемых групп (которые конвертируются в
тотоксических молекул (NO и H2O2) становится
метанол или этанол в случае метил и этил про
более важным, чем обеспечение энергетической
изводных метаболитов соответственно). Полез
эффективности клеток, что приводит к их пов
ной стратегией являлось бы использование не
реждению и подавлению биоэнергетики.
гидролизуемых аналогов в качестве контроля,
В данном обзоре мы постарали осветить не
как это делается для различия энзиматических и
которые спорные механизмы (например, ОЭТ)
не энзиматических эффектов АТР.
не столько с целью критики, сколько для того,
чтобы привлечь интерес читателей к этим воп
росам. Рассмотренная область исследований, на
ЗАКЛЮЧЕНИЕ
перекрестье метаболизма и сигналов АФК, пол
на загадок и ошеломительных парадоксов. Она
Принципиальная разница между АФК зави
представляет собой поле непаханое для исследо
симой передачей сигнала и окислительным
вателей, готовых посвятить себя раскрытию де
стрессом является большей частью количест
талей невероятно запутанных и чрезвычайно
венной: «умеренная» продукция АФК участвует
важных механизмов иммунитета.
в регуляции сигнальных путей, в то время как
избыточная генерация АФК переполняет ем
кости систем антиоксидантной защиты и пов
Финансирование. Эта работа была частично
реждает клетки. На некотором промежуточном
поддержана National Institute of Health
уровне АФК могут инициировать механизмы,
(P01AG014930/AG/NIA NIH HSS).
приводящие к самодеструкции клетки в резуль
Конфликт интересов. Авторы заявляют об от
тате апоптоза.
сутствии какого либо конфликта интересов.
Крутая зависимость продукции АФК от ре
Соблюдение этических норм. Данная статья
докс статуса переносчиков в ЭТЦ указывает на
не содержит описания исследований, проводи
ее прямую связь с метаболическим состоянием
мых авторами с участием людей или животных
клеток. Это соображение побудило нас вклю
моделей.
СПИСОК ЛИТЕРАТУРЫ
1.
Kushnareva, Y., Murphy, A. N., and Andreyev, A. (2002)
4.
Brand, M. D. (2010) The sites and topology of mitochon
Complex I mediated reactive oxygen species generation:
drial superoxide production, Exp. Gerontol., 45, 466 472,
modulation by cytochrome c and NAD(P)+ oxidation
doi: 10.1016/j.exger.2010.01.003.
reduction state, Biochem. J., 368, 545 553, doi: 10.1042/
5.
Chouchani, E. T., Pell, V. R., Gaude, E., Aksentijevic, D.,
BJ20021121.
Sundier, S. Y., et al. (2014) Ischaemic accumulation of suc
2.
Andreyev, A. Y., Kushnareva, Y. E., and Starkov, A. A.
cinate controls reperfusion injury through mitochondrial
(2005) Mitochondrial metabolism of reactive oxygen
ROS, Nature, 515, 431 435, doi: 10.1038/nature13909.
species, Biochemistry (Moscow), 70, 200 214, doi: 10.1007/
6.
Niatsetskaya, Z. V., Sosunov, S. A., Matsiukevich, D.,
s10541 005 0102 7.
Utkina Sosunova, I. V., Ratner, V. I., Starkov, A. A., and
3.
Scialo, F., Fernandez Ayala, D. J., and Sanz, A. (2017)
Ten, V. S. (2012) The oxygen free radicals originating from
Role of mitochondrial reverse electron transport in ROS
mitochondrial complex I contribute to oxidative brain injury
signaling: potential roles in health and disease, Front.
following hypoxia ischemia in neonatal mice, J. Neurosci.,
Physiol., 8, 428, doi: 10.3389/fphys.2017.00428.
32, 3235 3244, doi: 10.1523/JNEUROSCI.6303 11.2012.
БИОХИМИЯ том 85 вып. 12 2020
АФК ЗАВИСИМАЯ ТРАНСДУКЦИЯ СИГНАЛОВ В ИММУНИТЕТЕ
1955
7.
Sahni, P. V., Zhang, J., Sosunov, S., Galkin, A.,
bolic requirements of cell proliferation, Science, 324, 1029
Niatsetskaya, Z., Starkov, A., Brookes, P. S., and Ten, V. S.
1033, doi: 10.1126/science.1160809.
(2018) Krebs cycle metabolites and preferential succinate
25.
Hume, D. A., Radik, J. L., Ferber, E., and Weidemann,
oxidation following neonatal hypoxic ischemic brain
M. J. (1978) Aerobic glycolysis and lymphocyte transfor
injury in mice, Pediatr. Res., 83, 491 497, doi: 10.1038/
mation, Biochem. J.,
174,
703709, doi:
10.1042/
pr.2017.277.
bj1740703.
8.
Boveris, A., and Chance, B. (1973) The mitochondrial
26.
Wang, A., Luan, H. H., and Medzhitov, R. (2019) An evo
generation of hydrogen peroxide. General properties and
lutionary perspective on immunometabolism, Science,
effect of hyperbaric oxygen, Biochem. J., 134, 707 716,
363, doi: 10.1126/science.aar3932.
doi: 10.1042/bj1340707.
27.
Chang, C. H., Curtis, J. D., Maggi, L. B., Jr., Faubert, B.,
9.
Boveris, A., Oshino, N., and Chance, B. (1972) The cellu
Villarino, A. V., et al. (2013) Posttranscriptional control of
lar production of hydrogen peroxide, Biochem. J., 128,
T cell effector function by aerobic glycolysis, Cell, 153,
617 630, doi: 10.1042/bj1280617.
1239 1251, doi: 10.1016/j.cell.2013.05.016.
10.
Zorov, D. B., Juhaszova, M., and Sollott, S. J. (2014)
28.
Peng, M., Yin, N., Chhangawala, S., Xu, K., Leslie, C. S.,
Mitochondrial reactive oxygen species (ROS) and ROS
and Li, M. O. (2016) Aerobic glycolysis promotes T helper
induced ROS release, Physiol. Rev.,
94,
909950,
1 cell differentiation through an epigenetic mechanism,
doi: 10.1152/physrev.00026.2013.
Science, 354, 481 484, doi: 10.1126/science.aaf6284.
11.
Andreyev, A. Y., Kushnareva, Y. E., Murphy, A. N., and
29.
Zhang, D., Tang, Z., Huang, H., Zhou, G., Cui, C., et al.
