БИОХИМИЯ, 2021, том 86, вып. 2, с. 200 - 213
УДК 577.29:576.5
УРОВЕНЬ мРНК ТРАНСПОРТЕРОВ ЦИНКА ЗАВИСИТ
ОТ СОДЕРЖАНИЯ ЦИНКА И ВОСПАЛИТЕЛЬНЫХ ПРОЦЕССОВ,
ИНДУЦИРОВАННЫХ HIV 1 Tat В ЛИНИЯХ КЛЕТОК МЫШЦ (RD)
И МОНОЦИТАХ (THP 1)*
© 2021
К. Аллури1, С.Р. Ятхапу2, Н.Б. Кондапалли3,
Р. Хемалатха3, К.М. Наир4**, С. Гош1**
1 Molecular Biology Division, ICMR National Institute of Nutrition,
500007 Hyderabad, India; электронная почта: bihongo@yahoo.com
2 Drug Toxicology Division, ICMR National Institute of Nutrition, 500007 Hyderabad, India
3 Microbiology and Immunology Division, ICMR National Institute of Nutrition, 500007 Hyderabad, India
4 Micronutrient Division, ICMR National Institute of Nutrition,
500007 Hyderabad, India; электронная почта: nairthayil@gmail.com
Поступила в редакцию 01.06.2020
После доработки 27.08.2020
Принята к публикации 15.09.2020
При недостатке цинка в моноцитах и мышечных клетках наблюдается функционально различное поведе
ние с точки зрения системы депонирования цинка (в мышцах, в отличие от моноцитов, уровень цинка со
храняется на обычном уровне). Целью настоящей работы было изучение влияния уровня цинка и HIV 1 Tat
опосредованного воспаления на экспрессию транспортеров цинка в этих типах клеток. Уровень экспрессии
транспортеров цинка (ZnTs, ZIPs и металлотионеина MT) в клетках RD и THP 1 определяли с помощью ме
тода qRT PCR как по отдельности, так и при совместном культивировании этих клеток. Уровни экспрессии
белка ZnT1 определяли с использованием метода вестерн блоттинга. Значительное повышение уровня
мРНК белков MT и ZnT1 при добавлении цинка и снижение при дефиците цинка указывает на важную роль
генов, кодирующих транспортеры цинка, в поддержании гомеостаза цинка в этих тканях. В клетках RD
ZIP10 продемонстрировал обратную корреляцию с уровнем цинка, в то время как в клетках THP 1 никакой
корреляции обнаружено не было. Tat индуцированное воспаление приводило к значительному повышению
уровня транскриптов MT, IL 6, ZIP7, ZIP8 и ZIP9 в клетках RD при сокультивировании, в то время как в
клетках THP 1 наблюдалось повышение уровня IL 1β и снижение уровня ZIP7 и ZIP14. Уровень цинка и
HIV 1 Tat индуцированное воспаление, по видимому, оказывают влияние на дифференциальную экспрес
сию MT, ZnTs и ZIPs в мышечных клетках и в моноцитах.
КЛЮЧЕВЫЕ СЛОВА: транспортеры цинка, ZIP, ZnT, MT, совместное культивирование, HIV 1 Tat,
THP 1, RD.
DOI: 10.31857/S0320972521020056
ВВЕДЕНИЕ
его дефиците в течение 8-12 недель также сни
жается [3]. Концентрация ионов цинка не изме
При недостатке цинка его уровень в скелет
няется при краткосрочном незначительном де
ной мышце, коже и сердце не изменяется, одна
фиците цинка (12-16 мкМ), однако при про
ко снижается в кости, моноцитах, печени, яич
должающемся дефиците Zn происходит сниже
ках и плазме крови [1, 2]. Содержание цинка в
ние его концентрации в плазме крови [4]. Гомео
лимфоцитах, гранулоцитах и тромбоцитах при
стаз цинка поддерживается в результате коорди
Принятые сокращения: BCR - рецептор B клеток; DMEM - модифицированная Дульбекко среда Игла;
DMSO - диметилсульфоксид; FBS - фетальная бычья сыворотка; HIV - вирус иммунодефицита человека; IKK - ΙκΒ ки
наза; IL - интерлейкин; MHC - основной комплекс гистосовместимости; MT - металлотионеин; MTF - металлотран
скрипционный фактор; Nef - негативный фактор экспрессии; PKB/AKT - протеинкиназа В; RD - клетки рабдомиосар
комы; SLC - семейство транспортеров растворенных веществ; Tat - трансактиватор транскрипции; TNF - фактор нек
роза опухолей α; TPEN - N,N,N′,N′ тетракис (2 пиридилметил) этилендиамин; ZIP - Zrt и Irt подобный белок;
ZnT - транспортер цинка.
biokhimiya, в рубрике «Papers in Press», BM20 125, 02.11.2020.
