БИОХИМИЯ, 2020, том 85, вып. 2, с. 155 - 164
УДК 577.15
ГРАНЗИМЫ И МИТОХОНДРИИ
Обзор
© 2020
Д.Б. Киселевский
Московский государственный университет им. М.В. Ломоносова, биологический факультет,
119991 Москва, Россия; электронная почта: dkiselevs@mail.ru
Поступила в редакцию 01.10.2019
После доработки 01.11.2019
Принята к публикации 04.11.2019
Цитотоксические Т лимфоциты и естественные киллеры избавляют организм от зараженных клеток путем
включения в них программы гибели (апоптоза). Это может происходить в результате высвобождения содер
жимого литических гранул клеток киллеров, в которых локализованы порообразующие белки перфорины
и протеолитические ферменты гранзимы, и их последующего проникновения в клетки мишени. Гранзим B
способен инициировать зависимый от митохондрий путь апоптоза несколькими способами: через 1) про
апоптозный белок Bid, 2) белки Mcl 1 и Bim или 3) белок р53. В результате из митохондрий в цитоплазму
выходит цитохром с, и образуются апоптосомы, обеспечивающие протеолитический каскад активации кас
паз. Гранзимы М, H и F вызывают гибель клеток, которая сопровождается выходом цитохрома с из митохон
дрий. Гранзим А индуцирует образование активных форм кислорода (АФК), которые способствуют транс
локации ассоциированного с эндоплазматическим ретикулумом комплекса SET в ядро клетки. В клеточном
ядре гранзим А расщепляет SET; это активирует нуклеазы, которые осуществляют одноцепочечные разры
вы ДНК. Гранзимы А и В проникают в митохондрии и разрезают субъединицы комплекса I дыхательной це
пи. Одна из субъединиц комплекса I является мишенью также и для каспазы 3. Гранзим зависимое повреж
дение комплекса I приводит к образованию АФК и гибели клеток.
КЛЮЧЕВЫЕ СЛОВА: активные формы кислорода, апоптоз, гранзимы, митохондрии, программируемая
клеточная смерть.
DOI: 10.31857/S0320972520020013
Если организм инфицирован, то основной
киллерах, и рецептора CD95 (Fas) на клетках
способ его защиты - избавление от собственных
мишенях [3, 4]. В результате происходит актива
зараженных клеток. Специализированные клет
ция каспаз, начиная с каспазы 8, и апоптоз [5].
ки иммунной системы
- цитотоксические
При определенных условиях связывание лиган
Т лимфоциты и естественные киллеры (NK,
дов с рецепторами может приводить не к апоп
natural killer) - осуществляют поиск и уничто
тозу, а к некроптозу - одной из форм програм
жение клеток, содержащих чужеродные компо
мируемого (регулируемого) некроза. Образуется
ненты: инфицированных, мутировавших или
рипоптосома - белковый комплекс (2 MДа), со
раковых клеток. Ликвидация таких клеток осу
держащий киназу RIPK1 (receptor interacting
ществляется посредством включения в них
protein kinase 1), адаптерный белок FADD (Fas
программы гибели клеток (апоптоза) и может
associated death domain), каспазу 8 и cFLIP (cel
происходить двумя путями [1, 2].
lular FLICE like inhibitory protein). Некроптоз
Первый путь реализуется через взаимодей
реализуется в условиях подавления активности
ствие лигандов и рецепторов суперсемейства
cIAPs (сellular inhibitors of apoptosis proteins), ко
фактора некроза опухоли (TNF, tumor necrosis
торые способствуют убиквитинированию и раз
factor), например, CD95L (FasL, Fas лиганд),
рушению киназы RIPK1. Образование рипопто
расположенного трансмембранно на клетках
сомы регулируется разными изоформами белка
cFLIP и обеспечивает последовательное фосфо
Принятые сокращения: АФК - активные формы
рилирование RIPK1, RIPK3 и белка MLKL
кислорода; O2 - супероксидный анион радикал; GAAD -
(mixed lineage kinase like protein), который оли
ДНКаза, активируемая гранзимом А; FeS кластеры - же
гомеризуется и образует каналы в плазматичес
лезо серные кластеры; FMN - флавинмононуклеотид;
-
кой мембране клеток [6-8].
NK - естественные киллеры; ΔμH+ - разность электрохи
мических потенциалов ионов водорода; Δψ - трансмем
Второй путь - это высвобождение содержи
бранная разность электрических потенциалов.
мого литических гранул клеток убийц (рис. 1).
155
156
КИСЕЛЕВСКИЙ
животных общая концентрация кальция варьи
рует от 2,1 до 2,6 мМ, доля свободного ионизи
рованного Ca2+ снаружи клеток обычно состав
ляет половину от этих значений, а внутри кле
ток его концентрация на ~5 порядков ниже [16].
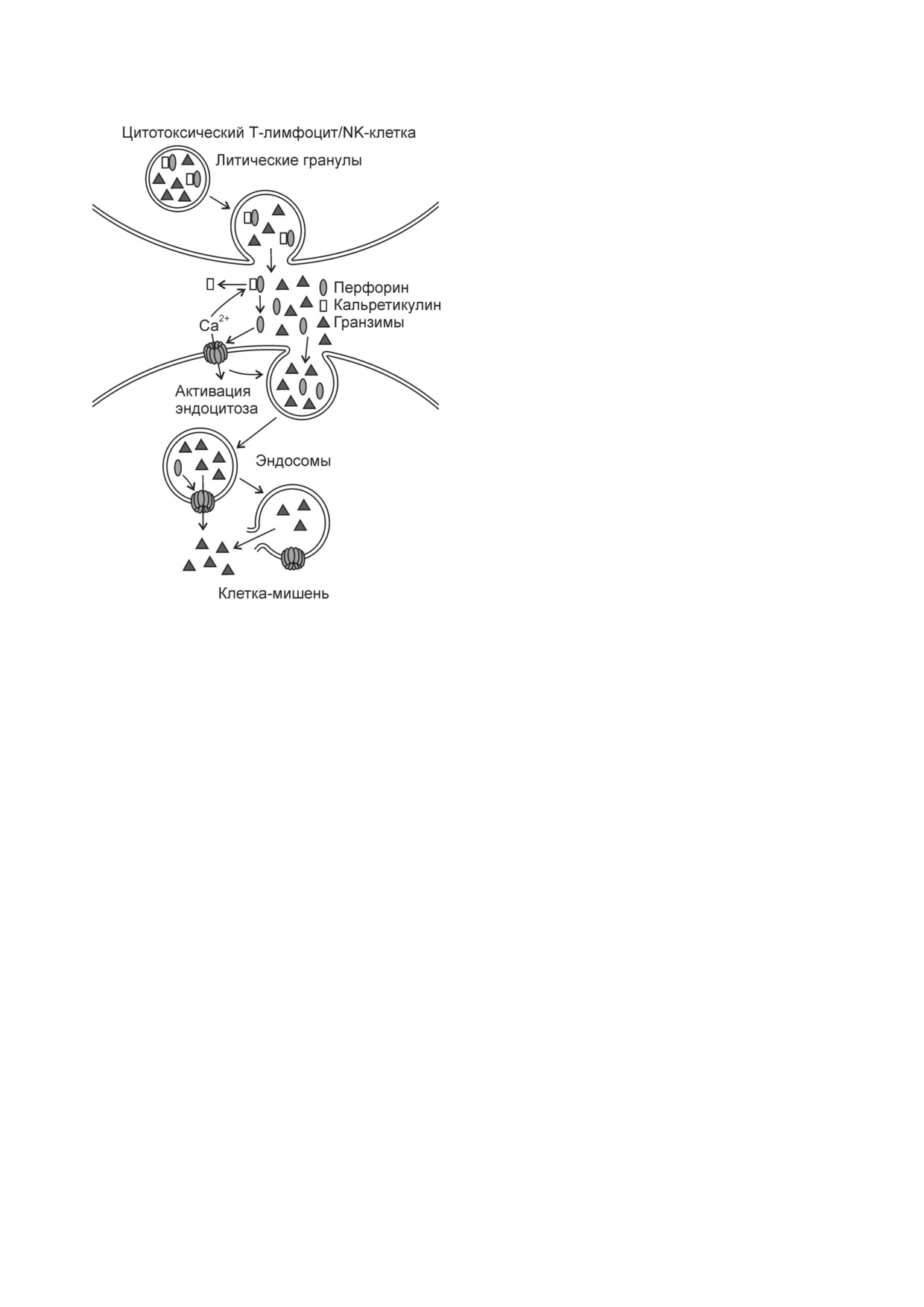
С помощью перфориновых пор внутрь кле
ток мишеней проникает еще один компонент
гранул - гранзимы - сериновые протеазы, раз
резающие внутриклеточные белки субстраты.
