БИОХИМИЯ, 2020, том 85, вып. 8, с. 1120 - 1132
УДК 577.23
ЭЛЕКТРИЧЕСКИЕ КАБЕЛИ КЛЕТОК.
II. БАКТЕРИАЛЬНЫЕ ЭЛЕКТРОННЫЕ ПРОВОДНИКИ
Обзор
© 2020
В.В. Птушенко1,2
1 НИИ физико химической биологии имени А.Н. Белозерского Московского государственного университета
имени М.В. Ломоносова, 119992 Москва, Россия; электронная почта: ptush@belozersky.msu.ru
2 Институт биохимической физики имени Н.М. Эмануэля РАН, 119334 Москва, Россия
Поступила в редакцию 17.05.2020
После доработки 12.07.2020
Принята к публикации 16.07.2020
Полвека назад была предложена концепция «электрических кабелей» клетки, участвующих в её биоэнерге
тических процессах [Skulachev, V. P. (1971) Curr. Top. Bioenerg., Elsevier, p. 127 190]. В течение многих десяти
летий считалось, что эту функцию выполняют исключительно мембранные стртуктуры клетки, причём
только для переноса электрохимического трансмембранного потенциала. Однако в последние десять-пят
надцать лет был обнаружен новый тип «электрических кабелей» живой клетки. В 2005 г. были открыты «на
нопровода» (электропроводящие пили) металл и сульфатредуцирующих бактерий, обеспечивающие про
текание электрического тока в масштабах десятков микрометров [Reguera, G. et al. (2005) Nature, 435, 1098
1101]. Ещё через пять лет были открыты бактериальные электрические токи, циркулирующие в простран
ственных масштабах порядка сантиметров [Nielsen, L. P. et al. (2010) Nature, 463, 1071 1074]. Новая группа
бактерий, обеспечивающих протекание электрических токов на макроскопические расстояния, позже по
лучила название «кабельных бактерий». Нанопровода и проводящие структуры кабельных бактерий служат
решению специфической задачи мембранной биоэнергетики - задачи соединения двух полуреакций окис
лительно восстановительной реакции. Иными словами, в отличие от мембранных «кабелей» функция элек
трических проводов этого типа - не передача готовой «энергетической валюты», а перенос электронов в хо
де её генерации в процессах окислительного фосфорилирования. Наиболее удивительной представляется
белковая природа некоторых из обнаруженных бактериальных проводов, на которую указывают последние
данные, поскольку о белковых проводах для дальнего транспорта электронов в живых системах ранее не бы
ло известно.
КЛЮЧЕВЫЕ СЛОВА: концепция «электрических кабелей» клетки, нанопровода, Geobacter sulfurreducens,
кабельные бактерии.
DOI: 10.31857/S0320972520080114
ВВЕДЕНИЕ
ΔμH+ также представляет собой электрический
ток, только уже в направлении вдоль мембраны,
Основные биоэнергетические процессы
а не поперёк.
клетки связаны с протеканием электрических
Из перечисленных электрических токов
токов. Как окислительное фосфорилирование,
только для последнего, как правило, характер
так и фотофосфорилирование включают пере
ны пространственные масштабы циркулирова
нос электронов от восстановителя к окислите
ния порядка микрометров и более. Когда пол
лю, которые пространственно разделены. С пе
века назад В.П. Скулачевым была предложена
реносом электронов сопряжён перенос прото
концепция «электрических кабелей» живой
нов через мембрану. В результате трансмембран
клетки, ответственных за транспорт энергии
ного переноса протонов возникает трансмем
между пространственно отдалёнными генера
бранный протонный электрохимический по
тором и потребителем [1], она предполагала в
тенциал (ΔμH+), который вызывает другие ион
качестве электрического тока именно перенос
ные токи через мембрану, а также распространя
ΔμH+ вдоль мембран. Впоследствии были най
ется далее вдоль мембраны. Распространение
дены примеры переноса ΔμH+ на макроскопи
ческие расстояния (порядка миллиметров) у
бактерий, простейших и животных [2]. Подроб
Принятые сокращения: ΔμH+ - трансмембранный
протонный электрохимический потенциал, ЭТЦ - элек
ное описание зарождения концепции «электри
трон транспортная цепь.
ческих кабелей» живой клетки, её эксперимен
1120
БАКТЕРИАЛЬНЫЕ ЭЛЕКТРОННЫЕ ПРОВОДНИКИ
1121
тального обоснования и наиболее ярких приме
для придатков, построенных из пилина. В то же
ров было приведено в первой части настоящего
время электропроводящие нитевидные придат
обзора [3].
ки бактериальной клетки могут обладать раз
Что же касается протонного и электронного
личной химической природой. В данной статье
токов в дыхательной или фотосинтетической
для внеклеточных электрически проводящих
электрон транспортной цепи (ЭТЦ), то харак
филаментов мы используем термин «нанопро
терные для них пространственные масштабы
вода», введенный Regula et al. [10], а также «пи
гораздо меньше. Транспорт электронов с по
ли подобные придатки». Однако термин «пили»
мощью растворимых переносчиков - напри
также широко используется в данной статье в
мер, белков пластоцианина или цитохрома c -
своём традиционном значении - в качестве об
происходит, как правило, в пределах от единиц
щего термина для нитевидных выростов на по
нанометров в суперкомплексах [4] до единиц
верхности клетки.
микрометров при пространственно разделён
Через пять лет, в 2010 г., были открыты бак
ных комплексах в тилакоидной мембране [5].
териальные электрические токи, циркулирую
Сопряжённый с ним транспорт протонов про
щие в пространственных масштабах порядка
исходит на расстояниях порядка толщины
сантиметров [11]. Позже была открыта новая
мембраны, т.е. единиц нанометров; примерно
группа бактерий, обеспечивающих протекание
такое же расстояние типично для переноса
этих электрических токов на макроскопичес
электрона на участке ЭТЦ внутри белка [6, 7].
кие расстояния [12], которая получила назва
Тем не менее среди всего разнообразия условий
ние «кабельные бактерии» [13]. Эти открытия
обитания живых организмов встречаются и та
необычайно расширили концепцию «электри
кие, при которых доноры и акцепторы электро
ческих кабелей» клетки. В настоящей статье
на находятся на достаточном удалении друг от
кратко описана история открытия нанопрово
друга в окружающей среде, и их использование
дов и электропроводящих филаментов кабель
организмом в энергодонорной окислительно
ных бактерий и приведены данные об их функ
восстановительной реакции требует ЭТЦ зна
ционировании и строении у разных организ
чительно бóльших пространственных размеров.
мов.
Такая ситуация возникает, например, при так
называемом внеклеточном дыхании, когда ко
нечный акцептор по каким либо причинам не
ОТКРЫТИЕ БАКТЕРИАЛЬНЫХ
может быть поглощён клеткой, и его восстанов
НАНОПРОВОДОВ
ление происходит внеклеточно [8]. Клетка
часто решает эту проблему использованием
Представления о «живых проводах» в пос
«электронных челноков» - соединений, кото
ледние полтора десятилетия получили развитие
рые выделяются во внеклеточную среду и могут
с неожиданной стороны - не из биоэнергетики,
быть затем окислены конечными неорганичес
а со стороны биогеохимии и микробиологии.
кими акцепторами, содержащимися в окружа
Две проблемы явились катализаторами поис
ющей среде [9].
ков, которые привели к открытию нового типа
Однако в течение последних полутора деся
биологических «электрических кабелей»: учас
тилетий были обнаружены белковые структуры,
тие в метаболизме микроорганизмов нераство
осуществляющие гораздо более эффективный
римых соединений, содержащихся в окружаю
перенос электронов из клетки во внешнюю сре
щей среде, и биогеохимия донных отложений.
ду или вдоль поверхности клеток на значитель
Исследования метаболизма металл и суль
ные расстояния. В 2005 г. были открыты элект
фатвосстанавливающих бактерий обнаружили
ропроводящие пили у некоторых металл и
их удивительную способность: восстанавливать
сульфатредуцирующих бактерий, передающие
ионы металлов (железа или марганца) независи
электрический ток на расстояния порядка еди
мо от того, присутствуют ли они в среде в виде
ниц-десятков микрометров [10]. Здесь необхо
растворимых или нерастворимых соединений
димо сказать о правомерности использования
(нерастворимые соединения, образующие агре
термина «пили», который традиционно исполь
гаты или микрокристаллы, не могут быть погло
зовали для обозначения разнообразных ните
щены клеткой и непосредственно взаимодей
видных придатков на поверхности бактерий и
ствовать с дыхательной цепью). Электронно
архей. Название клеточной структуры дало наз
микроскопические исследования показали, что
вание одному из составляющих её белков — пи
во втором случае (при наличии в среде лишь не
лину. Однако в настоящее время в терминоло
растворимых соединений металла) клетки обра
гии возникает обратное движение: термин «пи
зуют длинные тонкие выросты - пили, выходя
ли» предлагается использовать исключительно
щие за пределы клетки и имеющие, как мини
БИОХИМИЯ том 85 вып. 8 2020
1122
ПТУШЕНКО
мум, сопоставимые с ней размеры, т.е. порядка
держащегося в донных осадках сероводорода.
