БИОХИМИЯ, 2021, том 86, вып. 12, с. 1865 - 1874
УДК 577.121.7
БИОЭНЕРГЕТИКА ФИБРОЗА
Обзор
© 2021
Э.И. Якупова1,2, Д.Б. Зоров1,3*, Е.Ю. Плотников1,3*
1 НИИ физико химической биологии имени А.Н. Белозерского,
Московский государственный университет имени М.В. Ломоносова, 119991 Москва, Россия;
электронный адрес: zorov@belozersky.msu.ru; plotnikov@belozersky.msu.ru
2 Институт теоретической и экспериментальной биофизики РАН, 142290 Пущино, Московская обл., Россия
3 Национальный медицинский исследовательский центр акушерства, гинекологии
и перинатологии имени академика В.И. Кулакова, 117997 Москва, Россия
Поступила в редакцию 22.10.2021
После доработки 16.11.2021
Принята к публикации 16.11.2021
Известно, что развитие фиброза сопровождает многие заболевания, являясь как причиной, так и следстви
ем повреждения органов и тканей. Замещение функциональной ткани фиброзным рубцом может приво
дить к нарушению функционирования органов, что часто угрожает жизни больного. Разработка эффектив
ных способов ограничения развития или лечения фиброза требует понимания всех аспектов его патогенеза
в различных органах: от эпителиально мезенхимальной трансформации до экспансии фибробластов. При
чинами фиброза могут быть травма, ишемические повреждения, воспаление и многие другие состояния, ха
рактеризующиеся повторяющимися циклами повреждения и восстановления ткани. Энергетический мета
болизм является основой жизнедеятельности всех клеток организма, и именно его нарушения связаны с
развитием многих заболеваний, поэтому, как неоднократно было показано, воздействие на него может быть
основой терапии таких патологических процессов, как ишемия/реперфузия, эпилепсия, диабет, рак и нев
рологические расстройства. Развитие фиброза также связано с перестройками биоэнергетики клетки.
В данной работе мы проанализировали, какие изменения энергетического метаболизма наблюдаются при
развитии фиброза, и оценили возможность влияния на энергетический метаболизм в качестве подхода ан
тифиброзной терапии.
КЛЮЧЕВЫЕ СЛОВА: фиброз, фибробласты, энергетический метаболизм, гликолиз, дыхание, коллаген.
DOI: 10.31857/S0320972521120101
ВВЕДЕНИЕ
реакции репарации и ангиогенеза для восста
новления трёхмерной структуры и клеточного
Замещение функциональной ткани фиброз
состава ткани. На первой стадии развивается
ным рубцом сопровождает множество заболева
иммунный ответ и острое воспаление, что ха
ний, являясь как причиной, так и следствием
рактеризуется активацией резидентных макро
повреждения органов и тканей [1-4]. Процесс
фагов, нейтрофилов и дендритных клеток [5, 6].
развития фиброза, так называемый фиброгенез,
Продукция этими клетками в очаге поврежде
обычно состоит из двух стадий. В большинстве
ния цитокинов и хемокинов приводит к локаль
случаев фиброза первая стадия инициируется
ной активации фибробластов, превращающихся
разрушением паренхиматозных и стромальных
в миофибробласты, обладающие способностью
клеток органа, при котором гибель клеток вы
участвовать в образовании внеклеточного мат
зывается различными повреждающими агента
рикса (ВКМ) и дополнительно увеличивать
ми и механизмами [3, 4]. При этом запускаются
продукцию провоспалительных цитокинов, хе
мокинов и ангиогенных факторов. Отличитель
Принятые сокращения: ВКМ - внеклеточный мат
ной чертой этих клеток является высокая
рикс; ИЛФ - идиопатический легочный фиброз; КД - ке
экспрессия α актина гладких мышц (α SMA).
тогенная диета; ОЖК - окисление жирных кислот; ОКП - В активированные миофибробласты могут
ограничение калорийности питания; ATF4 - активирую
превращаться и другие популяции клеток под
щий фактор транскрипции 4; 3 BrPA - 3 бромпируват;
воздействием трансформирующего фактора
PHGDH - фосфоглицератдегидрогеназа; PPAR - рецеп
тор, активируемый пролифератором пероксисом; TGF β -
роста β (TGF β), уровень которого увеличивает
трансформирующий фактор роста β.
ся при развитии фиброза [7]. К таким клеткам
* Адресат для корреспонденции.
относятся: резидентные фибробласты ткани,
9
1865
1866
ЯКУПОВА и др.
циркулирующие фиброциты костного мозга,
Ниже будут рассмотрены особенности энер
различные субпопуляции эпителиальных и эн
гетического метаболизма в различных клетках
дотелиальных клеток (которые приобретают фе
при активации профибротических изменений, а
нотип миофибробластов в процессе, называе
также возможные пути воздействия на биоэнер
мом эпителиально мезенхимальным перехо
гетику для уменьшения развития фиброза в ор
дом), а также другие мезенхимальные клетки,
ганах при различных заболеваниях.
такие как перициты и резидентные мезенхи
мальные клетки [8].
Активированные миофибробласты в норме
ЭНЕРГЕТИЧЕСКИЙ МЕТАБОЛИЗМ
инициируют заживление ран и репарацию дру
ПРИ ФИБРОЗЕ
гих повреждений структуры ткани, секретируя
компоненты ВКМ (коллаген I/III, фибронектин
В норме клеточный анаболизм (синтез био
и ламинин). Параллельно в клетках синтезиру
молекул) и катаболизм (распад биомолекул)
ются белки семейства матриксных металлопро
взаимно сбалансированы и находятся под стро
теаз (MMP), которые контролируют уровень
гим контролем различных систем, в том числе и
секреции компонентов ВКМ для предотвраще
биоэнергетики. Развитие фиброза можно отнес
ния чрезмерного его образования [9, 10]. При
ти к сдвигу равновесия в соединительнотканных
сохранении повреждающего стимула стойкое
клетках в сторону анаболизма, поскольку ос
воспаление может усугубить гибель клеток орга
новная роль в развитии фиброза принадлежит
на, и продукты этой гибели, а также массивное
гиперпродукции белковых молекул ВКМ мио
высвобождение цитокинов провоцируют следу
фибробластами. Для обеспечения в миофибро
ющую стадию фиброгенеза. При этом сами мио
бластах повышенных пролиферации и синтеза
фибробласты выделяют аутокринные факторы
белка при фиброзе необходимы серьёзные мета
для поддержания активации с неконтролируе
болические изменения. Показано, что увеличе
мой секрецией и образованием ВКМ в ткани,
ние содержания коллагена в фибробластах соп
что в конечном итоге приводит к образованию
ровождается увеличением экспрессии генов, от
рубцов и дисфункции органа [3, 4].
