БИОХИМИЯ, 2021, том 86, вып. 12, с. 1739 - 1765
УДК 577.24
КОЭВОЛЮЦИЯ МОЗГА, КУЛЬТУРЫ
И ПРОДОЛЖИТЕЛЬНОСТИ ЖИЗНИ:
РЕЗУЛЬТАТЫ КОМПЬЮТЕРНОГО МОДЕЛИРОВАНИЯ
© 2021
А.В. Марков1,2*, М.А. Марков1
1 Московский государственный университет имени М.В. Ломоносова, биологический факультет,
119991 Москва, Россия; электронная почта: markov_a@inbox.ru
2 Палеонтологический институт РАН имени А.А. Борисяка, 117997 Москва, Россия
Поступила в редакцию 22.10.2021
После доработки 04.11.2021
Принято к публикации 04.11.2021
Люди обладают рядом признаков, редких или отсутствующих у других приматов, включая большой размер
мозга, культуру, язык, увеличенную продолжительность жизни (ПЖ) и длительный пострепродуктивный
период жизни. В работе мы использовали компьютерную модель TribeSim, изначально разработанную для
исследования автокаталитической совместной эволюции мозга и культуры у гоминин в рамках теории
«культурного драйва», чтобы выяснить, как культура и мозг могут эволюционировать вместе с ПЖ (или ско%
ростью старения). Мы показали, что в отсутствие культуры эволюция ПЖ зависит от интенсивности меж%
групповой конкуренции (МГК): сильная МГК приводит к более короткой ПЖ. Культура, однако, способ%
ствует генетической эволюции большей ПЖ даже при сильной МГК. Увеличение ПЖ, в свою очередь, спо%
собствует культурному развитию, создавая тем самым положительную обратную связь. Культурная эволю%
ция ПЖ (накопление знаний, способствующих или препятствующих выживанию) отличается от генетичес%
кой эволюции того же признака, частично потому, что «мемы» (идеи, навыки и варианты поведения), сни%
жающие риск смерти, имеют тенденцию накапливаться в пуле мемов, даже если они не приносят пользу ге%
нам. Следовательно, культурная эволюция старения имеет тенденцию приводить к большей ПЖ, чем гене%
тическая эволюция того же признака. Если ПЖ эволюционирует как генетически, так и культурно, типич%
ным результатом является общество, в котором молодые особи из%за своей генетической предрасположен%
ности ведут более рискованный образ жизни в обмен на возможность получить дополнительные ресурсы,
но с возрастом они накапливают полезные для выживания знания. Моделирование также показало, что
культурная эволюция адаптивного поведения может способствовать генетической эволюции длительного
пострепродуктивного периода жизни, например, если присутствие знающих долгожителей увеличивает
конкурентоспособность группы.
КЛЮЧЕВЫЕ СЛОВА: эволюция продолжительности жизни, старение, пострепродуктивный период, ко%
эволюция генов и культуры, культурный драйв, быстрая коэволюция мозга и культуры.
DOI: 10.31857/S0320972521120010
ВВЕДЕНИЕ
тельный пострепродуктивный период жизни,
особенно ярко выраженный у женщин [8]. Как и
Люди отличаются от других приматов во
почему эти признаки эволюционировали совме%
многих важных отношениях, включая большой
стно в ходе антропогенеза, является предметом
размер мозга, кумулятивную культуру, обеспе%
дискуссий [9-13]. В работе мы использовали
чиваемую языком или, в более широком смыс%
компьютерное моделирование для изучения
ле, улучшенными средствами социального обу%
возможных моделей эволюционного взаимо%
чения [1-3], увеличением продолжительности
действия между культурой, социальным обуче%
жизни (ПЖ) [4, 5] (вероятно, по причине «нео%
нием, размером мозга и ПЖ у социальных видов
тенического» замедления старения [6, 7]) и дли%
с высокой групповой сплоченностью, совмест%
ной добычей ресурсов, парными связями, высо%
кими родительскими инвестициями в потом%
Принятые сокращения: МГК - межгрупповая кон%
куренция; ОП - объем памяти; ПЖ - продолжительность ство и базовыми способностями к культурной
жизни; Сст - скорость старения; ЭО - эффективность передаче адаптивного поведения. Все эти приз%
охоты (полезный для группы поведенческий признак);
наки, вероятно, присутствовали у плейстоцено%
ЭОб - эффективность обучения; ЭЭП - эффективность
эгоистичного поступка (индивидуально полезный пове%
вых видов Homo и их прямых предков [14-16].
денческий признак); G - максимальный размер группы.
Объем мозга увеличился в три раза за пос%
* Адресат для корреспонденции.
ледние два миллиона лет эволюции Homo, не%
1739
1740
МАРКОВ А., МАРКОВ М.
смотря на очевидно высокие затраты [17-22].
уровень выживания и более сильный отбор на
Этот эволюционный паттерн быстрого увеличе%
усиление социального обучения [12, 29, 30].
ния размеров мозга, по%видимому, количествен%
Есть основания полагать, что ПЖ человека уве%
но уникален среди приматов [23]. Эти факты
личилась (а Сст уменьшилась) в ходе эволюции,
подразумевают, что отбор на увеличение разме%
и что это было связано как с генетическими из%
ров мозга в ходе эволюции человека, вероятно,
менениями, так и с развитием культуры [4, 6-7].
был обусловлен сильной положительной обрат%
Этот гипотетический механизм положитель%
ной связью, а не простыми экологическими
ной обратной связи в эволюции мозга, социаль%
факторами, такими как климат или доступность
ного обучения и ПЖ согласуется с тем фактом,
различных пищевых ресурсов (более подробное
что существуют значительные положительные
обсуждение см. в работе [13]). В текущем иссле%
связи между долголетием, объемом мозга (как
довании мы основываемся на одной из наиболее
абсолютным, так и относительным), склон%
обоснованных теорий, предложенных для объ%
ностью к социальному обучению и технически%
яснения необычайно быстрого увеличения моз%
ми инновациями у приматов [31-33] и китооб%
га у Homo: теории «культурного драйва» или
разных [34]. Несмотря на то что в настоящее
«культурного мозга», предполагающей, что сов%
время большое внимание уделяется исследова%
местная эволюция мозга, социального обучения
ниям эволюции параметров жизненного цикла в
и культуры может быть самоподдерживающейся
целом [35, 36], закономерности и последствия
или автокаталитической [1, 12]. В самой прос%
возможного коэволюционного взаимодействия
тейшей форме механизм положительной обрат%
между ПЖ и развитостью мозга, социальным
ной связи культурного драйва можно описать
обучением и культурой изучены недостаточно.
следующим образом: лучшее социальное обуче%
Для того чтобы исследовать эти закономер%
ние и познание → большее количество поведен%
ности, мы использовали компьютерную модель
ческих инноваций закрепляется в качестве
TribeSim, которая первоначально была разра%
культурных традиций; более богатая культура →
ботана для имитации динамики коэволюции
больше полезных навыков, которые можно пе%
мозга и культуры у высокосоциальных видов.
ренять от сородичей; повышенная полезность
Подробное описание модели и ее сравнение с
способностей к социальному обучению → более
отдаленно похожей моделью
«Культурный
сильный отбор на еще лучшее социальное обу%
мозг» [12] было опубликовано ранее [13]. Моде%
чения и познание. Увеличение размеров мозга
лирование подтвердило релевантность культур%
является побочным эффектом этого отбо%
ного драйва у социальных видов в социально%
ра [11-13, 24-27].
экологической ситуации, делающей возмож%
В этом контексте возможно существование
ным случайное изобретение нового полезного и
дополнительной петли положительной обрат%
требующего хороших когнитивных способнос%
ной связи через увеличение ПЖ (или замедле%
тей поведения. Культурный драйв, скорее всего,
ние старения) [1, 11, 13, 28]. ПЖ (или скорость
будет запущен, если некоторые из культурно
старения, Сст) может эволюционировать как ге%
передаваемых видов поведения индивидуально
нетически, так и культурно. В первом случае
полезны, в то время как другие полезны для
ожидается, что развитая культура будет способ%
группы. В этом случае культурный драйв возмо%
ствовать отбору на более медленное старение,
жен при различных уровнях межгрупповой
потому что у дольше живущих индивидов будет
конкуренции (МГК) и миграции. Моделирова%
больше времени, чтобы учиться у своих товари%
ние также показало, что увеличение размера
щей по группе и обучать других (или служить
мозга может получить дополнительный им%
образцом для подражания). Другими словами,
пульс, если развивающиеся механизмы соци%
индивиды, живущие дольше, извлекают больше
ального обучения являются дорогостоящими с
выгоды из культуры и в то же время лучше на%
точки зрения увеличения размера мозга (напри%
капливают и распространяют знания. Следова%
мер, опираются на сложные нейронные цепи) и
тельно, ожидается, что генетическая эволюция
устойчивыми к сложности передаваемой ин%
большей ПЖ будет способствовать культурному
формации, т.е. позволяют легко передавать
развитию, что, в свою очередь, может способ%
непростые навыки и концепции. Человеческий
ствовать отбору на увеличение ПЖ. Во втором
язык, вероятно, подходит под это описание.
случае (культурная эволюция ПЖ) возможна
Моделирование также подтвердило, что увели%
следующая петля обратной связи: развитая
чение ПЖ может ускорить коэволюцию мозга и
культура → повышенная выживаемость → бóль%
культуры [13]. Это согласуется с идеей о том, что
шая ПЖ → лучшие предпосылки для передачи
петля положительной обратной связи через уве%
знаний от поколения к поколению → еще более
личение ПЖ может еще сильнее способствовать
прогрессивная культура → еще более высокий
культурному развитию и увеличению размера
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1741
мозга. Тем не менее компьютерные симуляции
Более высокие значения ЭО приносят поль%
не показали, может ли культурная эволюция (и
зу группе (ЭО - «кооперативный признак»), по%
при каких обстоятельствах) вносить вклад в эво%
тому что количество ресурсов, приобретаемых
люцию увеличения (или снижения) ПЖ. В нас%
группой, пропорционально сумме индивидуаль%
тоящей работе мы использовали модифициро%
ных ЭО всех членов группы, которые идут на
ванную версию TribeSim для изучения эволю%
охоту («охотничье усилие» группы). Чем выше
ции ПЖ и старения (определяемого здесь как
охотничье усилие группы, тем меньше доля дру%
увеличение уровня смертности с возрастом) в
гих групп. Это приводит к межгрупповой конку%
контексте быстрой совместной эволюции куль%
ренции, которая, предположительно, была до%
туры и мозга.
вольно сильной у плейстоценовых гоминин
[11]. Внутри групп ресурсы распределяются в
соответствии с индивидуальными значениями
МАТЕРИАЛЫ И МЕТОДЫ
ЭЭП. ЭЭП - это «эгоистичный признак»: высо%
кая ЭЭП приносит пользу отдельному индиви%
TRIBESIM: описание модели. Подробное
ду, но не группе.
описание было опубликовано ранее [13], поэто%
Здесь мы вводим еще один фенотипический
му здесь мы ограничимся кратким изложением
признак, Сст (скорость старения), который мо%
основных принципов.
жет развиваться генетически и/или культурно, и
TribeSim - симуляционная модель с анали%
новую категорию мемов (мемы Сст). Сст%мемы
зом на уровне индивида, предназначенная для
влияют на возрастной риск смерти и могут как
имитации генетической и культурной эволю%
увеличивать, так и снижать выживаемость. Хотя
ции в популяции высокосоциальных животных.
Сст не оказывает прямого влияния на исход
Популяция состоит из конкурирующих соци%
конкуренции за ресурсы, она может влиять на
альных групп. Максимально возможный размер
него косвенно (см. ниже).
группы определяется параметром G; по дости%
В TribeSim отбор действует на трех уровнях:
жении этого предела группа разделяется на две
1) Групповой отбор. МГК за ресурсы и изби%
половины. Члены группы предпринимают сов%
рательное выживание, рост и разделение групп
местные усилия по извлечению ресурсов из ок%
приводят к групповому отбору, который способ%
ружающей среды, конкурируя с другими груп%
ствует развитию кооперативных признаков [41,
пами. Мы назвали это «коллективной охотой»
42]. Мы можем регулировать интенсивность
[37], хотя в этом контексте можно рассматри%
группового отбора, изменяя параметр G. На%
вать и другие виды сотрудничества, принося%
пример, когда G = 15, есть много небольших
щие пользу группе. Ресурсы, приобретенные
конкурирующих групп, МГК очень интенсивна,
группой, распределяются затем между членами
а групповой отбор силён. Когда G превышает
этой группы. Индивиды конкурируют друг с
максимально возможный размер популяции
другом, чтобы увеличить свою долю. Таким об%
(например, G = 2000), тогда вся популяция
разом, существует два уровня конкуренции за
представляет собой единую группу, и групповой
ресурсы: между группами и между отдельными
отбор не идет.
особями, что похоже на модель «перетягивания
2) Индивидуальный отбор. Внутригрупповая
каната», которая ранее использовалась для изу%
конкуренция за ресурсы приводит к избиратель%
чения влияния МГК на эволюцию внутригруп%
ному выживанию и размножению особей. Ин%
повой кооперации [38]. Результат конкуренции
дивидуальный отбор способствует развитию
зависит от двух поведенческих признаков, ко%
«эгоистичных признаков», которые приносят
торые могут развиваться генетически и (или)
пользу отдельному индивиду и могут быть вред%
культурно: «эффективности охоты» (ЭО) и «эф%
ными или нейтральными для группы. ЭЭП
фективности эгоистичного поступка» [«макиа%
обычно нейтральна для группы, потому что уси%
веллистского» поступка, поступка в духе Макиа%
ленное размножение особей с более высокой
велли, коварного, эгоистичного поступка, далее
ЭЭП компенсирует плохое воспроизводство
ЭЭП] [39, 40]. Оба признака зависят от культур%
особей с более низкой ЭЭП.
но передаваемых навыков («мемов»), которые
3) Отбор мемов. Мемы конкурируют за до%
могут быть изобретены с нуля (с фиксирован%
минирование в индивидуальной памяти и в
ной низкой вероятностью) или приобретены у
культуре группы (пул мемов). Отбор мемов от%
товарищей по группе через социальное обуче%
дает предпочтение мемам, которые распростра%
ние. Все индивиды рождаются с генетически
няются быстрее (тем, которые легче выучить
детерминированными начальными значениями
или которые требуют меньшего объема памяти
ЭО и ЭЭП (10 и 0 соответственно) и без мемов
для запоминания) [24]. Судьба мема также зави%
в памяти.
сит от его влияния на индивидуальные и груп%
БИОХИМИЯ том 86 вып. 12 2021
1742
МАРКОВ А., МАРКОВ М.
повые фенотипы. Некоторые категории мемов
ЭОб влияет на вероятность успешной передачи
могут приобретать «вирусные свойства», если
мема независимо от его размера. Причины это%
они помогают себе распространяться (т.е. если
го обсуждаются в [13]; предполагается, что ЭОб
они делают своего носителя более эффективной
при таком моделировании помогает понять эво%
машиной для распространения мемов). Обуча%
люцию языковых способностей.
ющие (учительские) навыки - один из приме%
Индивидуальный фенотип включает пять
ров таких мемов [13].
вариабельных признаков: ЭО (рассчитывается
Особи диплоидные и размножаются поло%
как сумма генотипического значения ЭО и эф%
вым путем. Пары формируются случайным об%
фективности всех известных индивиду мемов
разом в пределах групп (межгрупповая миграция
ЭО), ЭЭП (генотипическое значение ЭЭП плюс
представляет собой отдельный процесс); пара
суммарная эффективность всех известных ме%
дает одного потомка, если у родителей достаточ%
мов ЭЭП), Сст (рассчитывается аналогично;
но ресурсов; оба родителя вкладываются в пото%
эффективность мемов Сст может быть как поло%
мство; пары формируются каждый год заново.
жительной, так и отрицательной; это соответ%
Эволюционирующая часть генотипа вклю%
ствует мемам, ухудшающим и повышающим вы%
чает гены, отвечающие за объем памяти (ОП),
живаемость); ЭОб (определена генетически,
эффективность обучения (ЭОб), ЭО, ЭЭП и
ЭОб в целом аналогична точности социального
Сст. Каждому гену можно присвоить постоян%
обучения, которая, как полагают, важна для ко%
ное значение (в этом случае признак не может
эволюции мозга и культуры [1, 2, 12]; ОП (опре%
развиваться генетически) или разрешить мути%
делен генетически).
ровать и, следовательно, эволюционировать.
Жизнь моделируемой популяции состоит из
Каждая копия гена имеет «значение», которое
шагов (лет). Следующие события проходят каж%
напрямую транслируется в фенотип: например,
дый год:
если начальное значение гена ОП равно 0, тогда
1) Трата ресурсов на жизнеобеспечение. В
все индивидуумы в популяции изначально име%
новой версии TribeSim стоимость жизнеобеспе%
ют нулевую емкость памяти; гетерозиготный
чения зависит от Сст: нужно потратить допол%
индивидуум с двумя копиями гена ОП со значе%
нительные ресурсы, чтобы снизить риск смерти
ниями 0 и 0,2 имеет ОП, равный 0,1.
(например, сделав укрытие или одежду); в про%
Генотипические значения ОП и ЭОб связа%
тивном случае можно сэкономить немного ре%
ны с объемом мозга: их рост приводит к увели%
сурсов, ведя более рискованный образ жизни.
чению размера мозга (по умолчанию объем моз%
Это автоматически приводит к классическому
га равен 20 + ОП + 30 × ЭОб). Это согласуется с
компромиссу между размножением и выжива%
эмпирическими корреляциями между объемом
нием [45], потому что дополнительные ресурсы,
мозга и социальным обучением у приматов [32,
сэкономленные подверженным риску индиви%
33], китообразных [34] и, предположительно,
дуумом, скорее всего, будут потрачены на раз%
птиц [43]. Объем мозга - дорогостоящий приз%
множение. Введение ресурсной «платы» за по%
нак, потому что количество ресурсов, необходи%
вышение выживаемости важно для целей нас%
мых для рождения ребенка, пропорционально
тоящего исследования, потому что (i) в против%
объему мозга ребенка. Это согласуется с идеей,
ном случае отбор всегда будет отдавать пред%
что родительские инвестиции значительно уве%
почтение наименьшей возможной Сст, и (ii) все
личиваются в ходе эволюции гоминин наряду с
ограничения в TribeSim (например, ограниче%
энергетическими и когнитивными потребнос%
ния на рождаемость и увеличение размера моз%
тями быстрорастущего детского мозга [18, 44].