микрометров [10] или даже десятков микромет
Удивительным было то, что окисление серово
ров [14] (рис. 1). Идея о том, что эти нитевидные
дорода наблюдалось в анаэробной области дон
образования могут служить
«проводами»
ных отложений, отделённой от доступа кисло
(nanowires) для переноса электронов от дыха
рода сантиметровым слоем осадка, исключав
тельной цепи клетки к внеклеточным окислите
шим сколь либо заметную диффузию кислоро
лям - частицам оксида металла - довольно
да к месту реакции [11]. Протекание окисли
быстро получила первые подтверждения в экс
тельно восстановительной реакции в этом слу
периментах с мутантами, не образующими пили
чае можно было объяснить наличием электри
и одновременно потерявшими способность к
ческого тока, пересекающего толщу осадка и
восстановлению оксидов металла [10].
связывающего две пространственно разнесён
Осуществлённые в последнее время прямые
ные электродные полуреакции - окисление се
измерения электрической проводимости изоли
роводорода в глубине и восстановление кисло
рованных нанопроводов показали значения от
рода на поверхности осадка. Это открытие воз
51 мСм/см при нейтральных pH
[17] до
рождало гипотезу о природных электрических
1,4-4,3 См/см для Geobacter sulfurreducens [18]
токах в поверхностных слоях литосферы и о так
(бактерия, у которой нанопровода были впер
называемых «геобатареях», существовавшую в
вые открыты и наиболее изучены; приведённый
геофизике с 1960 х гг. Эта гипотеза была выска
диапазон значений соответствует сопротивле
зана Sato и Mooney [22] для объяснения анома
нию 730 МОм вдоль нанопровода длиной 1 мкм
лий спонтанного потенциала поверхности Зем
и вызван неопределённостью его диаметра, ис
ли и предполагала роль в их образовании элект
пользованного для расчётов: 2 нм или 3,5 нм по
рохимических процессов в земной коре, подоб
данным для иммобилизованного филамента
ных процессам в гальванической ячейке. При
или находящегося в растворе) и 277 См/см для
этом пласты земной поверхности с относитель
нанопроводов Geobacter metallireducens [16].
но высокой электронной проводимостью могли
Для иллюстрации этих данных можно ска
бы соединять области с различным окислитель
зать, что проводимость 1,4-4,3 См/см способна
но восстановительным потенциалом [23].
обеспечить скорость переноса электронов еди
Исходная модель геобатареи предполагала
ничной пилей длиной 1 мкм ∼ 9 × 108 электронов
крупномасштабные (порядка сотен метров-ки
в секунду (150 нА) при разумной разности потен
лометров) электрохимические системы, в кото
циалов в 100 мВ, что на два порядка превышает
рых высокая электрическая проводимость об
фактическую скорость восстановления нераст
ласти, связывающей «анодную» и «катодную»
воримого оксида железа (III) клеткой (∼ 9 × 106
зоны литосферы, была обусловлена соответ
электронов в секунду на одну клетку) [18]. При
ствующей минерализацией [23]. Позже был
мечательно, что этот механизм электронной
предложен биогеохимический вариант этой мо
проводимости обеспечивает величину силы то
дели - биогеобатарея, в которой различие в
ка, сравнимую с той, которая характерна для пе
электрохимических потенциалах анодной и ка
реноса энергии ΔμH+ вдоль цианобактериально
тодной областей создавалось за счёт жизнедея
го трихома (50 пА или 3 × 108 элементарных за
тельности бактерий, а проводимость разделяю
рядов в секунду) [19].
щего их слоя обеспечивали биоплёнки и осаж
В целом, для результатов измерений прово
дённые металлические частицы [24]. В случае
димости бактериальных проводов характерна
же, обнаруженном группой Nielsen et al. [11],
значительная вариабельность, что, вероятно,
как оказалось, не только обе электродные реак
обусловлено их неоднородностью, которая мо
ции, но и электрический ток между областями
жет быть существенна для проводимости в на
их протекания обеспечивали бактерии. Как вы
номасштабах [20]. У бактерии Shewanella onei
яснилось, это были длинные нитчатые много
densis более крупные пили подобные придатки
клеточные бактерии [12], содержащие тысячи
имели проводимость в диапазоне от 60 мСм/см
клеток и имеющие длину порядка сантиметров
до 1 См/см ([21]; см. также анализ в статье Tan et
[25]. Данные электронной микроскопии показа
al. [20]), однако они представляли комплекс
ли, что вдоль поверхности клеток тянутся длин
белка с липидами, что дополнительно затрудня
ные тяжи диаметром ~50 нм [26], уложенные па
ет стандартизацию исследуемого материала.
раллельно оси многоклеточной бактериальной
Однако ещё более впечатляющее открытие
нити, как натянутые струны вдоль грифа, и за
принесло изучение биогеохимических окисли
нимающие почти всю её поверхность [12]. Эти
тельно восстановительных процессов в донных
тяжи расположены в периплазматическом
отложениях. Группа исследователей из Дании и
пространстве и «укутаны» внешней мембраной;
Японии обнаружила необычное окисление со
причём они не прерываются и в межклеточной
БИОХИМИЯ том 85 вып. 8 2020
БАКТЕРИАЛЬНЫЕ ЭЛЕКТРОННЫЕ ПРОВОДНИКИ
1123
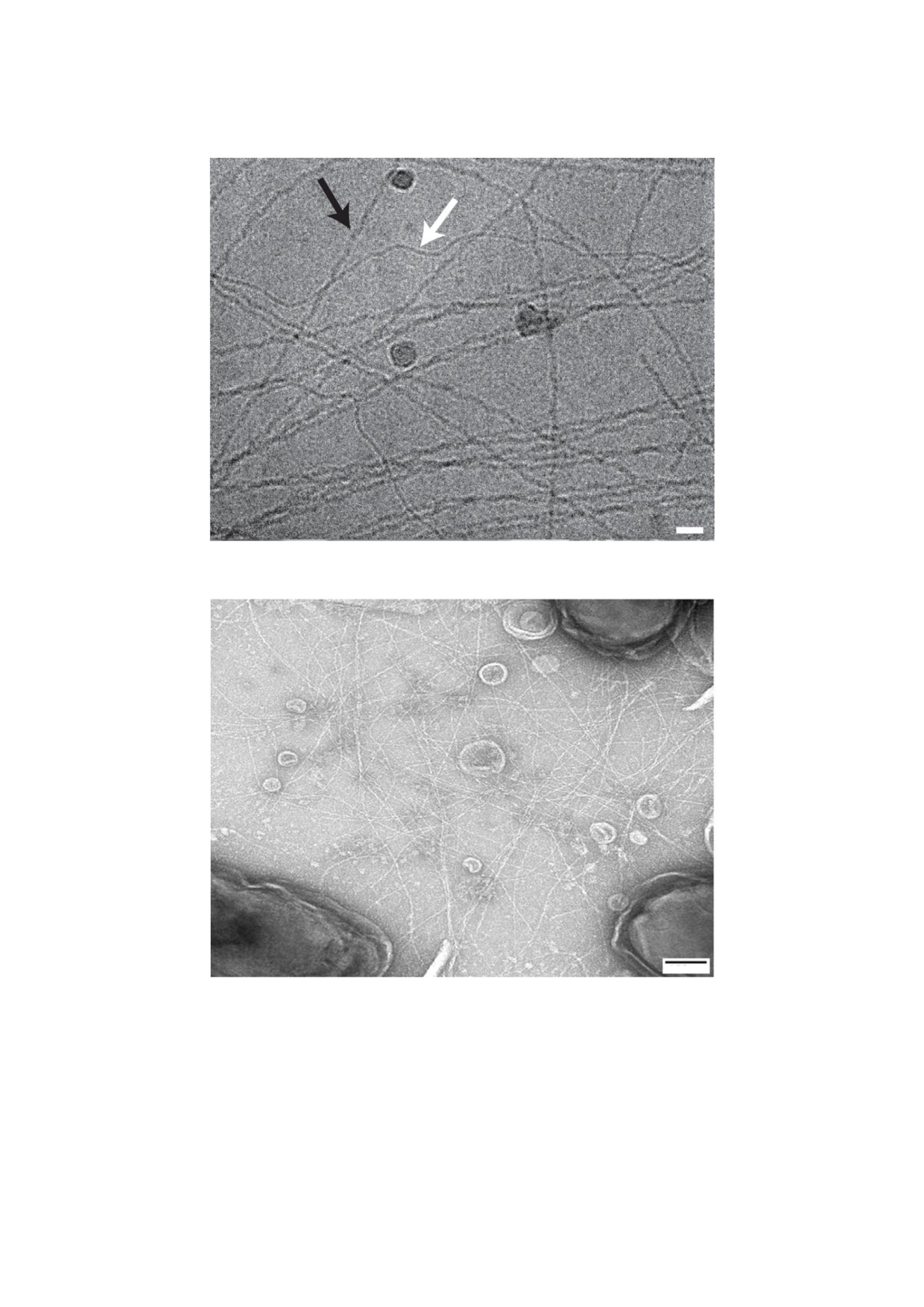
а
20 нм
б
100 нм
Рис. 1. a - Пили, изолированные из G. sulfurreducens, масштабный отрезок - 20 нм, криоэлектронная микроскопия, адап
тировано по [15]. б - Клетки штамма G. sulfurreducens, продуцирующие пили G. metallireducens, масштабный отрезок -
100 нм, просвечивающая криоэлектронная микроскопия, адаптировано по [16]
области, связывая между собой таким образом
проводами клетки, по которым она передаёт
все клетки (рис. 2). Уже с самого начала после их
электроны через толщу донного осадка. При
обнаружения было высказано предположение,
этом плотно окружающая их внешняя мембрана
что эти тяжи и служат теми электрическими
играет роль изолятора [12].
