БИОХИМИЯ, 2020, том 85, вып. 3, с. 348 - 359
УДК 577.2
РОЛЬ ГЕНОВ LAM В ГИБЕЛИ КЛЕТОК ДРОЖЖЕЙ S. cerevisiae,
ВЫЗВАННОЙ ВЫСОКОЙ КОНЦЕНТРАЦИЕЙ
ПОЛОВОГО ФЕРОМОНА*
© 2020
С.С. Соколов1**, К.В. Галкина1, Е.А. Литвинова2,
Д.А. Кнорре1,3, Ф.Ф. Северин1
1 НИИ физико химической биологии им. А.Н. Белозерского,
Московский государственный университет им. М.В. Ломоносова,
119991 Москва, Россия; электронная почта: sviatoslav.sokolov@gmail.com
2 Московский государственный университет им. М.В. Ломоносова,
биологический факультет, 119991 Москва, Россия
3 Первый Московский государственный медицинский университет имени И. М. Сеченова
Министерства здравоохранения Российской Федерации, 119992 Москва, Россия
Поступила в редакцию 03.11.2019
После доработки 21.01.2020
Принята к публикации 21.01.2020
Белки Lam1-Lam4 осуществляют невезикулярный транспорт стеринов из плазматической мембраны (ПМ)
в эндоплазматический ретикулум, нарушение их работы приводит к увеличению содержания стеринов в
ПМ. У животных гомологи белков Lam отвечают за интернализацию холестерина из плазмы крови. Биоло$
гическая роль дрожжевых белков Lam остается неясной, так как выраженные фенотипы у штаммов с деле$
цией отдельных генов LAM не наблюдаются. Однако известно, что делеция LAM1(YSP1) препятствует регу$
лируемой гибели клеток дрожжей Saccharomyces cerevisiae, вызванной половым феромоном. Мы исследова$
ли, будет ли мутация по гену LAM2 также влиять на гибель клеток в присутствии избытка полового феромо$
на и оценили характер генетических взаимодействий между LAM2 и генами, отвечающими за биосинтез эр$
гостерина. Мы показали, что делеция LAM2 частично предотвращает гибель клеток дрожжей лабораторно$
го штамма W303, вызванную половым феромоном. При этом делеция трех других генов LAM - LAM1, LAM3
и LAM4 - дополнительно не увеличивала этот эффект. Мутация в гене транскрипционного фактора UPC2,
приводящая к избыточному накоплению стеринов в клетке (UPC2 1), приводила к увеличению выживае$
мости клеток в присутствии α$фактора и проявляла аддитивность с делецией LAM2. В то же время в лабо$
раторном штамме BY4741 делеция LAM2 наоборот стимулировала гибель клеток, вызванную феромоном.
Мы обнаружили, что делеция генов биосинтеза эргостерина ERG2 и ERG6 снижала эффект от делеции
LAM2. В штамме Δerg4, в котором делетирован ген терминального этапа пути биосинтеза эргостерина, деле$
ция LAM2 заметно увеличивала долю мертвых клеток и снижала скорость роста суспензионной культуры
клеток этого штамма даже в отсутствие феромона. Мы предполагаем, что отсутствие эффекта от делеции ге$
на LAM2 в штаммах Δerg6 и Δerg2 указывает на неспособность Lam2p транспортировать некоторые интерме$
диаты биосинтеза эргостерина, такие как ланостерин. В совокупности наши данные говорят о том, что роль
белков Lam в клеточной смерти дрожжей, вызванной феромоном, обусловлена их воздействием на стери$
новый состав ПМ.
КЛЮЧЕВЫЕ СЛОВА: гены LAM, дрожжи, клеточная смерть, половой феромон, стерины.
DOI: 10.31857/S0320972520030057
У многоклеточных организмов запрограм$
росом. В случае хорошо изученного модельного
мированная гибель клеток (запрограммирован$
объекта - пекарских дрожжей Saccharomyces
ная клеточная смерть, ЗКС) играет важную роль
cerevisiae - принято различать: СКС (случайную
в индивидуальном развитии, иммунитете и под$
клеточную смерть, accidental cell death), вызван$
держании тканевого гомеостаза [1, 2]. Наличие
ную стрессорными факторами высокой интен$
и физиологическая роль ЗКС у одноклеточных
сивности и сопряженной с потерей барьерной
организмов до сих пор остается открытым воп$
функции плазматической мембраны (ПМ), и
Принятые сокращения: ЗКС - запрограммированная клеточная смерть, КОЕ - колониеобразующая единица;
ПМ - плазматическая мембрана; РКС - регулируемая гибель клеток; ЭР - эндоплазматический ретикулум.
* Первоначально английский вариант рукописи опубликован на сайте «Biochemistry» (Moscow) http://protein.
bio.msu.ru/biokhimiya, в рубрике «Papers in Press», BM19$339, 24.02.2020.
** Адресат для корреспонденции.
348
РОЛЬ ГЕНОВ LAM В СМЕРТИ КЛЕТОК ДРОЖЖЕЙ
349
РКС (регулируемую клеточную смерть) [3]. Под
мейству мембранных стерин$связывающих бел$
РКС подразумевают клеточную гибель, вызван$
ков [15]. Функция этих белков, предположи$
ную слабыми стрессорными условиями, внеш$
тельно, заключается в транспорте стеринов
ними или внутренними сигналами, которые
между ПМ и эндоплазматическим ретикулумом
можно искусственно предотвратить, например,
(ЭР). В случае многоклеточных животных такой
путем выключения работы определенных генов.
транспорт необходим для захвата холестерина
ЗКС является естественным (физиологическим)
из плазмы крови [16]. Для пекарских дрожжей
сценарием РКС. В настоящее время описано до$
было показано, что делеция LAM1 приводит к
вольно много индукторов РКС дрожжей и дру$
снижению резистентности клеток к антигриб$
гих одноклеточных организмов, однако физио$
ковому препарату амфотерицину B [15]. По$
логическая роль РКС во многих случаях остает$
скольку амфотерицин B способен встраиваться
ся неизвестной [4-8].
и нарушать функции только тех мембран, где
Ранее мы показали, что высокая концентра$
содержится эргостерин
[17], увеличение
ция полового феромона α$фактора (100 мкг/мл)
чувствительности делеционных мутантов Δlam1
вызывает РКС дрожжей а$типа спаривание [9, 10].
и Δlam2 к амфотерицину B указывает на то, что
Действие полового феромона сопровождается
нарушение транспорта стеринов в дрожжах при$
увеличением в цитоплазме концентрации каль$
водят к увеличению содержания эргостерина в
ция [11], поэтому было высказано предположе$
плазматической мембране. Недавно мы показа$
ние, что увеличение концентрации кальция в
ли, что делеции генов семейства LAM действи$
цитоплазме может быть фактором, принимаю$
тельно приводят к увеличению содержания сте$
щим участие в развитии цепи событий, приводя$
рина в плазматической мембране и во внутрен$
щей к гибели клеток [10]. Антиаритмический
них компартментах клетки [18].
препарат амиодарон также вызывает увеличение
Помимо LAM1 и LAM2 в геноме пекарских
концентрации кальция в цитоплазме [12] и акти$
дрожжей есть еще четыре паралогичных гена то$
вирует цепь событий внутри клетки, схожую с
го же семейства: LAM3, LAM4, LAM5 и LAM6.
той, которая включается в ответ на добавление
При этом синтезируемые ими белки Lam1,
100 мкг/мл феромона [10]. Следует отметить, что
Lam2, Lam3 и Lam4 локализованы в местах кон$
даже такие высокие концентрации α$фактора
тактов ПМ и ЭР, в то время как Lam5 и Lam6 -
приводят к гибели лишь определенной фракции
в местах контакта ЭР и митохондрий (см. обзо$
клеток в суспензии, в то время как существенная
ры Alli$Balogun и Levine [19] и Sokolov et al. [20]).
часть (50-80%) клеток сохраняют жизнеспособ$
Открытие роли белков Lam в транспорте
ность. Размер этой фракции зависит от штамма и
стеринов поставило вопрос о роли стеринового
дальнейшее увеличение концентрации феромо$
состава мембраны в РКС дрожжей. Возможно,
на не приводит к увеличению доли мертвых кле$
эффекты делеций генов LAM1 и LAM2, на РКС,
ток [13]. Механизмы, ограничивающие гибель
вызванную феромоном, амиодароном и ацета$
клеток в присутствии избытка полового феромо$
том, являются следствием изменения содержа$
на, до сих пор неизвестны. Мутация в гене каль$
ния эргостерина в ПМ. В этом случае другие
модулина cmd1 6 [9] или делеция гена метакас$
воздействия на клетку, повышающие или пони$
пазы дрожжей MCA1 [13] приводят к значитель$
жающие содержание стерина в дрожжах, пред$
ному увеличению количества мертвых клеток в
положительно, будут оказывать такое же воз$
присутствии избытка феромона.
