БИОХИМИЯ, 2020, том 85, вып. 2, с. 174 - 196
УДК 543.94
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
НУКЛЕИНОВЫХ КИСЛОТ И ИХ ПРИМЕНЕНИЕ В БИОАНАЛИЗЕ
Обзор
© 2020
О.Л. Бодулев, И.Ю. Сахаров*
Московский государственный университет им. М.В. Ломоносова, химический факультет,
119991 Москва, Россия; электронная почта: sakharovivan@gmail.com
Поступила в редакцию 13.08.2019
После доработки 01.11.2019
Принята к публикации 01.11.2019
В последние годы бурно развиваются методы изотермической амплификации нуклеиновых кислот как аль
тернатива полимеразной цепной реакции (ПЦР). Их использование позволяет проводить амплификацию
нуклеиновых кислот при постоянной температуре в отличие от ПЦР, для которой требуется циклическое
изменение температуры. Кроме того, при использовании изотермических методов амплификация может
проводиться непосредственно в живых клетках. В настоящем обзоре в краткой форме изложены принципы
известных изотермических методов амплификации, а также продемонстрирована их высокая эффектив
ность при конструировании новых высокочувствительных методов анализа как нуклеиновых кислот, так и
ферментов, ответственных за их модификацию. Более того, приведены данные об успешном использовании
изотермических методов амплификации в анализе клеток и биомолекул, определение которых проводится
с применением ДНК/РНК аптамеров.
КЛЮЧЕВЫЕ СЛОВА: нуклеиновые кислоты, амплификация, изотермическая, биоанализ, аптамеры.
DOI: 10.31857/S0320972520020037
Количественное и качественное определе
но, число исследований в этой области будет
ние нуклеиновых кислот является чрезвычайно
продолжать расти в следующие десятилетия.
важным направлением в современной биоло
Обнаружение ДНК/РНК патогенных бактерий
гии и медицине. Начиная с 1990 х гг., данная
и вирусов может иметь решающее значение для
область науки активно развивается, и, вероят
выбора правильной стратегии лечения пациен
тов. Недавние открытия указали на взаимо
Принятые сокращения: LAMP - петлевая изотер
связь между восприимчивостью людей к неко
мическая амплификация (Loop mediated isothermal
торым заболеваниям и однонуклеотидными
Amplification); NASBA - метод амплификации, основан
полиморфизмами или короткими вставка
ный на последовательности нуклеиновых кислот (Nucleic
ми/делециями. Также было показано, что гены
Acid Sequence Based Amplification); HDA - хеликаза зави
симая амплификация (Helicase Dependent Amplification);
микроРНК человека часто встречаются вблизи
EXPAR - экспоненциальная реакция амплификации нук
геномных регионов и сайтов, связанных с ра
леиновых кислот (Exponential Amplification Reaction);
ком. Уровень экспрессии некоторых мик
SDA - амплификация с замещением цепи (Strand
роРНК у пациентов с хроническим лимфолей
Displacement Amplification); RPA - рекомбиназная полиме
козом, колоректальной неоплазией, лимфомой
разная амплификация (Recombinase Polymerase Amplifica
tion); RCA - метод катящегося кольца (Rolling Circle
Беркитта, раком легких, крупноклеточными
Amplification); WGA - полногеномная амплификация
лимфомами, глиобластой и другими заболева
(Whole Genome Amplification); MDA - амплификация со ниями отличается от уровня экспрессии в нор
множественным замещением цепи (Multiple Displacement
мальных тканях.
Amplification); pWGA - полногеномный метод амплифика
ции с применением праймазы (Primase based Whole
Помимо врачей в развитии высокочувстви
Genome Amplification); EASA - метод экзонуклеаза III за
тельных методов анализа нуклеиновых кислот
висимой циклизации аналита (Exonuclease III Assisted чрезвычайно заинтересованы специалисты пи
Signal Amplification); ICSDP - амплификационный метод с щевой химии, т.к. эти методы с большой досто
полимеризацией и замещением (Isothermal Circular Strand
верностью и точностью позволяют проводить
Displacement Polymerization); HCR - реакция цепной гиб
ридизации (Hybridization Chain Reaction); CHA - метод ка
оценку качества пищевых продуктов [1]. Мето
талитической сборки шпилек (Catalytic Hairpin Assembly).
ды анализа нуклеиновых кислот также давно и
* Адресат для корреспонденции.
успешно используются в криминалистике [2].
174
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
175
Обнаружение последовательностей нуклеи
ется описание известных к настоящему времени
новых кислот без предварительной очистки в
изотермических методов амплификации, их
биологических образцах и непосредственно в
преимуществ и ограничений применения в био
живых организмах является крайне актуальной
анализе.
задачей. В основе таких методов анализа лежит
Описанные к настоящему времени изотер
реакция гибридизации, что определяет их высо
мические методы амплификации нуклеиновых
кую селективность. С учетом того, что концент
кислот можно разделить на две группы: 1) мето
рации нуклеиновых кислот в исследуемых об
ды амплификации, осуществляющие повыше
разцах практически всегда крайне низкие, а их
ние аналитического сигнала за счет увеличения
изменения в случае патологии могут быть не
концентрации аналита; 2) методы амплифика
значительными, требуются методы с чрезвычай
ции, позволяющие повышать аналитический
но высокой чувствительностью и низким преде
сигнал без изменения концентрации аналита.
лом обнаружения. Для этого в настоящее время
детектирующие методы анализа ДНК/РНК соп
рягаются с различными вариантами полимераз
МЕТОДЫ АМПЛИФИКАЦИИ,
ной цепной реакции (ПЦР). Данный метод ши
ОСУЩЕСТВЛЯЮЩИЕ ПОВЫШЕНИЕ
роко используется на практике и является высо
АНАЛИТИЧЕСКОГО СИГНАЛА ЗА СЧЕТ
коэффективным, т.к. он позволяет синтезиро
УВЕЛИЧЕНИЯ КОНЦЕНТРАЦИИ АНАЛИТА
вать до 109 копий (ампликонов) анализируемой
последовательности. Однако данный метод
Во всех методах изотермической амплифи
имеет ряд недостатков. Так, при проведении
кации, направленных на повышение концент
ПЦР возможна неспецифическая гибридиза
рации аналита, используются ферменты.
ция, приводящая к накоплению посторонних
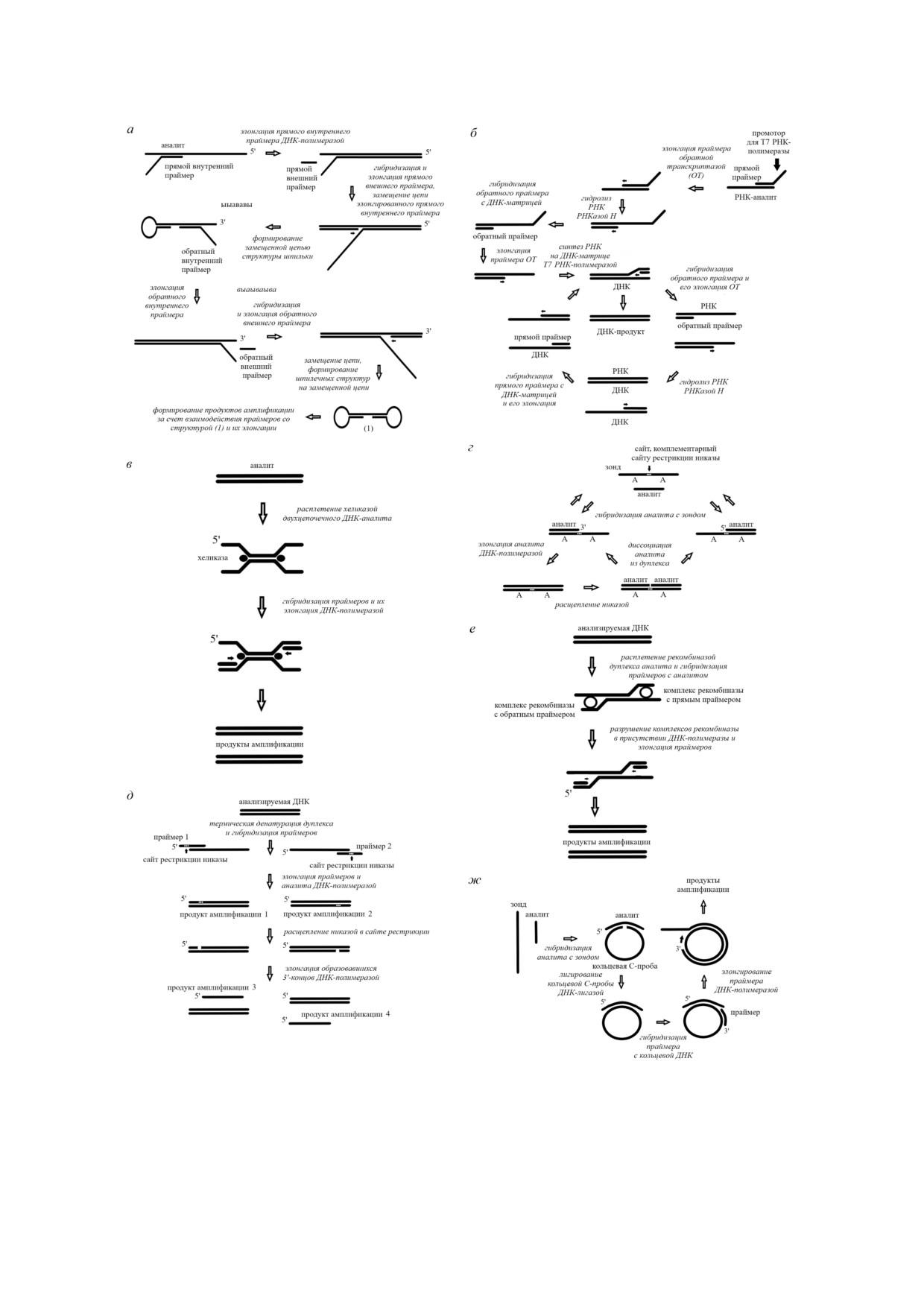
На рис. 1, а приведена схема петлевой изотер1
продуктов.
мической амплификации (LAMP). Данный метод
Для количественного определения нуклеи
впервые был описан Notomi et al. в 2000 г. [6]. В
новых кислот широко используется ПЦР с де
этом методе используются несколько прайме
текцией продукта в режиме реального времени
ров (чаще четыре, но иногда и шесть), компле
(ПЦР РВ) [3]. Существуют два основных вари
ментарных разным участкам определяемой
анта ПЦР РВ. В первом варианте используется
ДНК, а также ДНК полимеразы с высокой спо
Taq ДНК полимераза и линейный зонд (техно
собностью к замещению цепей. LAMP прово
логия TaqMan), во втором варианте - интерка
дится при температуре 60 °С.
лирующие красители, такие как SYBR Green,
Данная реакция инициируется прямым
Eva Green, BOXTO и др., флуоресценция кото
внутренним праймером, который на первом
рых значительно возрастает при их связывании
этапе гибридизуется с комплементарным фраг
с двухцепочечной молекулой ДНК. Линейный
ментом молекулы ДНК аналита, расположен
диапазон ПЦР РВ лежит в диапазоне от 10 до
ным в районе 5′ конца, а затем достраивается
5 . 109 копий анализируемой последовательности
ДНК полимеразой. На следующем этапе пря
[4]. Чувствительность метода сильно варьирует
мой внешний праймер гибридизируется с
и зависит от структуры использованных прай
5′ концовым фрагментом аналита, после чего
меров. Для ПЦР РВ характерна высокая вос
также элонгируется полимеразой, при этом вы
производимость.
тесняя ранее синтезированную последователь
Циклическое изменение температуры, тре
ность. Затем обратный внутренний праймер
буемое при проведении методов с применением
взаимодействует с комплементарным участком
ПЦР, способствует неспецифической гибриди
вновь синтезированной последовательности,
зации праймеров и ампликонов [5]. Более того,
расположенным вблизи 3′ конца. После достра
для осуществления циклического изменения
ивания этого праймера с синтезированной по
температуры в ходе ПЦР необходимо использо
следовательностью ДНК гибридизуется обрат
вание дорогостоящего оборудования. Следует
ный внешний праймер, который также фермен
также отметить, что в силу необходимости про
тативно достраивается, при этом вытесняя ра
ведения дуплексного плавления в ходе ПЦР, ко
нее синтезированную последовательность. На
торое проводится путем нагревания, данный
концах обеих синтезированных последователь
метод не может быть использован для обнару
ностей за счет комплементарных взаимодей
жения нуклеиновых кислот в живых клетках.
ствий формируются петлеобразные структуры.
Вышеупомянутые ограничения ПЦР стиму
Таким образом, в результате проведения указан
лировали разработку различных изотермичес
ных выше реакций синтезируются две структу
ких амплификационных платформ для обнару
ры с петлями на обоих концах. Получение таких
жения ДНК/РНК. Целью данного обзора явля
молекул инициирует последующие циклы амп
БИОХИМИЯ том 85 вып. 2 2020
176
БОДУЛЕВ, САХАРОВ
Рис. 1. Схемы изотермических методов амплификации нуклеиновых кислот с применением полимераз: а - петлевая изо
термическая амплификация ДНК (LAMP); б - метод амплификации, основанный на последовательности нуклеиновых
кислот (NASBA); в - хеликаза зависимая амплификация ДНК (HDA); г - экспоненциальная реакция амплификации
нуклеиновых кислот (EXPAR); д - метод амплификации с замещением цепи (SDA); е - рекомбиназная полимеразная
амплификация (РПА); ж - амплификация по принципу катящегося кольца (RCA)
БИОХИМИЯ том 85 вып. 2 2020
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
177
лификации с использованием тех же праймеров
зарегистрированы ложноположительные резуль
[7].
таты [15]. LAMP также был успешно применен
Амплификация LAMP носит экспоненци
для выявления in situ Salmonella в пищевых про
альный характер и позволяет получить до 109 ко
дуктах и stxA2 в Escherichia coli O157:H7 [15а, 15б].
пий ДНК в течение 15-60 мин. Использование
Недавно при исследовании штаммов
нескольких праймеров обеспечивает высокую
Burkholderia mallei и Burkholderia pseudomallei бы
специфичность копирования. При применении
ло показано, что с помощью LAMP праймеры
обратной транскрипции LAMP позволяет также
выявляли ДНК не всех анализируемых штам
амплифицировать молекулы РНК.
мов, к генам мишеням которых они были смо
Продукты реакции, образующиеся в LAMP,
делированы, но были способны направлять
чаще всего регистрируются либо с помощью
синтез фрагментов генов гетерологичных штам
электрофореза, либо по измерению мутности
мов [16]. По мнению авторов, неудовлетвори
среды, которая изменяется в результате образо
тельные результаты, полученные при проведе
вания пирофосфата магния [8]. Концентрация
нии LAMP, могут быть обусловлены наличием
образующегося в ходе LAMP пирофосфата мо
GC богатых областей в геноме исследованных
жет также оцениваться с помощью флуоресцент
бактерий и формированием вторичных структур
ного красителя кальцеина (флуорексона) [9].
при температуре проведения LAMP.
Для детекции образовавшейся ДНК использует
Следует отметить, что LAMP проявляет бо
ся интеркалирование различных красителей [10,
лее высокую специфичность относительно ПЦР
11]. Благодаря своей простоте, LAMP прекрасно
за счет использования как минимум шести
сочетается с микрофлюидными технологиями,
участков связывания праймера с анализируемой
что позволяет автоматизировать метод с одно
последовательностью. Утверждается, что
временным уменьшением времени анализа, а
чувствительность LAMP выше чувствительнос
также сократить расход реагентов [11].
ти ПЦР на порядок. Более того, LAMP менее
LAMP широко используется как скрининго
чувствительна к ингибиторам, присутствующим
вый метод, т.к. он проводится при постоянной
в биологических образцах [17]. Серьезным не
температуре, а также является высокоэффек
достатком данного метода является высокий
тивным и специфичным. В литературе описано
риск контаминации, часто приводящий к полу
применение LAMP для выявления палочки Ко
чению ложноположительных результатов в от
ха, герпеса, тяжелого острого респираторного
рицательных контролях [18].
синдрома, сибирской язвы, а также вирусов
Другим изотермическим методом амплифи
гриппа человека, птицы и др. [12, 13]. Метод,
кации нуклеиновых кислот, приводящим к по
разработанный для определения Leptospira с
вышению концентрации аналита, является ме1
применением LAMP, позволил детектировать
тод амплификации, основанный на последова1
ДНК данного патогена в концентрации от 200
тельности нуклеиновых кислот* (NASBA). В этом
пг/мл. Специфичность, оценка которой прово
методе, разработанном Дж. Комптоном в 1991 г.,
дилась с использованием 172 бактериальных
амплификация молекул РНК проводится при
штаммов, составила 100% [12].
участии трех ферментов, таких как обратная
С помощью LAMP диагностируются не
транскриптаза вируса миелобластоза птиц,
только бактериальные патогены и вирусы, но
РНКаза H и Т7 РНК полимераза [19]. NASBA
также и заболевания, вызываемые простейши
включает в себя две стадии: ацикличную и цик
ми. Одним из таких патогенов является маля
личную (рис. 1, б).
рийный плазмодий. В исследовании Poschl et al.