БИОХИМИЯ том 85 вып. 8 2020
1124
ПТУШЕНКО
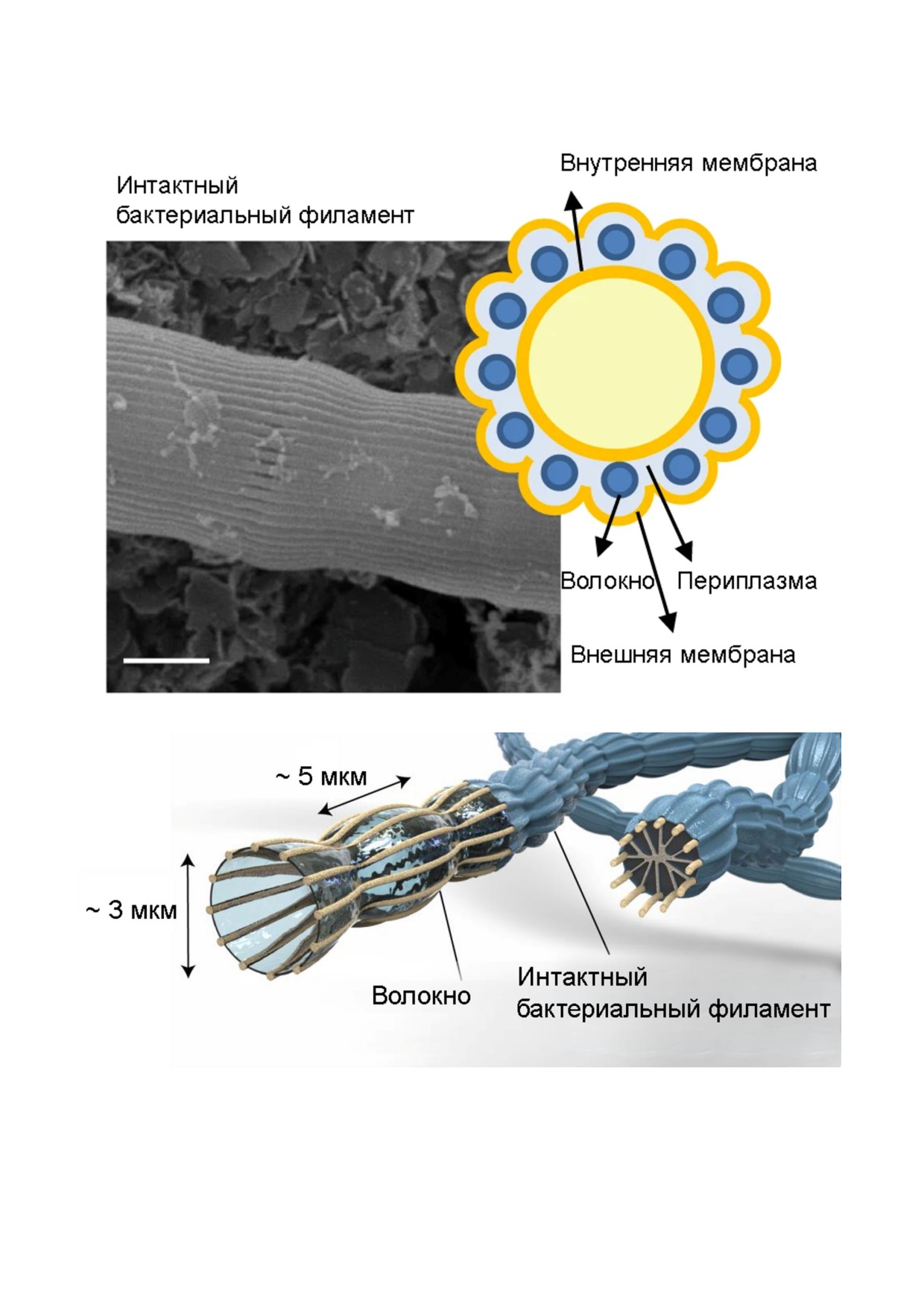
а
б
Рис. 2. Строение филамента кабельной бактерии (семейство Desulfobulbaceae). Интактный филамент (сканирующая
электронная микроскопия, масштабный отрезок - 1 мкм), схематичное изображение поперечного сечения филамента (a)
и филамента в целом (б). Адаптировано по: (a) - [27]; (б) - [28]. (С цветными вариантами рис. 2-4 можно ознакомиться
БИОХИМИЯ том 85 вып. 8 2020
БАКТЕРИАЛЬНЫЕ ЭЛЕКТРОННЫЕ ПРОВОДНИКИ
1125
СТРУКТУРА ЭЛЕКТРОПРОВОДЯЩИХ
«бусы» из пилина или цитохрома, нанизанных
БАКТЕРИАЛЬНЫХ ОБРАЗОВАНИЙ
на углеводную нить [25].
Однако существующие представления о
При всей значимости микробиологического,
структуре электропроводящих пилей металл и
экологического и биогеохимического аспектов
сульфатредуцирующих бактерий, на которые
этих открытий наиболее интригующим выгля
опираются гипотезы о химической природе на
дит механизм электрической проводимости на
нопроводов кабельных бактерий, в действитель
нопроводов. Можно предполагать, что у нано
ности не вполне надёжны. До сегодняшнего дня
проводов, образуемых разными бактериями,
они являются предметом интенсивных исследо
есть много общего. При кардинальном разли
ваний и постоянно сменяющих друг друга тео
чии в длине (десятки микрометров для нанопро
рий. Так, для бактерии G. sulfurreducens, у кото
водов и десятки миллиметров для кабельных
рой впервые обнаружили электропроводящие
бактерий) поперечные размеры этих структур
пили, предполагали, что их образует белок пи
близки: периплазматические волокна кабель
лин - продукт гена pilA [10]. На это указывали
ных бактерий имеют диаметр 50 нм [26], попе
результаты делеции гена pilA из генома G. sul
речный размер пили подобных придатков
furreducens - клетки оказывались неспособными
S. oneidensis также был оценен в десятки нано
формировать пили и восстанавливать оксид же
метров (50-150 нм) [14]. Диаметр пилей G. sul
леза. В то же время вставка функциональной ко
furreducens существенно меньше (3 нм), а для
пии гена pilA в геном pilA дефицитного мутанта
мутанта с модифицированным пилином - всего
восстанавливала эту способность [10]. Пред
1,5 нм [20]. Однако на электронно микроскопи
ставление о пилине (конкретно, пилине типа IV),
ческих изображениях выростов S. oneidensis в
как белке, формирующем пили, было естествен
местах их обрыва были видны слагающие их от
но также в силу того, что оно было хорошо изве
дельные тонкие волокна диаметром 3-5 нм [14],
стным фактом для других бактерий [30, 31], хотя
т.е. близкие к размерам пилей G. sulfurreducens.