** Адресат для корреспонденции.
200
РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ ТРАНСПОРТЕРОВ ЦИНКА
201
нированной регуляции активности транспорте
образом облегчая встраивание вирусной ДНК в
ров цинка, которые принадлежат к двум основ
ДНК клетки хозяина [16, 17]. Более того, химио
ным семействам: белки ZnTs (транспортеры
терапевтическая эффективность некоторых аро
цинка) и белки ZIPs (Zrt и Irt подобные белки)
матических C нитрозосоединений основана на
вместе с белком MT (металлотионеин), который
их способности удалять цинк из мотивов цинко
представляет собой внутриклеточный цинк
вого пальца нуклеокапсида HIV 1, таким обра
связывающий белок. Белки ZnTs (ZnT1-ZnT10)
зом предотвращая инфицирование вирусом [18].
кодируются семейством генов SLC30 (solute car
Было показано, что такие белки HIV 1, как
rier) и участвуют в высвобождении внутрикле
гликопротеин 120 (gp120), негативный фактор
точного Zn; в то время как белки транспортеры
экспрессии (Nef), трансактиватор транскрип
ZIPs (ZIP1-ZIP14), кодируемые семейством ге
ции (Tat) и регулятор экспрессии белков вирио
нов SLC39, ведут к повышению внутриклеточ
на (Rev), могут вызывать атрофию мышц, ката
ной концентрации цинка.
ракту, нефропатию, поражения кожи и иммуно
Кроме того, экспрессия белков ZIPs изменя
дефицитные состояния [19, 20]. Tat индуциро
ется с возрастом, а также регулируется посттран
ванная запрограммированная гибель клеток
скрипционно. Было показано, что уровень цин
приводит к снижению числа лимфоцитов у
ка per se сильнее влияет на экспрессию ZIP1,
ВИЧ инфицированных больных [21]. Напро
ZIP2 и ZIP3 в лимфоцитах молодых людей по
тив, Tat обладает также и митогенной актив
сравнению с пожилыми [5]. Токсичность цинка
ностью в клетках эпителия молочной железы и
предотвращается эндоцитозом транспортера
амниона и способствует передаче вируса от ма
ZIP4 из плазматической мембраны или его
тери к ребенку [22]. Интересно, что цинк явля
убиквитин опосредованной деградацией [6].
ется необходимым для активности белка Tat, чье
Когда цинк находится в доступности, происхо
действие обязательно для репликации HIV [23].
дит усиление трансляции ZIP5, опосредованное
Известно, что Tat стимулирует секрецию жид
взаимодействием консервативной шпильки и
кости от серозной к люминальной стороне энте
двух перекрывающихся сайтов связывания
роцитов, в то время как цинк предотвращает та
микроРНК в 3′ нетранслируемом регионе [7].
кую активность [24].
Цинк играет незаменимую роль в развитии,
Tat обладает ингибирующим действием в от
созревании и функционировании NK клеток
ношении основного комплекса гистосовмести
(natural killers) и NKT клеток (natural killer T
мости (MHC - major histocompatibility complex)
cells) [8]. Известно, что активация тучных клеток
I го и II го класса и индуцирует экспрессию
при аллергическом ответе замедленного типа
провоспалительных цитокинов, таким образом
происходит при участии белка ZnT5 [9]. Было
модулируя иммунный ответ на инфицирование
показано, что у мышей при сепсисе/бактериаль
HIV 1 [25, 26]. Кроме того, значительное коли
ной инфекции повышается экспрессия ZIP8 в
чество данных подтверждает, что развитию ка
ответ на активацию NF κB. ZIP8 опосредован
хексии, или болезни похудания (slim disease), у
ный приток цинка вызывает ингибирование ак
больных ВИЧ/СПИД способствует ограничен
тивности IKK (IκB киназы) и тем самым подав
ный MHC I полиомиозит, опосредованный ци
ляет воспаление [10]. Аналогично известно, что
тотоксическим действием T клеток [27, 28].