Проникновение гранзимов в клетку происходит
не напрямую через перфориновые каналы в
плазматической мембране. Длительное наруше
ние барьерной функции наружной мембраны
клеток, вызванное перфорином, может приво
дить к гибели клеток по типу некроза, который
сопровождается воспалением, тогда как гран
зим зависимая гибель клеток - это апоптоз,
происходящий, как правило, без воспаления
[17]. Перфориновые поры в плазматической
мембране вызывают временный приток Ca2+ в
клетку мишень (рис. 1), который длится не
сколько минут. Увеличение внутриклеточной
концентрации Ca2+ активирует механизм вос
становления повреждений плазматической
мембраны, включающий эндоцитоз для удале
ния поврежденных участков мембраны. Перфо
рин и гранзимы попадают в эндосомы. Гранзи
Рис. 1. Путь доставки гранзимов из литических гранул ци
мы могут медленно выходить из эндосом через
тотоксических Т лимфоцитов и NK клеток в цитоплазму
перфориновые поры в их мембране. Затем, при
клетки мишени
мерно через 15 мин после воздействия на клетки
перфорином и гранзимом, мембрана эндосом
разрывается, и гранзимы высвобождаются в ци
Чтобы сконцентрировать содержимое литичес
топлазму клетки мишени (рис. 1) [18].
ких гранул и обезопасить от него соседние клет
Название гранзимы (от словосочетания
ки, между клеткой киллером и клеткой ми
«гранулярные энзимы») было впервые предло
шенью формируется иммунный синапс - мик
жено для двух обнаруженных протеаз - гранзи
роскопическая межклеточная щель, ограничен
мов А (35 кДа) и В (29 кДа) - в 1986 г. в работе
ная плазматическими мембранами взаимодей
Masson et al. [19]. В настоящее время идентифи
ствующих клеток, в которую высвобождаются
цированы 12 гранзимов (A, B, C, D, E, F, G, H,
компоненты литических гранул. Размеры им
J, K, M и N), из них пять (A, B, H, K и M) най
мунного синапса достигают нескольких мик
дены у человека [20-22]. В литических гранулах
рон, а ширина щели между клетками - порядка
цитотоксических Т лимфоцитов и NK клеток
десятков нанометров [9-11]. В гранулах содер
~90% массы приходится на долю гранзимов [23].
жится перфорин с мембраноатакующим комп
лексом, подобным С9 компоненту системы
комплемента [12], создающий каналы в мембра
ВЫСВОБОЖДЕНИЕ ЦИТОХРОМА С
не диаметром в среднем ~16 нм [13, 14].
ИЗ МИТОХОНДРИЙ
Другой компонент литических гранул -
кальретикулин. В отсутствие Ca2+ он связывает
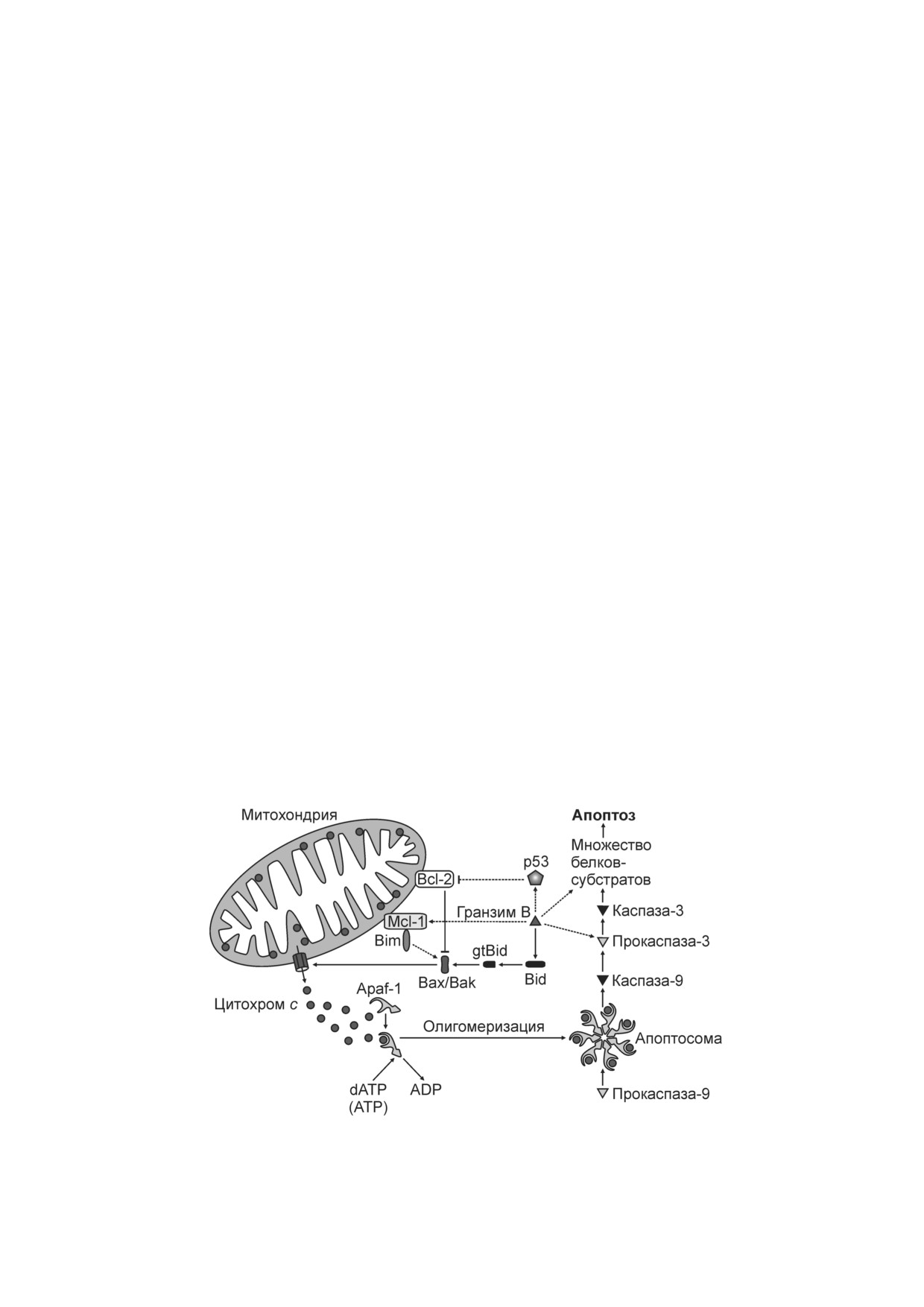
Гранзим B инициирует зависимый от мито
перфорин (рис. 1), а при увеличении концентра
хондрий путь апоптоза с участием проапоптоз
ции Ca2+ до 0,1 мМ и выше комплекс кальрети
ного белка Bid (BH3 interacting domain death ago
кулин-перфорин диссоциирует. Предполагает
nist). При входе в цитоплазму из эндосомально
ся, что кальретикулин препятствует образова
го компартмента гранзим B расщепляет Bid с
нию перфориновых пор в мембране литических
образованием gtBid (granzyme truncated Bid),
гранул, предохраняя цитотоксические Т лим
приводя к апоптозу (рис. 2). Участок разрезания
фоциты и естественные киллеры от опасного
белка Bid гранзимом В отличается от места его
содержимого гранул [15]. В жидкостях и тканях
расщепления каспазой 8. Сверхэкспрессия в
БИОХИМИЯ том 85 вып. 2 2020
ГРАНЗИМЫ И МИТОХОНДРИИ
157
митохондриях антиапоптозного белка Bcl 2 (B
зуется. Образовавшийся гептамер - апоптосома,
cell lymphoma 2) может блокировать проявления
являющаяся платформой для активации каспа
клеточной гибели, вызванной гранзимом B
зы 9. Апоптосома напрямую связывает молеку
[24-26]. Белок gtBid (14 кДа) транслоцируется в
лы прокаспазы 9 через гомотипическое взаимо
митохондрии и рекрутирует белки Bax (Bcl 2
действие доменов CARD (сaspase activation and
associated
X protein) или Bak (Bcl 2
recruitment domain), расположенных в централь
antagonist/killer), которые интегрируются в
ной части апоптосомы и на прокаспазе 9 [32,
мембрану и индуцируют высвобождение цито
33]. Удаление домена CARD существенно увели
хрома с (рис. 2) [27, 28]. Согласно одной из мо
чивает каталитическую активность прокаспазы 9,
делей взаимодействия белков семейства Bcl 2,
поэтому роль апоптосомы в активации каспазы 9
ингибирующее действие белка Bcl 2 может быть
может заключаться не только в олигомеризации
обусловлено его прямым связыванием с Bax,
прокаспаз 9 и их автопротеолизе [34, 35], но и в
Bak или gtBid. Другая модель предполагает, что в
устранении ингибирующего эффекта домена
норме белок Bax, имеющий сродство к липид
CARD [36]. Каспаза 9 инициирует протеолити
ной мембране, постоянно транслоцируется с на
ческий каскад, активирующий эффекторные
ружной мембраны митохондрии в цитоплазму
каспазы 3 и
7 [34]. Каспаза 3 может активиро
белком Bcl 2. При апоптозе gtBid (а также дру
ваться напрямую гранизимом В (рис. 2), но не
гие белки из группы проапоптозных белков се
гранзимом А [37, 38]. Субстратами эффекторных
мейства Bcl 2, содержащих только домен BH3)
каспаз, в т.ч. каспазы 3, являются сотни белков
препятствует действию антиапоптозного Bcl 2,
клетки. Среди них белки ядра и цитоскелета,
в результате чего Bax связывается с мембраной,
протеолиз которых приводит к фрагментации
димеризуется, а затем олигомеризуется, форми
клеточного ядра и образованию везикул на плаз
руя поры, через которые цитохром с выходит из
матической мембране - характерным проявле
межмембранного пространства митохондрий в
ниям апоптоза [39, 40].