Starkov, A. A. (2015) Mitochondrial ROS metabolism: 10
(2019) Metabolic regulation of gene expression by histone
years later, Biochemistry (Moscow),
80,
517531,
lactylation, Nature, 574, 575 580, doi: 10.1038/s41586
doi: 10.1134/S0006297915050028.
019 1678 1.
12.
Starkov, A. A., Fiskum, G., Chinopoulos, C., Lorenzo, B.
30.
Phan, A. T., Goldrath, A. W., and Glass, C. K. (2017)
J., Browne, S. E., Patel, M. S., and Beal, M. F. (2004)
Metabolic and epigenetic coordination of T cell and
Mitochondrial alpha ketoglutarate dehydrogenase com
macrophage immunity, Immunity,
46,
714729,
plex generates reactive oxygen species, J. Neurosci., 24,
doi: 10.1016/j.immuni.2017.04.016.
7779 7788, doi: 10.1523/JNEUROSCI.1899 04.2004.
31.
Millet, P., Vachharajani, V., McPhail, L., Yoza, B., and
13.
Kareyeva, A. V., Grivennikova, V. G., Cecchini, G., and
McCall, C. E. (2016) GAPDH binding to TNF alpha
Vinogradov, A. D. (2011) Molecular identification of the
mRNA contributes to posttranscriptional repression in
enzyme responsible for the mitochondrial NADH sup
monocytes: a novel mechanism of communication between
ported ammonium dependent hydrogen peroxide produc
inflammation and metabolism, J. Immunol., 196, 2541
tion, FEBS Lett., 585, 385 389, doi: 10.1016/j.febslet.
2551, doi: 10.4049/jimmunol.1501345.
2010.12.019.
32.
Colell, A., Green, D. R., and Ricci, J. E. (2009) Novel roles
14.
Buck, M. D., O’Sullivan, D., and Pearce, E. L. (2015) T
for GAPDH in cell death and carcinogenesis,
cell metabolism drives immunity, J. Exp. Med., 212, 1345
Cell Death Differ., 16, 1573 1581, doi: 10.1038/cdd.2009.137.
1360, doi: 10.1084/jem.20151159.
33.
Nagy, E., and Rigby, W. F. (1995) Glyceraldehyde 3 phos
15.
Almeida, L., Lochner, M., Berod, L., and Sparwasser, T.
phate dehydrogenase selectively binds AU rich RNA in the
(2016) Metabolic pathways in T cell activation and lineage
NAD+ binding region (Rossmann fold), J. Biol. Chem.,
differentiation, Semin. Immunol., 28, 514 524, doi: 10.1016/
270, 2755 2763, doi: 10.1074/jbc.270.6.2755.
j.smim.2016.10.009.
34.
Rodriguez Pascual, F., Redondo Horcajo, M., Magan
16.
Diskin, C., and Palsson McDermott, E. M.
(2018)
Marchal, N., Lagares, D., Martinez Ruiz, A.,
Metabolic modulation in macrophage effector function,
Kleinert, H., and Lamas, S. (2008) Glyceraldehyde 3
Front. Immunol., 9, 270, doi: 10.3389/fimmu.2018.00270.
phosphate dehydrogenase regulates endothelin 1 expres
17.
O’Neill, L. A., Kishton, R. J., and Rathmell, J. (2016) A
sion by a novel, redox sensitive mechanism involving
guide to immunometabolism for immunologists, Nat. Rev.
mRNA stability, Mol. Cell. Biol.,
28,
71397155,
Immunol., 16, 553 565, doi: 10.1038/nri.2016.70.
doi: 10.1128/MCB.01145 08.
18.
Ramalho, R., Rao, M., Zhang, C., Agrati, C., Ippolito, G.,
35.
Liberti, M. V., Dai, Z., Wardell, S. E., Baccile, J. A.,
Wang, F. S., Zumla, A., and Maeurer, M. (2020) Immuno
Liu, X., et al. (2017) A predictive model for selective tar
metabolism: new insights and lessons from antigen direct
geting of the Warburg effect through GAPDH inhibition
ed cellular immune responses, Semin. Immunopathol., 42,
with a natural product, Cell Metab., 26, 648 659 e648,
279 313, doi: 10.1007/s00281 020 00798 w.
doi: 10.1016/j.cmet.2017.08.017.
19.
Escoll, P., and Buchrieser, C. (2018) Metabolic reprogram
36.
Pathria, G., Scott, D. A., Feng, Y., Sang Lee, J., Fujita, Y.,
ming of host cells upon bacterial infection: why shift to a
et al. (2018) Targeting the Warburg effect via LDHA inhi
Warburg like metabolism? FEBS J., 285, 21462160,
bition engages ATF4 signaling for cancer cell survival,
doi: 10.1111/febs.14446.
EMBO J., 37, doi: 10.15252/embj.201899735.
20.
Wang, T., Marquardt, C., and Foker, J. (1976) Aerobic gly
37.
Yang, X., Xia, R., Yue, C., Zhai, W., Du, W., et al. (2018)
colysis during lymphocyte proliferation, Nature, 261, 702
ATF4 regulates CD4+ T cell immune responses through
705, doi: 10.1038/261702a0.
metabolic reprogramming, Cell Rep., 23, 17541766,
21.
Bennett, W. E., and Cohn, Z. A. (1966) The isolation and
doi: 10.1016/j.celrep.2018.04.032.
selected properties of blood monocytes, J. Exp. Med., 123,
38.
Dietl, K., Renner, K., Dettmer, K., Timischl, B.,
145 160, doi: 10.1084/jem.123.1.145.
Eberhart, K., et al. (2010) Lactic acid and acidification
22.
Gudmundsdottir, H., Wells, A. D., and Turka, L. A. (1999)
inhibit TNF secretion and glycolysis of human monocytes,
Dynamics and requirements of T cell clonal expansion
J. Immunol., 184, 1200 1209, doi: 10.4049/jimmunol.
in vivo at the single cell level: effector function is linked to
0902584.
proliferative capacity, J. Immunol., 162, 5212 5223.
39.
Haas, R., Cucchi, D., Smith, J., Pucino, V., Macdougall,
23.
Delmastro Greenwood, M. M., and Piganelli, J. D. (2013)
C. E., and Mauro, C. (2016) Intermediates of metabolism:
Changing the energy of an immune response, Am. J. Clin.
from bystanders to signalling molecules, Trends Biochem.
Exp. Immunol., 2, 30 54.
Sci., 41, 460 471, doi: 10.1016/j.tibs.2016.02.003.
24.
Vander Heiden, M. G., Cantley, L. C., and Thompson,
40.