На первой стадии денатурированная при
[14] в качестве метода сравнения был использо
65 °С РНК взаимодействует со специфическим
ван метод ПЦР, специфичность LAMP для
праймером, содержащим промоторную после
Plasmodium falciparum составила 100%. Все ПЦР
довательность Т7 РНК полимеразы. В присут
отрицательные образцы для данного возбудите
ствии обратной транскриптазы фермент синте
ля также были отрицательными при диагности
зирует последовательность ДНК по одноцепо
ке с помощью LAMP. При диагностике
чечной РНК, формируя гибрид РНК/ДНК.
Plasmodium vivax методом LAMP обнаружены 22
Данная реакция, как и все последующие, прово
из 23 ПЦР положительных образцов. Все 82
дится обычно при 41 оС. Полученный гибрид
ПЦР отрицательных образца также были отри
цательными в анализе с использованием LAMP.
* Хотя, по мнению авторов, название данного метода явля
ется чрезвычайно неинформативным, именно оно исполь
Таким образом, для диагностики видов
зуется в научной англоязычной литературе. Более верно, с
Plasmodium применение метода LAMP так же
нашей точки зрения, было бы назвать его методом изотер*
оправданно, как и ПЦР. К сожалению, при ис
мической амплификации РНК, основанным на последова*
пользовании LAMP в некоторых случаях были
тельности нуклеиновых кислот.
БИОХИМИЯ том 85 вып. 2 2020
178
БОДУЛЕВ, САХАРОВ
подвергается расщеплению РНКазой H, а обра
к ингибиторам, присутствующим в биологичес
зующаяся однонитевая ДНК реагирует со вторым
ких образцах, чем ПЦР [25].
праймером, который достраивается с образова
Хеликаза1зависимая амплификация (HDA)
нием двухцепочечной ДНК. Далее при участии
является аналогом ПЦР, расплетение цепей
Т7 РНК полимеразы на матрице ДНК синтези
ДНК проводится не за счет повышения темпе
руются РНК, комплементарные анализируемой
ратуры, а под действием хеликазы (рис. 1, в)
ДНК. Образующиеся молекулы РНК взаимодей
[26]. В этом методе используются хеликаза,
ствуют со вторым праймером, в процессе достра
ДНК полимераза и белки, связывающие одно
ивания он образует гибрид РНК/ДНК, который,
цепочечные ДНК. На первом этапе амплифика
в свою очередь, расщепляется РНКазой Н. На
ционного цикла к обоим концам двухцепочеч
этом этапе образующаяся одноцепочечная ДНК
ной ДНК присоединяются молекулы хеликазы,
реагирует с первым праймером, который удли
что приводит к расплетению дуплекса. С высво
няется в присутствии обратной транскриптазы,
божденной из дуплекса однонитевой ДНК, ста
а Т7 РНК полимераза синтезирует копии исход
билизированной ДНК связывающими белка
ной РНК, благодаря чему происходит запуск
ми, гибридизуются прямой и обратный прайме
следующего цикла амплификации.
ры, после чего в присутствии ДНК полимеразы
Преимуществом NASBA относительно ПЦР
осуществляется их элонгация. Таким образом,
с обратной транскрипцией является использо
при завершении первого цикла HDA из одной
вание одной и той же реакционной среды для
молекулы ДНК образуется две. В дальнейшем
реакции обратной транскрипции и последую
этот процесс многократно повторяется, приводя
щей реакции амплификации. Показано, что
к увеличению концентрации исследуемого ана
NASBA обладает большей чувствительностью и
лита. Данный метод характеризуется экспонен
является менее затратным по времени, чем
циальной кинетикой амплификации, его эф
ПЦР. Эффективность этого экспоненциального
фективность достигает 107 копий аналита [27].
метода составляет 109 копий аналита. Следует
Следует отметить, что HDA обычно проводится
отметить, что, несмотря на высокую эффектив
при двух температурах: при 37 °С или в интерва
ность экспоненциальных амплификационных
ле между 60 и 65 °С. При пониженной темпера
методов, для них характерна неспецифическая
туре применяется репаративный белок MutL, в
амплификация, которая приводит к получению
то время как при более высоких температурах
ложноположительных результатов.
использование данного белка не требуется. Сле
Сразу после разработки NASBA был приме
дует отметить, что понижение температуры про
нен для диагностики ВИЧ инфекции в сыво
ведения амплификации повышает концентра
ротке крови пациентов [20]. В настоящее время
цию неспецифических продуктов амплифика
данный метод широко используется при выяв
ции, что, в свою очередь, может приводить к по
лении бактерий рода Salmonella, вирусов гепати
явлению ложноположительных результатов
та и папилломы, энтеровирусов человека.
[28].
NASBA был также применен для определения
Важно отметить, что степень фоновой амп
мРНК и микроРНК [21, 22].
лификации в HDA выше, чем в ПЦР [29]. Для
Обычно продукты NASBA регистрируются
минимизации взаимодействия праймеров меж
методом электрофореза с использованием эти
ду собой описаны способы их модификации
дия бромида. Также применяются микрофлюид
[30]. Другой подход для минимизации фоновой
ные системы и биочипы с использованием зон
амплификации - использование таких соедине
дов, меченных флуоресцентными красителями
ний, как диметилсульфоксид, бетаин, сорбитол,
или пероксидазой [11, 21]. С использованием в
которые, к сожалению, также могут ингибиро
качестве метки пероксидаза подобного ДНКзи
вать и полимеразу. Для той же цели используют
ма NASBA был применен для детекции и диф
ся высокомолекулярные краудинг агенты, та
ференциации штаммов вируса чумы свиней
кие как, например, полиэтиленгликоль. Они
[23]. NASBA используется и в сочетании с мето
повышают эффективность полимеразы, одно
дом планшетного олигонуклеотидного анализа.
временно уменьшая степень взаимодействия
Такой подход был успешно применен для опре
праймеров между собой [28].
деления реовируса белого амура [24]. Разрабо
При сравнении методов HDA с флуоресцент
танный метод способен специфически детекти
ной детекцией и ПЦР РВ было показано, что
ровать 14 копий/мкл в течение 5 ч.
при использовании ПЦР РВ наблюдаются
Следует отметить, что в случае применения
меньшие значения предела обнаружения анали
NASBA довольно часто регистрируются ложно
та, а полученные концентрационные зависи
положительные и ложноотрицательные резуль
мости лучше линеаризуются. Так, при анализе
таты. В то же время NASBA менее чувствителен
ДНК Mycobacterium tuberculosis методом HDA
БИОХИМИЯ том 85 вып. 2 2020
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
179
предел обнаружения составил 1 фМ, в то время
присоединен к 3′ концу зонда, что позволяет
как для ПЦР РВ - 100 аМ [28].
ДНК полимеразе элонгировать анализируемую
Описано применение метода HDA в клини
последовательность. При образовании дуплекса
ческом анализе для выявления различных бак
завершается формирование специфического
териальных и вирусных инфекций, а также па
сайта расщепления никазы. Последующий фер
тогенов в воде и пище. Обычно продукты HDA
ментативный гидролиз дуплекса никазой при
регистрируются методом электрофореза с при
водит к его диссоциации с появлением допол
менением этидия бромида. Кроме того, приме
нительной копии аналита. В EXPAR широко ис
няются латеральные проточные методы, биочи
пользуются никазы Nt.BbvCI, Nb.BbvCI, AlwI,
пы и электрохимические биосенсоры. Так, в ра
Nt.AlwI, Nb.BssSI, Nt.BsmAI и Nb.BtsI.
боте Tang et al. [31] метод HDA был использован
В следующем цикле повторно происходит
в сочетании с латеральным проточным методом
элонгация аналита ДНК полимеразой и рас
для определения Salmonella typhimurium в воде,
щепление синтезированной последовательнос
предел обнаружения составил 100 КОЕ/мл.
ти никазой. В EXPAR используются такие ком
Метод HDA с электрохимической детекцией
мерчески доступные ферменты, как полимераза
сигнала был использован для количественного
phi 29, фрагмент Кленова, полимеразы Vent и
определения бактерий рода Salmonella [32]. Мо
Bst, у которых отсутствует 3′-5′ экзонуклеазная
лекулы захватывающей ДНК иммобилизовали
активность. Хотя оптимальным температурным
на поверхности электрода, а один из праймеров
интервалом для проведения EXPAR является
модифицировали флуоресцеином. В качестве
55-60 °С, эта реакция может проводиться и при
детектирующей системы использовали конъю
37 °С. В последнем случае предпочтительно ис
гат антител против флуоресцеина с пероксида
пользовать фрагмент Кленова [36]. Более высо
зой хрена. Предел обнаружения составил 10 ко
кая температура амплификации повышает эф
пий ДНК.
фективность и специфичность EXPAR.
Данный метод был применен также для детек
Таким образом, EXPAR осуществляет экспо
ции патогена Phytophthora kernoviae в листьях рас
ненциальное накопление молекул аналита. Сле
тений с использованием ДНК чипов [33]. В этом
дует отметить, что данный метод амплификации
случае предел обнаружения составил 10 нг/мл.
может использоваться исключительно для амп
Объединение HDA и планшетного олигофермент
лификации коротких олигонуклеотидов и поз
ного анализа позволило разработать колоримет
воляет достигать эффективности 108 копий ана
рический метод для идентификации и количест
лита [27].
венного определения Karlodinium veneficum и
В отличие от других методов амплификации,
Karlodinium armiger [34]. Предел обнаружения
в которых короткие праймеры используются в
данного метода составил 50 КОЕ/мл. Разрабо
высокой концентрации, в данном методе в вы
танный метод был применен для выявления
сокой концентрации применяется зонд, по дли
Karlodinium spp. в морской воде.
не превосходящий анализируемую последова
Кроме детекции патогенов HDA был успешно
тельность в 2 раза. В связи с этим возможна ди
использован при разработке метода анализа мик
меризация зонда с последующим образованием
роРНК, перспективных онкомаркеров. Так, Ma
неспецифических продуктов амплификации.
et al. [35] описали чувствительный флуоресцент
Степень фоновой амплификации зависит от
ный метод определения микроРНК miR 21. Пре
строения используемых зондов. Так, наличие в
дел обнаружения этого метода составил 12,8 фМ,
зонде фрагментов, обогащенных нуклеотидами
линейный диапазон - от 100 фМ до 10 нМ.
G и A, может усугублять фоновую амплифика
В методе, получившем название экспоненци1
цию в силу связывания полимераз с пуриновы
альная реакция амплификации нуклеиновых кис1
ми основаниями. Отмечается, что фоновая амп
лот (EXPAR) [36], используется зонд, который
лификация может быть также следствием фор
представляет собой две одинаковые последова
мирования зондом структуры шпильки. На се
тельности, комплементарные анализируемой
годняшний день не разработан эффективный
нуклеиновой кислоте и соединенные между со
способ подавления фоновой амплификации в
бой через сайт рестрикции никазы (рис. 1, г). В
EXPAR [36].
процессе определения аналит гибридизуется с
Впервые использование EXPAR для детек
одной из копий зонда с образованием дуплексов
ции ДНК олигонуклеотидов было описано в
двух типов. Один из дуплексов, в котором ана
2003 г. [37]. Впоследствии EXPAR был применен
лит присоединен к 5′ концу зонда, не может
для повышения чувствительности флуоресцент
быть элонгирован полимеразой и достаточно
ного метода количественного определения
быстро диссоциирует при температуре проведе
фрагмента мРНК гена p53. Предел обнаружения
ния EXPAR (60 °С). В другом дуплексе аналит
при его детекции в реальном времени составил
БИОХИМИЯ том 85 вып. 2 2020
180
БОДУЛЕВ, САХАРОВ
10 фМ, рабочий диапазон - от 10 фМ до 10 нМ
перспективность его использования в медико
[38]. В работе Li et al. [39] сочетание колоримет
биологических исследованиях.
рического метода детекции с использованием
За счет своей высокой специфичности дан
золотых наночастиц и EXPAR было применено
ный метод амплификации был успешно приме
для анализа микроРНК. Достигнутый предел
нен для выявления однонуклеотидных полимор
обнаружения составил 46 фМ, линейный диапа
физмов. Так, в работе Shi et al. [45] описан хеми
зон - от 50 фМ до 10 нМ. Кроме того, EXPAR
люминесцентный метод детекции точечных му
был успешно применен для детекции метилиро
таций. Использование в данном методе магнит
вания ДНК и мутаций РНК.
ных частиц позволило значительно понизить ве
EXPAR с использованием аптамеров был
личину предела обнаружения (0,1 фМ).
применен для определения белковых молекул, а
SDA может применяться для определения
именно тромбина, тромбоцитарного фактора
высокомолекулярных РНК, состоящих из сотен
роста и муцина 1 [36, 40]. Этот метод также ис
и даже тысяч нуклеотидов, таких как вирусная
пользовался для определения катионов ртути,
РНК, мРНК и рРНК. Zhao et al. предложили
предел обнаружения которых составил 100 пМ
двухэтапные колориметрический и флуоресцент
[41].
ный методы, в которых сначала проводится рас
Метод EXPAR позволяет повысить чувстви
щепление РНК ДНКзимом, а затем амплифика
тельность методов определения некоторых фер
ция с помощью SDA [46]. Этот метод был также
ментов. Описаны методы определения актив
применен для количественного определения
ности теломеразы (в клетках HeLa) [42], метил
микроРНК и раковых клеток [47, 48]. При де
трансферазы и урацил ДНК гликозидазы [36,
текции микроРНК предел обнаружения соста
43].
вил 16 фМ, линейный диапазон - от 16 фМ до
Механизм метода амплификации с замещени1
100 нМ. Предел обнаружения клеток Рамоса
ем цепи (SDA), впервые описанного в 1992 г., ос
был равен 45 клеток/мл, а линейный диапазон
нован на циклической реакции, включающей в
данного метода - 45-1000 клеток/мл.
себя стадии полимеризации, расщепления и за
На основе SDA Ding et al. [49] разработали
мещения [44]. Схема SDA представлена на
чувствительный метод определения активности
рис. 1, д. На первой стадии осуществляется тер
теломеразы. Этот флуоресцентный метод с ис
мическая денатурация двухцепочечной ДНК
пользованием молекулярного маяка позволил
при 95 °С (остальные этапы SDA проводятся при
измерить активность теломеразы, содержащей
37 °С). Это позволяет праймерам сформировать
ся в четырех клетках HeLa.
комплексы с каждой из дочерних цепей. Следу
В последние годы для аналитических целей
ет отметить, что фрагменты 5′ концов использу
активно применяются аптамеры. В связи с тем,
емых праймеров, не взаимодействующие с
что аптамеры по своей химической природе яв
ДНК аналитом, содержат сайт расщепления
ляются ДНК/РНК олигонуклеотидами, такие
никазы. Фрагмент Кленова (без экзонуклеазной
методы анализа пытаются комбинировать с ме
активности) катализирует удлинение 3′ концов
тодами амплификации нуклеиновых кислот.
последовательностей ДНК, приводя к образова
Описаны методы, основанные на взаимодей
нию дуплексов с активным сайтом расщепления
ствии аптамеров с тромбином и кокаином,
никазы. В результате ферментативного гидроли
чувствительность данных методов была сущест
за никазой образуются новые 3′ концевые по
венно повышена за счет применения SDA [50,
следовательности, что инициирует реакцию по
51]. Поскольку в настоящее время описано
лимеризации с одновременным замещением до
большое число аптамеров к различным молеку
черней анализируемой цепи. Данный процесс
лам, такой подход представляется достаточно
повторяется многократно. Таким образом, в хо
перспективным.
де SDA происходит экспоненциальная аккуму
Рекомбиназная полимеразная амплификация
ляция анализируемых последовательностей.
(RPA) - еще один изотермический метод ампли
При использовании в рамках данного метода
фикации нуклеиновых кислот [27, 52]. Начиная
одного праймера вместо двух происходит ли
с 2006 г., когда Найл Армес впервые описал RPA
нейная амплификация. SDA позволяет дости
[53], интерес к нему растет год от года. В этом
гать эффективности, равной 107 копий аналита
методе на первом этапе амплификационного
[27].
цикла происходит образование комплекса ре
Как и другие методы амплификации, SDA
комбиназы с прямым и/или обратным прайме
применяется для детекции геномной ДНК. Ана
ром (рис. 1, е). В присутствии комплементарной
лиз таких сложных образцов, содержащих бак
праймеру последовательности фермент распле
терии и вирусы, как кровь человека, успешно
тает дуплекс, что позволяет праймеру специфи
проводится с помощью SDA, что указывает на
чески вступать во взаимодействие с аналитом.
БИОХИМИЯ том 85 вып. 2 2020
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
181
Осуществлению этой реакции содействуют
расщепления с помощью эндонуклеазы в RPA
ДНК связывающие белки. В присутствии ДНК
служит дополнительным этапом, позволяющим
полимеразы со способностью к замещению це
уменьшить фоновый сигнал. Отмечается, что
пи происходит разрушение рекомбиназного
RPA и ПЦР сопоставимы по эффективности.
комплекса, а полимераза связывается с двухце
Метод позиционируется как наиболее быстрый
почечной ДНК и элонгирует праймер с 3′ кон
среди прочих методов амплификации [54].
ца. Вновь синтезированный дуплекс служит
Описано использование RPA в сочетании с
матрицей для следующего цикла. При использо
различными методами детекции для определе
вании двух праймеров (прямого и обратного) по
ния различных патогенов. Так, в работе
окончании реакции образуются два полукон
Mayboroda et al. [55] данный метод был исполь
сервативных дуплекса. Таким образом, RPA ха
зован для определения ДНК Y. pestis. При этом
рактеризуется экспоненциальным типом амп
прямой праймер был иммобилизован в лунках
лификации. Следует отметить, что при исполь
планшета для иммуноферментного анализа, а
зовании одного праймера наблюдается линей
обратный праймер модифицирован биотином,
ный тип амплификации.
