БИОХИМИЯ, 2021, том 86, вып. 2, с. 147 - 157
УДК 576.08/612.115.35/54 39
ИСПОЛЬЗОВАНИЕ КОНЪЮГИРОВАННОГО C ФЛУОРЕСЦЕНТНОЙ
МЕТКОЙ АПТАМЕРА ARC1779 ДЛЯ ОЦЕНКИ ВЛИЯНИЯ Н2О2
НА ЭКЗОЦИТОЗ ФАКТОРА ВИЛЛЕБРАНДА*
© 2021
П.П. Авдонин1, С.К. Труфанов1, А.А. Цитрина1, Е.Ю. Рыбакова1,
Н.В. Гончаров2,3, П.В. Авдонин1**
1 Институт биологии развития им. Н.К. Кольцова РАН, 119334 Москва, Россия;
электронная почта: pvavdonin@yandex.ru
2 Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН, 194223 Санкт Петербург, Россия
3 НИИ гигиены, профпатологии и экологии человека ФМБА России,
188663 пос. Кузьмоловский, Ленинградская обл., Россия
Поступила в редакцию 05.06.2020
После доработки 15.10.2020
Принята к публикации 15.10.2020
Предложен новый подход для количественного определения экспонированного на поверхности эндотели
альных клеток (ЭК) фактора Виллебранда (ФВ) с использованием аптамера ARC1779, взаимодействующе
го с доменом А1 в структуре ФВ. Для визуализации образующегося комплекса ФВ с аптамером к молекуле
последнего была присоединена флуоресцентная метка Cy5. Культивируемые ЭК из пупочной вены челове
ка прижизненно окрашивали конъюгатом ARC1779 Cy5, и снимали изображение с помощью флуоресцент
ного микроскопа. Анализ изображения проводили с помощью программы CellProfiler. Мультимерные
комплексы ФВ, секретируемые тельцами Вейбеля-Паладе, при окраске конъюгатом ARC1779 Cy5 видны
под микроскопом как яркие точечные структуры округлой и неправильной формы. Число этих структур
многократно увеличивается при воздействии на ЭК активаторов секреции ФВ, гистамина и тромбина. Кро
ме того, ARC1779 Cy5 выявляет на поверхности активированных ЭК гигантские мультимеры ФВ, развёрну
тые в виде длинных нитей. Секрецию ФВ в ЭК активируют вторичные мессенджеры - cAMP и Са2+. Есть
свидетельства, что пероксид водорода в ЭК также выполняет функцию вторичного мессенджера. Кроме то
го, в ЭК поступает экзогенный Н2О2, образующийся в лейкоцитах. Задачей работы было, используя пред
ложенный метод, определить, какое влияние Н2О2 оказывает на экспрессию ФВ на поверхности ЭК. Пока
зано, что пероксид водорода при концентрации 100 мкМ (ниже цитотоксической) в несколько раз увеличи
вает количество экспонированных на плазматической мембране точечных структур ФВ. Полученные дан
ные свидетельствуют о роли Н2О2 в качестве посредника, активирующего экзоцитоз телец Вейбеля-Паладе
и секрецию ФВ в эндотелии сосудов при воспалении и при индукции образования эндогенных активных
форм кислорода в ЭК.
КЛЮЧЕВЫЕ СЛОВА: фактор Виллебранда, аптамеры, пероксид водорода, эндотелиальные клетки,
секреция.
DOI: 10.31857/S0320972521020019
ВВЕДЕНИЕ
как правило, действуют на несколько мишеней
в клетках. С появлением методов генной инже
Более половины всех фармакологических
нерии и молекулярной иммунологии стало ин
препаратов являются лигандами мембранных
тенсивно развиваться новое направление в фар
рецепторов и других белков [1, 2]. Это в абсо
макологии - создание исключительно специ
лютном большинстве низкомолекулярные ве
фичных лекарственных препаратов на основе
щества природного происхождения или искус
гуманизированных антител. Разработаны анти
ственные синтетические соединения. Почти все
тела против рецепторов, компонентов системы
они не обладают полной избирательностью и,
комплемента и т.д. Присоединение к антителам
маркеров и токсических компонентов позволи
Принятые сокращения: ФВ - фактор Виллебранда;
ло использовать их одновременно и как диаг
ЭК - эндотелиальные клетки.
ностический препарат, и как лекарство. Данный
* Статья на английском языке опубликована в режиме
подход является основой для нового направле
Open Access (открытого доступа) на сайте издательства
ния в биомедицинских исследованиях - тера
вып. 2, 2021.
ностики. Антитело используется как «якорь», к
** Адресат для корреспонденции.
которому присоединяется конструкция, несу
147
148
АВДОНИН и др.
щая метку для визуализации патологического
лечении болезни Виллебранда типа 2В [15-17].
очага, и компонент, осуществляющий терапев
Метод SELEX усовершенствован благодаря ис
тическое воздействие [3].
пользованию при получении библиотек олиго
В 1990 г. был предложен метод SELEX
нуклеотидов со случайными (random) последо
(Systematic Evolution of Ligands by Exponential
вательностями искусственного пятого нуклео
Enrichment) для создания аптамеров - коротких
тида (ExSELEX), и таким образом был синтези
одноцепочечных олигонуклеотидов ДНК или
рован новый ещё более эффективный аптамер
РНК с уникальными последовательностями,
против ФВ [18].
способных с высоким сродством и специфич
Помимо применения в качестве фармпрепа
ностью связываться с мишенями разного типа:
ратов, низкомолекулярные лиганды и антитела
белками, полисахаридами, локализованными на
широко используются для проведения биологи
клеточной поверхности, с низкомолекулярны
ческих исследований мембранных рецепторов и
ми соединениями, ионами металлов и други
внутриклеточных белков. Метод иммунофлуо
ми [4-7]. По аффинности и избирательности
ресцентного окрашивания антителами позволя
действия аптамеры не уступают антителам. Раз
ет оценивать локализацию и уровень экспрес
работан подход для селекции аптамеров против
сии белков на плазматической мембране и внут
отдельных видов клеток, включая раковые [8].
ри клетки. Один из вопросов, требующих более
Проводятся клинические испытания аптамеров,
глубокого изучения, касается механизмов регу
предназначенных для лечения онкологических
ляции секреции ФВ эндотелиальными клетка
заболеваний и болезней крови [9, 10]. Аптамеры
ми. Антитела против ФВ позволяют оценивать
имеют ряд преимуществ по сравнению с антите
его экспонирование на поверхности плазмати
лами. Их структура воспроизводима, они тер
ческой мембраны. При этом проводится инку
мостабильны, менее иммуногенны и могут хра
бация фиксированных ЭК с первичными анти
ниться в сухом виде неограниченное время [10].
телами и после этого с флуоресцентно окрашен
Показана возможность использования аптаме
ными вторичными антителами [19]. В настоя
ров вместо антител в гистохимических исследо
щей работе нами впервые был применен подход
ваниях [11] и для проточной цитофлуоримет
с определением экзоцитоза ФВ с помощью флу
рии [12].
оресцентно меченного аптамера. С этой целью
Одной из мишеней для антител и аптамеров
был использован аптамер ARC1779 с присоеди
является фактор Виллебранда (ФВ). Это важ
ненным к его молекуле флуоресцентным краси
нейший гликопротеин плазмы крови, секрети
телем Cy5. В настоящее время разработано не
руемый эндотелиальными клетками (ЭК) (в
сколько аптамеров, присоединяющихся к ФВ
меньшей степени тромбоцитами) и обеспечива
по домену А1 и блокирующих его взаимодей
ющий поддержание гемостаза. ФВ поступает в
ствие с тромбоцитами [17, 18, 20, 21], однако по
кровь из ЭК в виде популяции гигантских ли
ка только ARC1779 получил применение в каче
нейных мультимерных молекул молекулярной
стве лекарственного препарата. Используя
массой до 20 000 кДа, которые образованы из
конъюгат ARC1779 Cy5, нам удалось наблюдать
мономеров ~250 кДа, соединённых дисульфид
за раскрытием на поверхности ЭК отдельных
ными связями [13]. В молекуле ФВ имеются ре
секреторных везикул и исследовать регуляцию
гулярно повторяющиеся структурные доме
этого процесса непосредственно в живых клет
ны А1, обеспечивающие его взаимодействие с
ках. Мы показали, что пероксид водорода в кон
гликопротеином 1b на мембране тромбоцитов.
центрациях, в которых он может накапливаться
Присоединение тромбоцитов к развернутым
в сосудах, стимулирует экзоцитоз телец Вейбе
молекулам ФВ по этому домену инициирует об
ля-Паладе и экспонирование ФВ на плазмати
разование тромбов в артериолах и капиллярах
ческой мембране ЭК.
при их повреждении и остановку кровотечения.
