БИОХИМИЯ, 2020, том 85, вып. 4, с. 547 - 555
УДК 577.214.6; 575.113.2
ВЫСОКИЙ УРОВЕНЬ ТРАНСКРИПЦИИ ГЕНА
НА СТАДИИ ЭМБРИОНА ПРИВОДИТ К ПОДАВЛЕНИЮ
ЕГО ГЕТЕРОХРОМАТИНОВОЙ транс ИНАКТИВАЦИИ
У ВЗРОСЛЫХ ОСОБЕЙ Drosophila melanogaster*,**
© 2020
А.А. Солодовников, В.А. Гвоздев, С.А. Лавров***
Институт молекулярной генетики РАН, 123182 Москва, Россия; электронная почта: slavrov@img.ras.ru
Поступила в редакцию 30.12.2019
После доработки 24.02.2020
Принята к публикации 24.02.2020
Перемещение гена в результате хромосомной перестройки из эухроматина к конститутивному гетерохрома
тину может сопровождаться эпигенетической инактивацией этого гена - цис инактивацией. При гетеро
хроматиновой транс инактивации происходит репрессия репортерных трансгенов в нормальной хромосо
ме под воздействием вызывающей цис инактивацию гомологичной хромосомы с перестройкой. Транс
инактивация является результатом соматического спаривания гомологов и перемещения участка нормаль
ной хромосомы в гетерохроматиновый компартмент ядра. Показано, что степень транс инактивация ре
портерного гена UAS%eGFP под влиянием перестройки In(2)A4 у взрослых мух зависит от уровня его тран
скрипции. В настоящей работе детально исследовали эпигенетическое наследование активного/репресси
рованного состояния транс инактивируемого репортерного гена при разных уровнях экспрессии путем из
мерения уровня флуоресценции eGFP в отдельных клетках мальпигиевых сосудов взрослых мух. Было об
наружено, что высокий уровень экспрессии на эмбриональной стадии препятствует транс инактивации
eGFP у взрослых особей даже при низком уровне экспрессии на постэмбриональных стадиях развития.
КЛЮЧЕВЫЕ СЛОВА: гетерохроматин, Drosophila, эффект положения, транс инактивация, экспрессия,
GAL4, GAL80ts.
DOI: 10.31857/S0320972520040077
Эффектами положения (ЭП) называются
стояния (сотни т.п.н.-миллион п.н.). При гете
нарушения экспрессии генов, происходящие
рохроматинизации в эухроматиновых районах
при изменении локализации генов в геноме без
появляются специфические для гетерохромати
нарушения их нуклеотидной последовательнос
на белки (HP1a и другие) и модификации гисто
ти. Частным случаем ЭП является гетерохрома
нов (Н3К9me2/3) [1-3], что и приводит к нару
тиновый эффект положения, при котором
шениям работы генов. При гетерохроматиновом
экспрессия эухроматиновых генов подавляется
ЭП гены могут инактивироваться лишь в части
при их перемещении к конститутивному гете
клеток на стадии эмбриона, при этом актив
рохроматину. В результате хромосомной пере
ное/репрессированное состояние способно
стройки эухроматин вблизи новых эу гетерох
эпигенетически наследоваться в клеточной ли
роматиновых границ приобретает свойства гете
нии, давая в результате мозаичную экспрессию в
рохроматина (гетерохроматинизация), этот про
пределах одной ткани.
цесс может распространяться на большие рас
В некоторых случаях перестроенная хромо
сома с новыми эу гетерохроматиновыми грани
Принятые сокращения: UAS%eGFP - ген eGFP под
цами способна инактивировать гены нормаль
контролем регуляторного элемента UAS, А4 - хромосомная
ной неперестроенной хромосомы [4-6]. Это яв
инверсия In(2)A4, ЭП - эффекты положения.
ление, транс инактивация, не может быть объ
* Приложение к статье на английском языке опубликова
яснено распространением гетерохроматина
но на сайте журнала «Biochemistry» (Moscow) и на сайте из
вглубь эухроматина от новой границы и связа
но, по современным представлениям, с переме
том 85, вып. 4, 2020.
щением эухроматинового района в гетерохрома
** Первоначально английский вариант рукописи опубли
тиновый компартмент ядра при соматическом
msu.ru/biokhimiya, в рубрике «Papers in Press», BM19 351,
спаривании нормальной хромосомы и гомоло
13.04.2020.
гичной хромосомы с перестройкой. При транс
*** Адресат для корреспонденции.
инактивации происходит формирование гете
547
548
СОЛОДОВНИКОВ и др.
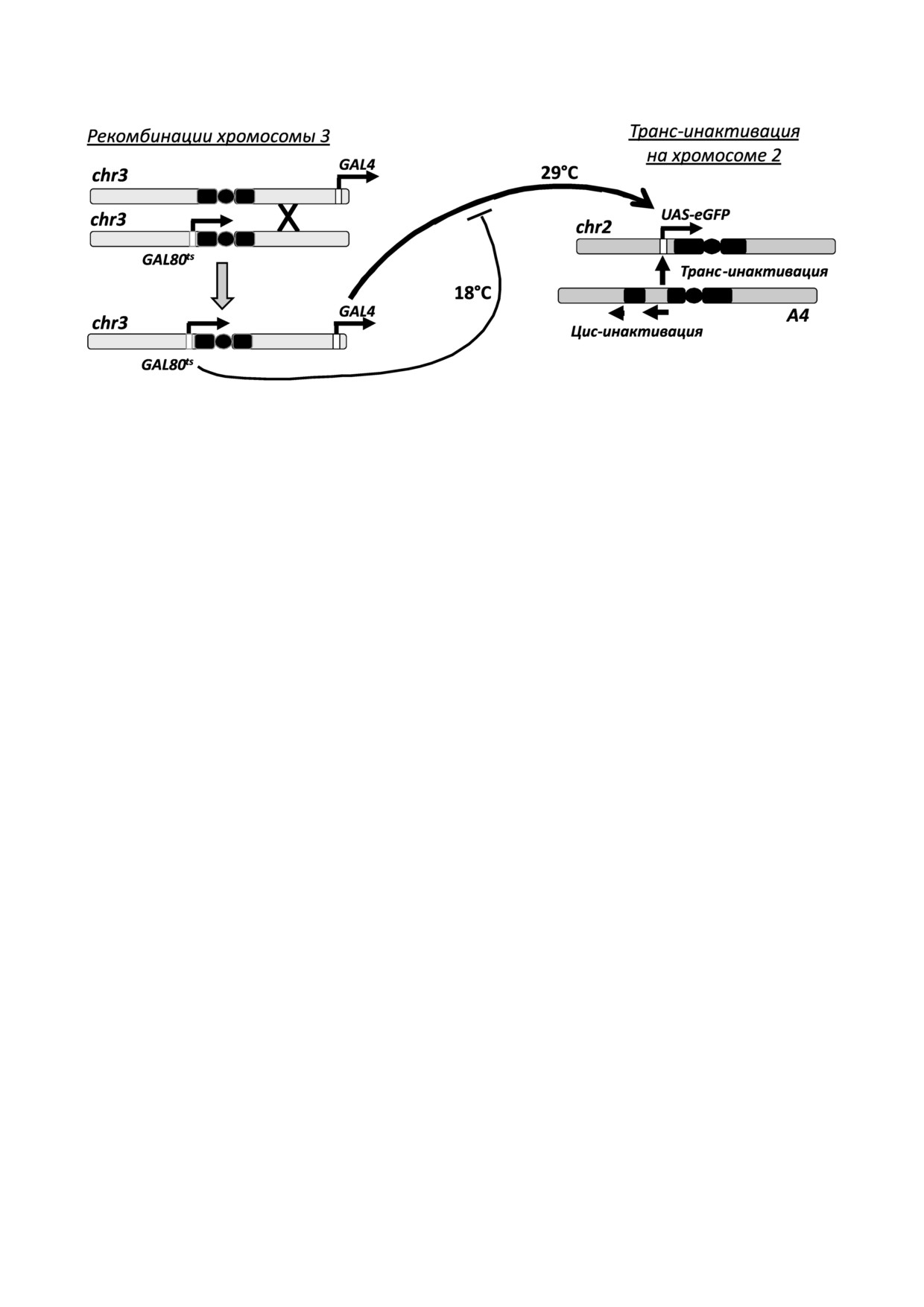
