БИОХИМИЯ, 2020, том 85, вып. 3, с. 324 - 334
УДК 577.21
НЕЙРОПРОТЕКТИВНЫЕ ЭФФЕКТЫ ПЕПТИДОВ В МОЗГЕ:
ТРАНСКРИПТОМНЫЕ ПОДХОДЫ К ИХ ИССЛЕДОВАНИЮ
(ПЕПТИДНАЯ РЕГУЛЯЦИЯ МОЗГА)
Обзор
© 2020
И.Б. Филиппенков*, Л.В. Дергунова, С.А. Лимборская, Н.Ф. Мясоедов
Институт молекулярной генетики РАН, 123182 Москва, Россия; электронная почта: Filippenkov@img.ras.ru
Поступила в редакцию 05.12.2019
После доработки 05.12.2019
Принята к публикации 11.12.2019
В настоящее время все большее значение придается изучению механизмов действия лекарственных средств
на основе естественных регуляторных пептидов. Особое внимание уделяется пептидным препаратам, спо"
собствующим восстановлению функций мозга после острого нарушения мозгового кровообращения (ин"
сульта), которое продолжает уже многие годы оставаться одной из главных угроз для здоровья человека. Од"
нако молекулярно"генетические изменения в мозге в ответ на ишемию, а также механизмы протективных
эффектов пептидов изучены недостаточно, что ограничивает их применение и затрудняет разработку новых
более эффективных препаратов со специализированным воздействием на функции мозга. Перспективным
подходом для изучения механизмов повреждающего действия ишемии мозга, а также механизмов нейро"
протективных эффектов пептидных препаратов является транскриптомный анализ. Помимо мРНК, направ"
ляющих синтез белка, большое значение имеет изучение роли регуляторных РНК при ишемии для разра"
ботки новых стратегий нейропротекции. Наибольший интерес представляют микроРНК, а также цикличес"
кие РНК (циклоРНК), которые имеют замкнутую структуру и преимущественно мозгоспецифическую
экспрессию. ЦиклоРНК могут взаимодействовать с микроРНК, нивелировать их активность и тем самым
препятствовать микроРНК"опосредованной репрессии мРНК. В последнее время становится очевидным,
что анализ системы взаимодействий циклоРНК-микроРНК-мРНК является важной составляющей для
детального изучения механизмов повреждения и восстановления. В данном обзоре представлены результа"
ты анализа изменений работы генов при развитии ишемического повреждения, изучения транскриптомно"
го профиля клеток мозга под действием пептидов в условиях экспериментальной ишемии мозга, а также
сформулированы основные принципы механизмов пептидной регуляции при ишемическом повреждении.
КЛЮЧЕВЫЕ СЛОВА: транскриптомика, регуляторные пептиды, ишемия мозга, мРНК, некодирующие
РНК, микроРНК, циклические РНК.
DOI: 10.31857/S0320972520030033
Регуляторные пептиды - универсальные эн"
кие как энкефалины, нейротензин, холецисто"
догенные биологические вещества пептидной
кинин, кортистатин, орексин, ангиотензин II,
природы, основное назначение которых заклю"
брадикинин, меланоцитстимулирующие гормо"
чается в интеграции нервной, эндокринной и
ны и др., могут активно функционировать в
иммунной систем в единый функциональный
ЦНС [2, 3]. В настоящее время успешно разви"
континуум [1]. Многочисленные пептиды, та"
ваются медицинские технологии, в т.ч. исполь"
Принятые сокращения: cAMP - циклический аденозин монофосфат; circDLGAP4 - циклическая РНК гена мем"
браносвязанной гуанилаткиназы Dlgap4; circHECTD1 - циклическая РНК гена убиквитинлигазы, содержащей HECT"до"
мен, Hectd1; CREB - транскрипционный фактор, опосредующий связь с cAMP (cAMP response element"binding protein);
E2 - 17β"эстрадиол; GLP"1 - инсулино"глюкагоноподобный пептид"1; GPCR - метаботропный рецептор, связанный с
G"белком; HMGB1 - амфотерин (high"mobility group protein B1); MSH - меланоцитстимулирующий гормон; N"AcPGP -
N"ацетил"Pro"Gly"Pro; NFAT - активность ядерного фактора активированных T"клеток; NF"κB - ядерный фактор κB;
OGD/R - кислородно"глюкозная депривация-реоксигенация; P - прогестерон; PACAP - полипептид, активирующий
аденилатциклазу гипофиза; PGP - Pro"Gly"Pro; pMCAO - модель необратимой окклюзии средней мозговой артерии
(permanent middle cerebral artery occlusion model); RNA"seq - технология высокопроизводительного секвенирования РНК;
tMCAO - модель обратимой окклюзии средней мозговой артерии (transient middle cerebral artery occlusion model); АКТГ -
адренокортикотропный гормон; днРНК - длинные некодирующие РНК (long non"coding RNAs, lncRNAs); кэРНК - кон"
курентные эндогенные РНК (competitive endogenous RNAs, ceRNAs); нкРНК - некодирующие РНК (non"coding RNAs,
ncRNAs); циклоРНК - циклические РНК (circular RNAs, circRNAs).
* Адресат для корреспонденции.
324
ПЕПТИДНАЯ РЕГУЛЯЦИЯ МОЗГА
325
зующие нейропротективные, противовоспали"
Для создания новых нейропротективных
тельные, нейростимулирующие и противострес"
препаратов необходимо полное понимание мо"
совые препараты, созданные на основе природ"
лекулярно"генетических и клеточных измене"
ных регуляторных пептидов. Такие лекарствен"
ний в мозге, которые происходят после ишеми"
ные препараты не имеют побочных эффектов и
ческого повреждения. На сегодняшний день по"
безопасны в применении. Уже имеющиеся на
казано, что в ответе на патологическое воздей"
сегодня пептидные лекарства оказывают значи"
ствие участвуют не только кодирующие мРНК,
тельное воздействие на работу мозга. Однако
но и различные типы некодирующих РНК
механизмы такого воздействия недостаточно
(нкРНК), в т.ч. микроРНК и длинные некоди"
изучены, что затрудняет разработку новых еще
рующие РНК (днРНК). Так, микроРНК, взаи"
более эффективных препаратов со специализи"
модействуя с сайтами"мишенями на мРНК,
рованным действием на функции мозга.
приводит к деградации мРНК или репрессии ее
Одной из наиболее распространенных при"
трансляции [11]. В последнее время активно
чин нарушений функций мозга является ише"
развивается представление о том, что днРНК
мический инсульт. Он возникает вследствие
могут взаимодействовать с микроРНК и ниве"
постоянного или временного снижения мозго"
лировать их активность [12]. Такие функции
вого кровотока, в большинстве случаев вызван"
приписывают в первую очередь циклическим
ного окклюзией мозговых артерий тромбом или
РНК (циклоРНК) - новому и активно изучае"
эмболом [4-6]. Ишемический инсульт приводит
мому в настоящее время типу РНК [13, 14]. Осо"
к гибели нейронов и клеток глии, сопровождаю"
бенности функционирования нкРНК под
щейся массированным воспалением. Таким об"
действием пептидных препаратов изучены не"
разом, исключительное значение приобретает
достаточно. Однако для дальнейшего совершен"
развитие новых стратегий нейропротекции и
ствования стратегии в разработке новых нейро"
восстановления нервных тканей после ишеми"
активных пептидов самое пристальное внима"
ческого инсульта, направленных на дальней"
ние начинает уделяться изучению роли нкРНК
шую разработку препаратов нейропротективно"
в формировании эффектов пептидных препара"
го действия.
тов. В последних работах показано, что мРНК,
Большие надежды при изучении особеннос"
циклоРНК и микроРНК, а также их взаимодей"
тей действия пептидных препаратов можно воз"
ствия играют важную роль в механизмах по"
лагать на исследования с помощью недавно по"
вреждения и регенерации при патологическом
явившихся транскриптомных подходов, в т.ч.
воздействии [15, 16].
основанных на полногеномном секвенирова"
В настоящем обзоре представлены результа"
нии клеточных РНК. Анализ транскриптома
ты анализа изменений работы генов при разви"
позволяет не только выявлять отдельные гены
тии ишемического повреждения, изучения
или РНК, которые специфичны для того или
транскриптомного профиля клеток мозга под
иного физиологического воздействия, но и пре"
действием пептидов в условиях эксперимен"
доставляет возможность изучать транскрипци"
тальной ишемии мозга, а также сформулирова"
онную реакцию всего генома единовременно, а
ны основные принципы механизмов пептидной
также обнаруживать сигнальные пути и биоло"
регуляции при ишемическом повреждении.