га) определяются в основном ограниченностью
Мемы хранятся в памяти и влияют на фено%
ресурсов, поэтому логично, что повышенная
типические признаки ЭО, ЭЭП и Сст. Мемы
выживаемость также должна быть ограничена
придумываются редко, их можно передать через
ресурсами. Идея затрат ресурсов на повышение
социальное обучение и/или забыть. Каждый
выживаемости ярко иллюстрируется историей
мем характеризуется своей категорией (ЭО,
трех поросят: два из них сэкономили свои ре%
ЭЭП или Сст), размером (величиной ОП, необ%
сурсы, построив дома из соломы и прутьев, но
ходимого для хранения мема) и эффектив%
рисковали своими жизнями, а третий потратил
ностью (изменением фенотипического призна%
много ресурсов на кирпичный дом, увеличивая
ка индивида, который знает мем). Размер и эф%
тем самым свои шансы на выживание. Стои%
фективность положительно коррелируют, но
мость жизнеобеспечения в год составляет
корреляция слаба [24]. Большие мемы могут
40 × (0,3 - Сст)2 + 2. Эта формула гарантирует,
быть изучены только особями с достаточным
что по мере замедления старения дальнейшая
количеством свободного ОП; таким образом,
эволюция в этом направлении становится все
размер мема ограничивает его распространение.
более дорогостоящей.
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1743
2) Спонтанное изобретение новых мемов.
пара сформирована, проверяется возможность
Индивид изобретает мем данной категории с
рождения ребенка. Чтобы произвести на свет
фиксированной низкой вероятностью (0,0001 в
ребенка, родители должны потратить количест%
год) независимо от количества разрешенных ка%
во ресурсов, равное предполагаемому объему
тегорий мемов.
мозга ребенка, умноженному на два. Сорок про%
3) Самопроизвольное забывание мемов.
центов этих ресурсов передаются ребенку. Если
Каждый индивид может забыть любой мем с ве%
у обоих родителей вместе не хватает ресурсов,
роятностью 0,01 в год.
попытка произвести на свет ребенка не удастся.
4) Коллективная охота. Все особи, обладаю%
После того как ребенок родился, при условии,
щие достаточными ресурсами, отправляются на
что у родителей остались какие%то ресурсы, 40%
охоту; стоимость действия - 2 ресурса.
из них также передаются ребенку, а оставшаяся
5) Раздел ресурсов. Ресурсы, полученные
часть распределяется поровну между родителя%
группой, распределяются между всеми членами
ми. Равенство родителей в TribeSim напоминает
группы. Это согласуется с традиционным пове%
предполагаемое усиление отцовской заботы,
дением некоторых охотников%собирателей [46],
снижение полового диморфизма и тенденцию к
и даже шимпанзе часто делятся мясом после ус%
моногамии и кооперативному размножению у
пешной охоты [47]. По умолчанию доли всех
гоминин [14, 16, 50]. Для простоты смоделиро%
равны. Однако если есть индивиды с ЭЭП > 0,
ванные особи в TribeSim не имеют фиксирован%
они выполняют «макиавеллистские трюки» [ма%
ного пола; любые две особи могут образовать
нипуляторские, коварные, эгоистичные поступ%
пару и произвести потомство.
ки], чтобы претендовать на бóльшую долю. За%
9) Деление групп. Если группа превышает
тем ресурсы распределяются согласно индиви%
свой верхний предел G, она распадается на две
дуальным значениям ЭЭП.
равные группы.
6) Обучение. Каждый индивид случайным
10) Межгрупповая миграция. Индивид мо%
образом выбирает себе товарища по группе и
жет покинуть свою группу и присоединиться к
пытается выучить имеющийся в его памяти мем.
другой (случайно выбранной) группе с опреде%
Мем выбирается случайным образом из мемов,
ленной вероятностью (0,001 в год по умолча%
известных потенциальному учителю, но не уче%
нию).
нику. Если таких мемов нет, или размер выбран%
Значения параметров (таблица) были выбра%
ного мема превышает размер свободного ОП
ны произвольно на основе логики модели, а так%
обучаемого, попытка не удалась. В противном
же опыта, полученного в результате предвари%
случае вероятность успеха равна ЭОб ученика.
тельных прогонов модели (например, генети%
7) Смерть. В новой версии TribeSim мы ис%
ческое значение ЭО по умолчанию было уста%
пользуем закон Гомпертца в его простейшей
новлено на 10, чтобы гарантировать, что попу%
форме: вероятность смерти в течение текущего
ляция может выжить даже в отсутствие адаптив%
года равна НУС × exp(Сст × (возраст-1)), где НУС
ных знаний). Не было предпринято никаких по%
(начальный уровень смертности) является конс%
пыток смоделировать какие%либо реальные ви%
тантой (в данном исследовании НУС = 0,008);
ды приматов или получить количественно точ%
Сст - фенотипический признак, который может
ные прогнозы.
развиваться генетически и культурно; возраст -
это возраст индивида в годах. Кроме того, инди%
вид может умереть от голода, если он или она не
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
имеет достаточно ресурсов для жизнеобеспече%
ния в течение двух лет подряд (один голодный
Генетическая эволюция скорости старения
год часто следует за рождением ребенка и не яв%
(Сст) и других адаптивных признаков (ЭО и
ляется смертельным).
ЭЭП). В первой серии симуляций мы отключи%
8) Размножение. Каждый индивид старше
ли культуру (вероятность изобретения мема бы%
шести лет пытается создать пару с другим чле%
ла установлена на ноль), чтобы исследовать вза%
ном группы и произвести на свет ребенка. Хотя
имное влияние генетической эволюции Сст и
возраст созревания имеет тенденцию зависеть
других адаптивных признаков. Некоторые ис%
от ПЖ [48, 49], в центре внимания настоящего
следователи утверждают, что старение может
исследования находится эволюция ПЖ; поэто%
развиваться как адаптация, направленная на по%
му для простоты мы смоделировали фиксиро%
вышение эволюционируемости, т.е. на ускоре%
ванный возраст созревания. Пары образуются
ние адаптивной эволюции других признаков [7,
только на один год (серийная моногамия). Если
51-54]. Один из гипотетических механизмов та%
в группе нет свободных, еще не образовавших
кого ускорения - более короткое время смены
пару особей, попытка не удалась. После того как
поколений. Мы задались вопросом, как быстро
БИОХИМИЯ том 86 вып. 12 2021
1744
МАРКОВ А., МАРКОВ М.
Значения параметров по умолчанию
Параметр
Значение по
Комментарии
умолчанию
Ресурсы окружающей среды,
6000
максимальный объем ресурсов, который можно получить из окру%
в год (R)
жающей среды в год; устанавливает ограничение на размер попу%
ляции. При параметрах по умолчанию размер популяции составля%
ет ~ 700-800 особей
Частота мутаций гена, ответ%
0,04; 0; 0,4
ген ОП мутирует с вероятностью 0,04 при передаче от родителя к
ственного за объем памяти
ребенку; к значению гена добавляется эффект мутации; если ре%
(ОП), среднее значение эф%
зультирующее значение выходит за пределы допустимого диапазо%
фекта, StDev
на (если ОП становится отрицательным), мутация отменяется и
попытка повторяется
Скорость мутирования гена
0,04; 0; 0,1
ЭОб варьирует от 0 (нулевой шанс узнать мем) до 1 (100%%ный
эффективности обучения (ЭОб),
успех)
среднее значение эффекта,
StDev
Скорость мутирования гена
0,04; 0; 0,002
при параметрах по умолчанию Сст обычно находится в диапазоне
скорости старения (Сст),
от 0 (старение отсутствует) до 0,25 (очень быстрое старение)
среднее значение эффекта,
StDev
Частота мутаций гена эффек%
0,002; 0; 0,5
тивности охоты (ЭО), среднее
значение эффекта, StDev
Частота мутаций гена эффек%
0,002; 0; 0,1
тивности эгоистичного пос%
тупка (ЭЭП), среднее значе%
ние эффекта, StDev
Основной объем мозга
20
минимально возможный объем мозга; объем мозга индивида ра%
вен 20, когда значения генов ОП и ЭОб являются равными нулю
Фактический объем мозга
20 + ОП + 30 × ЭОб
мозг становится больше с увеличением способностей к социально%
му обучению
Мемы ЭЭП: среднее значение
1; 1,5; 1; 2
когда изобретен мем ЭЭП, его эффективность устанавливается рав%
эффективности, StDev, C, R
ной нормально распределенному случайному значению (в случае
отрицательного результата попытка повторяется); размер мема - это
абсолютное значение эффективности мема, умноженное на C, плюс
случайное число с нулевым средним и стандартным отклонением R
Мемы ЭО: среднее значение
4; 6; 0,25; 2
то же, что и предыдущий; эффективность не может быть отрица%
эффективности, StDev, C, R
тельной
Сст%мемы: среднее значение
0; 0,03; 5; 2
эффективность мемов Сст может быть как положительной (мемы,
эффективности, StDev, C, R
препятствующие выживанию), так и отрицательной (мемы, способ%
ствующие выживанию)
Вероятность забыть мем
0,01
индивид забывает каждый мем с вероятностью 0,01 в год
Стоимость охоты
2
за участие в охоте у индивида забирается 2 ресурса
Шанс миграции
0,001
вероятность перехода в другую группу (на индивида в год)
Исходное состояние: числен%
10, 20
моделирование начинается с 10 индивидов, каждый из которых об%
ность популяции, ресурсы на
ладает 20 ресурсами
индивида
будут развиваться ЭО и ЭЭП при фиксирован%
люционной гонке вооружений друг с другом.
ной высокой (Сст
=
0,25) или низкой
Группы всегда конкурируют с другими группа%
(Сст = 0,03) скорости старения. В TribeSim ин%
ми, тем самым способствуя эволюции ЭО путем
дивиды и группы участвуют в бесконечной эво%
группового отбора. Внутри групп индивиды
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1745
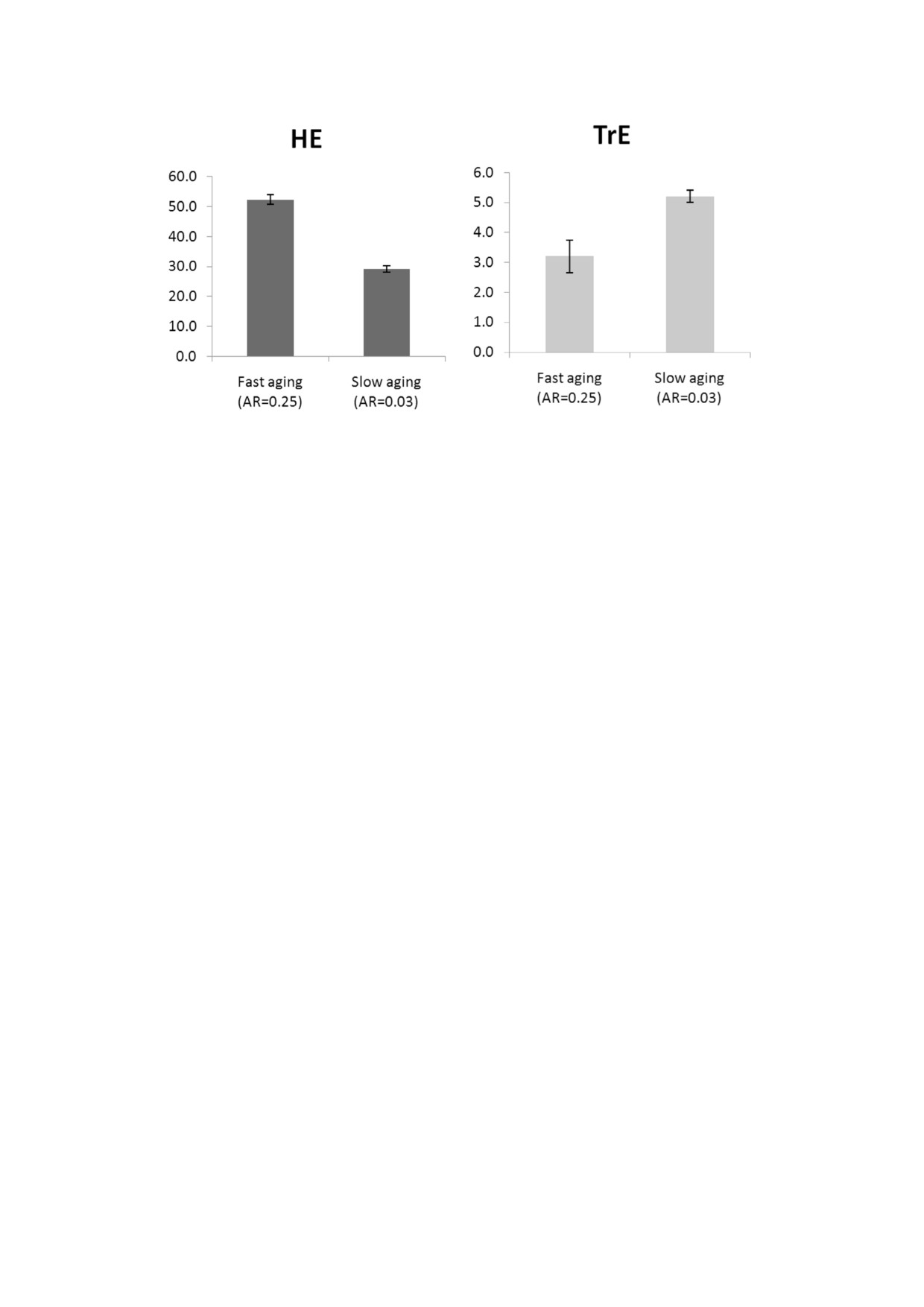
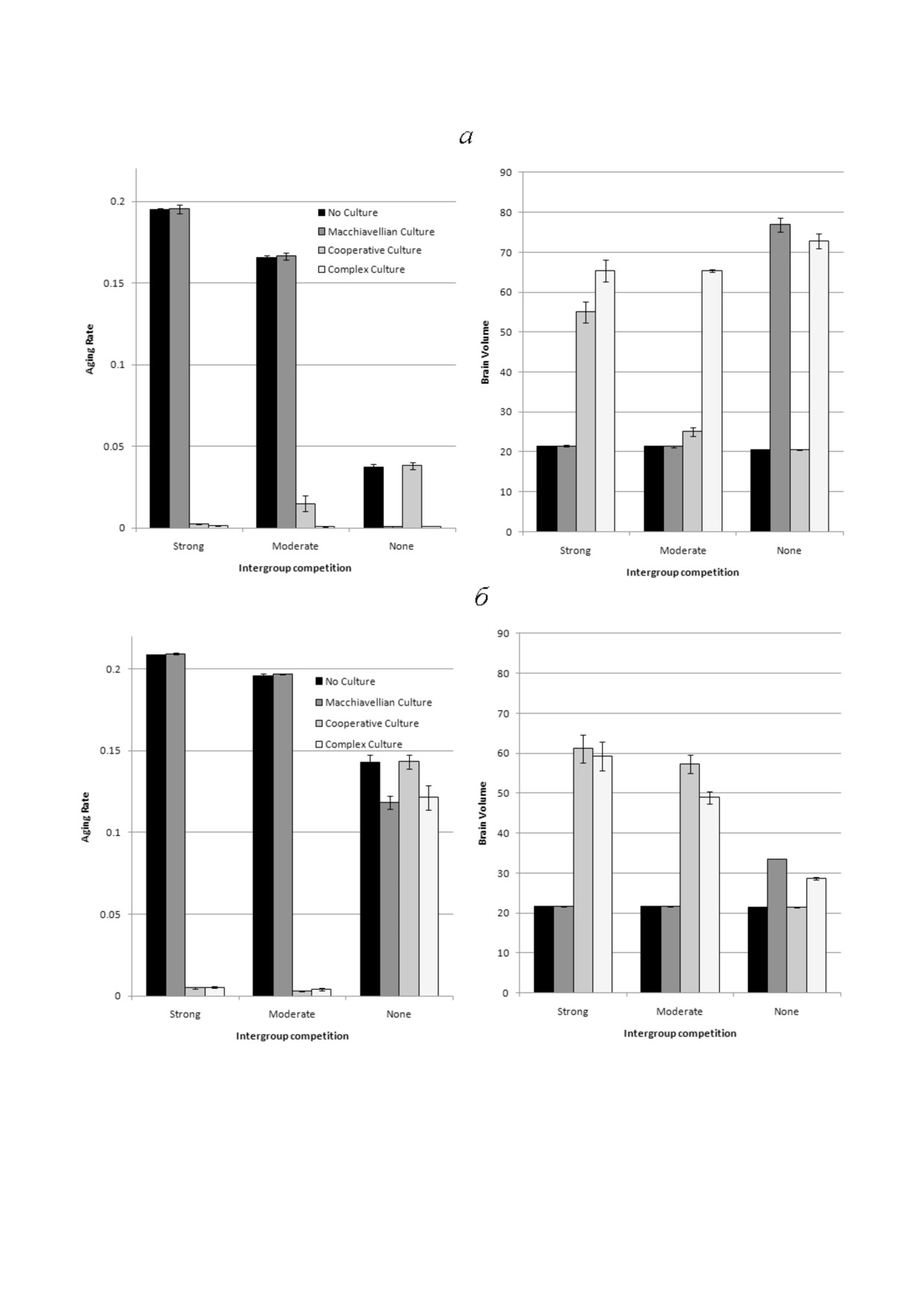
Рис. 1. Генетическая эволюция ЭО (HE) и ЭЭП (TrE) при быстром и медленном старении (AR - скорость старения, Сст)
(малое и большое время смены поколений). Приведены средние значения признаков за годы 50 000-80 000 из трех про%
гонов модели (± стандартное отклонение). Диаграммы показывают, что ЭО развивается быстрее при быстром старении,
а ЭЭП развивается быстрее при медленном старении. Параметры: генетическая эволюция - ЭО, ЭЭП; культура - нет;
G = 40
всегда конкурируют со своими товарищами по
жителей. При Сст = 0,03 средняя ПЖ составля%
группе, облегчая тем самым эволюцию ЭЭП
ет 38-40 лет при текущих параметрах; когда
посредством индивидуального отбора. Эта гон%
Сст = 0,25, средняя ПЖ составляет 12-13 лет. В
ка никогда не прекращается, и поэтому высокая
первом случае конкуренция за ресурсы более
эволюционируемость явно выгодна эволюцио%
острая, и процент смертей от голода выше
нирующим линиям, по крайней мере, в долго%
(17-18% всех смертей); при Сст = 0,25 смертей
срочной перспективе.