вечающих за контроль гликолиза, и подавлени
Известно, что воздействия, влияющие на
ем генов, ответственных за окисление жирных
энергетический обмен организма и контролиру
кислот (ОЖК) [19]. Активация фибробластов
ющие переключение доминирующих путей
увеличивает уровень ключевых гликолитичес
энергообеспечения клетки, обладают защитным
ких ферментов, таких как гексокиназа 2 и лак
эффектом при развитии патологических про
татдегидрогеназа (ЛДГ) [20], которые, как счи
цессов, обеспечивая их замедление или обраще
тается, сопровождают увеличение пролифера
ние вспять. К таким воздействиям относятся
ции клеток [20] и синтез коллагена [21]. В ре
низкоуглеводная и кетогенная диета (КД), огра
зультате усиления гликолитического пути обра
ничение калорийности питания (ОКП), а также
зуется больше пирувата, который превращается
фармакологические миметики таких алимен
в ацетил КоА в митохондриальном матриксе
тарных интервенций. Эти воздействия уже сей
[20] и поступает в цикл лимонной кислоты, что
час рассматриваются как перспективные тера
может увеличивать уровень таких метаболитов
певтические подходы при ишемии/реперфузии
как сукцинат, который, по ряду свидетельств,
органов [11, 12], эпилепсии [13], диабете [14],
как раз способствуют фиброзу [22].
онкологических заболеваниях [15, 16], острых и
Наоборот, катаболический фенотип фибро
хронических воспалительных процессах [17] и
бластов и деградация ВКМ ассоциированы с по
нейродегенеративных заболеваниях [18]. Отме
вышенной активностью путей окисления жир
тим, что некоторые из данных патологий так
ных кислот [19, 23]. Показано, что внутрикле
или иначе связаны с развитием фиброза, явля
точное ОЖК подавляется при тубулоинтерсти
ясь либо основой патогенеза заболевания, либо
циальном фиброзе у мышей и людей, а его вос
следствием повреждения ткани в острой фазе
становление защищает от фиброза [24]. При по
болезни. Несомненно, уменьшение поврежде
нижении окисления жирных кислот во время
ния органа во время патологии должно приво
фиброзного повреждения почек активируется
дить к уменьшению развития последующего
гликолиз, что, как описано выше, и могло яв
фиброза, и в этом смысле данные воздействия
ляться причиной усиленного прогрессирования
можно рассматривать как противофиброзные.
фиброза [25].
Однако намного важнее оценить возможность
Важно отметить, что была продемонстриро
влияния на энергетический метаболизм с целью
вана не просто корреляция определенного типа
уменьшения рубцовой трансформации ткани
энергетического метаболизма с анаболичес
уже после повреждения.
кой/катаболической активностью фиброблас
БИОХИМИЯ том 86 вып. 12 2021
БИОЭНЕРГЕТИКА ФИБРОЗА
1867
тов, а прямая причинно следственная связь
нокислот [30]. Этот фактор может обеспечивать
между этими процессами. Так, воздействие на
связь сигнализации, активируемой TGF β, и
сигнальные пути PPAR, активирующее окисле
метаболизма клетки, поскольку продукция
ние жирных кислот, приводило к снижению
ATF4 способствует транскрипции генов, коди
синтеза ВКМ, интернализации внеклеточных
рующих ферменты пути биосинтеза серина и
молекул коллагена и их лизосомальной деграда
глицина, а также транспортёра глюкозы
1
ции. Аналогичным эффектом обладало ингиби
(GLUT1). Другой активируемый TGF β путь
рование гликолиза в фибробластах, приводя к
связан с транскрипционным фактором Smad3 и
снижению транскрипции гена коллагена даже в
регуляцией комплекса mTORC1 [30], который
условиях нормальной (не усиленной) активнос
является известным сенсором метаболического
ти путей ОЖК. Поскольку эффективность гли
статуса клетки (реагируя на недостаток пита
колиза как источника АТФ невысока, он может
тельных веществ и энергетических субстратов),
иметь большую, по сравнению с окислительным
обеспечивая ещё один механизм метаболичес
фосфорилированием, значимость для маловас
кой регуляции функций миофибробластов при
куляризованных и потенциально гипоксичных
фиброзе [30].
тканей, таких как соединительная ткань и фиб
Также известно, что обработка культивируе
розный рубец. Однако, помимо обеспечения
мых фибробластов TGF β приводит к повы
механизма генерирования энергии, гликолиз
шенной экспрессии гликолитического фер
производит побочные продукты, которые регу
мента 6 фосфофрукто 2 киназы/фруктозо 2,6
лируют фиброз, например лактат. Снижение
бисфосфатазы 3 [31], а подавление гликолиза с
внеклеточного pH вследствие транспорта мо
помощью ингибитора фосфофруктокиназы,
лочной кислоты из клетки монокарбоксилат
3 (3 Пиридинил) 1 (4 пиридинил) 2 пропен
ным транспортёром способствует дифференци
1 он (3PO), препятствовало повышению синте
ровке фибробластов за счёт активации TGF β,
за белков ВКМ [32]. Аналогично, ингибитор
поскольку кислый рН индуцирует переход так
PHGDH (NCT 503) снижает индуцированный
называемого латентного внеклеточного TGF β в
TGF β синтез белка коллагена [28]. Использо
активную форму [26], кроме того, сам лактат
вание ещё одного ингибитора гликолиза, 3
может служить дополнительным источником
бромпирувата (3 BrPA), также приводило к
энергии для этих клеток [26, 27].
уменьшению индуцированной TGF β продук
Вторым важным для фиброза продуктом
ции коллагена в клеточной линии фибробластов
гликолиза является 3 фосфоглицериновая кис
почки NRK 49F in vitro [32]. При этом с по
лота. Поскольку именно коллаген является од
мощью молекулярного докинга было доказано,
ним из главных составляющих ВКМ, считается,
что 3 BrPA взаимодействует прежде всего с фер
что усиление гликолитического пути энергети
ментами аэробного гликолиза, снижая их актив
ческого обмена может способствовать выработ
ность, поэтому показанные антифибротические
ке ВКМ именно через обеспечение синтеза кол
эффекты можно однозначно связать именно с
лагена. Предполагается, что это связано с тем,
этим механизмом.