Избыточное образование ФВ или его повышен
ная активность, вызванная мутациями, приво
МАТЕРИАЛЫ И МЕТОДЫ
дит к патологическому тромбообразованию в
микрососудах и развитию тромботических мик
ЭК выделяли из пупочной вены человека по
роангиопатий. Создан фармакологический пре
методике, описанной ранее [22], и культивиро
парат «Caplacizumab» на основе наноантитела
вали в среде М199, содержащей 20% сыворотки
против домена А1 в молекуле ФВ [14], применя
эмбрионов крупного рогатого скота («Sigma
емый для лечения тромботической тромбоцито
Aldrich», США) и эндотелиальную ростовую до
пенической пурпуры. Параллельно с этим раз
бавку, полученную из замороженных мозгов
работан также связывающийся с А1 апта
кролика (ООО «Кролинфо», д. Новая, Московс
мер ARC1779, показавший эффективность при
кая обл., Россия) по методу Maciag et al. [23].
БИОХИМИЯ том 86 вып. 2 2021
КОНЪЮГАТ ARC1779 Cy5 ДЛЯ ОПРЕДЕЛЕНИЯ ФАКТОРА ВИЛЛЕБРАНДА
149
Клетки идентифицировали по морфологичес
щей интенсивности флуоресценции и средних
ким критериям: наличию ФВ, ангиотензин
значений числа частиц ФВ в пересчете на число
превращающего фермента и поверхностных
окрашенных ядер проводили после обработки
белков CD31, CD54, CD61 [24]. ЭК культивиро
мозаичных изображений с участков площадью
вали в атмосфере, содержавшей 5% СО2. В рабо
1,5 × 2 мм в шести контрольных и шести опытных
те использовали клетки 2-4 пассажей. Для сня
лунках. Число клеток на участке данной площа
тия клеток при пассировании применяли реа
ди составляло 2800-3000. Изображения анали
гент аккутазу («Sigma Aldrich», США).
зировались с помощью программы CellProfiler
Для окрашивания ФВ использовали апта
[25], находящейся в открытом доступе
мер ARC1779 (патент US20090203766A1,
«Archemix Corp.», США) с присоединённым к
описание применения этой программы для ана
нему по 5′ концу красителем Cy5 (Cy5 mGm
лиза изображения ЭК приведено ранее [26]. Об
CmGmUdGdCdAmGmUmGmCmCmUmUmCm
щее количество анализируемых кадров состав
GmGmCdCmG s dTmGdCdGdGdTmGmCd
ляло 96-128 (по 16 кадров из каждой лунки в 48
CmUdCdCmGmUdCmAmCmGmC, где «m» -
луночном планшете). Для получения приведен
2′ OMe, «s» - фосфоротиоатная связь). В каче
ных в статье данных проводилось не менее трех
стве контроля был взят олигонуклеотид Cy5
независимых экспериментов на разных препа
mGmGmCmCdAdGdCmCmUmCmUmCmCm
ратах ЭК. Проводилось определение общего ко
UmGmGmUmGdGmG s TmCdGdCdCTmAm
личества экспонированных на поверхности ЭК
GdGmUdCdGmGmGdCmCmCmCmU со слу
окрашенных ARC1779 Cy5 структур ФВ и их
чайной последовательностью, но с таким же со
средней яркости (интенсивность флуоресцен
отношением нуклеотидов, как у ARC1779. Оли
ции в пересчете на отдельную клетку). Данные
гонуклеотиды синтезировали в компании
представлены в относительных единицах. При
«ДНК синтез», Россия.
ведены средние значения ± стандартная ошибка
При определении действия гистамина
среднего. Достоверность отличий определяли с
(100 мкМ), тромбина (1 ед./мл) или пероксида
использованием критерия Стьюдента с по
водорода (100 мкМ) на экзоцитоз ФВ выращен
мощью программы Excel, а также однофактор
ные в 24 луночном планшете ЭК отмывали от
ного дисперсионного анализа с помощью прог
среды роста физиологическим солевым раство
раммы MedCalc («MedCalc Software Ltd», Бель
ром (PSS, pH 7,4), содержавшим 145 мМ NaCl,
гия).
5
мМ KCl, 10 мМ HEPES, 1 мМ MgCl2,
Для проведения иммунофлуоресцентного
1 мМ СaCl2 и 10 мМ глюкозы. Клетки инкубиро
окрашивания клетки фиксировались 4% ным
вали в отсутствии или в присутствии активатора
параформальдегидом в PBS в течение 20 мин на
при 32-34 °С в течение 20 мин. После этого
льду, затем их аккуратно отмывали раствором,
внеклеточную жидкость отбирали, и добавляли
содержащим 1% BSA. В этот же раствор вносили
в каждую лунку по 100 мкл раствора 250 нМ
первичные антитела против фактора Виллеб
ARC1779 Cy5 в среде гибридизации, содержа
ранда (Rabbit anti human vWF, «Dako», Фран
щей 0,1 мг/мл tRNA («RocheDiagnostics», Швей
ция, разведение 1/500 (исходная концентрация
цария), 1 мг/мл бычьего сывороточного альбу
антител 3,2 мг/мл)) и инкубировали ночь при
мина (BSA, «Amresco», США), 5 мМ MgCl2,
4 °С. После инкубации клетки отмывали и вно
25 мМ глюкозы, 2,67 мМ KCl, 1,47 мМ KH2PO4,
сили смесь из вторичных антител (goat anti rab
136,9 мМ NaCl, 8,10 мМ Na2HPO4, после чего
bit AlexaFluor 488 («Invitrogen», США), разведе
инкубировали 20 мин при 32 °С. Для окрашива
ние 1/1000), флуоресцентно меченного лектина
ния ядер в среду добавляли 1 мкг/мл Hoechst 33342
WGA (weat germ agglutinin AlexaFluor
488
(«ThermoFisher Scientific», США). По окон
(«Invitrogen», США), разведение 1/500) и ядер
чании инкубации клетки 2 раза промывали
ного красителя Hoechst 33258 («Sigma Aldrich»,
раствором фосфатно солевого буфера (PBS)
США) в концентрации 1 мкг/мл и далее инку
(«ThermoFisher Scientific», США) для удаления
бировали 1,5 ч при комнатной температуре. По
не связавшегося ARC1779 Cy5. К клеткам до
окончании инкубации клетки отмывали раство
бавляли
200
мкл FluoroBriteTM DMEM
ром PBS, содержащим 1% BSA, и помещали в
(«ThermoFisher Scientific», США), и сразу после
насыщенный раствор фруктозы. Готовые препа
этого снимали изображение окрашенных клеток
раты анализировали с помощью флуоресцент
с помощью флуоресцентного микроскопа
ного микроскопа Leica DMI 6000 («Leica», Гер
Leica DMI 6000 с использованием объектива
мания). Параметры захвата изображения на
HCX PL FLUOTAR L 20,0 × 0,40 DRY, диодных
страивались по самому яркому образцу и не меня
осветителей с длинами волн 385 и 620 нм и
лись в течение всего эксперимента. С централь
фильтров A4 и TX2 («Leica», ФРГ). Расчёт об
ного участка каждой лунки снимался мозаич
БИОХИМИЯ том 86 вып. 2 2021
150
АВДОНИН и др.