от голода практически не происходит. Более то%
Будут ли ЭО и ЭЭП развиваться быстрее при
го, в популяции долгожителей с наследствен%
быстром старении? В этом имитационном экс%
ными вариациями ЭЭП наблюдается сильное
перименте мы использовали промежуточный
ресурсное неравенство, что лишает особей с бо%
уровень МГК (G = 40), который гарантирует,
лее низкой ЭЭП возможности размножения.
что как групповой, так и индивидуальный отбор
Следовательно, в популяции долгожителей
достаточно эффективны. Результаты показаны
ЭЭП находится под более сильным отбором.
на рис. 1.
Оказывается, это более мощная эволюционная
Моделирование показало, что полезный для
сила, чем эффект более быстрой смены поколе%
группы признак, ЭО, развивается быстрее при
ний. Такая ситуация приводит к более быстрой
быстром старении, чем при медленном старе%
эволюции ЭЭП в популяции медленно старею%
нии, как и ожидалось. При этом индивидуально
щих индивидов.
полезный признак, ЭЭП, развивается быстрее,
Таким образом, мы видим, что ускорение
когда старение идет медленно. По%видимому, в
старения может ускорить эволюцию некоторых,
этом случае замедляющий эволюцию эффект
но не всех адаптивных признаков. Если положи%
более медленной смены поколений перевеши%
тельное влияние признака на приспособлен%
вается каким%то другим фактором. Природа
ность увеличивается с ростом ПЖ, то этот приз%
этого фактора становится ясной, если мы рас%
нак может даже быстрее эволюционировать в
смотрим, каким образом ЭЭП влияет на дарви%
популяции медленно стареющих индивидов.
новскую приспособленность индивидов. Инди%
Затем мы задались вопросом, может ли ста%
виды с более высокой ЭЭП получают бóльшую
рение эволюционировать быстрее (или медлен%
долю ресурсов, приобретаемых группой, чем их
нее) вследствие того, что оно ускоряет развитие
товарищи по группе с более низкой ЭЭП. Это
других адаптивных признаков. С этой целью мы
позволяет иметь больше потомства (сокращает
смоделировали популяцию, в которой два приз%
интервалы между родами) и снижает риск голо%
нака (Сст и ЭО или Сст и ЭЭП) эволюциониру%
да. Первое преимущество одинаково важно и
ют генетически, и сравнили ее с популяцией, в
для долгожителей, и для короткоживущих осо%
которой только Сст эволюционирует генетичес%
бей, но второе более важно в сообществе долго%
ки; культура отсутствует. Если более быстрое
БИОХИМИЯ том 86 вып. 12 2021
1746
МАРКОВ А., МАРКОВ М.
(или более медленное) старение может эволю%
ется за счет более быстрого старения. Напротив,
ционировать специально для ускорения эволю%
ЭЭП (индивидуально полезный признак) эво%
ции других признаков, то мы ожидали, что сов%
люционирует лучше в отсутствие МГК, когда
местная эволюция Сст и ЭО приведет к уско%
индивидуальный отбор является самым силь%
ренному старению, в то время как совместная
ным (G = 2000); медленное старение также спо%
эволюция Сст и ЭЭП приведет к более медлен%
собствует развитию ЭЭП.
ному старению по сравнению с популяцией, в
Влияние эгоистичных и кооперативных куль?
которой эволюционирует только Сст.
тур на генетическую эволюцию старения. Куль%
В этом эксперименте, а также в последую%
турные признаки отличаются от генетически
щих мы использовали три различных уровня
детерминированных несколькими важными
МГК: G = 15 (сильная МГК и групповой отбор,
особенностями [55], две из которых имеют осо%
слабый индивидуальный отбор), G = 40 (уме%
бое значение для текущего исследования.
ренная МГК, групповой и индивидуальный от%
Во%первых, культурные признаки наследуются
бор сопоставимой силы), G = 2000 (отсутствие
не только по вертикали (от родителей к детям),
МГК, отсутствие группового отбора, сильный
но и по горизонтали (от одного индивида к дру%
индивидуальный отбор). Результаты моделиро%
гому). Это означает, что размножение мемов не
вания показаны на рис. 2.
является неразрывно связанным с размножени%
Диаграммы показывают, что совместная эво%
ем генов. В результате мемы могут эволюцио%
люция с другими признаками не оказывает су%
нировать «эгоистично»: культурная эволюция
щественного влияния на эволюцию Сст. Незна%
может порождать признаки, которые снижают
чительные различия, видимые на диаграмме, не
дарвиновскую приспособленность индивидов
соответствуют вышеприведенным ожиданиям и
(т.е. ухудшают распространение их генов).
могут быть объяснены другими факторами (не%
Во%вторых, необходимо время, чтобы приоб%
большое количество прогонов модели и немно%
рести культурные признаки посредством соци%
го более низкая эффективность отбора по приз%
ального обучения. Индивиды рождаются с ну%
наку, когда отбирается более одного признака).
левыми знаниями и могут учиться на протяже%
Таким образом, моделирование не подтвердило
нии всей своей жизни. Чем дольше они живут,
идею о том, что скорость старения может изме%
тем больше знаний они могут получить и тем
няться специально для того, чтобы ускорить
сильнее будет фенотипическое выражение
эволюцию некоторых других адаптивных приз%
культурно обусловленных признаков. Напри%
наков.
мер, охотничье мастерство (ЭО) можно оттачи%
Диаграммы также показывают, что более вы%
вать в течение всей жизни, что делает пожилых
сокая МГК способствует более быстрому старе%
индивидов в среднем более эффективными
нию, а более низкая МГК способствует более
охотниками, чем молодые.
медленному старению. Это связано с тем, что
Нашей следующей задачей было понять, как
бóльшая ПЖ очень полезна для отдельных ин%
культура (эгоистичная, кооперативная и комп%
дивидов, но не для групп (конкурентоспособ%
лексная) влияет на эволюцию Сст. Для простоты
ность группы несильно зависит от того, на%
мы начали с моделирования не естественно эво%
сколько быстро сменяются поколения внутри
люционирующей культуры (которая, согласно
группы). Когда МГК сильна, как индивидуаль%
теории культурного драйва, эволюционирует
ный, так и групповой отбор работают против
вместе со способностью к социальному обуче%
чрезмерного потребления ресурсов, необходи%
нию), а стационарной, эволюционирующей
мых для повышения выживаемости, но когда
только в заданных изначально пределах. Для
МГК отсутствует, только индивидуальный отбор
этого мы установили фиксированные значения
выполняет свою работу. С точки зрения «блага
ОП = 5 (память, которая может вместить нес%
отдельного индивида» выгоды от долгой жизни
колько больших мемов или несколько десятков
перевешивают ущерб, причиненный затратами
маленьких) и ЭОб = 0,6 (при таком уровне эф%
на ее продление, в то время как с точки зрения
фективности обучения требуется около
«блага группы» эти затраты не компенсируются.
10-20 лет, чтобы заполнить память мемами).
Следовательно, при преобладании группового
Обе характеристики в этом эксперименте были
отбора эволюционирует более быстрое старе%
«бесплатными», т.е. не влияли на объем мозга,
ние, а при преобладании индивидуального от%
который всегда был равен 20. С этими парамет%
бора - более медленное старение.
рами пул мемов популяции быстро (в течение
Мы можем видеть, что ЭО (групповой полез%
первых сотен лет) заполняется достаточным ко%
ный признак) эволюционирует лучше, когда
личеством мемов, чтобы память индивидов не
МГК сильна (G = 15), как и ожидалось. В этом
оставалась пустой. Результаты моделирования
конкретном случае развитие ЭО также ускоря%
показаны на рис. 3, а.
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1747
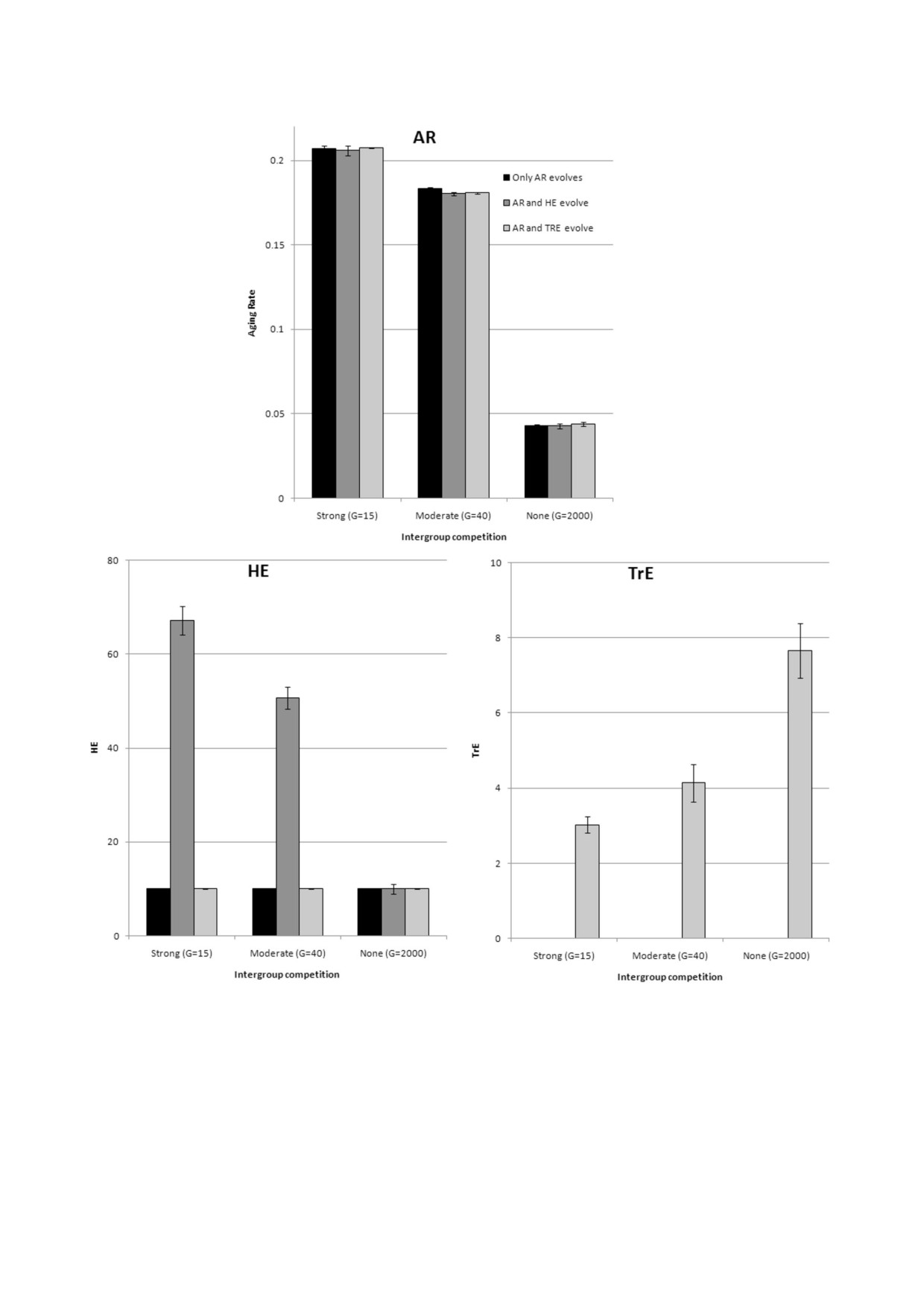
Рис. 2. Генетическая эволюция Сст (AR), ЭО (HE) и ЭЭП (TrE) в трех ситуациях: эволюционирует только Сст, эволюци%
онирует Сст и ЭО, эволюционирует Сст и ЭЭП; культура отсутствует. Показаны средние значения из трех прогонов мо%
дели за годы 50 000-80 000 (± стандартное отклонение)
На рис. 3 видно, что культура имеет тенден%
Эгоистичная культура больше способствует это%
цию способствовать замедлению старения (в
му, когда МГК минимальна (т.е. когда условия
каждой группе из четырех столбцов столбцы
для развития такой культуры наиболее благо%
2-4 в большинстве случаев ниже столбца 1).
приятны). И наоборот, кооперативная культура
БИОХИМИЯ том 86 вып. 12 2021
1748
МАРКОВ А., МАРКОВ М.
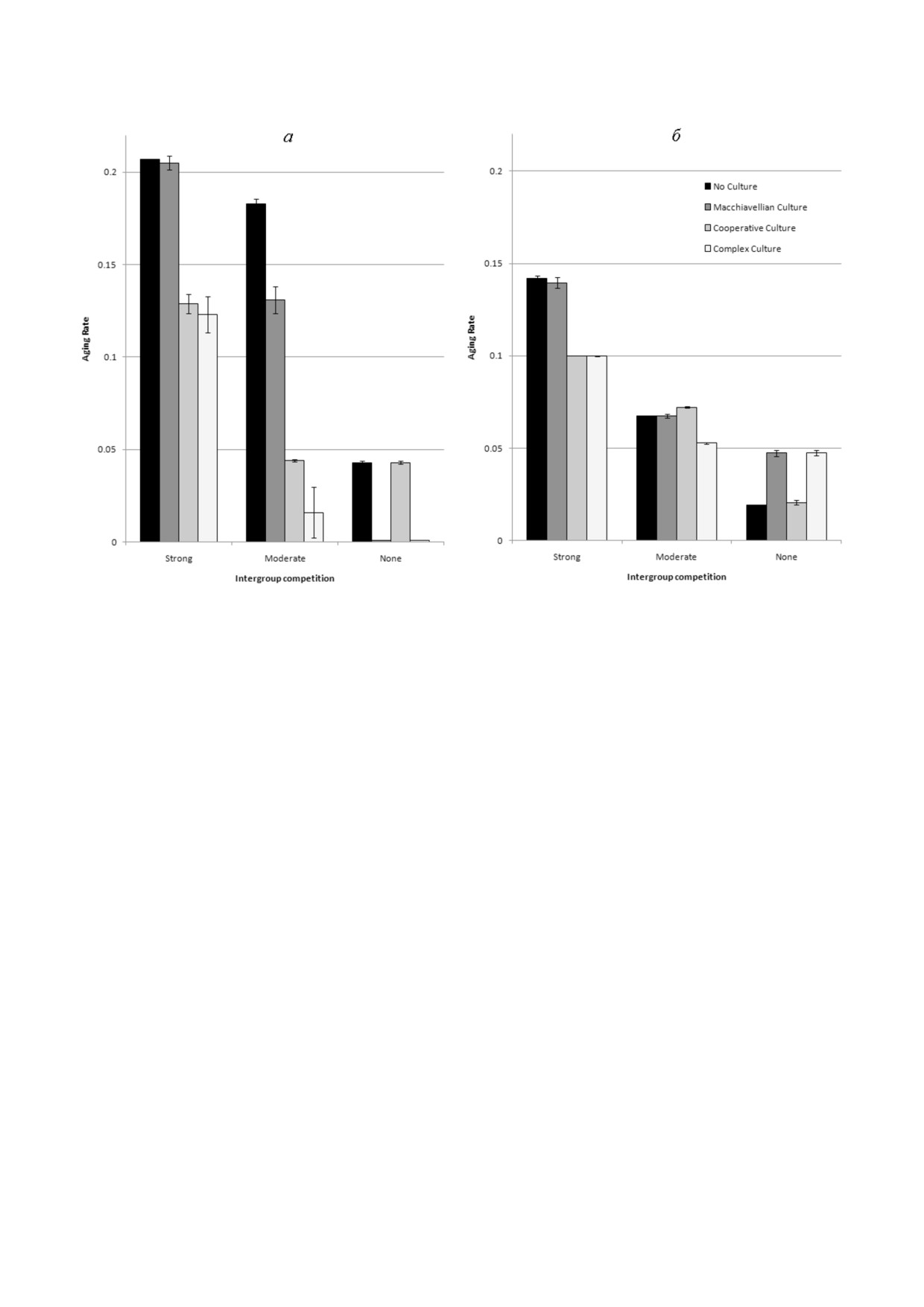
Рис. 3. Генетическая и культурная эволюция Сст. Мозг не эволюционирует (ОП = 5, ЭОб = 0,6, объем мозга = 20). Пока%
заны средние значения из трех прогонов модели за годы 50 000-80 000 (± стандартное отклонение). а - Генетическая эво%
люция Сст в четырех ситуациях: культура отсутствует; культура содержит только мемы ЭЭП (эгоистичная культура); куль%
тура содержит только мемы ЭО (кооперативная культура); культура содержит оба типа мемов (комплексная культура).
б - Культурная эволюция Сст в четырех ситуациях: разрешены только мемы Сст (культура отсутствует), мемы Сст и ЭЭП
(эгоистичная культура), мемы Сст и ЭО (кооперативная культура), мемы Сст, ЭЭП и ЭО (комплексная культура). Геноти%
пический (врожденный) уровень Сст - 0,1
способствует более медленному старению наи%
ожидать, никак не влияет на эволюцию старе%
более эффективно, когда МГК сильна (что оз%
ния.
начает, что условия благоприятны для развития
Почему культура, как правило, способствует
кооперативной культуры). Комплексная культу%
эволюции замедленного старения (рис. 3, а),
ра суммирует эффекты обеих специализирован%
тогда как генетическая эволюция тех же фено%
ных культур, усиливая отбор на замедление ста%
типических признаков (ЭО и ЭЭП) не влияет на
рения при любом уровне МГК.
эволюцию Сст (рис. 2)? Причина в том, что ге%
Два исключения легко объяснить. В первом
нетически детерминированные признаки явля%
случае (сильная МГК, эгоистичная культура,
ются врожденными, а для овладения культурно
столбец 2 в группе 1) культура развивается пло%
детерминированными признаками требуется
хо, потому что при сильной МГК эффектив%
время. При наличии кооперативной культуры
ность индивидуального отбора, поддерживаю%
(мемы ЭО) индивиды постепенно становятся
щего «эгоистичные» признаки, мала. Соответ%
все более и более умелыми охотниками на про%
ственно, влияние культуры на эволюцию Сст
тяжении всей своей жизни. В то же время они
незначительно. Во втором случае (без МГК,
становятся лучшими хранителями и распрост%
кооперативная культура, столбец 3 в группе 3)
ранителями полезных знаний (охотничьих на%
нет группового отбора для поддержки полез%
выков). Следовательно, чем дольше живут осо%
ных для группы признаков, таких как ЭО. Со%
би, тем лучше для группы: в ней будет больше
ответственно, культура развивается плохо, и,
хороших охотников и больше полезных знаний.
что наиболее важно, она не адаптивна в эволю%
Следовательно, групповой отбор способствует
ционном смысле, потому что ЭО не поддержи%
более медленному старению.