цитоплазму [29, 30].
У гранзима В, кроме белка Bid и прокаспаз 2,
Связывание цитохрома с в цитоплазме с мо
3,
7,
8,
9 и
10, есть множество других бел
номерным Apaf 1 (apoptotic protease activating
ков мишеней в клетке, включая белки теплово
factor 1) вызывает конформационные измене
го шока, белки цитоскелета (среди которых ак
ния последнего, приводящие к нуклеотидному
тин и тубулин), ингибитор каспаза активируе
обмену: ADP в нуклеотид связывающем домене
мой ДНКазы ICAD/DFF45 (inhibitor of caspase
Apaf 1 заменяется на dATP или ATP. В этой реак
activated DNase/DNA fragmentation factor, 45 кДа),
ции dATP более эффективен в сравнении с АТР,
ядерные белки (в т.ч. ламин, поли(ADP рибо
но внутриклеточная концентрация АТР выше,
за)полимераза и другие белки, связанные с ре
поэтому оба нуклеотида могут быть вовлечены в
парацией ДНК)
[10]. Митохондриальные
митохондриальный путь апоптоза [31]. Далее
субстраты гранзима В включают белок Mcl 1
Apaf 1, связанный с цитохромом с, олигомери
(myeloid cell leukemia 1), расположенный на
Рис. 2. Гранзим В активирует митохондриальный путь апоптоза
БИОХИМИЯ том 85 вып. 2 2020
158
КИСЕЛЕВСКИЙ
внешней мембране митохондрий [41, 42], и ди
клеток, вызванная гранзимом F, не включала
гидролипоилтрансацетилазу Е2 пируватдегид
расщепление Bid и активацию каспаз. Гранзим F
рогеназного комплекса [43], осуществляющего
нарушал транспорт электронов в дыхательной
окислительное декарбоксилирование пирувата с
цепи митохондрий, снижая скорость поглоще
образованием ацетил кофермента А и СО2, а
ния О2 и образование АТР в клетках [50].
также восстановлением NAD+ в матриксе мито
Гранзим А, подобно гранзиму В, способен
хондрий.
расщеплять множество клеточных белков, в т.ч.
При протеолизе Mcl 1 гранзимом В может
белки цитоскелета и ядра [51]. Губительное
осуществляться альтернативный, независимый
действие гранзима А на клетки связывают с ак
от белка Bid митохондриальный путь апоптоза
тивацией нуклеаз, производящих одноцепочеч
(рис. 2). Антиапоптозный белок Mcl 1 связыва
ные разрывы ДНК и способствующих гибели
ет проапоптозный белок Bim (или Bcl 2 like 11,
клеток с признаками апоптоза. С эндоплазмати
apoptosis facilitator). Оба этих белка, как и белки
ческим ретикулумом связан белковый комплекс
Bid, Bax и Bak, принадлежат к семейству белков
(270-420 кДа), содержащий ДНКазу NM23 H1
Bcl 2 [44]. Гранзим В высвобождает Bim из
(или GAAD, granzyme A activated DNase) и ее
комплекса Mcl 1-Bim, активируя его [10, 41].
ингибитор SET, который разрезается гранзимом А.
Как и в случае с Bid, обработанным гранзимом В,
В результате ДНКаза высвобождается и активи
белок Bim напрямую инициирует олигомериза
руется. Клетки с подавленной экспрессией
цию белков Вах и Bak, образование пор в мемб
NM23 H1 устойчивы к повреждению ДНК, выз
ранах митохондрий и выход цитохрома с. Bid
ванному гранзимом А, и цитолизу, а клетки со
преимущественно активирует Bak, тогда как
сверхэкспрессией NM23 H1, напротив, более
Bim предпочитает Bax [45].
чувствительны [52, 53]. Было обнаружено, что
В еще одном пути регуляции апоптоза гран
гранзим А вызывает образование активных форм
зимом B, связанном с митохондриями, участву
кислорода (АФК) и снижение разности электри
ет опухолевый супрессор р53. Активированный
ческих потенциалов (Δψ) в митохондриях. Пов
гранзимом B белок р53 транслоцируется к мито
реждение митохондрий является важным этапом
хондриям и взаимодействует с Bcl 2 на внешней
в апоптозе, вызванном гранзимом А [54].
митохондриальной мембране (рис. 2). Это уст
раняет ингибирующее воздействие Bcl 2 на Bax
и способствует образованию пор в митохондри
ОБРАЗОВАНИЕ АФК В МИТОХОНДРИЯХ
альной мембране [46].
Изученные в меньшей степени гранзимы М,
NADH:убихинон оксидоредуктаза (комп
H и F индуцировали гибель клеток с участием
лекс I дыхательной цепи митохондрий) катали
митохондрий, которая сопровождалась выходом
зирует окисление NADH убихиноном. Двухэлек
цитохрома с из митохондрий в цитоплазму.
тронное окисление одной молекулы NADH
Гранзим М напрямую не расщеплял прокаспазу 3
сопровождается трансмембранным переносом
и Bid, его действие опосредовано каспазой 8.