что треть аминокислотного состава коллагена
Наконец, ещё одним следствием усиления
представлена глицином, который частично вы
гликолиза является потенциальное изменение
рабатывается из продуктов метаболизма глюко
экспрессии ряда генов. Этот процесс обеспечи
зы [28]. Так, один из путей синтеза глицина в ор
вается через ацетилирование гистонов, основ
ганизме проходит через серин, который, в свою
ным источником ацетильной группы для кото
очередь, производится из 3 фосфоглицерино
рого является ацетил КoA, образующийся из
вой кислоты, образующейся в гликолизе. Хотя
пирувата [33]. Теоретически усиление гликолиза
считается, что данный путь синтеза глицина не
могло бы приводить к повышению количества
может покрыть всю потребность организма в
образующегося пирувата, а значит оно потенци
коллагене [29], известно, что профибротичес
ально способно увеличивать ацетилирование
кий ростовой фактор TGF β индуцирует
различных белков. Однако было показано, что
экспрессию de novo ферментов пути синтеза се
при развитии хронического фиброза происхо
рина и глицина, таких как фосфоглицератдегид
дит инактивация пируватдегидрогеназного
рогеназа (PHGDH), фосфосерин аминотранс
комплекса (ПДК) в митохондриях [34], вызывая
фераза 1 (PSAT1), фосфосеринфосфатаза (PSPH)
соответствующее снижение синтеза ацетил
и серингидроксиметилтрансфераза 2 (SHMT2)
КоА. В результате TGF β вызывает в фибро
[21, 28, 30]. Было показано, что TGF β также
бластах выраженное сокращение запасов аце
увеличивает продукцию активирующего факто
тил КоА, что ведёт к снижению ацетилирова
ра транскрипции 4 (ATF4), главного регулятора
ния лизина гистонов, а также многих ещё не
транскрипции компонентов метаболизма ами
идентифицированных белков в фибробластах
БИОХИМИЯ том 86 вып. 12 2021
9*
1868
ЯКУПОВА и др.
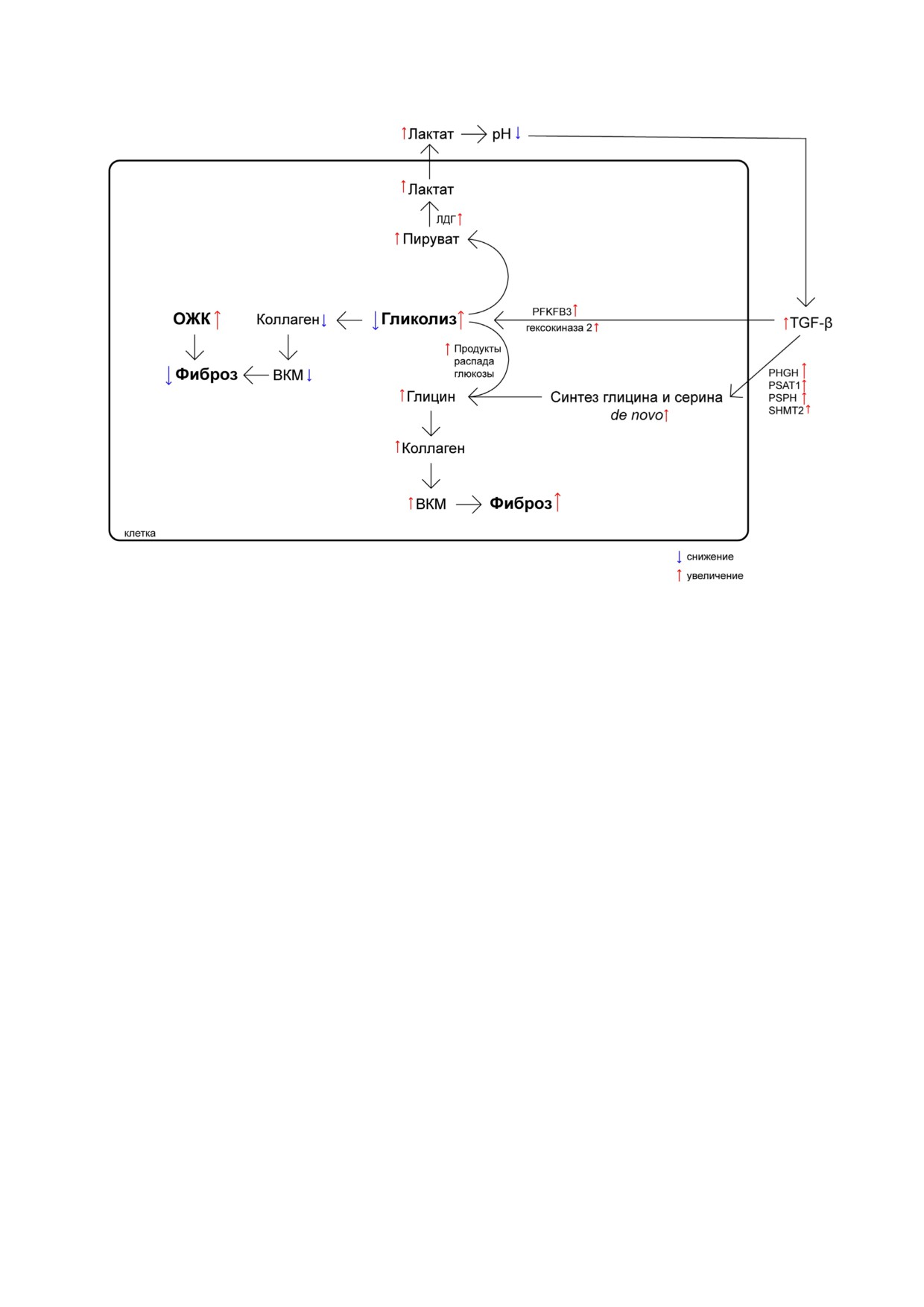
Рис. 1. Сдвиг в сторону того или иного пути энергетического метаболизма влияет на развитие фиброза. При повышении
ОЖК фиброз уменьшается, как и при понижении активности гликолиза, за счёт уменьшения продукции коллагена и
ВКМ в целом. Активация гликолиза увеличивает развитие фиброза за счёт поставки продуктов распада глюкозы в систе
му синтеза глицина, а далее синтеза коллагена/ВКМ. TGF β увеличивает активность ферментов синтеза глицина и сери
на de novo. Поставка пирувата гликолизом и увеличение активности ЛДГ повышает содержание лактата, что понижает pH,
и, как следствие, индуцирует TGF β, который вызывает повышение экспрессии ряда ферментов
почек крыс [34]. Таким образом, регуляторное
в том числе опухолевых [40]. С другой стороны,
ацетилирование обеспечивает связь между из
как и в случае гликолиза, не менее важным мо
менениями клеточного метаболизма и эпигене
жет быть обеспечение за счёт глутаминолиза
тической регуляцией транскрипции и функций
синтеза коллагена, поскольку пролин, составля
белков [35].
ющий вместе с глицином до 57% аминокислот
Обобщённая схема влияния энергетики
коллагена, напрямую продуцируется из глутама
клетки на развитие фиброза представлена на
та [41].
рис. 1.