который впоследствии использовался для при
RPA может проводиться при 22-45 °С, но
соединения конъюгата пероксидазы со стрепта
оптимальное значение температуры лежит в ин
видином. С использованием колориметричес
тервале 37-42 °С [52]. Для амплификации ис
кого субстрата пероксидазы (3,3′,5,5′ тетраме
пользуются как одноцепочечные, так и двухце
тилбензидина) достигнут предел обнаружения
почечные ДНК, а также метилированные ДНК.
0,3 фМ. Линейный диапазон при этом составлял
Как было продемонстрировано, RPA может
от 10 фМ до 10 нМ.
проводиться в присутствии ингибиторов ПЦР,
Следует отметить, что метод RPA часто соче
таких как гепарин, этанол и гемоглобин [53].
тается с электрохимическим методом детекции.
Это, в отличие от ПЦР, позволяет проводить
С помощью такого подхода Ng et al. [56] смогли
амплификацию непосредственно в биологичес
разработать амперометрический биосенсор,
ких образцах (таких как молоко, моча, кал,
позволяющий определять
1 КОЕ ДНК
плевральная жидкость) после предварительного
Mycobacterium tuberculosis. В другом электрохи
термического лизиса [54]. В то же время RPA
мическом методе в качестве медиатора исполь
ингибируется такими детергентами, как SDS и
зовали комплекс рутения [Ru(NH3)6]3+, который
СТАВ.
был интеркалирован в двухцепочечную ДНК
RPA проводится как в гомогенной, так и в
[57]. Предел обнаружения аналита предложен
гетерогенной средах. В случае гетерогенной
ным методом составил 11 КОЕ/мл.
амплификации один или оба праймера иммоби
Альтернативными электрохимическому ме
лизуются на твердой поверхности. Несмотря на
тоду детекции являются хемилюминесценция,
высокую скорость гомогенной амплификации,
флуоресценция и поверхностное комбинацион
гетерогенный подход активно развивается, т.к. в
ное рассеяние. Недавно с применением RPA
большинстве случаев он позволяет предотвра
был сконструирован флуоресцентный сенсор в
щать матричный эффект и развивать более
виде браслета, который способен определять
чувствительные методы анализа. Также отмеча
специфическую ДНК вируса Зика в условиях
ется, что применение RPA в гетерогенном фор
реального времени [58].
мате позволяет уменьшить степень неспецифи
Достаточно распространенным амплифика
ческой амплификации [55].
ционным методом, направленным на повыше
Хотя RPA - метод быстрый и чувствитель
ние концентрации детектируемой последова
ный, высокое значение фонового сигнала часто
тельности, является метод катящегося кольца
является проблемой при его использовании.
(RCA), разработанный в 1995 г. [59]. Как видно
Для устранения этого негативного эффекта ис
на рис. 1, ж, в основе этого метода лежит ис
пользуется праймер, который содержит сайт
пользование кольцевой ДНК. Кольцевая ДНК
расщепления для специфической эндонуклеазы
(С проба) формируется при взаимодействии
E. coli IV (Nfo), которая распознает и расщепля
анализируемой последовательности с одноце
ет этот участок в дуплексах [52]. Этот праймер
почечным ДНК зондом, содержащим на своих
может быть использован на стадии удлинения
концах последовательности, комплементарные
ДНК полимеразой только после того, как он
аналиту. При формировании такого комплекса
был расщеплен эндонуклеазой с появлением
5′ и 3′ концы соединяются в кольцо. Замыка
гидроксильной группы на 3′ конце. Праймер
ние кольца происходит в результате лигирова
может быть конъюгирован с флуорофором и ту
ния сближенных концов С пробы.
шителем, так что реакция его расщепления соп
В дальнейшем кольцевая ДНК гибридизует
ровождается усилением флуоресценции. Этап
ся с праймером, который в присутствии ДНК
БИОХИМИЯ том 85 вып. 2 2020
182
БОДУЛЕВ, САХАРОВ
полимеразы элонгируется, приводя к образова
ли концентрацию АТФ биолюминесцентным
нию последовательности, состоящей из много
методом с помощью люциферазы. Предел обна
численных участков, повторяющих ДНК ана
ружения модельного РНК аналита, определен
лит.
ный этим методом, составил 2 пМ, его линей
В RCA наиболее часто используются ДНК
ный диапазон - от 2 пМ до 1 нМ.
полимеразы phi 29 и Bst. Обычно данная реак
RCA использовался для повышения чувст
ция проводится при 30-37 °С [60]. Линейная
вительности метода определения активности
амплификация при постоянной температуре за
метилтрансферазы [68]. Разработанный метод
нимает от нескольких часов до нескольких дней,
позволил детектировать фермент с концентра
что приводит к синтезу многочисленных копий
цией 8,1 × 10-5 ед/мл. Линейный диапазон дан
аналита. Эффективность RCA оценивается в 103
ного метода лежит в диапазоне от 4 × 10-4 до
копий аналита.
1 × 10-2 ед/мл. Также RCA успешно был приме
Недавно описана модификация RCA, в ко
нен для детектирования метилирования ДНК
торой в реакционную смесь также добавляется
[69].
второй праймер, гибридизующийся c последо
Следует отметить, что метод RCA может
вательностью, элонгированной с использовани
быть направлен на повышение аналитического
ем первого праймера. Введение второго прайме
сигнала как за счет повышения концентрации
ра приводит к тому, что амплификация приоб
аналита, так и увеличения количества детекти
ретает экспоненциальный характер [60].
руемых меток.
Метод RCA широко используется при разра
Во всех методах амплификации, приводя
ботке методов определения бактериальных и
щих к повышению аналитического сигнала за
вирусных ДНК/РНК и микроРНК. Более того,
счет повышения концентрации аналита, ис
RCA является высокоспецифическим методом
пользуются ДНК полимеразы, уязвимые к
за счет применения ДНК лигазы, катализирую
действию ингибиторов, присутствующих в ана
щей лигирование лишь в случае точной коорди
лизируемых образцах. Часто это может приво
нации 3′ и 5′ концевых последовательностей
дить к получению ложноотрицательных резуль
[61, 62], в связи с чем данный метод позволяет
татов. С другой стороны, фоновая амплифика
выявлять однонуклеотидные полиморфизмы
ция, вызванная димеризацией используемых
[63].
праймеров/зондов, может быть причиной лож
В работе Schopf et al. [64] описано примене
ноположительных результатов. Минимизиро
ние RCA при определении геномной ДНК
вать указанные недостатки позволяет использо
Mycobacterium tuberculosis. На первом этапе ДНК
вание методов детекции, высокоспецифичных к
обрабатывали рестриктазами и денатурировали
анализируемой последовательности.
при повышенной температуре. Далее произво
Все вышеописанные методы амплификации
дили иммобилизацию ее фрагментов за счет их
были основаны на применении специфических
гибридизации с захватывающими олигонуклео
последовательностей. Вместе с тем были разра
тидами, ковалентно связанными с частицами
ботаны и изотермические методы усиления с
сефарозного геля. После проведения амплифи
использованием случайных праймеров. Такие
кации с помощью RCA в систему вносили комп
методы полногеномной амплификации (WGA)
лементарные анализируемым фрагментам ДНК
применяются для увеличения количества ДНК,
с флуоресцентными метками. Предел обнаруже
необходимого для ее секвенирования.
ния описанного метода составил 4,3 фМ анали
Первый вариант WGA, получивший название
зируемой ДНК и 104 КОЕ/мл M. tuberculosis.
амплификации со множественным замещением це1
Продукты RCA определялись также за счет ин
пи (MDA), был описан в 2001 г. [70]. Этот метод
теркалирования в дуплекс ДНК красителя SYBR
основан на использовании случайных гексамер
Green. В случае определения микроРНК miR
ных праймеров, взаимодействующих с кольце
let 7a предел обнаружения составил 10 фМ, ли
выми геномами, что проводит к формированию
нейный диапазон - от 25 фМ до 1 пМ аналита
многочисленных репликационных вилок. В
[65]. Кроме того, применение RCA позволило
MDA используется ДНК полимераза Phi 29, ко
визуализировать микроРНК семейства let 7 не
торая характеризуется повышенной процессив
посредственно в живых раковых клетках легких
ностью и низкой частотой ошибок. Протекание
A549 [66].
каскада реакций MDA приводит к экспоненци
Регистрация процесса полимеризации RCA
альному накоплению двухцепочечных ДНК и
возможна и по накоплению в реакционной сре
104 кратному увеличению концентрации плаз
де пирофосфата. В работе Mashimo et al. [67] с
мидной ДНК в течение нескольких часов.
помощью аденилил трансферазы пирофосфат
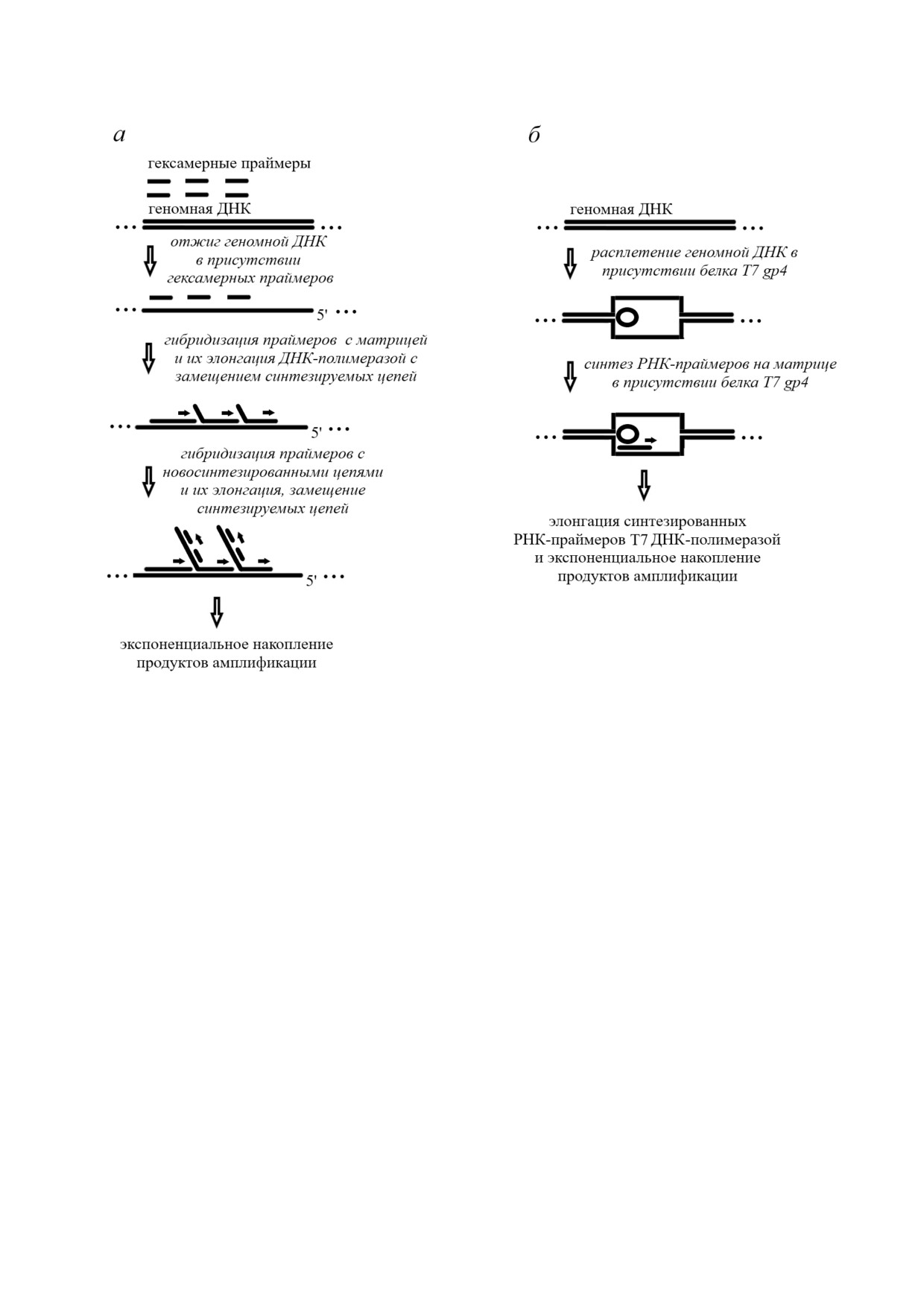
В 2002 г. этот метод был адаптирован для
трансформировали в АТФ, после чего определя
амплификации линейных геномов (рис. 2, а).
БИОХИМИЯ том 85 вып. 2 2020
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
183
Рис. 2. Полногеномный метод амплификации: а - амплификация с множественным замещением цепи (MDA); б - пол
ногеномный метод амплификации с применением праймазы
С применением данного метода наличие 1-10
применении pWGA удается избежать тепловой
копий геномной ДНК человека позволяет полу
денатурации геномной ДНК, которая проводит
чить ~20-30 мкг ДНК со средней длиной
ся в MDA.
~10 кб, которые могут быть использованы для
секвенирования и генотипирования.
Альтернативой WGA является полногеном1
МЕТОДЫ АМПЛИФИКАЦИИ,
ный метод амплификации с применением прайма1
ОСУЩЕСТВЛЯЮЩИЕ ПОВЫШЕНИЕ
зы (pWGA), который имитирует репликацию
АНАЛИТИЧЕСКОГО СИГНАЛА
ДНК бактериофага Т7 in vivo [71]. В присутствии
БЕЗ УВЕЛИЧЕНИЯ КОНЦЕНТРАЦИИ
бифункционального белка T7 gp4, обладающего
АНАЛИТА
активностями и хеликазы, и праймазы, геном
ная ДНК расплетается, после чего синтезирует
Для изотермических амплификационных
ся РНК праймер, комплементарный одноцепо
методов, позволяющих повысить аналитичес
чечной ДНК (рис. 2, б). Синтез ДНК катализи
кий сигнал без изменения концентрации анали
руется Т7 ДНК полимеразой, обладающей вы
та, характерна линейная кинетика амплифика
сокой процессивностью. Как правило, при ис
ции. Некоторые из них проводятся с примене
пользовании pWGA в течение 1 ч при 37 °С обра
нием ферментов, другие - без их участия, что,
зуется 1-10 нг геномной ДНК человека. Коль
естественно, делает последние методы более де
цевую ДНК также можно использовать в качест
шевыми, а также лишает их недостатков, типич
ве матрицы в pWGA. При введении в анализ все
ных для ферментативных методов. Далее будут
го лишь 100 копий коэффициент амплифика
рассмотрены и те, и другие методы, т.к. все они
ции достигает 108. Следует отметить, что при
имеют свои преимущества и недостатки и ис
БИОХИМИЯ том 85 вып. 2 2020
184
БОДУЛЕВ, САХАРОВ
пользуются при разработке аналитических ме
от экзонуклеазы III, катализируют отщепление
тодов.
мононуклеотидов не с 3′ конца, а с 5′ конца ис
Одним из изотермических амплификацион
пользуемого зонда.