гические процессы, активность которых может
быть связана с природой индуцируемого физио"
логического ответа клеток, органов и тканей.
ПЕПТИДНАЯ РЕГУЛЯЦИЯ ПРИ ИШЕМИИ
Применение транскриптомного анализа позво"
лило существенно детализировать изменения
Церебральная ишемия возникает в результа"
работы генов при развитии ишемического по"
те острого снижения мозгового кровотока, огра"
вреждения в мозге крыс в условиях транзиторной
ничения поступления в нервную ткань кислоро"
окклюзии средней мозговой артерии (transient
да и глюкозы, приводящего к истощению энер"
middle cerebral artery occlusion model, tMCAO)
гетических запасов, и каскада биохимических
[7], а также исследовать механизмы нейропро"
изменений в тканях мозга. После окклюзии со"
тективного действия синтетических пептидов, в
суда активируется глутамат"кальциевый каскад,
т.ч. успешно применяющихся в качестве лекар"
способствующий притоку ионов Ca2+, образова"
ственных препаратов при инсульте [8, 9]. В ми"
нию внутриклеточных медиаторов (фосфоино"
ровой практике имеются и другие примеры ус"
зитол, диацилглицерин), деполяризации мемб"
пешного использования транскриптомного
раны, накоплению глутамата и дальнейшему
подхода к изучению механизмов действия ряда
притоку Ca2+, приводящему к повреждению
пептидов: PACAP38, N"ацетил"Pro"Gly"Pro (N"
макромолекул и, в конечном итоге, к гибели
AcPGP) и др. [10].
клеток [17, 18]. В регуляции ишемического про"
БИОХИМИЯ том 85 вып. 3 2020
326
ФИЛИППЕНКОВ и др.
цесса также активно участвуют пептиды. Роль и
Пептидным препаратом, успешно зареко"
возможные механизмы протективного действия
мендовавшим себя в клинической практике при
ряда биологически активных пептидов, иссле"
лечении инсульта, является синтетический пеп"
дованные в условиях экспериментальной ише"
тид меланокортинового ряда семакс (Met"Glu"
мии мозга на животных, описаны в обзоре Wu et
His"Phe"Pro"Gly"Pro), N"конец которого содер"
al. [19].
жит фрагмент АКТГ(4-7), а С"конец стабилизи"
Было показано, что после ишемического
рован присоединением пептида Pro"Gly"Pro
повреждения эндогенные пептиды могут при"
(PGP). Семакс обладает ярко выраженным ноо"
нимать участие в регулировании кровотока
тропным, нейропротективным, противовоспа"
[20-24] и в работе гематоэнцефалического барь"
лительным и иммуномодулирующим эффектом
ера [25, 26], препятствовать развитию отека [27,
[41-43]. При действии семакса было показано
28], оказывать противовоспалительное иммуно"
существенное снижение размеров ишемическо"
модулирующее действие [29-31], уменьшать
го повреждения в условиях модели фотоиндуци"
повреждение нейронов и образование зоны ин"
рованного тромбоза у крыс [44].
фаркта [32]. Из ранних работ было известно, что
Все большее значение приобретают новые
к числу пептидов, активно функционирующих в
пептидные препараты, относящиеся к классу
условиях церебральной ишемии и обладающих
глипролинов. Они содержат в своем составе ди"
множеством эффектов при ишемии, относится
пептид Gly"Pro, имеют повышенную метаболи"
брадикинин. Показано, что при ишемическом
ческую стабильность, участвуют в различных
инсульте брадикинин оказывает антиоксидант"
биологических процессах, включая провоспали"
ное, противовоспалительное, антиапоптотичес"
тельную хемоаттракцию нейтрофилов при забо"
кое, антиэксайтотоксическое действие, подав"
леваниях легких, воспалительных заболеваниях
ляет митохондриально"опосредованный кле"
кишечника или при ишемическом инсульте
точный апоптоз, способствует выживанию кле"
[45]. Одним из наиболее простых глипролинов
ток глии в ядре и пенумбре [21-24, 33, 34].
является пептид PGP. В ряде работ было показа"
В последние годы активно разрабатываются
но, что пептид PGP оказывает противоязвенное
препараты на основе пептидов меланокортино"
и противовоспалительное действие [46, 47]. На
вого ряда. Меланокортины - это большое се"
основе данного пептида получен ряд синтети"
мейство нейропептидов, образованных из об"
ческих производных, которые проявляют актив"
щего предшественника - молекулы проопиоме"
ность в отношении широкого спектра патологи"
ланокортина, включающее в себя группу мела"
ческих и стрессовых состояний [48-51].
ноцитстимулирующих гормонов (α", β", γ"
Таким образом, регуляторные пептиды и ле"
MSH), адренокортикотропный гормон (АКТГ)
карственные препараты, созданные на их осно"
и некоторые фрагменты, содержащие, по край"
ве, являются полифункциональными, способ"
ней мере, аминокислотную последовательность
ными влиять сразу на несколько физиологичес"
His"Phe"Arg"Trp. Меланокортины принимают
ких функций, однако многие аспекты молеку"
активное участие в балансе дофаминергичес"
лярных механизмов их действия остаются неиз"
кой, холинергической и опиоидной систем.
вестными и требуют дальнейших исследований.
Кроме того, они проявляют противовоспали"
Можно полагать, что значительный вклад в изу"
тельные и иммуномодулирующие свойства, ока"
чение молекулярных механизмов действия пеп"
зывают влияние на процессы обучения и памя"
тидных препаратов внесет транскриптомный
ти, эмоциональное состояние, рост и регенера"
анализ.
цию нервов, пищевое и половое поведение и ряд
других функций организма [35, 36]. В моделях
ишемии и воспаления головного мозга сильные
ТРАНСКРИПТОМНЫЕ ИССЛЕДОВАНИЯ
противовоспалительные, нейрогенные и ней"
ПОВРЕЖДАЮЩЕГО ДЕЙСТВИЯ ИШЕМИИ
ропротективные эффекты проявляет α"MSH
[37-39]. α"MSH индуцирует активацию проте"
Острое снижение мозгового кровотока, вы"
инкиназы A, транскрипционного фактора,
зывающее каскад патофизиологических и био"
опосредующего связь с cAMP (CREB), подавля"
химических изменений, сопровождается немед"
ет активность ядерного фактора активирован"
ленными реакциями генома в клетках мозга
ных T"клеток (NFAT) и ядерного фактора κB
[18]. Реперфузия в сочетании с ишемическим
(NF"κB), снижает уровень провоспалительных
повреждением также приводит к многочислен"
цитокинов TNF"α, IL"1β, IL"6 и активность
ным нарушениям регуляции мРНК, участвую"
каспазы 3, а также индуцирует экспрессию ин"
щих в повреждении [52-54]. Технологии с ис"
терлейкина IL"10 и апоптотических белков Bcl"2
пользованием биочипов и высокопроизводи"
и Bcl"xL [39, 40].
тельного секвенирования РНК (RNA"seq) пре"
БИОХИМИЯ том 85 вып. 3 2020
ПЕПТИДНАЯ РЕГУЛЯЦИЯ МОЗГА
327
доставили возможность изучения реакции всего
(permanent middle cerebral artery occlusion model,
генома на подобные повреждения [7, 52, 53].
pMCAO model), полипептид, активирующий
В условиях tMCAO c помощью RNA"seq на"
аденилатциклазу гипофиза (PACAP), и продукт
ми были обнаружены сотни генов, уровень
его деградации PACAP38 существенно модули"
экспрессии которых изменился в подкорковых
ровали экспрессию генов передачи нервного
структурах мозга крыс, содержащих зону некро"
импульса (Gabra6), иммунного ответа (Crtam), а
за и пенумбру. Более четырехсот генов изменили
также генов семейства интерлейкинов. Автора"
экспрессию через 4,5 ч и в 4 раза больше - через
ми было сделано предположение, что PACAP
24 ч после tMCAO [7]. Сравнение дифференци"
активирует механизм защиты нейронов на ран"
ально экспрессированных генов, выявленных в
ней стадии ишемического повреждения [10].