действие. Альтернативой этому предположе$
Ранее мы провели два скрининга, направ$
нию является непосредственное участие белков
ленных на поиск генов - положительных регу$
Lam в трансдукции сигнала, приводящего к ги$
ляторов ЗКС дрожжей, вызванной феромоном и
бели клеток. Более того, до сих пор не известно
амиодароном. В результате были выявлены гены
способны ли паралогичные гены LAM дублиро$
LAM1 и LAM2, снижающие резистентность кле$
вать функции друг друга или каждый из них
ток к амиодарону [10, 14]. Первоначально эти
имеет свою узкую специализацию внутри клет$
гены были названы YSP1 и YSP2 (от Yeast
ки.
Suicidal Proteins), однако позднее переименова$
В своей работе мы исследовали РКС, выз$
ны в LAM1 и LAM2 (от LAM - Lipid transfer pro$
ванную высокой концентрацией полового фе$
tein Anchoredat Membrane contactsites, транспор$
ромона и амиодароном. Для этих индукторов
тер липидов, заякоренный в контактных сайтах
РКС мы оценили роль делеций гена LAM2 и му$
мембраны) соответственно. Делеция гена LAM1
таций, приводящих как к увеличению содержа$
также частично защищала клетки от гибели,
ния стерина (UPC2 1), так и к нарушению тер$
вызванной феромоном [10].
минальных стадий биосинтеза стерина (Δerg2,
Недавно было показано, что LAM1 и LAM2
Δerg3, Δerg4, Δerg6). Кроме того, мы сравнили
кодируют белки, принадлежащие одному се$
эффект от одиночной делеции гена LAM2 с эф$
БИОХИМИЯ том 85 вып. 3 2020
350
СОКОЛОВ и др.
фектом от делеции всех четырех генов LAM
Оценка эффективности скрещивания штам=
(LAM1-LAM4). Полученные нами и ранее опуб$
мов. Клетки обоих типов спаривания выращива$
ликованные данные указывают на то, что роль
ли 12-16 ч до логарифмической стадии роста и
генов LAM в РКС дрожжей опосредована изме$
разводили в жидкой YPD до конечной плотнос$
нениями в стериновом составе мембраны.
ти ~2×106 клеток/мл. Далее смешивали клетки
противоположных типов спаривания и инкуби$
ровали 6 ч при 30 °С. После инкубации смесь
МЕТОДЫ ИССЛЕДОВАНИЯ
клеток микроскопировали и подсчитывали до$
лю зигот на 250-300 клеток.
Штаммы, условия культивирования дрожжей
Проточная цитофлуориметрия, индукция РКС
и реагенты. Реактивы, используемые в исследо$
половым феромоном. Клетки выращивали 12-16 ч
вании: амиодарон, α$фактор, пропидия йодид,
в жидкой среде YPD до конечной плотности
синтетическая среда YNB, аминокислотные до$
4×106 клеток/мл. К суспензии добавляли α$фак$
бавки и галактоза («Sigma$Aldrich», США),
тор до конечной концентрации 100 мкг/мл и ин$
дрожжевой экстракт, пептон и агар («Difco»,
кубировали 4 ч на качалке («New Brunswick
США), глюкоза («Helicon», Россия). Клетки вы$
Scientific», США) при 200 об./мин 30 °С. В каче$
ращивали на средах: YPD
(1% дрожжевой
стве отрицательного контроля использовали
экстракт, 2% пептон, 2% глюкоза) и YPGal (1%
суспензию клеток, не обработанную α$факто$
дрожжевой экстракт, 2% пептон и 2% галакто$
ром, которую также инкубировали 4 ч на качал$
за). В твердые среды добавляли агар до 2%.
ке при 30 °С. В качестве положительного кон$
Штаммы UPC2 1, Δlam1, Δlam2, Δlam3, Δlam4 и
троля (мертвые клетки) использовали суспен$
Δlam1Δlam2Δlam3Δlam4 - производные W303.
зию клеток, которую инкубировали 3 ч при
Штаммы Δerg2, Δerg2Δysp2, Δerg3, Δerg3Δysp2,
50 °С. После инкубации все образцы разводили
Δerg4, Δerg4Δysp2, Δerg5, Δerg5Δysp2, Δerg6,
в
8 раз в фосфатно$солевом буфере PBS
Δerg6Δysp2 - производные штамма BY4741 (таб$
(«Gibco», США) и окрашивали пропидий йоди$
лица).
дом в концентрации 0,5 мкг/мл в течение
Смена типа спаривания. Для смены типа спа$
10 мин. Пропидий йодид - флуоресцентный
ривания мы трансформировали штаммы плаз$
зонд, не способный проникать в живые клетки и
мидой pGAL$HO, в которой закодирована экзо$
окрашивающий мертвые клетки [22]. Флуорес$
нуклеаза HO под контролем галактозо$регули$
ценцию клеток, окрашенных пропидий йоди$
руемого промотора PGAL, по методу, опублико$
дом, анализировали с помощью проточных ци$
ванному в работе Herskowitz и Jensen [21]. Полу$
тофлуориметров CytoFlex («Beckman», США) и
ченные штаммы инкубировали ночь в среде
Beckman Coulter FC 500 («Beckman», США) при
YPGal (2% галактоза) и переносили на твердую
длине волны возбуждения 488 нм и эмиссион$
среду YPD (2% глюкоза). Тип спаривания полу$
ном фильтре 690/50 нм. Каждый эксперимент
ченных таким образом отдельных колоний оп$
был выполнен не менее чем в трех независимых
ределяли контрольным скрещиванием со штам$
повторах, полученных в разные дни. В каждом
мами а и альфа типа. Колонии с нужным типом
эксперименте анализировали не менее 10 000 со$
спаривания сутки выращивали в жидкой среде
бытий (клеток).
YPD для потери плазмиды pGAL$HO. Потерю
плазмиды pGAL$HO проверяли по отсутствию
роста на селективной среде YNB$leu («Sigma$
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Aldrich», США).
Индукция РКС амиодароном, и оценка жиз=
Делеция гена LAM2 частично предотвращает
неспособности клеток дрожжей. Клетки выращи$
гибель клеток, вызванную избытком полового
вали 12-16 ч до логарифмической стадии роста
феромона, и не проявляет аддитивность с деле$
и разводили до конечной плотности 4×106 кле$
циями генов LAM1, LAM3 и LAM4.
ток/мл в YPD с добавлением 25 мМ MES (2$(N$
Как отмечалось выше, в геноме дрожжей со$
морфолино)этансульфоновая кислота) («Sigma$
держатся четыре гена LAM (LAM1-LAM4). Ко$
Aldrich», США), pH 5,5. К суспензии клеток до$
дируемые ими белки (Lam1-Lam4) локализова$
бавляли амиодарон до конечной концентрации
ны в местах контакта ПМ и ЭР. Ген LAM2 (YSP2)
60 мкМ. Инкубацию с амиодароном проводили
был ранее найден нами в результате генетичес$
в течение часа при 30 °С. Затем клетки разбавля$
кого скрининга, направленного на поиск мута$
ли и переносили на твердую среду YPD. Через
ций, снижающих чувствительность к амиодаро$
24 ч измеряли долю выживших клеток, как со$
ну [14]. Этот ген является паралогом гена LAM1
отношение колониеобразующих единиц (КОЕ)
(YSP1) [15], делеция которого частично пред$
после стресса к КОЕ до стресса.