ный кадр величиной в 25 микроскопических по
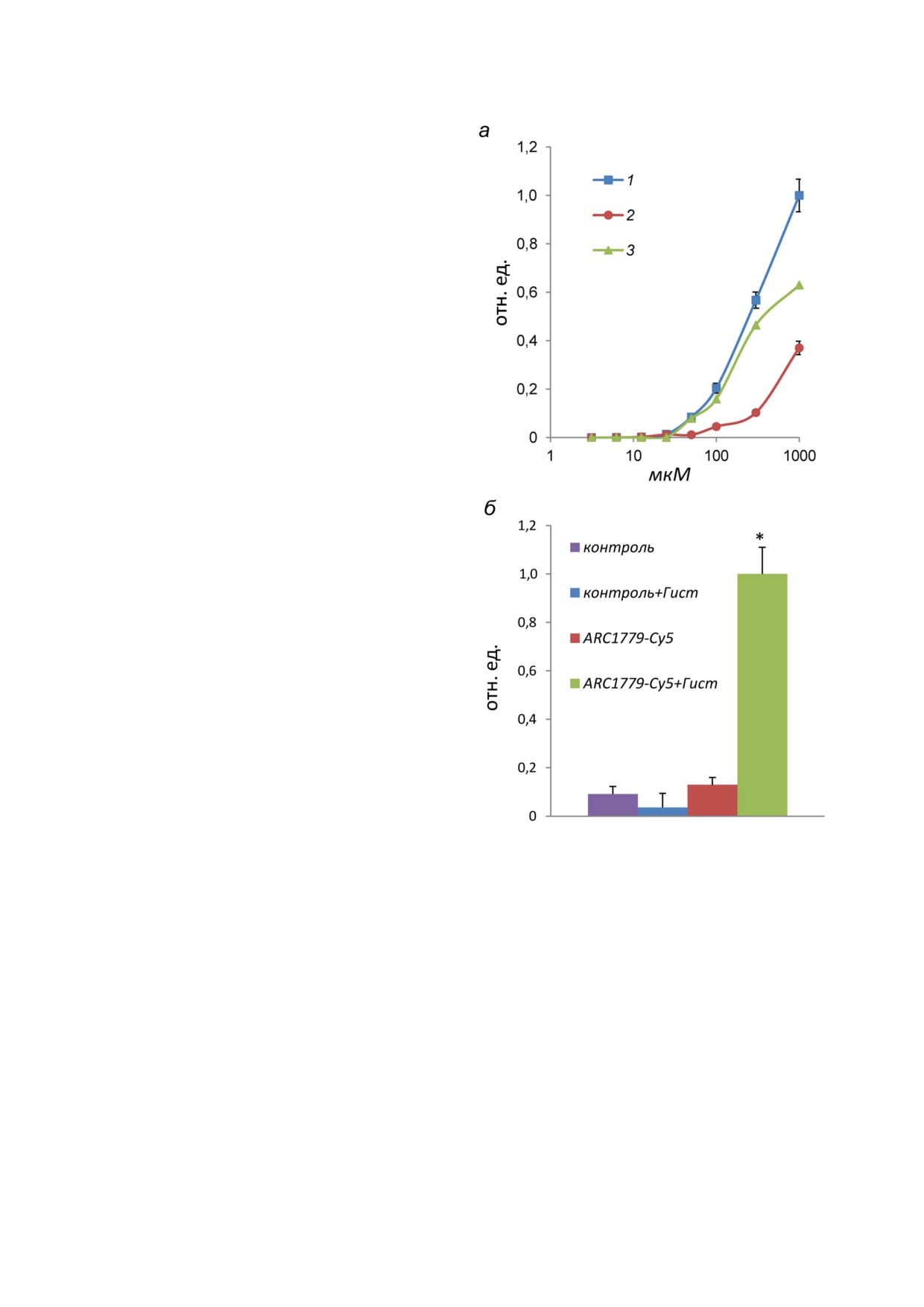
концентрациях от 3,1 нМ до 1 мкМ. С помощью
лей и имеющий физические размеры 1,5 × 2 мм.
программы CellProfiler рассчитывали суммар
Съемка велась с использованием высокоскорост
ную яркость окрашенных структур ФВ (objects)
ного адаптивного автофокуса. С каждого образ
на поверхности клеток. Видно, что окрашива
ца получали 25 изображений (по 16 кадров в
ние аптамером ARC1779 Cy5 ФВ на мембране
изображении), содержащих 3 флуоресцентных
ЭК становится заметным, начиная с концентра
канала с кодировкой глубины цвета 8 бит. С по
ции 50 нМ (рис. 2, а, кривая 1). Яркость окра
мощью программы CellProfiler определяли ин
шивания возрастает при повышении концент
тенсивность флуоресценции окрашенного ФВ
рации ARC1779 Cy5 до 1 мкМ. Неспецифичес
на каждом кадре в ЭК в контроле и после акти
кое связывание метки определяли при соответ
вации их Н2О2 или тромбином.
ствующих концентрациях конъюгата контроль
ного олигонуклеотида с Cy5. Конъюгированный
с меткой контрольный олигонуклеотид почти не
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
окрашивает клетки при концентрациях до
100 нМ. При дальнейшем повышении его кон
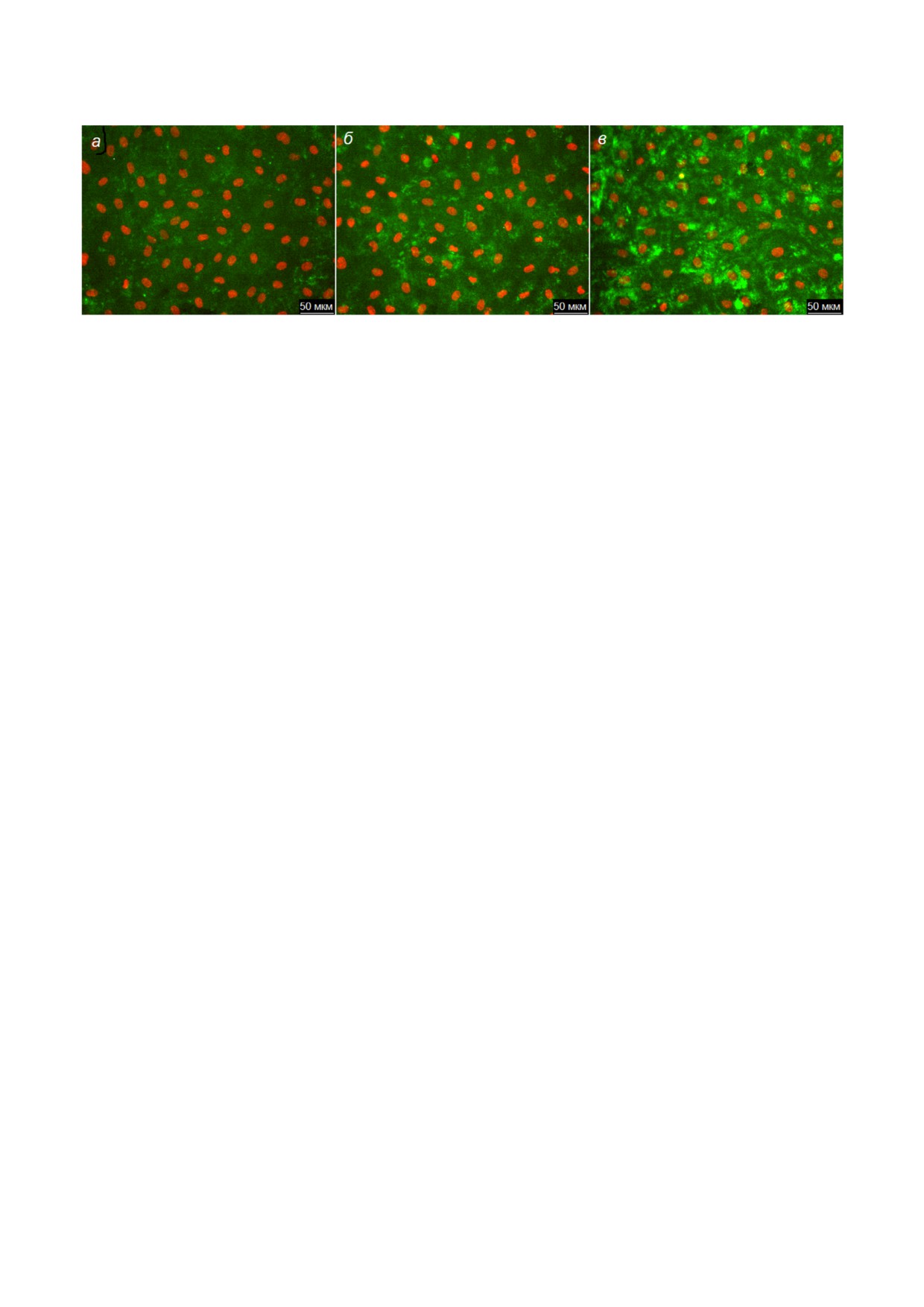
На рис. 1, а показаны эндотелиальные клет
центрации интенсивность флуоресценции на
ки, окрашенные ARC1779 Cy5 (50 нМ). Перед
чинает возрастать из за увеличения неспецифи
добавлением аптамера клетки инкубировали в
ческого связывания (рис. 2, а, кривая 2). Изоб
течение 20 мин с индуктором секреции ФВ -
раженная на рис. 2, а кривая 3 представляет со
гистамином. Для выявления клеточных ядер ис
бой разность между интенсивностями флуорес
пользовали Hoechst 33342. ARC1779 Cy5 окра
ценции ARC1779 Cy5 и контрольного олиго
шивает небольшие структуры, имеющие в ос
нуклеотида и отражает специфическое взаимо
новном округлую или неправильную форму. Для
действие ARC1779 Cy5 с ФВ. ЕС50 для связыва
оценки специфичности окрашивания ФВ апта
ния ARC1779 Cy5 с ФВ составило 287 нМ (95%
мером ARC1779 Cy5 в качестве отрицательного
ный доверительный интервал 155-589 нМ). В
контроля был использован конъюгированный с
экспериментах мы использовали концентрацию
Cy5 олигонуклеотид такой же длины и с таким
ARC1779 Cy5 50 нМ, при которой неспецифи
же соотношением нуклеотидов как у ARC1779,
ческое связывание метки невелико. Относи
но со случайной нуклеотидной последователь
тельные значения яркости флуоресценции не
ностью (рис. 1, б). Контрольный олигонуклео
активированных и активированных гистамином
тид практически не связывается с живыми клет
клеток после окраски их аптамером ARC1779
ками. Одиночные крупные яркие пятна, кото
Cy5 или контрольным олигонуклеотидом в кон
рые видны на рис. 1, б, представляют собой ок
центрации 50 нМ показаны на рис. 2, б. Видно,
рашенные безъядерные фрагменты мертвых
что гистамин не влияет на неспецифическое
клеток.