двух временных точках, позволило нам изучить
Применение транскриптомного анализа поз"
динамику изменений содержания транскриптов
волило нам в условиях модели pMCAO сущест"
в процессе развития повреждения. В подкорко"
венно детализировать механизмы действия пеп"
вых структурах мозга крыс в условиях ише"
тидного препарата семакс и его С"концевого
мии/реперфузии была обнаружена активация
пептида PGP. Анализ экспрессии более 22 000 ге"
огромного числа генов, участвующих в воспале"
нов с использованием микрочипов Illumina
нии, иммунном ответе, апоптозе, в ответе на
RatRef"12 Expression BeadChip позволил нам вы"
стресс, в функционировании рибосом, реплика"
явить изменение содержания транскриптов не"
ции ДНК и других процессах (Hspa1a, Hspb1,
скольких десятков генов, определяющих работу
Lrg1, Jun, Socs3, Cish, Cd14, Cd63, Cd74, Ccl6, Ccl9,
важнейших метаболических систем клеток моз"
Nfkb2 и др.) [7]. Наряду с этим было показано
га, связанных с функционированием семакса в
массированное угнетение генов, обеспечиваю"
условиях pMCAO [8, 9]. В первые часы после ок"
щих работу нейротрансмиттерных систем
клюзии под действием семакса было обнаруже"
(Chrm1, Chrm4, Cplx2, Drd2, Gabra5, Gria3, Grm3,
но существенное увеличение экспрессии генов
Htr6, Neurod6, Ntsr2, Drd1, Grm5 и др.), были вы"
транскрипционных факторов (Fos, Junb, Egr2,
явлены десятки сигнальных путей, с функцио"
Egr4), белковые продукты которых запускают
нированием которых связан ответ клеток мозга
сигнальные пути, корректирующие деструктив"
на повреждение [7]. Таким образом, анализ
ные процессы при ишемии. Семакс оказал поло"
транскриптома дает новое понимание тран"
жительное воздействие на экспрессию генов
скриптомных функциональных изменений ге"
факторов роста, участвующих в трофических и
нома в тканях мозга в ответ на ишемическое
протективных процессах (Bdnf, Nov), в васкуля"
повреждение.
ризации поврежденных тканей (Cyr61, Atf3, Klf4,
Adamts1, Nfil3, Ptgs2, Cox2 и др.). Введение семак"
са оказало существенное влияние на экспрессию
РОЛЬ ТРАНСКРИПТОМА В ПЕПТИДНОЙ
генов, ассоциированных с процессами иммун"
РЕГУЛЯЦИИ ПРИ ИШЕМИИ МОЗГА
ного ответа. Спустя 24 ч после окклюзии было
выявлено увеличение экспрессии генов, кодиру"
Анализ транскриптома может являться од"
ющих иммуноглобулины, белки комплекса гис"
ним из наиболее эффективных подходов в ис"
тосовместимости (RT1$Ba, RT1$A3, RT1$A1), а
следовании механизмов действия регуляторных
также белки, участвующие в регуляции синтеза
пептидов и препаратов на их основе в норме и
антител и миграции иммунных клеток [55, 56].
при различных физиологических и патологи"
Нами также был проведен полногеномный ана"
ческих состояниях. Основанием для этого слу"
лиз изменений транскриптома мозга крыс в ус"
жит представление о том, что при взаимодей"
ловиях pMCAO под воздействием пептида PGP
ствии пептида с рецепторами на клеточной
[55]. Поскольку пептид PGP является одним из
мембране сигнал передается по системе сиг"
преобладающих производных семакса в плазме
нальный путей (сAMP, PI3K, ERK и др.) в ядро
крови и тканях мозга животных [57], то можно
клетки. Далее происходит активация тран"
было ожидать, что наблюдаемые эффекты се"
скрипционных факторов (Fos, Jun, Creb и др.) и
макса и PGP будут схожи. Однако влияние PGP
инициация транскрипционного ответа клетки,
на экспрессию генов факторов роста лишь час"
связанного с эффектом пептида. Транскриптом"
тично совпадало с действием семакса [58]. Было
ные исследования действия регуляторных пеп"
показано увеличение экспрессии генов, ответ"
тидов в значительной степени подтвердили эту
ственных за процессы пролиферации, диффе"
гипотезу. Так, Hori et al. показали, что в услови"
ренцировки, миграции, выживаемости и гибели
ях модели экспериментальной ишемии мозга,
клеток (Bdnf, Nos3, Nts, Ttr и др.) [8, 56]. Однако
вызванной электрокоагуляционной окклюзией
под действием PGP, в отличие от семакса, была
дистального участка средней мозговой артерии
существенно снижена экспрессия генов, ассо"
БИОХИМИЯ том 85 вып. 3 2020
328
ФИЛИППЕНКОВ и др.
циированных с функцией иммунного ответа
регуляторов в механизмах повреждения и ней"
(Adora3, C1qa, C1qc, C2, Cd74, Cxcl1, Cxcl11,
ропротекции [63]. В недавних исследованиях
Cxcl13, Cxcl9 и др.) [8, 9, 55]. В число белков, ко"
были выявлены микроРНК, вовлеченные в пост"
дируемых генами, экспрессия которых умень"
ишемическое повреждение нейронов и тромбоз
шилась под действием PGP, вошли цитокины,
[64, 65], а также показано, что мРНК генов
транспортные белки, факторы транскрипции и
HMGB1, YWHAZ, PIK3R1, STAT3, MAPK1, CBX5,
трансмембранные рецепторы. Сравнительный
CAPZB, THBS1, TNFRSF10B и RCOR1, связан"
анализ действия PGP и семакса в условиях
ных с воспалением, свертыванием крови и акти"
pMCAO показал специфичность воздействия
вацией тромбоцитов, являются мишенями для
этих пептидов на транскриптом при ишемии.
микроРНК при инсульте [66]. Несколько обзо"
Таким образом, применение транскриптомного
ров посвящено использованию микроРНК в ка"
анализа позволяет выявить большее количество
честве диагностических и прогностических мар"
генов, которые связаны с протективным эффек"
керов, а также возможных терапевтических
том пептидов в условиях ишемии.
агентов при инсульте [64-68].
В одной из последних работ Herzog et al. изу"
чили роль стероидных гормонов 17β"эстрадиола
РЕГУЛЯТОРНЫЕ нкРНК -
(E2) и прогестерона (P) в мозге в качестве регу"
НОВЫЕ МИШЕНИ В МЕХАНИЗМАХ
ляторных факторов для miR"223"3p, miR"200c"3p,
ПОВРЕЖДЕНИЯ И НЕЙРОПРОТЕКЦИИ
miR"375"3p, miR"199"3p и miR"214"3p и их це"
левых генов при ишемии в условиях модели
В настоящее время показано, что в ответе на
tMCAO [69]. Было показано, что упомянутые
патологическое и стрессовое воздействия участ"
микроРНК демонстрировали увеличение уров"
вуют не только мРНК, но и различные типы
ня экспрессии через 12 или 72 ч после воздей"
нкРНК, которые обладают значительным регу"
ствия tMCAO. При этом Е2 и Р избирательно
ляторным потенциалом. Большое внимание ис"
подавляли miR"223 и miR"214, но дополнитель"
следователей уделяется сегодня изучению осо"
но повышали содержание miR"375. Интересно,
бенностей функционирования микроРНК и
что экспрессия генов Nr2b и Gria2, которые яв"
днРНК. Так, микроРНК представляют собой
ляются мишенями для miR"223, была снижена
молекулы нкРНК длиной 20-22 нуклеотидов.
после воздействия tMCAO, а применение E2 и P
Они действуют путем непосредственного взаи"
отменяло этот эффект. Кроме того, стероидная
модействия с сайтами"мишенями на мРНК, ко"
терапия ингибировала индуцированное tMCAO
торое приводит к деградации мРНК или репрес"
увеличение экспрессии генов Bcl"2 и Rad1, яв"
сии ее трансляции [11]. В настоящее время ак"
ляющихся мишенями для miR"375. Таким обра"
тивно развивается концепция, связанная с ни"
зом, была показана роль E2 и P в качестве кос"
велированием действия микроРНК с помощью
венных микроРНК"опосредованных регулято"
днРНК, выполняющих функции конкурентных
ров трансляции проапоптотических и провоспа"
эндогенных РНК (кэРНК). кэРНК конкуриру"
лительных генов. В результате функционирова"
ют с мРНК за связывание с микроРНК и ниве"
ние E2 и P приводило к ослаблению ишемичес"
лируют действие последних на транскрипцион"
кого повреждения ткани [69].
ном и посттранскрипционном уровнях регуля"
Bai et al. показали, что циклоРНК гена мем"
ции экспрессии генов [12]. В ряде последних ра"
браносвязанной гуанилаткиназы Dlgap4
бот отмечается, что эффективные кэРНК долж"
(circDLGAP4) функционирует как «губка» для
ны иметь множество сайтов связывания мик"
микроРНК, связывая их на себе и удаляя тем са"
роРНК, а также высокий уровень экспрессии
мым из клеточного пула. Ее нивелирующая ак"
или повышенную стабильность [12, 14].