отвращает гибель клеток, вызванную высокой
БИОХИМИЯ том 85 вып. 3 2020
РОЛЬ ГЕНОВ LAM В СМЕРТИ КЛЕТОК ДРОЖЖЕЙ
351
Штаммы, использованные в исследовании
Штамм
Генотип
Родительский штамм
или источник
W303 1A
MATa ade2 101 his3 11 trp1 1 ura3 52 can1 100 leu2 3
Laboratory of A. Hyman
Δlam2
MATa ade2 101 his3 11 trp1 1 ura3 52 can1 100 leu2 3 Δlam2::TRP1
[14]
Δlam1Δlam2
MATa ade2 101 his3 11 trp1 1 ura3 52 can1 100 leu2 3 MATa ade2 101 his3 11 trp1 1
[35]
Δlam3Δlam4
ura3 52 can1 100 leu2 3 Δlam3::kanMX4 Δlam2::TRP1 Δlam1::NAT Δlam4::loxP
UPC2 1 MATa
MATa UPC2 1 ura3 1 his3 11, 15 leu2 3, 112 trp1 1
UPC2 1 MATalpha
UPC2 1 Δlam2
MATa UPC2 1 ura3 1 his3 11, 15 leu2 3, 112 trp1 1 Δlam2::HIS3
UPC2 1 MATa
PGAL LAM2
MATa ade2 101 his3 11 trp1 1 ura3 52 can1 100 leu2 3 PGAL LAM2::kanMX4
W303 1A
cmd1 6
[10]
BY4741
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0
EUROSCARF
BY4742
MATalpha his3Δ1 leu2Δ0 met15Δ0 ura3Δ0
EUROSCARF
Δerg2MATa
Δerg2 MATalpha
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 erg2::kanMX4
EUROSCARF
Δerg3MATa
Δerg3 MATalpha
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 erg3::kanMX4
EUROSCARF
Δerg4MATa
Δerg4 MATalpha
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 erg4::kanMX4
EUROSCARF
Δerg5MATa
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 erg5::kanMX4
Δerg5 MATalpha
EUROSCARF
Δerg6MATa
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 erg6::kanMX4
Δerg6 MATalpha
EUROSCARF
Δerg2 Δlam2
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 erg2::kanMX4 Δlam2::HIS3
Δerg2 MATa
Δerg3 Δlam2
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 erg3::kanMX4 Δlam2::HIS3
Δerg3 MATa
Δerg4 Δlam2
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 erg4::kanMX4 Δlam2::HIS3
Δerg4 MATa
Δerg5 Δlam2
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 erg5::kanMX4 Δlam2::HIS3
Δerg5 MATa
Δerg6 Δlam2
MATa his3Δ1 leu2Δ0 met15Δ0 ura3Δ0 erg6::kanMX4 Δlam2::HIS3
Δerg6 MATa
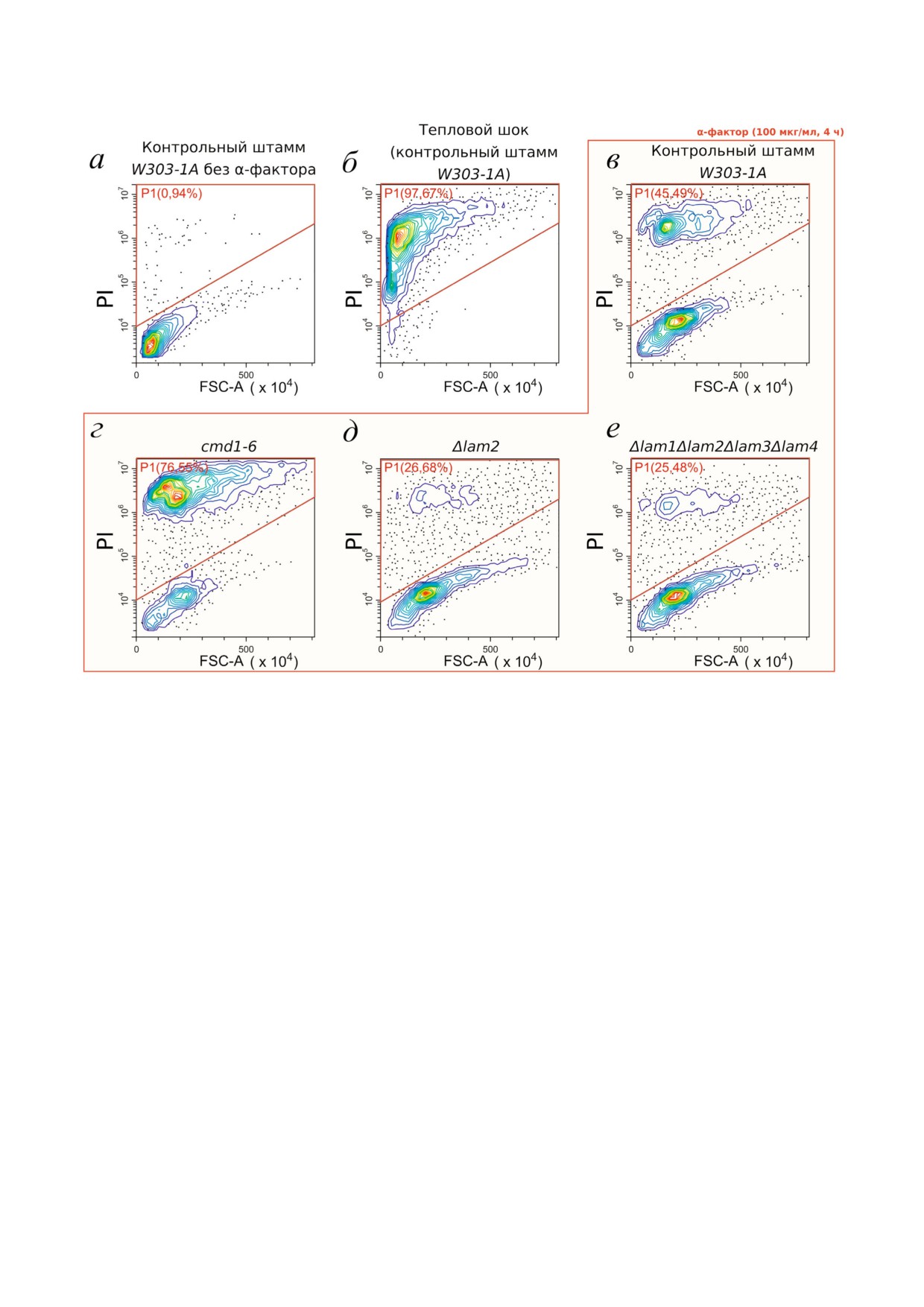
концентрацией α$фактора [10]. Мы проверили,
~45% клеток (рис. 1, в), а мутация в гене кальмо$
будет ли делеция остальных трех генов (LAM1,
дулина увеличивала чувствительность клеток к
LAM3 и LAM4) ингибировать РКС, вызванную
воздействию α$фактора (рис.1, г), что соответ$
феромоном. Для этого мы сравнили соотноше$
ствует данным литературы [23]. Мы обнаружи$
ние живых и мертвых клеток в суспензионной
ли, что делеция гена LAM2 частично предотвра$
культуре клеток дрожжей, подверженных
щала гибели клеток под действием α$фактора
действию α$фактора (100 мкг/мл, 4 ч). Мы раз$
(рис. 1, д). Одновременно с оценкой сигнала от
личали живые и мертвые клетки по накоплению
пропидий йодида мы оценивали линейные раз$
ими пропидий йодида с помощью проточной
меры клеток по светорассеянию. Как видно из
цитофлуорометрии (рис. 1, а, б). Пропидий йодид
рис. 1, α$фактор вызывал значительное увели$
способен проникать только в мертвые клетки, в
чение линейного светорассеяния (FSC$A). Это
то время как живые клетки с целостной плазма$
увеличение размера индивидуальных клеток, по
тической мембраной остаются неокрашенными.
всей видимости, обусловлено образованием
Добавление α$фактора приводило к гибели
«шму» - половых выростов и увеличением объ$
БИОХИМИЯ том 85 вып. 3 2020
352
СОКОЛОВ и др.
Рис. 1. Распределение линейных размеров (FSC$A) клеток и интенсивности сигнала от пропидий йодида (PI). a - Кон$
трольные клетки W303 1A в отсутствие стресса (отрицательный контроль на мертвые клетки); б - клетки W303 1A под$
вергнутые действию теплового шока - 50°С, 3 ч (положительный контроль на мертвые клетки); в-е - клетки, подвергну$
тые действию α$фактора (100 мкг/мл, 4 ч) различных штаммов: родительский штамм (в); штамм с мутацией в гене каль$
модулина cmd1 6 (г); штамм с делецией гена LAM2 (д); штамм с делециями всех четырех генов LAM (LAM1-LAM4) (e). На
графиках обозначена зона (gate), событие в которой расценивались нами как мертвые клетки. Для каждого из условий
указан процент мертвых клеток от общего числа клеток. Данные репрезентативных экспериментов, по 10 000 событий в
каждом эксперименте.
biokhsm/
ема клеток. Делеция гена LAM2 не препятство$
Также, как и в случае с α$фактором, нарушение
вала увеличению линейного светорассеяния,
LAM2 оказывало максимальный эффект, и мы
что свидетельствует о чувствительности клеток к
не обнаружили значимой разницы в резистент$
половому феромону.
ности к амиодарону между штаммом Δlam2 и
Как видно из рис. 2, делеция LAM2 оказыва$
штаммом Δlam1Δlam2Δlam3Δlam4 с делецией че$
ет наибольший эффект из проверенных генов
тырех генов LAM (рис. 3, а).
LAM на устойчивость к α$фактору, делеция до$
РКС, вызванная половым феромоном, сниже=
полнительно трех генов LAM на фоне делеции
на в штамме UPC2 1. Штамм с частично доми$
LAM2 не привела к увеличению выживаемости
нантным аллелемUPC2 1 демонстрирует повы$
клеток в присутствии полового феромона. Ана$
шенную скорость захвата стеринов клетками
логичный результат мы получили в случае с дру$
[24] и активирует транскрипцию генов ERG, что
гим индуктором РКС дрожжей — амиодароном.
приводит к увеличению содержания стеринов в
Ранее мы показали, что добавление амиодарона
клетке и ПМ в частности [25]. Делеция генов
к клеткам вызывает каскад событий, что и при
LAM также приводит к увеличению концентра$
добавлении избытка полового феромона [10].