четырех протонов и генерацией на внутренней
Ассоциированный с рецептором смерти CD95
мембране митохондрий разности электрохими
-
(Fas) адаптерный белок FADD (Fas associated
ческих потенциалов ионов водорода (ΔμH+) [55].
protein with death domain) специфически рас
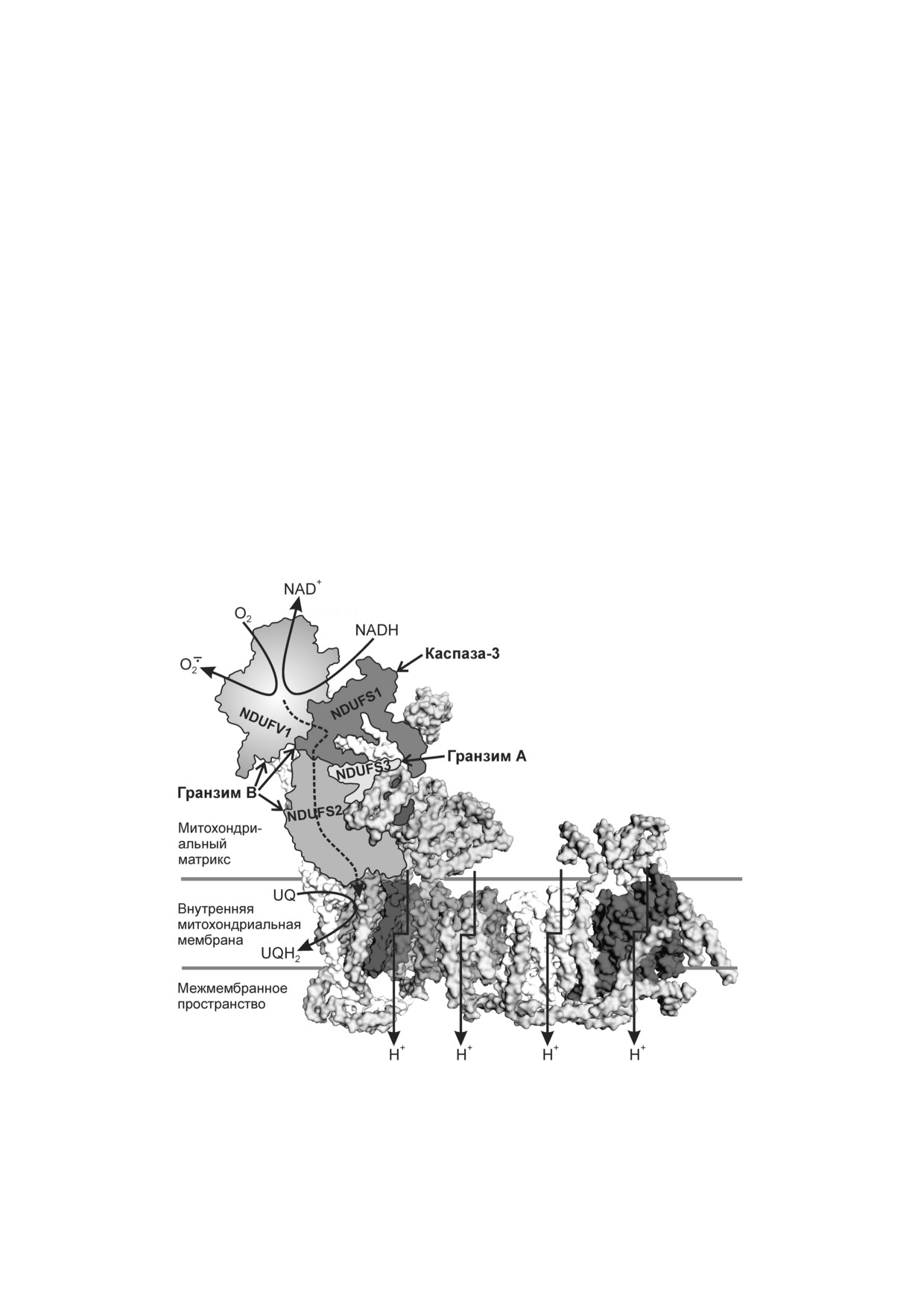
Комплекс I - один из крупнейших (970 кДа)
щеплялся гранзимом М после остатка Met196,
комплексов белков в клетке, связанных с мемб
образуя усеченный белок (tFADD). Это облегча
раной. Комплекс I млекопитающих содержит 45
ло рекрутирование прокаспазы 8. Индуциро
субъединиц: 14 основных субъединиц консерва
ванной гранзимом М гибели клеток сопутство
тивны и имеются у живых организмов от бакте
вало высвобождение цитохрома с из митохонд
рий до человека; 31 субъединица специфична
рий в цитоплазму [47]. Гранзим Н вызывал ги
для млекопитающих [56, 57].
бель клеток при участии Bcl 2, однако также не
Гранзим А действует непосредственно на ми
осуществлял прямого расщепления Bid. Выз
тохондрии, вызывая образование АФК и сниже
ванная гранзимом H гибель клеток сопровожда
ние Δψ на внутренней мембране митохондрий.
лась выходом цитохрома с из митохондрий. Ак
После 2 мин обработки гранзимом А в комбина
тивация каспаз и защитный эффект ингибито
ции с перфорином митохондрии клеток HeLa
ров каспаз наблюдались, однако были неболь
начинали генерировать супероксидный анион
шими при сравнении гранзима H c другими ин
радикал (O2 ), который детектировали с по
дукторами апоптоза. Гранзим Н, подобно гран
мощью MitoSOX Red, митохондриального флуо
зиму В, непосредственно взаимодействовал с
ресцентного индикатора O2 . Показано, что
ICAD/DFF45, что, вероятно, приводило к пов
гранзим А разрезает субъединицу комплекса I
реждению ДНК и гибели клеток [48, 49]. Смерть
NDUFS3 после остатка Lys56 [58]. Гранзим B,
БИОХИМИЯ том 85 вып. 2 2020
ГРАНЗИМЫ И МИТОХОНДРИИ
159
который подобно гранзиму А в сочетании с пер
Проникновение в митохондрии гранзима В
форином вызывал образование митохондриаль
происходит независимо от транслоказы внеш
ных АФК, расщепляет субъединицы NDUFV1,
ней митохондриальной мембраны Tom (translo
NDUFS1 и NDUFS2 [59]. Примечательно, что
case of the outer mitochondrial membrane), вместо
NDUFS1 разрезается также каспазой 3 - цент
этого используется Sam50 (sorting and assembly
ральным протеолитическим ферментом при
machinery). Через внутреннюю мембрану гран
апоптозе. В NDUFS1 есть участок из четырех ос
зим В проходит с помощью Tim22 (translocase of
татков аминокислот (DVMD у млекопитающих),
the inner mitochondrial membrane) при участии
являющийся субстратом каспазы [60]. На рис. 3
митохондриального белка теплового шока 70
отмечены субъединицы комплекса I, подверга
(mtHsp70). Гранзим A и каспаза 3 доставляются
ющиеся протеолизу каспазой 3 (NDUFS1),
в митохондрии по похожему пути. Для проник
гранзимами А (NDUFS3) или В (NDUFV1,
новения гранзимов и каспазы 3 митохондрии
NDUFS1 и NDUFS2). Все эти субъединицы из
должны обладать Δψ на внутренней мембране
числа основных субъединиц расположены в
[63, 64].
гидрофильном (немембранном) домене комп
Генерация O2 в комплексе I дыхательной це
лекса I, в котором происходит перенос электро
пи митохондрий зависит от Δψ, концентрации
нов с NADH на убихинон по цепочке FeS клас
и соотношения NADH/NAD+, соотношения
теров. NDUFV1 содержит флавинмононуклео
восстановленного и окисленного убихонона и
тид (FMN) и FeS кластер N3; NDUFS1 - FeS
концентрации O2 [65-68]. Считается, что ос
кластеры N1b, N4 и N5; NDUFS2 и NDUFS3
новным кофактором, ответственным за генера
содержат FeS кластеры, не принадлежащие к
цию O2 , является FMN, но не исключено и об
основному пути переноса электронов от NADH
разование АФК на участке связывания убихи
к убихинону [57].
нона. Предположительно, существуют два пути
Рис. 3. Структура комплекса I дыхательной цепи митохондрий (по статьям Hirst и Roessler [61], Rodenburg [62]). Обозна
чены субъединицы, расщепляемые каспазой 3 (NDUFS1), гранзимом А (NDUFS3) или гранзимом B (NDUFV1, NDUFS1
и NDUFS2). Пунктирной линией показан перенос электронов от FMN к участку связывания убихинона по цепочке FeS
кластеров
БИОХИМИЯ том 85 вып. 2 2020
160
КИСЕЛЕВСКИЙ
образования O2 . Первый путь: NADH → FMN в
связанного с ним цитохрома с в цитоплазму [69,
комплексе I
→ O2. Высокий уровень
70]. Антиоксиданты подавляли гибель клеток,
NADH/NAD+ приводит к восстановлению
вызванную гранзимом А и перфорином [54].
FMN. FMN (полностью восстановленный или в
АФК способствуют транслокации ассоцииро
форме семихинона) реагирует с О2, образуя O2 .
ванного с эндоплазматическим ретикулумом
Второй путь (обратный перенос электронов):
комплекса SET в ядро, где этот комплекс рас
восстановленный убихинон → цепь FeS класте
щепляется гранзимом А с высвобождением двух
ров в комплексе I → FMN в комплексе I → O2 .