Рис. 1. Система контроля экспрессии репортерного гена. Хромосома 2 с перестройкой A4 вызывает транс инактивацию
репортера UAS%eGFP в гомологичной нормальной хромосоме. Хромосома 3 получена путем рекомбинации и несет источ
ники GAL4 (tubGal4) и GAL80ts (tubGal80ts). Повышение температуры приводит к разрушению репрессора GAL80ts, повы
шению содержания активного GAL4 и усилению транскрипции UAS eGFP
рохроматина de novo в нормальной хромосоме
Center», США), номера линий соответствуют
[7-10].
идентификаторам в коллекции: линия 7018 (w*;
Хромосомная инверсия In(2)A4 (далее А4,
snaSco/CyO; P{tubP%GAL80ts}7) содержит на хро
рис. 1) вызывает цис инактивацию генов рядом с
мосоме 3 источник белка GAL80ts под промото
новыми эу гетерохроматиновыми границами и
ром alphaTub84B [13]; линия 5138 (y1 w*; P{tubP%
транс инактивацию репортерных конструкций
GAL4}LL7/TM3 Sb1 Ser1) содержит источник
в гомологичной хромосоме [6, 9-12]. С целью
GAL4 под промотором alphaTub84B [14].
изучения наследуемости эпигенетических состо
Линия w*; UAS%eGFP/CyO; P{tubP%GAL4}LL7;
яний транс инактивируемого гена в процессе
P{tubP%GAL80ts}7/ TM3 Sb1 Ser1 содержит репор
развития, были получены мухи, содержащие ре
терный ген UAS%eGFP на хромосоме 2 [11] и ре
портерный ген UAS%eGFP в нормальной хромо
гуляторные гены GAL4 и GAL80ts на хромосоме 3.
соме 2, инверсию A4 в гомологичной хромосоме
Хромосома 3 с GAL4 и GAL80ts получена путем
и комбинацию трансактиватора GAL4 и термо
рекомбинации между линиями 7018 (w*; snaSco/
чувствительной формы репрессора GAL80
CyO; P{tubP%GAL80ts}7) и 5138 (y1 w*; P{tubP%
(GAL80ts) в хромосоме 3. Комбинация GAL4 и
GAL4}LL7/TM3 Sb1 Ser1). Описание скрещива
GAL80ts позволяет регулировать экспрессию ре
ний приведено в Приложении к статье.
портерного гена путем изменения температуры
Влияние активации eGFP на разных стадиях
развития мух. Генетическая система была ис
развития на транс инактивацию, регистрируемую
пользована для исследования зависимости сте
у взрослых мух. Для исследования зависимости
пени транс инактивации репортерного гена у
степени транс инактивации eGFP от уровня его
взрослых мух от уровня его экспрессии на раз
экспрессии на разных стадиях развития было
ных стадиях развития. Показано, что высокий
проведено скрещивание w*; UAS%eGFP; GAL80ts
уровень экспрессии на эмбриональной стадии
GAL4/TM3 Sb1 Ser1 × w*; In(2)A4/CyO; +/+. В по
приводит к эпигенетически наследуемой в ходе
томстве от скрещивания появляются мухи четы
дальнейшего развития устойчивости репортер
рех генотипов. Мухи w*; UAS%eGFP/In(2)A4;
ного гена к транс инактивации.
GAL80ts GAL4/+ (опыт) и w*; UAS%eGFP/CyO;
GAL80ts GAL4/+ (контроль) содержат GAL4 и
GAL80ts, и при повышении температуры у них
МАТЕРИАЛЫ И МЕТОДЫ
повышается экспрессия eGFP. В опыте UAS%
eGFP подвергается транс инактивации под вли
Использованные в работе линии мух. Линия
янием In(2)A4, в контроле транс инактивация
w67c23; In(2)A4/CyO, содержащая вызывающую
отсутствует. Температуру развития особей варь
транс инактивацию хромосомную перестройку
ировали, согласно представленным схемам (таб
In(2)A4 (A4), получена и исследуется в ОМГК
лица и рис. 2, а).
ИМГ РАН [6].
Флуоресценцию eGFP в отдельных клетках
Следующие линии получены из коллекции
мальпигиевых сосудов анализировали у двух
Bloomington
(«Bloomington Drosophila Stock
дневных взрослых самок. Причиной выбора
БИОХИМИЯ том 85 вып. 4 2020
ЭФФЕКТ ПОЛОЖЕНИЯ И ТРАНСКРИПЦИЯ
549
мальпигиевых сосудов, как объекта исследова
активатор GAL4, и eGFP экспрессируется на
ния, было то, что в мальпигиевых сосудах моза
уровне фона. Сравнение экспрессии eGFP при
ичный ЭП был хорошо выражен, а морфология
транс инактивации (UAS%eGFP/In(2)A4) и без
ткани (крупные изолированные клетки, форми
нее (UAS%eGFP/CyO) при разных температурах
рующие однослойные трубки) существенно об
позволяет оценить влияние температуры непо
легчала количественные измерения.