вается отбором в отсутствие МГК. Такая «адап%
В случае «макиавеллистской» (далее - эго%
тивно нейтральная» культура, как и следовало истичной) культуры (мемов ЭЭП) индивиды в
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1749
течение жизни становятся все более и более
Именно это и происходит, например, в ситуа%
изощренными манипуляторами, эгоистами и
ции «эгоистичная культура, G = 40» (столбец 2 в
интриганами. Это позволяет им получать боль%
группе 2, рис. 3, а): групповой отбор не позволя%
ше ресурсов и, следовательно, производить
ет слишком сильно замедлить старение.
больше потомства и с меньшей вероятностью
Культурная эволюция скорости старения. По%
умереть от голода. Следовательно, при наличии
смотрев, как культура влияет на генетическую
эгоистичной культуры (но не врожденных «эго%
эволюцию Сст, мы повторили предыдущий экс%
истичных инстинктов») дарвиновская приспо%
перимент, на этот раз позволив признаку разви%
собленность индивидов растет с большим уско%
ваться культурно, а не генетически. Для этого
рением по мере того, как увеличивается их ПЖ.
мы запретили мутации гена Сст (фиксирован%
Следовательно, индивидуальный отбор способ%
ное значение генотипа Сст = 0,1) и ввели новую
ствует увеличению ПЖ.
категорию мемов: мемы Сст. Эти мемы могут
Интересно, что в отсутствие МГК чрезмер%
как увеличивать, так и уменьшать Сст. Первое
ное замедление старения в сочетании с эгоис%
соответствует рискованному поведению, кото%
тичной культурой (столбец
2 в группе
3,
рое позволяет индивиду экономить ресурсы
рис. 3, a) приводит к вымиранию популяции
или, что то же самое, получать дополнительные
при параметрах, используемых по умолчанию
ресурсы в обмен на повышенный риск смерти
(уровень врожденной ЭО равен 10). Чтобы за%
(«мемы соломенных хижин»). Второе соответ%
вершить этот эксперимент, нам пришлось удво%
ствует трате дополнительных ресурсов для ми%
ить уровень врожденной ЭО (ЭО = 20), что ана%
нимизации риска смерти («мемы кирпичных
логично искусственному кормлению популя%
домов»).
ции, неспособной прокормить себя. Механизм
Мы смоделировали те же три уровня МГК
вымирания следующий: увеличение ПЖ усили%
(G = 15, 40, 2000) и четыре варианта культуры:
вает конкуренцию за ресурсы, а эгоистичная
культура отсутствует (только Сст%мемы), эгоис%
культура делает распределение этих ресурсов
тичная культура (мемы Сст и ЭЭП), коопера%
чрезвычайно неравномерным (сильное неравен%
тивная культура (мемы Сст и ЭО), комплексная
ство ресурсов). Молодые особи почти ничего не
культура (мемы Сст, ЭЭП и ЭО). Остальные па%
получают, пока не выучат достаточно мемов
раметры были такими же, как в предыдущем
ЭЭП. Те немногие, кому посчастливилось быст%
эксперименте.
ро изучить эффективные мемы ЭЭП, живут дол%
Мы уже знаем, к каким стабильным уровням
го и успешно размножаются, но большинство
стремится генетическая эволюция Сст с этими
молодых индивидов умирают из%за нехватки ре%
параметрами (рис. 3, a). Другими словами, мы
сурсов. Возникает общество «алчных и хитрых
знаем эволюционные оптимумы (или эволюци%
долгожителей и голодающей молодежи». Острая
онно стабильные состояния) Сст с точки зрения
нехватка ресурсов приводит к снижению доли
генов. Будет ли культурная эволюция Сст при%
особей, участвующих в коллективной охоте.
водить к тем же стабильным состояниям? Ре%
Охота - дорогостоящее мероприятие, на кото%
зультаты моделирования показаны на рис. 3, б.
рое индивид тратит два ресурса; если у него нет
Мы видим, что МГК влияет на культурную и
этих ресурсов, он не ходит на охоту. Уменьше%
генетическую эволюцию Сст аналогичным об%
ние количества охотников можно было бы ком%
разом: сильная МГК способствует культурной
пенсировать увеличением охотничьего мастер%
эволюции высокой Сст (это позволяет группе
ства, но в данном случае это невозможно, так
экономить ресурсы), а слабая МГК способству%
как врожденный уровень ЭО не развивается, и
ет культурной эволюции низкой Сст. Однако то,
нет никаких мемов ЭО. В результате срабатыва%
как эгоистичные и кооперативные культуры
ет деструктивная положительная обратная связь
влияют на культурную эволюцию Сст, сильно
(увеличивается голод → меньше охотников →
отличается от того, что мы наблюдали в случае
голод увеличивается еще больше), что заканчи%
генетической эволюции Сст. У этих различий
вается вымиранием популяции. Катастрофа на%
есть несколько причин.
чинается в тот момент, когда общее количество
Первая причина заключается в том, что в
охотничьих усилий популяции становится не%
культурной эволюции важную роль играет кон%
достаточным для добычи всех 6000 ресурсов,
куренция между мемами за доминирование в па%
ежегодно предоставляемых окружающей сре%
мяти индивидов и в групповом пуле мемов. Кос%
дой. Конечно, это возможно только при отсут%
венные аналоги этому можно найти в генетичес%
ствии МГК. Если групповой отбор работает, он
кой эволюции в природе (например, компро%
предотвратит катастрофу, благоразумно отбра%
мисс между различными адаптивными призна%
ковав группы, в которых замедление старения и
ками), но в TribeSim это не так. Например, алле%
неравенство ресурсов зашли слишком далеко.
ли гена ЭЭП конкурируют друг с другом, но не с
БИОХИМИЯ том 86 вып. 12 2021
1750
МАРКОВ А., МАРКОВ М.
аллелями генов ЭО или Сст, и эволюция врож%
же, чем при генетической эволюции, потому что
денных эгоистичных способностей никоим об%
«мемы соломенных хижин» сокращают жизнь
разом не ограничивает эволюцию врожденных
своих хозяев, тем самым препятствуя своему
охотничьих способностей. Мемы разных кате%
собственному распространению. Поэтому им
горий, напротив, конкурируют за одни и те же
сложно конкурировать с «мемами кирпичных
ресурсы: носителей (индивидов), объем их па%
домов», которые, наоборот, сами помогают себе
мяти и время, которое они тратят на обучение.
распространяться (являются вирусными).
Вторая причина заключается в том, что оп%
№ 2: Сильная МГК; разрешены мемы Сст и
ределенные категории мемов могут стать вирус%
ЭЭП. Сильная МГК приводит к слабому разви%
ными, т.е. они могут ускорить свое собственное
тию «макиавеллистской» (эгоистичной) культу%
распространение, превратив своих носителей в
ры. В результате мемы Сст с положительным
более эффективные «машины для хранения и
эффектом («мемы соломенных хижин»), под%
распространения мемов». Сст%мемы с негатив%
держиваемые мощным групповым отбором, вы%
ным эффектом («мемы кирпичных домов») яв%
тесняют мемы ЭЭП из пула мемов. Следова%
ляются типичным примером вирусных мемов,
тельно, эгоистичная культура не влияет на ко%
поскольку они продлевают жизнь индивидов.
нечный уровень Сст, который оказывается та%
Индивиды%долгожители накапливают в своей
ким же, как и в предыдущем случае (около 0,14).
памяти больше мемов, дольше хранят их и пере%
№№ 3, 4: Сильная МГК; разрешены мемы
дают большему количеству подражателей (уче%
Сст и ЭО или Сст, ЭЭП и ЭО. Сильный культур%
ников). Следовательно, при прочих равных,
ный групповой отбор поддерживает группы с
«мемы кирпичных домов» распространяются
эффективной кооперативной (охотничьей)
быстрее и побеждают в конкуренции «мемы со%
культурой. В результате эта культура сильно раз%
ломенных хижин». Из%за этого культурная эво%
вивается, и мемы ЭО почти полностью вытесня%
люция Сст часто приводит к более низким фе%
ют мемы Сст и ЭЭП из группового пула мемов.
нотипическим значениям Сст, чем это опти%
Культура Сст не может развиваться вообще, а
мально с точки зрения генов (ниже эволюцион%
фенотипический уровень Сст остается на исход%
но стабильного состояния, характерного для ге%
ном, генетически детерминированном уровне
нетической эволюции Сст).
(0,1). Этому также способствует слабость груп%
Третья причина заключается в том, что боль%
пового отбора в пользу «мемов соломенных хи%
шая ПЖ способствует культурному развитию, а
жин», поскольку эволюционный оптимум в ге%
короткая - тормозит [13]. Следовательно, при
нетической эволюции Сст с этими параметрами
прочих равных условиях в обществе долгожите%
(кооперативная культура и G = 15) составляет
лей любые культурно обусловленные фенотипи%
примерно 0,12-0,13 (рис. 3, a, столбцы 2 и 3),
ческие признаки развиваются сильнее, чем в об%
что лишь немного превышает врожденный ге%
ществе короткоживущих особей.
нотипический уровень Сст в текущем экспери%
Давайте рассмотрим более подробно 12 ситу%
менте (0,1). Поэтому групповой отбор на сниже%
аций, показанных на рис. 3, б (столбцы 1-12),
ние ПЖ в этой ситуации не очень силен. Более
чтобы понять, как работают эти факторы. Сле%
важны охотничьи навыки, для приобретения
дует помнить, что уровень генотипической
которых требуется время (т.е. более продолжи%
(врожденной) Сст фиксирован и равен 0,1, а
тельная жизнь).
значение фенотипической Сст можно изменить
№ 5: Умеренная МГК; разрешены только ме%
только путем изучения мемов Сст.
мы Сст. В этом случае устанавливается уровень
№ 1: Сильная МГК; разрешены только мемы
Сст ≈ 0,07, что ниже исходного генотипического
Сст. Как мы знаем, в этой ситуации генетичес%
уровня (0,1) и намного ниже уровня, к которому
кая эволюция Сст стремится к Сст ≈ 0,21
стремится генетическая эволюция Сст в тех же
(рис. 3, а, столбец 1). Сильная МГК делает эко%
условиях (0,18, рис. 3, а, столбец 5). Первое оз%
номию ресурсов очень актуальной для групп.
начает, что группы развили культуру «кирпич%
Групповой отбор поддерживает группы с низкой
ных домов». Второе означает, что эта культура
ПЖ, потому что короткоживущие особи тратят
развивалась, несмотря на тот факт, что для от%
меньше ресурсов за год. В ходе культурной эво%
дельных особей и групп было бы более выгодно
люции (рис. 3, б, столбец 1) устанавливается бо%
(с точки зрения дарвиновской приспособлен%
лее низкий уровень Сст (Сст ≈ 0,14), но он все
ности) иметь более низкую ПЖ, т.е. иметь
еще выше исходного генотипического уровня,
«культуру соломенных хижин». Очевидно, это
равного 0,1. Это означает, что в конкурирующих
результат «эгоизма» или вирусных свойств «ме%
группах под влиянием культурного группового
мов каменных домов» (см. выше).
отбора [42] формируется «культура соломенных
№№ 6-8: Умеренная МГК; разрешены ме%
хижин». Конечный уровень Сст получается ни%
мы Сст и ЭЭП, Сст и ЭО или Сст, ЭЭП и ЭО. В
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1751
случае генетической эволюции Сст с этими па%
числа смертей за год (при отсутствии эгоистич%
раметрами любая культура (эгоистичная, коопе%
ной культуры в ситуациях 9 и 11 она составляет
ративная или комплексная) приводит к сниже%
менее 1%). В таких обстоятельствах лучшая за%
нию Сст (на рис. 3, а столбцы 6-8 ниже столб%
щита от преждевременной смерти обеспечива%
ца 5). При культурной эволюции этого не наб%
ется не «мемами кирпичных домов» (которые
людается: конечный уровень Сст остается при%
фактически увеличивают потребность индивида
мерно таким же, как и в предыдущей ситуации
в ресурсах), а мемами ЭЭП, которые предостав%
(0,05-0,07; рис. 3, б, столбец 5). Причина кроет%
ляют индивиду дополнительные ресурсы. Эго%
ся в балансе двух эволюционных сил. С одной
истичные мемы, продлевая жизнь индивидов,
стороны, эгоистичная и/или охотничья культу%
приобретают вирусные свойства и распростра%
ра способствует индивидуальному и/или груп%
няются еще быстрее, вытесняя Сст%мемы из пу%
повому отбору на долголетие. С другой стороны,
ла мемов и тем самым препятствуя развитию
эгоистичные и охотничьи мемы, поддерживае%
культуры «кирпичных домов».
мые отбором, распространяются в пуле мемов и,
При отсутствии охотничьих мемов (ситуа%
таким образом, ограничивают возможности
ция 10) популяция вымирает так же, как и в
распространения «мемов кирпичных домов».
случае генетической эволюции Сст с теми же
№№ 9, 11: Отсутствие МГК; допускаются
параметрами (см. выше). Для завершения этого
мемы Сст или Сст и ЭО. В отсутствие МГК
модельного эксперимента нам пришлось повы%
культура охоты не поддерживается отбором, ме%
сить уровень врожденной ЭО (с 10 до 20). Если
мы ЭО не распространяются и не влияют на ко%
охотничьи мемы разрешены (ситуация 12), то
нечный уровень Сст, поэтому эти две ситуации
популяция выживает. Хотя охотничья культура
похожи. Конечный уровень Сст очень низкий
в отсутствие МГК не поддерживается отбором,
(0,019-0,020). Это значительно ниже, чем в слу%
некоторые мемы ЭО все же распространяются
чае генетической эволюции Сст в тех же услови%
из%за того, что можно назвать «культурным
ях (0,043). Причина опять же в «эгоизме»
дрейфом», феноменом, который особенно
Сст%мемов с негативным эффектом (мемы
сильно выражен в отсутствие МГК [13]. В ре%
«кирпичных домов»). Продлевая жизнь индиви%
зультате средний уровень ЭО превышает 10, и
дов, эти мемы помогают себе распространяться.
популяция не достигает фатального порога,
В результате индивиды живут дольше, чем было
когда общее охотничье усилие популяции ста%
бы оптимально с точки зрения их дарвиновской
новится недостаточным для извлечения всех
приспособленности.
6000 ресурсов, ежегодно обеспечиваемых окру%
№№ 10, 12: Отсутствие МГК; разрешены ме%
жающей средой.
мы Сст и ЭЭП или Сст, ЭЭП и ЭО. В отсутствие
Генно?культурная эволюция Сст. До сих пор
МГК мемы ЭО не поддерживаются отбором и не
мы сравнивали генетическую и культурную эво%
влияют на окончательный уровень Сст, как уже
люцию Сст, рассматривая их по отдельности.
упоминалось, поэтому эти две ситуации похо%
Однако в реальном антропогенезе эти процес%
жи. Конечный уровень Сст составляет
сы, скорее всего, протекали параллельно. Есть
0,047-0,048, что значительно выше, чем в слу%
основания полагать, что существовала как гене%
чае генетической эволюции Сст в этих условиях
тическая эволюция Сст (например, максималь%
(0,001). Это также выше, чем в случае культур%
ная ПЖ Homo sapiens более чем вдвое выше, чем
ной эволюции Сст в отсутствие эгоистичной
максимальная ПЖ других человекообразных
культуры (0,019-0,020, см. выше). Причина в
обезьян [56]), так и культурная эволюция. Оче%
чрезвычайно сильном развитии эгоистичной
видно, что человечество накопило огромное ко%
культуры с этими параметрами. Мемы ЭЭП за%
личество знаний, обычаев, навыков, социаль%
полняют значительную часть пула мемов (объем
ных практик и норм, влияющих на выживание и
которого ограничен фиксированными значени%
жизнеспособность [4-7].
ями ОП и ЭОб) и тем самым препятствуют раз%
В TribeSim, если Сст разрешено эволюцио%
витию культуры «кирпичных домов». Сильное
нировать обоими способами одновременно,
развитие эгоистичной культуры при этих пара%
разворачивается сложный процесс генно%куль%
метрах объясняется тем, что долгая жизнь инди%
турной эволюции признака. Генетические изме%
видов приводит к нехватке ресурсов, а сама эго%
нения в Сст влияют на развитие культурных эле%
истичная культура создает сильное ресурсное
ментов, влияющих на Сст, и наоборот. Рассмот%
неравенство («общество жадных, хитрых долго%
рим ход генно%культурной эволюции Сст в двух
жителей и голодающей легкомысленной моло%
ситуациях, различающихся направлением «эво%
дежи»). Сочетание этих двух факторов приводит
люционных интересов» (положением эволюци%
к резкому увеличению доли смертей от голода.
онных стабильных состояний или оптимумов)
Эта доля увеличивается до 62-65% от общего
генов Сст и мемов Сст.
БИОХИМИЯ том 86 вып. 12 2021
1752
МАРКОВ А., МАРКОВ М.