Важно отметить, что параллельно со сдвигом
метаболизма в сторону гликолиза при фиброзе
МЕТАБОЛИЧЕСКОЕ ВОЗДЕЙСТВИЕ
происходит активация метаболизма глутамина
НА ФИБРОЗ В ЭКСПЕРИМЕНТАЛЬНЫХ
(деградации его до глутамата в процессе глута
МОДЕЛЯХ
минолиза), причём эти изменения напрямую
связаны с действием на фибробласты TGF β.
Хотя исследования in vitro являются простой
Показано in vitro, что метаболизм глутамина
и часто используемой моделью для изучения
усиливается при воздействии TGF β на фибро
фиброза (табл. 1), ряд исследователей отмечает
бласт подобные клетки лёгких [36-38] и ко
некоторые ограничения работы на клеточных
жи [39]. Роль усиления глутаминолиза до конца
культурах: 1) культуры клеток не воспроизводят
неясна. С одной стороны, предполагается что
полностью патофизиологию тканевого фиброза
это может быть альтернативный путь обеспече
in vivo, что было подтверждено при сравнении
ния энергией, поскольку гликолиз не может
профилей экспрессии генов [42]; 2) монослой
полностью удовлетворить потребности в синте
ные клеточные культуры не дают представления
зе АТФ. Усиление глутаминолиза характерно
о трёхмерном фиброзном микроокружении [43];
для многих активно пролиферирующих клеток,
3) миофибробласты из идиопатического лёгоч
БИОХИМИЯ том 86 вып. 12 2021
БИОЭНЕРГЕТИКА ФИБРОЗА
1869
Таблица 1. Моделирование фиброза in vitro
Способ
Клеточная линия/модель
активации
Ключевые результаты работы
Ссылки
фиброза
Фибробласты лёгких
инкубация
обработка клеток TGF β стимулирует аэробный гликолиз; TGF β
[47]
плода человека (IMR 90
клеток в
индуцированное метаболическое перепрограммирование является
клетки)
присутствии
зависимым от митоген активируемой протеинкиназы p38
TGF β
Первичная культура фиб
инкубация
TGF β подавляет биосинтез ацетил КоА посредством регуляции
[34]
робластов, выделенная
клеток в
комплекса пируватдегидрогеназы (ПДК)
из фиброзных почек (3 дня
присутствии
после односторонней
TGF β
обструкции мочеточни
ка) (UUOF) и нормаль
ных почек (NRKF) сам
цов крыс Sprague Dawley
Первичные звездчатые
инкубация
ингибирование ацетил КoA карбоксилазы снижает активацию TGF β
[48]
клетки печени
клеток в
стимулированных первичных звёздчатых клеток печени; такое инги
присутствии
бирование препятствовало метаболическому переключателю, необ
TGF β
ходимому для индукции гликолиза и окислительного фосфорилиро
вания во время активации; показано, что клетки нуждаются в липо
генезе de novo для активации
Выделенные фиброблас
образцы па
келоидные фибробласты продемонстрировали сдвиг своего метабо
[49]
ты из нормальной кожи
циентов с
лического фенотипа с окислительного фосфорилирования на аэроб
и кожи из келоидных
келоидным
ный гликолиз; анализ генов и белков показал, что экспрессия глико
рубцов
рубцом
литических ферментов повышена в келоидных фибробластах по
сравнению с нормальными фибробластами; кроме того, наблюдался
более высокий приток глюкозы и продукция лактата в келоидных
фибробластах, а их пролиферация подавлялась дозозависимым и за
висящим от времени образом после ингибирования гликолиза
2 дезоксиглюкозой
Линия фибробластов
инкубация
дифференцировка миофибробластов лёгких сопровождается усиле
[31]
легких человека MRC 5;
клеток в
нием гликолитического пути энергетического обмена;
миофибробласты лёгких
присутствии
повышенная экспрессия гликолитического фермента 6 фосфофрук
пациентов с идиопати
TGF β
то 2 киназы/фруктозо 2,6 бифосфатазы 3 (PFKFB3) связана с уси
ческим легочным фибро
лением гликолиза, который наблюдался при обработке клеток фиб
зом (ИЛФ)
робластов TGF β; эти изменения коррелировали со степенью диффе
ренцировки фибробластов в миофибробласты;
ингибирование PFKFB3 способствует обратной дифференцировке
миофибробластов от пациентов с ИЛФ
Изолированные мезен
три вида
все три способа активации фибробластов были связаны с подавлени
[19]
химальные клетки, выде
воздействия:
ем генов передачи сигналов рецептора, активируемого пролиферато
ленные из подушек лап
гипоксия
ром пероксисом (PPAR), и с активацией генов гликолиза, тогда как
мышей C3H или B6;
(2% О2),
PPAR является индуктором пути ОЖК;
культура первичных фиб
присутствие
генетическое перепрограммирование клеток (с помощью CRISPR
робластов кожи человека
TGF β или
Cas9) для подавления ОЖК вызывало повышение внеклеточных
тромбоци
уровней фибронектина, коллагена 1 и PAI1;
тарного
подавление гликолиза, наоборот, снижает уровень белков ВКМ
фактора
роста BB
(PDGF BB)
Культура первичных
инкубация
TGF β индуцирует экспрессию гликолитических генов и увеличивает
[21]
фибробластов лёгких че
клеток в
гликолитический поток;
ловека
присутствии
TGF β индуцирует экспрессию ферментов пути синтеза de novo сери
TGF β
на и глицина, что необходимо для продукции коллагена
Нормальные фиброблас
инкубация
обработка фибробластов ингибитором PHGDH (NCT 503) снижает
[28]
ты лёгких человека
клеток в
индуцированный TGF β синтез коллагена
(Lonza);
присутствии
фибробласты NIH 3T3
TGF β
БИОХИМИЯ том 86 вып. 12 2021
1870
ЯКУПОВА и др.
Окончание таблицы 1
Способ
Клеточная линия/модель
активации
Ключевые результаты работы
Ссылки
фиброза
Культура первичных
инкубация
TGF β увеличивает продукцию активирующего фактора транскрип
[30]
фибробластов человека;
клеток в
ции 4 (ATF4), главного регулятора транскрипции метаболизма ами
клеточная линия pHLF,
присутствии
нокислот;
экспрессирующая доми
TGF β
ATF4 усиливает транскрипцию генов, кодирующих ферменты пути
нантно негативную му
биосинтеза серина и глицина de novo, и транспортёра глюкозы 1
тантную форму белка
(GLUT1);
4E BP1
индуцированная TGF β продукция ATF4 зависела от кооперации
между канонической передачей сигналов TGF β через Smad3 и акти
вацией комплекса mTORC1 и его нижележащей мишени, эукариоти
ческого фактора инициации трансляции 4E связывающего белка 1
(4E ВР1)
ного фиброза (ИЛФ) представляют собой очень
воляют дать однозначный ответ об антифиброз
гетерогенную популяцию, что не учитывается в
ной эффективности подавления гликолиза.