средственно на транс инактивацию.
У мух w*; UAS%eGFP/In(2)A4; TM3 Sb1 Ser1/+
Измерения уровня флуоресценции eGFP в
и w*; UAS%eGFP/CyO; TM3 Sb1 Ser1/+ отсутствует клетках мальпигиевых сосудов. Изображения
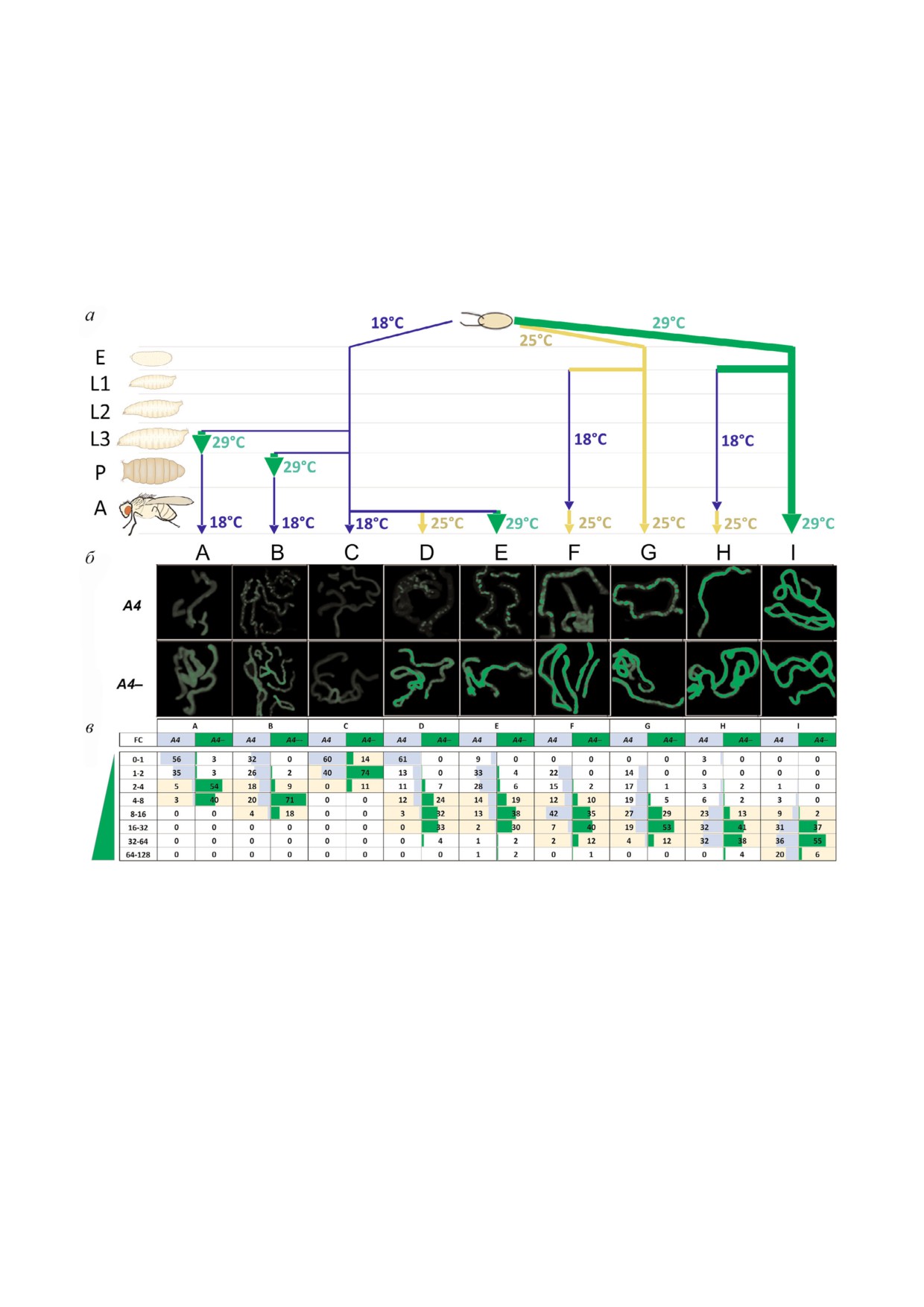
Рис. 2. Транс инактивация репортерного гена UAS eGFP у взрослых мух после модуляции его экспрессии на разных ста
диях развития. а - Схема вариантов температурных воздействий (от A до I) (cм. также таблицу) в ходе развития мух
eGFP/A4; GAL80ts GAL4/+ (A4, транс инактивация) и eGFP/CyO; GAL80ts GAL4/+ (A4-, контроль). E - эмбрионы, L1-L3 -
личинки возрастов 1-3, P - куколки, A - взрослые мухи). б - Флуоресценция eGFP в мальпигиевых сосудах мух с хро
мосомой A4 и без нее (A4-, контроль). В вариантах B и D-G (A4) заметна существенная неравномерность флуоресценции
от клетки к клетке, отражающая гетерохроматиновую транс инактивацию. в - Распределение интенсивностей флуорес
ценции eGFP в клетках мальпигиевых сосудов у мух с транс инактивацией (A4) и в контроле (A4-). Измеряли интенсив
ность флуоресценции в каждой клетке мальпигиевого сосуда и нормировали на модальный (встречающийся чаще всего)
уровень флуоресценции в мальпигиевых сосудах мух A4- при 18 °С, принятый за единицу (столбец С, A4-). Полученные
значения (FC, fold change) показывают во сколько раз уровень флуоресценции выше принятого за единицу. Весь диапа
зон уровней флуоресценции был разбит на поддиапазоны (указаны в крайнем левом столбце), и для каждого варианта об
работки подсчитан процент клеток, попадающий в поддиапазон (цифры в ячейках). Желтым цветом выделен диапазон
флуоресценции, в который попадает большинство клеток мальпигиевых сосудов у мух без транс инактивации (A4-), т.е.
нормальное значение флуоресценции при данном варианте температурной обработки. Величины достоверности разли
чий между уровнями флуоресценции в каждом из вариантов (Р величины) представлены в табл. S1 в Приложении, зна
чения погрешностей - в табл. S2 и на рис. S1 в Приложении.
journal/biokhsm/
БИОХИМИЯ том 85 вып. 4 2020
550
СОЛОДОВНИКОВ и др.