ных методов без изменения концентрации ана
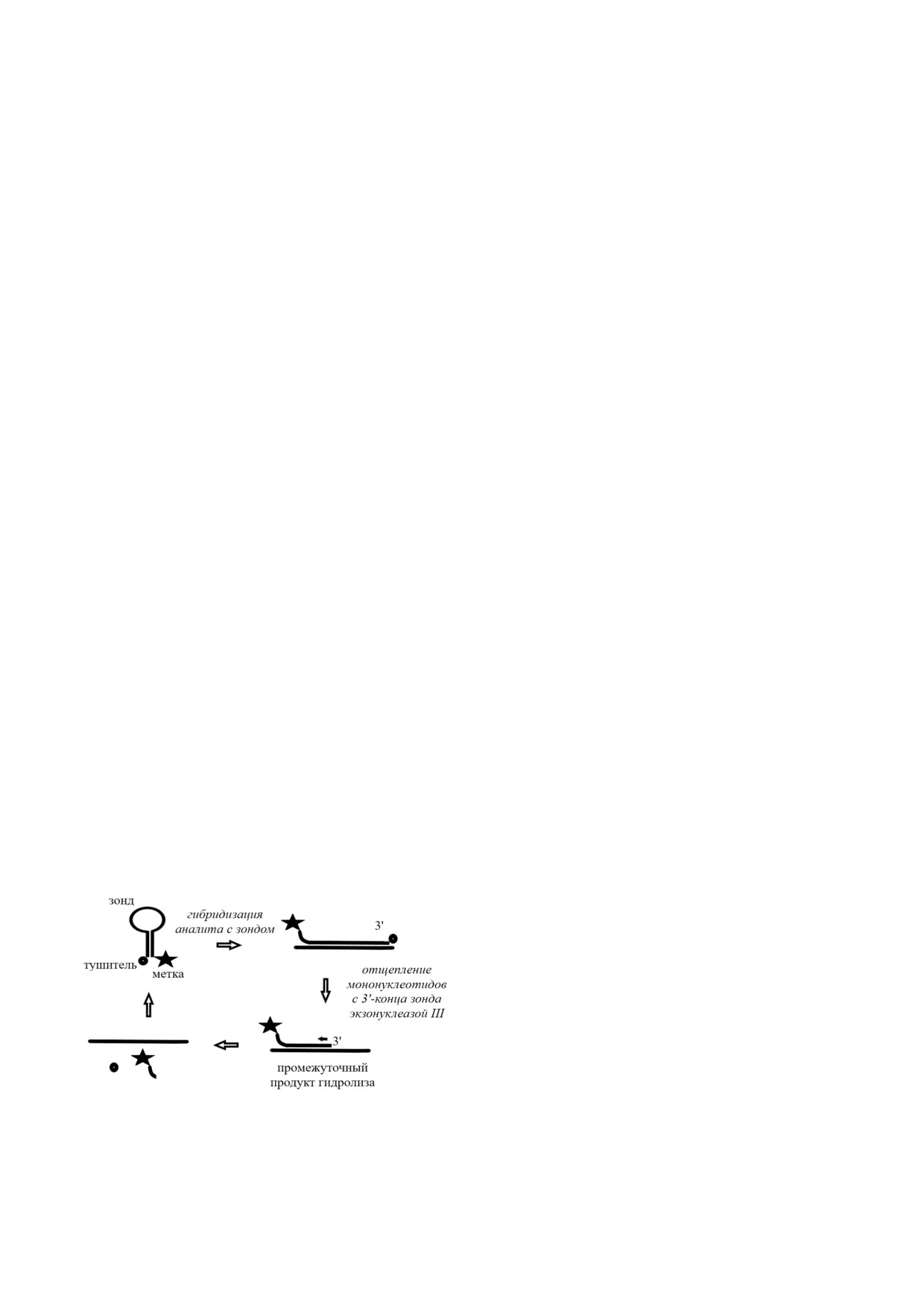
В пионерской работе по EASA данный метод
лита является метод экзонуклеаза III1зависимой
был применен для количественного определе
циклизации аналита (EASA) [7, 72]. Экзонуклеа
ния модельного ДНК олигонуклеотида [74]. В
за III - фермент семейства нуклеаз, катализиру
этой работе был использован молекулярный ма
ющий поэтапное удаление мононуклеотидов с
як, который содержал флуоресцентный краси
3′ гидроксилированных концов двухцепочеч
тель на 5′ конце и тушитель на одном из нукле
ной ДНК путем гидролиза фосфодиэфирных
отидов, расположенных во внутренней области
связей [73], т.е. экзонуклеаза III обладает неспе
последовательности маяка. При этом за счет са
цифической 3′-5′ экзонуклеазной активностью.
могибридизации формировалась шпилька с
Подходящими субстратами для проявления ак
выступающим концом, устойчивым к гидролизу
тивности такого фермента являются ДНК с ту
экзонуклеазой III. Более того, закрытая струк
пыми или утопленными 3′ концами. Следует
тура шпильки удерживала флуорофор в непос
отметить, что субстраты с выступающими 3′
редственной близости от тушителя, что приво
концами при их удлинении на четыре или более
дило к регистрации лишь слабого сигнала флуо
нуклеотидов ферментативному расщеплению
ресценции. В присутствии ДНК аналита шпи
экзонуклеазой III не подвергаются.
лечная структура маяка раскрывалась с образо
В 2010 г. Plaxco et al. [74] впервые описали
ванием двухцепочечной структуры с тупым 3′
принцип EASA (рис. 3). Суть данного амплифи
концом, что позволяло экзонуклеазе III катали
кационного метода заключается в том, что ис
зировать удаление мононуклеотидов с этого
следуемая нуклеиновая кислота гибридизуется с
конца, что, в свою очередь, приводило к высво
ДНК зондом с образованием двухцепочечной
бождению флуорофора и повышению интен
структуры с тупым 3′ концом, что позволяет эк
сивности флуоресценции. Одновременно с этим
зонуклеазе III катализировать поэтапное удале
освобожденный аналит взаимодействовал с дру
ние мононуклеотидов с 3′ конца используемого
гой молекулой молекулярного маяка, иници
зонда. В результате ферментативного гидролиза
ируя следующий цикл EASA. Таким образом,
молекула аналита высвобождается и вступает во
применение EASA позволило разработать прос
взаимодействие с другой молекулой зонда, что
той флуоресцентный метод обнаружения ДНК
инициирует следующий цикл амплификации.
аналита.
Таким образом, одна молекула аналита может
Позднее было опубликовано большое число
привести к образованию большого числа моле
работ [76-80], в которых EASA применен для
кул, образующихся при гидролизе зонда. Если
повышения чувствительности анализа нуклеи
используемый зонд содержит в своей структуре
новых кислот, при этом на стадии детекции ис
некую метку, то регистрируемый аналитический
пользовалась не только флуоресценция, но и
сигнал будет амплифицирован. Амплификация
электрохимия, колориметрия и хемилюминес
обычно проводится при 25 или 37 °С.
ценция.
Позднее подобные амплификационные ме
Используя в качестве индикаторной реакции
тоды были развиты с применением T7 экзонук
взаимодействие Hg2+ с тиминами, разработан
леазы и λ экзонуклеазы [75], которые, в отличие
метод с применением EASA для количественно
го определения ионов ртути, предел обнаруже
ния которого составил 1 пМ Hg2+, а линейный
диапазон - от 10 пМ до 100 нМ [79]. Объедине
ние в одном методе EASA и аптамеров позволи
ло разработать методы для определения АТФ,
лизоцима и тромбина.
Следует особо отметить, что в некоторых ра
ботах по определению нуклеиновых кислот пре
дел обнаружения достигал фемто , а в каких то
случаях и аттомолярных значений. По видимо
му, к указанным для EASA столь низким значе
ниям предела обнаружения следует относиться с
некоторой долей осторожности, т.к. уже в пио
нерской работе [74] авторами было отмечено,
Рис. 3. Схема количественного анализа ДНК олигонукле
отида с помощью амплификационного метода экзонукле
что экзонуклеаза III проявляет значительную
аза III зависимой циклизации аналита (EASA)
каталитическую активность по отношению к
БИОХИМИЯ том 85 вып. 2 2020
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
185
а
б
Рис. 4. Схемы методов изотермической амплификации нуклеиновых кислот с формированием Y структур: а - метод, ос
нованный на формировании и последующем ферментативном расщеплении Y зонда; б - безферментный метод с приме
нением Y зонда
одноцепочечным последовательностям ДНК,
ло показано, что экзонуклеаза III способна гид
что должно приводить к ферментативному гид
ролизовать ДНК, имеющую структуру G квад
ролизу анализируемой последовательности. На
руплекса (неопубликованные данные). Другие
личие этой побочной активности отмечалось и
экзонуклеазы также катализируют указанные
другими авторами [80, 81]. Более того, нами бы
побочные реакции. Отмеченный факт сущест
3 БИОХИМИЯ том 85 вып. 2 2020
186
БОДУЛЕВ, САХАРОВ
венно ограничивает развитие метода амплифи
ферментативного расщепления Y зонда исполь
кации с использованием таких экзонуклеаз.
зовалась эндонуклеаза HaeIII. Для регистрации
EASA применяется также для повышения
процесса расщепления Y зонда одна из его по
чувствительности аналитических методов опре
следовательностей, иммобилизованная на поверх
деления активности ряда ферментов, таких как
ности электрода, была конъюгирована с метиле
T4 полинуклеотидкиназа, метилтрансфераза,
новым синим. Данное вещество было простран
теломераза [82-84], при этом используются как
ственно удалено от поверхности электрода и по
экзонуклеаза III, так и λ экзонуклеаза.
этой причине электрохимически не окислялось.
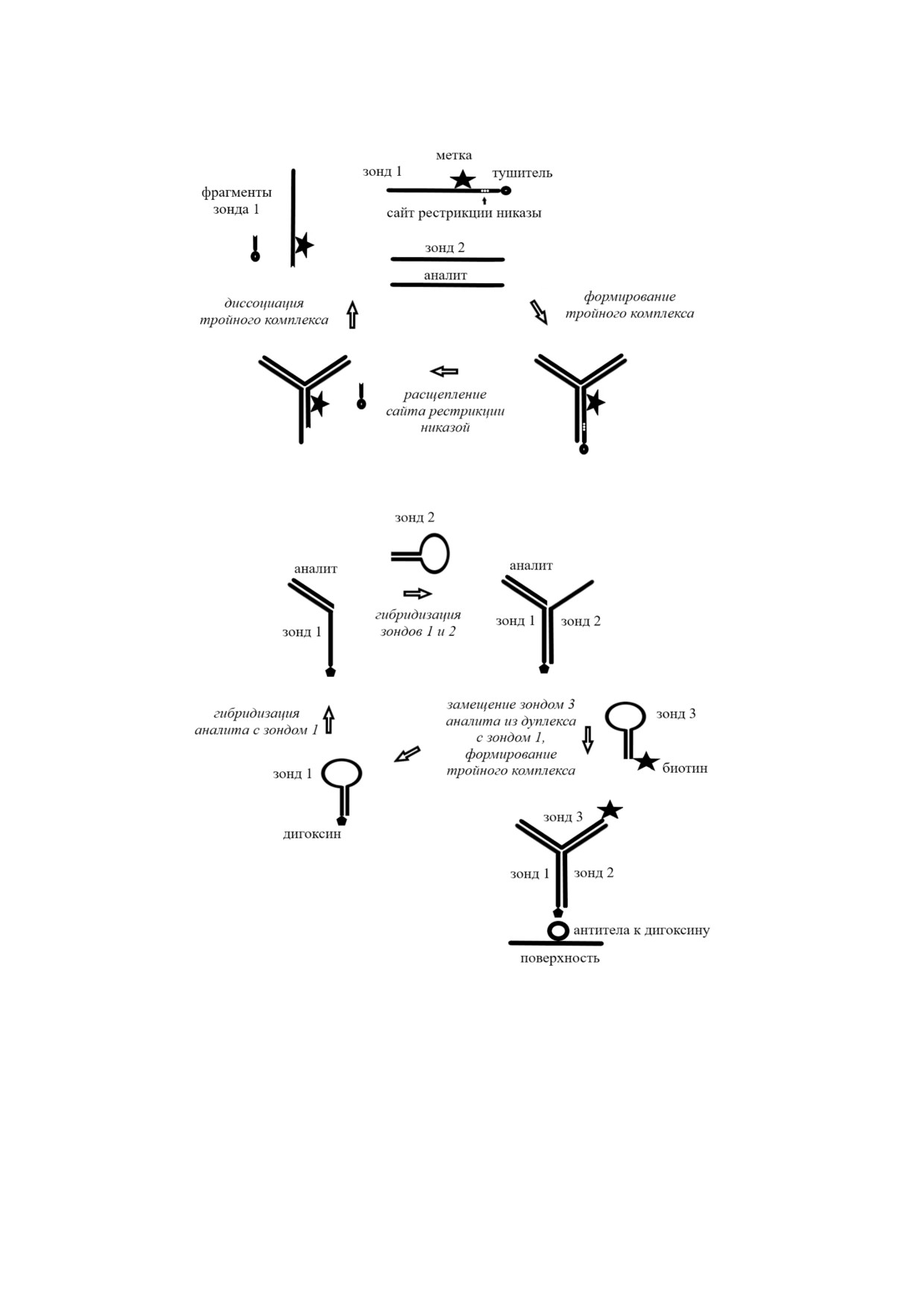
Одним из новых изотермических амплифи
В результате гидролиза сайта рестрикции Y зон
кационных методов является метод с формиро1
да данное соединение обретало способность
ванием Y1структур [7]. При формировании таких
мигрировать к поверхности электрода и там
структур используются два частично компле
окисляться, что приводило к повышению реги
ментарных друг другу зонда (обычно 4-6 п.н.),
стрируемого значения тока, которое было про
которые из за ограниченной комплементарнос
порционально концентрации аналита. Предел
ти не связываются друг с другом. В то же время
обнаружения данного метода составил 14 пМ.
в присутствии анализируемой последователь
Описан также и латеральный проточный ме
ности, которая частично комплементарна обо
тод с колориметрической детекцией для опреде
им зондам, формируются устойчивые Y комп
ления микроРНК miR 16 с применением Y зон
лексы (рис. 4, а). Более того, фрагмент таких
да [89]. В данном случае реакцию амплифика
комплексов, образованных при взаимодействии
ции проводили в гомогенной среде. Y Зонд об
зондов между собою, содержит сайт расщепле
разовывался как результат взаимодействия мо
ния рестриктазами. Введение в реакционную
лекулярного маяка, вспомогательного ДНК
среду рестриктазы, соответствующей последо
олигонуклеотида и miR 16. В присутствии эндо
вательности сайта расщепления, приводит к
нуклеазы Nt.BbvCI молекулярный маяк в соста
ферментативному расщеплению дуплекса с по
ве полученного Y зонда расщеплялся на два
следующей диссоциацией Y структуры и, соот
фрагмента. Это приводило к тому, что вспомога
ветственно, высвобождением анализируемой
тельный олигонуклеотид и miR 16 реагировали
последовательности, которая используется для
со следующей молекулой молекулярного маяка,
формирования нового Y зонда. Образующийся
и процесс повторялся вновь. Концентрацию об
Y зонд принимает участие в следующем цикле
разовавшихся фрагментов маяка определяли ла
амплификации, что в конечном счете приводит
теральным методом. Предел обнаружения miR 16
к образованию многочисленных гидролизован
составил 0,1 пМ, линейный диапазон - от 0,1 пМ
ных фрагментов используемых зондов. Наличие
до 10 нМ.
большого числа известных рестриктаз (~3500)
Также был описан модифицированный ме
позволяет смоделировать и получить разнооб
тод амплификации с применением Y зондов без
разные Y зонды с различными сайтами расщеп
использования каких либо ферментов [90]. На
ления. Sintim et al. показали, что архитектура
первом этапе молекула аналита за счет компле
зондов оказывает существенное влияние на ско
ментарного взаимодействия образует комплекс
рость ферментативного гидролиза Y зондов
с двумя шпильками (рис. 4, б), одна из которых
рестриктазами, что необходимо учитывать при
содержит присоединенный дигоксин. Надо под
моделировании структуры используемых зондов
черкнуть, что сами шпильки не способны гиб
[85]. Следует отметить, что некоторые авторы
ридизоваться между собой в отсутствие аналита.
вместо рестриктаз для гидролиза Y зонда ус
Затем, при добавлении меченной биотином
пешно использовали никазы [86]. Температура
третьей шпильки, которая вытесняет из комп
проведения этого типа амплификации в различ
лекса молекулу аналита, формируется Y зонд.
ных работах варьируется в диапазоне 25-37 °С.
Высвободившийся аналит вновь формирует
Как следует из схемы данного анализа
комплекс со шпильками 1 и 2, инициируя нача
(рис. 4, а), метод амплификации с применением
ло следующего цикла амплификации, что в ре
Y зондов может быть использован для опреде
зультате позволяет без изменения концентра
ления как ДНК, так и РНК, а также аналитов
ции аналита получить Y зонд, концентрация
другой химической природы (например, анти
которого значительно превышает концентра
биотиков), при этом в качестве распознающих
цию аналита. После проведения реакции амп
соединений используются аптамеры [87].
лификации концентрацию образовавшегося Y
С применением Y зонда ранее был разрабо
зонда, содержащего в своей структуре дигоксин
тан электрохимический метод определения 28
и биотин, определяли электрохимически. Для
членного ДНК олигонуклеотида, являющегося
этого за счет взаимодействия дигоксина с его
модельным аналитом [88]. В данном методе для
антителами, сорбированными на поверхности
БИОХИМИЯ том 85 вып. 2 2020
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
187
электрода, проводили иммобилизацию Y зонда,
а последовательное добавление конъюгата
стрептавидина с пероксидазой и 3,3′,5,5′ тетра
метилбензидина позволило регистрировать ве
личину тока, пропорциональную концентрации
Y зонда. При использовании ДНК олигонукле
отида в качестве модельного аналита предел об
наружения составил 10,9 аМ, линейный диапа
зон - от 100 аМ до 1 мкМ.
Никазы применяются не только в методе
изотермической амплификации нуклеиновых
Рис. 5. Схема метода с применением никазы, основанного
кислот с формированием Y зонда, но также и в
на формировании и последующем ферментативном рас
методе, основанном на применении специфи
щеплении ДНК дуплекса
ческих дуплексов [91]. Схема такого метода
представлена на рис. 5. Анализируемая последо
вательность формирует специфический дуплекс
щей молекулой зонда. Для данного метода пре
с ДНК зондом, создавая одновременно сайт
дел обнаружения фрагмента гена p53 составил
рестрикции, распознаваемый одной из никаз.
1 пМ, рабочий диапазон - 1-100 пМ.
На стадии синтеза в последовательность зонда
Данный метод амплификации был применен
вводится некая метка, с помощью которой впо
также в комбинации с электрохимическим мето
следствии проводится оценка выхода фермента
дом детекции нуклеиновых кислот. Chen et al.
тивного гидролиза зонда. В дальнейшем специ
[93] описали приготовление ДНК зонда шпиль
фическая никаза расщепляет последователь
ки с ковалентно связанным ферроценом, кото
ность зонда, что приводит к диссоциации дуп
рый был иммобилизован на поверхность золото
лекса. Высвобожденная анализируемая молеку
го электрода. После ферментативного расщепле
ла взаимодействует со следующей молекулой
ния сайта рестрикции, образованного при его
зонда, и этот процесс повторяется многократно.