ции эргостерина в плазматической мембране
БИОХИМИЯ том 85 вып. 3 2020
РОЛЬ ГЕНОВ LAM В СМЕРТИ КЛЕТОК ДРОЖЖЕЙ
353
[15, 25]. Если резистентность делеционных
бы проверить это предположение мы исследо$
штаммов по генам LAM обусловлена изменения$
вали гибель дрожжей штамма UPC2 1 и дикого
ми концентрации эргостерина в ПМ, то аллель
типа под действием α$фактора (100 мкг/мл). На
UPC2 1 также должен предотвращать гибель
рис. 4 приведена типичная контурная диаграм$
клеток, вызванную половым феромоном. Что$
ма, где по одной оси отложена интенсивность
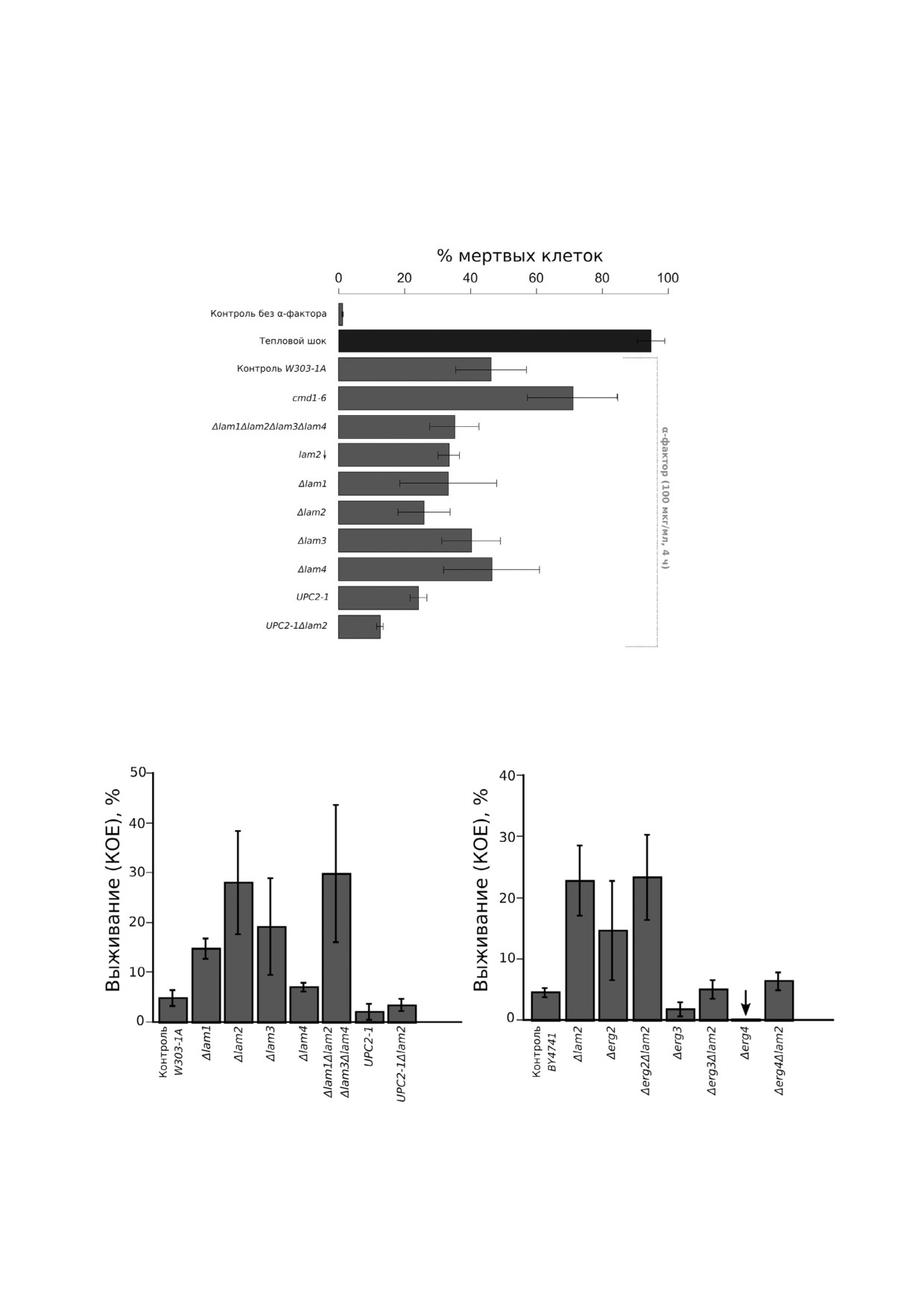
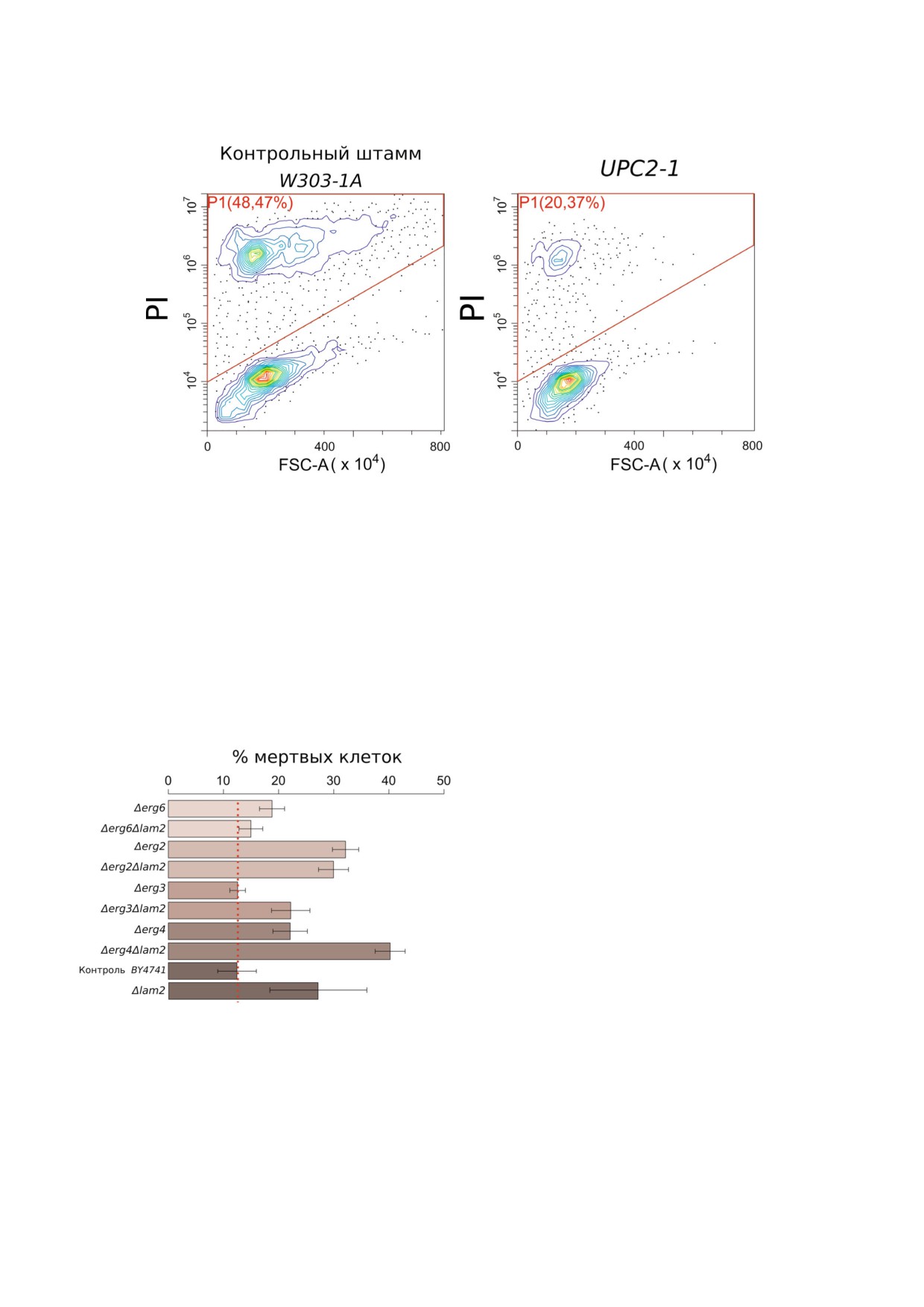
Рис. 2. РКС клеток дрожжей с нарушенным транспортом и накоплением эргостерина, вызванным добавлением α$факто$
ра (100 мкг/мл). Соотношение мертвых и живых клеток оценивали с помощью пропидий йодида и проточной цитофлуо$
риметрии. На столбчатых диаграммах указано среднее значение ± стандартное отклонение (n = 3)
а
б
Рис. 3. Выживание клеток в присутствии индуктора РКС дрожжей амиодарона (60 мкМ, 1 ч инкубации). За 100% прини$
мали количество клеток (КОЕ) в начальный момент времени (до добавки амиодарона)
БИОХИМИЯ том 85 вып. 3 2020
354
СОКОЛОВ и др.