Измерение температуры развития, Т °C
Стадия
Вариант
Эмбрион
Личинка
Личинка перед
Куколка
Куколка 2 дня →
Взрослые
Взрослые
0-24 ч
L1-L3
окукливанием 24 ч
0-24 ч
вылупление
2 дня
48-72 ч
A
18
18
29
18
18
18
18
B
18
18
18
29
18
18
18
C
18
18
18
18
18
18
18
D
18
18
18
18
18
18
25
E
18
18
18
18
18
18
29
F
25
18
18
18
18
18
25
G
25
25
25
25
25
25
25
H
29
18
18
18
18
18
25
I
29
29
29
29
29
29
29
Использованные варианты изменений температуры на разных стадиях развития особей (к рис. 1). Пример (вариант B):
развитие при 18 °С до окукливания, куколки развивались сутки при 29 °С, далее развитие продолжалось при 18 °С до
взрослых двухдневных мух, у которых в клетках мальпигиевых сосудов оценивали уровни флуоресценции. L1-L3 - пери
од личиночного развития (личинки первого, второго и третьего возраста).
мальпигиевых сосудов (рис. 2, б) были получены
но ~100 записей измерений флуоресценции
с использованием флуоресцентного микроскопа
eGFP, соответствующих отдельным клеткам.
Leica DM6000 («Leica Microsystems», ФРГ), ос
Полученные результаты использовали для по
нащенного камерой ORCA ER («Hamamatsu
строения гистограмм распределений интенсив
Photonics», Япония) и набором фильтров для де
ности флуоресценции eGFP и подсчета статис
текции флуоресценции. Мальпигиевые сосуды
извлекали в PBT (1× PBS с 0,1% (v/v) Tween 20),
project.org/). Результаты получены для двух не
инкубировали в PBT с DAPI 15 мин при комнат
зависимых скрещиваний, для каждого генотипа
ной температуре и фотографировали. Все изобра
и варианта температурной обработки анализи
жения получены при увеличении 5× и при оди
ровали 5-10 мух.
наковых параметрах съемки (экспозиция, сте
Количественная ПЦР. Выделение РНК из
пень усиления, разрешение), экспозицию под
взрослых мух w*; UAS%eGFP/In(2)A4; GAL80ts
бирали, избегая насыщения изображения на на
GAL4/+ (опыт), w*; UAS%eGFP/CyO; GAL80ts
иболее ярких образцах. Съемку проводили в ка
GAL4/+ (контроль), w*; UAS%eGFP/In(2)A4 (ис
налах для GFP (488 нм) и DAPI (405 нм), изо
следование влияния температуры выращивания
бражения сохраняли в виде 16bit TIFF и обраба
непосредственно на транс инактивацию), об
тывали в программе ImageJ, загружая изображе
ратную транскрипцию и количественную ПЦР с
ния сосуда в каналах DAPI и GFP. Изображение
праймерами к гену eGFP проводили, как описа
ядер (DAPI) использовали для сегментации
но ранее [11]. Количество мРНК eGFP нормиро
изображения (Инструмент Threshold). Получен
вали на количество мРНК гена домашнего хо
ный набор областей интереса (ROI) переносили
зяйства Rpl32.
на изображение с eGFP, для вычитания фона
выделяли область, не содержащую объектов.
Для набора областей измеряли площадь, мо
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
дальную величину плотности изображения (зна
чение, наблюдаемое в большинстве клеток), а
Использованная в работе репортерная
также стандартное отклонение интенсивности
конструкция не содержит каких либо функцио
пикселей в области и ее сферичность (Инстру
нальных элементов кроме UAS%eGFP, который
мент MultiMeasure в ImageJ). Полученные дан
без трансактиватора GAL4 экспрессируется на
ные переносили в Excel, где проводили филь
низком уровне на всех стадиях развития мухи.
трацию записей по площади (30-300 пикселей),
Экспрессию UAS%eGFP можно менять при помо
величине стандартного отклонения (не более
щи активатора транскрипции GAL4 и его тер
интенсивности области) и сферичности (>0,2).
мочувствительного репрессора GAL80ts [11].
В итоге для каждого изображения было получе
Трансактиватор GAL4 и репрессор GAL80ts на
БИОХИМИЯ том 85 вып. 4 2020
ЭФФЕКТ ПОЛОЖЕНИЯ И ТРАНСКРИПЦИЯ
551
ходятся под контролем промоторов повсемест
стадиях развития, начиная с эмбриональной.
но активного альфа тубулина, поэтому экспрес
Степень транс инактивации репортера оцени
сируются на сходном высоком уровне на всех
вали по уровню флуоресценции eGFP в отдель
стадиях развития. Репрессор GAL80ts блокирует
ных клетках мальпигиевых сосудов взрослых
активаторный домен GAL4 при 18 °С, но разру
мух при изменении уровня экспрессии репорте
шается при повышении температуры, освобож
ра на разных стадиях развития. Общая схема ва
дая активатор GAL4 и приводя к увеличению
риантов температурных воздействий (варианты
экспрессии eGFP [13]. Понижение температуры
A-I) в ходе развития представлена на рис. 2, a и
приводит к накоплению активной формы
в таблице, а визуальная и количественная оцен
GAL80ts, выключению GAL4 и понижению
ки интенсивности флуоресценции представле
уровня экспрессии репортера. Процесс актива
ны, соответственно, на рис. 2, б и в. Значения P
ции репортерного гена не требует запуска
величин при попарном сравнении распределе
экспрессии GAL4, а определяется межбелковы
ний интенсивностей флуоресценции и стандарт
ми взаимодействиями GAL4 и GAL80ts, уже
ные отклонения уровней флуоресценции между
присутствующих в клетках. Таким образом,
экспериментами приведены в табл. S1 и S2 и на
комбинация воздействия GAL4 и GAL80ts поз
рис. S2 в Приложении.