ДНКаз (NM23 H1 или GAAD и TREX1), кото
Сукцинат, субстрат комплекса II дыхательной
рые разрезают ядерную ДНК при гибели кле
цепи, восстанавливающего убихинон, вызывал
ток, опосредованной гранзимом А [54, 71]. Ан
генерацию O2 в субмитохондриальных части
тиоксиданты также уменьшали проявление
цах. Она подавлялась ротеноном, ингибирую
признаков апоптоза, индуцированного гранзи
щим взаимодействие убихинона с комплексом
мом В [59].
I, и протонофорным разобщителем, снимаю
Литические гранулы клеток киллеров у че
щим Δψ. NADH Зависимое образование АФК,
ловека (но не у грызунов), в дополнение к пер
напротив, могло усиливаться ротеноном и быть
форину, кальретикулину и гранзимам, содержат
нечувствительным к действию разобщителя.
антимикробный пептид гранулизин, который
NADH в миллимолярной концентрации (близ
избирательно разрушает микробные мембраны
кой к физиологической) и NAD+ подавляли об
с низким содержанием холестерина. Этот пеп
разование O2 , по видимому, конкурируя с О2 за
тид способствует защите организма от внутри
связывание с восстановленным FMN. Есть
клеточных паразитических простейших (трипа
предположение, что FMN комплекса I взаимо
носомы, токсоплазмы, лейшмании). Перфорин
действует по меньшей мере с двумя разными
обеспечивает проникновение гранзимов и гра
участками связывания нуклеотидов: в одном
нулизина в инфицированные клетки, а затем
осуществляется окисление NADH, а в другом
гранулизин доставляет гранзимы к внутрикле
может происходить восстановление NAD+ или
точным паразитам. Показано, что гранзимы за
О2 [65, 66].
пускали образование АФК и инактивировали
Неясно, каким образом опосредованный
ферменты антиоксидантной защиты для унич
гранзимами протеолиз субъединиц комплекса I
тожения паразита. Гибель паразитических прос
приводит к образованию АФК. Поскольку
тейших была нечувствительна к ингибиторам
действие гранзимов сопровождается снижением
каспаз, но по ряду признаков напоминала апоп
Δψ, можно предположить, что разрезание субъ
тоз млекопитающих [72].
единиц, в т.ч. и тех, в которых находятся компо
Таким образом, цитотоксические клетки
ненты электронтранспортной цепи (NDUFV1 и
киллеры, использующие гранзимы, могут ини
NDUFS1), нарушает перенос электронов на
циировать множество путей активации клеточ
убихинон, и генерация O2 происходит по перво
ной гибели. По видимому, это необходимо для
му пути: NADH → FMN → O2 . Однако это про
противодействия патогенам, которые могут ис
тиворечит наблюдению, что ротенон подавлял
пользовать разные способы уклонения от имму
образование АФК и гибель клеток, вызванную
нитета [49].
гранзимом А в комбинации с перфорином [58].
Ингибирование процесса ротеноном - признак
В последние годы растет интерес к T или
обратного переноса электронов, от убихинона
NK клеткам киллерам. Это связано как с их
на FMN. По видимому, при обработке клеток
участием в противовирусной защите организма,
гранзимами основной путь переноса электронов
так и с возможным применением в иммунотера
сохраняется. Возможно, протеолиз субъединиц
пии опухолей. Цитотоксические Т и NK клет
комплекса I способствует доступу О2 к скрытым
ки вырабатывают интерферон γ, который обес
внутри белка FMN и FeS центрам и их окисле
печивает включение ряда различных механиз
нию с образованием O2 . Наряду с глутамат и
мов противовирусной защиты [73]. Цитотокси
малат зависимым поглощением О2, гранзим В
ческие T лимфоциты распознают и уничтожают
подавлял дыхание с сукцинатом, а также актив
клетки, зараженные вирусом и презентирующие
ность комплекса III дыхательной цепи, поэтому
вирусные пептиды на молекулах главного комп
не исключено взаимодействие гранзимов с дру
лекса гистосовместимости класса I (MHC I,
гими комплексами дыхательной цепи [59].
major histocompatibility complex class I) [74]. Это
АФК играют важную роль при внедрении в
свойство может иметь практическое примене
клетку гранзимов. Образование АФК в митохон
ние в медицине. Так, например, трансплантация
дриях может провоцировать окисление кардио
сопряжена с ослаблением иммунитета и повы
липина и высвобождение электростатически
шенным риском инфицирования. При этом ис
БИОХИМИЯ том 85 вып. 2 2020
ГРАНЗИМЫ И МИТОХОНДРИИ
161
пользование специфичных к вирусным антиге
является многообещающей стратегией избавле
нам цитотоксических Т лимфоцитов, получен
ния от злокачественных новообразований, ус
ных с помощью МНС, которые несут синтети
тойчивых к традиционным методам лечения.
ческие вирусные пептиды - это возможная аль
Отсутствие МНС I на поверхности клеток мо
тернатива высокотоксичным противовирусным
жет быть следствием опухолевой трансформа
препаратам [75].
ции. Утилизация таких клеток естественными
Для ускользания от противовирусной реак
киллерами дополняет уничтожение клеток опу
ции цитотоксических Т клеток вирусы могут
холи цитотоксическими T лимфоцитами [77,
препятствовать презентации антигена: подав
78].
лять функции протеасом, транспорт пептидов,
Ликвидация инфицированных или мутиро
транслокацию молекул МНС I из эндоплазма
вавших клеток - основа защитной реакции,
тического ретикулума на поверхность клеток
осуществляемой цитотоксическими Т лимфо
[74]. Естественные киллеры способны выявлять
цитами и естественными киллерами и направ
и уничтожать инфицированные клетки, в кото
ленной на поддержание гомеостаза организма.
рых нарушена презентация антигенов, т.е. нет
Существование множества альтернативных пу
МНС I на клеточной поверхности. Вклад цито
тей инициации гибели клеток, которые описа
литической функции NK клеток, обеспечивае
ны в настоящей работе, по видимому, важно,
мой перфорином и гранзимами, в противови
чтобы: 1) препятствовать механизмам ускольза
русную защиту зависит от многих факторов и
ния от иммунных реакций, возникающим у па
может быть неодинаковым в разных тканях ор
тогенов; 2) осуществлять клеточную смерть да
ганизма [76].
же в случае мутаций отдельных компонентов пу
Перспективно использование клеток килле
тей, приводящих к ней. Как и в случае зависи
ров в иммунотерапии рака. Есть свидетельства
мого от каспаз апоптоза, большинство выявлен
успешного применения адоптивной терапии
ных путей гибели клетки, включаемых гранзи
опухолей, при которой у пациента берут клетки
мами, ассоциированы с митохондриями.
киллеры, подвергают их обработке цитокинами
и/или генно инженерным модификациям для
активации и усиления противоопухолевого от
Финансирование. Работа выполнена при фи
вета и вводят обратно в организм [77]. Особый
нансовой поддержке научно исследовательской
интерес представляют модифицированные
работы (НИР) из средств федерального бюджета
клетки с химерными антигенными рецепторами
(регистрационный номер НИР в ЦИТИС:
(CAR, chimeric antigen receptors) - синтетичес
АААА А16 116021660081 0).
кими рецепторами, содержащими антигенсвя
Благодарности. Автор благодарен д.б.н. про
зывающие, активирующие и костимуляторные
фессору В.Д. Самуилову за внимательное про
домены, в которых антигенсвязывающий сайт
чтение рукописи статьи и ценные замечания.
рецептора Т клеток (TCR, T cell receptor) заме
Конфликт интересов. Автор заявляет об отсут
няется сайтом иммуноглобулина с высокой аф
ствии конфликта интересов.
финностью и специфичностью к целевому опу
Соблюдение этических норм. Настоящее ис
холевому антигену. Терапия на основе CAR поз
следование проводили без использования жи
воляет устранить необходимость МНС зависи
вотных и без привлечения людей в качестве ис
мой презентации антигена клеткам киллерам и
пытуемых.
СПИСОК ЛИТЕРАТУРЫ
1.
Cullen, S.P., and Martin, S.J. (2008) Mechanisms of gran
5.