тивность для miR"143 приводит к ингибирова"
На сегодняшний день имеются данные о
нию экспрессии гомологов E6"AP C"концевого
том, что значительная часть днРНК существует
домена E3 убиквитинлигазы 1 в условиях цереб"
в виде циклической формы [59-61]. ЦиклоРНК
ральной ишемии
[16]. При этом уровень
не подвергаются действию экзонуклеаз и вслед"
circDLGAP4 был значительно снижен даже в
ствие своей повышенной устойчивости могут
плазме пациентов с острым ишемическим ин"
более эффективно играть роль кэРНК [13, 14].
сультом, а также в условиях модели tMCAO в
Большинство циклоРНК человека и грызунов
мозге у мышей. Было показано, что увеличение
обладают мозгоспецифической экспрессией
экспрессии circDLGAP4 значительно ослабляло
[13, 61, 62], что может указывать на их особые
неврологический дефицит, уменьшало размер
функции в мозге.
ядра инфаркта и повреждение гематоэнцефали"
В последние годы активно изучаются осо"
ческого барьера в модели инсульта у мышей. В
бенности функционирования нкРНК в качестве
работе Han et al. (2018) также сообщается, что в
БИОХИМИЯ том 85 вып. 3 2020
ПЕПТИДНАЯ РЕГУЛЯЦИЯ МОЗГА
329
мозге мышей в условиях модели tMCAO, в клет"
G"белком (GPCR), составляют самое большое
ках глиобластомы человека A172 в условиях кис"
семейство рецепторов клеточной поверхности,
лородно"глюкозной депривации-реоксигена"
которые опосредуют многочисленные пути пе"
ции (OGD"R) и в крови пациентов с ишемичес"
редачи сигналов в клетках, участвуют в разнооб"
ким инсультом повышена экспрессия цик"
разных физиологических процессах и являются
лоРНК гена убиквитинлигазы, содержащей
мишенями, на которые нацелены лекарствен"
HECT"домен, Hectd1 (circHECTD1) [70]. Было
ные средства [73, 74]. Молекулярный механизм
показано, что circHECTD1 участвует в регуля"
передачи сигналов семейства рецепторов GPCR
ции регенеративных механизмов клеток мозга
изложен в ряде обзоров [75-77].
при ишемии [63, 68].
Недавние исследования показали, что меха"
низмы функционирования пептидов могут не
ограничиваться ортостерическим взаимодей"
РОЛЬ нкРНК В ПЕПТИДНОЙ
ствием с рецепторами [78, 79]. Обнаружено, что
РЕГУЛЯЦИИ МОЗГА
один и тот же регуляторный пептид может иметь
собственные высокоспецифичные места связы"
Некодирующие РНК могут также влиять на
вания на поверхности клетки, а также способен
функционирование пептидов. Так, Shang et al.
влиять на работу сразу нескольких рецепторных
исследовали потенциальное участие микроРНК
групп, относящихся к различным системам кле"
в эффекте инсулино"глюкагоноподобного пеп"
точной сигнализации [80, 81]. Эксперименталь"
тида"1 (GLP"1) на глюкозостимулированную
ные данные по специфическому связыванию
секрецию [71]. Было показано, что GLP"1 уве"
пептидов позволили выдвинуть предположение
личивает экспрессию miR"132 и miR"212 через
об аллостерическом действии пептидов на раз"
протеинкиназа A/cAMP"зависимый путь в
личные рецепторные системы. Аллостерическое
панкреатических β"клетках поджелудочной же"
связывание приводит к изменению влияния эн"
лезы [71]. В работе Madadi et al. (2019) описана
догенных ортостерических лигандов на рецеп"
роль микроРНК в ингибировании трансляции
тор и специфическим ответам клеток [82]. Это
мРНК, кодирующих ключевые белки путей
предположение объясняет полифункциональ"
транспорта β"амилоида из клетки, что способ"
ность пептидов, а также специфичность и «мяг"
ствует его накоплению при болезни Альцгейме"
кость» их действия.
ра. Так, miR"34a и miR"29b ингибируют по
Чтобы объяснить множественное влияние
меньшей мере три пути транспорта β"амилоида
пептидов на межмолекулярные процессы, была
и могут быть в перспективе использованы для
предложена гипотеза, согласно которой в осно"
лечения болезни Альцгеймера [72].
ве действия регуляторных пептидов лежит си"
Несмотря на то, что днРНК, в первую оче"
нактонный механизм [82]. Суть механизма зак"
редь циклоРНК, играют критическую роль в
лючается в том, что помимо самого пептида
развитии и прогрессировании ишемического
можно выделить функциональное ядро, предс"
инсульта [15, 16], особенности их функциони"
тавленное основными продуктами его метабо"
рования под действием пептидных препаратов
лизма. Эти вторичные пептиды могут иметь
остаются неясными. Однако в последнее время
свои собственные сайты связывания и вместе с
становится очевидным, что анализ системы вза"
родительским пептидом составлять единый
имодействий циклоРНК-микроРНК-мРНК
комплекс биорегуляторов, действующих в опре"
является важной составляющей для детального
деленной последовательности и во взаимодей"
изучения механизмов повреждения и регенера"
ствии, - синактон. В частности, примером си"
ции при патологическом или стрессовом состо"
нактонного механизма является действие се"
янии и действии терапевтических средств, в т.ч.
макса. В общем пуле его метаболитов, который
пептидной природы.
включает продукты расщепления родительской
молекулы, можно выделить функциональное
ядро, представленное пептидами HFPGP и PGP -
СОВРЕМЕННОЕ ПРЕДСТАВЛЕНИЕ
основными продуктами его метаболизма. Эф"
О МЕХАНИЗМАХ ДЕЙСТВИЯ
фект пептидов, которые действуют в синактоне,
ПЕПТИДОВ В МОЗГЕ
значительно расширяет регуляторный потенци"
ал исходной молекулы. В результате, помимо
Механизмы действия регуляторных пепти"
ортостерического связывания, наблюдается ал"
дов обусловлены, по"видимому, их специфичес"
лостерическое взаимодействие с различными
кими лиганд"рецепторными взаимодействиями
типами рецепторов, что указывает на множест"
на плазматических мембранах клеток"мишеней.
венный (плейотропный) характер действия пеп"
Нейропептидные рецепторы, связанные с
тидов.
3 БИОХИМИЯ том 85 вып. 3 2020
330
ФИЛИППЕНКОВ и др.