воляет предсказуемо регулировать уровень
Для количественной оценки степени транс
экспрессии гена путем изменения температуры
инактивации уровни флуоресценции в отдель
[11]. Использование eGFP в качестве репортер
ных клетках мальпигиевых сосудов в опыте (му
ного гена позволяет наблюдать эффект положе
хи UAS%eGFP/A4, A4 на рис. 2, б, в) и контроле
ния in vivo, а измерение относительного уровня
(мухи UAS%eGFP/CyO, A4- на рис. 2, б, в) были
флуоресценции eGFP является прямым методом
измерены и нормированы на принятый за еди
оценки уровня экспрессии и в целом коррели
ницу модальный (встречающийся чаще всего)
рует с количеством его мРНК (рис. S1 в Прило
уровень флуоресценции в мальпигиевых сосудах
жении). Проведенные ранее измерения количе
контрольных мух, развивавшихся при 18 °С (ва
ства мРНК eGFP при помощи ПЦР показали,
риант С, A4-). Полученная величина показыва
что в образцах РНК из целых взрослых мух
ет, во сколько раз уровень флуоресценции в об
экспрессия репортерного гена при 18 °С возрас
разце больше принятого за единицу. На основа
тает в ~3 раза, при 25 °С - в ~50 раз и при 29 °С -
нии полученных данных для всех вариантов
в ~100 раз относительно мух без источника
температурных обработок для опыта и контроля
GAL4 (уровень фона) [11]. Уровни активации
были построены гистограммы, отражающие
транскрипции eGFP при разных температурах у
процент клеток образца, попадающих в задан
взрослых мух и на стадии эмбрионов сходны
ные диапазоны интенсивности флуоресценции.
(неопубликованные данные). Экспрессия eGFP
К примеру, в случае D (рис. 2, в) контрольные и
при 18 °С обозначается далее как слабая (~3 ра
опытные мухи развивались до взрослой стадии
за), при 25 °С - как умеренная (~50 раз) и при
при 18 °С (слабая активация eGFP), затем у
29 °С - как сильная (~100 раз).
взрослых особей репортер был умеренно акти
Использованная в работе генетическая сис
вирован при 25 °С перед измерением флуорес
тема с рекомбинантной хромосомой 3, несущей
ценции. При этом, при транс инактивации
оба регуляторных трансгена, представлена на
(D, столбец А4) 61% клеток имеет уровень флуо
рис. 1. У мух генотипа UAS%eGFP/A4; GAL4
ресценции меньше или равный принятому за
GAL80ts/+ (eGFP/A4) репортерный ген транс
единицу, тогда как в контроле без транс инак
инактивируется под влиянием инверсии A4, а
тивации (D, столбец А4%) 89% клеток показыва
мухи UAS%eGFP/CyO; GAL4 GAL80ts/+ (eGFP/+),
ют уровень экспрессии, превышающий приня
лишенные хромосомы A4, служат контролем. В
тый за единицу, в 4-32 раза. Распределения ин
мальпигиевых сосудах взрослых мух транс
тенсивностей флуоресценции в опыте и контро
инактивация проявляется в появлении клеток
ле практически не перекрываются, это пример
со сниженным уровнем или отсутствием флуо
сильной транс инактивации под влиянием ин
ресценции eGFP (рис. 2, б, варианты B, D-G).
версии A4.
Представляло интерес оценить устойчивость
Анализ полученных данных позволяет сде
эпигенетического наследования состояния
лать следующие заключения. Низкий уровень
экспрессии гена, испытывающего гетерохрома
экспрессии репортера на ранних стадиях разви
тинизацию, индуцированную гомологичной пе
тия сопровождается его сильной транс инакти
рестроенной хромосомой А4. Мы исследовали,
вацией у взрослых мух (варианты A-D), во всех
как степень транс инактивации у взрослых мух
случаях обнаруживается популяция клеток с
может зависеть от уровня экспрессии репортера
флуоресценцией eGFP на уровне фона. При
(слабая, умеренная и сильная) на более ранних
умеренной и сильной активации экспрессии на
БИОХИМИЯ том 85 вып. 4 2020
552
СОЛОДОВНИКОВ и др.
стадии эмбриона у взрослых мух отсутствуют
дим высокий уровень экспрессии именно на
клетки с флуоресценцией eGFP на уровне фона
эмбриональной стадии развития, тогда как на
(варианты F-I). При умеренной экспрессии в
постэмбриональных стадиях активная тран
течение всего развития (25 °С, вариант G) ~45%
скрипция репортера необязательна.
клеток показывают транс инактивацию eGFP,
Хорошо известно, что степень выраженнос
их уровень флуоресценции ниже минимального
ти гетерохроматинового эффекта положения,
уровня флуоресценции в контроле. Сильная
как правило, снижается при повышении темпе
экспрессия eGFP в течение всего развития пре
ратуры в процессе развития [1, 3]. Поскольку в
пятствует транс инактивации в основной массе
исследуемой нами системе уровень экспрессии
клеток, распределение уровней флуоресценции
репортерного гена регулировали путем измене
в опыте и контроле достоверно не отличаются
ния температуры, то такое воздействие могло
(вариант I). Однако ~10% клеток содержат
влиять на степень транс инактивации непосред
транс инактивированный репортер и, что не
ственно, а не только через активацию экспрес
ожиданно, в ~10% клеток мух UAS%eGFP/A4 уро
сии репортера с участием GAL4. Однако оценка
вень флуоресценции превышает максимальный
с помощью количественной ПЦР не выявила
уровень флуоресценции в контроле. Репресси
заметных различий в содержании мРНК eGFP у
рованное состояние репортерного гена, устано
мух UAS%eGFP/A4, лишенных источника GAL4
вившееся на ранних стадиях развития, может
(UAS%eGFP/A4; +/+), развивавшихся при 18 °С и
быть преодолено путем сильной активации
29 °С (рис. S3 в Приложении). Также известно,
транскрипции. Так, в варианте D eGFP слабо
что при ЭП зависимость репрессии от темпера
экспрессируется на протяжении всего развития
туры развития особи варьирует в разных пере
(18 °С) и затем умеренно активируется у взрос
стройках, иногда влияние температуры отсут
лых мух (25 °С), тогда как в варианте Е у взрос
ствует [3, 15]. Представляется более вероятным,
лых мух репортер активируется сильно (29 °С).
что наблюдаемое нами влияние уровня экспрес
В итоге в случае E обнаруживается лишь девять
сии репортерного гена на степень его транс
процентов клеток с полной репрессией репорте
инактивации обусловлено именно активацией
ра по сравнению с вариантом D, где число таких
транскрипции с участием GAL4.
клеток достигает 60%. Иными словами, при
сильной активации транскрипции у взрослых
мух до 80% клеток с транс инактивированным
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
eGFP начинают его активно экспрессировать
(рис. 2, в, вариант E). При этом уровень флуо
Исследования репрессии генов при гетеро
ресценции в случае E у мух с транс инактиваци
хроматиновом эффекте положения в ходе инди
ей был в целом ниже контрольного (А4-).