взаимодействии с аналитом, фрагменты зонда,
В результате одна молекула аналита позволяет
модифицированные ферроценом, отдалялись от
произвести большое число фрагментов исполь
поверхности электрода. Предел обнаружения мо
зуемого зонда, что, в свою очередь, приводит к
дельного ДНК аналита, измеренный с помощью
повышению регистрируемого сигнала. Темпера
электрохимического биосенсора, составил
тура проведения амплификации должна быть
68 аМ, а линейный диапазон - от 0,1 до 100 фМ.
достаточно высокой для быстрой диссоциации
Недавно была опубликована работа, в кото
расщепленного зонда, но при этом она ограни
рой метод амплификации с использованием ни
чена термостабильностью используемой ника
казы Nt. Alwl применили в количественном ана
зы. При первом описании данного метода, в ко
лизе катионов ртути [94]. Используя образова
тором использовалась никаза Nt.AlwI, темпера
ние комплекса тимин-Hg2+-тимин как индика
турный оптимум составлял 58 °С [91].
торную реакцию, Vijayan et al. разработали метод
В зависимости от типа вводимой в зонд мет
анализа с пределом обнаружения 0,14 нМ. Авто
ки возможно использование различных методов
ры показали, что разработанный метод может
регистрации сигнала. Так, в работе Lin et al. [92]
применяться для определения ртути в питьевой
ДНК зонд представлял из себя шпильку, при
воде. На основе амплификации с помощью ни
этом одна из цепей стебля шпильки была апта
казы также были успешно разработаны методы с
мером гемина. После реакции зонда с анализи
использованием аптамеров для определения ли
руемой последовательностью (19 членным оли
зоцима, карциноэмбрионального и простатспе
гонуклеотидом гена p53) формировался дуплекс
цифического антигенов, а также муцина 16 в
с сайтом рестрикции никазы Nt.BstNBI. После
сыворотке крови человека [95].
ферментативного гидролиза высвобождался ап
В работах с применением данного типа амп
тамер гемина, и после добавления в реакцион
лификации отмечается высокая специфичность
ную среду гемина происходило формирование
разрабатываемых методов анализа. Тем более
каталитически активного пероксидаза подоб
странно, что число публикаций с упоминанием
ного ДНКзима, активность которого определя
о применении данного метода амплификации
лась при окислении 2,2′ азино бис(3 этилбенз
значительно меньше, чем в случае других изо
тиазолин 6 сульфоната) в присутствии перок
термических амплификационных методов.
сида водорода. Одновременно происходило
В 2009 г. был описан изотермический ампли1
высвобождение аналита из дуплекса, что позво
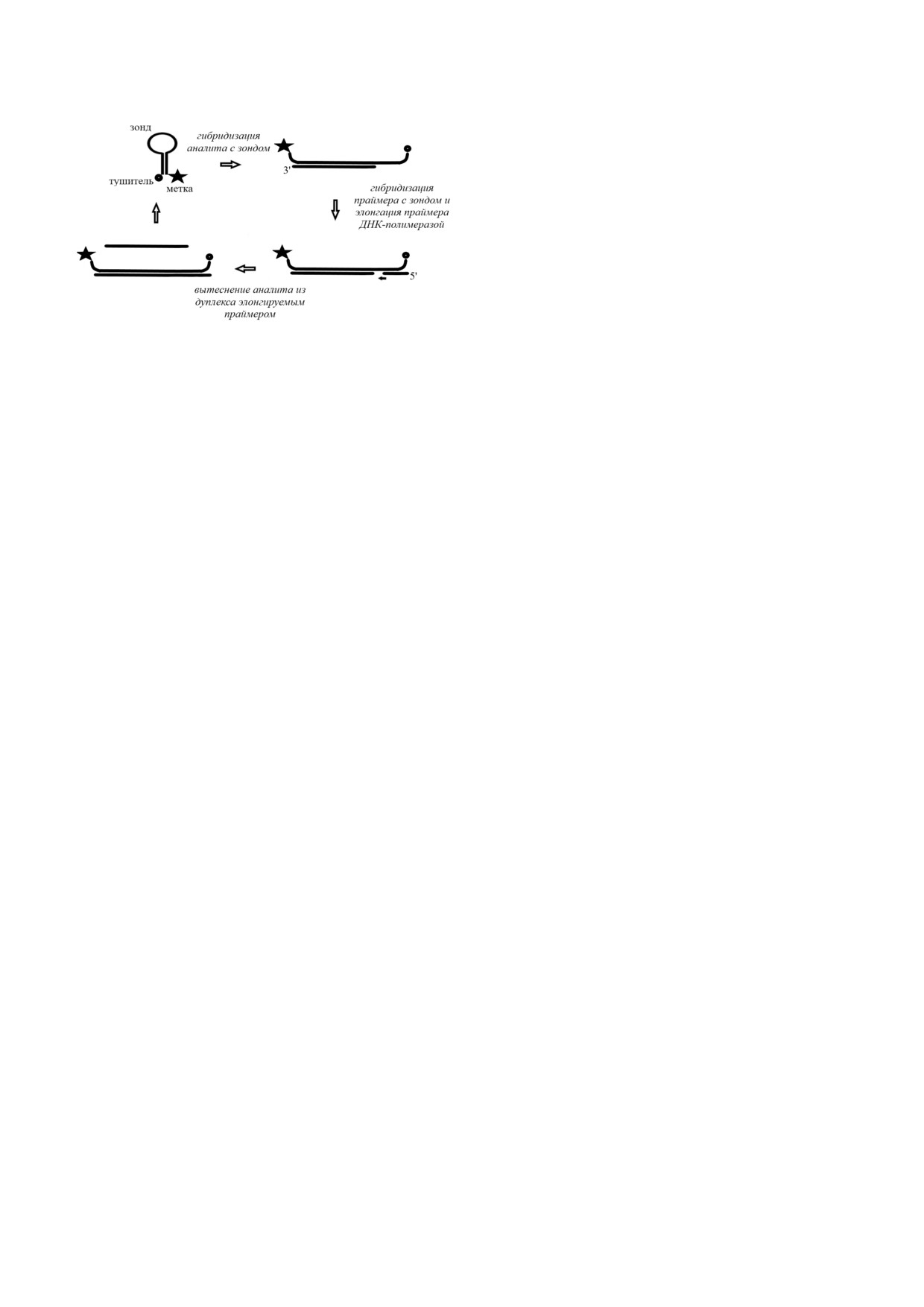
фикационный метод с полимеризацией и замеще1
ляло ему вступать во взаимодействие со следую
нием (ICSDP) [96]. Принцип метода представ
БИОХИМИЯ том 85 вып. 2 2020
3*
188
БОДУЛЕВ, САХАРОВ
ровалась фрагментом Кленова. Для определе
ния модельного 26 членного ДНК олигонукле
отида был развит флуоресцентный гомогенный
метод с ICSDP, предел обнаружения которого
составил 6,4 фM.
Аналогичный принцип был использован при
разработке метода определения микроРНК
miR 210 [97]. Предел обнаружения описанного
метода составил 50 пМ. Следует отметить, что
линейный диапазон определения микроРНК
для данного метода оказался достаточно узким
(от 330 пМ до 1,66 нМ). С помощью разработан
Рис. 6. Схема метода с использованием амплификации с
ного метода был проведен анализ miR 210 в
полимеризацией и замещением (ICSDP)
трансфицированных клетках К562.
ICSDP был также использован при разработ
лен на рис. 6. Этот метод основан на примене
ке гетерогенных методов определения нуклеи
нии ДНК шпильки (зонда), короткого прайме
новых кислот. Так, Gao et al. [98] на поверхнос
ра и ДНК полимеразы. Структура зонда вклю
ти золотого электрода иммобилизовали шпиль
чает в себя некую метку, позволяющую оцени
ку, модифицированную ферроценом. В присут
вать концентрацию данной молекулы. Посколь
ствии аналита происходило ее раскрытие и выс
ку последовательность, расположенная на 5′
вобождение комплементарного праймеру фраг
конце и формирующая стебель зонда и часть его
мента, модифицированного метиленовым голу
петли, комплементарна анализируемой после
бым. После гибридизации праймера происходи
довательности, это позволяет им специфически
ла его элонгация в присутствии ДНК полимера
взаимодействовать друг с другом с образовани
зы со способностью к замещению цепи. Предел
ем дуплекса и раскрытием структуры шпильки.
обнаружения электрохимического биосенсора,
Праймер, обычно представляющий собой по
разработанного для определения модельного
следовательность длиной в восемь нуклеотидов,
ДНК аналита (31 нуклеотид), составил 28 фМ.
комплементарен 3′ концу стебля зонда. При от
Рабочий диапазон метода находился в диапазо
сутствии аналита праймер с зондом не взаимо
не от 100 фМ до 10 нМ. Аналогичный подход
действует, тогда как в присутствии молекулы
был применен для определения гена mecA в ме
аналита после образования дуплекса праймер
тициллин резистентных штаммах Staphylococcus
связывается с освободившимся 3′ концом зонда.
aureus [99].
Связывание праймера инициирует реакцию
Следует отметить, что поскольку в ICSDP
его элонгации в присутствии ДНК полимеразы
используется ДНК полимераза, данный метод
и дезоксинуклеозид 5′ трифосфатов. В процессе
может быть чувствителен к компонентам, инги
удлинения праймера синтезируемая последова
бирующим ДНК полимеразу, что может приво
тельность вытесняет аналит, переводя его в не
дить к получению ложноотрицательных резуль
связанное состояние. Это, в свою очередь, поз
татов. К тому же фоновая амплификация, выз
воляет ему гибридизоваться с другой молекулой
ванная неспецифической гибридизацией, в
зонда шпильки, что запускает следующий цикл
свою очередь, может приводить к ложнополо
полимеризации/вытеснения. Следовательно,
жительным результатам.
молекула аналита выступает в качестве триггера
В отличие от описанных выше изотермичес
реакции полимеризации. Параллельно с расхо
ких методов амплификации нуклеиновых кис
дованием зонда происходит накопление дуплек
лот, в реакции цепной гибридизации (HCR) не ис
са, состоящего из зонда и синтезированной по
пользуются какие либо ферменты. Данный ме
следовательности, концентрация которого мо
тод был разработан Дирксом и Пирсом в 2004 г.
жет быть измерена различными физико хими
[100]. В основе метода HCR лежит реакция об
ческими методами. Таким образом, изотерми
разования двухцепочечной ДНК в результате
ческий ICSDP, который проводится обычно при
взаимодействия двух шпилек, которая иници
37 °С, позволяет амплифицировать сигнал при
ируется ДНК/РНК аналитом (рис. 7). Следует
постоянной концентрации аналита.
отметить, что структура шпилек 1 и 2 выбирает
Guo et al. [96] в качестве зонда использовали
ся таким образом, чтобы их комплементарные
молекулярный маяк, у которого на концах
взаимодействия между собою были кинетичес
шпильки находились флуоресцеин как флуорес
ки затруднены. В присутствии аналита образо
центная метка и DABCYL как тушитель флуо
вание дуплекса между шпилькой 1 и анализиру
ресценции. Реакция полимеризации катализи
емой последовательностью приводит к высво
БИОХИМИЯ том 85 вып. 2 2020
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
189
Рис. 7. Схема безферментного изотермического метода амплификации с применением реакции цепной гибридизации (HCR)
бождению не вступающей в это взаимодействие
ная двухцепочечная ДНК [102]. Таким образом,
последовательности стебля шпильки 1, что, в
в случае HCR аналит может инициировать фор
свою очередь, позволяет этому одноцепочечно
мирование дуплексов ДНК с наличием разры
му фрагменту вступить в реакцию со шпилькой
вов в обеих цепях, и увеличение интенсивности
2. Последняя реакция приводит к образованию
аналитического сигнала происходит из за того,
дуплекса между используемыми шпильками и
что сформировавшаяся ДНК содержит в своей
одновременному высвобождению непрореаги
структуре большое число введенных меток.
ровавшей последовательности стебля шпильки
При использовании HCR в биоанализе часто
2, которая затем вступает в реакцию с другой
применяются шпильки (одна или две), содержа
молекулой шпильки 1. Введение детектирую
щие биотин. Так, в работе Yang et al. [103] иссле
щих меток в структуру шпилек или интеркали
дуемая ДНК (модельный аналит) инициировала
рование красителей в структуру дуплекса позво
проведение HCR с формированием дуплексной
ляет зарегистрировать образующуюся ДНК. Та
цепочки, содержащей биотин, т.к. одна из ис
ким образом, в результате проведения HCR
ходных шпилек была модифицирована данным
формируется двухцепочечная ДНК с разрывами
соединением. За счет присутствия в дуплексе
в каждой цепи, при этом длина молекулы опре
биотиновых остатков он способен взаимодей
деляется количеством шпилек в реакционном
ствовать со стрептавидином, сорбированным на
растворе. Обычно амплификация проводится
магнитных частицах. Поскольку не все биотины
при 25 или 37 °С.
дуплекса были задействованы в процессе иммо
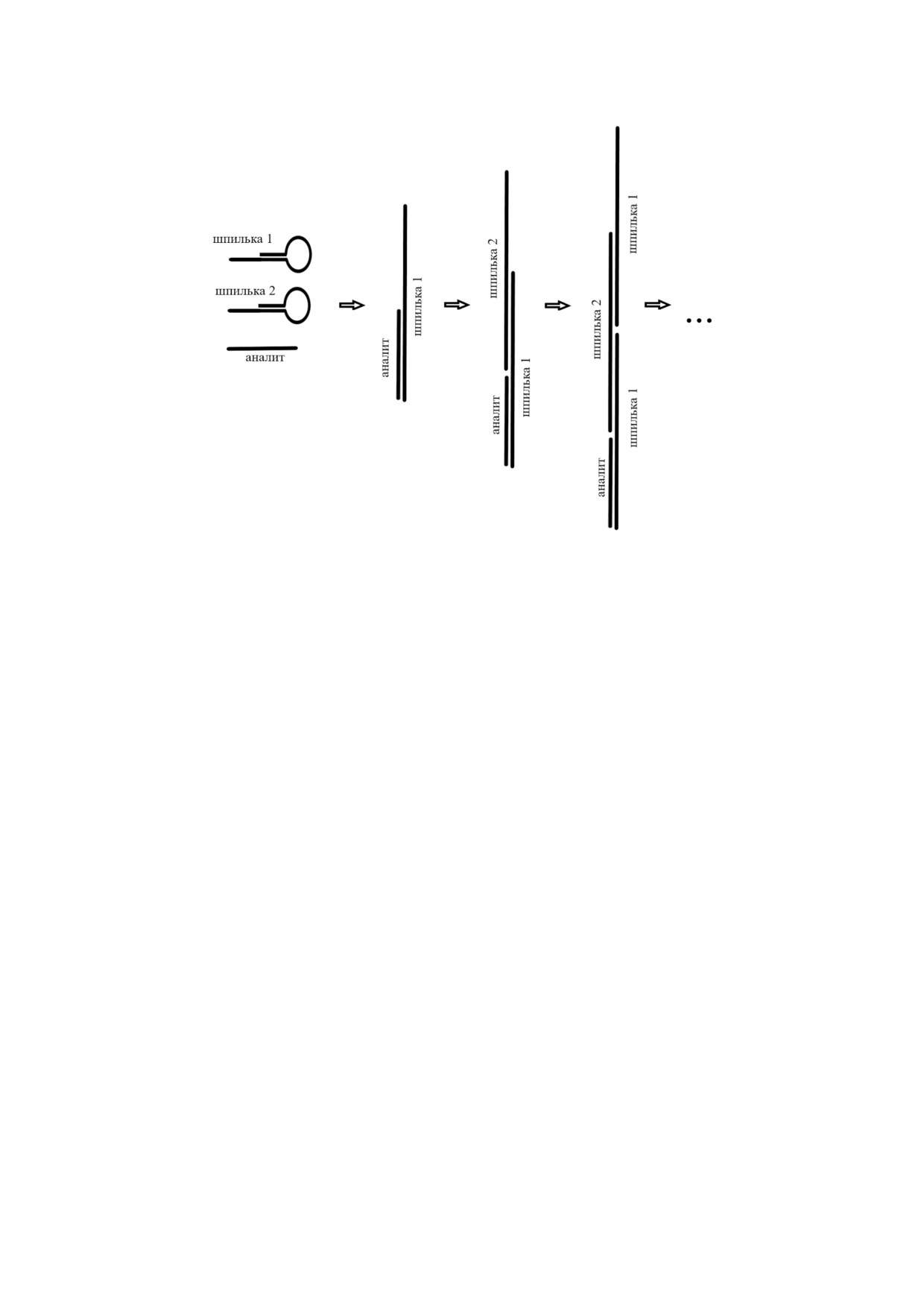
Для усиления эффекта амплификации были
билизации на частицах, другая часть остатков
разработаны методы с использованием шпиль
биотина была использована для взаимодействия
ки с двумя и большим количеством петель, что
с конъюгатом авидина и глюкозооксидазы. В
позволяло получать разветвленную структуру
присутствии глюкозооксидазы глюкоза окисля
двуцепочечной ДНК [101]. С использованием
лась, что приводило к образованию пероксида
четырех шпилек и двух дополнительных одно
водорода. В его присутствии происходило трав
цепочечных ДНК описана модификация метода
ление поверхности наночастиц серебра, что ре
HCR, названная гиперразветленной HCR
гистрировалось методом поверхностного плаз
(hyperbranched HCR). Кинетика амплификации
монного резонанса. Предел обнаружения ана
данной модификации экспоненциальная, а
лита в этом случае составил 6 фМ, линейный
продуктом амплификации является разветвлен
диапазон - от 10 фМ до 100 пМ.