связывание флуоресцентной метки с ЭК. В от
Зависимость связывания ARC1779 Cy5 с
сутствии гистамина флуоресценция лишь не
клетками от концентрации аптамера показана
значительно превышает фоновое значение, что
на рис. 2. ЭК инкубировали с ARC1779 Cy5 в
говорит о низком уровне экспрессии ФВ на
Рис. 1. Окрашивание аптамером ARC1779 Cy5 (а) и конъюгированным с флуоресцентной меткой Cy5 контрольным оли
гонуклеотидом (б) эндотелиальных клеток, активированных гистамином (100 мкМ). Концентрации ARC1779 Cy5 и конт
рольного олигонуклеотида - 50 нМ. (С цветными вариантами рис. 1-5 можно ознакомиться в электронной версии статьи
БИОХИМИЯ том 86 вып. 2 2021
КОНЪЮГАТ ARC1779 Cy5 ДЛЯ ОПРЕДЕЛЕНИЯ ФАКТОРА ВИЛЛЕБРАНДА
151
плазматической мембране ЭК в отсутствии ак
тиваторов.
Второй задачей работы было изучение регу
ляции экзоцитоза ФВ с помощью вышеописан
ного экспериментального подхода. Экзоцитоз
ФВ в ЭК стимулируют нейроэндокринные фак
торы, включая тромбин [27], вазопрессин [28],
гистамин [29], серотонин [30], адреналин [31],
пуриновые нуклеотиды [32], цитокины и фак
торы роста [33, 34]. В реализации их действия
участвуют вторичные посредники - cAMP и
Са2+ [35]. Известно, что пероксид водорода так
же выполняет в ЭК функцию вторичного мес
сенджера [36]. Кроме того, на функционирова
ние ЭК оказывает влияние экзогенный Н2О2,
образующийся в нейтрофилах. Ранее нами бы
ло показано, что добавленный извне Н2О2 про
никает в эндотелиальные клетки [26]. Извест
ным механизмом действия Н2О2 как вторично
го мессенджера является ингибирование тиро
зиновых протеинфосфатаз [37]. В ЭК Н2О2 из
бирательно усиливает действие агонистов 5
НТ1В и 5 НТ2В рецепторов [38] и стимулиру
ет двупоровые эндолизосомальные кальциевые
каналы [39]. Данные о влиянии Н2О2 на секре
цию ФВ весьма противоречивы. Ранее было по
казано, что экзогенный супероксид анион уве
личивает секрецию ФВ во внешнюю среду
культивируемыми ЭК, однако достоверного
эффекта Н2О2, добавленного в концентрациях
вплоть до 0,5 мМ, выявлено не было [40]. Нап
ротив, в работе Yang et al. [41] приведены дан
ные об увеличении секреции ФВ в 1,5-2 раза
под влиянием 0,5 мМ Н2О2. В любом случае в
этих исследованиях было установлено действие
цитотоксической концентрации Н2О2. Ранее
методами иммуноцитофлуориметрии и имму
ноферментного анализа нами было показано,
что пероксид водорода в концентрации
100 мкМ (что ниже порога цитотоксичности
Рис. 2. а - Зависимости яркости флуоресценции ЭК от
для культивируемых ЭК из пупочной вены) вы
концентрации ARC1779 Cy5 (кривая 1) или конъюгата
зывает увеличение количества экспонирован
контрольного олигонуклеотида с Cy5 (кривая 2). Кривая 3
ного на поверхности ЭК ФВ и его секрецию во
представляет собой разность между значениями на кри
внеклеточную среду [26]. В данной работе мы
вой 1 (общее связывание) и кривой 2 (неспецифическое
связывание) и отражает взаимодействие ARC1779 Cy5 с
продолжили эти исследования, применив но
ФВ. б
- Яркость флуоресценции ЭК, окрашенных
вый подход с использованием аптамера
ARC1779 Cy5 (ARC1779 Cy5, ARC1779 Cy5 + Гист.) или
ARC1779, к молекуле которого присоединен
контрольным олигонуклеотидом (контроль, конт
флуоресцентный краситель Cy5. В отличие от
роль + Гист.) в концентрациях 50 нМ в отсутствии или в
присутствии 100 мкМ гистамина (Гист.). *р < 0,01 - Досто
иммунофлуоресцентной метки, в случае конъ
верность отличия ARC1779 Cy5 + Гист. от ARC1779 Cy5 и
югата ARC1779 Cy5 окрашивание ЭК проводи
от контролей
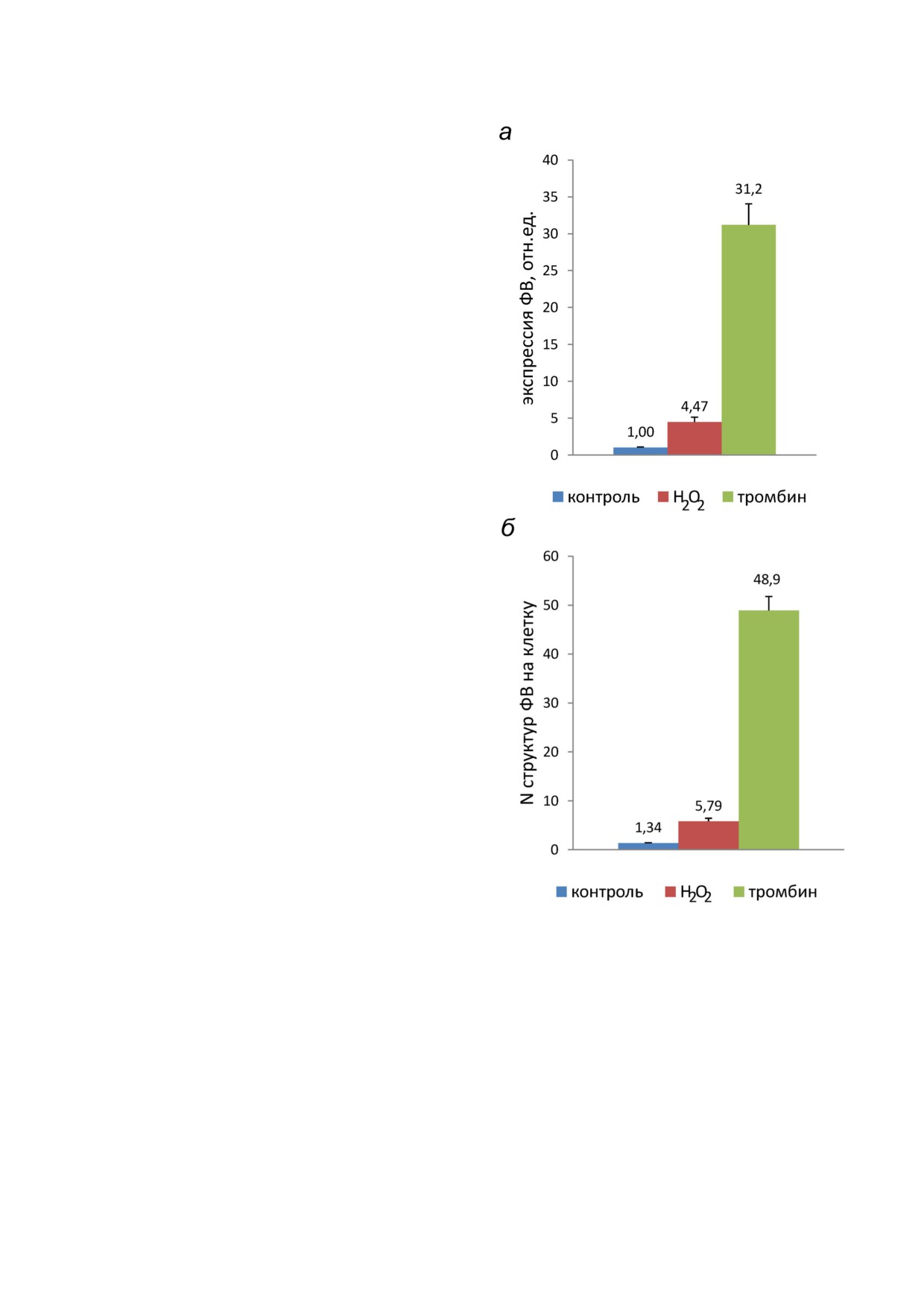
лось на живых клетках. Видно, что после воз
действия 100 мкМ Н2О2 в течение 20 мин коли
чество окрашенных ARC1779 Cy5 клеток резко
ФВ в пересчете на одну клетку при действии
возросло (рис. 3, б) по сравнению с контролем
Н2О2 составило 5,79, что в 4,3 раза превышает
(рис. 3, а). Средняя интенсивность флуоресцен
число структур ФВ в контроле (рис. 4, б). Сте
ции (рис. 4, а) выросла в 4,47 раза (p < 0,01).