видуального развития были ранее проведены в
Сравнение вариантов развития D, F и H по
ряде работ [16-19]. В двух из них [18, 19] изучали
казало, что активация репортера на стадии эм
поведение репортерной конструкции, содержа
брионов приводит к формированию устойчиво
щей UAS%eGFP (а также другие функциональные
го в развитии активного эпигенетического имп
элементы), под влиянием гетерохроматина, при
ринтинга в присутствии хромосомы A4. На ста
чем в работе Sage et al. [19] исследовали именно
дии эмбриона в варианте D eGFP активировался
транс действующий эффект положения.
слабо (18 °С), в варианте F - умеренно (25 °С) и
В работе Ahmad et al. [18] была использована
в варианте H - сильно (29 °С). Дальнейшее раз
репортерная конструкция, содержащая гены
витие во всех случаях проходило при 18 °С (сла
GFP (под контролем UAS) и mini%white.
бая экспрессия репортера) вплоть до вылупле
Конструкция подвергалась ЭП, а экспрессия
ния, после чего экспрессию репортера умеренно
GFP могла быть запущена на разных стадиях
активировали
(25
°С) для анализа степени
развития при помощи источников GAL4 под
транс инактивации. Оказалось, что доли клеток
ткане или стадия специфичным промотором.
мальпигиевых сосудов, в которых eGFP был реп
Анализировались глазные имагинальные диски
рессирован, существенно различались между
у личинок и глаза взрослых мух. Было показано,
вариантами. В случае D наблюдается сильный
что сильная активация экспрессии репортерно
эффект положения (85% клеток c нарушенной
го гена GFP на стадии эмбриона (драйвер GAL4
активацией репортера), в случае F репрессию
под актиновым промотором) приводит к суп
демонстрируют ~ 40% клеток, а в случае H -
рессии ЭП, однако при активации экспрессии в
только 12% клеток. Cравнение вариантов D, F и
меньшей степени (GAL4 под контролем слабо
H показывает, что для поддержания активного
го, но активного в большинстве тканей и на всех
состояния хроматина (устойчивости к транс
стадиях развития промотора гена armadillo, arm)
инактивации) до стадии взрослых мух необхо
эффект положения сохраняется. ЭП наблюдали
БИОХИМИЯ том 85 вып. 4 2020
ЭФФЕКТ ПОЛОЖЕНИЯ И ТРАНСКРИПЦИЯ
553
также при активации транскрипции на пост
дах взрослых мух). Мальпигиевые сосуды фор
эмбриональных стадиях развития. Дерепрессию
мируются из эктодермы и мезодермы на стадиях
репортерного гена (появление экспрессирую
5-13 развития эмбриона, клеточные деления за
щих GFP клеток) наблюдали в процессе диффе
канчиваются на стадии 13, далее число клеток
ренцировки имагинальных дисков у личинок,
не меняется. Мальпигиевые сосуды не подвер
причем дерепрессия происходила как в деля
гаются гистолизу в ходе метаморфоза [20]. По
щихся, так и в уже дифференцированных клет
литенные хромосомы в ядрах клеток мальпигие
ках. Это наблюдение означает, что для реактива
вых сосудов в ходе развития проходят до 8 цик
ции репрессированного при ЭП гена не требует
лов эндорепликации ДНК без цитокинеза и мо
ся прохождение клеточного цикла.
гут содержать до 256 ти сестринских хроматид
В работе Sage et al. [19] была исследована
[21]. Мальпигиевые сосуды состоят из терми
транс инактивация репортерной конструкции,
нально дифференцированных клеток и схожи с
содержащей UAS%eGFP и mini%white, под влияни
клетками слюнных желез, но в отличие от пос
ем аллеля BwD на соседней хромосоме. BwD
ледних сохранены у взрослых мух.
представляет собой вставку сателлитной ДНК в
Согласно нашим данным, транс инактива
ген Brown. Экспрессию UAS%eGFP активировали
ция репортерного гена под влиянием инверсии
при помощи источников GAL4 под контролем
A4 демонстрирует свойства, обнаруженные в
промоторов разной силы и специфичности (в
упомянутых выше более ранних работах и отра
том числе актина и arm, как и в работе Ahmad et
жающие, видимо, характерные черты гетерохро
al. [18]), а затем анализировали степень транс
матиновой репрессии в целом. Так, на стадии
инактивации репортера в тканях личинок 3 воз
эмбриона экспрессия UAS%eGFP на высоком
раста и у взрослых мух. Транс инактивация UAS%
уровне супрессирует наблюдаемую у взрослых
eGFP в данной работе демонстрировала замет
мух транс инактивацию. При 100× повышении
ную тканеспецифичность и наблюдалась в тех
экспрессии репортера на стадии эмбриона
случаях, когда экспрессия репортерного гена
транс инактивация у взрослых мух достоверно
была на низком уровне на стадиях развития,
не наблюдалась, но при активации в 30-50 раз
предшествующих исследованным (личинки 3
репрессия была хорошо выражена. При актива
возраста и взрослые мухи).
ции в 100 раз уровень экспрессии eGFP состав
Анализ полученных в ранних работах дан
лял ~17% от уровня экспрессии гена домашнего
ных позволяет отметить следующие общие осо
хозяйства Rpl32 [11]. Это наблюдение позволяет
бенности проявления инактивации генов, об
предполагать, что гены, экспрессирующиеся на
щие для цис и транс действующего гетерохро
сходных или более высоких уровнях на стадии
матинового ЭП в процессе развития. Репрессия
эмбриона, не будут подвергаться гетерохрома
при ЭП начинается со стадии гаструляции у
тиновой репрессии. В двух работах, в которых
эмбрионов. Если ген экспрессируется на высо
исследовали уровень транскрипции генов при
ком уровне, начиная со стадии эмбриона (высо
цис действующем ЭП [9, 22], было обнаружено,
кий уровень обеспечивает, к примеру, промотор
что гетерохроматиновой репрессии подвергает
актина), то такой ген устойчив к ЭП. Если ген
ся лишь несколько генов из десятков, находя
экспрессируется, начиная со стадии эмбриона
щихся в области распространения эффекта по
на низком/среднем уровне (уровень промотора
ложения. Достаточно высокий уровень тран
arm), то он может быть репрессирован. Также
скрипции этих генов на эмбриональной стадии
репрессируются гены, экспрессия которых
может служить объяснением этого наблюдения.