моделях in vitro [44-46].
Уже упоминавшийся 3 BrPA оказался до
Поэтому несмотря на положительные эф
вольно эффективным при экспериментальном
фекты подавления гликолиза, полученные на
фиброзе у животных. Его использование в каче
клеточных моделях фиброза, такие подходы
стве ингибитора гликолиза при внутрибрюшин
должны быть проверены in vivo. Таких работ по
ном введении мышам с унилатеральной
ка очень мало (табл. 2), и их результаты не поз
обструкцией мочеточников существенно подав
Таблица 2. Моделирование фиброза in vivo
Способ
Модель
активации
Ключевые результаты работы
Ссылки
фиброза
Фиброз кожи, индуциру
у мышей после об
транскриптомный анализ показал, что во время гипоксии наи
[19]
емый гамма облучением
лучения и людей,
более значимо подавляются гены белков, участвующих в
задней конечности мы
больных раком и
окислительном фосфорилировании, ОЖК и цикле трикарбо
шей C3H/HeJ или мы
получавших луче
новых кислот;
шей B6.129S1
вую терапию, наб
в фибробластах пациентов с фиброзом кожи наблюдалось
Cd36tm1Mfe/J;
людаются призна
значительное усиление процессов гликолиза, включая усиле
фиброз кожи человека
ки фиброза кожи
ние транспорта глюкозы и подавление глюконеогенеза
после лучевой терапии
(первичные фибробласты)
Индуцированный лёгоч
мышам интратра
ингибирование гликолиза с помощью 3PO (ингибитор
[31]
ный фиброз у мышей
хеально вводили
PFKFB3) приводило к снижению лёгочного фиброза у мышей
C57BL/6
блеомицин или
с индукцией фиброза;
TGF β
отмечалось увеличение лактата в лёгких
Ткани лёгких, полученные
ИЛФ
экспрессия ферментов пути синтеза серина de novo активиру
[21]
от пациентов с ИЛФ при
ется во время фиброза in vivo
трансплантации лёгких
Индуцированный лёгоч
интратрахеально
у мышей, получавших ингибитор фосфоглицератдегидрогена
[28]
ный фиброз у мышей
вводили раствор
зы (PHGDH), через 7 дней после интратрахеального введения
C57BL/6
блеомицина
блеомицина наблюдалось ослабление фиброза лёгких
Четыре модели неалко
модели фиброза
ингибирование ацетил КoA in vivo значительно снижает фиб
[48]
гольного стеатогепатита
печени грызунов,
роз в четырех моделях неалкогольного стеатогепатита
мышей и крыс
вызвали диетой
или гепатотокси
ном, диэтилнитро
замином
БИОХИМИЯ том 86 вып. 12 2021
БИОЭНЕРГЕТИКА ФИБРОЗА
1871
Рис. 2. Схематическое изображение влияния баланса биоэнергетических путей на фиброз
ляло фиброзное повреждение почек и выработ
возрастные заболевания [55, 56], к которым от
ку интерстициального коллагена, причём этот
носится острое и хроническое повреждение по
эффект был дозозависимым [32].
чек [12]. Во время ОКП активируются много
Хотя в ряде работ было показано, что инги
численные сигнальные пути, включая те, кото
бирование ферментов может использоваться
рые влияют на рост, метаболизм, аутофагию, ре
для противофиброзной терапии (табл. 2), этот
акцию на окислительные вызовы, стресс, реге
подход также не лишён недостатков [50]: 1) де
нерацию и воспаление [57]. Благоприятные эф
леция гена Phgdh у мышей вызывает эмбрио
фекты КД также описаны [11, 12-16, 18], но
нальную летальность из за множественных де
есть исследования, в которых предупреждается,
фектов развития, особенно в центральной
что мы должны с осторожностью использо
нервной системе [51]; 2) низкомолекулярный
вать КД, например, потому что она может уве
ингибитор PHGDH, NCT 503, нуждается в тес
личить сердечный фиброз [58]. Было показано,
тировании на биодоступность, в том числе на
что в сердечной ткани пациентов с фибрилля
проникновение через гематоэнцефалический
цией предсердий выявляется повышение содер
барьер, если планируется его применение по
жания бета оксибутирата, кетогенных амино
предотвращению фиброза в головном мозге;
кислот и глицина [59]. В исследовании, имити
3) необходимы исследования, оценивающие
рующем КД у крыс посредством введения экзо
долгосрочные эффекты ингибирования PHGDH
генного β оксибутирата, было продемонстриро
и других ферментов гликолиза [50], поскольку
вано снижение биогенеза митохондрий и усиле
фиброз развивается в течение длительного вре
ние сердечного фиброза [58]. Как ОКП, так
мени, а значит и применение таких веществ
и КД влияют на процесс развития фиброза, а
должно быть довольно продолжительным;
влияют ли они на уже сформировавшийся фиб
4) хотя ингибирование PHGDH может оказать
роз, до сих пор неизвестно.
ся полезным для уменьшения накопления кол
лагена во время фиброгенеза за счёт снижения
доступности глицина, оно также может приво
ЗАКЛЮЧЕНИЕ
дить к снижению уровня глутатиона, необходи
мого для поддержания окислительно восстано
Таким образом, несмотря на ограниченное
вительного гомеостаза в тканях, что актуально,
количество экспериментальных данных, совре
в частности, для лёгких [52], характеризующих
менные представления сводятся к идее, что гли
ся, с одной стороны, высоким содержанием
колитический путь энергетического метаболиз
коллагена и высокой вероятностью развития
ма может быть критической точкой регуляции
фиброза [53], а с другой - высокой вероят
синтеза коллагена и развития фиброза при раз
ностью окислительного стресса из за повышен
личных патологиях. Было показано, что инги
ного парциального давления кислорода в тка
бирование некоторых ферментов гликолиза
ни.
приводит к уменьшению фиброза как в клеточ
Исследования показывают, что ингибирова
ных моделях, так и у животных. Кроме того,
ние дифференцировки в миофибробласты воз
важным является соотношение различных пу
можно путём обеднения среды глутамином, так
тей энергообеспечения клетки (рис. 2), по
же, как и ингибирование глутаминазы в покоя
скольку показано, что, например, подавление
щихся звёздчатых клетках печени [54].
пути ОЖК приводит к повышению синтеза бел
В контексте метаболических модуляций
ков ВКМ даже при неизменной активности гли
фиброза можно отметить, что одним из спосо
колиза.