пень прироста ФВ позитивных структур прак
Среднее число окрашенных точечных структур
тически совпадает с показателем увеличения
БИОХИМИЯ том 86 вып. 2 2021
152
АВДОНИН и др.
Рис. 3. Влияние Н2О2 и тромбина на экзоцитоз ФВ в ЭК. Показаны ядра клеток и окрашенный с помощью ARC1779 Cy5
ФВ в ЭК в контроле (а), после воздействия 100 мкМ Н2О2 (б) или 1 ед./мл тромбина (в). Концентрация ARC1779 Cy5 -
50 нМ
флуоресценции под влиянием Н2О2. Из этого
вышает экспрессию ФВ за счёт увеличения ко
следует, что увеличение экспонирования ФВ на
личества раскрытых секреторных везикул. По
поверхности ЭК под действием Н2О2 происхо
мимо точечных структур можно видеть одиноч
дит главным образом в результате увеличения
ные окрашенные нити длиной в десятки мик
количества раскрытых секреторных везикул -
рон, которые образуются при слиянии в виде
телец Вейбеля-Паладе. Ранее в работах иссле
спирали сверхкрупных мультимеров ФВ
дователей из Нидерландов [19, 42] с помощью
(рис. 3, в).
корреляционного иммунофлуоресцентного и
Иммуноцитофлуоресцентный метод являет
электронно микроскопического анализа изобра
ся стандартным подходом для определения
жения было продемонстрировано, что во время
экспрессии ФВ на поверхности ЭК. В данном
секреции ФВ происходит слияние телец Вейбе
исследовании для окрашивания ФВ вместо ан
ля-Паладе с образованием более крупных сек
титела был использован конъюгированный с
реторных капсул, которые раскрываются на
флуоресцентной меткой Cy5 аптамер ARC1779.
плазматической мембране с образованием пор
Принципиальным отличием предложенного
диаметром 0,5-5 мкм. Размеры выявляемых на
подхода от иммунофлуоресцентного метода яв
ми точечных структур ФВ лежат в рамках этого
ляется окрашивание живых, а не фиксирован
диапазона. Их средние максимальный и мини
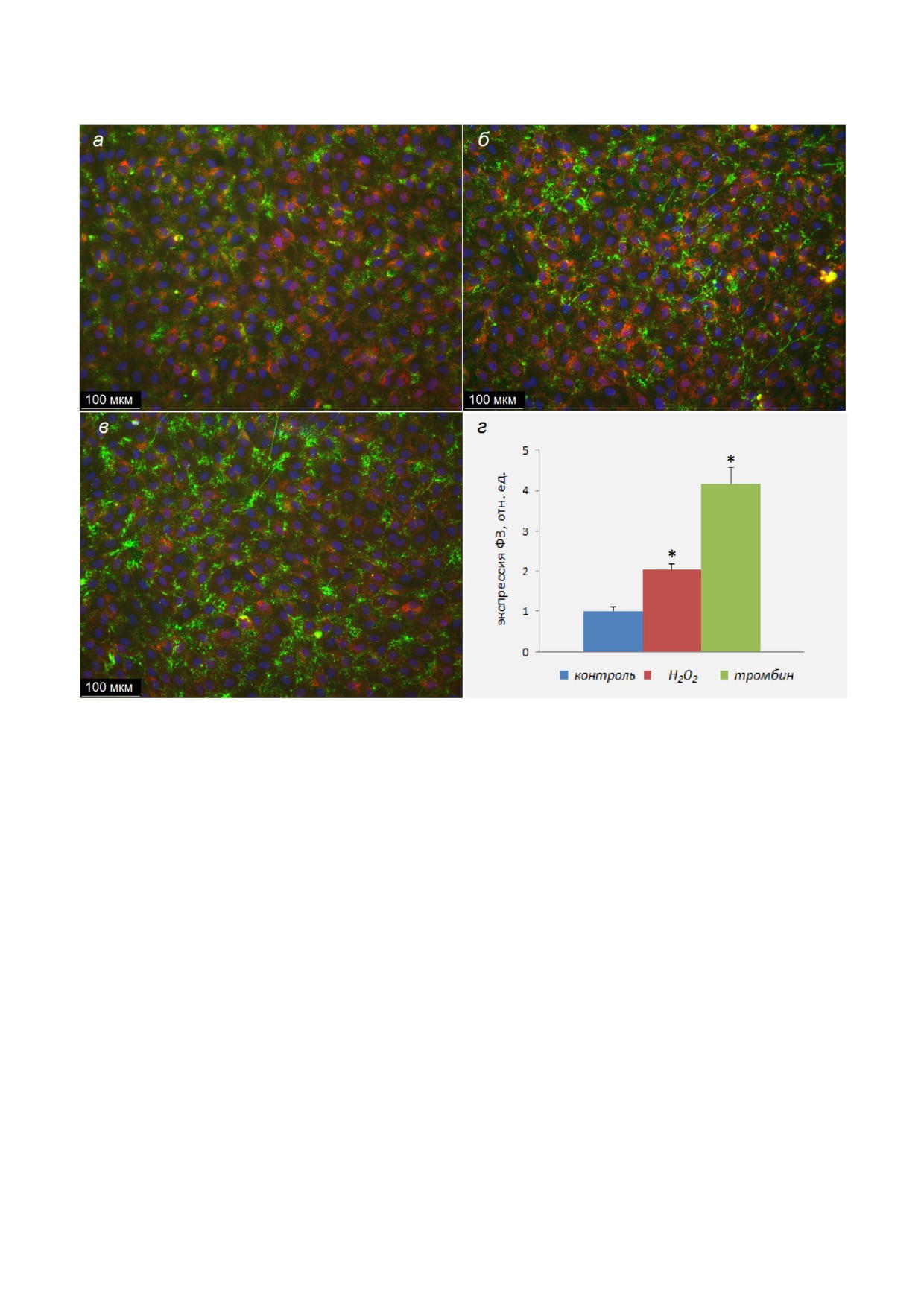
ных клеток. Мы провели сравнение данного ме
мальный диаметры Ферета составляют в конт
тода с иммунофлуоресцентным. На рис. 5 пока
роле 0,97 и 0,45 мкм соответственно, а с Н2О2 -
заны ЭК, окрашенные антителами против ФВ, в
1,02 и 0,48 мкм соответственно.
контроле и после воздействия Н2О2 или тромби
При определении эффекта Н2О2 методом
на. Был использован тот же препарат ЭК, как и
иммуноцитофлуориметрии мы регистрировали
в опыте с ARC1779 Cy5 (рис. 3), и такие же усло
не более чем 2× увеличение количества ФВ, экс
вия инкубации. Конечная концентрация пер
понированного на поверхности клеток [26].
вичных антител против ФВ составляла
Причиной более выраженного эффекта в случае
40 нМ (разведение 1/500 исходного препарата
использования ARC1779 Cy5, как мы предпола
IgG c концентрацией 3,2 мг/мл или 20 мкМ).
гаем, является исключение стадии фиксации,
Полученные изображения (рис. 5, а-в) соответ
при которой возможно дополнительное арте
ствуют тем, которые были получены с помощью
фактное слияние секреторных пузырьков с
ARC1779 Cy5 (рис. 3). Четкость изображения
мембраной. При сравнении данных, приведён
при использовании антител выше, т.к. вторич
ных на рис. 2, б и 4, а, видно, что эффект Н2О2 на
ные антитела дают более яркое окрашивание от
экспрессию ФВ сопоставим с эффектом
носительно фоновой флуоресценции, чем
100 мкМ гистамина. В качестве положительного
ARC1779 Cy5. Однако флуоресценция окра
контроля мы использовали также тромбин.