включается на постэмбриональных стадиях раз
В случаях транс инактивации эпигенетичес
вития. Ген, репрессированный на ранних этапах
кий эффект ассоциируется с изменениями внут
развития, может реактивироваться на более
риядерной локализации репортерного гена. Ра
поздних стадиях (обычно в ходе метаморфоза).
нее было показано [7-9, 23], что репрессия со
Реактивация не требует прохождения цикла
провождается переносом гена в гетерохромати
клеточного деления [18, 19].
новый компартмент ядра, и наоборот, сохране
Используемая нами система позволяла
ние активного состояния приводит к локализа
предсказуемо и количественно регулировать
ции гена в эухроматине. Локальная активация
уровень экспрессии репортера [11], тогда как
UAS%eGFP может приводить к нарушению спари
источники GAL4, применяемые для активации
вания транс инактивируемого района нормаль
транскрипции в ранних работах, имели лишь
ной хромосомы с инверсией и к переносу облас
качественно охарактеризованные профили
ти расположения репортерного гена в эухрома
экспрессии в ходе развития в разных типах тка
тиновый компартмент ядра. Представляет инте
ней. Кроме того, транс инактивацию детекти
рес при помощи FISH гибридизации сравнить
ровали в одном типе ткани (мальпигиевых сосу
локализацию малоактивного и активированного
7 БИОХИМИЯ том 85 вып. 4 2020
554
СОЛОДОВНИКОВ и др.
в 100 раз репортера в отдельных клетках мальпи
хроматиновые гены, то есть включают как мар
гиевых сосудов политенных хромосом, демон
керы гетерохроматина (НР1a и Н3K9me2/3), так
стрирующих разную степень транс инактива
и маркеры активной транскрипции, например,
ции.
H3K4me3 [24].
Мы обнаружили, что установившуюся на
В настоящей работе мы продемонстрирова
стадии эмбриона транс инактивацию репорте
ли возможность исследования эпигенетически
ра можно преодолеть сильной активацией тран
наследуемых изменений экспрессии генов при
скрипции у молодых взрослых мух (рис. 2, б, ва
гетерохроматиновой транс инактивации в гене
риант E, A4). Температурный уровень для акти
тической системе, в которой осуществляли
вации экспрессии совпадает с тем, который не
контролируемое изменение экспрессии гена
обходим для предотвращения транс инактива
репортера в процессе развития и следили за на
ции на стадии эмбриона. По видимому, дости
следованием этого состояния у взрослых осо
гается пороговый уровень экспрессии репорте
бей, выключая индуктор транскрипции. Обна
ра, при котором гетерохроматинизированное
ружено, что сильная активация экспрессии гена
состояние в районе гена становится нестабиль
на стадии эмбриона приводит к переключению
ным, не формируется у эмбрионов и разрушает
репрессированного состояния на активное у
ся у взрослых мух. Так как мальпигиевые сосуды
взрослых мух. Предложенная в работе генети
представляют собой неделящуюся терминально
ческая система может быть использована для
дифференцированную ткань, то изменение ор
дальнейших исследований гетерохроматиново
ганизации хроматина/положения в ядре репор
го эффекта положения, включая выявление но
тера при реактивации достигается без прохож
вых генов, вовлеченных в процессы эпигенети
дения клеточного деления и в политенизиро
ческого наследования, а также механизмов уста
ванных ядрах.
новления гетерохроматиновой репрессии.
При сильной активации репортера на протя
жении всего развития (29 °С) на фоне инверсии
A4 до ~10% клеток мальпигиевых сосудов демон
Финансирование. Работа выполнена при фи
стрируют уровень флуоресценции более высо
нансовой поддержке Российского научного
кий, чем в контроле (рис. 2, вариант I). Это оз
фонда (грант № 19 74 20178) в части синтеза ге
начает, что гетерохроматиновое окружение мо
нотипов мух с регулируемой транс инактиваци
жет поддерживать высокий уровень экспрессии
ей с дальнейшим использованием в исследова
и, более того, усиливать транскрипцию. Пред
ниях и Российского фонда фундаментальных
ставляется, что гетерохроматин не является не
исследований (гранты № 17 04 01984 А и № 17
специфическим репрессором эухроматиновых
00 00282 КОМФИ) в части молекулярно биоло
генов - часть из них может вести себя аналогич
гических экспериментов и биоинформационно
но генам гетерохроматина, демонстрирующим
го анализа полученных данных.
высокий уровень транскрипции именно в гете
Конфликт интересов. Авторы заявляют, что у
рохроматиновом окружении. Можно предполо
них нет конфликта интересов.
жить, что гиперактивированные репортерные
Соблюдение этических норм. Настоящая
гены локализуются в гетерохроматиновом ком
статья не содержит описания выполненных ав
партменте ядра, а по организации хроматина
торами исследований с участием людей или ис
похожи на сильно экспрессирующиеся гетеро
пользованием животных в качестве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Elgin, S. C., and Reuter, G. (2013) Position effect variega
inactivation of Drosophila white transgenes, Genetics, 147,
tion, heterochromatin formation, and gene silencing in
671 677.
Drosophila, Cold Spring Harb. Perspect. Biol., 5, a017780,
6.
Абрамов Ю. А., Кибанов М. В., Гвоздев В. А., Лавров С. А.
doi: 10.1101/cshperspect.a017780.
(2011) Генетический и молекулярный анализы явле
2.
Weiler, K. S., and Wakimoto, B. T. (1995) Heterochromatin
ния транс инактивации генов у Drosophila
and gene expression in Drosophila, Annu. Rev. Genet., 29,
melanogaster, обусловленного эу гетерохроматиновой
577 605, doi: 10.1146/annurev.ge.29.120195.003045.
перестройкой гомологичной хромосомы, Доклады
3.
Spofford, J. B., (1976) Position effect variegation in
Академии Наук, 437, 261 265.
Drosophila, in The Genetics and Biology of Drosophila,
7.
Thakar, R., and Csink, A. K. (2005) Changing chromatin
Academic press. p. 955.
dynamics and nuclear organization during differentiation
4.
Csink, A. K., Bounoutas, A., Griffith, M. L., Sabl, J. F., and
in Drosophila larval tissue, J. Cell Sci., 118, 951 960,
Sage, B. T. (2002) Differential gene silencing by trans hete
doi: 10.1242/jcs.01684.
rochromatin in Drosophila melanogaster, Genetics, 160, 257 269.
8.
Thakar, R., Gordon, G. and Csink, A. K. (2006) Dynamics and
5.