Tummers, B., and Green, D.R. (2017) Caspase 8: regulat
ule dependent killing, Cell Death Differ., 15, 251-262,
ing life and death, Immunol. Rev.,
277,
76-89,
doi: 10.1038/sj.cdd.4402244.
doi: 10.1111/imr.12541.
2.
Cullen, S.P., Brunet, M., and Martin, S.J.
(2010)
6.
Vanden Berghe, T., Linkermann, A., Jouan Lanhouet, S.,
Granzymes in cancer and immunity, Cell Death Differ., 17,
Walczak, H., and Vandenabeele, P. (2014) Regulated necrosis:
616-623, doi: 10.1038/cdd.2009.206.
the expanding network of non apoptotic cell death pathways,
3.
Wajant, H. (2014) Principles and mechanisms of CD95
Nat. Rev. Mol. Cell Biol., 15, 135-147, doi: 10.1038/nrm3737.
activation, Biol. Chem., 395, 1401-1416, doi: 10.1515/hsz
7.
Tsuchiya, Y., Nakabayashi, O., and Nakano, H. (2015) FLIP
2014 0212.
the switch: regulation of apoptosis and necroptosis by cFLIP,
4.
Siegmund, D., Lang, I., and Wajant, H. (2017) Cell death
Int. J. Mol. Sci., 16, 30321-30341, doi: 10.3390/ijms161226232.
independent activities of the death receptors CD95,
8.
Zhang, Y., Chen, X., Gueydan, C., and Han, J. (2018)
TRAILR1, and TRAILR2, FEBS J., 284, 1131-1159,
Plasma membrane changes during programmed cell deaths,
doi: 10.1111/febs.13968.
Cell Res., 28, 9-21, doi: 10.1038/cr.2017.133.
БИОХИМИЯ том 85 вып. 2 2020
162
КИСЕЛЕВСКИЙ
9.
Grakoui, A., Bromley, S.K., Sumen, C., Davis, M.M.,
Roberts, D.L., Hunter, A.M., Korneluk, R., and
Shaw, A.S., Allen, P.M., and Dustin, M.L. (1999) The
Bleackley, R.C. (2003) Granzyme B induced apoptosis
immunological synapse: a molecular machine controlling T
requires both direct caspase activation and relief of caspase
cell activation, Science, 285, 221-227, doi: 10.1126/sci
inhibition, Immunity, 18, 355-365, doi: 10.1016/s1074
ence.285.5425.221.
7613(03)00032 3.
10.
Rousalova, I., and Krepela, E. (2010) Granzyme B induced
26.
Wowk, M.E., and Trapani, J.A. (2004) Cytotoxic activity of
apoptosis in cancer cells and its regulation (review), Int. J.
the lymphocyte toxin granzyme B, Microbes Infect., 6,
Oncol., 37, 1361-1378, doi: 10.3892/ijo_00000788.
752-758, doi: 10.1016/j.micinf.2004.03.008.
11.
Woodsworth, D.J., Dunsing, V., and Coombs, D. (2015)
27.
Heibein, J.A., Goping, I.S., Barry, M., Pinkoski, M.J.,
Design parameters for granzyme mediated cytotoxic lym
Shore, G.C., Green, D.R., and Bleackley, R.C. (2000)
phocyte target cell killing and specificity, Biophys. J., 109,
Granzyme B mediated cytochrome c release is regulated by
477-488, doi: 10.1016/j.bpj.2015.06.045.
the Bcl 2 family members Bid and Bax, J. Exp. Med., 192,
12.
Podack, E.R., and Munson, G.P. (2016) Killing of microbes
1391-1402, doi: 10.1084/jem.192.10.1391.
and cancer by the immune system with three mammalian
28.
Wang, G.Q., Wieckowski, E., Goldstein, L.A., Gastman, B.R.,
pore forming killer proteins, Front. Immunol., 7, 464,
Rabinovitz, A., Gambotto, A., Li, S., Fang, B., Yin, X.M.,
doi: 10.3389/fimmu.2016.00464.
and Rabinowich, H. (2001) Resistance to granzyme B
13.
Stewart, S.E., D’Angelo, M.E., and Bird, P.I.
(2012)
mediated cytochrome c release in Bak deficient cells, J.
Intercellular communication via the endo lysosomal sys
Exp. Med., 194, 1325-1337, doi: 10.1084/jem.194.9.1325.
tem: translocation of granzymes through membrane barri
29.
Cosentino, K., and Garcia Saez, A.J. (2017) Bax and Bak
ers, Biochim. Biophys. Acta, 1824, 59-67, doi: 10.1016/
pores: Are we closing the circle? Trends Cell Biol., 27,
j.bbapap.2011.05.020.
266-275, doi: 10.1016/j.tcb.2016.11.004.
14.
Voskoboinik, I., Whisstock, J.C., and Trapani, J.A. (2015)
30.
Kale, J., Osterlund, E.J., and Andrews, D.W. (2018) BCL
Perforin and granzymes: function, dysfunction and human
2 family proteins: changing partners in the dance towards
pathology, Nat. Rev. Immunol., 15, 388-400, doi: 10.1038/
death,
Cell
Death
Differ.,
25,
65-80,
nri3839.
doi: 10.1038/cdd.2017.186.
15.
Andrin, C., Pinkoski, M.J., Burns, K., Atkinson, E.A.,
31.
Li, P., Nijhawan, D., Budihardjo, I., Srinivasula, S.M.,
Krahenbuhl, O., Hudig, D., Fraser, S.A., Winkler, U.,
Ahmad, M., Alnemri, E.S., and Wang, X.
(1997)
Tschopp, J., Opas, M., Bleackley, R.C., and Michalak, M.
Cytochrome c and dATP dependent formation of Apaf
(1998) Interaction between a Ca2+ binding protein calretic
1/caspase 9 complex initiates an apoptotic protease cas
ulin and perforin, a component of the cytotoxic T cell gran
cade, Cell,
91,
479-489, doi:
10.1016/s0092
ules, Biochemistry,
37,
10386-10394, doi:
10.1021/
8674(00)80434 1.
bi980595z.
32.
Li, J., and Yuan, J. (2008) Caspases in apoptosis and beyond,
16.
Carafoli, E., and Krebs, J. (2016) Why calcium? How calci
Oncogene, 27, 6194-6206, doi: 10.1038/onc.2008.297.
um became the best communicator, J. Biol. Chem., 291,
33.
Dorstyn, L., Akey, C.W., and Kumar, S. (2018) New
20849-20857, doi: 10.1074/jbc.R116.735894.
insights into apoptosome structure and function, Cell Death
17.
Davidovich, P., Kearney, C.J., and Martin, S.J. (2014)
Differ., 25, 1194-1208, doi: 10.1038/s41418 017 0025 z.
Inflammatory outcomes of apoptosis, necrosis and necrop
34.
Srinivasula, S.M., Ahmad, M., Fernandes Alnemri, T., and
tosis, Biol. Chem., 395, 1163-1171, doi: 10.1515/hsz 2014
Alnemri, E.S. (1998) Autoactivation of procaspase 9 by
0164.
Apaf 1 mediated oligomerization, Mol. Cell, 1, 949-957,
18.
Thiery, J., Keefe, D., Boulant, S., Boucrot, E., Walch, M.,
doi: 10.1016/s1097 2765(00)80095 7.
Martinvalet, D., Goping, I.S., Bleackley, R.C.,
35.
Chang, H.Y., and Yang, X. (2000) Proteases for cell suicide:
Kirchhausen, T., and Lieberman, J. (2011) Perforin pores in
functions and regulation of caspases, Microbiol. Mol. Biol.
the endosomal membrane trigger the release of endocytosed
Rev., 64, 821-846, doi: 10.1128/mmbr.64.4.821 846.2000.
granzyme B into the cytosol of target cells, Nat. Immunol.,
36.
Li, Y., Zhou, M., Hu, Q., Bai, X. C., Huang, W.,
12, 770-777, doi: 10.1038/ni.2050.
Scheres, S.H.W., and Shi, Y. (2017) Mechanistic insights
19.
Masson, D., Nabholz, M., Estrade, C., and Tschopp, J.
into caspase 9 activation by the structure of the apoptosome
(1986) Granules of cytolytic T lymphocytes contain two
holoenzyme, Proc. Natl. Acad. Sci. USA, 114, 1542-1547,
serine esterases, EMBO J., 5, 1595-1600.
doi: 10.1073/pnas.1620626114.