а
б
Рис. 4. Распределение линейных размеров (FSC$A) клеток и интенсивности сигнала от пропидий йодида (PI) в клетках
дрожжей дикого типа (а) и мутантного штамма UPC2 1 (б). Клетки инкубировали 4 ч в присутствии 100 мкг/мл α$факто$
ра. Данные репрезентативных экспериментов, 10 000 событий в каждом эксперименте.
biokhsm/
сигнала от пропидий йодидом, а по другой
значит эта мутация ингибирует гибель клеток,
оси - линейный размер клеток, который мы
вызванную α$фактором (см. также рис. 2, где
оценивали по амплитуде линейного светорассея$
представлены усредненные данные трех экспе$
ния (FSC$A). Как видно из рисунка, мутация
риментов). В отношении амиодарона штамм с
UPC2 1 приводила к снижению относительного
мутацией UPC2 1 напротив демонстрировал по$
числа клеток, окрашенных пропидий йодидом,
вышенную чувствительность (рис. 3, а). Срав$
нение частоты образования зигот при скрещи$
вании штаммов UPC2 1 MATa с UPC2
1MATalpha (2,2%), Δlam1Δlam2Δlam3Δlam4MATa
с Δlam1Δlam2Δlam3Δlam4MATalpha (1,8%) и ди$
кого типа BY4741 (MATa) с BY4742 (MATalpha)
(2.3%) не выявило различий, а значит повы$
шенная
устойчивость
UPC2 1
и
Δlam1Δlam2Δlam3Δlam4 к α$фактору не связана
со снижением ответа клеток на него.
Генетические взаимодействия LAM2 и генов
биосинтеза эргостерина. В следующей серии экс$
периментов мы исследовали РКС дрожжей,
вызванную феромоном и амиодароном, на клет$
ках дрожжей с делециями генов ERG2-ERG6.
Эти гены кодируют ферменты, катализирующие
терминальные этапы биосинтеза эргостерина.
Нарушение биосинтеза эргостерина, как прави$
Рис. 5. РКС клеток дрожжей с мутациями по генам био$
синтеза эргостерина, вызванная добавлением α$фактора
ло, летально [26], однако делеция генов, кото$
(100 мкг/мл). Соотношение мертвых и живых клеток оце$
рые кодируют ферменты терминальных стадий
нивали с помощью пропидий йодида и проточной цито$
биосинтеза от ланостерина до эргостерина, не
флуориметрии. На столбчатых диаграммах указано среднее приводит к потере клетками жизнеспособности
значение ± стандартное отклонение (n = 3).
[26]. Из нашего анализа мы исключили ген
С цветным вариантом рис. 5 можно ознакомиться в элек$
ERG5, так как штамм Δerg5 с высокой частотой
journal/biokhsm/
(до 80%) терял митохондриальную ДНК, а поте$
БИОХИМИЯ том 85 вып. 3 2020
РОЛЬ ГЕНОВ LAM В СМЕРТИ КЛЕТОК ДРОЖЖЕЙ
355
ря митохондриальной ДНК значительно снижа$
вательности, соответствующей реакциям био$
ет чувствительность клеток к α$фактору [9, 10].
синтеза эргостерина ферментами, закодирован$
Следует также отметить, что в этой серии опы$
ными этими генами. Как видно из рисунка, де$
тов мы использовали другой родительский
леция генов ERG не снижала, а иногда даже уве$
штамм (BY4741), тогда как эксперименты с
личивала долю мертвых клеток в присутствии α$
UPC2 1 проводили на штаммах, полученных на
фактора. При этом делеция LAM2 в штаммах
основе лабораторного штамма W303. Чувстви$
Δerg6 и Δerg2 не оказывала значительного эф$
тельность этих лабораторных штаммов к феро$
фекта на РКС, индуцированную феромоном
мону значительно различалась.
(рис. 5). В то же время мы не наблюдали сниже$
Мы оценили процент мертвых клеток дрож$
ния РКС дрожжей от делеции в штаммах с нару$
жей в штамме дикого типа и в штаммах с деле$
шенным синтезом эргостерина. Делеция LAM2 в
цией генов: ERG6, ERG2, ERG3 и ERG4 (рис. 5).
штамме Δerg4 и в контрольном штамме BY4741
На рис. 5 результаты с разными делеционными
приводила к существенному увеличению доли
мутантами по генам ERG приведены в последо$
мертвых клеток в присутствии α$фактора.
а
в
б
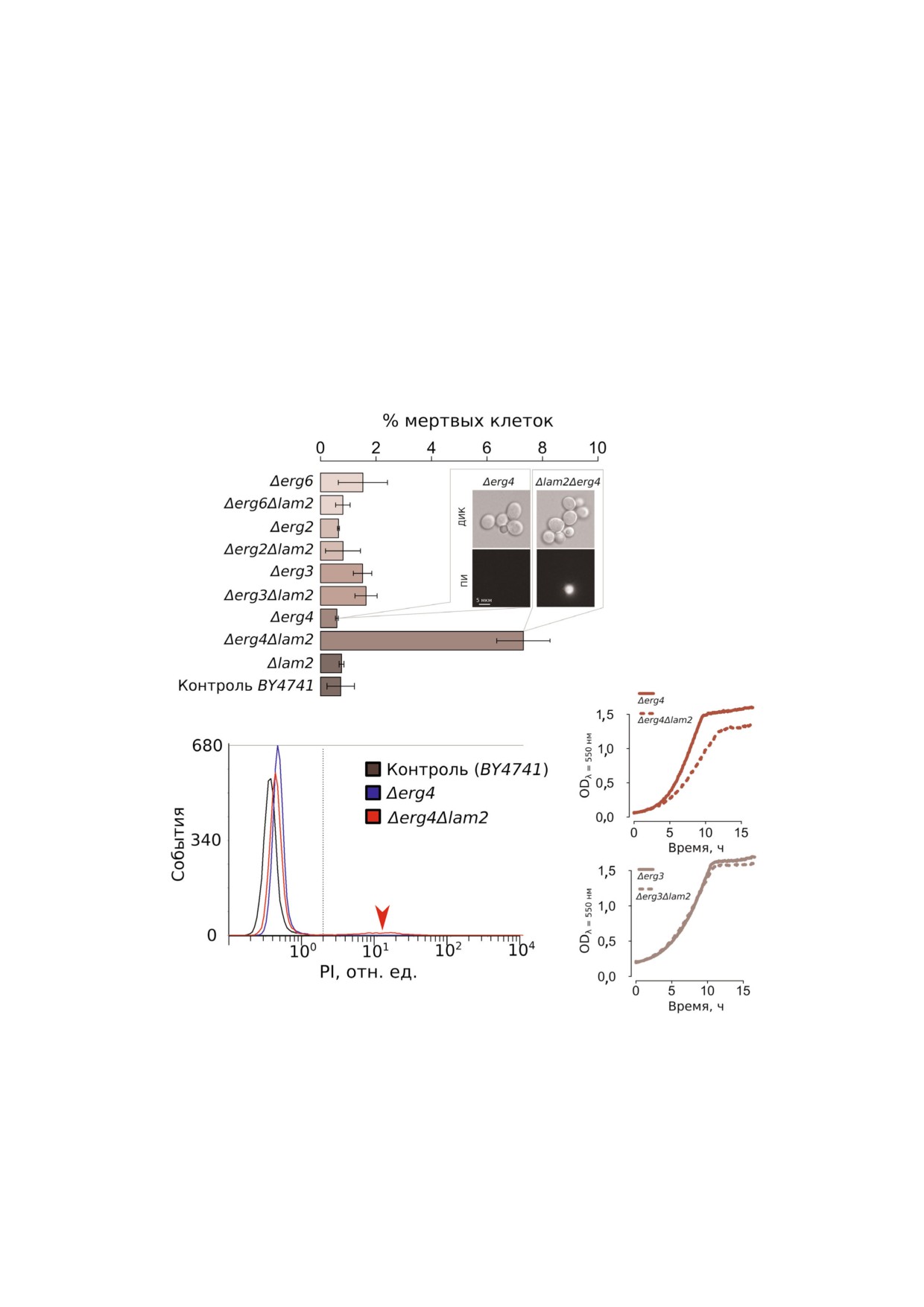
Рис. 6. Штамм Δerg4Δlam2 с двойной делецией содержит субпопуляцию мертвых клеток, окрашиваемых пропидий йоди$
дом, и обладает сниженной скоростью роста. а - Анализ выживаемости штаммов в контрольных условиях (среднее ±
± стандартное отклонение, n = 3); во вставке представлена фотография клеток штаммов Δerg4Δlam2 и Δerg4, окрашенных
пропидий йодидом. б - Репрезентативная гистограмма эксперимента с проточного цитофлуориметра. в - Сравнение ско$
ростей роста штаммов Δerg4 и Δerg4Δlam2 (сверху), Δerg3 и Δerg3Δlam2 (снизу) по увеличению светорассеяния (длина вол$
ны λ = 550 нм).
biokhsm/
БИОХИМИЯ том 85 вып. 3 2020
356
СОКОЛОВ и др.