бов повлиять на энергетический метаболизм ор
На сегодняшний день подход, связанный с
ганизма являются манипуляции характера пита
влиянием на энергетический обмен для умень
ния (алиментарные подходы), наиболее извест
шения эффектов фиброза, представляется пер
ными и изученными из которых являются КД
спективной стратегией, однако имеет свои огра
и ОКП. ОКП, как известно, облегчает патологи
ничения, что требует более глубокого экспери
ческие процессы [12, 17, 55, 56], в частности,
ментального анализа.
БИОХИМИЯ том 86 вып. 12 2021
1872
ЯКУПОВА и др.
Финансирование. Работа выполнена при фи
Соблюдение этических норм. Настоящая
нансовой поддержке Российского научного
статья не содержит описания каких либо иссле
фонда (грант № 21 75 30009).
дований с участием людей или животных в каче
Конфликт интересов. Авторы заявляют об от
стве объектов.
сутствии конфликта интересов.
СПИСОК ЛИТЕРАТУРЫ
1.
Plotnikov, E. Y., Brezgunova, A. A., Pevzner, I. B., Zorova,
cancer xenografts, Clin. Cancer Res., 19, 39053913,
L. D., Manskikh, V. N., et al. (2018) Mechanisms of LPS
doi: 10.1158/1078 0432.CCR 12 0287.
induced acute kidney injury in neonatal and adult rats,
16.
Otto, C., Kaemmerer, U., Illert, B., Muehling, B.,
Antioxidants, 7, 105, doi: 10.3390/antiox7080105.
Pfetzer, N., et al. (2008) Growth of human gastric cancer
2.
Popkov, V. A., Andrianova, N. V., Manskikh, V. N.,
cells in nude mice is delayed by a ketogenic diet supple
Silachev, D. N., Pevzner, I. B., et al. (2018) Pregnancy pro
mented with omega 3 fatty acids and medium chain
tects the kidney from acute ischemic injury, Sci. Rep., 8,
triglycerides, BMC Cancer, 8, 122, doi: 10.1186/1471
14534, doi: 10.1038/s41598 018 32801 8.
2407 8 122.
3.
Henderson, N. C., Rieder, F., and Wynn, T. A. (2020)
17.
González, O. A., Tobia, C., Ebersole, J. L., and Novak,
Fibrosis: from mechanisms to medicines, Nature, 587,
M. J. (2012) Caloric restriction and chronic inflammatory
555 566, doi: 10.1038/s41586 020 2938 9.
diseases, Oral Diseases, 18, 16 31, doi: 10.1111/j.1601
4.
Weiskirchen, R., Weiskirchen, S., and Tacke, F. (2019)
0825.2011.01830.x.
Organ and tissue fibrosis: molecular signals, cellular mech
18.
Stafstrom, C. E., and Rho, J. M. (2012) The ketogenic diet
anisms and translational implications, Mol. Aspects Med.,
as a treatment paradigm for diverse neurological disorders,
65, 2 15, doi: 10.1016/j.mam.2018.06.003.
Front. Pharmacol., 3, 59, doi: 10.3389/fphar.2012.00059.
5.
Martin, P., and Leibovich, S. J. (2005) Inflammatory cells
19.
Zhao, X., Psarianos, P., Ghoraie, L. S., Yip, K.,
during wound repair: the good, the bad and the ugly, Trends
Goldstein, D., et al. (2019) Metabolic regulation of dermal
Cell Biol., 15, 599 607, doi: 10.1016/j.tcb.2005.09.002.
fibroblasts contributes to skin extracellular matrix homeo
6.
Wynn, T. A., and Ramalingam, T. R. (2012) Mechanisms
stasis and fibrosis, Nat. Metab., 1, 147 157, doi: 10.1038/
of fibrosis: therapeutic translation for fibrotic disease, Nat.
s42255 018 0008 5.
Med., 18, 1028 1040, doi: 10.1038/nm.2807.
20.
Liu, G., and Summer, R. (2019) Cellular metabolism in
7.
Zeisberg, M., and Kalluri, R. (2013) Cellular mechanisms
lung health and disease, Annu. Rev. Physiol., 81, 403 428,
of tissue fibrosis. 1. Common and organ specific mecha
doi: 10.1146/annurev physiol 020518 114640.
nisms associated with tissue fibrosis, Am. J. Physiol. Cell
21.
Nigdelioglu, R., Hamanaka, R. B., Meliton, A. Y.,
Physiol., 304, 216 225, doi: 10.1152/ajpcell.00328.2012.
O’Leary, E., Wittet, L. J., et al. (2016) Transforming
8.
Di Carlo, S. E., and Peduto, L. (2018) The perivascular
Growth Factor (TGF) β promotes de novo serine synthesis
origin of pathological fibroblasts, J. Clin. Invest., 128, 54
for collagen production, J. Biol. Chem., 291, 27239 27251,
63, doi: 10.1172/JCI93558.
doi: 10.1074/jbc.M116.756247.
9.
Giannandrea, M., and Parks, W. C. (2014) Diverse func
22.
Park, S. Y., Le, C. T., Sung, K. Y., Choi, D. H., and Cho,
tions of matrix metalloproteinases during fibrosis, Disease
E. H. (2018) Succinate induces hepatic fibrogenesis by
Models Mechanisms, 7, 193203, doi: 10.1242/dmm.
promoting activation, proliferation, and migration, and
012062.
inhibiting apoptosis of hepatic stellate cells, Biochem.
10.
Yue, L., Shi, Y., Su, X., Ouyang, L., Wang, G., et al. (2021)
Biophys. Res. Commun., 496, 673 678, doi: 10.1016/j.bbrc.
Matrix metalloproteinases inhibitors in idiopathic pul
2018.01.106.
monary fibrosis: medicinal chemistry perspectives, Eur. J.
23.
Rabinowitz, J. D., and Mutlu, G. M. (2019) A metabolic
Med. Chem., 224, 113714, doi: 10.1016/j.ejmech.2021.
strategy to reverse fibrosis? Nat. Metab., 1, 1213,
113714.
doi: 10.1038/s42255 018 0013 8.
11.
Lin, J., Huang, Z., Liu, J., Huang, Z., Liu, Y., et al. (2020)
24.
Kang, H. M., Ahn, S.H., Choi, P., Ko, Yi An, Han, S. H.,
Neuroprotective effect of ketone metabolism on inhibiting
et al. (2015) Defective fatty acid oxidation in renal tubular
inflammatory response by regulating macrophage polariza
epithelial cells has a key role in kidney fibrosis develop
tion after acute cervical spinal cord injury in rats, Front.
ment, Nat. Med., 21, 37 46, doi: 10.1038/nm.3762.
Neurosci., 14, 583611, doi: 10.3389/fnins.2020.583611.
25.
Wei, Q., Su, J., Dong, G., Zhang, M., Huo, Y.,et al. (2019)
12.