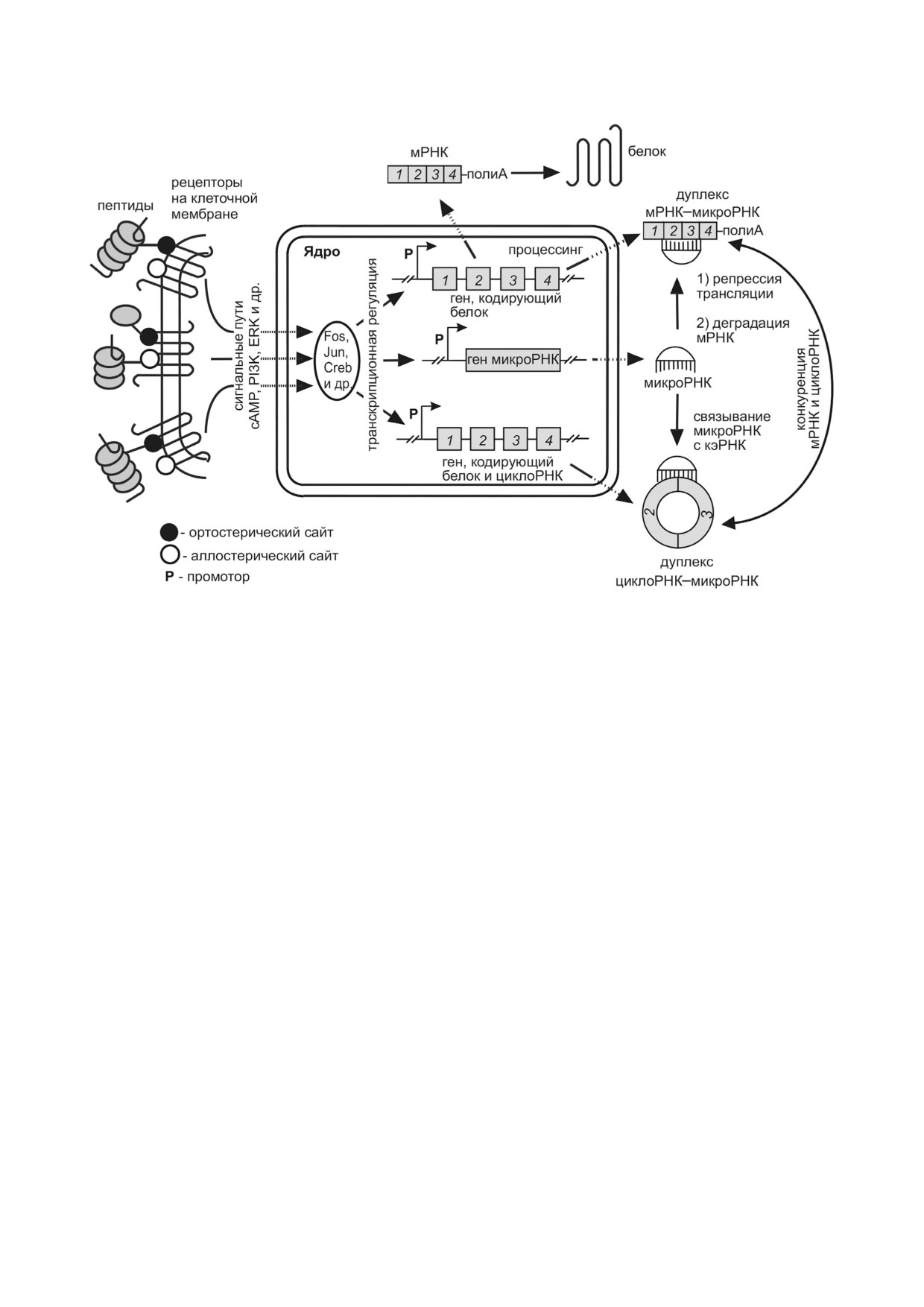
Модель пептидной регуляции с участием некодирующих РНК в мозге. Экзоны показаны пронумерованными блоками,
интроны - линиями, соединяющими экзоны
С учетом вышеизложенных данных мы пред"
человека. В борьбе с ишемией большие надеж"
лагаем модель пептидной регуляции в мозге с
ды возлагаются на новые препараты пептидной
участием нкРНК на основе известных механиз"
природы, которые обладают широким спект"
мов их функционирования (рисунок). При взаи"
ром действия и минимальным побочным воз"
модействии пептида с рецептором на мембране
действием на организм. Обзор эксперимен"
сигнал передается в ядро клетки по системе сиг"
тальных данных свидетельствует о том, что
нальных путей. Активация транскрипционных
большое число пептидов обладает нейропро"
факторов приводит к транскрипции генов, на"
тективными полифункциональными свойства"
правляющих синтез мРНК, микроРНК и цик"
ми, однако многие аспекты молекулярных ме"
лоРНК. Численность активных мРНК, способ"
ханизмов их действия, а также метаболические
ных обеспечивать синтез белка, регулируется
звенья и возможные мишени зачастую остают"
взаимодействием кэРНК циклической природы
ся неизвестными и требуют дальнейших иссле"
с микроРНК и удалением последних из клеточ"
дований. В формировании физиологических
ного пула. Таким образом, действие пептидов на
ответов пептидов существенное значение име"
транскриптом может быть обусловлено систе"
ет регуляция на уровне транскриптома. Важ"
мой взаимодействий циклоРНК-микроРНК-
ную роль в регулировании внутриклеточных
мРНК, которая, в конечном итоге, может при"
процессов помимо мРНК играют нкРНК, сре"
водить клетки мозга в иное функциональное
ди которых особый интерес представляют мик"
состояние. Изучение данной системы взаимо"
роРНК и циклоРНК. В настоящее время роль
действий позволит сформулировать основные
нкРНК в пептидной регуляции в мозге остает"
принципы пептидной регуляции и на основе
ся недостаточно изученной. Однако становит"
этого определить новые подходы для дальней"
ся очевидным, что изучение механизмов
шей разработки препаратов направленного ней"
действия терапевтических средств, в т.ч. пеп"
ропротективного действия.
тидной природы, а также определение страте"
гии в достижении нейропротективного эффек"
В современном мире церебральная ишемия
та препаратов невозможно без учета вклада
является одной из главных угроз для здоровья нкРНК.
БИОХИМИЯ том 85 вып. 3 2020
ПЕПТИДНАЯ РЕГУЛЯЦИЯ МОЗГА
331
Финансирование. Работа выполнена при под"
Соблюдение этических норм. Настоящая
держке Российского научного фонда (проект
статья не содержит описания выполненных ав"
№ 19"14"00268).
торами исследований с участием людей и ис"
Конфликт интересов. Авторы заявляют об от"
пользованием животных в качестве объектов.
сутствии конфликта интересов.
СПИСОК ЛИТЕРАТУРЫ
1.
Ашмарин И.П., Ляпина Л.A., Пасторова В.Е. (1996)
dance, Mol. Cell, 54, 766-776, doi: 10.1016/j.molcel.
Модуляция гемостатических реакций in vitro и in vivo
2014.03.045.
представителями семейств регуляторных пептидов,
15.
Saugstad, J.A. (2015) Non"coding RNAs in stroke and
Вестник Российской академии медицинских наук, 6,
neuroprotection, Front. Neurol., 6, 50, doi: 10.3389/
50-57.
fneur.2015.00050.
2.
Snyder, S. (1980) Brain peptides as neurotransmitters,
16.
Bai, Y., Zhang, Y., Han, B., Yang, L., Chen, X., Huang, R.,
Science, 209, 976-983, doi: 10.1126/science.6157191.
Wu, F., Chao, J., Liu, P., Hu, G., Zhang, J.H., and Yao, H.
3.
Hoyer, D., and Bartfai, T. (2012) Neuropeptides and neu"
(2018) Circular RNA DLGAP4 ameliorates ischemic
ropeptide receptors: drug targets, and peptide and non"
stroke outcomes by targeting miR"143 to regulate endothe"
peptide ligands: a tribute to Prof. Dieter Seebach, Chem.
lial"mesenchymal transition associated with blood"brain
Biodivers., 9, 2367-2387, doi: 10.1002/cbdv.201200288.
barrier integrity, J. Neurosci., 38, 32-50, doi: 10.1523/
4.
Kalaria, R.N., and Ballard, C. (2001) Stroke and cogni"
JNEUROSCI.1348"17.2017.
tion, Curr. Atheroscler. Rep., 3, 334-339, doi: 10.1007/
17.
Pellegrini"Giampietro, D.E., Bennett, M.V., and Zukin, R.S.
s11883"001"0028"5.
(1992) Are Ca2+"permeable kainate/AMPA receptors more
5.
Seshadri, S., Beiser, A., Kelly"Hayes, M., Kase, C.S., Au, R.,
abundant in immature brain? Neurosci. Lett., 144, 65-69,
Kannel, W.B., and Wolf, P.A. (2006) The lifetime risk of
doi: 10.1016/0304"3940(92)90717"l.
stroke: estimates from the Framingham Study, Stroke, 37,
18.
Гусев Е.И., Скворцова В.И. (2001) Ишемия головного
345-350, doi: 10.1161/01.STR.0000199613.38911.b2.
мозга, Медицина, Москва.
6.
Mukherjee, D., and Patil, C.G. (2011) Epidemiology and
19.
Wu, D., Wang, J., Wang, H., Ji, A., and Li, Y. (2017)
the global burden of stroke, World Neurosurg., 76,
Protective roles of bioactive peptides during ischemia"
S85-S90, doi: 10.1016/j.wneu.2011.07.023.
reperfusion injury: from bench to bedside. Life Sci., 180,
7.
Dergunova, L.V., Filippenkov, I.B., Stavchansky, V.V.,
83-92, doi: 10.1016/j.lfs.2017.05.014.
Denisova, A.E., Yuzhakov, V.V., Mozerov, S.A., Gubsky, L.V.,
20.
Miyazaki, T., Otani, K., Chiba, A., Nishimura, H.,
and Limborska, S.A. (2018) Genome"wide transcriptome
Tokudome, T., Takano"Watanabe, H., Matsuo, A.,
analysis using RNA"Seq reveals a large number of differen"
Ishikawa, H., Shimamoto, K., Fukui, H., Kanai, Y.,
tially expressed genes in a transient MCAO rat model,
Yasoda, A., Ogata, S., Nishimura, K., Minamino, N., and
BMC Genomics, 19, 655, doi: 10.1186/s12864"018"5039"5.
Mochizuki, N. (2018) A new secretory peptide of natri"
8.
Medvedeva, E.V., Dmitrieva, V.G., Povarova, O.V.,
uretic peptide family, osteocrin, suppresses the progression
Limborska, S.A., Skvortsova, V.I., Myasoedov, N.F., and
of congestive heart failure after myocardial infarction, Circ.