Мы обратили внимание, что в штамме
этого предположения требует дополнительных
Δerg4Δlam2 с двойной делецией даже в отсут$
исследований. С другой стороны, штаммы с
ствии полового феромона количество мертвых
одиночными делециями Δlam3 и Δlam4 не вызы$
(пропидий йодид положительных) клеток зна$
вали устойчивости к α$фактору (рис. 2). Учиты$
чительно увеличено по сравнению с родитель$
вая данные широкомасштабных масс$спектро$
ским штаммом и со всеми другими исследован$
метрических протеомных анализов, что Lam2р
ными нами двойными и одиночными мутанта$
содержится в клетках в больших количествах,
ми (рис. 6, а, б). Мы показали, что делеция
чем другие белки Lam [28, 29], можно предполо$
LAM2 существенно снижает скорость роста
жить, что отсутствие эффекта делеций LAM3 и
штамма, в котором делетирован ген ERG4, в то
LAM4 на резистентность к РКС, вызванной α$
время как при делеции гена ERG3, находящего$
фактором, связано с разницей в концентрации
ся на один шаг выше по каскаду в метаболичес$
этих белков в стандартных условиях. Однако в
ком пути биосинтеза стерина, такого эффекта
этих анализах количество Lam2р не превышало
не наблюдается (рис. 6, в). Таким образом, наши
суммарного количества его трех паралогов, а в
данные указывают на эпистаз между геном
другой работе с упрощенным протоколом подго$
LAM2 и геном ERG4.
товки проб к масс$спектрометрии Lam1p даже
Исследование устойчивости к амиодарону
был обнаружен в больших количествах [30]. Та$
показало, что делеция LAM2 повышает выживае$
ким образом, мы считаем, что отсутствие эффек$
мость клеток на фоне всех испытанных мутаций
та от делеции генов LAM1, LAM3, LAM4 в штам$
(Δerg2, Δerg3, Δerg4 и UPC2 1) (рис. 3).
ме Δlam2 не связано с низким содержанием этих
белков в клетке, а отражает их функциональную
взаимосвязь. Распределение вклада делеций ге$
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
нов LAM в устойчивость к амиодарону оказалось
сходным: LAM2 вносит основной вклад, а допол$
Исследование генов LAM в пекарских дрож$
нительное нарушение генов LAM1, LAM3, LAM4
жах затруднено из$за того, что они представле$
не увеличивает устойчивость.
ны паралогичными парами [20]. В связи с этим
Лабораторный штамм BY4741 оказался зна$
до сих пор было описано довольно мало фено$
чительно более резистентным к РКС, вызван$
типов для штаммов, в которых нокаутированы
ной α$фактором, чем штамм W303. При этом де$
отдельные гены LAM. Так, например, нами ра$
леция гена LAM2 не уменьшала, а наоборот уве$
нее было показано, что делеция LAM2 приводит
личивала долю мертвых клеток в нашей экспе$
к увеличению раздробленности митохондриаль$
риментальной модели с половым феромоном
ной сети в клетках, находящихся в стационар$
(рис. 5). В то же время эффект делеции LAM2 на
ной фазе роста [27]. Исключениями являются
резистентность к амиодарону не зависел от ге$
устойчивость к амиодарону, показанная нами
нетического фона, который мы использовали
ранее для Δlam1 и Δlam2 [10, 14], и устойчивость
(рис. 4). Считается, амиодарон вызывает тот же
к α$фактору, которая показана для штамма
каскад событий в клетке, что и половой феро$
Δlam1 [10]. В этой работе мы показали, что деле$
мон, но на более поздних стадиях - амиодарон
ция LAM2 также демонстрирует заметное сни$
стимулирует вход избытка кальция в цитоплаз$
жение чувствительности штамма W303 S. cere
му клетки [10, 12]. По$видимому, это происхо$
visiae к РКС, вызванной α$фактором (рис. 1, 2).
дит из$за гиперполяризации ПМ вследствие на$
Далее, воспользовавшись этим выраженным
рушения функционирования ионных каналов
фенотипом в качестве тест$системы для изуче$
[31]. Вероятно, вызванное делецией LAM2 по$
ния генетических взаимодействий, мы исследо$
вышение эргостерина на ПМ влияет как на ак$
вали РКС, вызванную феромоном в штамме
тивность ионных каналов ПМ, так и на актив$
Δlam1Δlam2Δlam3Δlam4 с четверной делецией.
ность феромонового рецептора. При этом эф$
Делеция дополнительных генов LAM не привела
фект гена LAM2 на каскад событий, вызванный
к снижению чувствительности штамма к α$фак$
половым феромоном, сильно зависит от генети$
тору дополнительно к тому, что наблюдалось в
ческого фона. Картирование мутаций, обеспе$
штамме с одиночной делецией Δlam2. Получен$
чивающих различие штаммов BY4741 и W303 по
ный результат говорит о том, что другие белки
отношению к РКС, вызванной половым феро$
Lam не проявляют функции, связанной с кле$
моном, требует дополнительных исследований.
точным ответом на половой феромон, если но$
Однако на основании представленных в этой
каутирован ген LAM2. Можно предположить,
работе результатов можно заключить, что роль
что белки Lam функционируют в качестве гете$
генов LAM в РКС, вызванной феромоном, не
родимеров и наличие LAM2 является необходи$
консервативна и зависит от других генетических
мым фактором для их работы, однако проверка
факторов.
БИОХИМИЯ том 85 вып. 3 2020
РОЛЬ ГЕНОВ LAM В СМЕРТИ КЛЕТОК ДРОЖЖЕЙ
357
Мы обнаружили аддитивность между эффек$
минимум двух генов MAP$киназного каскада,
тами от делеций генов LAM2 и ERG4 (рис. 5, 6).
инициируемого также феромоном [36]. Возни$
ERG4 кодирует C$24(28) стерин редуктазу, кото$
кает предположение, что в мутантах по LAM по$
рая катализирует последнюю реакцию метабо$
вышенный эргостерол ПМ предотвращает РКС,
лического пути биосинтеза эргостерина в
снижая активность Upc2. Наши данные свиде$
дрожжах [32]. При этом в штамме Δerg3, где от$
тельствуют против этого предположения.
сутствует фермент, катализирующий предпос$
Действительно, гиперактивация Upc2 (мутация
леднюю реакцию этого же пути, эффект от де$
UPC2 1) не увеличивает, а снижает феромон$за$
леции LAM2 был меньше в абсолютном выра$
висимую РКС (рис. 1, 2). Поскольку Upc2 явля$
жении. На фоне делеций генов ERG2 и ERG6
ется основным активатором синтеза эргостери$
изменений резистентности к феромону мы не
на, можно предположить, что увеличение концен$
обнаружили. Делеция генов биосинтеза эрго$
трации эргостерина в ПМ влияет на ионные по$
стерина приводит к изменению стеринового
токи через ПМ, и это приводит к устойчивости.
состава клеточных мембран - эргостерин заме$
Действительно, известно, что мутации по генам
щается на интермедиаты его биосинтеза [33].
биосинтеза эргостерина приводят к нарушениям
Делеция ERG6 и ERG2 обрывает биосинтез на
работы различных транспортеров ПМ [37].
зимостерине и фекостерине соответственно
Повышенная чувствительность клеток
[34], однако нам не известны работы, в которых
штамма UPC2 1 к амиодарону также хорошо
бы определяли количественное распределение
согласуется с данными литературы. Амиодарон
интермедиатов биосинтеза стерина в делецион$
вызывает увеличение концентрации кальция в
ных штаммах. Мы предполагаем, что отсут$
цитоплазме [12], что вызывает дальнейший кас$
ствие эффекта от делеции LAM2 в штаммах
кад событий, приводящий к гибели клеток [10].
Δerg2 и Δerg6 связано с тем, что LAM2 не спосо$
При этом повышение уровня эргостерина вызы$
бен полностью или частично транспортировать
вает нарушения в вакуолях [38]. Нормальное
зимостерин, фекостерин и ланостерин - ран$
функционирование вакуолей, в свою очередь,
ние интермедиаты биосинтеза. В этом случае
обеспечивает устойчивость к амиодарону: вакуо$
делеция гена LAM2 никак не может повлиять на
ли способны удалять избыточный Ca2+ из ци$
фенотип клеток. В то же время согласно пред$
топлазмы, и это их свойство критично для ус$
лагаемой нами модели, поздние интермедиаты
тойчивости к амиодарону - токсичность амио$
биосинтеза и эргостерин транспортируются
дарона опосредована подъемом Ca2+ [39].
LAM2.