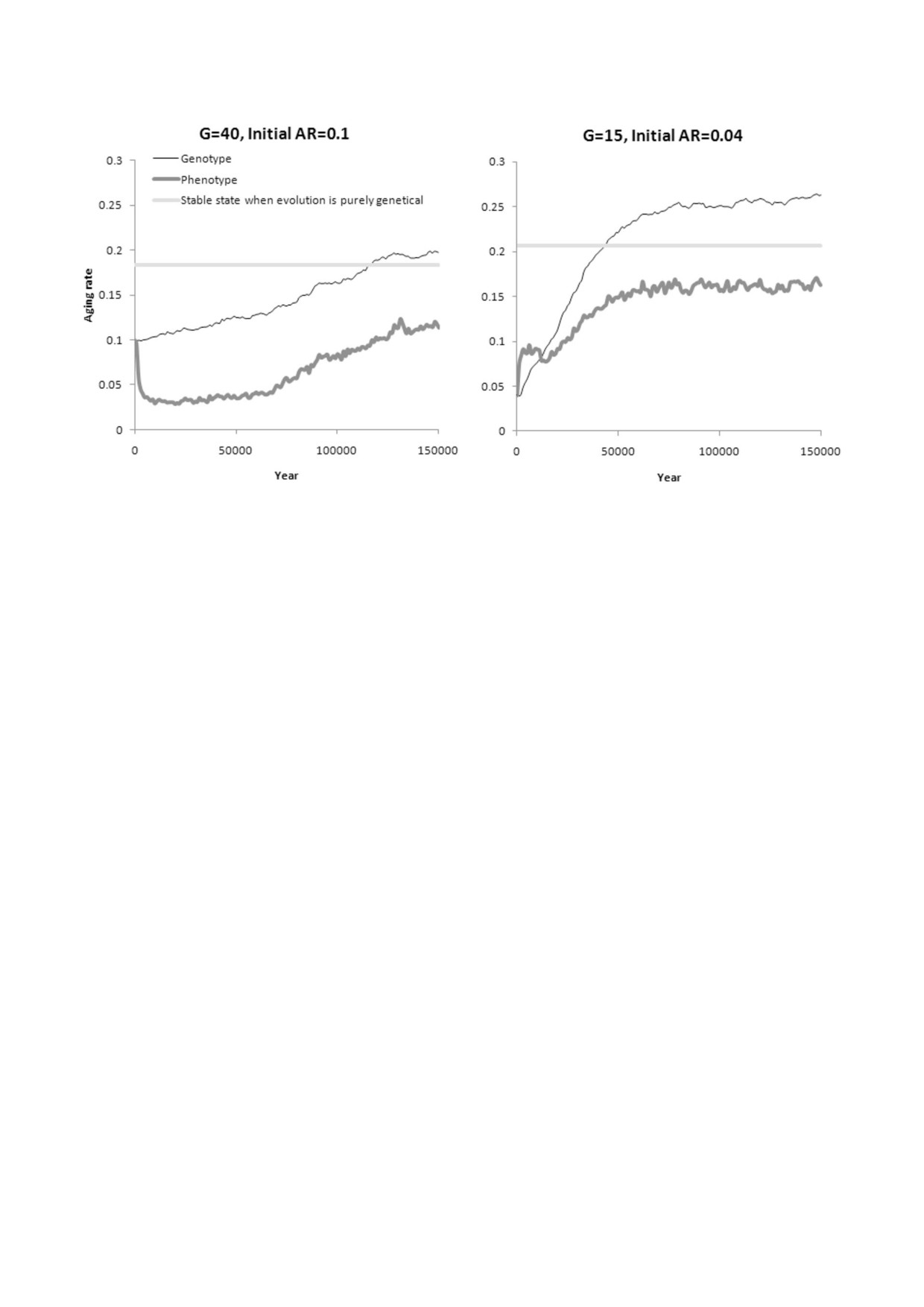
Рис. 4. Генно%культурная эволюция Сст (AR). Тонкая линия - генотипическое значение Сст, жирная линия - фенотипи%
ческое значение Сст. Прямая горизонтальная линия отмечает стабильное состояние признака, к которому генетическая
эволюция Сст стремится в тех же условиях без культуры. Разница между фенотипом и генотипом отражает влияние куль%
туры. Эта разница положительная (фенотипическое значение Сст выше генотипического), когда преобладает «культура
соломенных хижин», и отрицательная, когда преобладает «культура кирпичных домов». Показаны средние значения из
трех прогонов модели
1)
«Интересы» генов и мемов изначально
состояние, отчасти потому, что культура увели%
разнонаправлены. Параметры: генетическая
чивает изменчивость фенотипического прояв%
эволюция Сст (остальные гены фиксированы);
ления признака, и, таким образом, отбор оказы%
культурная эволюция Сст (другие мемы запре%
вается менее эффективным. В конце концов, ге%
щены); исходный генотипический уровень
нотипическое значение признака поднимается
Сст равен 0,1; G = 40 (умеренная МГК).
выше уровня, к которому стремится чисто гене%
Из предыдущих экспериментов мы уже зна%
тическая эволюция Сст в тех же условиях. В
ем, что при таких параметрах генетическая эво%
этом случае воздействие культуры всегда на%
люция Сст, если ей не мешают другие факторы,
правлено на снижение фенотипического значе%
стремится к равновесному уровню около 0,18,
ния Сст, т.е. формируется и сохраняется культу%
в то время как культурная эволюция из%за спо%
ра «кирпичных домов».
собности «мемов кирпичных домов» облегчать
2) «Интересы» генов и мемов изначально од%
собственное распространение стремится к более
нонаправлены. Параметры: генетическая эво%
низкому уровню, около 0,07.
люция Сст (остальные гены фиксированы);
В случае генно%культурной эволюции мемы
культурная эволюция Сст (другие мемы запре%
начинают сдвигать признак вниз, в то время как
щены); исходный генотипический уровень Сст
гены пытаются сдвинуть его вверх и компенси%
равен 0,04; G = 15 (сильная МГК).
ровать влияние культуры (рис. 4, график слева).
В этом случае генетическая эволюция Сст в
Культура в этом случае играет роль неадаптив%
отсутствие культуры стремится к 0,21, а культур%
ной фенотипической пластичности, что способ%
ная эволюция в отсутствие генетической эволю%
ствует усиленной генетической эволюции приз%
ции стремится к 0,14. Оба уровня значительно
нака в направлении, противоположном направ%
выше, чем исходный генотипический уровень
лению пластических изменений [57]. Другими
Сст в текущем эксперименте (0,04). Следова%
словами, поскольку культура уменьшает значе%
тельно, на начальном этапе и мемы, и гены
ние признака, гены вынуждены увеличивать его
должны стремиться к увеличению Сст.
еще сильнее. Это явление известно как генети%
Действительно, в этой ситуации сначала
ческая компенсация [58]. Генно%культурный
быстро эволюционирует культура «соломенных
эволюционный процесс требует значительного
хижин» (рис. 4: в левой части второго графика
времени для того, чтобы прийти в стационарное
фенотипический уровень Сст выше, чем гено%
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1753
типический уровень). Генетический уровень Сст
индивидов действительно способствует увели%
также растет, хотя и более медленными темпа%
чению размера мозга и культурному разви%
ми. Достаточно скоро этот рост приводит к тому,
тию [13], но еще не моделировали эволюцию
что культура сначала становится нейтральной
ПЖ (или скорости старения Сст) в контексте
(мемы «соломенных хижин» и «кирпичных до%
культурного драйва.
мов» уравновешивают друг друга), а затем раз%
Чтобы посмотреть, как коэволюция мозга и
вивается полноценная культура «кирпичных до%
культуры взаимодействует с эволюцией Сст, мы
мов». Это происходит задолго до того, как гено%
использовали следующие параметры: генетичес%
типическое или фенотипическое значение Сст
кая эволюция Сст, ОП и ЭОб; начальные значе%
приближается к 0,21 или даже 0,14, что объясня%
ния: Сст = 0,1, ОП = 0, ЭОб = 0. ОП и ЭОб явля%
ется вирусными свойствами мемов «кирпичных
ются дорогостоящими признаками в том смыс%
домов» (эти мемы, как мы помним, помогают
ле, что их рост приводит к увеличению объема
себе распространяться, потому что продлевают
мозга (который равен 20 + ОП + 30 × ЭОб), а
жизнь их носителям). Эгоистичная эволюция
увеличение объема мозга повышает ресурсную
мемов сдвигает уровень Сст ниже генетического
стоимость размножения.
эволюционного оптимума.
Мы смоделировали процесс при трех раз%
В результате в обоих случаях, показанных на
личных уровнях МГК (G = 15, 40 и 2000) и четы%
рис. 4, формируется общество, в котором моло%
рех вариантах культуры: без культуры, эгоистич%
дые индивиды ведут рискованный образ жизни,
ной, кооперативной и комплексной. Сначала
тем самым получая дополнительные ресурсы, и
рассмотрим подробно ход процесса при G = 40 и
у них есть генетическая предрасположенность к
комплексной культуре (рис. 5), а затем более
этой рискованной жизни. Но с возрастом инди%
кратко остановимся на остальных 11 комбина%
виды приобретают знания и навыки, которые
циях параметров (рис. 6, а).
делают жизнь безопаснее, хотя и требуют бóль%
На рис. 5 показано, что при эволюциониру%
ших ресурсных затрат. Это похоже на то, что мы
ющей Сст возникает гораздо более мощный
наблюдаем во многих человеческих популяциях.
культурный драйв, чем при Сст, зафиксирован%
Генетическая эволюция Сст и культурный
ной на относительно высоком уровне
драйв. До сих пор мы исследовали влияние куль%
(Сст = 0,1): социальное обучение (ОП и ЭОб, от
туры на эволюцию ПЖ и старение с фиксиро%
которых зависит объем мозга) и культура, осо%
ванными (а значит, не эволюционирующими)
бенно эгоистичная, эволюционируют сильнее.
значениями объема памяти (ОП), эффектив%
Это связано, прежде всего, с увеличением ПЖ
ности обучения (ЭОб) и объема мозга. Однако в
(долгожители лучше хранят и распространяют
реальном антропогенезе специфические для че%
знания), а во%вторых, с усилением развития
ловека эволюционные изменения ПЖ и Сст,
эгоистичной культуры. Последнее на опреде%
скорее всего, происходили на фоне быстро эво%
ленном этапе формирует положительную об%
люционирующего мозга и развивающейся куль%
ратную связь с увеличением доли смертей от го%
туры.
лода. Мемы ЭЭП усугубляют неравенство ре%
Нашей следующей задачей было смоделиро%
сурсов, что приводит к увеличению доли смер%
вать эволюцию Сст в контексте культурного
тей от голода. Это, в свою очередь, приводит к
драйва (самоподдерживающейся коэволюции
появлению сильного «жизнеспасающего» эф%
мозга, культуры и социального обучения).
фекта у мемов ЭЭП и, как следствие, к еще
Программа TribeSim изначально была разрабо%
большему распространению мемов ЭЭП и еще
тана специально для изучения культурного
большему неравенству ресурсов. В то же время
драйва [13].
развитие эгоистичной культуры стимулирует
Было высказано предположение, что в авто%
отбор на улучшение способностей к социально%
каталитической совместной эволюции мозга и
му обучению, что приводит к увеличению раз%
культуры может присутствовать дополнитель%
мера мозга.
ная петля положительной обратной связи через
В данном случае культурный драйв сначала
увеличение ПЖ. Культурное развитие ведет к
запускается мемами ЭО, которые повышают
увеличению ПЖ (как за счет культурно переда%
конкурентоспособность групп. Объем памяти и
ваемых навыков, которые продлевают жизнь,
эффективность обучения изначально находятся
так и за счет отбора на долголетие, которому
на нулевом уровне, но гены ОП и ЭОб мутиру%
способствует культура). В свою очередь, долгая
ют, и поэтому в популяции появляются особи с
жизнь особей способствует еще большему раз%
ненулевыми значениями этих признаков. Пока
витию культуры, что стимулирует дальнейшее
в пуле мемов нет адаптивных знаний, гены ОП и
развитие мозга [1, 11, 12, 28-30]. Ранее мы пока%
ЭОб с ненулевыми значениями ведут себя как
зали с помощью TribeSim, что увеличение ПЖ
слабовредные аллели в равновесии мутации-от%
2 БИОХИМИЯ том 86 вып. 12 2021
1754
МАРКОВ А., МАРКОВ М.
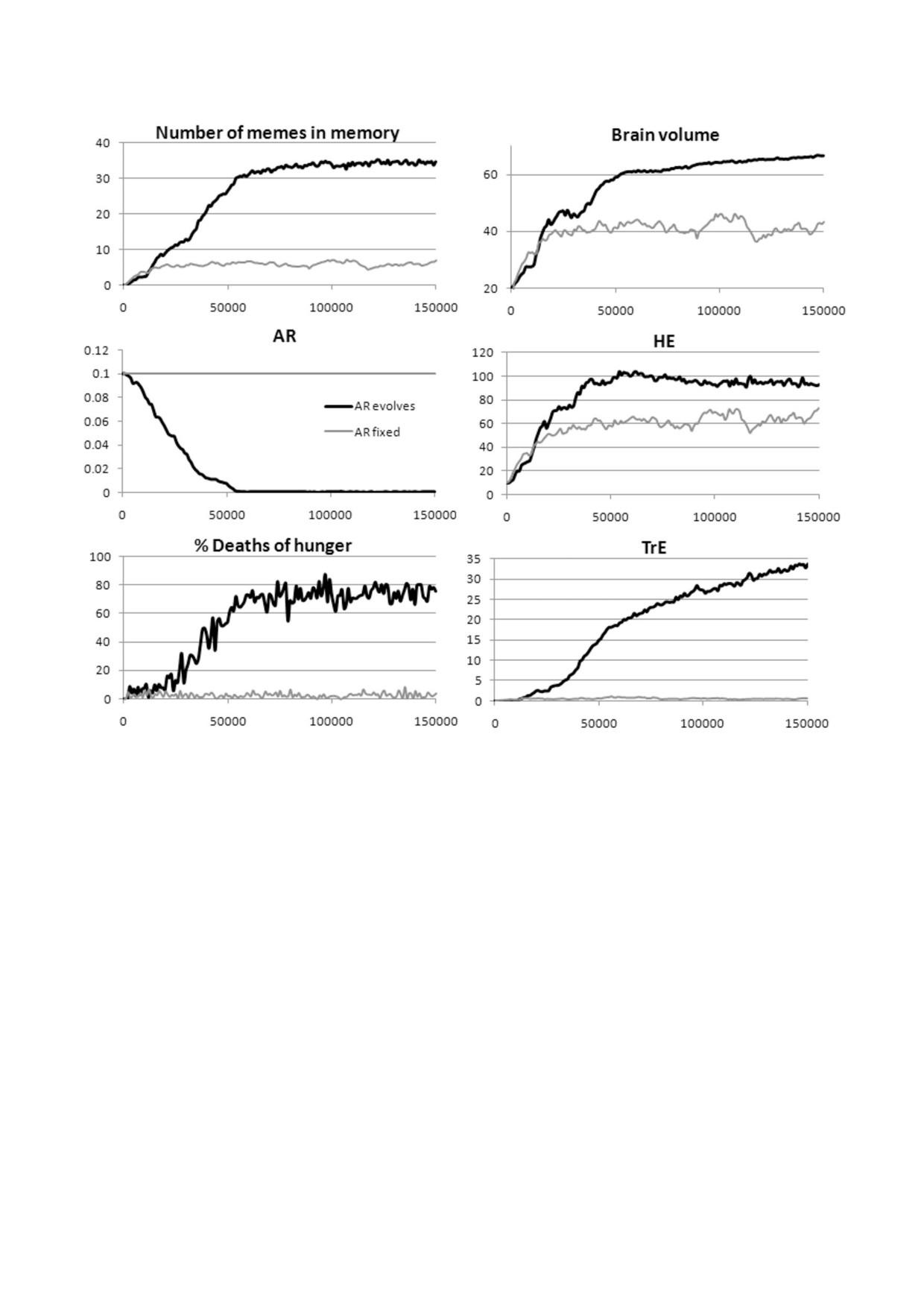
Рис. 5. Культурный драйв (автокаталитическая совместная эволюция мозга, социального обучения и культуры) с фикси%
рованной или эволюционирующей Сст (AR). Параметры: генетическая эволюция ОП, ЭОб (или ОП, ЭОб и Сст); куль%
турная эволюция ЭЭП (TrE) и ЭО (HE); G = 40; начальный уровень Сст равен 0,1. Показаны средние значения из трех
прогонов модели
бор: они немного снижают приспособленность
и Сст начинает быстро снижаться. Чуть позже
(учиться нечему, но размножение обходится до%
(через 20 000-30 000 лет после начала моделиро%
роже), но отбор не может вычистить все слабо%
вания) старение замедляется настолько, что воз%
вредные аллели из генофонда. Как только хотя
никают предпосылки для развития эгоистичной
бы один достаточно простой (умещающийся в
культуры (долгоживущие особи получают боль%
небольшой памяти), но эффективный мем ЭО
ше пользы от этой культуры, чем короткоживу%
появляется в пуле мемов, может начаться отбор
щие). Эгоистичная культура еще больше усили%
на лучшее социальное обучение (группы, в ко%
вает отбор на увеличение ПЖ, потому что она
торых есть индивиды, которые могут выучить и
делает долгую жизнь более полезной, и Сст про%
запомнить этот мем, получают преимущество, и
должает быстро сокращаться. Снижение Сст в
гены более высоких значений ОП и ЭОб начи%
сочетании с развитием эгоистичной культуры
нают распространяться).
приводит к дефициту ресурсов и увеличению
На начальном этапе (первые
10
000-
ресурсного неравенства, что проявляется в уве%
20 000 лет) культурный драйв поддерживается
личении доли смертей от голода. Это создает по%
распространением мемов ЭО и групповым отбо%
ложительную обратную связь, о которой гово%
ром. Развитие кооперативной культуры приво%
рилось выше, что дает дополнительный импульс
дит к более сильному отбору на увеличение ПЖ,
развитию эгоистичной культуры. Обе культуры,
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1755
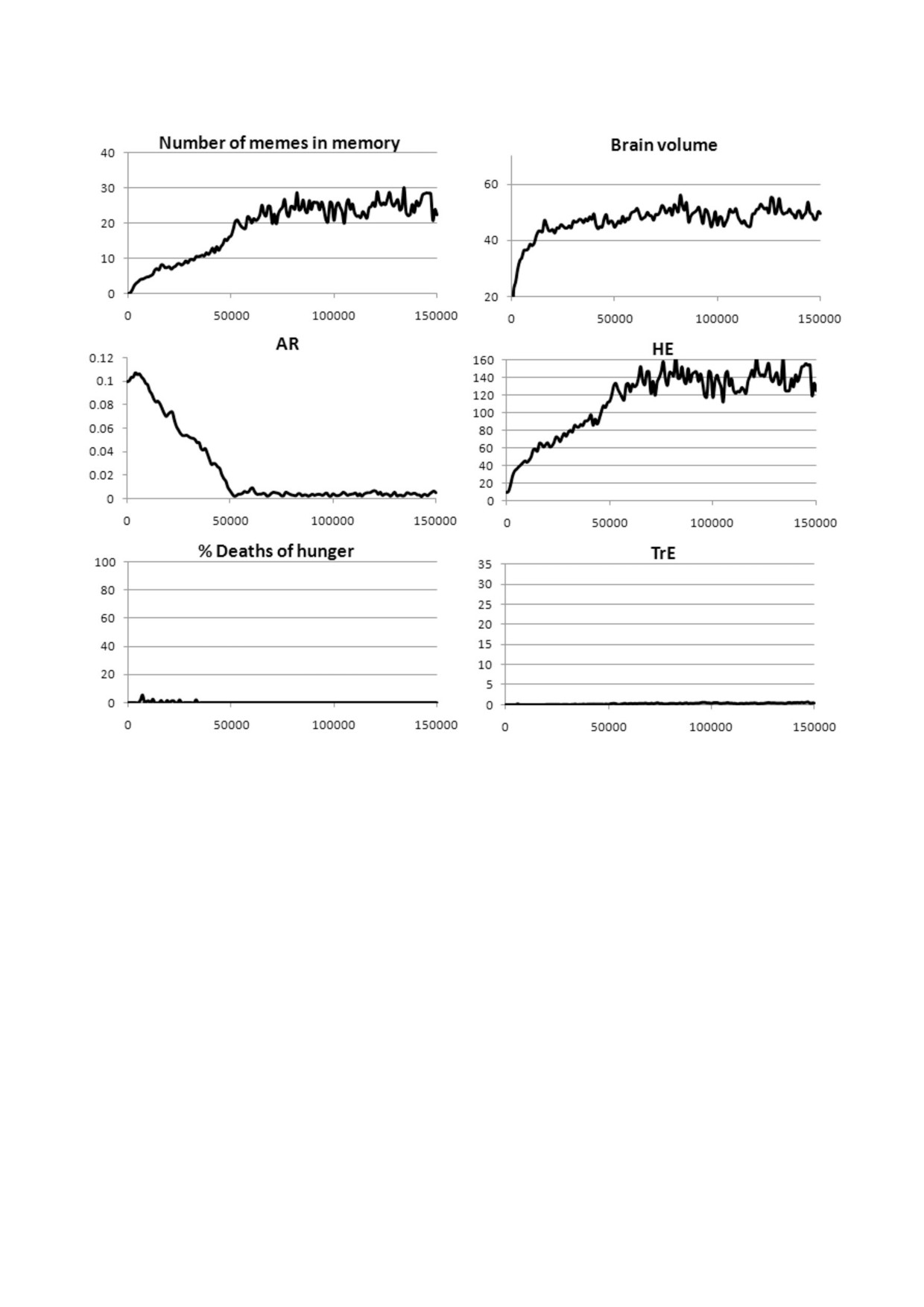
Рис. 6. Культурный драйв (автокаталитическая коэволюция мозга и культуры) и генетическая эволюция Сст. Показаны
средние значения Сст (левые диаграммы) и объема мозга (20 + ОП + 30 ×ЭОб, правые диаграммы) при трех уровнях МГК
и четырех вариантах культуры за годы 100 000-150 000 из трех прогонов модели (± стандартное отклонение). а - Особи
размножаются на протяжении всей жизни (репродуктивный период от 6 лет до смерти), б - репродуктивный период от 6
до 18 лет
БИОХИМИЯ том 86 вып. 12 2021
2*
1756
МАРКОВ А., МАРКОВ М.