ZIP9 способствует повышению уровня внутри
Состояние кахексии в дальнейшем усугубляется
клеточного цинка, приводя к активации проте
действием провоспалительных цитокинов, та
инкиназы В (PKB/AKT), фосфорилированию
ких как интерлейкин 1β и фактор некроза опу
киназы, регулируемой внеклеточными сигнала
холей TNF α, которые секретируются ВИЧ ин
ми ERK (extracellular signal regulated kinase), и ак
фицированными моноцитами/макрофагами
тивации рецептора В клеток (BCR) [11]. В раз
[29]. Возникновение атрофии скелетных мышц
личных работах показано, что повышенные кон
у больных ВИЧ/СПИД может быть вызвано из
центрации Zn вызывают ингибирование тран
менениями циркулирующих уровней цитоки
скрипции вируса иммунодефицита человека
нов и миостатина в крови больных [30, 31].
(HIV 1) наряду с формированием новых инфек
С учетом данных, представленных выше,
ционных вирионов. В то же время применение
целью настоящей работы было изучение влия
Zn совместно с Зидовудином (Zidovudine,
ния уровня цинка и воспаления в клетках, из ко
ZDV/AZT) приводит к снижению заболеваемос
торых цинк высвобождается (моноциты), и
ти, вызванной оппортунистическими инфекция
клетках, в которых депонируется (мышечные
ми, и диареи у ВИЧ инфицированных детей
клетки). Мы предположили, что за различный
[12-15]. Хорошо известно, что ВИЧ является
ответ этих клеток на дефицит цинка и развитие
цинк зависимым ретровирусом, в котором Zn
воспаления ответственны транспортеры цинка.
стимулирует вирусный фермент интегразу, таким
Поэтому мы провели эксперименты на ткане
БИОХИМИЯ том 86 вып. 2 2021
202
АЛЛУРИ и др.
специфичных репрезентативных линиях кле
Поглощение этилового эфира Zinquin в культу
ток, а именно THP 1 (моноциты) и клетки раб
рах клеток RD и THP 1. Как было описано в
домиосаркомы (RD) мышцы человека. Был изу
предыдущем разделе, клетки инкубировали с
чен ответ этих клеток на добавление экзогенно
25 мкМ цинка в виде сульфата цинка или 5 мкМ
го цинка, его дефицит, опосредованный
TPEN в течение 4 ч в бессывороточной среде.
действием TPEN - хелатора внутриклеточного
Клетки промывали PBS и затем инкубировали с
цинка (N,N,N′,N′ тетракис (2 пиридилметил)
10 мкМ цинк специфичного флуоресцентного
этилендиамин), а также воспаление, индуциро
зонда этилового эфира Zinquin в течение 30 мин
ванное HIV 1 Tat.
[33]. Флуоресцентный комплекс Zn-Zinquin ви
зуализировали, считывая флуоресцентную
эмиссию на 490 нм при возбуждении на 370 нм с
МАТЕРИАЛЫ И МЕТОДЫ
использованием флуоресцентного микроскопа с
фильтром GFP (EVOS FLC «Life Technologies»,
Реагенты. Реагенты для культивирования
США).
клеток, среда DMEM (Dulbecco’s Modified Eagle
Определение внутриклеточного цинка. Сус
Medium), среда RPMI 1640 (Roswell Park
пензию клеток (5 × 106/мл) THP 1 и трипсини
Memorial Institute Medium), FBS (фетальная
зированных клеток RD инкубировали с 10 мкМ
бычья сыворотка), глутамакс, смесь антибиоти
этилового эфира Zinquin при 37 °C в течение
ков и противогрибковых агентов, поли L ли
30 мин [34]. Клетки трижды промывали PBS и
зин, этиловый эфир Zinquin (zinquin ethyl ester),
ресуспендировали в PBS. Флуоресценцию проб
DMSO (диметилсульфоксид) и TPEN были за
объемом 2 мл (5 × 106 клеток/мл) в кюветах реги
казаны в компании «Sigma Aldrich» (США). На
стрировали при длине волны 490 нм при воз
бор пептидов ВИЧ (HIV 1 Consensus B Tat
буждении на 370 нм с помощью спектрофлуори
Peptide Pool; Cat:12706) был любезно предостав
метра Jasco FP 6500 («Jasco», Япония).
лен по программе Национального Института
Вестерн блоттинг для определения уровня
Здоровья по борьбе с ВИЧ (США).