шенного ФВ при этом возрастала под действием
Тромбин оказывает гораздо более мощный эф
Н2О2 в 2 раза и тромбина - в 4 раза по отноше
фект - экспрессия ФВ на плазматической
нию к контролю (рис. 5, в), в отличие от 4,5× и
мембране ЭК возрастает в 31,2 раза, а число экс
31× увеличения в эксперименте с использовани
понированных структур ФВ в пересчете на ко
ем ARC1779 Cy5 (рис. 4, а). В случае гистамина
личество ядер (т.е. на отдельную клетку) увели
в опытах с окраской клеток антителами был бо
чивается в 36,5 раз. Близость этих значений ука
лее слабый эффект, чем при окраске ФВ аптаме
зывает, что, как и в случае с Н2О2, тромбин по
рами (данные не приводятся). Как уже отмечено
БИОХИМИЯ том 86 вып. 2 2021
КОНЪЮГАТ ARC1779 Cy5 ДЛЯ ОПРЕДЕЛЕНИЯ ФАКТОРА ВИЛЛЕБРАНДА
153
выше, мы предполагаем, что менее выраженный
эффект агонистов на экспрессию ФВ на поверх
ности ЭК при иммунофлуоресцентном окраши
вании объясняется тем, что во время фиксации
происходит дополнительный артефактный эк
зоцитоз секреторных гранул. Таким образом,
применение ARC1779 Cy5 позволяет исследо
вать более приближенные к реальности процес
сы регуляции экзоцитоза ФВ по сравнению с
антителами, несмотря на то что получаемое с
ARC1779 Cy5 изображение по контрастности
уступает иммунофлуоресцентному окрашива
нию.
Мы предполагаем, что можно увеличить от
ношение специфического окрашивания ФВ к
фоновому окрашиванию, если использовать ап
тамеры, которые были разработаны уже после
ARC1779 и проходят сейчас клинические испы
тания, как возможные препараты для лечения
тромботической микроангиопатии. Есть дан
ные, что аптамер TAGX 0004, в нуклеотидной
последовательности которого имеется одно ис
кусственное основание, более эффективно по
давляет агрегацию тромбоцитов по сравнению с
ARC1779 [17]. Пегилированный аптамер BT200
подавляет активность ФВ в плазме с IC50 70 нМ
[43]. Данные об эффективности этих двух апта
меров получены в тесте агрегации тромбоцитов.
Тромбоциты также секретируют ФВ и связыва
ют его на своей поверхности, поэтому конъюгат
ARC1779 Cy5 может быть использован для
оценки их активации методом проточной ци
тофлуориметрии.
С помощью ARC1779 Cy5 нам удалось вы
явить хорошо выраженную активацию экзоци
тоза ФВ под действием Н2О2. В условиях in vivo
пероксид водорода образуется эндогенно в
ЭК [36], а также выделяется нейтрофилами, ад
гезированными на поверхности активированно
го эндотелия [44]. Согласно расчетам, приве
денным в работе Jones [45], скорость образова
ния Н2О2 в организме может достигать величи
ны 500 мкмоль/кг/мин. В результате измерений
концентрации Н2О2 в стационарных условиях в
Рис. 4. Определение с помощью ARC1779 Cy5 экспрессии
цельной крови и в плазме крови человека и кры
ФВ на поверхности ЭК (а) и среднего количества точечных
сы, проведённых в разных лабораториях, полу
структур ФВ в отдельной клетке (б) в контроле при воздей
чены данные в широком диапазоне концентра
ствии 100 мкМ Н2О2 или 1 ед./мл тромбина
ций - от единиц до десятков микромолей на
литр плазмы и до сотен микромолей на литр
крови [46, 47], последнее - за счет высокой кон
рации до 150 мкМ не оказывает на них заметно
центрации Н2О2 в эритроцитах и особенно в
го токсического действия [48, 49]. Таким обра
нейтрофилах, даже с учетом разницы в количе
зом, используемая в настоящих экспериментах
стве этих клеток. Локальная концентрация Н2О2
концентрация 100 мкМ не является токсичной
вблизи эндотелия может быть значительно вы
для ЭК в культуре и соответствует физиологи
ше средней концентрации в плазме крови. На
ческим условиям. Ранее показано, что в концен
препарате культивируемых ЭК из пупочной ве
трациях 50-100 мкМ Н2О2 вызывает в ЭК по
ны показано, что воздействие Н2О2 в концент
верхностную экспрессию главного комплекса
БИОХИМИЯ том 86 вып. 2 2021
154
АВДОНИН и др.
Рис. 5. Окрашивание с помощью антител ФВ на поверхности ЭК в контроле (а), при воздействии 100 мкМ Н2О2 (б) или
1 ед./мл тромбина (в). Уровень экспрессии ФВ на поверхности ЭК, определенный иммунофлуоресцентным методом, в
контроле и при активации ЭК Н2О2 или тромбином (г). Достоверность отличия от контроля, *p < 0,01. Для выявления гра
ниц клеток проводилась окраска лектином WGA AlexaFluor 594. Окраска в псевдоцветах - ФВ представлен зеленым, яд
ра - синим, лектин - пурпурным цветом
гистосовместимости, MHC1, и молекулы меж
торы (VEGFR2) происходит активация секре
клеточной адгезии, ICAM 1 [50]. Приведенные
ции ФВ [51]. Провоспалительные цитокины IL
данные дополняют представления о влиянии
6, IL 9, TNFalpha также инициируют два про
Н2О2 на процессы секреции в ЭК.
цесса - экзоцитоз ФВ [34] и образование актив
Полученные результаты укладываются в
ных форм кислорода, включая Н2О2 [52]. Одна
рамки концепции о роли Н2О2 как вторичного
ко во всех этих случаях возможная связь между
мессенджера, регулирующего, в дополнение к
стимуляцией секреции ФВ и образованием Н2О2
cAMP и Са2+, экзоцитоз ФВ в ЭК. В связи с этим
не исследовалась. Недавно нами было показано,
встаёт вопрос: в действии каких нейроэндо
что блокатор NADPH оксидаз, VAS2870, подав
кринных факторов на данный процесс в ЭК
ляет секрецию ФВ, вызванную гистамином [53].
принимает участие Н2О2. Прямые данные, ука
Таким образом, для ответа на вопрос о роли
зывающие на участие Н2О2 в рецептор зависи
Н2О2 в передаче сигналов агонистов на обмен
мой регуляции экзоцитоза ФВ, в литературе от
ФВ требуются дальнейшие исследования. Раз
сутствуют, однако есть косвенные свидетельства
работанный подход прижизненного окрашива
в пользу этой гипотезы. Известно, что рецепто
ния ФВ, экспонированного на поверхности ЭК,
ры фактора роста VEGF взаимодействуют с
как нам представляется, будет способствовать
NADPH оксидазой NOX4, и через эти же рецеп
прогрессу в этой области.
БИОХИМИЯ том 86 вып. 2 2021
КОНЪЮГАТ ARC1779 Cy5 ДЛЯ ОПРЕДЕЛЕНИЯ ФАКТОРА ВИЛЛЕБРАНДА
155
Финансирование. Работа выполнена при под
Соблюдение этических норм. Все процедуры,
держке Российского научного фонда (грант
выполненные в исследовании с участием людей,
№ 18 15 00417), а также в рамках госзадания
соответствуют этическим стандартам институ
№ 0088 2021 0008 с использованием оборудова
ционального и национального комитета по ис
ния ЦКП ИБР им Н.К. Кольцова РАН.
следовательской этике и Хельсинкской декла
Конфликт интересов. Авторы подтверждают
рации 1964 года и ее последующим изменениям
отсутствие конфликта интересов.
или сопоставимым нормам этики.
СПИСОК ЛИТЕРАТУРЫ
1.
Langmead, C. J., and Summers, R. J. (2018) Molecular
16.
Jilma Stohlawetz, P., Knobl, P., Gilbert, J. C., and Jilma, B.
pharmacology of GPCRs, Br. J. Pharmacol., 175, 4005
(2012) The anti von Willebrand factor aptamer ARC1779
4008, doi: 10.1111/bph.14474.
increases von Willebrand factor levels and platelet counts in
2.
Sriram, K., and Insel, P. A. (2018) G protein coupled
patients with type 2B von Willebrand disease, Thromb.
receptors as targets for approved drugs: how many targets
Haemost., 108, 284 290, doi: 10.1160/TH11 12 0889.
and how many drugs? Mol. Pharmacol., 93, 251 258,
17.
Sakai, K., Someya, T., Harada, K., Yagi, H., Matsui, T.,
doi: 10.1124/mol.117.111062.
and Matsumoto, M. (2019) Novel aptamer to von
3.
Deyev, S. M., and Lebedenko, E. N. (2015) Supramolec
Willebrand factor A1 domain (TAGX 0004) shows total
ular agents for theranostics, Russ. J. Bioorg. Khim., 41, 481
inhibition of thrombus formation superior to ARC1779 and
493, doi: 10.1134/s1068162015050052.
comparable to caplacizumab, Haematologica, 105, 2631
4.
Ellington, A. D., and Szostak, J. W. (1990) In vitro selec
2638, doi: 10.3324/haematol.2019.235549.
tion of RNA molecules that bind specific ligands, Nature,
18.
Matsunaga, K. I., Kimoto, M., and Hirao, I. (2017) High
346, 818 822, doi: 10.1038/346818a0.