геопилины (пилины бактерий рода Geobacter) и
На этом, однако, известные нам сведения о
отличались довольно существенно от соответ
сходстве и различии этих проводящих систем
ствующих пилинов ( пилинов типа IV). Для про
практически заканчиваются. Кабельные бакте
теобактерии S. oneidensis MR 1, образующей пи
рии пока не удаётся культивировать в чистой
ли подобные электропроводящие придатки,
культуре, для них получены лишь черновые ге
также было показано участие в образовании пи
номные последовательности (draft genomes) на
лей продуктов гена pilA, наряду с псевдопилина
основании метагеномных данных и геномики
ми (gspG) и манноза чувствительными гемаг
отдельных бактериальных филаментов [25]. На
глютининами (mshA) [14]. Однако наряду с пи
основании анализа последовательностей генов
лином в качестве обязательного компонента для
диссимиляционной сульфитредуктазы и 16S
переноса электрона по нанопроводам также с
рРНК индивидуальных филаментов был выпол
самого начала рассматривались цитохромы. Так,
нен филогенетический анализ и предложены
шести и четырёхгемовый цитохромы c внеш
два кандидатных рода, относящихся к семейству
ней мембраны G. sulfurreducens, OmcS и OmcE,
сульфатредуцирующих и серо диспропорцио
экспрессировались только при наличии в среде
нирующих протеобактерий Desulfobulbaceae:
нерастворимого оксида железа (III) (но не раст
Candidatus Electrothrix, содержащий четыре пре
воримого цитрата!) и были необходимы для его
имущественно морских кандидатных вида, и
восстановления [32] - т.е. проявляли те же осо
Candidatus Electronema с двумя преимущественно
бенности образования и функционирования в
пресноводными кандидатными видами [29]. На
клетке, что и пили. Позже для OmcS также было
основании электронно микроскопических, не
показано, что этот белок связан с пилями и рас
полногеномных и метапротеомных данных и по
полагается вдоль них кластерами [33], а другие
аналогии с бактериями, образующими элект
мультигемовые цитохромы также оказались ли
ропроводящие пили, было высказано предполо
бо экскретируемыми во внешнее пространство
жение, что нанопровода у кабельных бактерий
и слабо связанными с наружными клеточными
образованы пилином (PilA), который является
образованиями, как OmcZ [34], либо связанны
наиболее распространённым белком у изучен
ми с внешней мембраной и экспонированными
ных кабельных бактерий [25]. При этом предпо
наружу, как OmcB [35] или OmcE [36]. У S. onei
лагается, что пилин мог бы образовывать пили
densis проводимость пили подобных придатков
подобные тонкие (3-5 нм) нити, которые, в
существенно падала при делеции генов десяти
свою очередь, сплетаются в электропроводящие
гемовых цитохромов MtrC и OmcA [14]. Нако
волокна, расположенные в периплазме. В каче
нец, были получены данные, свидетельствую
стве альтернативной гипотезы предполагались
щие, вопреки представлениям предшествующих
БИОХИМИЯ том 85 вып. 8 2020
1126
ПТУШЕНКО
восьми лет исследований, что эти придатки у
выше [10]. Наконец, (5) были получены генно
S. oneidensis, по видимому, никак не связаны с
инженерные пили, которые демонстрировали
пилином, а являются выростами периплазмати
проводимость, сопоставимую с проводимостью
ческого пространства и внешней мембраны, со
природных пилей (сопротивление рекомби
держащей целый ряд мультигемовых цитохро
нантного белкового провода длиной 6 мкм со
мов (MtrC, OmcA, MtrA, MtrB) [37].
ставляло ∼900 МОм, в то время как для природ
ной пили в том же исследовании была получена
величина 925 МОм [42]).
МЕХАНИЗМЫ ЭЛЕКТРОПРОВОДИМОСТИ
За «цитохромно/суперобменный» механизм
НАНОПРОВОДОВ
говорили данные о том, что наличие наиболее
массовых цитохромов клетки, для которых
В связи с приведёнными выше данными о
предполагалась связь с пилями, критично для
строении нанопроводов возникли две альтерна
проводимости как пилей, так и биоплёнок, фор
тивные гипотезы механизма переноса по ним
мируемых бактериями [14, 21, 43]. Правда, в
электрона: гипотеза металлоподобной проводи
части работ были получены противоположные
мости [38] и гипотеза, основанная на супероб
результаты - сохранение или даже увеличение
менном взаимодействии редокс кофакторов
проводимости пилей и биоплёнок, образуемых
[39]. В соответствии с первой из них перенос
бактериями с инактивированными цитохрома
электрона происходит по системе остатков аро
ми [38], а также отсутствие положительной кор
матических аминокислот белка пилина в пред
реляции (или отрицательная корреляция) между
положении, что ароматические кольца уложены
содержанием цитохромов и проводимостью у
в белке параллельно друг другу, и π системы со
разных штаммов [44]. Ещё на один существен
седних колец перекрываются. Проводимость
ный недостаток суперобменной гипотезы ука
пилей в этом случае аналогична проводимости
зывали теоретические модели переноса элект
полимеров полианилина или полипиррола. При
рона: слишком малое расстояние (не более 1 нм)
этом молекулы цитохрома, ассоциированные с
между соседними гемами и предельно малые
пилями, осуществляют непосредственный пере
значения энергии реорганизации переноса
нос электронов от пилей к металлу [40] либо к
электрона, необходимые для достижения экспе
пилям от дыхательной цепи [36]. Суперобмен
риментально наблюдаемой силы тока [45]. При
ная теория, наоборот, предполагает активное
этом фактическое расстояние между соседними
участие цитохромов в переносе электрона вдоль
молекулами цитохрома OmcS, которое было
пили за счёт последовательных «перескоков»
оценено по данным электронной микроскопии
электрона между соседними гемами. Таким об
с окрашиванием антителами, конъюгированны
разом, две изначальные альтернативные гипоте
ми с коллоидным золотом, составляло 30 нм в
зы различались как по предполагаемому физи
кластерах и 100-200 нм между кластерами ([33];
ческому механизму проводимости, так и по
см. также анализ в статье Malvankar et al. [44]),
представлениям о роли разных белков в её обес
т.е. намного больше требуемых величин. Тем не
печении.
менее и здесь теоретические оценки, сделанные
В поддержку «пилиновой/ароматической»
разными группами, существенно расходились
гипотезы был получен ряд разнообразных дан
между собой, и некоторые из них предсказыва
ных: (1) мутации по ароматическим остаткам,
ли значения силы тока, которые могут быть
т.е. снижающие их количество, приводили к
обеспечены суперобменным механизмом при
снижению проводимости пилей [18, 41]; (2) му
вполне реалистичных параметрах редокс реак
тации, заменяющие фенилаланин и тирозин на
ций и плотностях расположения гемов вдоль
обеспечивающий более высокую проводимость
длины пили, близкие к наблюдаемым [39].
триптофан, наоборот, увеличивали проводи
В свете этих дискуссий, в ходе которых был
мость [20]; (3) сопоставление природных пили
получен огромный массив результатов, совер
нов из разных видов рода Geobacter показывало
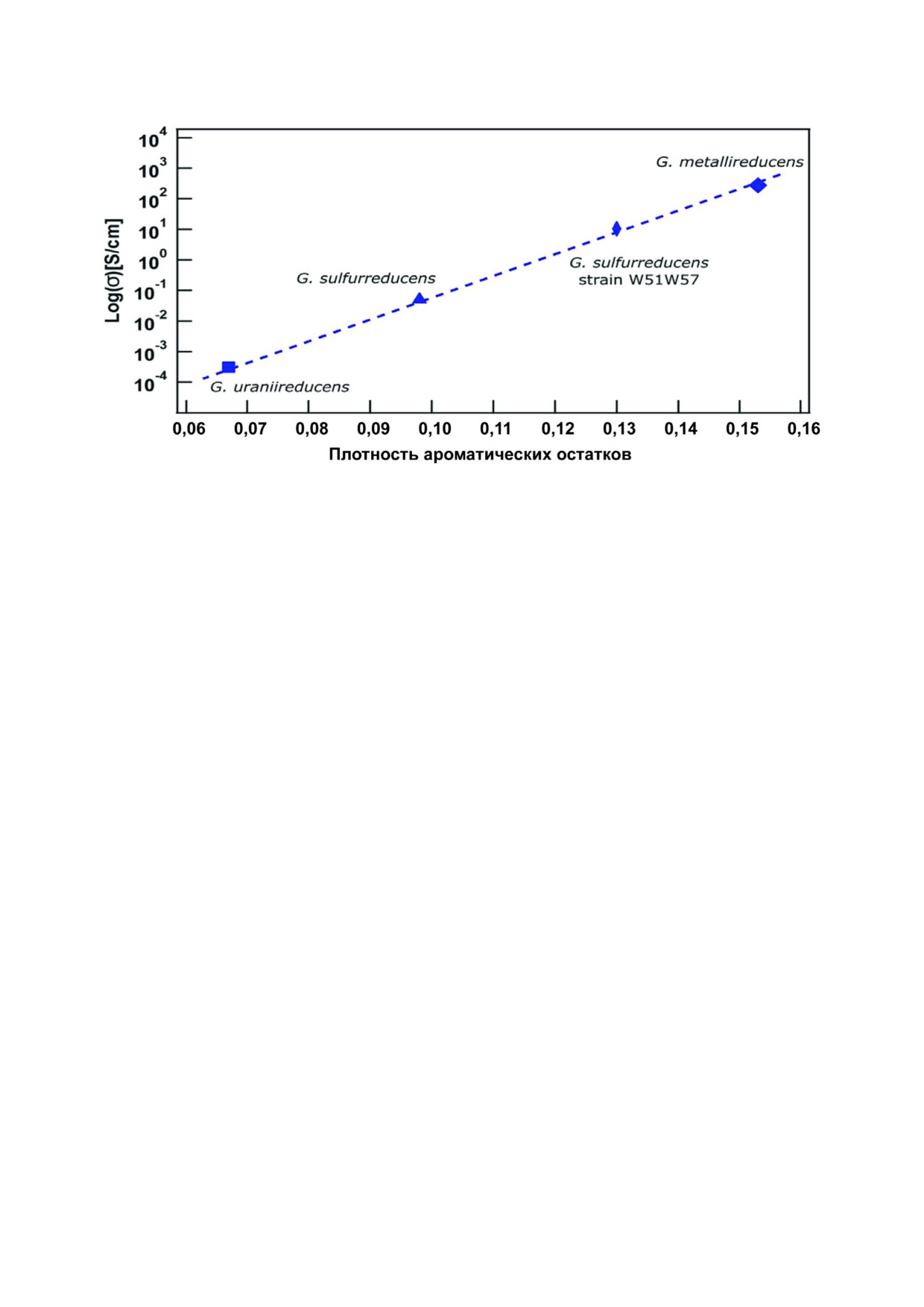
шенно неожиданными оказались недавно полу
сильную экспоненциальную зависимость между
ченные структурные данные. Как показали дан
содержанием в них ароматических остатков и
ные криоэлектронной микроскопии, нанопро
проводимостью пилей, образуемых бактериями
вода G. sulfurreducens образованы не пилином, а
(рост проводимости в 105 раз при удвоении со
шестигемовым цитохромом c - OmcS [15, 46].