Таким образом, по нашему мнению, наибо$
Сам факт того, что делеции LAM снижают
лее предпочтительным объяснением роли генов
смерть дрожжевых клеток от высоких концен$
LAM в РКС, вызванной феромоном, является
траций полового феромона и амиодарона, из$
следующее: гены LAM влияют на процессы, про$
вестен уже более десяти лет [10, 14]. Тем не менее
текающие внутри клетки опосредованно. Эф$
молекулярный механизм участия белков Lam в
фекты их делеций связаны с изменением содер$
этих типах РКС оставался неясен. Планируя на$
жания эргостерина в плазматической мембране,
ше исследование, мы рассматривали две воз$
которое, в свою очередь, меняет активность
можности: (1) мутации по генам LAM влияют на
мембранных белков и обусловленных ими ион$
РКС опосредованно, влияя на эргостерин ПМ;
ных потоков между внешней средой и цитоплаз$
(2) белки Lam непосредственно взаимодей$
мой.
ствуют с сигнальными молекулами ПМ. Наши
данные указывают на первую возможность, так
как все другие вмешательства в гомеостаз стери$
Финансирование. Работа выполнена при под$
нов в клетке также сильно влияли на устойчи$
держке Российского научного фонда (грант
вость клеток к этим типам РКС. Вероятно, деле$
№ 18$14$00151) (рис. 1, 2, 4, 5, 6) и Российского
ции генов LAM приводят к увеличению эргосте$
фонда фундаментальных исследований (грант
рина в ПМ. Это следует из приведенных выше
№ 18$04$01183) (рис. 3).
данных по чувствительности к амфотерицину В,
Благодарности. Выражаем благодарность Ва$
а также из наших недавних работ по чувстви$
силине Ефимовой за участие в получении двой$
тельности к липофильным катионам [35] и изу$
ных Δerg Δlam2 мутантов.
чению влияния генов LAM на устойчивость к
Конфликт интересов. Авторы заявляют об от$
стрессу [18].
сутствии конфликта интересов.
Как повышенный эргостерин ПМ может
Соблюдение этических норм. Настоящая
влиять на РКС? Известно, что высокий эргосте$
статья не содержит каких$либо исследований с
рин предотвращает перемещение Upc2 в ядро.
участием людей или использованием животных
При этом Upc2 активирует транскрипцию как
в качестве объектов исследований.
БИОХИМИЯ том 85 вып. 3 2020
358
СОКОЛОВ и др.
СПИСОК ЛИТЕРАТУРЫ
1.
Nagata, S. (2018) Apoptosis and clearance of apoptotic
Hartman, C.L., Strugatsky, D., Lee, S.D., He, C., Hong, C.,
cells, Annu. Rev. Immunol., 36, 489-517, doi: 10.1146/
Jiang, H., Bentolila, L.A., Gatta, A.T., Levine, T.P., Ferng, A.,
annurev$immunol$042617$053010.
Lee, R., Ford, D.A., Young, S.G., Ikonen, E.,
2.
Fuchs, Y., and Steller, H. (2015) Live to die another way:
Schwabe, J.W.R., and Tontonoz, P. (2018) Aster proteins
modes of programmed cell death and the signals emanating
facilitate nonvesicularplasma membrane to ER cholesterol
from dying cells, Nat. Rev. Mol. Cell Biol., 16, 329-344,
transport in mammalian cells, Cell, 175, 514-529.e20,
doi: 10.1038/nrm3999.
doi: 10.1016/j.cell.2018.08.033.
3.
Carmona$Gutierrez, D., Bauer, M.A., Zimmermann, A.,
17.
Huang, W., Zhang, Z., Han, X., Tang, J., Wang, J., Dong, S.,
Aguilera, A., Austriaco, N., et al. (2018) Guidelines and
and Wang, E. (2002) Ion channel behavior of amphotericin
recommendations on yeast cell death nomenclature,
B in sterol$free and cholesterol$ or ergosterol$containing
Microb. Cell Fact., 5, 4-31, doi: 10.15698/mic2018.01.607.
supported phosphatidylcholine bilayer model membranes
4.
Gordeeva, A.V., Labas, Y.A., and Zvyagilskaya, R.A. (2004)
investigated by electrochemistry and spectroscopy, Biophys.
Apoptosis in unicellular organisms: mechanisms and evo$
J., 83, 3245-3255, doi: 10.1016/S0006$3495(02)75326$5.
lution, Biochemistry (Moscow),
69,
1055-1066,
18.
Sokolov, S., Vorobeva, M., Smirnova, E., Smirnova, A.,
doi: 10.1023/b:biry.0000046879.54211.ab.
Trushina, N., Galkina, K., Severin, F., and Knorre, D.
5.
Severin, F.F., Meer, M.V., Smirnova, E.A., Knorre, D.A.,
(2020) LAM genes contribute to environmental stress toler$
and Skulachev, V.P. (2008) Natural causes of programmed
ance but sensibilise yeast cells to azoles, Front. Microbiol.,
death of yeast Saccharomyces cerevisiae, Biochim. Biophys.
11, 38, doi: 10.3389/fmicb.2020.00038.
Acta, 1783, 1350-1353, doi: 10.1016/j.bbamcr.2008.
19.
Alli$Balogun, G.O., and Levine, T.P. (2019) Regulation of
02.001.
targeting determinants in interorganelle communication,
6.
Carmona$Gutierrez, D., Eisenberg, T., Büttner, S.,
Curr. Opin. Cell Biol., 57, 106-114, doi: 10.1016/j.ceb.
Meisinger, C., Kroemer, G., and Madeo, F.
(2010)
2018.12.010.
Apoptosis in yeast: triggers, pathways, subroutines, Cell
20.
Sokolov, S.S., Trushina, N.I., Severin, F.F., and Knorre, D.A.
Death Differ., 17, 763-773, doi: 10.1038/cdd.2009.219.
(2019) Ergosterol turnover in yeast: an interplay between
7.
Sukhanova, E.I., Rogov, A.G., Severin, F.F., and
biosynthesis and transport, Biochemistry (Moscow), 84,
Zvyagilskaya, R.A.
(2012) Phenoptosis in yeasts,
346-357, doi: 10.1134/S0006297919040023.
Biochemistry (Moscow), 77, 761-775, doi: 10.1134/
21.
Herskowitz, I., and Jensen, R.E. (1991) Putting the HO
S0006297912070097.
gene to work: practical uses for mating$type switching, in
8.
Aouacheria, A., Cunningham, K.W., Hardwick, J.M.,
Methods in Enzymology, Academic Press, pp. 132-146,
Palková, Z., Powers, T., Severin, F.F., and Váchová. L.
doi: 10.1016/0076$6879(91)94011$z.
(2018) Comment on «Sterilizing immunity in the lung
22.
Sträuber, H., and Müller, S. (2010) Viability states of bac$
relies on targeting fungal apoptosis$like programmed cell
teria - specific mechanisms of selected probes, Cytometry
death», Science, 360, doi: 10.1126/science.aar6910.
A, 77, 623-634, doi: 10.1002/cyto.a.20920.
9.
Severin, F.F., and Hyman, A.A. (2002) Pheromone induces
23.
Moser, M.J., Geiser, J.R., and Davis, T.N. (1996) Ca2+$
programmed cell death in S. cerevisiae, Curr. Biol., 12,
calmodulin promotes survival of pheromone$induced
R233-R235, doi: 10.1016/j.cellbi.2005.10.023.
growth arrest by activation of calcineurin and Ca2+$
10.
Pozniakovsky, A.I., Knorre, D.A., Markova, O.V.,
calmodulin$dependent protein kinase, Mol. Cell. Biol., 16,
Hyman, A.A., Skulachev, V.P., and Severin, F.F. (2005)
4824-4831, doi: 10.1128/mcb.16.9.4824.
Role of mitochondria in the pheromone$ and amiodarone$
24.
Crowley, J.H., Leak, F.W. Jr., Shianna, K.V., Tove, S., and
induced programmed death of yeast, J. Cell Biol., 168,
Parks, L.W. (1998) A mutation in a purported regulatory
257-269, doi: 10.1083/jcb.200408145.
gene affects control of sterol uptake in Saccharomyces cere
11.
Ohsumi, Y., and Anraku, Y. (1985) Specific induction of
visiae, J. Bacteriol., 180, 4177-4183.
Ca2+ transport activity in MATa cells of Saccharomyces
25.
Joshua, I.M., and Höfken, T. (2017) From lipid homeosta$
cerevisiae by a mating pheromone, alpha factor, J. Biol.
sis to differentiation: old and new functions of the zinc
Chem., 260, 10482-10486.
cluster proteins Ecm22, Upc2, Sut1 and Sut2, Int. J. Mol.
12.
Gupta, S.S., Ton, V.$K., Beaudry, V., Rulli, S.,
Sci., 18, doi: 10.3390/ijms18040772.
Cunningham, K., and Rao, R. (2003) Antifungal activity of
26.