Dergunova, L.V. (2014) The peptide semax affects the
Res., 122, 742-751, doi: 10.1161/CIRCRESAHA.117.
expression of genes related to the immune and vascular sys"
312624.
tems in rat brain focal ischemia: genome"wide transcrip"
21.
Zhang, H.Y., Li, J., Guo, N., and Zhang, B.Y. (2019)
tional analysis, BMC Genomics, 15, 228, doi: 10.1186/
Brain functions and unusual β"amyloid accumulation in
1471"2164"15"228.
the hypertensive white matter lesions of rats, J. Biol. Regul.
9.
Medvedeva, E.V., Dmitrieva, V.G., Limborska, S.A.,
Homeost. Agents, 33, 1073-1084.
Myasoedov, N.F., and Dergunova, L.V. (2017) Semax, an
22.
Martins, A.H., Zayas"Santiago, A., Ferrer"Acosta, Y.,
analog of ACTH(4"7), regulates expression of immune response
Martinez"Jimenez, S.M., Zueva, L., Diaz"Garcia, A., and
genes during ischemic brain injury in rats, Mol. Genet.
Inyushin, M. (2019) Accumulation of amyloid beta (Aβ)
Genomics, 292, 635-653, doi: 10.1007/s00438"017"1297"1.
peptide on blood vessel walls in the damaged brain after
10.
Hori, M., Nakamachi, T., Shibato, J., Rakwal, R., Shioda, S.,
transient middle cerebral artery occlusion, Biomolecules, 9,
and Numazawa, S. (2015) Unraveling the specific ischemic
350, doi: 10.3390/biom9080350.
core and penumbra transcriptome in the permanent middle
23.
Guo, S., Barringer, F., Zois, N.E., Goetze, J.P., and
cerebral artery occlusion mouse model brain treated with
Ashina, M. (2014) Natriuretic peptides and cerebral hemo"
the neuropeptide PACAP38, Microarrays,
4,
2-24,
dynamics, Regul. Pept., 192-193, 15-23, doi: 10.1016/
doi: 10.3390/microarrays4010002.
j.regpep.2014.07.003.
11.
Bartel, D.P. (2009) MicroRNAs: target recognition and
24.
James, M.L., Wang, H., Venkatraman, T., Song, P.,
regulatory
functions,
Cell,
136,
215-233,
Lascola, C.D., and Laskowitz, D.T. (2010) Brain natriuret"
doi: 10.1016/j.cell.2009.01.002.
ic peptide improves long"term functional recovery after
12.
Broderick, J.A., and Zamore, P.D. (2014) Competitive
acute CNS injury in mice, J. Neurotrauma, 27, 217-228,
endogenous RNAs cannot alter microRNA function in
doi: 10.1089/neu.2009.1022.
vivo, Mol. Cell,
54,
711-713, doi:
10.1016/j.mol"
25.
Liu, Z., Liu, Q., Cai, H., Xu, C., Liu, G., and Li, Z. (2011)
cel.2014.05.023.
Calcitonin gene"related peptide prevents blood"brain bar"
13.
Hansen, T.B., Jensen, T.I., Clausen, B.H., Bramsen, J.B.,
rier injury and brain edema induced by focal cerebral
Finsen, B., Damgaard, C.K., and Kjems, J. (2013) Natural
ischemia reperfusion, Regul. Pept.,
171,
19-25,
RNA circles function as efficient microRNA sponges,
doi: 10.1016/j.regpep.2011.05.014.
Nature, 495, 384-388, doi: 10.1038/nature11993.
26.
Huang, Q., Zhong, W., Hu, Z., and Tang, X. (2018)
14.
Denzler, R., Agarwal, V., Stefano, J., Bartel, D.P., and
A review of the role of cav"1 in neuropathology and neural
Stoffel, M. (2014) Assessing the ceRNA hypothesis with
recovery after ischemic stroke, J. Neuroinflammation, 15,
quantitative measurements of miRNA and target abun"
348, doi: 10.1186/s12974"018"1387"y.
БИОХИМИЯ том 85 вып. 3 2020
3*
332
ФИЛИППЕНКОВ и др.
27.
Wang, Y.F., and Parpura, V. (2018) Astroglial modulation
42.
Grivennikov, I.A., Dolotov, O.V, and Gol’dina, I.I. (1999)
of hydromineral balance and cerebral edema, Front. Mol.
Peptide factors in processes of proliferation, differentia"
Neurosci., 11, 204, doi: 10.3389/fnmol.2018.00204.
tion, and extended viability of mammalian nervous system
28.
Jia, S.W., Liu, X.Y., Wang, S.C., and Wang, Y.F. (2016)
cells, Mol. Biol. (Mosk.), 33, 120-126.
Vasopressin hypersecretion"associated brain edema forma"
43.
Kolomin, T., Shadrina, M., Slominsky, P., Limborska, S.,
tion in ischemic stroke: underlying mechanisms, J. Stroke
and Myasoedov, N. (2013) A new generation of drugs: syn"
Cerebrovasc. Dis., 25, 1289-1300, doi: 10.1016/j.jstroke"
thetic peptides based on natural regulatory peptides,
cerebrovasdis.2016.02.002.
Neurosci. Med., 4, 223-252, doi: 10.4236/nm.2013.44035
29.
Waschek, J.A. (2013) VIP and PACAP: neuropeptide mod"
44.
Romanova, G.A., Silachev, D.N., Shakova, F.M.,
ulators of CNS inflammation, injury, and repair, Br. J.
Kvashennikova, Y.N., Viktorov, I.V., Shram, S.I., and
Pharmacol., 169, 512-523, doi: 10.1111/bph.12181.
Myasoedov, N.F. (2006) Neuroprotective and antiamnesic
30.
Lakhan, S.E., Kirchgessner, A., and Hofer, M. (2009)
effects of Semax during experimental ischemic infarction
Inflammatory mechanisms in ischemic stroke: therapeutic
of the cerebral cortex, Bull. Exp. Biol. Med., 142, 663-666,
approaches, J. Transl. Med., 7, 97, doi: 10.1186/1479"
doi: 10.1007/s10517"006"0445"0.
5876"7"97.
45.
Misiura, M., and Miltyk, W. (2019) Proline"containing
31.
La, M., Tailor, A., D’Amico, M., Flower, R.J., and
peptides - new insight and implications: a review,
Perretti, M. (2001) Analysis of the protection afforded by
Biofactors, 45, 857-866, doi: 10.1002/biof.1554.
annexin 1 in ischaemia-reperfusion injury: focus on neu"
46.
Бондаренко Н.С., Шнейдерман А.Н., Гусева А.А.,
trophil recruitment, Eur. J. Pharmacol., 429, 263-278,
Умарова Б.А. (2017) Пептид пролил-глицил-пролин
doi: 10.1016/S0014"2999(01)01325"5.
(PGP) препятствует повышению проницаемости крове"
32.
Wang, M., Wang, J., Liu, Z., Guo, X., Wang, N, Jia, N,
носных сосудов при воспалении, Acta Naturae, 1, 55-59.
Zhang, Y., and Yuan, J. (2018) Effects of intermedin on
47.
Безуглов В.В., Грецкая Н.М., Васильева Т.М., Петру"
autophagy in cerebral ischemia/reperfusion injury,
хина Г.Н., Андреева Л.А., Мясоедов Н.Ф., Макаров В.А.
Neuropeptides, 68, 15-21, doi: 10.1016/j.npep.2017.
(2014) Антиагрегационная активность конъюгатов
10.004.
арахидоновой кислоты и нейротропных пептидов
33.
Ji, B., Cheng, B., Pan, Y., Wang, C., Chen, J., and Bai, B.
проглипрола и cемакса, Экспериментальная и клини$
(2017) Neuroprotection of bradykinin/bradykinin B2
ческая фармакология, 77, 30-32.
receptor system in cerebral ischemia, Biomed.
48.
Мясоедов Н.Ф., Рочев Д.Л., Ляпина Л.А., Оберган Т.Ю.,
Pharmacother., 94, 1057-1063, doi: 10.1016/j.biopha.
Андреева Л.А. (2013) Лейцинсодержащие глипролины
2017.08.042.
(PRO"GLY"PRO"LEU и LEU"PRO"GLY"PRO): учас"
34.