Haas, R., Smith, J., Rocher Ros, V., Nadkarni, S.,
C. B. (2009) Understanding the Warburg effect: the meta
Montero Melendez, T., et al. (2015) Lactate regulates
БИОХИМИЯ том 85 вып. 12 2020
15*
1956
АНДРЕЕВ и др.
metabolic and pro inflammatory circuits in control of T
56.
Salkowski, C. A., Detore, G., McNally, R., van Rooijen,
cell migration and effector functions, PLoS Biol., 13,
N., and Vogel, S. N. (1997) Regulation of inducible nitric
e1002202, doi: 10.1371/journal.pbio.1002202.
oxide synthase messenger RNA expression and nitric oxide
41.
Metallo, C. M., and Vander Heiden, M. G. (2010)
production by lipopolysaccharide in vivo: the roles of
Metabolism strikes back: metabolic flux regulates cell sig
macrophages, endogenous IFN gamma, and TNF recep
naling, Genes Dev., 24, 2717 2722, doi: 10.1101/gad.
tor 1 mediated signaling, J. Immunol., 158, 905 912.
2010510.
57.
Strelko, C. L., Lu, W., Dufort, F. J., Seyfried, T. N., Chiles,
42.
Weinberg, S. E., Sena, L. A., and Chandel, N. S. (2015)
T. C., Rabinowitz, J. D., and Roberts, M. F.
(2011)
Mitochondria in the regulation of innate and adaptive
Itaconic acid is a mammalian metabolite induced during
immunity, Immunity, 42, 406 417, doi: 10.1016/j.immuni.
macrophage activation, J. Am. Chem. Soc., 133, 16386
2015.02.002.
16389, doi: 10.1021/ja2070889.
43.
Palmieri, E. M., Spera, I., Menga, A., Infantino, V.,
58.
Mills, E. L., Ryan, D. G., Prag, H. A., Dikovskaya, D.,
Porcelli, V., et al. (2015) Acetylation of human mitochon
Menon, D., et al. (2018) Itaconate is an anti inflammato
drial citrate carrier modulates mitochondrial citrate/
ry metabolite that activates Nrf2 via alkylation of KEAP1,
malate exchange activity to sustain NADPH production
Nature, 556, 113 117, doi: 10.1038/nature25986.
during macrophage activation, Biochim. Biophys. Acta,
59.
Newton, R., Priyadharshini, B., and Turka, L. A. (2016)
1847, 729 738, doi: 10.1016/j.bbabio.2015.04.009.
Immunometabolism of regulatory T cells, Nat. Immunol.,
44.
Williams, N. C., and O’Neill, L. A. J. (2018) A role for the
17, 618 625, doi: 10.1038/ni.3466.
Krebs cycle intermediate citrate in metabolic reprogram
60.
Lindsten, T., June, C. H., and Thompson, C. B. (1988)
ming in innate immunity and inflammation, Front.
Multiple mechanisms regulate c myc gene expression dur
Immunol., 9, 141, doi: 10.3389/fimmu.2018.00141.
ing normal T cell activation, EMBO J., 7, 2787 2794.
45.
Loftus, R. M., and Finlay, D. K. (2016) Immunometa
61.
Weichhart, T., Hengstschlager, M., and Linke, M. (2015)
bolism: cellular metabolism turns immune regulator,
Regulation of innate immune cell function by mTOR, Nat.
J. Biol. Chem., 291, 1 10, doi: 10.1074/jbc.R115.693903.
Rev. Immunol., 15, 599 614, doi: 10.1038/nri3901.
46.
Chinopoulos, C. (2020) Acute sources of mitochondrial
62.
Saravia, J., Raynor, J. L., Chapman, N. M., Lim, S. A.,
NAD+ during respiratory chain dysfunction, Exp. Neurol.,
and Chi, H. (2020) Signaling networks in immunometabo
327, 113218, doi: 10.1016/j.expneurol.2020.113218.
lism, Cell Res., 30, 328 342, doi: 10.1038/s41422 020
47.
Hillgartner, F. B., Salati, L. M., and Goodridge, A. G.
0301 1.
(1995) Physiological and molecular mechanisms involved
63.
Muller, M. R., and Rao, A. (2010) NFAT, immunity and
in nutritional regulation of fatty acid synthesis, Physiol.
cancer: a transcription factor comes of age, Nat. Rev.
Rev., 75, 47 76, doi: 10.1152/physrev.1995.75.1.47.
Immunol., 10, 645 656, doi: 10.1038/nri2818.
48.
Tannahill, G. M., Curtis, A. M., Adamik, J., Palsson
64.
Seo, H., Chen, J., Gonzalez Avalos, E., Samaniego
McDermott, E. M., McGettrick, A. F., et al. (2013) Succi
Castruita, D., Das, A., et al. (2019) TOX and TOX2 tran
nate is an inflammatory signal that induces IL 1beta
scription factors cooperate with NR4A transcription fac
through HIF 1alpha, Nature, 496, 238 242, doi: 10.1038/
tors to impose CD8+ T cell exhaustion, Proc. Natl. Acad.
nature11986.
Sci. USA,
116,
1241012415, doi:
10.1073/pnas.
49.
Gill, T., and Levine, A. D. (2013) Mitochondria derived
1905675116.
hydrogen peroxide selectively enhances T cell receptor ini
65.
Wang, R., Dillon, C. P., Shi, L. Z., Milasta, S., Carter, R.,
tiated signal transduction, J. Biol. Chem., 288, 26246
et al. (2011) The transcription factor Myc controls meta
26255, doi: 10.1074/jbc.M113.476895.
bolic reprogramming upon T lymphocyte activation,
50.
Selak, M. A., Armour, S. M., MacKenzie, E. D.,
Immunity,
35,
871882, doi:
10.1016/j.immuni.
Boulahbel, H., Watson, D. G., et al. (2005) Succinate links
2011.09.021.
TCA cycle dysfunction to oncogenesis by inhibiting HIF
66.
Afonina, I. S., Zhong, Z., Karin, M., and Beyaert, R.
alpha prolyl hydroxylase, Cancer Cell,
7,
7785,
(2017) Limiting inflammation the negative regulation of
doi: 10.1016/j.ccr.2004.11.022.
NF kappaB and the NLRP3 inflammasome, Nat.
51.
Cordes, T., Wallace, M., Michelucci, A., Divakaruni, A. S.,
Immunol., 18, 861 869, doi: 10.1038/ni.3772.
Sapcariu, S. C., et al. (2016) Immunoresponsive gene 1 and
67.