20.
Masson, D., and Tschopp, J. (1987) A family of serine
37.
Darmon, A.J., Nicholson, D.W., and Bleackley, R.C.
esterases in lytic granules of cytolytic T lymphocytes, Cell,
(1995) Activation of the apoptotic protease CPP32 by cyto
49, 679-685, doi: 10.1016/0092 8674(87)90544 7.
toxic T cell derived granzyme B, Nature, 377, 446-448,
21.
Susanto, O., Trapani, J.A., and Brasacchio, D.
(2012)
doi: 10.1038/377446a0.
Controversies in granzyme biology, Tissue Antigens, 80,
38.
Quan, L.T., Tewari, M., O’Rourke, K., Dixit, V.,
477-487, doi: 10.1111/tan.12014.
Snipas, S.J., Poirier, G.G., Ray, C., Pickup, D.J., and
22.
Vahedi, F., Fraleigh, N., Vlasschaert, C., McElhaney, J.,
Salvesen, G.S. (1996) Proteolytic activation of the cell
and Hanifi Moghaddam, P. (2014) Human granzymes:
death protease Yama/CPP32 by granzyme B, Proc. Natl.
related but far apart, Med. Hypotheses, 83, 688-693,
Acad. Sci. USA,
93,
1972-1976, doi:
10.1073/
doi: 10.1016/j.mehy.2014.09.019.
pnas.93.5.1972.
23.
Trapani, J.A. (2001) Granzymes: a family of lymphocyte
39.
Luthi, A.U., and Martin, S.J. (2007) The CASBAH: a
granule
serine
proteases,
Genome Biol.,
2,
searchable database of caspase substrates, Cell Death Differ.,
reviews3014.1-3014.7, doi:
10.1186/gb 2001 2 12
14, 641-650, doi: 10.1038/sj.cdd.4402103.
reviews3014.
40.
Julien, O., and Wells, J.A. (2017) Caspases and their sub
24.
Sutton, V.R., Wowk, M.E., Cancilla, M., and Trapani, J.A.
strates, Cell Death Differ., 24, 1380-1389, doi: 10.1038/
(2003) Caspase activation by granzyme B is indirect, and
cdd.2017.44.
caspase autoprocessing requires the release of proapoptotic
41.
Han, J., Goldstein, L.A., Gastman, B.R., Froelich, C.J.,
mitochondrial factors, Immunity,
18,
319-329,
Yin, X.M., and Rabinowich, H. (2004) Degradation of
doi: 10.1016/s1074 7613(03)00050 5.
Mcl 1 by granzyme B: implications for Bim mediated
25.
Goping, I.S., Barry, M., Liston, P., Sawchuk, T.,
mitochondrial apoptotic events, J. Biol. Chem., 279,
Constantinescu, G., Michalak, K.M., Shostak, I.,
22020-22029, doi: 10.1074/jbc.M313234200.
БИОХИМИЯ том 85 вып. 2 2020
ГРАНЗИМЫ И МИТОХОНДРИИ
163
42.
Han, J., Goldstein, L.A., Gastman, B.R., Rabinovitz, A.,
56.
Zhu, J., Vinothkumar, K.R., and Hirst, J. (2016) Structure
and Rabinowich, H. (2005) Disruption of Mcl 1⋅Bim com
of mammalian respiratory complex I, Nature,
536,
plex in granzyme B mediated mitochondrial apoptosis, J.
354-358, doi: 10.1038/nature19095.
Biol. Chem.,
280,
16383-16392, doi:
10.1074/jbc.
57.
Fiedorczuk, K., Letts, J.A., Degliesposti, G., Kaszuba, K.,
M411377200.
Skehel, M., and Sazanov, L.A. (2016) Atomic structure of
43.
Matsumura, S., Van De Water, J., Kita, H., Coppel, R.L.,
the entire mammalian mitochondrial complex I, Nature,
Tsuji, T., Yamamoto, K., Ansari, A.A., and Gershwin, M.E.
538, 406-410, doi: 10.1038/nature19794.
(2002) Contribution to antimitochondrial antibody produc
58.
Martinvalet, D., Dykxhoorn, D.M., Ferrini, R., and
tion: cleavage of pyruvate dehydrogenase complex E2 by
Lieberman, J. (2008) Granzyme A cleaves a mitochondrial
apoptosis related proteases, Hepatology,
35,
14-22,
complex I protein to initiate caspase independent cell
doi: 10.1053/jhep.2002.30280.
death, Cell, 133, 681-692, doi: 10.1016/j.cell.2008.03.032.
44.
Siddiqui, W.A., Ahad, A., and Ahsan, H. (2015) The mys
59.
Jacquemin, G., Margiotta, D., Kasahara, A., Bassoy, E.Y.,
tery of BCL2 family: Bcl 2 proteins and apoptosis: an
Walch, M., Thiery, J., Lieberman, J., and Martinvalet, D.
update, Arch. Toxicol., 89, 289-317, doi: 10.1007/s00204
(2015) Granzyme B induced mitochondrial ROS are
014 1448 7.
required for apoptosis, Cell Death Differ., 22, 862-874,
45.
Sarosiek, K.A., Chi, X., Bachman, J.A., Sims, J.J.,
doi: 10.1038/cdd.2014.180.
Montero, J., Patel, L., Flanagan, A., Andrews, D.W.,
60.
Ricci, J.E., Muñoz Pinedo, C., Fitzgerald, P., Bailly
Sorger, P., and Letai, A. (2013) BID preferentially activates
Maitre, B., Perkins, G.A., Yadava, N., Scheffler, I.E.,
BAK while BIM preferentially activates BAX, affecting
Ellisman, M.H., and Green, D.R. (2004) Disruption of
chemotherapy response, Mol. Cell,
51,
751-765,
mitochondrial function during apoptosis is mediated by
doi: 10.1016/j.molcel.2013.08.048.
caspase cleavage of the p75 subunit of complex I of the elec
46.
Ben Safta, T., Ziani, L., Favre, L., Lamendour, L., Gros, G.,
tron transport chain, Cell, 117, 773-786, doi: 10.1016/
Mami Chouaib, F., Martinvalet, D., Chouaib, S., and
j.cell.2004.05.008.
Thiery, J. (2015) Granzyme B activated p53 interacts with
61.
Hirst, J., and Roessler, M.M. (2016) Energy conversion,
Bcl 2 to promote cytotoxic lymphocyte mediated apoptosis,
redox catalysis and generation of reactive oxygen species by
J. Immunol., 194, 418-428, doi: 10.4049/jimmunol.1401978.
respiratory complex I, Biochim. Biophys. Acta, 1857,
47.
Wang, S., Xia, P., Shi, L., and Fan, Z. (2012) FADD cleav
872-883, doi: 10.1016/j.bbabio.2015.12.009.
age by NK cell granzyme M enhances its self association to
62.
Rodenburg, R.J. (2016) Mitochondrial complex I linked
facilitate procaspase 8 recruitment for auto processing
disease, Biochim. Biophys. Acta,
1857,
938-945,
leading to caspase cascade, Cell Death Differ., 19, 605-615,
doi: 10.1016/j.bbabio.2016.02.012.
doi: 10.1038/cdd.2011.130.
63.
Chiusolo, V., Jacquemin, G., Yonca Bassoy, E., Vinet, L.,
48.
Hou, Q., Zhao, T., Zhang, H., Lu, H., Zhang, Q., Sun, L.,
Liguori, L., Walch, M., Kozjak Pavlovic, V., and
and Fan, Z. (2008) Granzyme H induces apoptosis of target
Martinvalet, D. (2017) Granzyme B enters the mitochon
tumor cells characterized by DNA fragmentation and Bid
dria in a Sam50 , Tim22 and mtHsp70 dependent manner
dependent mitochondrial damage, Mol. Immunol., 45,
to induce apoptosis, Cell. Death Differ., 24, 747-758,
1044-1055, doi: 10.1016/j.molimm.2007.07.032.
doi: 10.1038/cdd.2017.3.
49.
Ewen, C.L., Kane, K.P., and Bleackley, R.C. (2013)
64.