Li, M., Chen, S., Shi, X., Lyu, C., Zhang, Y., Tan, M.,
тие в реакциях гемостаза in vitro и in vivo в условиях на"
Wang, C., Zang, N., Liu, X., Hu, Y., Shen, J., Zhou, L.,
рушений процессов свертывания крови и жирового
and Gu, Y. (2018) Cell permeable HMGB1"binding hep"
обмена у крыс, Доклады Академии наук, 453, 457-460.
tamer peptide ameliorates neurovascular complications
49.
Ляпина Л.А., Мясоедов Н.Ф., Андреева Л.А.,
associated with thrombolytic therapy in rats with transient
Ульянов А.М., Оберган Т.Ю., Шубина Т.А., Пасторова В.Е.
ischemic stroke, J. Neuroinflammation,
15,
237,
(2010) Фибринолитический и гипогликемический эф"
doi: 10.1186/s12974"018"1267"5.
фекты пептида Pro"Gly"Pro"Leu при развитии инсу"
35.
De Wied, D. (1999) Behavioral pharmacology of neu"
линзависимого диабета у крыс, Известия РАН Сер. би$
ropeptides related to melanocortins and the neurohy"
ол., 3, 375-379.
pophyseal hormones, Eur. J. Pharmacol., 375, 1-11,
50.
Ashmarin, I.P., Samonina, G.E., Lyapina, L.A.,
doi: 10.1016/s0014"2999(99)00339"8.
Kamenskii, A.A., Levitskaya, N.G., Grivennikov, I.A.,
36.
Catania, A., Gatti, S., Colombo, G., and Lipton, J.M.
Dolotov, O.V., Andreeva, L.A., and Myasoedov, N.F.
(2004) Targeting melanocortin receptors as a novel strategy
(2005). Natural and hybrid (“chimeric”) stable regulatory
to control inflammation, Pharmacol. Rev., 56, 1-29,
glyproline peptides, Pathophysiology,
11,
179-185,
doi: 10.1124/pr.56.1.1.
doi: 10.1016/j.pathophys.2004.10.001.
37.
Giuliani, D., Ottani, A., Neri, L., Zaffe, D., Grieco, P.,
51.
Filatova, E., Kasian, A., Kolomin, T., Rybalkina, E.,
Jochem, J., Cavallini, G.M., Catania, A., and Guarini, S.
Alieva, A., Andreeva, L., Limborska, S., Myasoedov, N.,
(2017) Multiple beneficial effects of melanocortin MC4
Pavlova, G., Slominsky, P., and Shadrina, M.
(2017)
receptor agonists in experimental neurodegenerative disor"
GABA, Selank, and olanzapine affect the expression of
ders: therapeutic perspectives. Prog. Neurobiol., 148,
genes involved in GABAergic neurotransmission in IMR"
40-56, doi: 10.1016/j.pneurobio.2016.11.004.
32 cells, Front. Pharmacol., 8, 89, doi: 10.3389/fphar.
38.
Lisak, R.P., and Benjamins, J.A. (2017) Melanocortins,
2017.00089.
melanocortin receptors and multiple sclerosis, Brain Sci.,
52.
Ford, G., Xu, Z., Gates, A., Jiang, J., and Ford, B.D.
7, 104, doi: 10.3390/brainsci7080104.
(2006) Expression analysis systematic explorer (EASE)
39.
Mykicki, N., Herrmann, A.M., Schwab, N., Deenen, R.,
analysis reveals differential gene expression in permanent
Sparwasser, T., Limmer, A., Wachsmuth, L., Klotz, L.,
and transient focal stroke rat models, Brain Res., 1071,
Kohrer, K., Faber, C., Wiendl, H., Luger, T.A., Meuth, S.G.,
226-236, doi: 10.1016/j.brainres.2005.11.090.
and Loser, K. (2016) Melanocortin"1 receptor activation is
53.
Wang, C., Liu, M., Pan, Y., Bai, B., and Chen, J. (2017)
neuroprotective in mouse models of neuroinflammatory
Global gene expression profile of cerebral ischemia-reper"
disease, Sci. Transl. Med., 8, 362ra146, doi: 10.1126/sci"
fusion injury in rat MCAO model, Oncotarget,
8,
translmed.aaf8732.
74607-74622, doi: 10.18632/oncotarget.20253.
40.
Giuliani, D., Minutoli, L., Ottani, A., Spaccapelo, L.,
54.
DeGracia, D.J. (2017) Regulation of mRNA following
Bitto, A., Galantucci, M., Altavilla, D., Squadrito, F., and
brain ischemia and reperfusion, Wiley Interdiscip. Rev.
Guarini, S. (2012) Melanocortins as potential therapeutic
RNA, 8, 4, doi: 10.1002/wrna.1415.
agents in severe hypoxic conditions, Front. Neuroendocrinol.,
55.
Медведева Е.В., Дмитриева В.Г., Поварова О.В., Лим"
33, 179-193, doi: 10.1016/j.yfrne.2012.04.001.
борская С.А., Скворцова В.И., Мясоедов Н.Ф., Дергу"
41.
De Wied, D. (1997) Neuropeptides in learning and memo"
нова Л.В. (2014) Трипептид Pro"Gly"Pro влияет на тран"
ry processes, Behav. Brain Res., 83, 83-90, doi: 10.1016/
скриптом коры головного мозга крыс в условиях фо"
s0166"4328(97)86050"0.
кальной ишемии, Молекулярная биология, 48, 277-287.
БИОХИМИЯ том 85 вып. 3 2020
ПЕПТИДНАЯ РЕГУЛЯЦИЯ МОЗГА
333
56.
Medvedeva, E.V., Dmitrieva, V.G., Povarova, O.V.,
terone on inflammatory and apoptotic microRNA expres"
Limborska, S.A., Skvortsova, V.I., Myasoedov, N.F., and
sion after ischemia in a rat model, J. Steroid Biochem. Mol.
Dergunova, L.V. (2013) Effect of semax and its C"terminal
Biol., 167, 126-134, doi: 10.1016/j.jsbmb.2016.11.018.
fragment Pro"Gly"Pro on the expression of VEGF family
70.
Han, B., Zhang, Yuan, Zhang, Yanhong, Bai, Y., Chen, X.,
genes and their receptors in experimental focal ischemia of
Huang, R., Wu, F., Leng, S., Chao, J., Zhang, J.H.,
the rat brain, J. Mol. Neurosci.,
49,
328-333,
Hu, G., and Yao, H. (2018) Novel insight into circular
doi: 10.1007/s12031"012"9853"y.
RNA HECTD1 in astrocyte activation via autophagy by
57.
Dolotov, O.V., Zolotarev, I.A., Dorokhova, E.M.,
targeting MIR142"TIPARP: implications for cerebral
Andreeva, L.A., Alfeeva, L.I., Grivennikov, I.A., and
ischemic stroke, Autophagy, 14, 1164-1184, doi: 10.1080/
Miasoedov, N.F. (2004) The binding of Semax, ACTH 4"
15548627.2018.1458173.
10 heptapeptide, to plasma membranes of the rat forebrain
71.
Shang, J., Li, J., Keller, M.P., Hohmeier, H.E., Wang, Y.,
basal nuclei and its biodegradation, Bioorg. Khim., 30,
Feng, Y., Zhou, H.H., Shen, X., Rabaglia, M., Soni, M.,
241-246, doi: 10.1023/b:rubi.0000030127.46845.f0.
Attie, A.D., Newgard, C.B., Thornberry, N.A., Howard, A.D.,
58.
Дмитриева В.Г., Дергунова Л.В., Поварова О.В.,
and Zhou, Y.P. (2015) Induction of miR"132 and miR"212
Скворцова В.И., Лимборская С.А., Мясоедов Н.Ф.
expression by glucagon"like peptide 1 (GLP"1) in rodent
(2008) Действие семакса и его С"концевого трипепти"
and human pancreatic β"cells, Mol. Endocrinol., 29,
да PGP на экспрессию генов факторов роста и их ре"
1243-1253, doi: 10.1210/me.2014"1335.
цепторов в условиях экспериментальной ишемии
72.
Madadi, S., Schwarzenbach, H., Saidijam, M., Mahjub, R.,
мозга крыс, Доклады Академии наук, 422, 258-261.
and Soleimani, M. (2019) Potential microRNA"related
59.
Lasda, E., and Parker, R. (2014) Circular RNAs: diversity
targets in clearance pathways of amyloid"β: novel thera"
of form and function, RNA, 20, 1829-1842, doi: 10.1261/
peutic approach for the treatment of Alzheimer’s disease,
rna.047126.114.
Cell Biosci., 9, 91, doi: 10.1186/s13578"019"0354"3.
60.
Filippenkov, I.B., Kalinichenko, E.O., Limborska, S.A.,
73.