Zhong, Z., Umemura, A., Sanchez Lopez, E., Liang, S.,
itaconate inhibit succinate dehydrogenase to modulate
Shalapour, S., et al. (2016) NF kappaB restricts inflamma
intracellular succinate levels, J. Biol. Chem., 291, 14274
some activation via elimination of damaged mitochondria,
14284, doi: 10.1074/jbc.M115.685792.
Cell, 164, 896 910, doi: 10.1016/j.cell.2015.12.057.
52.
Lampropoulou, V., Sergushichev, A., Bambouskova, M.,
68.
Jones, R. G., and Pearce, E. J. (2017) MenTORing immu
Nair, S., Vincent, E. E., et al. (2016) Itaconate links inhi
nity: mTOR signaling in the development and function of
bition of succinate dehydrogenase with macrophage
tissue resident immune cells, Immunity, 46, 730742,
metabolic remodeling and regulation of inflammation,
doi: 10.1016/j.immuni.2017.04.028.
Cell Metab., 24, 158 166, doi: 10.1016/j.cmet.2016.06.
69.
Codo, A. C., Davanzo, G. G., Monteiro, L. B., de Souza,
004.
G. F., Muraro, S. P., et al. (2020) Elevated glucose levels
53.
O’Neill, L. A. J., and Artyomov, M. N. (2019) Itaconate:
favor SARS CoV 2 infection and monocyte response
the poster child of metabolic reprogramming in
through a HIF 1alpha/glycolysis dependent axis, Cell
macrophage function, Nat. Rev. Immunol., 19, 273 281,
Metab., 32, 437446 e435, doi: 10.1016/j.cmet.2020.
doi: 10.1038/s41577 019 0128 5.
07.007.
54.
Michelucci, A., Cordes, T., Ghelfi, J., Pailot, A.,
70.
Mills, E. L., Kelly, B., Logan, A., Costa, A. S. H.,
Reiling, N., et al. (2013) Immune responsive gene 1 pro
Varma, M., et al. (2016) Succinate dehydrogenase supports
tein links metabolism to immunity by catalyzing itaconic
metabolic repurposing of mitochondria to drive inflamma
acid production, Proc. Natl. Acad. Sci. USA, 110, 7820
tory macrophages, Cell, 167, 457 470 e413, doi: 10.1016/
7825, doi: 10.1073/pnas.1218599110.
j.cell.2016.08.064.
55.
Dennis, E. A., Deems, R. A., Harkewicz, R.,
71.
Van den Bossche, J., Baardman, J., Otto, N. A., van der
Quehenberger, O., Brown, H. A., et al. (2010) A mouse
Velden, S., Neele, A. E., et al. (2016) Mitochondrial dysfunc
macrophage lipidome, J. Biol. Chem., 285, 39976 39985,
tion prevents repolarization of inflammatory macrophages,
doi: 10.1074/jbc.M110.182915.
Cell Rep., 17, 684 696, doi: 10.1016/j.celrep.2016.09.008.
БИОХИМИЯ том 85 вып. 12 2020
АФК ЗАВИСИМАЯ ТРАНСДУКЦИЯ СИГНАЛОВ В ИММУНИТЕТЕ
1957
72.
Stuehr, D. J., and Nathan, C. F. (1989) Nitric oxide. A
87. Loschen, G., Flohe, L., and Chance, B.
(1971)
macrophage product responsible for cytostasis and respira
Respiratory chain linked H2O2 production in pigeon heart
tory inhibition in tumor target cells, J. Exp. Med., 169,
mitochondria, FEBS Lett., 18, 261 264, doi: 10.1016/
1543 1555, doi: 10.1084/jem.169.5.1543.
0014 5793(71)80459 3.
73.
Brown, G. C., and Borutaite, V. (1999) Nitric oxide,
88. Korshunov, S. S., Skulachev, V. P., and Starkov, A. A.
cytochrome c and mitochondria, Biochem. Soc. Symp., 66,
(1997) High protonic potential actuates a mechanism of
17 25, doi: 10.1042/bss0660017.
production of reactive oxygen species in mitochondria,
74.
Clementi, E., Brown, G. C., Feelisch, M., and
FEBS Lett.,
416,
1518, doi:
10.1016/s0014
Moncada, S. (1998) Persistent inhibition of cell respira
5793(97)01159 9.
tion by nitric oxide: crucial role of S nitrosylation of mito
89. Zhong, Z., Liang, S., Sanchez Lopez, E., He, F.,
chondrial complex I and protective action of glutathione,
Shalapour, S., et al. (2018) New mitochondrial DNA syn
Proc. Natl. Acad. Sci. USA, 95, 7631 7636, doi: 10.1073/
thesis enables NLRP3 inflammasome activation, Nature,
pnas.95.13.7631.
560, 198 203, doi: 10.1038/s41586 018 0372 z.
75.
Palmieri, E. M., Gonzalez Cotto, M., Baseler, W. A.,
90. Jazayeri, M., Andreyev, A., Will, Y., Ward, M., Anderson, C. M.,
Davies, L. C., Ghesquiere, B., et al. (2020) Nitric oxide
and Clevenger, W. (2003) Inducible expression of a domi
orchestrates metabolic rewiring in M1 macrophages by tar
nant negative DNA polymerase gamma depletes mito
geting aconitase 2 and pyruvate dehydrogenase, Nat.
chondrial DNA and produces a rho0 phenotype, J. Biol.
Commun., 11, 698, doi: 10.1038/s41467 020 14433 7.
Chem., 278, 9823 9830, doi: 10.1074/jbc.m211730200.
76.
Mookerjee, S. A., Gerencser, A. A., Nicholls, D. G., and
91. Martinez Reyes, I., Diebold, L. P., Kong, H.,
Brand, M. D. (2017) Quantifying intracellular rates of gly
Schieber, M., Huang, H., et al. (2016) TCA cycle and
colytic and oxidative ATP production and consumption
mitochondrial membrane potential are necessary for
using extracellular flux measurements, J. Biol. Chem., 292,
diverse biological functions, Mol. Cell, 61, 199209,
7189 7207, doi: 10.1074/jbc.M116.774471.
doi: 10.1016/j.molcel.2015.12.002.
77.
Nicholls, D. G., Ferguson, S. J. (2001) Bioenergetics 3,
92. Bell, E. L., Klimova, T. A., Eisenbart, J., Moraes, C. T.,
Academic Press, San Diego.
Murphy, M. P., Budinger, G. R., and Chandel, N. S.
78.
Lehninger, A. L. (1976) Biochemistry, 2nd edition, Worth
(2007) The Qo site of the mitochondrial complex III is
Publishers Inc., New York, N.Y.
required for the transduction of hypoxic signaling via reac
79.