Martinvalet, D. (2019) Mitochondrial entry of cytotoxic
Granzyme H induces cell death primarily via a Bcl 2 sensi
proteases: a new insight into the granzyme B cell death
tive mitochondrial cell death pathway that does not require
pathway, Oxid. Med. Cell. Longev.,
2019,
9165214,
direct Bid activation, Mol. Immunol., 54, 309-318,
doi: 10.1155/2019/9165214.
doi: 10.1016/j.molimm.2012.12.020.
65.
Grivennikova, V.G., and Vinogradov, A.D.
(2006)
50.
Shi, L., Wu, L., Wang, S., and Fan, Z. (2009) Granzyme F
Generation of superoxide by the mitochondrial complex I,
induces a novel death pathway characterized by Bid inde
Biochim. Biophys. Acta, 1757, 553-561, doi: 10.1016/
pendent cytochrome c release without caspase activation,
j.bbabio.2006.03.013.
Cell Death Differ.,
16,
1694-1706, doi:
10.1038/
66.
Murphy, M.P. (2009) How mitochondria produce reactive
cdd.2009.101.
oxygen species, Biochem. J., 417, 1-13, doi: 10.1042/
51.
Van Damme, P., Maurer Stroh, S., Hao, H., Colaert, N.,
BJ20081386.
Timmerman, E., Eisenhaber, F., Vandekerckhove, J., and
67.
Korge, P., Calmettes, G., and Weiss, J.N. (2016) Reactive
Gevaert, K. (2010) The substrate specificity profile of
oxygen species production in cardiac mitochondria after
human granzyme A, Biol. Chem.,
391,
983-997,
complex I inhibition: modulation by substrate dependent
doi: 10.1515/BC.2010.096.
regulation of the NADH/NAD+ ratio, Free Radic. Biol.
52.
Beresford, P.J., Zhang, D., Oh, D.Y., Fan, Z., Greer, E.L.,
Med., 96, 22-33, doi: 10.1016/j.freeradbiomed.2016.04.002.
Russo, M.L., Jaju, M., and Lieberman, J. (2001) Granzyme
68.
Robb, E.L., Hall, A.R., Prime, T.A., Eaton, S., Szibor, M.,
A activates an endoplasmic reticulum associated caspase
Viscomi, C., James, A.M., and Murphy, M.P. (2018)
independent nuclease to induce single stranded DNA
Control of mitochondrial superoxide production by reverse
nicks, J. Biol. Chem., 276, 43285-43293, doi: 10.1074/
electron transport at complex I, J. Biol. Chem., 293,
jbc.M108137200.
9869-9879, doi: 10.1074/jbc.RA118.003647.
53.
Fan, Z., Beresford, P.J., Oh, D.Y., Zhang, D., and
69.
Shidoji, Y., Hayashi, K., Komura, S., Ohishi, N., and Yagi,
Lieberman, J. (2003) Tumor suppressor NM23 H1 is a
K.
(1999) Loss of molecular interaction between
granzyme A activated DNase during CTL mediated apop
cytochrome c and cardiolipin due to lipid peroxidation,
tosis, and the nucleosome assembly protein SET is its
Biochem. Biophys. Res. Commun.,
264,
343-347,
inhibitor, Cell,
112,
659-672, doi:
10.1016/s0092
doi: 10.1006/bbrc.1999.1410.
8674(03)00150 8.
70.
Kagan, V.E., Tyurin, V.A., Jiang, J., Tyurina, Y.Y., Ritov, V.B.,
54.
Martinvalet, D., Zhu, P., and Lieberman, J.
(2005)
Amoscato, A.A., Osipov, A.N., Belikova, N.A., Kapralov, A.A.,
Granzyme A induces caspase independent mitochondrial
Kini, V., Vlasova, I.I., Zhao, Q., Zou, M., Di, P.,
damage, a required first step for apoptosis, Immunity, 22,
Svistunenko, D.A., Kurnikov, I.V., and Borisenko, G.G.
355-370, doi: 10.1016/j.immuni.2005.02.004.
(2005) Cytochrome c acts as a cardiolipin oxygenase
55.
Гривенникова В.Г., Виноградов А.Д. (2003) Митохондри
required for release of proapoptotic factors, Nat. Chem.
альный комплекс I, Успехи биологической химии, 43, 19-58.
Biol., 1, 223-232, doi: 10.1038/nchembio727.
БИОХИМИЯ том 85 вып. 2 2020
164
КИСЕЛЕВСКИЙ
71. Lucken Ardjomande, S., and Martinou, J. C.
(2008)
кова Е.Н., Ефимов Г.А., Савченко В.Г. (2016) Примене
Granzyme A, a stealth killer in the mitochondrion, Cell,
ние рекомбинантных МНС тетрамеров для изоляции
133, 568-570, doi: 10.1016/j.cell.2008.04.031.
вирусспецифичных CD8+ клеток здоровых доноров:
72. Dotiwala, F., Mulik, S., Polidoro, R.B., Ansara, J.A.,
потенциальный подход к клеточной терапии посттранс
Burleigh, B.A., Walch, M., Gazzinelli, R.T., and
плантационной цитомегаловирусной инфекции, Био(
Lieberman, J. (2016) Killer lymphocytes use granulysin,
химия, 81, 1628-1642, doi: 10.1134/ S0006297916110146.
perforin and granzymes to kill intracellular parasites, Nat.
76. Biron, C.A., and Brossay, L. (2001) NK cells and NKT cells
Med., 22, 210-216, doi: 10.1038/nm.4023.
in innate defense against viral infections, Curr. Opin.
73. Kang, S., Brown, H.M., and Hwang, S. (2018) Direct
Immunol.,
13,
458-464, doi:
10.1016/s0952
antiviral mechanisms of interferon gamma, Immune Netw.,
7915(00)00241 7.
18, e33, doi: 10.4110/in.2018.18.e33.
77. Ruella, M., and Kalos, M. (2014) Adoptive immunothera
74. Hansen, T.H., and Bouvier, M. (2009) MHC class I antigen
py for cancer, Immunol. Rev., 257, 14-38, doi: 10.1111/
presentation: learning from viral evasion strategies, Nat.
imr.12136.
Rev. Immunol., 9, 503-513, doi: 10.1038/nri2575.
78. Kim, N., Lee, H.H., Lee, H.J., Choi, W.S., Lee, J., and
75. Вдовин А.С., Филькин С.Ю., Ефимова П.Р., Шитиков С.А.,
Kim, H.S. (2019) Natural killer cells as a promising thera
Капранов Н.М., Давыдова Ю.О., Егоров Е.С., Хамага
peutic target for cancer immunotherapy, Arch. Pharm. Res.,
нова Е.Г., Дроков М.Ю., Кузьмина Л.А., Паровични
42, 591-606, doi: 10.1007/s12272 019 01143 y.
GRANZYMES AND MITOCHONDRIA
Review
D. B. Kiselevsky
Lomonosov Moscow State University, Faculty of Biology, 119991 Moscow, Russia; E(mail: dkiselevs@mail.ru
Received October 1, 2019
Revised November 1, 2019
Accepted November 4, 2019
Cytotoxic T lymphocytes and natural killers eliminate infected cells from an organism by triggering programmed cell
death (apoptosis). The contents of the lytic granules of killer cells, including pore forming proteins perforins and pro
teolytic enzymes granzymes, are released with subsequent penetration into target cells. Granzyme B initiates mito
chondria dependent apoptosis via (i) proapoptotic Bid protein, (ii) Mcl 1 and Bim proteins, or (iii) p53 protein. As
a result, cytochrome c is released from mitochondria into cytoplasm, causing formation of apoptosomes that provide
proteolytic cascade of caspases’ activation. Granzymes M, H, and F cause cell death, which is accompanied by the
release of cytochrome c from mitochondria. Granzyme A induces generation of reactive oxygen species, which pro
mote translocation of the endoplasmic reticulum associated SET complex to cell nucleus. In cell nucleus granzyme
A cleaves the SET thus activating nucleases that cause single strand DNA breaks. Granzymes A and B penetrate mito
chondria and cut the complex I subunits of the respiratory chain. One of the complex I subunits is a target for the cas
pase 3 as well. Granzyme dependent damage to complex I leads to ROS generation and cell death.
Keywords: reactive oxygen species, apoptosis, granzymes, mitochondria, programmed cell death
БИОХИМИЯ том 85 вып. 2 2020