кооперативная и эгоистичная, стимулируют
Интересно, что в ходе совместной эволюции
эволюцию социального обучения, что, в свою
мозга, культуры и ПЖ (Сст) различные компо%
очередь, способствует дальнейшему культурно%
ненты комплексной культуры «помогают» друг
му развитию.
другу развиваться. Например, рис. 5 показывает,
Если Сст не эволюционирует, эгоистичная
что культурный драйв сначала был иницииро%
культура при этих параметрах не развивается,
ван кооперативным компонентом культуры (ме%
второй этап культурного драйва не наступает,
мами ЭО). Со временем это создало предпосыл%
мозг и социальное обучение эволюционируют
ки для быстрого развития «эгоистичного» ком%
слабее, и их развитие раньше выходит на стацио%
понента (мемов ЭЭП), что, в свою очередь, спо%
нарный уровень.
собствовало дальнейшему развитию коопера%
Как мы видели из результатов предыдущих
тивного компонента. Эволюция мозга может
экспериментов, с этими параметрами и без
быть стимулирована одним из компонентов
культуры Сст стабилизируется на уровне около
культуры (наиболее выгодным на данный мо%
0,18 (рис. 3, а, столбец 5). При комплексной
мент), но когда дорогостоящий мозг уже развит,
культуре, но без эволюции мозга, Сст стабили%
это может быть использовано другим компонен%
зируется на уровне 0,016 (рис. 3, a, столбец 8).
том культуры для своей эволюции. Например,
Здесь мы видим, что в контексте культурного
если с теми же параметрами, что и на рис. 5 (и
драйва Сст снижается почти до нуля (рис. 5).
рис. 6, a, столбец 8), разрешена только эгоис%
Это означает, что не только увеличение ПЖ ве%
тичная культура, то культурный драйв вообще
дет к более сильному культурному развитию, но
не начинается, и культура не эволюционирует
и культурное развитие способствует эволюции
(рис. 6, a, столбец 6). Если разрешена только ко%
более долгой жизни. Таким образом, моделиро%
оперативная культура, то культурный драйв за%
вание подтвердило теоретическую возможность
пускается, но работает плохо, и мозг остается
петли положительной обратной связи, которая
довольно маленьким (рис. 6, а, столбец 7). Если
способствует развитию мозга и социального
оба компонента культуры разрешены одновре%
обучения за счет эволюции более медленного
менно, то совместная эволюция мозга, социаль%
старения или более длительной жизни. Этот
ного обучения, культуры и долголетия будет
цикл работает, даже если лишь небольшой части
происходить очень быстро и может привести к
индивидов удается реализовать свою врожден%
впечатляющим результатам (рис. 5; рис. 6, а,
ную способность жить долго, а большинство (в
столбец 8).
данном случае около 75%) индивидов умирает
Генетическая и культурная эволюция Сст на
преждевременно из%за нехватки ресурсов.
фоне культурного драйва. В предыдущем разделе
На рис. 6, а показаны значения Сст и объема
мы смоделировали ситуацию, когда на фоне
мозга (средние уровни за 100 000-150 000 лет)
совместной эволюции мозга, социального обу%
для симуляций с культурным драйвом, эволю%
чения и культуры также происходит генетичес%
ционирующей Сст, трех уровней МГК и четырех
кая эволюция Сст (или ПЖ). Однако в реальном
вариантов культуры.
антропогенезе почти наверняка имела место не
Культурный драйв имел место в 6 случаях из
чисто генетическая эволюция этого признака, а
12 (на что указывает объем мозга, значительно
генно%культурная.
превышающий исходный уровень, равный 20).
Нашей следующей задачей было смоделиро%
В трех случаях (столбцы 1, 5, 9) культурный
вать генно%культурную эволюцию Сст в рамках
драйв не мог начаться из%за отсутствия культу%
культурного драйва. Мы использовали тот же
ры. В случаях 2 и 6 (сильная или умеренная
сбалансированный (и, вероятно, реалистичный)
МГК, эгоистичная культура) культурный драйв
набор параметров, что и на рис. 5: умеренная
не начался, потому что мемы ЭЭП в этих ситуа%
МГК, комплексная культура. Единственная раз%
циях недостаточно полезны для запуска отбора
ница в том, что сейчас, наряду с генетической
на более крупный мозг, по крайней мере, пока
эволюцией Сст, также разрешена культурная
ПЖ остается низкой. В случае 11 (нет МГК, ко%
эволюция Сст. Результаты моделирования
оперативная культура) культурный драйв не на%
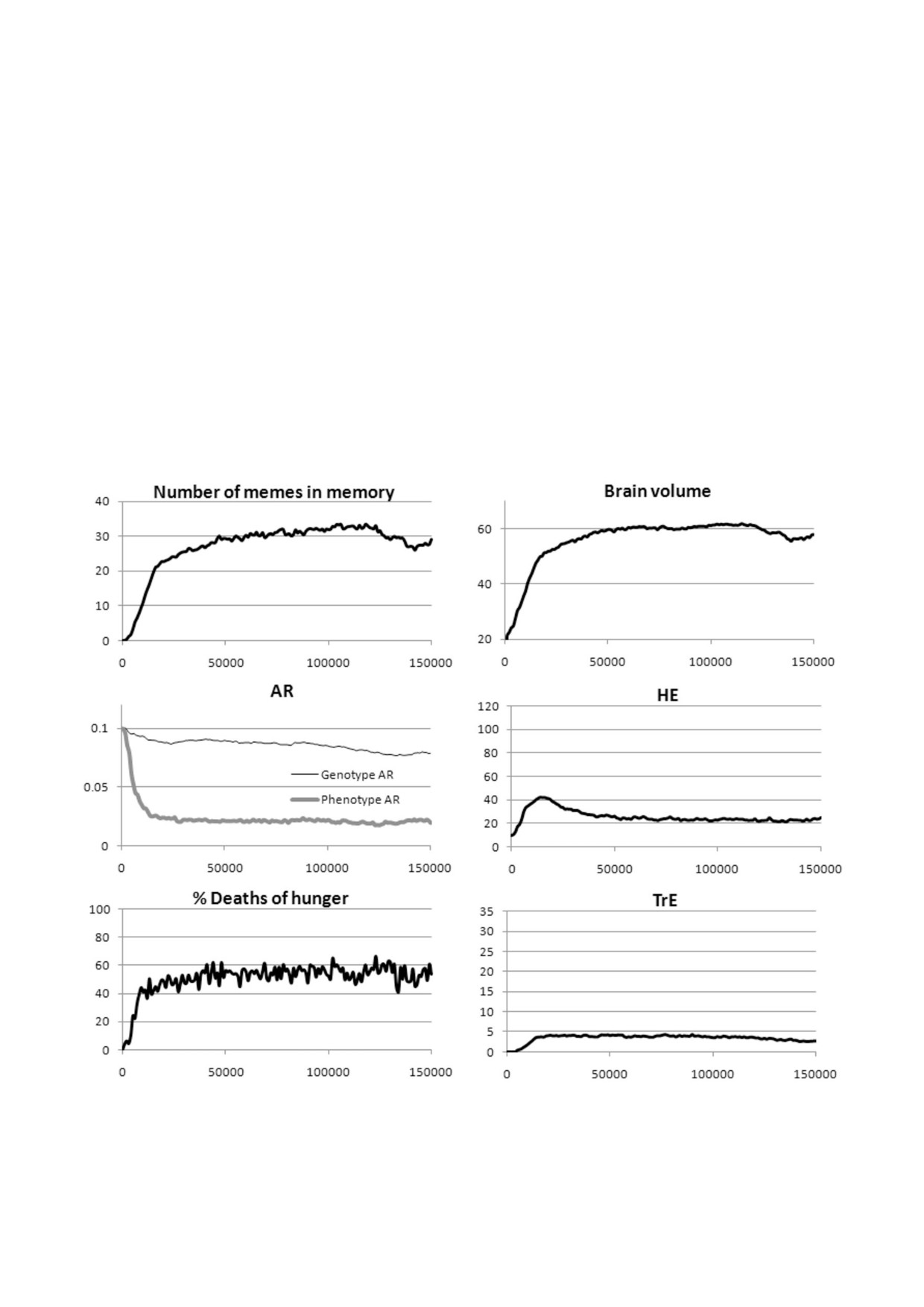
представлены на рис. 7.
чался, потому что мемы ЭО не поддерживаются
Мы видим, что культурный драйв быстро
индивидуальным отбором, а групповой отбор не
стартовал, и мозг в конечном итоге увеличился
работает в отсутствие МГК.
почти до того же объема, что и в случае чисто ге%
Во всех случаях, когда культурный драйв на%
нетической эволюции Сст. Основные отличия
чинался, он приводил к резкому снижению Сст,
заключаются в следующем.
которое было более выраженным, чем при на%
Культура «кирпичных домов» сильно разви%
личии «бесплатной», изначально ограниченной
лась: в пуле мемов накопились продляющие
культуры (рис. 3, а).
жизнь мемы Сст. Эта культура играет роль эво%
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1757
люционного буфера (адаптивной фенотипичес%
витию мощной эгоистичной культуры. Коопера%
кой пластичности), что ослабляет давление от%
тивная культура, которая спровоцировала куль%
бора на генетический компонент Сст. В резуль%
турный драйв вначале, достигла пика примерно
тате генетическая эволюция в сторону более
через 20 000 лет, а затем пришла в упадок, по%
низкого уровня врожденной Сст не зашла очень
скольку мемы Сст постепенно вытесняли мемы
далеко. Получившаяся популяция состоит из
ЭО из пула мемов. И кооперативная, и эгоистич%
рискованной, безрассудной молодежи и осто%
ная культуры не исчезли полностью, а остались в
рожных индивидов старшего возраста. Возмож%
стабильном, умеренно развитом состоянии.
но, нечто подобное имело место и в реальном
Средняя ПЖ стабилизировалась на отметке
антропогенезе: генетически скорость старения
41 год. Интересно, что при чисто генетической
не сильно снизилась по сравнению с другими
эволюции Сст средняя ПЖ составляет всего
обезьянами, но реальный («фенотипический»)
около 31 года, несмотря на более низкое фено%
потенциал долголетия значительно увеличился
типическое значение Сст. Объясняется это тем,
благодаря достижениям культуры (в реальных
что при чисто генетической эволюции Сст чрез%
человеческих культурах есть множество элемен%
вычайно высокоразвитая эгоистичная культура
тов, похожих на продляющие жизнь мемы Сст).
порождает «общество жадных, хитрых долгожи%
Благодаря своим вирусным свойствам мемы
телей и голодающей молодежи». В таком обще%
Сст заполонили пул мемов и препятствовали раз% стве значительная часть молодых индивидов,
Рис. 7. Культурный драйв с генно%культурной эволюцией скорости старения (Сст). Параметры: генетическая эволюция
ОП, ЭОб и Сст; культурная эволюция ЭЭП (TrE), ЭО (HE) и Сст (AR); G = 40; исходный генетический уровень Сст ра%
вен 0,1. Показаны средние значения из восьми прогонов модели
БИОХИМИЯ том 86 вып. 12 2021
1758
МАРКОВ А., МАРКОВ М.
Рис. 8. Коэволюция мозга (объем мозга), культуры (ЭО [HE], ЭЭП [TrE]) и возрастной динамики вероятности смерти
(Сст [AR]) в случае прекращения размножения в определенном возрасте. Все параметры - как на рис. 5, за исключением
того, что индивиды не могут производить потомство после 18 лет. Показаны средние значения из трех прогонов модели
которым не удалось быстро выучить достаточ%
тах репродуктивный потенциал особей не сни%
ное количество мемов ЭЭП, умирает от нехват%
жался с возрастом. Однако у людей фертиль%
ки ресурсов. Это приводит к уменьшению сред%
ность снижается с возрастом, особенно у жен%
ней ПЖ.
щин, репродуктивная функция которых пол%
В целом развитие, которое мы наблюдаем с
ностью прекращается в определенном возрасте
этим набором параметров (умеренная МГК,
(менопауза) [59, 60]. Нашей следующей задачей
комплексная культура, генетическая эволюция
было смоделировать возрастное снижение ре%
дорогостоящих средств социального обучения
продуктивного потенциала, чтобы ответить на
[ОП и ЭОб], генно%культурная эволюция возра%
следующие вопросы: (1) как это повлияет на ко%
стной динамики вероятности смерти), вероят%
эволюцию мозга, культуры и возрастной дина%
но, является адекватным приближением к ре%
мики вероятности смерти; (2) способен ли отбор,
альным эволюционным процессам, которые
управляемый и опосредуемый культурой, при%
имели место в ходе антропогенеза (хотя эффект
вести к эволюции длительного пострепродуктив%
Сст%мемов, вытесняющих другие мемы из пула
ного периода, и если да, то при каких условиях?
мемов, возможно, преувеличен).
Мы повторили моделирование, описанное в
Культурный драйв и эволюция пострепродук?
разделе «Генетическая эволюция Сст и культур%
тивного периода. В описанных выше эксперимен% ный драйв», с одним только изменением: инди%
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1759
виды больше не могли производить потомство
тате объем мозга стабилизируется на более низ%
после 18 лет (репродуктивный период от 6 до
ком уровне, чем при продолжении размножения
18 лет; ранее репродуктивный период составлял
до смерти.
от 6 лет до смерти).
Эти выводы подтверждаются результатами
На рис. 8 показаны подробные результаты
моделирования, представленными на рис. 6, б.
для G = 40 и комплексной культуры. На рис. 6, б
Сравнение рис. 6, а и б показывает, что прекра%
представлены результаты для трех уровней
щение размножения в определенном возрасте
МГК и четырех типов культуры. Эти рисунки
приводит к следующим эволюционным послед%
можно сравнить с рис. 5 и 6, а, чтобы увидеть
ствиям (по сравнению с размножением на про%
влияние прекращения размножения в опреде%
тяжении всей жизни):
ленном возрасте на совместную эволюцию моз%
1) Индивидуальный отбор на долголетие в
га, культуры и возрастной динамики вероятнос%
целом слабее. Особенно это заметно в отсут%
ти смерти.
ствие МГК, когда работает только индивидуаль%
На рис. 8 видно, что при умеренной МГК и
ный отбор. Например, при G = 2000 и эгоистич%
комплексной культуре (мемы ЭЭП и ЭО) от%
ной [«макиавеллистской»] культуре скорость
ключение репродуктивной функции в возрасте
старения Сст составит 0,037 (медленное старе%
18 лет не препятствует ни культурному драйву,
ние, средняя ПЖ = 39,6 лет), если размножение
ни эволюции долгой жизни. Более того, в этом
продолжается до смерти (рис. 6, a), и 0,143
случае особи живут в среднем намного дольше
(быстрое старение, ПЖ = 18,1 года), если раз%
(в среднем ПЖ = 88,6 лет), чем в случае возмож%
множение прекращается к 18 годам (рис. 6, б).
ности размножения до самой смерти (рис. 5,
2) Индивидуально выгодная эгоистичная
ПЖ = 30,7 лет), потому что в популяции почти
культура развивается слабо, потому что в случае
нет смертей от голода. Отсутствие смертей от го%
короткого репродуктивного периода она прино%
лода объясняется тем, что после 18 лет индиви%
сит относительно мало пользы для индивидов.
ды перестают тратить ресурсы на размножение,
Эта культура по%прежнему способна запускать
а рождаемость в целом низкая. Это приводит к
культурный драйв в отсутствие МГК, но даже в
относительному изобилию ресурсов в популя%
этом случае, получающийся в результате объем
ции. К тому же эгоистичная культура практи%
мозга меньше, чем с теми же параметрами и по%
чески не эволюционирует (см. ниже), а значит,
жизненным размножением. Эгоистичная куль%
ресурсного неравенства в группах почти нет.
тура не способствует эволюции длительного
Почему отбор поддерживает долгую жизнь
пострепродуктивного периода. Например, при
особей, если она не способствует их репродук%
G = 2000 и эгоистичной культуре скорость ста%
тивному успеху? В данном случае это происхо%
рения (Сст) составит 0,118, а средняя ПЖ - 20,1
дит исключительно по той причине, что присут%
года. С теми же параметрами, но с пожизнен%
ствие пожилых особей, у которых было доста%
ным размножением, скорость старения стре%
точно времени, чтобы накопить много мемов
мится к нулю (Сст < 0,001), а средняя ПЖ будет
ЭО (т.е. умелых охотников), значительно увели%
равна 54,8 года.