Divakaruni, A. S., Wallace, M., Buren, C., Martyniuk, K.,
tive oxygen species production, J. Cell Biol., 177, 1029
Andreyev, A. Y., et al. (2017) Inhibition of the mitochondr
1036, doi: 10.1083/jcb.200609074.
ial pyruvate carrier protects from excitotoxic neuronal
93. Son, Y., Kim, S., Chung, H. T., and Pae, H. O. (2013)
death, J. Cell Biol., 216, 1091 1105, doi: 10.1083/jcb.
Reactive oxygen species in the activation of MAP kinases,
201612067.
Methods Enzymol., 528, 27 48, doi: 10.1016/B978 0 12
80.
Wang, A., Huen, S. C., Luan, H. H., Yu, S., Zhang, C.,
405881 1.00002 1.
Gallezot, J. D., Booth, C. J., and Medzhitov, R. (2016)
94. Dard, L., Blanchard, W., Hubert, C., Lacombe, D., and
Opposing effects of fasting metabolism on tissue tolerance
Rossignol, R. (2020) Mitochondrial functions and rare dis
in bacterial and viral inflammation, Cell, 166,
1512
eases, Mol. Aspects Med., 71, 100842, doi: 10.1016/j.mam.
1525.e1512, doi: 10.1016/j.cell.2016.07.026.
2019.100842.
81.
Wang, A., Pope, S. D., Weinstein, J. S., Yu, S., Zhang, C.,
95. Sena, L. A., and Chandel, N. S. (2012) Physiological roles
Booth, C. J., and Medzhitov, R. (2019) Specific sequences
of mitochondrial reactive oxygen species, Mol. Cell, 48,
of infectious challenge lead to secondary hemophago
158 167, doi: 10.1016/j.molcel.2012.09.025.
cytic lymphohistiocytosis like disease in mice, Proc. Natl.
96. Franchina, D. G., Dostert, C., and Brenner, D. (2018)
Acad. Sci. USA, 116, 22002209, doi: 10.1073/pnas.
Reactive oxygen species: involvement in T cell signaling
1820704116.
and metabolism, Trends Immunol.,
39,
489502,
82.
Vats, D., Mukundan, L., Odegaard, J. I., Zhang, L.,
doi: 10.1016/j.it.2018.01.005.
Smith, K. L., et al. (2006) Oxidative metabolism and
97. Flohe, L., Brigelius Flohe, R., Saliou, C., Traber, M. G.,
PGC 1beta attenuate macrophage mediated inflamma
and Packer, L. (1997) Redox regulation of NF kappa B
tion, Cell Metab., 4, 13 24, doi: 10.1016/j.cmet.2006.
activation, Free Radic. Biol. Med.,
22,
11151126,
05.011.
doi: 10.1016/s0891 5849(96)00501 1.
83.
Huang, S. C., Everts, B., Ivanova, Y., O’Sullivan, D.,
98. Liu, T., Zhang, L., Joo, D., and Sun, S. C. (2017) NF
Nascimento, M., et al. (2014) Cell intrinsic lysosomal
kappaB signaling in inflammation, Signal Transduct.
lipolysis is essential for alternative activation of
Target. Ther., 2, doi: 10.1038/sigtrans.2017.23.
macrophages, Nat. Immunol., 15, 846 855, doi: 10.1038/
99. Son, Y., Cheong, Y. K., Kim, N. H., Chung, H. T., Kang,
ni.2956.
D. G., and Pae, H. O. (2011) Mitogen activated protein
84.
Divakaruni, A. S., Hsieh, W. Y., Minarrieta, L., Duong,
kinases and reactive oxygen species: how can ROS activate
T. N., Kim, K. K. O., et al. (2018) Etomoxir inhibits
MAPK pathways? J. Signal Transduct., 2011, 792639,
macrophage polarization by disrupting CoA homeostasis,
doi: 10.1155/2011/792639.
Cell Metab., 28, 490 503.e497, doi: 10.1016/j.cmet.2018.
100. Chandel, N. S., Vander Heiden, M. G., Thompson, C. B.,
06.001.
and Schumacker, P. T. (2000) Redox regulation of p53 dur
85.
Tan, Z., Xie, N., Cui, H., Moellering, D. R., Abraham, E.,
ing hypoxia, Oncogene, 19, 3840 3848, doi: 10.1038/sj.onc.
Thannickal, V. J., and Liu, G. (2015) Pyruvate dehydroge
1203727.
nase kinase 1 participates in macrophage polarization via
101. Liu, B., Chen, Y., and St Clair, D. K. (2008) ROS and p53:
regulating glucose metabolism, J. Immunol., 194, 6082
a versatile partnership, Free Radic. Biol. Med., 44, 1529
6089, doi: 10.4049/jimmunol.1402469.
1535, doi: 10.1016/j.freeradbiomed.2008.01.011.
86.
Faris, R., Fan, Y. Y., De Angulo, A., Chapkin, R. S.,
102. Nathan, C., and Cunningham Bussel, A. (2013) Beyond
deGraffenried, L. A., and Jolly, C. A.
(2014)
oxidative stress: an immunologist’s guide to reactive oxygen
Mitochondrial glycerol 3 phosphate acyltransferase 1 is
species, Nat. Rev. Immunol., 13, 349 361, doi: 10.1038/
essential for murine CD4+ T cell metabolic activation,
nri3423.
Biochim. Biophys. Acta, 1842, 1475 1482, doi: 10.1016/
103. Zhang, H., Wang, L., and Chu, Y. (2019) Reactive oxygen
j.bbalip.2014.07.009.
species: the signal regulator of B cell, Free Radic. Biol.
БИОХИМИЯ том 85 вып. 12 2020
1958
АНДРЕЕВ и др.
Med., 142, 1622, doi:
10.1016/j.freeradbiomed.2019.
tion of type I interferons by mitochondrial DNA, Cell, 159,
06.004.
1563 1577, doi: 10.1016/j.cell.2014.11.037.
104. Sena, L. A., Li, S., Jairaman, A., Prakriya, M.,
120. Liu, X., Kim, C. N., Yang, J., Jemmerson, R., and
Ezponda, T., et al. (2013) Mitochondria are required for
Wang, X. (1996) Induction of apoptotic program in cell
antigen specific T cell activation through reactive oxygen
free extracts: requirement for dATP and cytochrome c,
species signaling, Immunity, 38, 225 236, doi: 10.1016/
Cell, 86, 147 157, doi: 10.1016/s0092 8674(00)80085 9.
j.immuni.2012.10.020.