Affinity DNA aptamer generation targeting von Willebrand
5.
Tuerk, C., and Gold, L. (1990) Systematic evolution of li
factor A1 domain by genetic alphabet expansion for system
gands by exponential enrichment: RNA ligands to bacte
atic evolution of ligands by exponential enrichment using
riophage T4 DNA polymerase, Science, 249, 505 510,
two types of libraries composed of five different bases, J. Am.
doi: 10.1126/science.2200121.
Chem. Soc., 139, 324 334, doi: 10.1021/jacs.6b10767.
6.
Wang, J., and Li, G. (2011) Aptamers against cell surface
19.
Valentijn, K. M., van Driel, L. F., Mourik, M. J., Hendriks,
receptors: selection, modification and application, Curr. Med.
G. J., Arends, T. J., Koster, A. J., and Valentijn, J. A. (2010)
Chem., 18, 4107 4116, doi: 10.2174/092986711797189628.
Multigranular exocytosis of Weibel-Palade bodies in vas
7.
Spiridonova, V. A., Novikova, T. M., Sizov, V. A.,
cular endothelial cells, Blood,
116,
18071816,
Shashkovskaya, V. S., Titaeva, E. V., Dobrovolsky, A. B.,
doi: 10.1182/blood 2010 03 274209.
Zharikova, E. B., and Mazurov, A. V. (2019) DNA aptamers
20.
Kovacevic, K. D., Buchtele, N., Schoergenhofer, C.,
to thrombin exosite I. Structure function relationships and
Derhaschnig, U., Gelbenegger, G., Brostjan, C., Zhu, S.,
antithrombotic effects, Biochemistry (Moscow), 84, 1521
Gilbert, J. C., and Jilma, B. (2020) The aptamer BT200
1528, doi: 10.1134/S0006297919120113.
effectively inhibits von Willebrand factor (VWF) dependent
8.
Rahimizadeh, K., Al Shamaileh, H., Fratini, M.,
platelet function after stimulated VWF release by desmo
Chakravarthy, M., Stephen, M., Shigdar, S., and Veedu,
pressin or endotoxin, Sci. Rep.,
10,
11180,
R. N. (2017) Development of cell specific aptamers:
doi: 10.1038/s41598 020 68125 9.
recent advances and insight into the selection procedures,
21.
Nimjee, S. M., Dornbos, D., 3rd, Pitoc, G. A., Wheeler,
Molecules, 22, 2070, doi: 10.3390/molecules22122070.
D. G., Layzer, J. M., et al. (2019) Preclinical development
9.
Zhuo, Z., Yu, Y., Wang, M., Li, J., Zhang, Z., et al. (2017)
of a vWF aptamer to limit thrombosis and engender arteri
Recent advances in SELEX technology and aptamer appli
al recanalization of occluded vessels, Mol. Ther., 27, 1228
cations in biomedicine, Int. J. Mol. Sci., 18, 2142,
1241, doi: 10.1016/j.ymthe.2019.03.016.
doi: 10.3390/ijms18102142.
22.
Goncharov, N. V., Sakharov, I., Danilov, S. M., and
10.
Zhang, Y., Lai, B. S., and Juhas, M. (2019) Recent
Sakandelidze, O. G. (1987) Use of collagenase from the
advances in aptamer discovery and applications, Molecules,
hepatopancreas of the Kamchatka crab for isolating and
24, 941, doi: 10.3390/molecules24050941.
culturing endothelial cells of the large vessels in man, Bull.
11.
Bauer, M., Macdonald, J., Henri, J., Duan, W., and
Eksp. Biol. Med., 104, 376 378.
Shigdar, S. (2016) The application of aptamers for
23.
Maciag, T., Cerundolo, J., Ilsley, S., Kelley, P. R., and
immunohistochemistry, Nucleic Acid Ther., 26, 120 126,
Forand, R. (1979) An endothelial cell growth factor from
doi: 10.1089/nat.2015.0569.
bovine hypothalamus: identification and partial character
12.
Zhang, P., Zhao, N., Zeng, Z., Feng, Y., Tung, C. H.,
ization, Proc. Natl. Acad. Sci. USA, 76, 56745678,
Chang, C. C., and Zu, Y. (2009) Using an RNA aptamer
doi: 10.1073/pnas.76.11.5674.
probe for flow cytometry detection of CD30 expressing
24.
Kudryavtsev, I. V., Garnyuk, V. V., Nadeev, A. D., and
lymphoma cells, Lab. Invest., 89, 1423 1432, doi: 10.1038/
Goncharov, N. V. (2014) Hydrogen peroxide modulates
labinvest.2009.113.
expression of surface antigens by human umbilical vein
13.
Sadler, J. E. (1998) Biochemistry and genetics of von
endothelial cells in vitro, Biochemistry (Moscow) Supp.
Willebrand factor, Annu. Rev. Biochem., 67, 395 424,
Ser. A Membr. Cell Biol., 8, 97102, doi: 10.1134/
doi: 10.1146/annurev.biochem.67.1.395.
S1990747813050103.
14.
Duggan, S. (2018) Caplacizumab: first global approval,
25.
Bray, M. A., Vokes, M. S., and Carpenter, A. E. (2015)
Drugs, 78, 1639 1642, doi: 10.1007/s40265 018 0989 0.
Using CellProfiler for automatic identification and mea
15.
Bae, O. N. (2012) Targeting von Willebrand factor as
surement of biological objects in images, Curr. Protoc. Mol.
a novel anti platelet therapy; application of ARC1779,
Biol., 109, 1 18, doi: 10.1002/0471142727.mb1417s109.
an anti vWF aptamer, against thrombotic risk, Arch.
26.
Avdonin, P. V., Tsitrina, A. A., Mironova, G. Y., Avdonin,
Pharm. Res., 35, 1693 1699, doi: 10.1007/s12272 012
P. P., Zharkikh, I. L., Nadeev, A. D., and Goncharov, N. V.
10003.
(2017) Hydrogen peroxide stimulates exocytosis of von
БИОХИМИЯ том 86 вып. 2 2021
156
АВДОНИН и др.
Willebrand factor in human umbilical vein endothelial
40.
Vischer, U. M., Jornot, L., Wollheim, C. B., and Theler,
cells, Biol. Bull.,
44,
531537, doi:
10.1134/
J. M. (1995) Reactive oxygen intermediates induce regulat
s106235901705003x.
ed secretion of von Willebrand factor from cultured human
27.
Hattori, R., Hamilton, K. K., Fugate, R. D., McEver, R.
vascular endothelial cells, Blood, 85, 3164 3172.
P., and Sims, P. J. (1989) Stimulated secretion of endothe
41.
Yang, S., Zheng, Y., and Hou, X. (2019) Lipoxin A4
lial von Willebrand factor is accompanied by rapid redistri
restores oxidative stress induced vascular endothelial cell
bution to the cell surface of the intracellular granule mem
injury and thrombosis related factor expression by its
brane protein GMP 140, J. Biol. Chem., 264, 7768 7771.
receptor mediated activation of Nrf2 HO 1 axis, Cell.
28.
Kaufmann, J. E., Oksche, A., Wollheim, C. B., Gunther,
Signal., 60, 146 153, doi: 10.1016/j.cellsig.2019.05.002.
G., Rosenthal, W., and Vischer, U. M. (2000) Vasopressin
42.
Mourik, M. J., Valentijn, J. A., Voorberg, J., Koster, A. J.,
induced von Willebrand factor secretion from endothelial
Valentijn, K. M., and Eikenboom, J.
(2013) Von
cells involves V2 receptors and cAMP, J. Clin. Invest., 106,
Willebrand factor remodeling during exocytosis from vas
107 116, doi: 10.1172/JCI9516.
cular endothelial cells, J. Thromb. Haemost., 11, 2009
29.
Esposito, B., Gambara, G., Lewis, A. M., Palombi, F.,
2019, doi: 10.1111/jth.12401.
D’Alessio, A., et al. (2011) NAADP links histamine H1
43.
Zhu, S., Gilbert, J. C., Hatala, P., Harvey, W., Liang, Z.,
receptors to secretion of von Willebrand factor in human
Gao, S., Kang, D., and Jilma, B. (2020) The development
endothelial cells, Blood, 117, 4968 4977, doi: 10.1182/
and characterization of a long acting anti thrombotic von
blood 2010 02 266338.
Willebrand factor (VWF) aptamer, J. Thromb. Haemost.,
30.
Schluter, T., and Bohnensack, R. (1999) Serotonin
18, 1113 1123, doi: 10.1111/jth.14755.
induced secretion of von Willebrand factor from human
44.
Shappell, S. B., Toman, C., Anderson, D. C., Taylor, A. A.,
umbilical vein endothelial cells via the cyclic AMP signal
Entman, M. L., and Smith, C. W.