держания ароматических остатков в PilA [16];
Иными словами, молекулы цитохрома не свя
рис. 3); (4) зависимость способности образовы
зываются с поверхностью пилей, образованных
вать пили и восстанавливать нерастворимые со
пилином, как предполагалось ранее, а сами об
единения металла у бактерий от наличия актив
разуют нанопровода (рис. 4). При этом гемы
ного гена пилина pilA, о чём уже упоминалось
шестигемовых цитохромов образуют непрерыв
БИОХИМИЯ том 85 вып. 8 2020
БАКТЕРИАЛЬНЫЕ ЭЛЕКТРОННЫЕ ПРОВОДНИКИ
1127
Рис. 3. Корреляция между содержанием ароматических аминокислотных остатков в PilA и проводимостью индивидуаль
ной пили при pH 7. Каждая точка соответствует мутантному (W51W57) или одному из природных пилинов, принадлежа
щих бактериям G. metallireducens, G. sulfurreducens или Geobacter uraniireducens, адаптировано по [16]
ную цепь с расстояниями между соседними ге
вительной реакции в целом. При таком объяс
мами не более 0,41 нм. Тем самым внутри белка
нении, однако, остаётся загадкой идеальная
образуется проводящий канал, и в наблюдаемой
экспоненциальная зависимость проводимости
картине фактически совмещаются основные
пилей от содержания ароматических остатков в
представления двух разных исходных гипотез о
пилине, показанная на рис. 3 [16]. Наконец,
механизме переноса электрона.
нельзя исключить возможность сосуществова
Эта изящная картина, однако, порождает
ния в клетке нанопроводов различного типа, в
много вопросов о том, как интерпретировать те
частности, образованных пилином или цито
разнообразные данные, которые ранее рассмат
хромом. Однако в связи с этим возникает воп
ривались в качестве несомненных свидетельств
рос: почему клетке необходимо иметь несколько
в пользу «пилиновой» и против «цитохромной»
различных структур для обеспечения одной и
модели. В первую очередь это относится к дан
той же функции? Возможно, нынешний пово
ным, полученным на мутантах с инактивиро
рот в наших представлениях о природе бактери
ванными пилинами или цитохромами: почему
альных нанопроводов не последний, и у пилина
первые из них теряли способность образовывать
найдётся ещё какая то функция, кроме секре
нанопровода, а вторые, наоборот, сохраняли её?
торной, которая даст более логичное объясне
Одно из возможных объяснений заключается в
ние найденным ранее зависимостям проводи
том, что инактивация гена цитохрома OmcS
мости пилей от присутствия в клетках пилина и
приводит к повышению уровня экспрессии дру
его молекулярного строения (содержания аро
гого мультигемового цитохрома, OmcZ, гипер
матических групп в белке).
продукция которого могла бы скомпенсировать
Отметим ещё один вопрос, возникающий в
отсутствие OmcS. Что же касается пилина PilA,
связи со структурой нанопроводов - проводов
то его в рамках новых преставлений предполага
без изоляции. Каким образом возможно функ
ют компонентом секреторной системы клетки,
ционирование проводов без электрической изо
от которого зависит сборка электропроводящих
ляции в высокопроводящей среде, каковой яв
цитохромных нанопроводов. При этом процесс
ляется морская вода? Как видно из структурных
сборки и конечная конформация нанопроводов
данных, цитохромный нанопровод представля
каким то образом меняются при точечных мута
ет собой чисто белковую структуру, не имеющую
циях в пилине [46]. Можно также предполо
мембранной «оплётки». При этом расстояние от
жить, что электронная проводимость нанопро
гемов до границы белок-вода сравнимо с рас
вода не является лимитирующим фактором для
стояниями между соседними гемами. Следова
скорости протекания окислительно восстано
тельно, можно ожидать, что токи «утечки» через
БИОХИМИЯ том 85 вып. 8 2020
1128
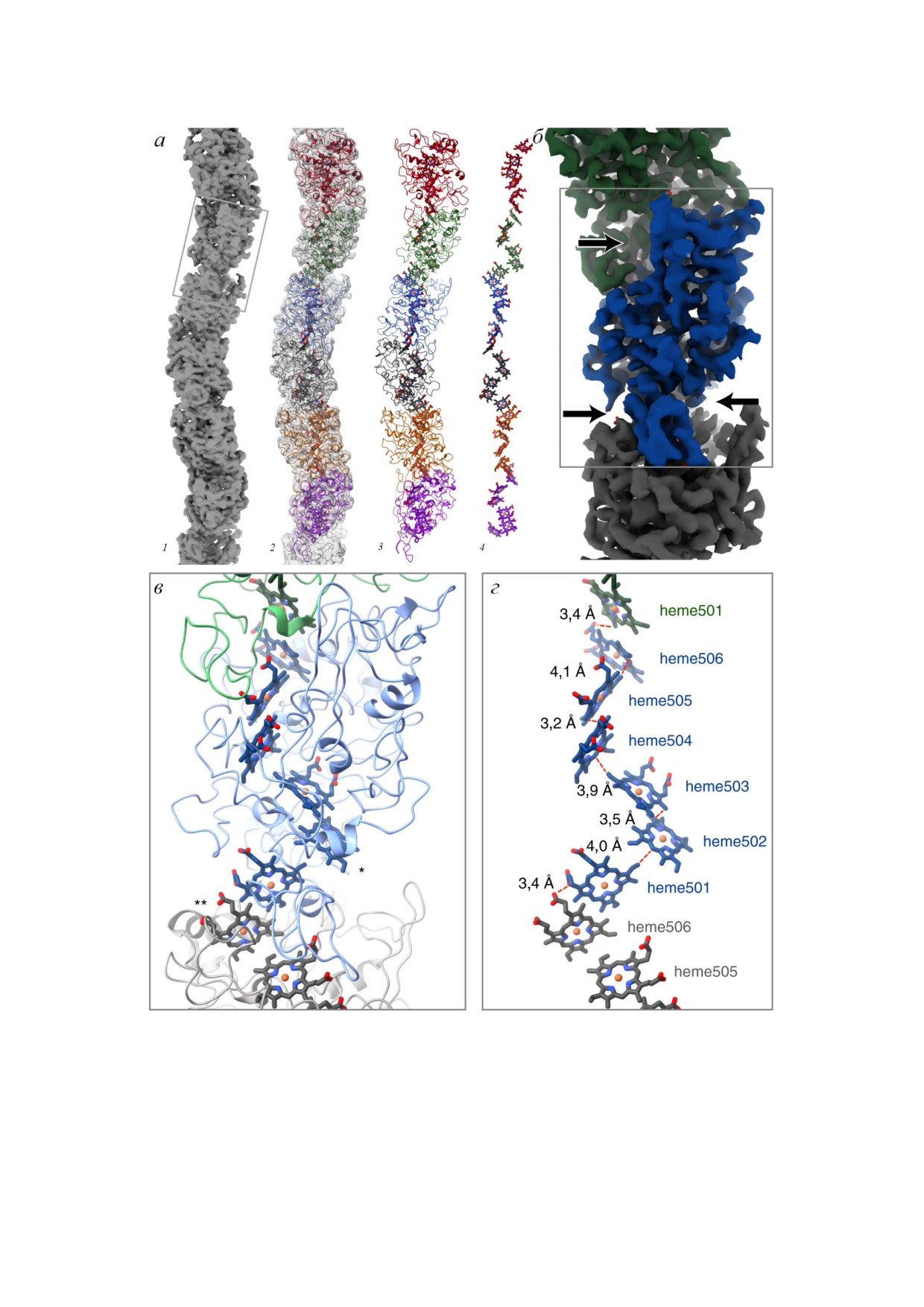
ПТУШЕНКО
Рис. 4. Строение нанопровода металл и сульфатредуцирующих бактерий G. sulfurreducens по данным криоэлектронной
микроскопии. a - Визуализация поверхности пили (1), контуров поверхности с наложением атомной модели белка (2),
атомной модели, включающей пептидную цепь белка и гемы (3), а также только гемы (4). б - Визуализация поверхности
короткого участка пили, синим цветом выделена одна субъединица цитохрома OmcS, стрелки указывают её области кон
такта с соседними субъединицами. в - Атомная модель, показывающая укладку пептидной цепи вокруг гемов; звездочка
ми указаны гемы, экспонированные в водную фазу: heme502 (*) и heme506 (**) соседних субъединиц. г - Атомная модель
гемов, расположенных в соседних субъединицах цитохрома OmcS вдоль длины филамента; указаны кратчайшие рассто
яния (edge to edge) между соседними гемами. Адаптировано по [15]
БИОХИМИЯ том 85 вып. 8 2020
БАКТЕРИАЛЬНЫЕ ЭЛЕКТРОННЫЕ ПРОВОДНИКИ
1129
боковую поверхность нанопровода будут значи
должны снижаться. Интересно сравнить эти ве
тельны, и поддержание движения электронов
личины с аналогичными величинами, характер
вдоль нанопровода будет затруднено. Таким об
ными для переноса энергии ΔμH+ вдоль циано
разом, функционирование подобной белковой
бактериальных трихомов. Для цианобактерий
структуры в качестве электрического провода на
типичные значения электрического тока состав
первый взгляд может показаться невозможным
ляли ∼50 пА (3 × 108 элементарных зарядов в се
в электропроводящей среде. Однако, по види
кунду) [19], а характерное расстояние, на кото
мому, здесь мы сталкиваемся с тем случаем, ког
ром происходит потеря 50% энергии, было тео
да недостаток конструкции оказывается её дос
ретически оценено на основе данных о геомет
тоинством в силу её физиологических функций.