Martin Morris, L. E., Csink, A. K., Dorer, D. R., Talbert,
anchoring of heterochromatic loci during development, J. Cell
P. B., and Henikoff, S. (1997) Heterochromatic trans
Sci., 119, 4165 4175, doi: 10.1242/jcs.03183.
БИОХИМИЯ том 85 вып. 4 2020
ЭФФЕКТ ПОЛОЖЕНИЯ И ТРАНСКРИПЦИЯ
555
9.
Abramov, Y. A., Shatskikh, A. S., Maksimenko, O. G.,
17. Weiler, K. S., and Wakimoto, B. T. (1998) Chromosome
Bonaccorsi, S., Gvozdev, V. A., and Lavrov, S. A. (2016)
rearrangements induce both variegated and reduced, uni
The differences between cis and trans gene inactivation
form expression of heterochromatic genes in a develop
caused by heterochromatin in Drosophila, Genetics, 202,
ment specific manner, Genetics, 149, 1451 1464.
93 106, doi: 10.1534/genetics.115.181693.
18. Ahmad, K., and Henikoff, S. (2001) Modulation of a tran
10.
Shatskikh, A. S., Abramov, Y. A. and Lavrov, S. A. (2017)
scription factor counteracts heterochromatic gene silenc
Trans inactivation: repression in a wrong place, Fly
ing in Drosophila, Cell, 104, 839 847.
(Austin), 11, 96 103, doi: 10.1080/19336934.2016.1225634.
19. Sage, B. T., Wu, M. D. and Csink, A. K. (2008) Interplay
11.
Shatskikh, A. S., Olenkina, O. M., Solodovnikov, A. A.,
of developmentally regulated gene expression and hete
and Lavrov, S. A. (2018) Regulated gene expression as a
rochromatic silencing in trans in Drosophila, Genetics, 178,
tool for analysis of heterochromatin position effect in
749 759, doi: 10.1534/genetics.107.083105.
Drosophila, Biochemistry (Moscow),
83,
542551,
20. Gautam, N. K., Verma, P., and Tapadia, M. G., (2017)
doi: 10.1134/S0006297918050073.
Drosophila malpighian tubules: a model for understanding
12.
Лавров С. А., Шацких А. С., Кибанов М. В., Гвоздев В. А.
kidney development, function, and disease, in Kidney
(2013) Транскрипционная инактивация генов при эф
Development and Disease, (R.K. Miller, Editor.), Springer
фекте положения у D. melanogaster коррелирует на
International Publishing: Cham, pp. 3 25.
уровне отдельных клеток с их перемещением в гете
21. Lamb, M. J. (1982) The DNA content of polytene nuclei
рохроматиновый компартмент ядра, Молекулярная био%
in midgut and Malpighian tubule cells of adult Drosophila
логия, 47, 1 6.
melanogaster, Wilhelm Roux’s archives of developmental
13.
Mcguire, S. E., Le, P. T., Osborn, A. J., Matsumoto, K.,
biology, 191, 381 384, doi: 10.1007/bf00879628.
and Davis, R. L. (2003) Spatiotemporal rescue of memory
22. Vogel, M. J., Pagie, L., Talhout, W., Nieuwland, M.,
dysfunction in Drosophila, Science, 302, 17651768,
Kerkhoven, R. M., and Van Steensel, B. (2009) High res
doi: 10.1126/science.1089035.
olution mapping of heterochromatin redistribution in a
14.
Lee, T., and Luo, L. (1999) Mosaic analysis with a repress
Drosophila position effect variegation model, Epigenetics
ible cell marker for studies of gene function in neuronal
Chromatin, 2, 1, doi: 10.1186/1756 8935 2 1.
morphogenesis, Neuron, 22, 451 461, doi: 10.1016/s0896
23. Harmon, B., and Sedat, J. (2005) Cell by cell dissection of
6273(00)80701 1.
gene expression and chromosomal interactions reveals
15.
Lloyd, V. K., Dyment, D., Sinclair, D. A., and Grigliatti, T. A.
consequences of nuclear reorganization, PLoS Biol., 3,
(2003) Different patterns of gene silencing in position effect
e67, doi: 10.1371/journal.pbio.0030067.
variegation, Genome, 46, 1104 1117, doi: 10.1139/g03 070.
24. Saha, P., Sowpati, D. T. and Mishra, R. K.
(2019)
16.
Lu, B. Y., Bishop, C. P., and Eissenberg, J. C. (1996)
Epigenomic and genomic landscape of Drosophila
Developmental timing and tissue specificity of heterochro
melanogaster heterochromatic genes, Genomics, 111, 177
matin mediated silencing, EMBO J., 15, 1323 1332.
185, doi: 10.1016/j.ygeno.2018.02.001.
THE HIGH LEVEL OF GENE TRANSCRIPTION AT THE EMBRYO STAGE
LEADS TO THE SUPPRESSION OF ITS HETEROCHROMATIС
trans INACTIVATION IN Drosophila melanogaster ADULTS*,**
A. A. Solodovnikov, V. A. Gvozdev, and S. A. Lavrov***
Institute of Molecular Genetics of the Russian Academy of Sciences,
123182 Moscow, Russia; E%mail: slavrov@img.ras.ru
Received December 30, 2019
Revised February 24, 2020
Accepted February 24, 2020
The movement of the gene as a result of chromosomal rearrangement from euchromatin to constitutive heterochro
matin in some cases is accompanied by epigenetic inactivation of this gene cis inactivation. In the case of trans inac
tivation, the transgenes in the normal chromosome are repressed by the cis inactivation causing homologous
rearranged chromosome. Trans inactivation is the result of somatic pairing of homologs and movement of the normal
chromosome region into the heterochromatiс compartment of the nucleus. Previously, we showed that the degree of
trans inactivation of the UAS%eGFP reporter gene in adult flies depends on the level of its transcription, which was reg
ulated by temperature changes on the background of GAL4 transcription activator and GAL80ts temperature sensi
tive inhibitor presenting in the same genome. In this paper we investigated in detail the epigenetic inheritance of the
active/repressed state of the trans inactivated reporter gene at different expression levels by measuring the fluores
cence level of eGFP in individual cells of Malpigian tubes in adult flies. It was found that the high level of expression
at the embryonic stage protects the eGFP gene from trans inactivation in adults, and the activated state is inherited
in the continuation of the entire development and differentiation while the activating effect of GAL4 is turned off.
Keywords: heterochromatin, Drosophila, PEV, trans inactivation, expression, GAL4, GAL80ts
БИОХИМИЯ том 85 вып. 4 2020
7*