БИОХИМИЯ том 85 вып. 2 2020
190
БОДУЛЕВ, САХАРОВ
Рис. 8. Схема безферментного изотермического метода амплификации с применением метода каталитической сборки
шпилек (CHA)
HCR была применена при конструировании
са. При этом домен шпильки 1, комплементар
электрохимического биосенсора, предназна
ный последовательности хвоста шпильки 2, ста
ченного для определения микроРНК в лизате
новится доступен для такого взаимодействия.
клеток HUVEC, HK 2, HeLa и MCF 7 [104].
Следует отметить, что структура используемых
Достигнутый с помощью вольтамперометричес
шпилек зондов моделируется с учетом того, что
кой детекции предел обнаружения микроРНК
они должны быть комплементарны друг другу,
Hsa miR 17 5p составил 2 аМ, линейный диапа
но их взаимодействие между собой должно быть
зон - от 100 аМ до 100 пМ.
кинетически затруднено.
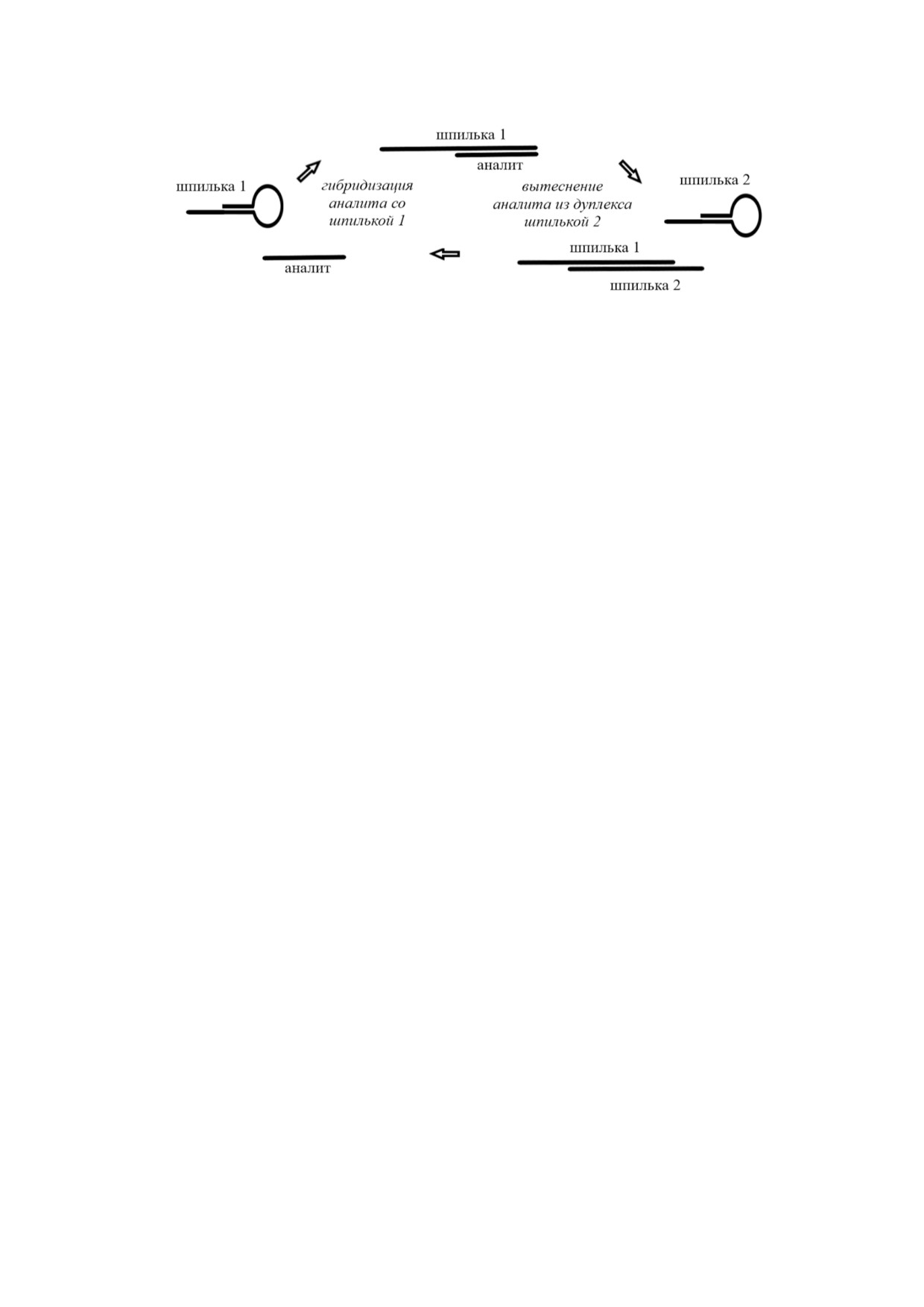
Более того, было показано, что HCR может
Доступность освободившегося домена при
быть эффективна как метод для внутриклеточ
водит к осуществлению его реакции со шпиль
ной визуализации РНК. Так, в работе Wu et al.
кой 2. Такая реакция сопровождается вытесне
[105] описан флуоресцентный метод для оценки
нием аналита из первичного дуплекса, что ини
экспрессии мРНК в пикомолярном интервале
циирует начало следующего цикла амплифика
концентраций. Следует также отметить, что ис
ции. Таким образом, одна молекула аналита
пользование данного метода не требует приме
способна инициировать образование большего
нения реакции обратной транскрипции.
числа дуплексов. При использовании CHA
Помимо анализа нуклеиновых кислот, опи
чрезвычайно важно уделять серьезное внимание
сано применение метода HCR в анализе ионов
предотвращению взаимодействия шпилек меж
ртути, АТФ, а также белков. Guo et al. [106] в ка
ду собою в отсутствие аналита, т.к. протекание
честве модельных белковых биомаркеров ис
такой реакции будет повышать фоновый сигнал
пользовали α фетопротеин и простат специфи
анализа и тем самым понижать его чувствитель
ческий антиген. В присутствии аналитов на по
ность. Таким образом, в отличие от HCR, кото
верхности электрода происходило формирова
рая, как и CHA, протекает без использования
ние иммунных комплексов. Вторичные антите
какого либо фермента, в случае CHA повыше
ла были ковалентно связаны с ДНК олигонук
ние интенсивности аналитического сигнала
леотидом, запускающим HCR. Предел обнару
происходит из за того, что молекула аналита
жения, полученный при вольтамперометричес
многократно участвует в проведении индика
кой детекции, для α фетопротеина составил
торной реакции.
0,25 пг/мл, для простат специфического анти
На сегодняшний день большая часть иссле
гена - 0,17 пг/мл.
дований с использованием CHA нацелена на
С помощью HCR успешно проводилась
разработку методов анализа микроРНК, кото
идентификация опухолевых клеток [107]. Мето
рые считаются перспективными биомаркерами
ды данного направления основаны на специфи
для диагностики онкологических заболеваний
ческом взаимодействии аптамеров с маркерами,
[109]. Продукты CHA могут быть зарегистриро
расположенными на поверхности исследуемых
ваны различными физико химическими мето
клеток.
дами. Так, в работе Zhang et al. [110] для детек
Как и HCR, метод каталитической сборки
ции микроРНК miR 21 с применением CHA ис
шпилек (CHA) не требует применения каких ли
пользовали электрофорез. Для данного метода
бо ферментов. Как видно на рис. 8, в основе
предел обнаружения микроРНК составил
CHA лежит использование двух олигонуклеоти
10 пМ. Электрохимический метод анализа с
дов шпилек [108]. За счет комплементарности
применением CHA использован в работе Shuai
первая шпилька может взаимодействовать с
et al. [111], в которой шпилька 1 была иммоби
аналитом, что приводит к образованию дуплек
лизована на поверхности электрода, а шпилька 2
БИОХИМИЯ том 85 вып. 2 2020
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
191
была модифицирована биотином/конъюгатом
Благодаря этому многие аналиты могут опреде
щелочной фосфатазы со стрептавидином. Пре
ляться в фемто и пикомолярных концентраци
дел детекции miR 21 составил 50 аМ, линейный
ях. Описаны также методы, позволяющие де
диапазон - от 0,1 фМ до 100 пМ.
тектировать исследуемые вещества в аттомоляр
Флуоресцентный гомогенный метод опреде
ных концентрациях. Такая чувствительность ме
ления микроРНК miR let 7a с применением мо
тодов позволяет детектировать практически все
лекулярного маяка и CHA для амплификации
значимые аналиты в реальных образцах.
регистрируемого сигнала был описан в работе
В то же время существует достаточное коли
Jiang et al. [112]. Данный метод позволил опре
чество вопросов, которые должны быть разре
делить анализируемую микроРНК с концентра
шены в будущем. В первую очередь это касается
цией, равной 1 пМ. Линейный диапазон метода
ложноположительных и ложноотрицательных
находился в интервале от 1 пМ до 2 нМ.
сигналов, получаемых в некоторых случаях при
Следует заметить, что некоторые авторы,
работе с реальными образцами. Другой важный
стремясь дополнительно повысить чувствитель
вопрос, ожидающий своего разрешения, заклю
ность аналитических методов, используют не
чается в том, что исследователи из различных
один метод, а комбинацию двух разных ампли
научных групп, используя идентичные методы
фикационных методов. Данные комбинации по
амплификации и детекции, разрабатывают ме
лучили название каскадных методов амплифи
тоды количественного определения аналитов с
кации [24]. Так, в работах Dong et al. [113] и Xu et
пределами обнаружения, которые часто отлича
al. [114] были объединены, соответственно, CHA
ются между собой на несколько порядков. При
и RCA c методом амплификации с использова
чины таких расхождений должны быть обяза
нием никазы. Сочетание RCA и EASA было ис
тельно выяснены в ближайшее время. Следует
пользовано для идентификации ДНК генетичес
отметить, что многие описанные методы с низ
ки модифицированной сои MON89788. Предел
кими значениями предела обнаружения имеют
обнаружения этого метода составил 45 аМ [115].
невысокую чувствительность, что не позволяет
Для повышения чувствительности анализа RCA
их использовать для определения аналитов,
использовалось также его сопряжение с CHA
концентрация которых при патологии не силь
[116]. Однако предел обнаружения в этом случае
но отличается от их концентрации в норме, на
был достаточно высок (100 фМ).
пример, микроРНК miR 21 [120]. Нужно также
Флуоресцентный биосенсор был разработан
заметить, что количество опубликованных ра
для определения бисфенола А с использованием
бот по применению аналитических методов,
сочетания метода с формированием Y структур
сопряженных с методами амплификации, для
и EASA [117]. Предел обнаружения бисфенола А
количественного определения биомаркеров
составил 5 фМ, линейный диапазон - от 10 фМ
аналитов в реальных образцах невысоко, а боль
до 10 нМ. Метод определения аденозина с при
шинство работ проводилось по их определению
менением HCR и EASA был опубликован Sun et
лишь в буферных растворах. Все вышеперечис
al. [118]. В некоторых случаях применялся дру
ленное указывает на необходимость продолже
гой подход, когда один и тот же метод использо
ния интенсивных научных исследований по
вался несколько раз для амплификации различ
оценке применимости в биоанализе и усовер
ных промежуточных соединений [119].
шенствованию изотермических методов ампли
фикации, а также разработке на их основе высо
Таким образом, в данном обзоре представле
кочувствительных и высокоселективных мето
ны данные по изотермическим методам ампли
дов анализа с высокой точностью измерений,
фикации нуклеиновых кислот, которые были
которые так необходимы для практического
разработаны как альтернатива ПЦР. В настоя
применения.
щее время эти методы активно и успешно при
меняются в биоанализе для повышения детек
тируемости и чувствительности количественно
Финансирование. Работа выполнена при под
го определения анализируемых соединений -
держке Российского научного фонда (грант 17
как нуклеиновых кислот, так и веществ другой
14 01042).
химической природы (белков, ферментов, анти
Конфликт интересов. Авторы заявляют об от
биотиков, наркотиков и т.д.), определение кото
сутствии конфликта интересов.
рых построено на применении ДНК/РНК апта
Соблюдение этических норм. Настоящая
меров. Большинство методов с использованием
статья не содержит описания каких либо иссле
изотермической амплификации нуклеиновых
дований с участием людей или использованием
кислот обладают высокой чувствительностью.
животных в качестве объектов.
БИОХИМИЯ том 85 вып. 2 2020
192
БОДУЛЕВ, САХАРОВ
СПИСОК ЛИТЕРАТУРЫ
1.
Manzanares Palenzuela, C.L., de los Santos Alvarez, N.,
northern Thailand, Am. J. Tropical Med. Hygiene, 83,
Lobo Castanon, M.J., and Lopez Ruiz, B.
(2015)
56-60, doi: 10.4269/ajtmh.2010.09 0630.
Multiplex electrochemical DNA platform for femtomolar
15.
Gao, X., Sun, B., and Guan, Y. (2019) Pullulan reduces
level quantification of genetically modified soybean,
the non specific amplification of loop mediated isother
Biosens. Bioelectron., 68, 259-265, doi: 10.1016/j.bios.
mal amplification (LAMP), Anal. Bioanal. Chem., 411,
2015.01.007.
1211-1218, doi: 10.1007/s00216 018 1552 2.
2.
Cavanaugh, S.E., and Bathrick, A.S. (2018) Direct PCR
15а.
Yang, Q., Domesle, K.J., and Ge, B. (2018) Loop mediat
amplification of forensic touch and other challenging DNA
ed isothermal amplification for Salmonella detection in
samples: a review, Foressic Sci. Int. Genet., 32, 40-49,
food and feed: current applications and future directions,
doi: 10.1016/j.fsigen.2017.10.005.
Foodborne Pathogens and Disease, 15, 309-331, doi:
3.
Higuchi, R., Dollinger, G., Walsh, P.S., and Griffith, R.
10.1089/fpd.2018.2445.
(1992) Simultaneous amplification and detection of specif
15б. Maruyama, F., Kenzaka, T., Yamaguchi, N., Tani, K., and
ic DNA sequences, Biotechnology (NY), 10, 413-417.
Nasu, M. (2003) Detection of bacteria carrying the stx2
4.
Алексеев Я.И., Белов Ю.В., Варламов Д.А., Конова
gene by in situ loop mediated isothermal amplification,
лов С.В., Курочкин В.Е., Маракушин Н.Ф., Петров А.И.,
Appl. Environ. Microbiol., 69, 5023-5028, doi: 10.1128/
Петряков А.О., Румянцев Д.А., Скоблилов Е.Ю., Со
aem.69.8.5023 5028.2003.
колов В.Н., Фесенко В.А., Чернышев А.В. (2006) При
16.
Щит И.Ю., Игнатов К.Б., Бикетов С.Ф. (2018) Cрав
боры для диагностики биологических объектов на ос
нительный анализ методов LAMP и ПЦР РВ для об
нове метода полимеразной цепной реакции в реаль
наружения возбудителей сапа и мелиоидоза, Клинич.
ном времени (ПЦР РВ), Научное приборостроение, 16,
лаб. диагностика, 63.
132-136.
17.
Макарова Ю.А., Зотиков А.А., Белякова Г.А., Алексеев Б.Я.,
5.
Borst, A., Box, A.T.A., and Fluit, A.C. (2004) False posi
Шкурников М.Ю. (2018) Изотермическая петлевая
tive results and contamination in nucleic acid amplification
амплификация: эффективный метод экспресс диаг
assays: suggestions for a prevent and destroy strategy, Eur. J.
ностики в онкологии, Онкоурология,
14,
88-99,
Clin. Microbiol. Infect. Dis., 23, 289-299, doi: 10.1007/
doi: 10.17650/1726 9776 2018 14 2 88 99.
s10096 004 1100 1.
18.
Wong, Y.P., Othman, S., Lau, Y.L., Radu, S., and Chee, H.Y.
6.
Notomi, T., Okayama, H., Masubuchi, H., Yonekawa, T.,
(2018) Loop mediated isothermal amplification (LAMP):
Watanabe, K., Amino, N., and Hase, T. (2000) Loop
a versatile technique for detection of microorganisms,
mediated isothermal amplification of DNA, Nucleic Acids
J. Appl. Microbiol., 124, 626-643, doi: 10.1111/jam.13647.
Res., 28, E63, doi: 10.1093/nar/28.12.e63.
19.
Compton, J. (1991) Nucleic acid sequence based amplifi
7.
Yan, L., Zhou, J., Zheng, Y., Gamson, A.S., Roembke, B.T.,
cation, Nature, 350, 91-92, doi: 10.1038/350091a0.
Nakayama, S., and Sintim, H.O. (2014) Isothermal ampli
20.
Kievits, T., van Gemen, B., van Strijp, D., Schukkink, R.,
fied detection of DNA and RNA, Mol. Biosyst., 10,
Dircks, M., Adriaanse, H., Malek, L., Sooknanan, R., and
970-1003, doi: 10.1039/c3mb70304e.
Lens, P. (1991) NASBA isothermal enzymatic in vitro
8.
Mori, Y., Kitao, M., Tomita, N., and Notomi, T. (2004)
nucleic acid amplification optimized for the diagnosis of
Real time turbidimetry of LAMP reaction for quantifying
HIV 1 infection, J. Virol. Methods, 35, 273-286.
template DNA, J. Biochem. Biophys. Methods,
59,
21.