рии трихома и электрической проводимости
Поскольку задача нанопровода - сбросить
мембраны и цитоплазмы, как ∼7 мм [48]. Таким
электрон на какой либо экзогенный акцептор,
образом, механизм электронной проводимости
то «утечка» электронов с него вполне отвечает
у кабельных бактерий, по видимому, превосхо
его задаче. Более того, в условиях, при которых
дит механизм переноса энергии ΔμH+ вдоль циа
клетка образует электропроводящие пили, мож
нобактериального трихома как по величине
но говорить скорее о дефиците акцепторов
проводимого электрического тока (десятки на
электрона в среде. Напомним, что когда подхо
ноампер для кабельных бактерий [27] и 50 пА -
дящие акцепторы (растворимые соединения ио
для цианобактерий [19]), так и по дальности пе
нов переходных металлов) присутствуют в сре
реноса энергии, ограниченной электрическими
де, бактерии не образуют электропроводящие
потерями.
пили [10].
В завершение этого раздела вернёмся к ка
бельным бактериям, которые пока остаются
ЗАКЛЮЧЕНИЕ
менее изученными, чем описанные выше пи
ли образующие бактерии. Как показывают
Представление об «электрических кабелях»,
последние данные [27], сопротивление одной
связывающих биоэнергетические процессы в
клетки в составе всей нити составляет ∼5 кОм.
разных частях живой клетки или надклеточной
Если оценивать проводимость индивидуаль
системы, появилось полвека назад и было связа
ных периплазматических волокон (которых, в
но с развитием мембранной биоэнергетики -
зависимости от размеров клеток, может содер
научной дисциплины, формировавшейся в тот
жаться несколько десятков в одном филамен
момент и вступавшей в период особенно интен
те), то она составляет в физиологических усло
сивного развития. Это представление было свя
виях ∼20 См/см, т.е. приближается к той, что
зано, прежде всего, с мембранами как структу
наблюдается для наиболее высокопроводящих
рами, обеспечивающими распространение
пилей G. metallireducens. При этом длина воло
электрического тока, и с ионами, создающими
кон на три порядка превосходит длину пилей.
трансмембранный электрохимический потен
Величина тока, который течёт через филамент
циал, как его носителями. Электрический ток, в
при напряжениях, характерных для физиологи
котором носителями являются электроны, а
ческих условий, может составлять до десятков
средой - белки, также всегда был одним из глав
наноампер, а для одного волокна она оценена в
ных объектов исследований биоэнергетики, од
1,3 нА (∼8 × 109 электронов в секунду). При этом
нако воспринимался в основном как «локаль
температурная зависимость проводимости со
ный», существующий в масштабах нанометров в
ответствует закону Аррениуса, что характерно
пределах отдельных белковых комплексов. От
для полупроводников, но не для металлов, и
крытие в последние полтора десятилетия бакте
исключает механизм металлоподобной прово
риальных «нанопроводов» и проводящих воло
димости [28].
кон кабельных бактерий, имеющих, по видимо
Падение напряжения вдоль волокна в интакт
му, белковую природу и осуществляющих пере
ных клетках в условиях, близких к естествен
нос электронов на расстояния в микрометровой
ным, было оценено в 14 ± 4 мВ/мм. Даже для от
и даже сантиметровой шкале, необычайно рас
носительно короткого волокна длиной 20 мм
ширили концепцию «электрических кабелей» в
это соответствует падению напряжения до 0,3 В,
живых системах. Мы пишем «по видимому»,
т.е. полные резистивные потери в нанопроводе
так как исследования бактериальных электрон
достаточно велики (сравнимы с теоретическим
ных проводников происходят интенсивно и
пределом падения электрохимического потен
постоянно приносят новые данные, порой резко
циала при окислении сероводорода, ∼1 В) [47],
меняющие представления, которые успели о
хотя при уменьшении силы электрического тока
них сложиться за предшествующие десять-пят
через бактериальный филамент они также
надцать лет. Так, пили подобные придатки
10 БИОХИМИЯ том 85 вып. 8 2020
1130
ПТУШЕНКО
S. oneidensis оказались в итоге мембранными об
P. thermopropionicum и архебактерии Methano
разованиями [37]. Какую роль играет в этом слу
thermobacter thermautotrophicus [49]. Связывать
чае мембрана - только ли «чехла» для всей
клетки бактерий разных видов могут и сугубо
структуры, предохраняющего её от потери раст
белковые пили, например в смешанной культу
воримых компонентов, или носителя мембран
ре G. metallireducens и G. sulfurreducens; явление,
ных белков, или же обладает также свойствами
обеспечиваемое такими связями, получило наз
сопрягающей мембраны - вопрос пока откры
вание прямого межвидового переноса электро
тый. Что же касается проводящих структур ка
нов (direct interspecies electron transfer) [50]. Даже
бельных бактерий, то для них показана тесная
эти немногие примеры дают основание ожидать
связь с внешней мембраной бактериальных кле
в скором времени существенное расширение
ток, которая «укутывает» каждое из отдельных
наших представлений о роли «электрических
волокон по всей его длине [26]. Служит ли при
кабелей» не только в биоэнергетических про
этом мембрана лишь «оплёткой», изолирующей
цессах, но и в процессах функционирования и
белковое волокно от растворённых в среде ио
коэволюции биологических сообществ.
нов, или же сама принимает какое то участие в
передаче электрического тока - также вопрос
будущих исследований.
Финансирование. Исследование выполнено в
Однако что ясно уже сейчас и что является
рамках бюджетного финансирования Московс
наиболее существенным элементом картины -
кого государственного университета имени
широкая распространённость, практически
М.В. Ломоносова, гостема АААА А17
универсальность «электрических кабелей» сре
1171205700114, и Института биохимической
ди живых организмов. Мембранные кабели (или
физики имени Н.М. Эмануэля РАН, гостема
сети) оказались характерными как для многих
01201253314.
тканей животных организмов, так и для циано
Благодарности. Автор благодарен D. Lovley,
бактерий, простейших и растений, хотя прост
M. Strauss, J.V. Manca, J. Oemig и Ф.О. Каспа
ранственные масштабы циркулирования элект
ринскому за любезное предоставление рисунков
рических токов во всех этих случаях могут быть
для публикации. Автор благодарен анонимным
разные. Бактериальные электронные проводни
рецензентам, чьи конструктивные замечания
ки также не ограничиваются лишь узкими груп
позволили улучшить статью.
пами металл , сульфатредуцирующих и «кабель
Конфликт интересов. Автор заявляет об отсут
ных» бактерий. Так пили подобные образова
ствии конфликта интересов в финансовой или
ния, связывающие между собой несколько кле
какой либо иной сфере.
ток, оказались характерными для разных групп
Соблюдение этических норм. Настоящая
бактерий. Они были обнаружены у цианобакте
статья не содержит описания выполненных ав
рий Synechocystis PCC6803 [14] и у грамположи
тором исследований с участием людей или ис
тельных пропионат окисляющих бактерий
пользованием животных в качестве объектов.
Pelotomaculum thermopropionicum [49]. Более того,
Приведённые в настоящей статье рисунки адап
оказалось, что подобные структуры могут свя
тированы из статей, указанных в подрисуноч
зывать клетки разных видов, даже предельно от
ных подписях, оригинальные рисунки опубли
далённых друг от друга - например эубактерии
кованы по лицензии CC BY 4.0.
СПИСОК ЛИТЕРАТУРЫ
1.