121. Kluck, R. M., Bossy Wetzel, E., Green, D. R., and
105. Rashida Gnanaprakasam, J. N., Wu, R., and Wang, R.
Newmeyer, D. D. (1997) The release of cytochrome c from
(2018) Metabolic reprogramming in modulating T cell
mitochondria: a primary site for Bcl 2 regulation of apop
reactive oxygen species generation and antioxidant capaci
tosis, Science, 275, 11321136, doi: 10.1126/science.
ty, Front. Immunol.,
9,
1075, doi:
10.3389/fimmu.
275.5303.1132.
2018.01075.
122. Kushnareva, Y., and Newmeyer, D. D.
(2010)
106. Belikov, A. V., Schraven, B., and Simeoni, L. (2015) T cells
Bioenergetics and cell death, Ann. N. Y. Acad. Sci., 1201,
and reactive oxygen species, J. Biomed. Sci., 22, 85,
50 57, doi: 10.1111/j.1749 6632.2010.05633.x.
doi: 10.1186/s12929 015 0194 3.
123. Vince, J. E., De Nardo, D., Gao, W., Vince, A. J.,
107. Previte, D. M., O’Connor, E. C., Novak, E. A., Martins,
Hall, C., et al. (2018) The mitochondrial apoptotic effec
C. P., Mollen, K. P., and Piganelli, J. D. (2017) Reactive
tors BAX/BAK activate caspase 3 and
7 to trigger
oxygen species are required for driving efficient and sus
NLRP3 inflammasome and caspase 8 driven IL 1beta
tained aerobic glycolysis during CD4+ T cell activation,
activation, Cell Rep., 25, 2339 2353.e2334, doi: 10.1016/
PLoS One, 12, e0175549, doi: 10.1371/journal.pone.
j.celrep.2018.10.103.
0175549.
124. Zorov, D. B. (1996) Mitochondrial damage as a source of
108. Reth, M. (2002) Hydrogen peroxide as second messenger
diseases and aging: a strategy of how to fight these, Biochim.
in lymphocyte activation, Nat. Immunol., 3, 1129 1134,
Biophys. Acta,
1275,
1015, doi:
10.1016/0005
doi: 10.1038/ni1202 1129.
2728(96)00042 4.
109. Devadas, S., Zaritskaya, L., Rhee, S. G., Oberley, L., and
125. Patrushev, M., Kasymov, V., Patrusheva, V., Ushakova, T.,
Williams, M. S. (2002) Discrete generation of superoxide
Gogvadze, V., and Gaziev, A. (2004) Mitochondrial per
and hydrogen peroxide by T cell receptor stimulation:
meability transition triggers the release of mtDNA frag
selective regulation of mitogen activated protein kinase
ments, Cell. Mol. Life Sci., 61, 3100 3103, doi: 10.1007/
activation and fas ligand expression, J. Exp. Med., 195, 59
s00018 004 4424 1.
70, doi: 10.1084/jem.20010659.
126. Yu, C. H., Davidson, S., Harapas, C. R., Hilton, J. B.,
110. Kaminski, M. M., Sauer, S. W., Klemke, C. D., Suss, D.,
Mlodzianoski, M. J., et al. (2020) TDP 43 triggers mito
Okun, J. G., Krammer, P. H., and Gulow, K. (2010)
chondrial DNA release via mPTP to activate
Mitochondrial reactive oxygen species control T cell activa
cGAS/STING in ALS, Cell,
183,
636649 e618,
tion by regulating IL 2 and IL 4 expression: mechanism of
doi: 10.1016/j.cell.2020.09.020.
ciprofloxacin mediated immunosuppression, J. Immunol.,
127. Guo, W., Liu, W., Chen, Z., Gu, Y., Peng, S., et al. (2017)
184, 4827 4841, doi: 10.4049/jimmunol.0901662.
Tyrosine phosphatase SHP2 negatively regulates NLRP3
111.
Gong, T., Liu, L., Jiang, W., and Zhou, R. (2020) DAMP
inflammasome activation via ANT1 dependent mitochon
sensing receptors in sterile inflammation and inflammato
drial homeostasis, Nat. Commun., 8, 2168, doi: 10.1038/
ry diseases, Nat. Rev. Immunol., 20, 95 112, doi: 10.1038/
s41467 017 02351 0.
s41577 019 0215 7.
128. Kushnareva, Y. E., and Sokolove, P. M. (2000) Prooxidants
112. Meylan, E., Tschopp, J., and Karin, M.
(2006)
open both the mitochondrial permeability transition pore
Intracellular pattern recognition receptors in the host
and a low conductance channel in the inner mitochondri
response, Nature, 442, 39 44, doi: 10.1038/nature04946.
al membrane, Arch. Biochem. Biophys., 376, 377 388,
113. Zhou, R., Yazdi, A. S., Menu, P., and Tschopp, J. (2011) A
doi: 10.1006/abbi.2000.1730.
role for mitochondria in NLRP3 inflammasome activa
129. Ichas, F., Jouaville, L. S., and Mazat, J. P.
(1997)
tion, Nature, 469, 221 225, doi: 10.1038/nature09663.
Mitochondria are excitable organelles capable of generat
114. Shimada, K., Crother, T. R., Karlin, J., Dagvadorj, J.,
ing and conveying electrical and calcium signals, Cell, 89,
Chiba, N., et al. (2012) Oxidized mitochondrial DNA acti
1145 1153, doi: 10.1016/s0092 8674(00)80301 3.
vates the NLRP3 inflammasome during apoptosis,
130. Xu, Y., Shen, J., and Ran, Z. (2020) Emerging views of
Immunity, 36, 401 414, doi: 10.1016/j.immuni.2012.01.009.
mitophagy in immunity and autoimmune diseases,
115. Nakahira, K., Haspel, J. A., Rathinam, V. A., Lee, S. J.,
Autophagy, 16, 3 17, doi: 10.1080/15548627.2019.1603547.
Dolinay, T., et al. M. (2011) Autophagy proteins regulate
131. Chandel, N. S., Maltepe, E., Goldwasser, E., Mathieu,
innate immune responses by inhibiting the release of mito
C. E., Simon, M. C., and Schumacker, P. T. (1998)
chondrial DNA mediated by the NALP3 inflammasome,
Mitochondrial reactive oxygen species trigger hypoxia
Nat. Immunol., 12, 222 230, doi: 10.1038/ni.1980.
induced transcription, Proc. Natl. Acad. Sci. USA, 95,
116. Banoth, B., and Cassel, S. L. (2018) Mitochondria in
11715 11720, doi: 10.1073/pnas.95.20.11715.
innate immune signaling, Transl. Res., 202, 5268,
132. Schofield, C. J., and Ratcliffe, P. J. (2004) Oxygen sensing
doi: 10.1016/j.trsl.2018.07.014.
by HIF hydroxylases, Nat. Rev. Mol. Cell Biol., 5, 343 354,
117. West, A. P., Khoury Hanold, W., Staron, M., Tal, M. C.,
doi: 10.1038/nrm1366.