Edward Zhou, X., Melcher, K., and Eric Xu, H. (2019)
and Dergunova, L.V. (2017) Circular RNAs - one of the
Structural biology of G protein"coupled receptor signaling
enigmas of the brain, Neurogenetics,
18,
1-6,
complexes, Protein Sci., 28, 487-501, doi: 10.1002/pro.3526.
doi: 10.1007/s10048"016"0490"4.
74.
Левченко М.Е., Поройков В.В., Канехиса М. (2004)
61.
Filippenkov, I.B., Sudarkina, O.Y., Limborska, S.A., and
Пептидные рецепторы, сопряженные с G"белками, и
Dergunova, L.V. (2015) Circular RNA of the human
их эндогенные лиганды в геноме человека, Биомеди$
sphingomyelin synthase 1 gene: multiple splice variants,
цинская химия, 50, 149-158.
evolutionary conservatism and expression in different tis"
75.
Duc, N.M., Kim, H.R., and Chung, K.Y.
(2015)
sues, RNA Biol., 12, 1030-1042, doi: 10.1080/15476286.
Structural mechanism of G protein activation by G pro"
2015.1076611.
tein"coupled receptor, Eur. J. Pharmacol., 763, 214-222,
62.
Rybak"Wolf, A., Stottmeister, C., Glazar, P., Jens, M.,
doi: 10.1016/j.ejphar.2015.05.016.
Pino, N., Giusti, S., Hanan, M., Behm, M., Bartok, O.,
76.
Culhane, K.J., Liu, Y., Cai, Y., and Yan, E.C. (2015)
Ashwal"Fluss, R., Herzog, M., Schreyer, L., Papavasileiou, P.,
Transmembrane signal transduction by peptide hormones
Ivanov, A., Ohman, M., Refojo, D., Kadener, S., and
via family B G protein"coupled receptors, Front.
Rajewsky, N. (2015) Circular RNAs in the mammalian brain
Pharmacol., 6, 264, doi: 10.3389/fphar.2015.00264.
are highly abundant, conserved, and dynamically expressed,
77.
Lohse, M.J., Maiellaro, I., and Calebiro, D.
(2014)
Mol. Cell, 58, 870-885, doi: 10.1016/j.molcel.2015.03.027.
Kinetics and mechanism of G protein"coupled receptor
63.
Heydari, E., Alishahi, M., Ghaedrahmati, F., Winlow, W.,
activation, Curr. Opin. Cell Biol., 27, 87-93, doi: 10.1016/
Khoshnam, S.E., and Anbiyaiee, A. (2019) The role of
j.ceb.2013.11.009.
non"coding RNAs in neuroprotection and angiogenesis
78.
Vyunova, T.V., Andreeva, L., Shevchenko, K., and
following ischemic stroke, Metab. Brain Dis.,
Myasoedov, N. (2018) Peptide"based anxiolytics: the mol"
doi: 10.1007/s11011"019"00485"2.
ecular aspects of heptapeptide Selank biological activity,
64.
He, W., Chen, S., Chen, X., Li, S., and Chen, W. (2016)
Protein. Pept. Lett.,
25,
914-923, doi:
10.2174/
Bioinformatic analysis of potential microRNAs in ischemic
0929866525666180925144642.
stroke, J. Stroke Cerebrovasc. Dis.,
25,
1753-1759,
79.
Lee, S.M., Hay, D.L., and Pioszak, A.A. (2016) Calcitonin
doi: 10.1016/j.jstrokecerebrovasdis.2016.03.023.
and amylin receptor peptide interaction mechanisms:
65.
Dewdney, B., Trollope, A., Moxon, J., Thomas
insights into peptide"binding modes and allosteric modula"
Manapurathe, D., Biros, E., and Golledge, J.
(2018)
tion of the calcitonin receptor by receptor activity"modify"
Circulating microRNAs as biomarkers for acute ischemic
ing proteins, J. Biol. Chem., 291, 8686-8700, doi: 10.1074/
stroke: a systematic review, J. Stroke Cerebrovasc. Dis., 27,
jbc.M115.713628.
522-530, doi: 10.1016/j.jstrokecerebrovasdis.2017.09.058.
80.
Вьюнова Т.В., Андреева Л.А., Шевченко К.В., Шев"
66.
Eyileten, C., Wicik, Z., De Rosa, S., Mirowska"Guzel, D.,
ченко В.П., Бобров М.Ю., Безуглов В.В., Мясоедов
Soplinska, A., Indolfi, C., Jastrzebska"Kurkowska, I.,
Н.Ф. (2018) Взаимодействие трипептида Pro"Gly"Pro,
Czlonkowska, A., and Postula, M. (2018) MicroRNAs as
меченного по С"концевому остатку пролина, с плазма"
diagnostic and prognostic biomarkers in ischemic stroke -
тическими мембранами мозга крыс, Доклады Академии
a comprehensive review and bioinformatic analysis, Cells,
наук, 419, 136-137, doi: 10.1017/CBO9781107415324.004.
7, 249, doi: 10.3390/cells7120249.
81.
Вьюнова Т.В., Андреева Л.А., Шевченко К.В., Шев"
67.
Khoshnam, S.E., Winlow, W., Farbood, Y., Moghaddam, H.F.,
ченко В.П., Бобров М.Ю., Безуглов В.В., Мясоедов Н.Ф.
and Farzaneh, M. (2017) Emerging roles of microRNAs in
(2014) Особенности специфического связывания пен"
ischemic stroke: as possible therapeutic agents, J. Stroke,
тапептида HFPGP, меченного по C"концевому остат"
19, 166-187, doi: 10.5853/jos.2016.01368.
ку пролина, с плазматическими мембранами нервных
68.
Wang, S.W., Liu, Z., and Shi, Z.S. (2018) Non"coding
клеток головного мозга крысы, Доклады Академии на$
RNA in acute ischemic stroke: mechanisms, biomarkers
ук, 456, 490-493, doi: 10.7868/s0869565214160312.
and therapeutic targets, Cell Transplant., 27, 1763-1777,
82.
Vyunova, T.V., Andreeva, L.A., Shevchenko, K.V., and
doi: 10.1177/0963689718806818.
Myasoedov, N.F. (2017) Synacton and individual activity
69.
Herzog, R., Zendedel, A., Lammerding, L., Beyer, C., and
of synthetic and natural corticotropins, J. Mol. Recognit.,
Slowik, A. (2017) Impact of 17β"estradiol and proges"
30, e2597, doi: 10.1002/jmr.2597.
БИОХИМИЯ том 85 вып. 3 2020
334
ФИЛИППЕНКОВ и др.
NEUROPROTECTIVE EFFECTS OF PEPTIDES IN THE BRAIN:
TRANSCRIPTOME APPROACH
Review
I. B. Filippenkov*, L. V. Dergunova, S. A. Limborska, and N. F. Myasoedov
Institute of Molecular Genetics, Russian Academy of Sciences, 123182 Moscow, Russia; E$mail: Filippenkov@img.ras.ru
Received December 5, 2019
Revised December 5, 2019
Accepted December 11, 2019
The importance of studying the action mechanisms of drugs based on natural regulatory peptides is commonly rec"
ognized. Particular attention is paid to the peptide drugs that contribute to the restoration of brain functions after
acute cerebrovascular accidents (stroke), which for many years continues to be one of the main problems and threats
to human health. However, molecular genetic changes in the brain in response to ischemia, as well as the mechanisms
of protective effects of peptides, have not been sufficiently studied. This limits the use of neuroprotective peptides and
makes it difficult to develop new, more efficient drugs with targeted action on brain functions. Transcriptome analy"
sis is a promising approach for studying the mechanisms of the damaging effects of cerebral ischemia and neuropro"
tective action of peptide drugs. Beside investigating the role of mRNAs in protein synthesis, the development of new
neuroprotection strategies requires studying the involvement of regulatory RNAs in ischemia. Of greatest interest are
microRNAs (miRNAs) and circular RNAs (circRNAs), which are expressed predominantly in the brain. CircRNAs
can interact with miRNAs and diminish their activity, thereby inhibiting miRNA"mediated repression of mRNAs. It
has become apparent that analysis of the circRNA/miRNA/mRNA system is essential for deciphering the mecha"
nisms of brain damage and repair. Here, we present the results of studies on the ischemia"induced changes in the
activity of genes and peptide"mediated alterations in the transcriptome profiles in experimental ischemia and formu"
late the basic principles of peptide regulation in the ischemia"induced damage.
Keywords: transcriptomics, regulatory peptides, brain ischemia, mRNA, non"coding RNA, microRNA, circular RNA
БИОХИМИЯ том 85 вып. 3 2020