Skulachev, V. P. (1971) Energy transformations in the res
consequences on cyclic electron flow, Photosynth. Res.,
piratory chain, Curr. Top. Bioenerg., Elsevier, pp. 127 190.
129, 307 320.
2.
Skulachev, V. P. (1980) Integrating functions of biomem
6.
Ptushenko, V. V., Cherepanov, D. A., Krishtalik, L. I., and
branes. Problems of lateral transport of energy, metabolites
Semenov, A. Y. (2008) Semicontinuum electrostatic calcu
and electrons, Biochim. Biophys. Acta, 604, 297 320.
lations of redox potentials in photosystem I, Photosynth.
3.
Ptushenko, V. V. (2020) Electric cables of living cells. I.
Res., 97, 55 74.
Energy transfer along coupling membranes, Biochemistry
7.
Ptushenko, V. V., and Krishtalik, L. I. (2018) Reorganiza
(Moscow), 85, 820 832, doi: 10.1134/S000629792007010X.
tion energies of the electron transfer reactions involving
4.
Minagawa, J. (2016) A supercomplex of cytochrome bf and
quinones in the reaction center of Rhodobacter sphaeroides,
photosystem I for cyclic electron flow, in Cytochrome
Photosynth. Res., 138, 167 175.
Complexes: Evolution, Structures, Energy Transduction, and
8.
Gralnick, J. A., and Newman, D. K. (2007) Extracellular
Signaling, (Cramer, W. A., and Kallas, T., eds) Springer, pp.
respiration, Mol. Microbiol., 65, 1 11.
453 462.
9.
Turick, C. E., Tisa, L. S., and Caccavo, F., Jr. (2002) Melanin
5.
Dumas, L., Chazaux, M., Peltier, G., Johnson, X., and
production and use as a soluble electron shuttle for Fe (III)
Alric, J. (2016) Cytochrome b 6 f function and localization,
oxide reduction and as a terminal electron acceptor by
phosphorylation state of thylakoid membrane proteins and
Shewanella algae BrY, Appl. Environ. Microbiol., 68, 2436 2444.
БИОХИМИЯ том 85 вып. 8 2020
БАКТЕРИАЛЬНЫЕ ЭЛЕКТРОННЫЕ ПРОВОДНИКИ
1131
10.
Reguera, G., McCarthy, K. D., Mehta, T., Nicoll, J. S.,
M., Nielsen, P. H., Nielsen, L. P., and Schramm, A. (2019)
Tuominen, M. T., and Lovley, D. R. (2005) Extracellular elec
On the evolution and physiology of cable bacteria, Proc.
tron transfer via microbial nanowires, Nature, 435, 1098 1101.
Natl. Acad. Sci. USA, 116, 19116 19125.
11.
Nielsen, L. P., Risgaard Petersen, N., Fossing, H.,
26.
Cornelissen, R., Bøggild, A., Thiruvallur Eachambadi, R.,
Christensen, P. B., and Sayama, M. (2010) Electric cur
Koning, R. I., Kremer, A., Hidalgo Martinez, S.,
rents couple spatially separated biogeochemical processes
Zetsche, E. M., Damgaard, L. R., Bonné, R.,
in marine sediment, Nature, 463, 1071 1074.
Drijkoningen, J., Geelhoed, J. S.,Boesen, T., Boschker, H. T. S.,
12.
Pfeffer, C., Larsen, S., Song, J., Dong, M., Besenbacher, F.,
Valcke, R., Nielsen, L. P., D’Haen, J., Manca, J. V., and
Meyer, R. L., Kjeldsen, K. U., Schreiber, L., Gorby, Y. A.,
Meysman, F. J. R. (2018) The cell envelope structure of
El Naggar, M. Y., Leung, K. M., Schramm, A., Risgaard
cable bacteria, Front. Microbiol., 9, 3044.
Petersen, N., and Nielsen, L. P. (2012) Filamentous bacte
27.
Meysman, F. J. R., Cornelissen, R., Trashin, S., Bonné, R.,
ria transport electrons over centimetre distances, Nature,
Martinez, S. H., van der Veen, J., Blom, C. J., Karman, C.,
491, 218.
Hou, J. L., Thiruvallur Eachambadi, R., Geelhoed, J. S.,
13.
Schauer, R., Risgaard Petersen, N., Kjeldsen, K. U.,
De Wael, K., Beaumont, H. J. E., Bart Cleuren, B., Valcke, R.,
Bjerg, J. J. T., Jørgensen, B. B., Schramm, A., and
van der Zant, H. S. J., Boschker, H. T. S., and Manca, J. V.
Nielsen, L. P. (2014) Succession of cable bacteria and elec
(2019) A highly conductive fibre network enables centime
tric currents in marine sediment, ISME J., 8, 1314 1322.
tre scale electron transport in multicellular cable bacteria,
14.
Gorby, Y. A., Yanina, S., McLean, J. S., Rosso, K. M.,
Nat.Commun., 10, 1 8.
Moyles, D., Dohnalkova, A., Beveridge, T. J., Chang, I. S.,
28.
Bonné, R., Hou, J. L., Hustings, J., Meert, M., Hidalgo
Kim, B. H., Kim, K. S., Culley, D. E., Reed, S. B.,
Martinez, S., Cornelissen, R., D’Haen, J., Thijs, S.,
Romine, M. F., Saffarini, D. A., Hill, E. A., Shi, L.,
Vangronsveld, J., Valcke, R., Cleuren, B., Meysman, F. J. R.,
Elias, D. A., Kennedy, D. W., Pinchuk, G., Watanabe, K.,
and Manca, J. V. (2019) Cable bacteria as long range bio
Ishii, S., Logan, B., Nealson, K. H., and Fredrickson, J. K.
logical semiconductors, arXiv Preprint arXiv:191206224.
(2006) Electrically conductive bacterial nanowires pro
29.
Trojan, D., Schreiber, L., Bjerg, J. T., Bøggild, A., Yang, T.,
duced by Shewanella oneidensis strain MR 1 and other
Kjeldsen, K. U., and Schramm, A. (2016) A taxonomic
microorganisms, Proc. Natl Acad. Sci. USA, 103, 11358
framework for cable bacteria and proposal of the candidate
11363.
genera Electrothrix and Electronema, Syst. Appl. Microbiol.,
15.
Filman, D. J., Marino, S. F., Ward, J. E., Yang, L., Mester, Z.,
39, 297 306.
Bullitt, E., Lovley, D. R., and Strauss, M. (2019) Cryo EM
30.
Parge, H. E., Forest, K. T., Hickey, M. J., Christensen, D. A.,
reveals the structural basis of long range electron transport
Getzoff, E. D., and Tainer, J. A. (1995) Structure of the
in a cytochrome based bacterial nanowire, Commun. Biol.,
fibre forming protein pilin at 2.6 Å resolution, Nature, 378,
2, 219, doi: 10.1038/s42003 019 0448 9.
32 38.
16.
Tan, Y., Adhikari, R. Y., Malvankar, N. S., Ward, J. E.,
31.
Craig, L., Taylor, R. K., Pique, M. E., Adair, B. D., Arvai, A. S.,
Woodard, T. L., Nevin, K. P., and Lovley, D. R. (2017)
Singh, M., Lloyd, S. J., Shin, D. S., Getzoff, E. D., Yeager,
Expressing the Geobacter metallireducens PilA in Geobacter
M., Forest, K. T., and Tainer, J. A. (2003) Type IV pilin
sulfurreducens yields pili with exceptional conductivity,
structure and assembly: X ray and EM analyses of Vibrio
mBio, 8, e02203 6, doi: 10.1128/mBio.02203 16.
cholerae toxin coregulated pilus and Pseudomonas aerugi
17.
Adhikari, R. Y., Malvankar, N. S., Tuominen, M. T., and
nosa PAK pilin, Mol. Cell, 11, 1139 1150.
Lovley, D. R. (2016) Conductivity of individual Geobacter
32.
Mehta, T., Coppi, M. V., Childers, S. E., and Lovley, D. R.
pili, RSC Adv., 6, 8354 8357.
(2005) Outer membrane c type cytochromes required for
18.
Lampa Pastirk, S., Veazey, J. P., Walsh, K. A., Feliciano, G. T.,
Fe (III) and Mn (IV) oxide reduction in Geobacter sulfurre
Steidl, R. J., Tessmer, S. H., and Reguera, G. (2016)
ducens, Appl. Environ. Microbiol., 71, 8634 8641.
Thermally activated charge transport in microbial protein
33.
Leang, C., Qian, X., Mester, T., and Lovley, D. R. (2010)
nanowires, Sci. Rep., 6, 23517, doi: 10.1038/srep23517.
Alignment of the c type cytochrome OmcS along pili of
19.
Потапова Т., Кокшарова О. (2020) Нитчатые циано
Geobacter sulfurreducens, Appl. Environ. Microbiol., 76,
бактерии как прототип многоклеточных организмов,
4080 4084.
Физиология Растений, 67, 20 34.
34.