Andrianova, N. V., Zorova, L. D., Pevzner, I. D., Popkov,
Glycolysis inhibitors suppress renal interstitial fibrosis via
V. A., Chernikov, V. P., et al. (2020) Resemblance and dif
divergent effects on fibroblasts and tubular cells, Am. J.
ferences in dietary restriction nephroprotective mecha
Physiol. Ren. Physiol., 316, F1162 F1172, doi: 10.1152/
nisms in young and old rats, Aging (Albany. NY), 12, 18693
ajprenal.00422.2018.
18715, doi: 10.18632/aging.103960.
26.
Kottmann R. M., Kulkarni,A. A., Smolnycki, K. A., Lyda,
13.
Neal, E. G., Chaffe, H., Schwartz, R. H., Lawson, M. S.,
E., et al. (2012) Lactic acid is elevated in idiopathic pul
Edwards, N., et al. (2008) The ketogenic diet for the treat
monary fibrosis and induces myofibroblast differentiation
ment of childhood epilepsy: a randomised controlled trial,
via pH dependent activation of transforming growth fac
Lancet Neurol.,
7,
500506, doi:
10.1016/S1474
tor β, Am. J. Respir. Crit. Care Med., 186, 740751,
4422(08)70092 9.
doi: 10.1164/rccm.201201 0084OC.
14.
Nielsen, J. V., and Joensson, E. A. (2008) Low carbohy
27.
Faubert, B., Kulkarni, A. A., Smolnycki, K. A., Lyda, E.,
drate diet in type 2 diabetes: stable improvement of body
Dahanayake, T., et al. (2017) Lactate Metabolism in
weight and glycemic control during 44 months follow up,
human lung tumors, Cell, 171, 358 371.e9, doi: 10.1016/
Nutr. Metab., 5, 14, doi: 10.1186/1743 7075 5 14.
j.cell.2017.09.019.
15.
Allen, B. G., Bhatia, S. K., Buatti, J. M., Brandt, K. E.,
28.
Hamanaka, R. B., Nigdelioglu, R., Meliton, A. Y.,
Lindholm, K. E., et al. (2013) Ketogenic diets enhance
Tian, Y., Witt, L. J., et al. (2018) Inhibition of phospho
oxidative stress and radio chemo therapy responses in lung
glycerate dehydrogenase attenuates bleomycin induced
БИОХИМИЯ том 86 вып. 12 2021
БИОЭНЕРГЕТИКА ФИБРОЗА
1873
pulmonary fibrosis, Am. J. Respir. Cell Mol. Biol., 58, 585
Trends Biotechnol., 38, 623 636, doi: 10.1016/j.tibtech.
593, doi: 10.1165/rcmb.2017 0186OC.
2019.12.010.
29.
Meléndez Hevia, E., De Paz Lugo, P., Cornish
44.
Akamatsu, T., Arai, Y., Kosugi, I., Kawasaki, H.,
Bowden, A., and Cárdenas, M. L. (2009) A weak link in
Meguro, S., et al. (2013) Direct isolation of myofibroblasts
metabolism: the metabolic capacity for glycine biosynthe
and fibroblasts from bleomycin injured lungs reveals their
sis does not satisfy the need for collagen synthesis,
functional similarities and differences, Fibrogenes Tissue
J. Biosci., 34, 853 872, doi: 10.1007/s12038 009 0100 9.
Repair, 6, 15, doi: 10.1186/1755 1536 6 15.
30.
Selvarajah, B., Azuelos, I., Platé, M., Guillotin, D.,
45.
Wynn, T. A. (2011) Integrating mechanisms of pulmonary
Fortyet, E. J., et al. (2019) MTORC1 amplifies the ATF4
fibrosis, J. Exp. Med., 208, 1339 1350, doi: 10.1084/jem.
dependent de novo serine glycine pathway to supply
20110551.
glycine during TGF 1 induced collagen biosynthesis, Sci.
46.
Kis, K., Liu, X., and Hagood, J. S. (2011) Myofibroblast
Signal., 12, eaav3048, doi: 10.1126/scisignal.aav3048.
differentiation and survival in fibrotic disease, Expert Rev.
31.
Xie, N., Tan, Z., Banerjee, S., Cui, H., Ge, J., et al. (2015)
Mol. Med., 13, e27, doi: 10.1017/s1462399411001967.
Glycolytic reprogramming in myofibroblast differentiation
47.
Bernard, K., Logsdon, N. J., Ravi, S., Xie, Na, Persons,
and lung fibrosis, Am. J. Respir. Crit. Care Med., 192, 1462
B. P., et al. (2015) Metabolic reprogramming is required
1474, doi: 10.1164/rccm.201504 0780OC.
for myofibroblast contractility and differentiation, J. Biol.
32.
Yu, H., Zhu, J., Chang, L., Liang, C., Li, X., et al. (2021)
Chem.,
290,
25427 25438, doi:
10.1074/jbc.M115.
3 Bromopyruvate decreased kidney fibrosis and fibroblast
646984.
activation by suppressing aerobic glycolysis in unilateral
48.
Bates, J., Vijayakumar, A., Ghoshal, S., Marchand, B.,
ureteral obstruction mice model, Life Sci., 272, 119206,
Yi, S., et al. (2020) Acetyl CoA carboxylase inhibition dis
doi: 10.1016/j.lfs.2021.119206.
rupts metabolic reprogramming during hepatic stellate cell
33.
Kori, Y., Sidoli, S., Yuan, Z. F., Lund, P. J., Zhao, X.,
activation, J. Hepatol., 73, 896 905, doi: 10.1016/j.jhep.
et al. (2017) Proteome wide acetylation dynamics in
2020.04.037.
human cells, Sci. Rep., 7, 10296, doi: 10.1038/s41598
49.
Li, Q., Qin, Z., Nie, F., Bi, H., Zhao, R., et al. (2018)
017 09918 3.
Metabolic reprogramming in keloid fibroblasts: aerobic
34.
Smith, E. R., and Hewitson, T. D. (2020) TGF beta1 is a
glycolysis and a novel therapeutic strategy, Biochem.
regulator of the pyruvate dehydrogenase complex in fibrob
Biophys. Res. Commun., 496, 641 647, doi: 10.1016/j.bbrc.
lasts, Sci. Rep., 10, 17914, doi: 10.1038/s41598 020
2018.01.068.
74919 8.
50.
Bernard, K. (2018) Collagen biosynthesis in pulmonary
35.
Hewitson, T. D., and Smith, E. R. (2021) A metabolic
fibrosis: unraveling the metabolic web, Am. J. Respir. Cell
reprogramming of glycolysis and glutamine metabolism is a
Mol. Biol., 58, 545 546, doi: 10.1165/rcmb.2017 0350ED.
requisite for renal fibrogenesis - why and how? Front.