Pineda, C. M., et al. (2015) Mitochondrial DNA stress
133. Hirsila, M., Koivunen, P., Gunzler, V., Kivirikko, K. I.,
primes the antiviral innate immune response, Nature, 520,
and Myllyharju, J. (2003) Characterization of the human
553 557, doi: 10.1038/nature14156.
prolyl 4 hydroxylases that modify the hypoxia inducible
118. White, M. J., McArthur, K., Metcalf, D., Lane, R. M.,
factor, J. Biol. Chem., 278, 30772 30780, doi: 10.1074/
Cambier, J. C., et al. (2014) Apoptotic caspases suppress
jbc.M304982200.
mtDNA induced STING mediated type I IFN pro
134. Bell, E. L., Klimova, T. A., Eisenbart, J., Schumacker, P.
duction, Cell, 159, 1549 1562, doi: 10.1016/j.cell.2014.11.
T., and Chandel, N. S. (2007) Mitochondrial reactive oxy
036.
gen species trigger hypoxia inducible factor dependent
119. Rongvaux, A., Jackson, R., Harman, C. C., Li, T., West,
extension of the replicative life span during hypoxia, Mol.
A. P., et al. (2014) Apoptotic caspases prevent the induc
Cell. Biol., 27, 5737 5745, doi: 10.1128/MCB.02265 06.
БИОХИМИЯ том 85 вып. 12 2020
АФК ЗАВИСИМАЯ ТРАНСДУКЦИЯ СИГНАЛОВ В ИММУНИТЕТЕ
1959
135. Oshino, N., Jamieson, D., and Chance, B. (1975) The
151. Polster, B. M., Nicholls, D. G., Ge, S. X., and Roelofs, B.
properties of hydrogen peroxide production under hyper
A. (2014) Use of potentiometric fluorophores in the mea
oxic and hypoxic conditions of perfused rat liver,
surement of mitochondrial reactive oxygen species,
Biochem. J., 146, 53 65, doi: 10.1042/bj1460053.
Methods Enzymol., 547, 225 250, doi: 10.1016/B978 0 12
136. Hoffman, D. L., Salter, J. D., and Brookes, P. S. (2007)
801415 8.00013 8.
Response of mitochondrial reactive oxygen species gener
152. Roelofs, B. A., Ge, S. X., Studlack, P. E., and Polster,
ation to steady state oxygen tension: implications for
B. M. (2015) Low micromolar concentrations of the super
hypoxic cell signaling, Am. J. Physiol. Heart Circ. Physiol.,
oxide probe MitoSOX uncouple neural mitochondria and
292, H101 H108, doi: 10.1152/ajpheart.00699.2006.
inhibit complex IV, Free Radic. Biol. Med., 86, 250 258,
137. Stepanova, A., Konrad, C., Manfredi, G., Springett, R.,
doi: 10.1016/j.freeradbiomed.2015.05.032.
Ten, V., and Galkin, A. (2019) The dependence of brain
153. Zielonka, J., and Kalyanaraman, B. (2010) Hydroethi
mitochondria reactive oxygen species production on oxy
dine and MitoSOX derived red fluorescence is not a reli
gen level is linear, except when inhibited by antimycin A,
able indicator of intracellular superoxide formation: anoth
J. Neurochem., 148, 731 745, doi: 10.1111/jnc.14654.
er inconvenient truth, Free Radic. Biol. Med., 48, 983
138. Panday, A., Sahoo, M. K., Osorio, D., and Batra, S. (2015)
1001, doi: 10.1016/j.freeradbiomed.2010.01.028.
NADPH oxidases: an overview from structure to innate
154. Parasassi, T., Brunelli, R., Costa, G., De Spirito, M.,
immunity associated pathologies, Cell. Mol. Immunol., 12,
Krasnowska, E., Lundeberg, T., Pittaluga, E., and
5 23, doi: 10.1038/cmi.2014.89.
Ursini, F. (2010) Thiol redox transitions in cell signaling: a
139. Brown, D. I., and Griendling, K. K. (2009) Nox proteins
lesson from N acetylcysteine, ScientificWorldJournal, 10,
in signal transduction, Free Radic. Biol. Med., 47, 1239
1192 1202, doi: 10.1100/tsw.2010.104.
1253, doi: 10.1016/j.freeradbiomed.2009.07.023.
155. Samuni, Y., Goldstein, S., Dean, O. M., and Berk, M.
140. Cemerski, S., Cantagrel, A., Van Meerwijk, J. P., and
(2013) The chemistry and biological activities of N acetyl
Romagnoli, P. (2002) Reactive oxygen species differential
cysteine, Biochim. Biophys. Acta,
1830,
41174129,
ly affect T cell receptor signaling pathways, J. Biol. Chem.,
doi: 10.1016/j.bbagen.2013.04.016.
277, 19585 19593, doi: 10.1074/jbc.M111451200.
156. Benrahmoune, M., Therond, P., and Abedinzadeh, Z.
141. Belikov, A. V., Schraven, B., and Simeoni, L. (2014) TCR
(2000) The reaction of superoxide radical with N acetyl
triggered extracellular superoxide production is not
cysteine, Free Radic. Biol. Med.,
29,
775782,
required for T cell activation, Cell Commun. Signal., 12,
doi: 10.1016/s0891 5849(00)00380 4.
50, doi: 10.1186/s12964 014 0050 1.
157. Winterbourn, C. C., and Metodiewa, D. (1999) Reactivity
142. Martinon, F. (2010) Signaling by ROS drives inflamma
of biologically important thiol compounds with superoxide
some activation, Eur. J. Immunol.,
40,
616619,
and hydrogen peroxide, Free Radic. Biol. Med., 27, 322
doi: 10.1002/eji.200940168.
328, doi: 10.1016/s0891 5849(99)00051 9.