Mader, A., Riehle, U., Brandstetter, T., Stickeler, E., and
145-157, doi: 10.1016/j.jbbm.2003.12.005.
Ruehe, J. (2012) Universal nucleic acid sequence based
9.
Tomita, N., Mori, Y., Kanda, H., and Notomi, T. (2008)
amplification for simultaneous amplification of
Loop mediated isothermal amplification (LAMP) of gene
messengerRNAs and microRNAs, Anal. Chim. Acta, 754,
sequences and simple visual detection of products, Nat.
1-7, doi: 10.1016/j.aca.2012.09.045.
Protoc., 3, 877-882, doi: 10.1038/nprot.2008.57.
22.
Ma, Y., Dai, X., Hong, T., Munk, G.B., and Libera, M.
10.
Pang, B., Yao, S., Xu, K., Wang, J., Song, X.L., Mu, Y.,
(2017) A NASBA on microgel tethered molecular beacon
Zhao, C., and Li, J. (2019) A novel visual mixed dye for
microarray for real time microbial molecular diagnostics,
LAMP and its application in the detection of foodborne
Analyst, 142, 147-155, doi: 10.1039/c6an02192a.
pathogens, Anal. Biochem., 574, 1-6, doi: 10.1016/j.ab.
23.
Lu, X., Shi, X., Wu, G., Wu, T., Qin, R., and Wang, Y.
2019.03.002.
(2017) Visual detection and differentiation of Classic Swine
11.
Troger, V., Niemann, K., Gartig, C., and Kuhlmeier, D.
Fever Virus strains using nucleic acid sequence based
(2015) Isothermal amplification and quantification of
amplification (NASBA) and G quadruplex DNAzyme
nucleic acids and its use in microsystems, J. Nanomed.
assay, Scientific Reports, 7, 44211, doi: 10.1038/srep44211.
Nanotechnol.,
6,
282-298, doi:
10.4172/2157
24.
Zeng, W., Yao, W., Wang, Y., Li, Y., Bermann, S.M., Ren, Y.,
7439.1000282.
Shi, C., Song, X., Huang, Q., Zheng, S., and Wang, Q.
12.
Najian, A.B.N., Foo, P.C., Ismail, N., Kim Fatt, L., and
(2017) Molecular detection of genotype II grass carp
Yean, C.Y. (2019) Probe specific loop mediated isother
reovirus based on nucleic acid sequence based amplifica
mal amplification magnetogenosensor assay for rapid and
tion combined with enzyme linked immunosorbent assay
specific detection of pathogenic Leptospira, Mol. Cell.
(NASBA ELISA), J. Virol. Methods,
243,
92-97,
Probes, 44, 63-68, doi: 10.1016/j.mcp.2019.03.001.
doi: 10.1016/j.jviromet.2017.02.001.
13.
Shchit, I.Yu., Ignatov, K.B., Kudryavtseva, T.Yu.,
25.
Honsvall, B.K., and Robertson, L.J. (2017) From research
Shishkova, N.A., Mironova, R.I., Marinin, L.I.,
lab to standard environmental analysis tool: will NASBA
Mokrievich, A.N., Kramarov, V.M., Biketov, S.F., and
make the leap? Water Res., 109, 389-397, doi: 10.1016/
Dyatlov, I.A. (2017) The use of loop mediated isothermal
j.watres.2016.11.052.
DNA amplification for the detection and identification of
26.
Vincent, M., Xu, Y., and Kong, H. (2004) Helicase depen
the anthrax pathogen, Mol. Genet. Microbiol. Virol., 32,
dent isothermal DNA amplification, EMBO Reports, 5,
100-108, doi: 10.3103/S0891416817020094.
795-800, doi: 10.1038/sj.embor.7400200.
14.
Poschl, B., Waneesorn, J., Thekisoe, O., Chutipongvivate, S.,
27.
Zhao, Y., Chen, F., Li, Q., Wang, L., and Fan, C. (2015)
and Panagiotis, K. (2010) Comparative diagnosis of malar
Isothermal amplification of nucleic acids, Chem. Rev., 115,
ia infections by microscopy, nested PCR, and LAMP in
12491-12545, doi: 10.1021/acs.chemrev.5b00428.
БИОХИМИЯ том 85 вып. 2 2020
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
193
28.
Barreda Garcia, S., Miranda Castro, R., de los Santos
41.
Jia, H., Wang, Z., Wang, C., Chang, L., and Li, Z. (2014)
Alvarez, N., Miranda Ordieres, A.J., and Lobo Castanon, M.J.
Real time fluorescence detection of Hg2+ ions with high
(2018) Helicase dependent isothermal amplification: a
sensitivity by exponentially isothermal oligonucleotide
novel tool in the development of molecular based analyti
amplification, RSC Adv., 4, 9439-9444, doi: 10.1039/
cal systems for rapid pathogen detection, Anal. Bioanal.
C3RA45986A.
Chem., 410, 679-693, doi: 10.1007/s00216 017 0620 3.
42.
Tian, L., and Weizmann, Y. (2013) Real time detection of
29.
Mahalanabis, M., Do, J., ALMuayad, H., Zhang, J.Y., and
telomerase activity using the exponential isothermal ampli
Klapperich, C.M. (2011) An integrated disposable device
fication of telomere repeat assay, J. Am. Chem. Soc., 135,
for DNA extraction and helicase dependent amplification,
1661-1664, doi: 10.1021/ja309198j.
Biomed. Microdevices, 13, 353-359, doi: 10.1007/s10544
43.
Wang, L., Ren, M., Zhang, Q., Tang, B., and Zhang, C.
009 9391 8.
(2017) Excision repair initiated enzyme assisted bicyclic
30.
Ao, W., Aldous, S., Woodruff, E., Hicke, B., Rea, L.,
cascade signal amplification for ultrasensitive detection of
Kreiswirth, B., and Jenison, R. (2012) Rapid detection of
uracil DNA glycosylase, Anal. Chem., 89, 4488-4494,
rpoB gene mutations conferring rifampin resistance in
doi: 10.1021/acs.analchem.6b04673.
Mycobacterium tuberculosis, J. Clin. Microbiol.,
50,
44.
Walker, G.T., Fraiser, M.S., Schram, J.L., Little, M.C.,
2433-2440, doi: 10.1128/JCM.00208 12.
Nadeau, J.G., and Malinowski, D.P. (1992) Strand dis
31.
Tang, R., Yang, H., Gong, Y., You, M., Liu, Z., Choi, J.R.,
placement amplification - an isothermal, in vitro DNA
Wen, T., Qu, Z., Mei, Q., and Xu, F. (2017) A fully dispos
amplification technique, Nucleic Acids Res.,
20,
able and integrated paper based device for nucleic acid
1691-1696, doi: 10.1093/nar/20.7.1691.
extraction, amplification and detection, Lab on a Chip, 17,
45.
Shi, C., Ge, Y., Gu, H., and Ma, C. (2011) Highly sensitive
1270-1279, doi: 10.1039/c6lc01586g.
chemiluminescent point mutation detection by circular
32.
Barreda Garcia, S., Miranda Castro, R., de Los Santos
strand displacement amplification reaction, Biosens.
Alvarez, N., Miranda Ordieres, A.J., and Lobo Castanon, M.J.
Bioelectron., 26, 4697-4701, doi: 10.1016/j.bios.2011.05.017.
(2017) Solid phase helicase dependent amplification and
46.
Zhao, Y., Zhou, L., and Tang, Z. (2013) Cleavage based
electrochemical detection of Salmonella on highly stable
signal amplification of RNA, Nat. Commun., 4, 1493,
oligonucleotide modified ITO electrodes, Chem. Commun.,
doi: 10.1038/ncomms2492.
53, 9721-9724, doi: 10.1039/c7cc05128j.
47.
Shi, C., Liu, Q., Ma, C., and Zhong, W.
(2014)
33.
Schwenkbier, L., Pollok, S., Rudloff, A., Sailer, S., Cialla
Exponential strand displacement amplification for detec
May, D., Weber, K., and Popp, J. (2015) Non instrument
tion of microRNAs, Anal. Chem.,
86,
336-339,
ed DNA isolation, amplification and microarray based
doi: 10.1021/ac4038043.
hybridization for a rapid on site detection of devastating
48.
Ren, R., Leng, C., and Zhang, S. (2010) Detection of
Phytophthora kernoviae, Analyst,
140,
6610-6618,
DNA and indirect detection of tumor cells based on circu
doi: 10.1039/c5an00855g.
lar strand replacement DNA polymerization on electrode,
34.
Toldra, A., Jauset Rubio, M., Andree, K.B., Fernandez
Chem. Commun., 46, 5758-5760, doi: 10.1039/C002466J.
Tejedor, M., Diogene, J., Katakis, I., O’Sullivan, C.K., and
49.
Ding, C., Li, X., Ge, Y., and Zhang, S. (2010) Fluorescence
Campas, M. (2018) Detection and quantification of the
detection of telomerase activity in cancer cells based on
toxic marine microalgae Karlodinium veneficum and
isothermal circular strand displacement polymerization
Karlodinium armiger using recombinase polymerase ampli
reaction, Anal. Chem., 82, 2850-2855, doi: 10.1021/
fication and enzyme linked oligonucleotide assay, Anal.
ac902818w.
Chim. Acta, 1039, 140-148, doi: 10.1016/j.aca.2018.07.057.
50.
Zhu, C., Wen, Y., Li, D., Wang, L., Song, S., Fan, C., and
35.
Ma, F., Liu, M., Tang, B., and Zhang, C.Y. (2017) Sensi
Willner, I. (2009) Inhibition of the in vitro replication of
tive quantification of microRNAs by isothermal helicase
DNA by an aptamer protein complex in an autonomous
dependent amplification, Anal. Chem., 89, 6183-6188,
DNA machine, Chemistry, 15, 11898-11903, doi: 10.1002/
doi: 10.1021/acs.analchem.7b01113.
chem.200901275.
36.
Reid, M.S., Le, X.C., and Zhang, H. (2018) Exponential
51.
Li, Y., Zeng, Y., Mao, Y., Lei, C., and Zhang, S. (2014)
isothermal amplification of nucleic acids and assays for
Proximity dependent isothermal cycle amplification for
proteins, cells, small molecules, and enzyme activities: an
small molecule detection based on surface enhanced
EXPAR example, Angew. Chem. Int. Ed.,
57,
Raman scattering, Biosens. Bioelectron., 51, 304-309,
11856-11866, doi: 10.1002/anie.201712217.
doi: 10.3390/bios9020057.
37.
Van Ness, J., Van Ness, L.K., and Galas, D.J. (2003)
52.
Li, J., Macdonald, J., and von Stetten, F. (2019) Review: a
Isothermal reactions for the amplification of oligonu
comprehensive summary of a decade development of the
cleotides, Proc. Natl. Acad. Sci. USA, 100, 4504-4509,
recombinase polymerase amplification, Analyst, 144,
doi: 10.1073/pnas.0730811100.
31-67, doi: 10.1039/c8an01621f.
38.
Wang, H., Wang, H., Duan, X., Wang, X., Sun, Y., and Li, Z.
53.
Kersting, S., Rausch, V., Bier, F.F., and von Nickisch
(2017) Sensitive detection of mRNA by using specific
Rosenegk, M. (2014) Rapid detection of Plasmodium falci*
cleavage mediated isothermal exponential amplification
parum with isothermal recombinase polymerase amplifica
reaction, Sens. Actuat. B: Chemical, 252, 215-221,
tion and lateral flow analysis, Malar. J.,
13,
99,
doi: 10.1016/j.snb.2017.06.008.
doi: 10.1186/1475 2875 13 99.
39.
Li, R.D., Yin, B.C., and Ye, B.C. (2016) Ultrasensitive,
54.
Lobato, I.M., and O’Sullivan, C.K. (2018) Recombinase
colorimetric detection of microRNAs based on isothermal
polymerase amplification: basics, applications and recent
exponential amplification reaction assisted gold nanopar
advances, Trends Anal. Chem.,
98,
19-35,
ticle amplification, Biosens. Bioelectron., 15, 1011-1016,
doi: 10.1016/j.trac.2017.10.015.
doi: 10.1016/j.bios.2016.07.042.
55.
Mayboroda, O., Gonzalez Benito, A., Sabate del Rio, J.,
40.
Liu, H., Zhang, L., Xu, Y., Chen, J., Wang, Y., Huang, Q.,
Svobodova, M., Julich, S., Tomaso, H., O’Sullivan, C.K.,
Che, X., Liu, Y., Da, Z., Zou, X., and Li, Z. (2019)
and Katakis, I. (2016) Isothermal solid phase amplifica
Sandwich immunoassay coupled with isothermal exponen
tion system for detection of Yersinia pestis, Anal. Bioanal.
tial amplification reaction: an ultrasensitive approach for
Chem., 408, 671-676, doi: 10.1007/s00216 015 9177 1.
determination of tumor marker MUC1, Talanta, 204,
56.
Ng, B.Y., Xiao, W., West, N.P., Wee, E.J., Wang, Y., and
248-254, doi: 10.1016/j.talanta.2019.06.001.
Trau, M. (2015) Rapid, single cell electrochemical detec
БИОХИМИЯ том 85 вып. 2 2020
194
БОДУЛЕВ, САХАРОВ
tion of Mycobacterium tuberculosis using colloidal gold
using Phi29 DNA polymerase and multiply primed rolling
nanoparticles, Anal. Chem.,
87,
10613-10618,
circle amplification, Genome Res.,
11,
1095-1099,
doi: 10.1021/acs.analchem.5b03121.
doi: 10.1101/gr.180501.
57.
Tsaloglou, M.N., Nemiroski, A., Camci Unal, G.,
71.
Li, Y., Kim, H.J., Zheng, C., Chow, W.H.A., Lim, J.,
Christodouleas, D.C., Murray, L.P., Connelly, J.T., and
Keenan, B., Pan, X., Lemieux, B., and Kong, H. (2008)
Whitesides, G.M. (2017) Handheld isothermal amplifica
Primase based whole genome amplification, Nucleic Acids
tion and electrochemical detection of DNA in resource
Res., 36, e79, doi: 10.1093/nar/gkn377.
limited settings, Anal. Biochem.,
543,
116-121,
72.
Miao, P., Tang, Y., Wang, B., Yin, J., and Ning, L. (2015)
doi: 10.1016/j.ab.2017.11.025.
Signal amplification by enzymatic tools for nucleic acids,
58.
Yang, B., Kong, J., and Fang, X. (2019) Bandage like
Trends Anal. Chem., 67, 1-15, doi: 10.1016/j.trac.2014.
wearable flexible microfluidic recombinase polymerase
12.006.
amplification sensor for the rapid visual detection of nucle
73.
Mol, C.D., Kuo, C.F., Thayer, M.M., Cunningham, R.P.,
ic acids, Talanta, 204, 685-692, doi: 10.1016/j.talan
and Tainer, J.A. (1995) Structure and function of the mul
ta.2019.06.031.
tifunctional DNA repair enzyme exonuclease III, Nature,
59.
Fire, A., and Xu, S.Q. (1995) Rolling replication of short
374, 381-386, doi: 10.1038/374381a0.
DNA circles, Proc. Nat. Acad. Sci. USA, 92, 4641-4645,
74.
Zuo, X., Xia, F., Xiao, Y., and Plaxco, K.W.
(2010)
doi: 10.1073/pnas.92.10.4641.
Sensitive and selective amplified fluorescence DNA detec
60.
Goo, N. I., and Kim, D. E. (2016) Rolling circle amplifi
tion based on exonuclease III aided target recycling, J. Am.
cation as isothermal gene amplification in molecular diag
Chem. Soc., 132, 1816-1818, doi: 10.1021/ja909551b.
nostics, BioChip J., 10, 262-271, doi: 10.1007/s13206
75.
Yan, M., Bai, W., Zhu, C., Huang, Y., Yan, J., and Chen, A.
016 0402 6.
(2016) Design of nuclease based target recycling signal
61.
Nilsson, M., Barbany, G., Antson, D.O., Gertow, K., and
amplification in aptasensors, Biosens. Bioelectron., 77,
Landegren, U. (2000) Enhanced detection and distinction
613-623, doi: 10.1016/j.bios.2015.10.015.
of RNA by enzymatic probe ligation, Nat. Biotechnol., 18,
76.
Yang, W., Tian, J., Ma, Y., Wang, L., Zhao, Y., and Zhao, S.
791-793, doi: 10.1038/77367.
(2015) A label free fluorescent probe based on DNA tem
62.
Marciniak, J., Kummel, A.C., Esener, S.C., Heller, M.J.,
plated silver nanoclusters and exonuclease III assisted
and Messmer, B.T. (2008) Coupled rolling circle amplifica
recycling amplification detection of nucleic acid, Anal.
tion loop mediated amplification for rapid detection of
Chim. Acta, 900, 90-96, doi: 10.1016/j.aca.2015.10.015.
short DNA sequences, Biotechniques, 45, 275-280,
77.
Xu, L., Shen, X., Li, B., Zhu, C., and Zhou, X. (2017) G
doi: 10.2144/000112910.
Quadruplex based Exo III assisted signal amplification
63.
Li, X.H., Zhang, X.L., Wu, J., Lin, N., Sun, W.M., Chen, M.,
aptasensor for the colorimetric detection of adenosine,
Ou, Q.S., and Lin, Z.Y. (2019) Hyperbranched rolling cir
Anal. Chim. Acta,
980,
58-64, doi:
10.1016/j.aca.
cle amplification (HRCA) based fluorescence biosensor
2017.05.015.
for ultrasensitive and specific detection of single
78.