(1990) Mac 1
ing systems independent of increased cytoplasmic calcium
(CD11b/CD18) mediates adherence dependent hydrogen
concentration, Biochem. Pharmacol., 57, 1191 1197.
peroxide production by human and canine neutrophils,
31.
Vischer, U. M., and Wollheim, C. B. (1997) Epinephrine
J. Immunol., 144, 2702 2711.
induces von Willebrand factor release from cultured
45.
Jones, D. P. (2008) Radical free biology of oxidative stress,
endothelial cells: involvement of cyclic AMP dependent
Am. J. Physiol. Cell. Physiol.,
295, C849 C868,
signalling in exocytosis, Thromb. Haemost., 77, 1182 1188.
doi: 10.1152/ajpcell.00283.2008.
32.
Vischer, U. M., and Wollheim, C. B. (1998) Purine
46.
Forman, H. J., Bernardo, A., and Davies, K. J. (2016)
nucleotides induce regulated secretion of von Willebrand
What is the concentration of hydrogen peroxide in blood
factor: involvement of cytosolic Ca2+ and cyclic adenosine
and plasma? Arch. Biochem. Biophys.,
603,
4853,
monophosphate dependent signaling in endothelial exocy
doi: 10.1016/j.abb.2016.05.005.
tosis, Blood, 91, 118 127.
47.
Varma, S. D., and Devamanoharan, P. S. (1991) Hydrogen
33.
Matsushita, K., Yamakuchi, M., Morrell, C. N., Ozaki,
peroxide in human blood, Free Radic. Res. Commun., 14,
M., O’Rourke, B., Irani, K., and Lowenstein, C. J. (2005)
125 131, doi: 10.3109/10715769109094124.
Vascular endothelial growth factor regulation of
48.
Nadeev, A. D., Kudryavtsev, I. V., Serebriakova, M. K.,
Weibel-Palade body exocytosis, Blood, 105, 207214,
Avdonin, P. V., Zinchenko, V. P., and Goncharov, N. V.
doi: 10.1182/blood 2004 04 1519.
(2016) Dual proapoptotic and pronecrotic effect of hydro
34.
Bernardo, A., Ball, C., Nolasco, L., Moake, J. F., and
gen peroxide on human umbilical vein endothelial cells,
Dong, J. F. (2004) Effects of inflammatory cytokines on
Cell Tiss. Biol.,
10,
145151, doi:
10.1134/
the release and cleavage of the endothelial cell derived
S1990519X16020097.
ultralarge von Willebrand factor multimers under flow,
49.
Yang, D., Liu, X., Liu, M., Chi, H., Liu, J., and Han, H.
Blood, 104, 100 106, doi: 10.1182/blood 2004 01 0107.
(2015) Protective effects of quercetin and taraxasterol
35.
Vischer, U. M. (2006) Von Willebrand factor, endothelial
against H2O2 induced human umbilical vein endothelial
dysfunction, and cardiovascular disease, J. Thromb.
cell injury in vitro, Exp. Ther. Med., 10, 1253 1260,
Haemost., 4, 1186 1193, doi: 10.1111/j.1538 7836.2006.
doi: 10.3892/etm.2015.2713.
01949.x.
50.
Bradley, J. R., Johnson, D. R., and Pober, J. S. (1993)
36.
Breton Romero, R., and Lamas, S. (2014) Hydrogen per
Endothelial activation by hydrogen peroxide. Selective
oxide signaling in vascular endothelial cells, Redox Biol., 2,
increases of intercellular adhesion molecule 1 and major
529 534, doi: 10.1016/j.redox.2014.02.005.
histocompatibility complex class I, Am. J. Pathol., 142,
37.
Rhee, S. G., Kang, S. W., Jeong, W., Chang, T. S., Yang,
1598 1609.
K. S., and Woo, H. A. (2005) Intracellular messenger func
51.
Xiong, Y., Huo, Y., Chen, C., Zeng, H., Lu, X., et al.
tion of hydrogen peroxide and its regulation by peroxire
(2009) Vascular endothelial growth factor (VEGF) recep
doxins, Curr. Opin. Cell Biol.,
17,
183189,
tor 2 tyrosine 1175 signaling controls VEGF induced von
doi: 10.1016/j.ceb.2005.02.004.
Willebrand factor release from endothelial cells via phos
38.
Avdonin, P. V., Nadeev, A. D., Mironova, G. Y., Zharkikh,
pholipase C gamma 1 and protein kinase A dependent
I. L., Avdonin, P. P., and Goncharov, N. V.
(2019)
pathways, J. Biol. Chem., 284, 23217 23224, doi: 10.1074/
Enhancement by hydrogen peroxide of calcium signals in
jbc.M109.019679.
endothelial cells induced by 5 HT1B and 5 HT2B recep
52.
Mo, S. J., Son, E. W., Rhee, D. K., and Pyo, S. (2003)
tor agonists, Oxid. Med. Cell. Longev., 2019, 1701478,
Modulation of TNF alpha induced ICAM 1 expression,
doi: 10.1155/2019/1701478.
NO and H2O2 production by alginate, allicin and ascorbic
39.
Avdonin, P. V., Nadeev, A. D., Tsitrin, E. B., Tsitrina, A.
acid in human endothelial cells, Arch. Pharm. Res., 26,
A., Avdonin, P. P., Mironova, G. Y., Zharkikh, I. L., and
244 251.
Goncharov, N. V. (2017) Involvement of two pore chan
53.
Avdonin, P. V., Rybakova, E. Y., Avdonin, P. P., Trufanov,
nels in hydrogen peroxide induced increase in the level of
S. K., Mironova, G. Y., Tsitrina, A. A., and Goncharov,
calcium ions in the cytoplasm of human umbilical vein
N. V. (2019) VAS2870 Inhibits histamine induced calcium
endothelial cells, Dokl. Biochem. Biophys., 474, 209 212,
signaling and vWF secretion in human umbilical vein
doi: 10.1134/S1607672917030152.
endothelial cells, Cells, 8, 196, doi: 10.3390/cells8020196.
БИОХИМИЯ том 86 вып. 2 2021
КОНЪЮГАТ ARC1779 Cy5 ДЛЯ ОПРЕДЕЛЕНИЯ ФАКТОРА ВИЛЛЕБРАНДА
157
THE USE OF FLUORESCENTLY LABELED ARC1779 APTAMER
FOR ASSESSING THE EFFECT OF H2O2
ON VON WILLEBRAND FACTOR EXOCYTOSIS*
P. P. Avdonin1, S. K. Trufanov1, E. Yu. Rybakova1, A. A. Tsitrina1,
N. V. Goncharov2,3, and P. V. Avdonin1**
1 Koltsov Institute of Development Biology Russian Academy of Sciences,
119334 Moscow, Russia; E mail: pvavdonin@yandex.ru
2 Sechenov Institute of Evolutionary Physiology and Biochemistry Russian Academy of Sciences,
194223 St. Petersburg, Russia
3 Scientific Research Institute of Hygiene, Occupational Pathology and Human Ecology,
FMBA, 188663 Kuzmolovsky, Leningrad Region, Russia
Here, we propose a new approach for quantitative estimation of von Willebrand factor (vWF) exposed on the surface
of endothelial cells (ECs) using the ARC1779 aptamer that interacts with the vWF A1 domain. To visualize complex
formation between vWF and the aptamer, the latter was conjugated with the Cy5 fluorescent label. Cultured human
umbilical vein endothelial cells (HUVEC) were stained with the ARC1779 Cy5 conjugate and imaged with a fluores
cence microscope. The images were analyzed with the CellProfiler software. vWF released from the Weibel-Palade
bodies was observed as bright dot like structures of round and irregular shape, the number of which increased several
times after HUVEC exposure to histamine or thrombin. Staining with ARC1779 Cy5 also revealed long filamentous
vWF structures on the surface of activated HUVEC. vWF secretion by ECs is activated by the second messengers
cAMP and Ca2+. There is evidence that hydrogen peroxide also acts as a second messenger in ECs. In addition, exoge
nous H2O2 formed in leukocytes can enter ECs. The aim of our study was to determine the effect of H2O2 on the vWF
exposure at the surface of HUVEC using the proposed method. It was shown that hydrogen peroxide at concentration
100 μM, which is lower than the cytotoxicity threshold of H2O2 for cultured HUVEC, increased several times the
number of dot like structures and total amount of vWF exposed on plasma membrane of HUVEC, which suggest that
H2O2 acts as a mediator that activates exocytosis of Weibel-Palade bodies and vWF secretion in the vascular endothe
lium during inflammation and upon elevated generation of endogenous reactive oxygen species in ECs.
Keywords: von Willebrand factor, aptamers, hydrogen peroxide, endothelial cells, secretion
БИОХИМИЯ том 86 вып. 2 2021