51.
Yoshida, K., Furuya, S., Osuka, S., Mitoma, J.,
Physiol., 12, 645857, doi: 10.3389/fphys.2021.645857.
Shinoda, Y., et al. (2004) Targeted disruption of the mouse
36.
Bernard, K., Logsdon, N. J., Benavides, G. A.,
3 phosphoglycerate dehydrogenase gene causes severe
Sanders, Y., Zhang, J., et al. (2018) Glutaminolysis is
neurodevelopmental defects and results in embryonic
required for transforming growth factor β1 induced
lethality, J. Biol. Chem., 279, 3573 3577, doi: 10.1074/jbc.
myofibroblast differentiation and activation, J. Biol. Chem.,
C300507200.
293, 1218 1228, doi: 10.1074/jbc.RA117.000444.
52.
Hecker, L., Logsdon, N. J., Kurundkar, D., Kurundkar, A.,
37.
Ge, J., Cui, H., Xie, Na, Banerjee, S., Guo, S., et al.
Bernard, K., et al. (2014) Reversal of persistent fibrosis in
(2018) Glutaminolysis promotes collagen translation and
aging by targeting Nox4 Nrf2 redox imbalance, Sci. Transl.
stability via α ketoglutarate mediated mTOR activation
Med., 6, 231ra47, doi: 10.1126/scitranslmed.3008182.
and proline hydroxylation, Am. J. Respir. Cell Mol. Biol.,
53.
Tsukui, T., Sun, K. H., Wetter, J. B., Wilson Kanamori,
58, 378 390, doi: 10.1165/rcmb.2017 0238OC.
J. R., Hazelwood, L. A., et al. (2020) Collagen producing
38.
Hamanaka, R. B., O’Leary, E. M., Witt, L. J., Tian, Y.,
lung cell atlas identifies multiple subsets with distinct local
Gökalp, G. A., et al. (2019) Glutamine metabolism is
ization and relevance to fibrosis, Nat. Commun., 11, 1920,
required for collagen protein synthesis in lung fibroblasts,
doi: 10.1038/s41467 020 15647 5.
Am. J. Respir. Cell Mol. Biol.,
61,
597606,
54.
Du, K., Hyun, J., Premont, R. T., Choi, S. S., Michelotti,
doi: 10.1165/rcmb.2019 0008OC.
G. A., et al. (2018) Hedgehog YAP signaling pathway reg
39.
Henderson, J., Duffy, L., Stratton, R., Ford, D.,
ulates glutaminolysis to control activation of hepatic stel
O’Reilly, S. (2020) Metabolic reprogramming of glycolysis
late cells, Gastroenterology,
154,
14651479.e1413,
and glutamine metabolism are key events in myofibroblast
doi: 10.1053/j.gastro.2017.12.022.
transition in systemic sclerosis pathogenesis, J. Cell Mol.
55.
Robertson, L. T., and Mitchell, J. R. (2013) Benefits of
Med., 24, 14026 14038, doi: 10.1111/jcmm.16013.
short term dietary restriction in mammals, Exp. Gerontol.,
40.
Fernandez de Cossio Diaz, J., and Vazquez, A. (2017)
48, 1043 1048, doi: 10.1016/j.exger.2013.01.009.
Limits of aerobic metabolism in cancer cells, Sci. Rep., 7,
56.
Spingler, S. R., (2010) Caloric restriction: from soup to
13488, doi: 10.1038/s41598 017 14071 y.
nuts, Ageing Res. Rev., 9, 324 353, doi: 10.1016/j.arr.2009.
41.
Li, P., and Wu, G. (2018) Roles of dietary glycine, proline,
10.003.
and hydroxyproline in collagen synthesis and animal
57.
López Lluch, G., and Navas, P. (2016) Calorie restriction
growth, Amino Acids, 50, 29 38, doi: 10.1007/s00726 017
as an intervention in ageing, J. Physiol., 594, 2043 2060,
2490 6.
doi: 10.1113/JP270543.
42.
Minicis, S., Seki, E., Uchinami, H., Kluwe, J., Zhang, Y.,
58.
Ribarič, S. (2012) Diet and aging, Oxid. Med. Cell. Longev.,
et al. (2007) Gene expression profiles during hepatic stel
2012, 741468, doi: 10.1155/2012/741468.
late cell activation in culture and in vivo, Gastroenterology,
59.
Xu, S., Tao, H., Cao, W., Cao, Li, Lin, Y., et al. (2021)
132, 1937 1946, doi: 10.1053/j.gastro.2007.02.033.
Ketogenic diets inhibit mitochondrial biogenesis and
43.
Sacchi, M., Bansal, R., and Rouwkema, J.
(2020)
induce cardiac fibrosis, Signal Transduct. Target. Ther., 6,
Bioengineered 3D models to recapitulate tissue fibrosis,
54, doi: 10.1038/s41392 020 00411 4.
БИОХИМИЯ том 86 вып. 12 2021
1874
ЯКУПОВА и др.
BIOENERGETICS OF THE FIBROSIS
Review
E. I. Yakupova1,2, D. B. Zorov1,3*, and E. Y. Plotnikov1,3*
1 Belozersky Institute of Physico Chemical Biology, Lomonosov Moscow State University,
119991 Moscow, Russia; E mail: zorov@belozersky.msu.ru; plotnikov@belozersky.msu.ru
2 Institute of Theoretical and Experimental Biophysics, Russian Academy of Sciences,
142290 Pushchino, Moscow Region, Russia
3 Kulakov National Medical Research Center of Obstetrics, Gynecology and Perinatology, 117997 Moscow, Russia
It is known that the development of fibrosis is associated with many diseases, being both a cause and effect of damage
to organs and tissues. Replacement of functional tissue with a scar can lead to organ dysfunction, which is often a life
threatening condition. The development of effective approaches for the prevention or treatment of fibrosis requires an
in depth understanding of all aspects of its pathogenesis, from epithelial mesenchymal transformation to fibroblast
proliferation. Fibrosis can be induced by trauma, ischemic injury, inflammation, and many other conditions accom
panied by repeated cycles of tissue damage and repair. Energetic metabolism is the basis of the functioning of all cells
in the organism and its impairment can be the target for the therapy of such pathological processes as ischemia/reper
fusion, epilepsy, diabetes, cancer, and neurological disorders. The emergence of fibrosis is also associated with the fea
tures of the cell bioenergetics, and in our study, we tried to analyze what changes in the energetic metabolism are
observed during fibrosis development and evaluate the possibility of affecting energetic metabolism as an anti fibrotic
approach.
Keywords: fibrosis, fibroblasts, energy metabolism, glycolysis, respiration, collagen
БИОХИМИЯ том 86 вып. 12 2021