Gao, Y., and Li, B. (2013) G Quadruplex DNAzyme
nucleotide polymorphism genotyping associated with the
based chemiluminescence biosensing strategy for ultrasen
therapy of chronic hepatitis B virus infection, Talanta, 191,
sitive DNA detection: combination of exonuclease III
277-282, doi: 10.1016/j.talanta.2018.08.064.
assisted signal amplification and carbon nanotubes assisted
64.
Schopf, E., Liu, Y., Deng, J.C., Yang, S., Cheng, G., and
background reducing, Anal. Chem., 85, 11494-11500,
Chen, Y. (2011) Mycobacterium tuberculosis detection via
doi: 10.1021/ac402728d.
rolling circle amplification, Anal. Methods, 3, 267-273,
79.
Gan, X., Zhao, H., Chen, S., and Quan, X. (2015)
doi: 10.1039/C0AY00529K.
Electrochemical DNA sensor for specific detection of
65.
Cheng, Y., Zhang, X., Li, Z., Jiao, X., Wang, Y., and
picomolar Hg (II) based on exonuclease III assisted recy
Zhang, Y. (2009) Highly sensitive determination of
cling signal amplification, Analyst, 140, 2029-2036,
microRNA using target primed and branched rolling cir
doi: 10.1039/C5AN00082C.
cle amplification, Angew. Chem. Int. Ed. Engl., 48,
80.
Yang, Z., Sismour, A.M., and Benner, S.A.
(2007)
3268-3272, doi: 10.1002/anie.200805665.
Nucleoside α thiotriphosphates, polymerases and the
66.
Deng, R., Tang, L., Tian, Q., Wang, Y., Lin, L., and Li, J.
exonuclease III analysis of oligonucleotides containing
(2014) Toehold initiated rolling circle amplification for
phosphorothioate linkages, Nucleic Acids Res.,
35,
visualizing individual microRNAs in situ in single cells,
3118-3127, doi: 10.1093/nar/gkm168.
Angew. Chem. Int. Ed. Engl.,
53,
2389-2393,
81.
Xu, Q., Cao, A., Zhang, L. F., and Zhang, C. Y. (2012)
doi: 10.1002/anie.201309388.
Rapid and label free monitoring of exonuclease III assist
67.
Mashimo, Y., Mie, M., Suzuki, S., and Kobatake, E.
ed target recycling amplification, Anal. Chem.,
84,
(2017) Detection of small RNA molecules by a combina
10845-10851, doi: 10.1021/ac303095z.
tion of branched rolling circle amplification and biolumi
82.
Wang, Y., Wu, Y., Wang, Y., Zhou, B., and Wu, S. (2015) A
nescent pyrophosphate assay, Anal. Bioanal. Chem., 401,
sensitive immobilization free electrochemical assay for
221-227, doi: 10.1007/s00216 011 5083 3.
T4PNK activity based on exonuclease III assisted recy
68.
Cui, W., Wang, L., Xu, X., Wang, Y., and Jiang, W. (2017)
cling, RSC Adv.,
5,
75348-75353, doi:
10.1039/
A loop mediated cascade amplification strategy for highly
C5RA12849H.
sensitive detection of DNA methyltransferase activity,
83.
Li, W., Liu, X., Hou, T., Li, H., and Li, F.
(2015)
Sens.
Actuat.
B:
Chemical,
244,
599-605,
Ultrasensitive homogeneous electrochemical strategy for
doi: 10.1016/j.snb.2017.01.013.
DNA methyltransferase activity assay based on
69.
Qing, T., He, D., He, X., Wang, K., Xu, F., Wen, L.,
autonomous exonuclease III assisted isothermal cycling
Shangguan, J., Mao, Z., and Lei, Y. (2016) Nucleic acid
signal amplification, Biosens. Bioelectron., 70, 304-309,
tool enzymes aided signal amplification strategy for bio
doi: 10.1016/j.bios.2015.03.060.
chemical analysis: status and challenges, Anal. Bioanal.
84.
Min, X., Xia, L., Zhuang, Y., Wang, X., Du, J., Zhang, X.,
Chem.,
408,
2793-2811, doi:
10.1007/s00216 015
Lou, X., and Xia, F. (2017) An AIEgens and exonuclease
9240 y.
III aided quadratic amplification assay for detecting and
70.
Dean, F.B., Nelson, J.R., Giesler, T.L., and Lasken, R.S.
cellular imaging of telomerase activity, Sci. Bull., 62,
(2001) Rapid amplification of plasmid and phage DNA
997-1003, doi: 10.1016/j.scib.2017.06.008.
БИОХИМИЯ том 85 вып. 2 2020
ИЗОТЕРМИЧЕСКИЕ МЕТОДЫ АМПЛИФИКАЦИИ
195
85.
Yan, L., Nakayama, S., and Sintim, H.O. (2013) Probe
deoxyribonucleic acid sensor, Anal. Chim. Acta, 883,
design rules and effective enzymes for endonuclease based
67-73, doi: 10.1016/j.aca.2015.04.058.
detection of nucleic acids, Bioorg. Med. Chem., 21,
99. Wang, T., Zhang, Z., Li, Y., and Xie, G. (2015) Amplified
6181-6185, doi: 10.1016/j.bmc.2013.04.009.
electrochemical detection of mecA gene in methicillin
86.
Liu, S., Zhang, C., Ming, J., Wang, C., Liu, T., and Li, F.
resistant Staphylococcus aureus based on target recycling
(2013) Amplified detection of DNA by an analyte induced
amplification and isothermal strand displacement poly
Y shaped junction probe assembly followed with a nicking
merization reaction, Sens. Actuat. B: Chemical, 221,
endonuclease mediated autocatalytic recycling process,
148-154, doi: 10.1016/j.snb.2015.06.057.
Chem. Commun., 49, 7947-7949, doi: 10.1039/c3cc45211e.
100. Dirks, R.M., and Pierce, N.A. (2004) Triggered amplifica
87.
Chen, M., Gan, N., Li, T., Wang, Y., Xu, Q., and Chen, Y.
tion by hybridization chain reaction, Proc. Natl. Acad. Sci.
(2017) An electrochemical aptasensor for multiplex antibi
USA, 101, 15275-15278, doi: 10.1073/pnas.0407024101.
otics detection using Y shaped DNA based metal ions
101. Xuan, F., and Hsing, I.M. (2014) Triggering hairpin free
encoded probes with NMOF substrate and CSRP target
chain branching growth of fluorescent DNA dendrimers
triggered amplification strategy, Anal. Chim. Acta, 968,
for nonlinear hybridization chain reaction, J. Am. Chem.
30-39, doi: 10.1016/j.aca.2017.03.024.
Soc., 136, 9810-9813, doi: 10.1021/ja502904s.
88.
Wang, Q., Yang, L., Yang, X., Wang, K., He, L., Zhu, J.,
102. Bi, S., Chen, M., Jia, X., Dong, Y., and Wang, Z. (2015)
and Su, T. (2012) An electrochemical DNA biosensor
Hyperbranched hybridization chain reaction for triggered
based on the “Y” junction structure and restriction
signal amplification and concatenated logic circuits,
endonuclease aided target recycling strategy, Chem.
Angew. Chem. Int. Ed., 54, 8144-8148, doi: 10.1002/
Commun., 48, 2982-2984, doi: 10.1039/c2cc17679c.
anie.201501457.
89.
Huang, Y., Wang, W., Wu, T., Xu, L. P., Wen, Y., and
103. Yang, X., Yuebo, Yu, Y., and Gao, Z. (2014) A highly sen
Zhang, X. (2016) A three line lateral flow biosensor for
sitive plasmonic DNA assay based on triangular silver
logic detection of microRNA based on Y shaped junction
nanoprism etching, ACS Nano,
85,
4902-4907,
DNA and target recycling amplification, Anal. Bioanal.
doi: 10.1021/nn5008786.
Chem., 408, 8195-8202, doi: 10.1007/s00216 016 9925 x.
104. Miao, P., Tang, Y., and Yin, J. (2015) MicroRNA detection
90.
Zhao, Z., Chen, S., Wang, J., Su, J., Xu, J., Mathur, S.,
based on analyte triggered nanoparticle localization on a
Fan, C., and Song, S. (2017) Nuclease free target recycling
tetrahedral DNA modified electrode followed by
signal amplification for ultrasensitive multiplexing DNA
hybridization chain reaction dual amplification, Chem.
biosensing, Biosens. Bioelectron.,
94,
605-608,
Commun., 51, 15629-15632, doi: 10.1039/C5CC05499K.
doi: 10.1016/j.bios.2017.03.051.
105. Wu, Z., Liu, G. Q., Yang, X. L., and Jiang, J. H. (2015)
91.
Kiesling, T., Cox, K., Davidson, E.A., Dretchen, K.,
Electrostatic nucleic acid nanoassembly enables hybridiza
Grater, G., Hibbard, S., and Danielsen, M.
(2007)
tion chain reaction in living cells for ultrasensitive mRNA
Sequence specific detection of DNA using nicking
imaging, J. Am. Chem. Soc., 137, 6829-6836, doi: 10.1021/
endonuclease signal amplification (NESA), Nucleic Acids
jacs.5b01778.
Res., 35, e117, doi: 10.1093/nar/gkm654.
106. Guo, J., Wang, J., Zhao, J., Guo, Z., and Zhang, Y. (2016)
92.
Lin, Z., Yang, W., Zhang, G., Liu, Q., Qiu, B., Cai, Z., and
Ultrasensitive multiplexed immunoassay for tumor bio
Chen, G. (2011) An ultrasensitive colorimeter assay strate
markers based on DNA hybridization chain reaction
gy for p53 mutation assisted by nicking endonuclease signal
amplifying signal, ACS Appl. Materials Interfaces, 8,
amplification, Chem. Commun.,
47,
9069-9071,
6898-6904, doi: 10.1021/acsami.6b00756.
doi: 10.1039/C1CC13146J.
107. Jie, G., and Jie, G. (2016) Sensitive electrochemilumines
93.
Chen, J., Zhang, J., Li, J., Fu, F., Yang, H.H., and Chen, G.
cence detection of cancer cells based on a CdSe/ZnS
(2010) An ultrahighly sensitive and selective electrochemi
quantum dot nanocluster by multibranched hybridization
cal DNA sensor via nicking endonuclease assisted current
chain reaction on gold nanoparticles, RSC Adv., 6,
change amplification, Chem. Commun., 46, 5939-5941,
24780-24785, doi: 10.1039/C6RA00750C.
doi: 10.1039/c0cc00748j.
108. Choi, H.M.T., Chang, J.Y., Trinh, L.A., Padilla, J.E.,
94.
Vijayan, A.N., Liu, Z., Zhao, H., and Zhang, P. (2019)
Fraser, S.E., and Pierce, N.F. (2010) Programmable in situ
Nicking enzyme assisted signal amplifiable Hg2+ detection
amplification for multiplexed imaging of mRNA expression,
using upconversion nanoparticles, Anal. Chim. Acta, 1072,
Nat. Biotechnol., 28, 1208-1214, doi: 10.1038/nbt.1692.
75-80, doi: 10.1016/j.aca.2019.05.001.
109. Bertoli, G., Cava, C., and Castiglioni, I.
(2015)
95.
Xie, L.S., Li, T.H., Hu, F.T., Jiang, Q.L., Wang, Q.Q., and
MicroRNAs: new biomarkers for diagnosis prognosis, ther
Gan, N. (2019) A novel microfluidic chip and antibody
apy prediction and therapeutic tools for breast cancer,
aptamer based multianalysis method for simultaneous
Theranostics, 5, 1122-1143, doi: 10.7150/thno.11543.
determination of several tumor markers with polymeriza
110. Zhang, J., Zhang, W., and Gu, Y. (2018) Enzyme free
tion nicking reactions for homogenous signal amplifica
isothermal target recycled amplification combined with
tion, Microchem. J.,
147,
454-462, doi:
10.1016/
PAGE for direct detection of microRNA 21, Anal.
j.microc.2019.03.028.
Biochem., 550, 117-122, doi: 10.1016/j.ab.2018.04.024.
96.
Guo, Q., Yang, X., Wang, K., Tan, W., Li, W., Tang, H.,
111.
Shuai, H. L., Huang, K. J., Xing, L. L., and Chen, Y. X.
and Li, H. (2009) Sensitive fluorescence detection of
(2016) Ultrasensitive electrochemical sensing platform for
nucleic acids based on isothermal circular strand displace
microRNA based on tungsten oxide graphene composites
ment polymerization reaction, Nucleic Acids Res., 37, e20,
coupling with catalyzed hairpin assembly target recycling
doi: 10.1093/nar/gkn1024.
and enzyme signal amplification, Biosens. Bioelectron., 86,
97.
Giuffrida, M.C., Zanoli, L.M., D’Agata, R., Finotti, A.,
337-345, doi: 10.1016/j.bios.2016.06.057.
Gambari, R., and Spoto, G. (2015) Isothermal circular
112. Jiang, Z., Wang, H., Zhang, X., Liu, C., and Li, Z. (2014)
strand displacement polymerization of DNA and
An enzyme free signal amplification strategy for sensitive
microRNA in digital microfluidic devices, Anal. Bioanal.
detection of microRNA via catalyzed hairpin assembly,
Chem., 407, 1533-1543, doi: 10.1007/s00216 014 8405 4.
Anal. Methods, 6, 9477-9482, doi: 10.1039/C4AY02142H.
98.
Gao, F., Du, L., Zhang, Y., Tang, D., and Du, Y. (2015)
113. Dong, G., Dai, J., Jin, L., Shi, H., Wang, F., Zhou, C.,
Molecular beacon mediated circular strand displacement
Zheng, B., Guo, Y., and Dan Xiao, D. (2019) A rapid
strategy for constructing a ratiometric electrochemical
room temperature DNA amplification and detection strat
БИОХИМИЯ том 85 вып. 2 2020
196
БОДУЛЕВ, САХАРОВ
egy based on nicking endonuclease and catalyzed hairpin
117. Chen, J., and Zhou, S. (2016) Label free DNA Y junction
assembly, Anal. Methods, 11, 2537-2541, doi: 10.1039/
for bisphenol A monitoring using exonuclease III based
C9AY00507B.
signal protection strategy, Biosens. Bioelectron.,
77,
114. Xu, J., Guo, J., Maina, S.W., Yang, Y., Hu, Y., Li, X., Qiu,
277-283, doi: 10.1016/j.bios.2015.09.042.
J., and Xin, Z. (2018) An aptasensor for Staphylococcus
118. Sun, J., Jiang, W., Zhu, J., Li, W., and Wang, L. (2015)
aureus based on nicking enzyme amplification reaction and
Label free fluorescence dual amplified detection of
rolling circle amplification, Anal. Biochem., 549, 136-142,
adenosine based on exonuclease III assisted DNA cycling
doi: 10.1016/j.ab.2018.03.013.
and hybridization chain reaction, Biosens. Bioelectron., 70,
115. Chen, D., Zhang, M., Ma, M., Hai, H., Li, J., and Shan, Y.
15-20, doi: 10.1016/j.bios.2015.03.014.
(2019) A novel electrochemical DNA biosensor for transgenic
119. Bi, S., Li, L., and Cui, Y. (2012) Exonuclease assisted cas
soybean detection based on triple signal amplification, Anal.
caded recycling amplification for label free detection of
Chim. Acta, 1078, 24-31, doi: 10.1016/j.aca.2019.05.074.
DNA, Chem. Commun., 48, 1018-1020, doi: 10.1039/
116. Song, H., Yang, Z., Jiang, M., Zhang, G., Gao, Y., and
c1cc16684k.
Shen, Z., Wu, Z. S., and Lou, Y. (2019) Target catalyzed
120. D’Agata, R., and Spoto, G. (2019) Advanced methods for
hairpin structure mediated padlock cyclization for ultra
microRNA biosensing: a problem solving perspective,
sensitive rolling circle amplification, Talanta, 204, 29-35,
Anal. Bioanal. Chem., 411, 4425-4444, doi: 10.1007/
doi: 10.1016/j.talanta.2019.05.057.
s00216 019 01621 8.
ISOTHERMAL AMPLIFICATION METHODS OF NUCLEIC ACIDS
AND THEIR APPLICATION IN BIOANALYSIS
Review
O. L. Bodulev and I. Yu. Sakharov*
Lomonosov Moscow State University, Faculty of Chemistry, 119991 Moscow, Russia; E*mail: sakharovivan@gmail.com
Received August 13, 2019
Revised November 1, 2019
Accepted November 1, 2019
In recent years, isothermal amplification methods of nucleic acids are intensely developed as an alternative of poly
merase chain reaction (PCR). The advantage of these methods is that the amplification of nucleic acids is carried out
at a constant temperature, in contrast to PCR, which requires a cyclic change in temperature. In addition, isothermal
amplification of nucleic acids can be carried out directly in living cells. This review presents the principles of the
isothermal amplification methods and demonstrates their high efficiency in the construction of new highly sensitive
methods for the analysis of both nucleic acids and enzymes responsible for their modification. Moreover, data are pre
sented on their successful application in bioanalysis of cells and biomolecules with the use of DNA/RNA aptamers.
Keywords: nucleic acids, amplification, isothermal, bioanalysis, aptamers
БИОХИМИЯ том 85 вып. 2 2020