чивает конкурентоспособность группы. Такие
3) При наличии МГК и кооперативной куль%
особи полезны по двум причинам: во%первых,
туры, полезной для группы, групповой отбор
они хорошие охотники и получают много ресур%
поддерживает медленное старение, несмотря на
сов для группы, а во%вторых, они хорошие хра%
то что старые особи не размножаются. Это объ%
нители и распространители знаний. Следова%
ясняется тем, что долгожители приносят пользу
тельно, групповой отбор поддерживает группы с
группе, поскольку накапливают множество ме%
низкими значениями Сст.
мов ЭО и становятся эффективными охотника%
В отличие от охотничьей культуры, которая в
ми и распространителями полезных знаний. При
этой ситуации развивается сильнее, чем когда
некоторых комбинациях параметров (G = 40, ко%
размножение продолжается до самой смерти
оперативная культура) мозг эволюционирует да%
(рис. 5), эгоистичная культура практически не
же сильнее, чем в случае размножения до смерти.
развивается. Это связано с тем, что при корот%
Это связано с тем, что из%за отсутствия смертей
ком репродуктивном периоде мемы ЭЭП не мо%
от голода реальная ПЖ выше, что способствует
гут значительно повысить репродуктивный ус%
развитию культуры и усиливает культурный
пех индивида. Они могли бы защитить от голо%
драйв. Случаи смерти от голода отсутствуют, по%
да, но эта проблема и так не возникает по при%
тому что долгожители не тратят ресурсы на раз%
чинам, указанным выше. Следовательно, мемы
множение, а рождаемость в целом ниже.
ЭЭП в этой ситуации не приобретают вирусных
4) Эффект «взаимопомощи» между коопера%
свойств, не получают широкого распростране%
тивными и эгоистичными культурами исчезает:
ния и не стимулируют развитие мозга. В резуль%
поскольку эгоистичная культура становится ме%
БИОХИМИЯ том 86 вып. 12 2021
1760
МАРКОВ А., МАРКОВ М.
нее полезной, она больше не может значительно
нетическая эволюция Сст не приводит к форми%
улучшить развитие мозга. В результате на пер%
рованию длительного пострепродуктивного пе%
вый план выходит конкуренция между мемами:
риода, поскольку долгая жизнь не улучшает дар%
мемы ЭЭП, присутствующие в пуле мемов как
виновскую приспособленность индивидов, а
практически ненужный балласт, не помогают, а
групповой отбор отсутствует. Однако культурная
немного мешают развитию кооперативной
эволюция Сст приводит к появлению значи%
культуры.
тельного пострепродуктивного периода (сред%
Очевидно, что снижение важности эгоис%
няя ПЖ в 1,5 раза превышает возраст, в котором
тичной культуры, вызванное прекращением
заканчивается размножение), потому что это
размножения в определенном возрасте, будет
полезно для мемов Сст с негативным эффектом
менее выраженным, если пострепродуктивные
(«мемы кирпичных домов»). В результате мемы,
особи смогут передавать ресурсы или иным об%
соблюдая собственные «эгоистичные интере%
разом помогать своему потомству или другим
сы», заставляют индивидов жить дольше, чем
родственникам (т.е. при развитом «кумовстве»
было бы оптимально с точки зрения их генов.
[nepotism]). Действительно, модельные пост%
Интересно, что в этой ситуации (генетичес%
репродуктивные особи не тратят ресурсы на
кая эволюция ОП и ЭОб; Сст% и ЭЭП%мемы;
размножение, и поэтому, с одной стороны, они
прекращение репродукции в 18 лет; G = 2000)
обычно накапливают много ресурсов, а с другой
из%за вирусных свойств мемов Сст прогрессив%
стороны, эти избыточные ресурсы не повышают
ная совместная эволюция мозга и культуры по%
их дарвиновскую приспособленность (потому
ворачивает вспять, мозг в конечном итоге воз%
что они не размножаются). Если бы индивиды
вращается почти к своему первоначальному
умели передавать избыточные ресурсы
объему, и эгоистичная культура деградирует.
родственникам, это повысило бы их инклюзив%
Культурный драйв сначала запускается мемами
ную приспособленность, и эгоистичная культу%
ЭЭП, которые не очень полезны для индивидов,
ра снова стала бы очень полезной. В этом случае
которые прекращают размножаться после 18 лет,
ситуация, показанная на рис. 6, б, стала бы
но все же достаточно полезны, чтобы вызвать
больше похожей на ситуацию, изображенную на
отбор на более крупный мозг. Но как только ОП
рис. 6, a. Тогда длительный пострепродуктив%
и ЭОб начинают расти, продлевающие жизнь
ный период смог бы появиться не только при
мемы Сст («мемы кирпичных домов») быстро
сильной МГК и кооперативной культуре, но
распространяются в пуле мемов. Они не увели%
также при слабой МГК и эгоистичной культуре.
чивают дарвиновскую приспособленность ин%
Это согласуется с гипотезой эволюции менопау%
дивидов (поскольку размножение все равно
зы, называемой также «гипотезой бабушки», ко%
прекращается в 18), но они распространяются
торая предполагает, что пожилые женщины по%
благодаря своим вирусным свойствам и вытес%
вышают свою инклюзивную приспособлен%
няют мемы ЭЭП из культуры. После этого мозг
ность, помогая своему потомству [61-64].
начинает уменьшаться, потому что с этими па%
Другая возможность эволюции длительного
раметрами «в интересах» генов поддерживать
пострепродуктивного периода при слабой МГК
большой мозг ради мемов ЭЭП, но не ради ме%
связана с культурной эволюцией ПЖ. Как уже
мов Сст. С деградацией мозга культура, особен%
упоминалось, продлевающие жизнь мемы явля%
но эгоистичная, постепенно угасает. Остаточная
ются вирусными, потому что они делают своих
культура «кирпичных домов» всё же сохраняет
хозяев более эффективными распространителя%
среднюю ПЖ на уровне около 27,9 лет, что в
ми мемов. Этот эффект не зависит от того, спо%
1,5 раза превышает возраст, в котором прекра%
собны ли долгожители к размножению: доста%
щается размножение.
точно того, что они могут распространять зна%
Способность культурной эволюции генери%
ния. Например, если в ситуации, соответствую%
ровать длинный пострепродуктивный период
щей столбцу 10 на рис. 6, б (генетическая эволю%
независимо от дарвиновской приспособленнос%
ция ОП, ЭОб и Сст; мемы ЭЭП; прекращение
ти особей проявляется еще более отчетливо в
размножения в 18 лет; G = 2000), мы допускаем
идеализированной ситуации «бесплатной», но
культурную эволюцию Сст вместо генетической
ограниченной культуры и неэволюционирую%
(генетическая эволюция ОП и ЭОб; мемы Сст и
щего мозга (рис. 3, б). Например, если G = 40 и
ЭЭП; прекращение размножения в 18 лет;
разрешены только мемы Сст, а размножение
G = 2000), то вместо Сст ≈ 0,118 (средняя ПЖ
продолжается до смерти, то в результате величи%
равна 20,1 года) мы будем иметь Сст ≈ 0,074
на Сст составляет около 0,067, ПЖ ≈ 30,1
(средняя ПЖ равна 27,9 лет) (среднее значение
(рис. 3, б, столбец 5). В этом случае пострепро%
из трех прогонов модели для 100 000-150 000
дуктивного периода нет, т.к. размножение про%
лет). Другими словами, при этих параметрах ге%
должается на протяжении всей жизни. Если с те%
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1761
ми же параметрами размножение прекращается
ной наследственности» и моделями генно%куль%
после 18 лет, то в результате величина Сст сос%
турной коэволюции [1, 9, 11, 55, 65, 66]. В соот%
тавляет около 0,014, ПЖ ≈ 86,9 лет. Таким обра%
ветствии с этим представлением моделирование
зом, в ходе культурной эволюции появляется
выявило сложное взаимодействие между
очень длинный пострепродуктивный период. В
(1) культурной эволюцией полезных для группы
этом случае ни группа не нуждается в нем (нет
и индивидуально полезных поведенческих
охотничьей культуры, так что все индивиды охо%
признаков, (2) генетической эволюцией мозга и
тятся с одинаковой эффективностью, независи%
способностей к социальному обучению и (3) ге%
мо от возраста), ни гены индивидов (потому что
нетической и (или) культурной эволюцией ста%
долгожители не размножаются). Единственны%
рения и ПЖ у моделируемых социальных жи%
ми действующими агентами, которые «хотят»,
вотных.
чтобы индивиды жили дольше, являются мемы:
Особого упоминания заслуживают следую%
длинный пострепродуктивный период возника%
щие моменты.
ет в результате эгоистичной культурной (мемети%
Скорость старения, или увеличения вероят%
ческой) эволюции. Групповой отбор не мешает
ности смерти с возрастом (Сст), по%разному
этому, потому что (1) долгожители не тратят ре%
влияет на скорость эволюции других адаптив%
сурсы на размножение и поэтому никогда не
ных признаков. Моделирование подтвердило,
пропускают охоту вследствие нехватки ресурсов;
что Сст и ПЖ могут влиять на скорость развития
(2) уровень рождаемости низок, и поэтому кон%
других адаптивных признаков (как индивиду%
куренция за ресурсы не очень интенсивна;
ально полезных, так и выгодных для группы).
(3) из%за отсутствия эгоистичной культуры ре%
Направление этого влияния может быть раз%
сурсы в пределах групп делятся поровну между
ным. Быстрое старение может ускорить разви%
всеми членами группы; (4) молодые особи, кото%
тие некоторых признаков благодаря более быст%
рые еще не выучили много мемов Сст, не тратят
рой смене поколений, но замедлить эволюцию
дополнительные ресурсы на жизнеобеспечение,
других. Например, если признак более полезен
но расходуют их вместо этого на размножение.
(сильнее увеличивает дарвиновскую приспособ%
Таким образом, культура «кирпичных домов» не
ленность особей) в популяции долгожителей,
ухудшает конкурентоспособность группы, и
чем в популяции короткоживущих, то этот
групповой отбор не препятствует ее эволюции.
признак будет находиться под более сильным
Для сравнения, в ситуации с теми же параметра%
давлением отбора при низкой Сст. Этот эффект
ми, но с генетической (а не культурной) эволю%
может быть сильнее, чем эффект более длитель%
цией Сст, величина Сст стабилизируется на
ного времени смены поколений, и в результате
уровне около 0,185, ПЖ ≈ 15,5 (если размноже%
признак будет быстрее эволюционировать в по%
ние продолжается до смерти; рис. 3, а, столбец 5)
пуляции долгоживущих особей.
или Сст ≈ 0,208, ПЖ ≈ 14,4 (если размножение
Сст не эволюционирует ради ускорения эво%
прекращается после 18). Это консенсус, достиг%
люции других признаков (старение не является
нутый групповым и индивидуальным отбором
адаптацией для более быстрой адаптивной эво%
по вопросу об «эволюционных интересах генов».
люции). Моделирование не подтверждает, что
Как мы видим, гены в этой ситуации «не нужда%
старение может эволюционировать как средство
ются» в наличии пострепродуктивного периода.
для ускорения эволюции других адаптивных
Эти примеры показывают, что культурная
признаков [67]. Дело в том, что скорость старе%
эволюция Сст способна приводить к появле%
ния сама по себе является важным адаптивным
нию длинного пострепродуктивного периода в
признаком, который находится под прямым и
различных ситуациях, включая те, в которых ге%
очень строгим контролем естественного отбора.
нетическая эволюция Сст не способна этого
Конкретнее, увеличение ПЖ, при прочих рав%
сделать.
ных условиях, значительно повышает дарвино%
вскую приспособленность особей, в то время
как затраты ресурсов, необходимых для повы%
ЗАКЛЮЧЕНИЕ
шения выживаемости, уменьшают ее. В резуль%
тате устанавливается равновесный уровень Сст,
Многие специалисты полагают, что основ%
зависящий от параметров. На фоне этого пер%
ные механизмы дарвиновской эволюции (такие
вичного, прямого давления отбора на Сст, гипо%
как отбор, мутации, дрейф, миграция и реком%
тетические вторичные, косвенные эффекты,
бинация) лишь частично объясняют эволюцию
связанные с тем, что некоторые другие призна%
людей и других животных, способных к соци%
ки эволюционируют быстрее или медленнее при
альному обучению, и что некоторые аспекты их
разных уровнях Сст, не обнаруживаются (даже
эволюции лучше описываются теорией «двой%
при наличии сильного группового отбора).
БИОХИМИЯ том 86 вып. 12 2021
1762
МАРКОВ А., МАРКОВ М.
Равновесные (эволюционно устойчивые)
вирусные свойства, а культурная эволюция мо%
уровни Сст и ПЖ зависят от МГК. Если увели%
жет создавать признаки, снижающие дарвинов%
чение ПЖ связано с затратами ресурсов, а груп%
скую приспособленность индивидов. Типичным
пы конкурируют за ресурсы, то в отсутствие
примером являются мемы, которые снижают
культуры групповой отбор работает на уменьше%
риск смерти (мемы Сст с негативным эффек%
ние ПЖ. При слабой МГК индивидуальный от%
том): они могут распространяться в пуле мемов,
бор на увеличение ПЖ выходит на первый план,
даже если это невыгодно для генов, просто по%
и в ходе эволюции может развиться бóльшая
тому что эти мемы делают своих хозяев более
ПЖ, потому что на индивидуальном уровне вы%
эффективными распространителями мемов.
годы от продления жизни могут перевесить зна%
Из%за этого чисто культурная эволюция Сст час%
чительные затраты ресурсов.
то приводит к более низким значениям этого
Культура обычно способствует генетической
признака (и большей ПЖ), чем чисто генетичес%
эволюции медленного старения и долгой жиз%
кая эволюция того же признака. Во%вторых, ме%
ни, что, в свою очередь, способствует культур%
мы разных категорий конкурируют за одни и те
ному развитию. Культурная эволюция может
же ресурсы (объем памяти индивидов и время
создавать новые культурно сконструированные
обучения). Из%за этой конкуренции сильное
среды, которые приводят к новым направлени%
развитие одних аспектов культуры (например,
ям отбора, действующего на гены (создание
распространение мемов ЭО) может препятство%
культурной ниши) [9, 68, 69]. В соответствии с
вать развитию других (например, распростране%
этим мы показали, что культурное развитие (на%
нию мемов Сст).
копление адаптивных знаний, передаваемых пу%
Генетическая и культурная эволюции Сст
тем социального обучения) может привести, на%
(ПЖ) сложным образом влияют друг на друга.
ряду с другими факторами, к более сильному от%
Если ПЖ эволюционирует как генетически, так
бору на увеличение ПЖ (т.е. на более медленное
и культурно, как это, вероятнее всего, имело
старение). Долгая жизнь позволяет индивидам
место в реальной эволюции человека [4-7], то
накапливать больше знаний и, таким образом,
разворачивается сложный и довольно медлен%
делает культуру более полезной (т.е. усиливает
ный процесс генно%культурной коэволюции.
положительное влияние культуры на дарвинов%
Типичным результатом этого процесса является
скую приспособленность индивидов и конку%
общество, в котором молодые особи из%за гене%
рентоспособность групп). Другими словами, ес%
тической предрасположенности ведут более
ли в общем пуле мемов популяции много полез%
рискованный образ жизни в обмен на возмож%
ных знаний, то как для отдельных индивидов,
ность получить дополнительные ресурсы. Одна%
так и для групп выгоднее, чтобы индивиды жи%
ко с возрастом они накапливают знания, кото%
ли дольше. Индивидуально полезная (напри%
рые помогают снизить риск смерти за счет до%
мер, эгоистичная или макиавеллистская) куль%
полнительных затрат. Вероятно, это похоже на
тура сильнее способствует генетической эволю%
то, что происходило в реальной эволюции:
ции долгожительства, когда МГК слаба, в то
врожденная Сст снизилась у людей умеренно по
время как культура, полезная для группы (нап%
сравнению с другими обезьянами, но популя%
ример, охотничья) делает это, когда МГК силь%
ции накопили большое количество способству%
на. Более того, поскольку долгая ПЖ способ%
ющих выживанию знаний и способов поведе%
ствует развитию культурно обусловленных
ния. Последнее действовало как эволюционный
признаков (потому что долгожители накаплива%
буфер (аналог адаптивной фенотипической
ют больше мемов и передают их большему коли%
пластичности [57]), который замедлял генети%
честву реципиентов), может возникнуть поло%
ческую эволюцию низкой Сст и долгой жизни.
жительная обратная связь между генетической
Культурный драйв способствует развитию
эволюцией ПЖ и культурной эволюцией других
большей ПЖ, что, в свою очередь, усиливает
адаптивных признаков.
культурный драйв. Моделирование подтвердило
Культурная эволюция Сст (ПЖ) в целом
эффективность гипотетической петли положи%
подчиняется тем же законам, что и генетическая
тельной обратной связи, которая усиливает
эволюция, но имеет свои особенности. Разли%
культурный драйв (автокаталитическую совме%
чия между культурной эволюцией ПЖ и генети%
стную эволюцию мозга, социального обучения
ческой эволюцией того же признака объясняют%
и культуры) за счет продления жизни особей.
ся несколькими причинами, из которых две, ве%
Долгая жизнь способствует развитию культуры
роятно, являются наиболее важными. Во%пер%
и мозга, а развитая культура, в свою очередь,
вых, из%за того, что воспроизводство мемов не
способствует эволюции большей ПЖ.
связано неразрывно с воспроизводством генов,
Культура может вносить вклад в развитие
некоторые категории мемов могут приобретать
длительного пострепродуктивного периода.
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1763
Моделирование подтвердило возможность ге%
странением знаний, которые непосредственно
нетической эволюции длительного пострепро%
удлиняют жизнь. Такие мемы имеют вирусные
дуктивного периода за счет «культурного груп%
свойства: они распространяются в пуле мемов и
пового отбора». Это может произойти, если на%
«заставляют» индивидов жить дольше, даже ес%
личие долгожителей, которые с помощью соци%
ли это приводит к снижению дарвиновской
ального обучения накопили ценные знания,
приспособленности особей.
повышает конкурентоспособность группы.