Daum, G., Lees, N.D., Bard, M., and Dickson, R. (1998)
amiodarone is mediated by disruption of calcium homeo$
Biochemistry, cell biology and molecular biology of lipids
stasis, J. Biol. Chem., 278, 28831-28839, doi: 10.1074/
of Saccharomyces cerevisiae, Yeast,
14,
1471-1510,
jbc.M303300200.
doi: 10.1002/(SICI)1097$0061(199812)14:16<1471::AID$
13.
Zhang, N.$N., Dudgeon, D.D., Paliwal, S., Levchenko, A.,
YEA353>3.0.CO;2$Y.
Grote, E., and Cunningham, K.W. (2006) Multiple signal$
27.
Knorre, D.A., Ojovan, S.M., Saprunova, V.B., Sokolov, S.S.,
ing pathways regulate yeast cell death during the response
Bakeeva, L.E., and Severin, F.F. (2008) Mitochondrial
to mating pheromones, Mol. Biol. Cell, 17, 3409-3422,
matrix fragmentation as a protection mechanism of yeast
doi: 10.1091/mbc.e06$03$0177.
Saccharomyces cerevisiae, Biochemistry (Moscow), 73,
14.
Sokolov, S., Knorre, D., Smirnova, E., Markova, O.,
1254-1259, doi: 10.1134/s0006297908110126.
Pozniakovsky, A., Skulachev, V., and Severin, F. (2006)
28.
de Godoy, L.M.F., Olsen, J.V., Cox, J., Nielsen, M.L.,
Ysp2 mediates death of yeast induced by amiodarone or
Hubner, N.C., Fröhlich, F., Walther, T.C., and Mann, M.
intracellular acidification, Biochim. Biophys. Acta, 1757,
(2008) Comprehensive mass$spectrometry$based proteome
1366-1370, doi: 10.1016/j.bbabio.2006.07.005.
quantification of haploid versus diploid yeast, Nature, 455,
15.
Gatta, A.T., Wong, L.H., Sere, Y.Y., Calderón Noreña, D.M.,
1251-1254, doi: 10.1038/nature07341.
Cockcroft, S., Menon, A.K., and Levine, T.P. (2015) A
29.
Peng, M., Taouatas, N., Cappadona, S., van Breukelen, B.,
new family of StART domain proteins at membrane con$
Mohammed, S., Scholten, A., and Heck, A.J.R. (2012)
tact sites has a role in ER$PM sterol transport, Elife, 4,
Protease bias in absolute protein quantitation, Nat.
doi: 10.7554/eLife.07253.
Methods, 9, 524-525, doi: 10.1038/nmeth.2031.
16.
Sandhu, J., Li, S., Fairall, L., Pfisterer, S.G., Gurnett, J.E.,
30.
Kulak, N.A., Pichler, G., Paron, I., Nagaraj, N., and
Xiao, X., Weston, T.A., Vashi, D., Ferrari, A., Orozco, J.L.,
Mann, M. (2014) Minimal, encapsulated proteomic$sam$
БИОХИМИЯ том 85 вып. 3 2020
РОЛЬ ГЕНОВ LAM В СМЕРТИ КЛЕТОК ДРОЖЖЕЙ
359
ple processing applied to copy$number estimation in
sterols on the interaction of SDS, benzalkoniumchloride,
eukaryotic cells, Nat. Methods, 11, 319-324, doi: 10.1038/
and anovel compound, Kor105, with membranes,
nmeth.2834.
Biomolecules, 9, doi: 10.3390/biom9100627.
31. Maresova, L., Muend, S., Zhang, Y.$Q., Sychrova, H., and
36. Höfken, T. (2017) Ecm22 and Upc2 regulate yeast mating
Rao, R. (2009) Membrane hyperpolarization drives cation
through control of expression of the mating genes PRM1
influx and fungicidal activity of amiodarone, J. Biol.
and PRM4, Biochem. Biophys. Res. Commun., 493,
Chem., 284, 2795-2802, doi: 10.1074/jbc.M806693200.
1485-1490, doi: 10.1016/j.bbrc.2017.10.005.
32. Zweytick, D., Hrastnik, C., Kohlwein, S.D., and Daum, G.
37. Kodedová, M., and Sychrová, H. (2015) Changes in the
(2000) Biochemical characterization and subcellular local$
sterol composition of the plasma membrane affect mem$
ization of the sterol C$24(28) reductase, erg4p, from the
brane potential, salt tolerance and the activity of multidrug
yeast Saccharomyces cerevisiae, FEBS Lett., 470, 83-87,
resistance pumps in Saccharomyces cerevisiae, PLoS One,
doi: 10.1016/s0014$5793(00)01290$4.
10, e0139306, doi: 10.1371/journal.pone.0139306.
33. Liu, G., Chen, Y., Færgeman, N.J., and Nielsen, J. (2017)
38. Hongay, C., Jia, N., Bard, M., and Winston, F. (2002)
Elimination of the last reactions in ergosterol biosynthesis
Mot3 is a transcriptional repressor of ergosterol biosynthet$
alters the resistance of Saccharomyces cerevisiae to multiple
ic genes and is required for normal vacuolar function in
stresses, FEMS Yeast Res., 17, doi: 10.1093/femsyr/fox063.
Saccharomyces cerevisiae, EMBO J., 21, 4114-4124,
34. Parks, L.W., and Casey, W.M. (1995) Physiological implica$
doi: 10.1093/emboj/cdf415.
tions of sterol biosynthesis in yeast, Annu. Rev. Microbiol.,
39. Zhang, Y.$Q., Gamarra, S., Garcia$Effron, G., Park, S.,
49, 95-116, doi: 10.1146/annurev.mi.49.100195.000523.
Perlin, D.S., and Rao, R. (2010) Requirement for ergo$
35. Jiménez$Mungu a, I., Volynsky, P.E., Batishchev, O.V.,
sterol in V$ATPase function underlies antifungal activity of
Akimov, S.A., Korshunova, G.A., Smirnova, E.A., Knorre,
azole drugs, PLoS Pathog., 6, e1000939, doi: 10.1371/
D.A., Sokolov, S.S., and Severin, F.F. (2019) Effects of
journal.ppat.1000939.
THE ROLE OF LAM GENES IN SEX PHEROMONE=INDUCED
S. cerevisiae YEAST CELL DEATH*
S. S. Sokolov1**, K. V. Galkina1, E. A. Litvinova2, D. A. Knorre1,3, and F. F. Severin1
1 Belozersky Institute of Physico Chemical Biology, Lomonosov Moscow State University,
119991 Moscow, Russia; E mail: sviatoslav.sokolov@gmail.com
2 Lomonosov Moscow State University, Faculty of Biology, 119991 Moscow, Russia
3 Sechenov First Moscow State Medical University Ministry of Health of the Russian Federation,
119992 Moscow, Russia
Received November 3, 2019
Revised January 21, 2020
Accepted January 21, 2020
Lam1$Lam4 proteins shuttle sterols between plasma membrane and endoplasmic reticulum. Disruption of their func$
tion presumably increases plasma membrane sterol content. While in mammals the homologs of Lam proteins are
responsible for the internalization of cholesterol from blood plasma, the biological role of Lam proteins in yeast
remains unclear, since the deletions of individual LAM genes do not produce pronounced phenotypes. However, the
deletion of LAM1 (YSP1) interferes with regulated cell death (RCD) of Saccharomyces cerevisiae yeast caused by sex
pheromone. Here we studied the role of LAM2 deletion in pheromone$induced RCD and the genetic interactions
between LAM2 and the genes of ergosterol biosynthesis. The deletion of LAM2 gene partially prevented the
pheromone$induced cell death in the W303 laboratory yeast strain. Additional deletions of LAM1, LAM3, and LAM4
did not increase the effect of LAM2 deletion. A mutation in the Upc2 transcription factor gene (UPC2 1), leading to
increased accumulation of sterols in cells, also inhibited pheromone$induced RCD, the effect being additive with the
effect of LAM2 deletion. On the contrary, LAM2 deletion in another yeast laboratory strain, BY4741, stimulated
pheromone$induced RCD. Next, the deletions of ergosterol biosynthesis genes ERG2 and ERG6 decreased the effect
of the LAM2 deletion. Interestingly, in Δerg4 strain (with deletion of the gene encoding Erg4 protein catalyzing the
terminal stage of ergosterol biosynthesis), the deletion of LAM2 significantly increased the proportion of dead cells
and reduced the growth rate of cells in suspension even in the absence of pheromone. Possibly, the lack of the LAM2
deletion phenotype in Δerg6 and Δerg2 strains indicates the inability of Lam2 protein to transport upstream ergosterol
biosynthesis intermediates, such as lanosterol. Together, our data suggest that the role of Lam proteins in pheromone$
induced yeast RCD is mediated by their effect on the sterol composition of plasma membrane.
Keywords: LAM genes, yeast, regulated cell death, pheromone, sterols
БИОХИМИЯ том 85 вып. 3 2020