Inoue, K., Leang, C., Franks, A. E., Woodard, T. L.,
20.
Tan, Y., Adhikari, R. Y., Malvankar, N. S., Pi, S., Ward, J. E.,
Nevin, K. P., and Lovley, D. R. (2011) Specific localization
Woodard, T. L., Nevin, K. P., Xia, Q., Tuominen, M. T.,
of the c type cytochrome OmcZ at the anode surface in
and Lovley, D. R. (2016) Synthetic biological protein
current producing biofilms of Geobacter sulfurreducens,
nanowires with high conductivity, Small, 12, 4481 4485.
Environ. Microbiol. Rep., 3, 211 217.
21.
El Naggar, M. Y., Wanger, G., Leung, K. M., Yuzvinsky, T. D.,
35.
Qian, X., Reguera, G., Mester, T., and Lovley, D. R. (2007)
Southam, G., Yang, J., Lau, W. M., Nealson, K. H., and
Evidence that OmcB and OmpB of Geobacter sulfurre
Gorby, Y. A. (2010) Electrical transport along bacterial
ducens are outer membrane surface proteins, FEMS
nanowires from Shewanella oneidensis MR 1, Proc. Natl
Microbiol. Lett., 277, 21 27.
Acad. Sci. USA, 107, 18127 18131.
36.
Shi, L., Richardson, D. J., Wang, Z., Kerisit, S. N.,
22.
Sato, M., and Mooney, H. M. (1960) The electrochemical
Rosso, K. M., Zachara, J. M., and Fredrickson, J. K.
mechanism of sulfide self potentials, Geophysics, 25, 226
(2009) The roles of outer membrane cytochromes of
249.
Shewanella and Geobacter in extracellular electron transfer,
23.
Bigalke, J., and Grabner, E. W. (1997) The geobattery
Environ. Microbiol. Rep., 1, 220 227.
model - a contribution to large scale electrochemistry,
37.
Pirbadian, S., Barchinger, S. E., Leung, K. M., Byun, H. S.,
Electrochimica Acta, 42, 3443 3452.
Jangir, Y., Bouhenni, R. A., Reed, S. B., Romine, M. F.,
24.
Naudet, V., and Revil, A. (2005) A sandbox experiment to
Saffarini, D. A., Shi, L., Gorby, Y. A., Golbeck, J. H., and
investigate bacteria mediated redox processes on self
El Naggar, M. Y. (2014) Shewanella oneidensis MR 1
potential
signals,
Geophys.
Res.
Lett.,
32,
nanowires are outer membrane and periplasmic extensions
doi: 10.1029/2005GL022735.
of the extracellular electron transport components, Proc.
25.
Dueholm, M. S., Larsen, S., Risgaard Petersen, N.,
Natl. Acad. Sci. USA, 111, 12883 12888.
Nierychlo, M., Schmid, M., Bøggild, A., van de
38.
Malvankar, N. S., Vargas, M., Nevin, K. P., Franks, A. E.,
Vossenberg, J., Geelhoed, J. S., Meysman, F. J. R., Wagner,
Leang, C., Kim, B. C., Inoue, K., Mester, T., Covalla, S. F.,
БИОХИМИЯ том 85 вып. 8 2020
10*
1132
ПТУШЕНКО
Johnson, J. P., Rotello, V. M., Tuominen, M. T., and
nanowires of Geobacter sulfurreducens, Energy Environ.
Lovley, D. R. (2011) Tunable metallic like conductivity in
Sci., 5, 8651 8659.
microbial nanowire networks, Nat. Nanotechnol., 6, 573
45. Polizzi, N. F., Skourtis, S. S., and Beratan, D. N. (2012)
579.
Physical constraints on charge transport through bacterial
39.
Strycharz Glaven, S. M., Snider, R. M., Guiseppi Elie, A.,
nanowires, Faraday Discuss., 155, 43 61.
and Tender, L. M. (2011) On the electrical conductivity of
46. Wang, F., Gu, Y., O’Brien, J. P., Sophia, M. Y., Yalcin, S. E.,
microbial nanowires and biofilms, Energy Environ. Sci., 4,
Srikanth, V., Shen, C., Vu, D., Ing, N. L., Hochbaum, A. I.,
4366 4379.
Egelman, E. H., and Malvankar, N. S. (2019) Structure of
40.
Boesen, T., and Nielsen, L. P. (2013) Molecular dissection
microbial nanowires reveals stacked hemes that transport
of bacterial nanowires, mBio, 4, e00270 13, doi: 10.1128/
electrons over micrometers, Cell, 177, 361 369.
mBio.00270 13.
47. Bjerg, J. T., Boschker, H. T. S., Larsen, S., Berry, D.,
41.
Vargas, M., Malvankar, N. S., Tremblay, P. L., Leang, C.,
Schmid, M., Millo, D., Tataru, P., Meysman, F. J. R.,
Smith, J. A., Patel, P., Synoeyenbos West, O., Nevin, K. P.,
Wagner, M., Nielsen, L. P., and Schramm, A. (2018) Long
and Lovley, D. R. (2013) Aromatic amino acids required
distance electron transport in individual, living cable bac
for pili conductivity and long range extracellular electron
teria, Proc. Natl. Acad. Sci. USA, 115, 5786 5791.
transport in Geobacter sulfurreducens, mBio, 4, e00105 13,
48. Chailakhyan, L. M., Glagolev, A. N., Glagoleva, T. N.,
doi: 10.1128/mBio.00105 13.
Murvanidze, G. V., Potapova, T. V., and Skulachev, V. P.
42.
Cosert, K. M., Castro Forero, A., Steidl, R. J., Worden, R. M.,
(1982) Intercellular power transmission along trichomes of
and Reguera, G. (2019) Bottom up fabrication of protein
cyanobacteria, Biochim. Biophys. Acta, 679, 60 67.
nanowires via controlled self assembly of recombinant Geo
49. Ishii, S., Kosaka, T., Hori, K., Hotta, Y., and Watanabe, K.
bacter pilins, mBio, 10, e02721 19, doi: 10.1128/mBio. 02721 19.
(2005) Coaggregation facilitates interspecies hydrogen
43.
Holmes, D. E., Chaudhuri, S. K., Nevin, K. P., Mehta, T.,
transfer between Pelotomaculum thermopropionicum and
Methé, B. A., Liu, A., Ward, J. E., Woodard, T. L.,
Methanothermobacter thermautotrophicus, Appl. Environ.
Webster, J., and Lovley, D. R. (2006) Microarray and
Microbiol., 71, 7838 7845.
genetic analysis of electron transfer to electrodes in Geo
50. Summers, Z. M., Fogarty, H. E., Leang, C., Franks, A. E.,
bacter sulfurreducens, Environ. Microbiol., 8, 1805 1815.
Malvankar, N. S., and Lovley, D. R. (2010) Direct
44.
Malvankar, N. S., Tuominen, M. T., and Lovley, D. R.
exchange of electrons within aggregates of an evolved syn
(2012) Lack of cytochrome involvement in long range
trophic coculture of anaerobic bacteria, Science, 330,
electron transport through conductive biofilms and
1413 1415.
ELECTRIC CABLES OF LIVING CELLS.
II. BACTERIAL ELECTRON CONDUCTORS
Review
V. V. Ptushenko1,2
1 Belozersky Institute of Physico Chemical Biology, Lomonosov Moscow State University,
119992 Moscow, Russia; E mail: ptush@belozersky.msu.ru
2 Emanuel Institute of Biochemical Physics of Russian Academy of Sciences, 119334 Moscow, Russia
Received May 17, 2020
Revised July 12, 2020
Accepted July 16, 2020
The concept of “electric cables” involved in bioenergetic processes of a living cell was proposed half a century ago
[Skulachev, V. P. (1971) Curr. Top. Bioenerg., Elsevier, pp. 127 190]. For many decades, only cell membrane structures
have been considered as probable pathways for the electric current, namely, for the transfer of transmembrane elec
trochemical potential. However, the last ten to fifteen years have brought the discovery of bacterial “electric cables”
of a new type. In 2005, “nanowires” conducting electric current over distances of tens of micrometers were discov
ered in metal and sulphate reducing bacteria [Reguera, G. et al. (2005) Nature, 435, pp. 1098 1101]. The next five
years have witnessed the discovery of microbial electric currents over centimeter distances [Nielsen, L. P. et al. (2010)
Nature, 463, 1071 1074]. This new group of bacteria allowing electric currents to flow over macroscopic distances was
later called cable bacteria. Nanowires and conductive structures of cable bacteria serve to solve a special problem of
membrane bioenergetics: they connect two redox half reactions. In other words, unlike membrane “cables”, their
function is electron transfer in the course of oxidative phosphorylation for the generation of membrane energy rather
than of the end product. The most surprising is the protein nature of these cables (at least of some of them) indicat
ed by recent data, since no protein wires for the long distance electron transport had been previously known in living
systems.
Keywords: The concept of “electric cables” of a cell, nanowires, Geobacter sulfurreducens, cable bacteria
БИОХИМИЯ том 85 вып. 8 2020