В целом, моделирование показало много%
Предпосылками являются наличие МГК и
численные и разнообразные возможности слож%
культуры, полезной для группы. В отсутствие
ных взаимодействий между эволюцией мозга,
МГК культурный драйв стимулируется только
культуры и ПЖ. Потребуется много дополни%
индивидуально полезными знаниями и инди%
тельных исследований, чтобы понять, какие из
видуальным отбором, в то время как культуре,
этих взаимодействий действительно имели мес%
полезной для группы, отбор не благоприятству%
то в процессе эволюции человека и какова их
ет. Индивидуальный отбор не может обеспечить
относительная важность.
генетическую эволюцию пострепродуктивного
периода, т.к. индивидуальная приспособлен%
Благодарности. Мы благодарим В.И. Голо%
ность не увеличивается из%за того, что индиви%
щапова за оптимизацию программного кода,
ды остаются в живых после прекращения разм%
позволившую завершить моделирование в ра%
ножения независимо от наличия или отсут%
зумные сроки, Е.Б. Наймарк за содержательное
ствия культуры. Кроме того, индивидуально по%
обсуждение статьи, двух анонимных рецензен%
лезное знание перестает быть полезным с эво%
тов за полезные замечания, Г.А. Шиловского за
люционной точки зрения, когда размножение
неоценимую помощь при подготовке русского
прекращается. Однако очевидно, что если пост%
текста статьи.
репродуктивные особи могут делиться своими
Финансирование. Это исследование не полу%
ресурсами с родственниками, то пострепродук%
чало какого%либо специального финансирова%
тивный период может эволюционировать гене%
ния.
тически даже в отсутствие МГК, т.к. это увели%
Конфликт интересов. Авторы заявляют об от%
чивает инклюзивную (совокупную) приспособ%
сутствии конфликта интересов.
ленность [8, 64]. Другая возможность эволюции
Соблюдение этических норм. Настоящая
пострепродуктивного периода в отсутствие
статья не содержит описания каких%либо иссле%
МГК и полезной для группы культуры связана с
дований с участием людей или животных в каче%
культурной эволюцией Сст (ПЖ), т.е. с распро% стве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Laland, K. N. (2017) Darwin’s Unfinished Symphony. How
8. Johnstone, R. A., and Cant, M. A. (2010) The evolution of
Culture Made the Human Mind, Princeton & Oxford:
menopause in cetaceans and humans: the role of demogra%
Princeton University Press.
phy, Proc. R. Soc. B, 277, 3765%3771.
2.
Lewis, H. M., and Laland, K. N. (2012) Transmission
9. Ross, C. T., and Richerson, P. J. (2014) New frontiers in
fidelity is the key to the build%up of cumulative culture,
the study of human cultural and genetic evolution, Curr.
Phil. Trans. R. Soc. B, 367, 2171%2180, doi: 10.1098/
Opin. Genet. Dev., 29, 103%109.
rstb.2012.0119.
10. Kirkwood, T. B. L. (2003) Age differences in evolutionary
3.
Morgan, T. J. H., Uomini, N. T., Rendell, L. E.,
selection benefits, In Understanding Human Development
Chouinard%Thuly, L., Street, S. E., et al. (2015) Experi%
(Staudinger, U. M., and Lindenberger, U., eds) Springer,
mental evidence for the co%evolution of hominin tool%
Boston, MA, pp. 45%57.
making teaching and language, Nat. Commun., 6, 6029,
11. Henrich, J. (2015) The Secret of our Success: How Culture
doi: 10.1038/ncomms7029.
is Driving Human Evolution, Domesticating our Species,
4.
Finch, C. E. (2010) Evolution of the human lifespan and
and Making us Smarter, Princeton, Princeton University
diseases of aging: Roles of infection, inflammation, and
Press.
nutrition, Proc. Natl. Acad. Sci. USA, 107, 1718%1724.
12. Muthukrishna, M., Doebeli, M., Chudek, M., and
5.
Finch, C. E., and Stanford, C. B. (2004) Meat%adaptive
Henrich, J. (2018) The cultural brain hypothesis: how cul%
genes and the evolution of slower aging in humans, Q. Rev.
ture drives brain expansion, sociality, and life history, PLoS
Biol., 79, 3%50.
Comput. Biol., 14, e1006504, doi: 10.1371/journal.pcbi.
6.
Skulachev, V. P., Holtze, S., Vyssokikh, M. Y., Bakeeva,
1006504.
L. E., Skulachev, M. V., et al. (2017) Neoteny, prolonga%
13. Markov, A. V., and Markov, M. A. (2020) Runaway brain%
tion of youth: from naked mole rats to “Naked Apes”
culture coevolution as a reason for larger brains: exploring
(Humans), Physiol. Rev., 97, 699%720.
the “cultural drive” hypothesis by computer modeling,
7.
Skulachev, V. P., Shilovsky, G. A., Putyatina, T. S., Popov,
Ecol. Evol., 10, 6059%6077, doi: 10.1002/ece3.6350.
N. A., Markov, A. V., et al. (2020) Perspectives of Homo
14. Lovejoy, C. O. (2009) Reexamining human origins in light
sapiens lifespan extension: focus on external or internal
of Ardipithecus ramidus, Science,
326,
74%74.e8,
resources? Aging (Albany NY), 12, 5566.
doi: 10.1126/science.1175834.
БИОХИМИЯ том 86 вып. 12 2021
1764
МАРКОВ А., МАРКОВ М.
15.
Raghanti, M. A., Edler, M. K., Stephenson, A. R.,
34.
Fox, K. C. R., Muthukrishna, M., and Shultz, S. (2017)
Munger, E. L., Jacobs, B., et al. (2018) A neurochemical
The social and cultural roots of whale and dolphin brains,
hypothesis for the origin of hominids, Proc. Natl. Acad.
Nat. Ecol. Evol., 1, 1699%1705, doi: 10.1038/s41559%017%
Sci. USA,
115, E1108%E1116, doi:
10.1073/pnas.
0336%y.
1719666115.
35.
Frankenhuis, W. E., and Nettle, D. (2020) Current debates
16.
Stanyon, R., and Bigoni, F. (2014) Sexual selection and
in human life history research, Evol. Hum. Behav., 41, 469%
the evolution of behavior, morphology, neuroanatomy
473.
and genes in humans and other primates, Neurosci.
36.
Stearns, S. C., and Rodrigues, A. M. M. (2020) On the use
Biobehav. Rev., 46, 579%590, doi: 10.1016/j.neubiorev.
of “life history theory” in evolutionary psychology, Evol
2014.10.001.
Hum Behav, 41, 474%485.
17.
Holloway, R. L. (2015) The evolution of the hominid brain,
37.
Stanford, C. B. (1999) The Hunting Apes: Meat Eating and
In Handbook of Paleoanthropology (Henke, W., and
the Origins of Human Behavior, Princeton University Press,
Tattersall, I., eds) Berlin, Heidelberg, Springer%Verlag,
Princeton.
pp. 1961%1987.
38.
Reeve, H. K., and Hölldobler, B. (2007) The emergence of
18.
Leigh, S. R. (2012) Brain size growth and life history in
a superorganism through intergroup competition, Proc.
human evolution, Evol. Biol., 39, 587%599, doi: 10.1007/
Natl. Acad. Sci. USA, 104, 9736%9740, doi: 10.1073/pnas.
s11692%012%9168%5.
0703466104.
19.
Neubauer, S., and Hublin, J.%J. (2012) The evolution of
39.
Byrne, R. W., and Whiten, A. (1988) Machiavellian
human brain development, Evol. Biol., 39, 568%586,
Intelligence: Social Expertise and the Evolution of Intellect in
doi: 10.1007/s11692%011%9156%1.
Monkeys, Apes and Humans, Clarendon Press, Oxford.
20.
Rightmire, G. P. (2004) Brain size and encephalization in
40.
Humphrey, N. K. (1976) The social function of intellect, in
early to Mid%Pleistocene Homo, Am. J. Phys. Anthrop.,
Growing Points in Ethology (Bateson, P. P. G., and
124, 109%123, doi: 10.1002/ajpa.10346.
Hinde, R. A. eds) Cambridge University Press, Cambridge,
21.
Roth, G., and Dicke, U. (2005) Evolution of the brain and
pp. 303%317.
intelligence, Trends Cognit. Sci., 9, 250%257, doi: 10.1016/
41.
Darwin, C. R. (1874) The Descent of Man, and Selection in
j.tics.2005.03.005.
Relation to Sex (2nd Edn.) Appleton, New York, NY.
22.
Sherwood, C. C., Subiaul, F., and Zawidzki, T. W. (2008) A
42.
Richerson, P. J., Baldini, R., Bell, A. V., Demps, K.,
natural history of the human mind: tracing evolutionary
Frost, K., et al. (2016) Cultural group selection plays an
changes in brain and cognition, J. Anatomy, 212, 426%454,
essential role in explaining human cooperation: A sketch of
doi: 10.1111/j.1469%7580.2008.00868.x.
the evidence, Behav. Brain Sci., 39, e30.
23.
Miller, I. F., Barton, R. A., and Nunn, C. L. (2019)
43.
Van Schaik, C. P., Isler, K., and Burkart, J. M. (2012)
Quantitative uniqueness of human brain evolution revealed
Explaining brain size variation: from social to cultural
through phylogenetic comparative analysis, eLife, 8,
brain, Trends Cognit. Sci., 16, 277%284, doi: 10.1016/j.tics.
e41250, doi: 10.7554/eLife.41250.
2012.04.004.
24.
Gavrilets, S., and Vose, A. (2006) The dynamics of
44.
Hublin, J.%J., Neubauer, S., and Gunz, P. (2015) Brain
Machiavellian intelligence, Proc. Natl. Acad. Sci. USA,
ontogeny and life history in Pleistocene hominins, Phil.
103, 16823%16828, doi: 10.1073/pnas.0601428103.
Trans. R. Soc. B, 370, 20140062.
25.
Heyes, C. (2012) New thinking: the evolution of human
45.
Bolund, E. (2020) The challenge of measuring trade%offs in
cognition, Phil. Trans. R. Soc. B, 367, 2091%2096,
human life history research, Evol. Hum. Behav., 41, 502%
doi: 10.1098/rstb.2012.0111.
512.
26.
Laland, K. N., and Rendell, L. (2013) Cultural memory,
46.
Hawkes, K., O’Connell, J. F., and Blurton Jones, N. G.
Curr. Biol., 23, R736%R740, doi: 10.1016/j.cub.2013.07.
(2001) Hadza meat sharing, Evol. Hum. Behav., 22, 113%
071.
142, doi: 10.1016/S1090%5138(00)00066%0.
27.
Whiten, A., Ayala, F. J., Feldman, M. W., and Laland,
47.
Gilby, C. (2006) Meat sharing among the Gombe chim%
K. N. (2017) The extension of biology through culture,
panzees: Harassment and reciprocal exchange, Animal
Proc. Natl. Acad. Sci. USA, 114, 7775%7781, doi: 10.1073/
Behav., 71, 953%963, doi: 10.1016/j.anbehav.2005.09.009.
pnas.1707630114.
48.
Sear, R. (2020) Do human ‘life history strategies’ exist?
28.
Kaplan, H. S., and Robson, A. J. (2002) The emergence of
Evol. Hum. Behav., 41, 513%526.
humans: the coevolution of intelligence and longevity with
49.
Del Giudice, M. (2020) Rethinking the fast%slow continu%
intergenerational transfers, Proc. Natl. Acad. Sci. USA, 99,
um of individual differences, Evol. Hum. Behav., 41, 536%
10221%10226.
549.
29.
Caspari, R., and Lee, S.%H. (2004) Older age becomes
50.
Kramer, K. L., and Russell, A. F. (2015) Was monogamy a
common late in human evolution, Proc. Natl. Acad. Sci.
key step on the hominin road? Reevaluating the
USA, 101, 10895%10900.
monogamy hypothesis in the evolution of cooperative
30.
Crews, D. E. (2003) Human Senescence: Evolutionary and
breeding, Evol. Anthropol., 24, 73%83.
Biocultural Perspectives, Cambridge University Press,
51.
Skulachev, V. P. (1999) Phenoptosis: programmed death of
Cambridge.
an organism, Biochemistry (Moscow), 64, 1418%1426.
31.
Navarrete, A. F., Reader, S. M., Street, S. E., Whalen, A.,
52.
Weismann, A. (1889) Essays upon Heredity and Kindred
and Laland, K. N. (2016) The coevolution of innovation
Biological Problems, Clarendon press, Oxford.
and technical intelligence in primates, Phil. Trans. R.
53.
Longo, V. D., Mitteldorf, J., and Skulachev, V. P. (2005)
Soc. B, 371, 20150186, doi: 10.1098/rstb.2015.0186.
Programmed and altruistic ageing, Nat. Rev. Genet., 6, 866%
32.
Reader, S. M., and Laland, K. N. (2002) Social intelli%
872, doi: 10.1038/nrg1706.
gence, innovation, and enhanced brain size in primates,
54.
Goldsmith, T. C. (2008) Aging, evolvability, and the indi%
Proc. Natl. Acad. Sci. USA, 99, 4436%4441, doi: 10.1073/
vidual benefit requirement; medical implications of aging
pnas.062041299.
theory controversies, J. Theor. Biol., 252, 764%768,
33.
Street, S. E., Navarrete, A. F., Reader, S. M., and Laland,
doi: 10.1016/j.jtbi.2008.02.035.
K. N. (2017) Coevolution of cultural intelligence, extend%
55.
Creanza, N., Kolodny, O., and Feldman, M. W. (2017)
ed life history, sociality, and brain size in primates, Proc.
Cultural evolutionary theory: How culture evolves and why
Natl. Acad. Sci. USA, 114, 7908%7914.
it matters, Proc. Natl. Acad. Sci. USA, 114, 7782%7789.
БИОХИМИЯ том 86 вып. 12 2021
МОЗГ, КУЛЬТУРА И ПРОДОЛЖИТЕЛЬНОСТЬ ЖИЗНИ
1765
56.
Tacutu, R., Thornton, D., Johnson, E., Budovsky, A.,
menopause, and the evolution of human life histories,
Barardo, D., et al. (2018) Human Ageing Genomic
Proc. Natl. Acad. Sci. USA, 95, 1336%1339.
Resources: new and updated databases, Nucleic Acids Res.,
64. Sear, R, and Mace, R. (2008) Who keeps children alive? A
46, D1083%D1090.
review of the effects of kin on child survival, Evol. Hum.
57.
Ghalambor, C. K., Hoke, K. L., Ruell, E. W., Fischer,
Behav., 29, 1%18.
E. K., Reznick, D. N., et al. (2015) Non%adaptive plastici%
65. Whitehead, H., Laland, K. N., Rendell, L.,
ty potentiates rapid adaptive evolution of gene expression
Thorogood, R., and Whiten, A. (2019) The reach of gene%
in nature, Nature, 525, 372%375.
culture coevolution in animals, Nat. Commun., 10, 2405,
58.
Grether, G. F. (2005) Environmental change, phenotypic
doi: 10.1038/s41467%019%10293%y.
plasticity, and genetic compensation, Am. Nat., 166, E115%
66. Boyd, R., and Richerson, P. J. (1985) Culture and the
E123.
Evolutionary Process, The University of Chicago Press.
59.
Lahdenperä, M., Gillespie, D. O. S., Lummaa, V., and
67. Markov, A. V., Barg, M. A., and Yakovleva, E. Yu. (2018)
Russell, A. F. (2012) Severe intergenerational reproductive
Can aging develop as an adaptation to optimize natural
conflict and the evolution of menopause, Ecol. Lett., 15,
selection? (Application of computer modeling for search%
1283%1290, doi: 10.1111/j.1461%0248.2012.01851.x.
ing some conditions when the “fable about hares” is possi%
60.
Maklakov, A. A., and Lummaa, V. (2013) Evolution of sex
ble to explain the evolution of aging), Biochemistry
differences in lifespan and aging: causes and constraints,
(Moscow), 83, 1504%1516.
BioEssays, 35, 717%724.
68. Laland, K. N., and O’Brien, M. J. (2011) Cultural niche
61.
Williams, G. C. (1957) Pleiotropy, natural selection, and
construction: an introduction, Biol. Theory, 6, 191%202,
the evolution of senescence, Evolution, 11, 398%411.
doi: 10.1007/s13752%012%0026%6.
62.
Hamilton, W. D. (1966) The moulding of senescence by
69. Laland, K. N., Odling%Smee, J., and Feldman, M. W.
natural selection, J. Theor. Biol., 12, 12%45.
(2001) Cultural niche construction and human evolution,
63.
Hawkes, K., O’Connell, J. F., Blurton%Jones, N. G.,
J. Evol. Biol., 14, 22%33, doi: 10.1046/j.1420%9101.2001.
Alvarez, H., and Charnov, E. L. (1998) Grandmothering,
00262.x.
COEVOLUTION OF THE BRAIN, CULTURE, AND LIFESPAN:
INSIGHTS FROM COMPUTER SIMULATIONS
Alexander V. Markov1,2* and Mikhail A. Markov1
1 Lomonosov Moscow State University, Faculty of Biology, 119991 Moscow, Russia; e-mail: markov_a@inbox.ru
2 Paleontological Institute of the Russian Academy of Sciences, 117997 Moscow, Russia
Humans possess a number of traits which are rare or absent in other primates, including large brain size, culture, lan%
guage, extended lifespan (LS) and long post%reproductive period. Here, we use a computer model, TribeSim, origi%
nally designed to explore the autocatalytic coevolution of the hominin brain and culture within the framework of the
“cultural drive” theory, to find out how culture and brain could coevolve with LS (or rate of aging). We show that in
the absence of culture, the evolution of LS depends on the intensity of between%group competition (BGC): strong
BGC results in shorter LS. Culture, however, favors the genetic evolution of longer LS even if BGC is strong.
Extended LS, in turn, enhances cultural development, thus creating positive feedback. Cultural evolution of LS
(accumulation of survival%enhancing or survival%impairing knowledge) differs from the genetic evolution of the same
trait, partially because those “memes” (ideas, skills, and behaviors) that reduce the risk of death tend to spread in the
meme pool even if it is not beneficial to genes. Consequently, cultural evolution of aging tends to result in longer LS
than genetic evolution of the same trait. If LS evolves both genetically and culturally, the typical result is a society in
which young individuals, due to their genetic predisposition, lead a riskier lifestyle in exchange for a chance to gain
additional resources, but with age, they accumulate survival%enhancing knowledge. Simulations also showed that cul%
tural evolution of adaptive behaviors can contribute to the genetic evolution of a long post%reproductive period, e.g., if
the presence of knowledgeable long%livers increases the competitiveness of the group.
Keywords: evolution of lifespan, aging, post%reproductive period, gene%culture coevolution, cultural drive, runaway
brain%culture coevolution
БИОХИМИЯ том 86 вып. 12 2021