143. Aldieri, E., Riganti, C., Polimeni, M., Gazzano, E.,
158. Ezerina, D., Takano, Y., Hanaoka, K., Urano, Y., and
Lussiana, C., Campia, I., and Ghigo, D. (2008) Classical
Dick, T. P. (2018) N Acetyl cysteine functions as a fast
inhibitors of NOX NAD(P)H oxidases are not specific, Curr.
acting antioxidant by triggering intracellular H2S and sul
Drug Metab., 9, 686 696, doi: 10.2174/ 138920008786049285.
fane sulfur production, Cell Chem. Biol., 25, 447 459 e444,
144. Dostert, C., Petrilli, V., Van Bruggen, R., Steele, C.,
doi: 10.1016/j.chembiol.2018.01.011.
Mossman, B. T., and Tschopp, J. (2008) Innate immune
159. Kelso, G. F., Porteous, C. M., Coulter, C. V., Hughes, G.,
activation through Nalp3 inflammasome sensing of
Porteous, W. K., Ledgerwood, E. C., Smith, R. A., and
asbestos and silica, Science, 320, 674 677, doi: 10.1126/
Murphy, M. P. (2001) Selective targeting of a redox active
science.1156995.
ubiquinone to mitochondria within cells: antioxidant and
145. Wong, H. S., Benoit, B., and Brand, M. D. (2019)
antiapoptotic properties, J. Biol. Chem., 276, 4588 4596,
Mitochondrial and cytosolic sources of hydrogen peroxide
doi: 10.1074/jbc.M009093200.
in resting C2C12 myoblasts, Free Radic. Biol. Med., 130,
160. Skulachev, V. P. (2007) A biochemical approach to the
140 150, doi: 10.1016/j.freeradbiomed.2018.10.448.
problem of aging: “megaproject” on membrane penetrating
146. West, A. P., Brodsky, I. E., Rahner, C., Woo, D. K.,
ions. The first results and prospects, Biochemistry (Moscow),
Erdjument Bromage, H., et al. (2011) TLR signalling aug
72, 1385 1396, doi: 10.1134/s0006297907120139.
ments macrophage bactericidal activity through mitochon
161. Skulachev, V. P. (2013) Cationic antioxidants as a powerful
drial ROS, Nature,
472,
476480, doi:
10.1038/
tool against mitochondrial oxidative stress, Biochem.
nature09973.
Biophys. Res. Commun., 441, 275 279, doi: 10.1016/j.bbrc.
147. Vorobjeva, N., Prikhodko, A., Galkin, I., Pletjushkina, O.,
2013.10.063.
Zinovkin, R., Sud’ina, G., Chernyak, B., and Pinegin, B.
162. Antonenko, Y. N., Roginsky, V. A., Pashkovskaya, A. A.,
(2017) Mitochondrial reactive oxygen species are involved
Rokitskaya, T. I., Kotova, E. A., Zaspa, A. A., Chernyak,
in chemoattractant induced oxidative burst and degranula
B. V., and Skulachev, V. P. (2008) Protective effects of
tion of human neutrophils in vitro, Eur. J. Cell Biol., 96,
mitochondria targeted antioxidant SkQ in aqueous and
254 265, doi: 10.1016/j.ejcb.2017.03.003.
lipid membrane environments, J. Membr. Biol., 222, 141
148. Dikalov, S. (2011) Cross talk between mitochondria and
149, doi: 10.1007/s00232 008 9108 6.
NADPH oxidases, Free Radic. Biol. Med., 51, 1289 1301,
163. Sun, K. A., Li, Y., Meliton, A. Y., Woods, P. S., Kimmig,
doi: 10.1016/j.freeradbiomed.2011.06.033.
L. M., Cetin Atalay, R., Hamanaka, R. B., and Mutlu,
149. Pinegin, B., Vorobjeva, N., Pashenkov, M., and
G. M. (2020) Endogenous itaconate is not required for
Chernyak, B. (2018) The role of mitochondrial ROS in
particulate matter induced NRF2 expression or inflamma
antibacterial immunity, J. Cell. Physiol., 233, 3745 3754,
tory response, Elife, 9, doi: 10.7554/eLife.54877.
doi: 10.1002/jcp.26117.
164. ElAzzouny, M., Tom, C. T., Evans, C. R., Olson, L. L.,
150. Nicholls, D. G. (2012) Fluorescence measurement of
Tanga, M. J., Gallagher, K. A., Martin, B. R., and Burant,
mitochondrial membrane potential changes in cultured
C. F. (2017) Dimethyl itaconate is not metabolized into
cells, Methods Mol. Biol., 810, 119 133, doi: 10.1007/978
itaconate intracellularly, J. Biol. Chem., 292, 4766 4769,
1 61779 382 0_8.
doi: 10.1074/jbc.C117.775270.
БИОХИМИЯ том 85 вып. 12 2020
1960
АНДРЕЕВ и др.
METABOLIC ROS SIGNALING: TO IMMUNITY AND BEYOND
Review
A. Y. Andreyev1*, Y. E. Kushnareva2, N. N. Starkova3, and A. A. Starkov4
1 The Scripps Research Institute, 10550 La Jolla, CA 92037, USA; E mail: alex_andreyev@mitoexperts.com
2 La Jolla Institute for Allergy and Immunology, 9420 La Jolla, CA 92037, USA
3 State University of New York, Maritime College, New York, NY 10465, USA
4 Brain and Mind Research Institute, Weill Medical College of Cornell University,
Weill Medical College of Cornell University, New York, NY 10065, USA
Received September 11, 2020
Revised November 13, 2020
Accepted November 17, 2020
Metabolism is a critical determinant of immune cell functionality. Immunometabolism, by definition, is a multidis
ciplinary area of immunology research that integrates the knowledge of energy transduction mechanisms and bio
chemical pathways. An important concept in the field is metabolic switch, a transition of immune cells upon activa
tion to preferential utilization of select catabolic pathways for their energy needs. Mitochondria are not inert in this
process and contribute to the metabolic adaptation by different mechanisms which include increasing ATP produc
tion to match dynamic bioenergetic demands and serving as a signaling platform. The latter involves generation of
reactive oxygen species (ROS), one of the most intensively studied mitochondrial processes. While the role of mito
chondrial ROS in the context of oxidative stress is well established, ROS signaling in immunity is an emerging and
quickly changing field. In this review, we discuss ROS signaling and immunometabolism concepts from standpoint of
bioenergetics. We also provide a critical insight into the methodology for ROS assessment, outlining current chal
lenges in the field. Finally, based on our analysis of the literature data, we hypothesize that regulatory ROS produc
tion, as opposed to oxidative stress, is controlled by mitochondrial biogenesis rather than metabolic switches.
Keywords: immunometabolism, reactive oxygen species, glycolytic switch, macrophages, T cells, mitochondrial bio
genesis
БИОХИМИЯ том 85 вып. 12 2020