транспортера ZnT1. Клетки лизировали в буфе
Источники клеточных линий. Линия лейкеми
ре для анализа радиоиммунопреципитации
ческих моноцитов человека THP 1 была любез
RIPA, содержащем коктейль ингибиторов про
но предоставлена институтом CSIR IICT (Ин
теаз. Концентрацию белка в супернатанте опре
дия). Линия клеток RD была приобретена в
деляли с помощью набора Micro BCA («Thermo
ATCC (American type culture collection, США).
Fisher Scientific», США). Белки (50 мкг) разде
Клетки выдерживали при 37 °C в увлаженной ат
ляли с помощью электрофореза в 10% ном по
мосфере с 5% CO2, и культуральную среду сме
лиакриламидном геле в присутствии додецил
няли один раз в два дня. Клетки THP 1 и RD
сульфата натрия SDS (10% SDS PAGE), разде
культивировали в среде RPMI 1640 и DMEM со
ленные белки переносили на нитроцеллюлоз
ответственно с добавлением 10% FBS и по 1%
ные мембраны («Bio Rad», США). После бло
глутамакса и смеси антибиотиков и противог
кировки мембран 5% ным раствором сухого
рибковых средств. Клетки выдерживали в лога
молока, мембраны инкубировали в течение но
рифмической фазе роста, эксперименты прово
чи при 4 °C с антителами против ZnT1. Затем их
дили после достижения состояния ~70-80% ной
промывали и инкубировали в течение 1 ч с вто
конфлюэнтности. Культуральная среда, за иск
ричными антителами класса IgG, конъюгиро
лючением FBS, не содержала цинк.
ванными с пероксидазой хрена (HRP). Имму
Обработка цинком и TPEN клеток THP 1 и
ноблоты проявляли с помощью набора ECL
RD. Для удаления избыточного цинка из среды
detection kit («Bio Rad», США) и получали
клетки промывали фосфатным солевым раство
изображения с помощью системы визуализа
ром PBS. Клетки THP 1 и RD обрабатывали
ции G box («Syngene», США). В качестве конт
25 мкМ цинка в форме сульфата цинка в комп
ролей количества белка при анализе клеток
лексе с 25 мкМ бычьего сывороточного альбу
THP 1 и RD использовали β актин и ГАФДГ
мина BSA («Sigma», США) [32] или 5 мкМ
(глицеральдегид 3 фосфат дегидрогеназа) со
TPEN (хелатор внутриклеточного цинка) для
ответственно.
создания дефицита цинка, оцениваемого по
Совместное культивирование клеток RD и
жизнеспособности клеток (окрашивание трипа
THP 1 для определения роли HIV 1 Tat. Для оп
новым синим) в течение 4 ч в бессывороточной
ределения роли Tat индуцированного воспале
среде. Затем клетки THP 1 фиксировали на
ния на межклеточные взаимодействия было
покровных стеклах, покрытых 0,1 мг/мл поли
проведено совместное культивирование клеток
L лизина. Все эксперименты выполнялись
RD и THP 1. Вкратце, клетки RD были посаже
трижды как независимые повторения.
ны с плотностью 0,9 × 106/мл, через 24 ч среду
БИОХИМИЯ том 86 вып. 2 2021
РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ ТРАНСПОРТЕРОВ ЦИНКА
203
меняли на свежую DMEM. Моноциты сажали
дважды промывочным буфером и инкубировали
на монослой клеток RD в соотношении 1 : 1, до
с антителами PE Cy7 TNF α («BD Biosciences»,
бавляли Tat (100 нг/мл) и инкубировали в тече
США) при комнатной температуре в темноте в
ние 8 ч [35]. Суспензию клеток THP 1 помеща
течение 30 мин для определения уровня мембра
ли на клетки RD аналогичным образом и ис
носвязанного TNF α. Несвязавшиеся антитела
пользовали в качестве контроля для сокультиви
дважды отмывали буфером Perm/Wash («BD
руемых клеток. После инкубации суспендиро
Biosciences», США) и ресуспендировали в
ванные клетки THP 1 были аспирированы и от
400 мкл промывочного буфера. Полученные с
мыты с помощью PBS. Прикрепленные клетки
использованием проточного цитометра (BD
RD также тщательно промывались PBS для уда
FACS Aria II, «BD Biosciences», США) данные
ления суспендированных клеток THP 1. Эти
анализировали с помощью программы
снятые клетки были использованы для опреде
Flow Jo 10.7 («Flow Jo», США).
ления уровня экспрессии MT, ZnTs, ZIPs, IL 6 и
MHC I при воспалении. Совместно культи
IL 1β с использованием метода количественной
вируемые клетки RD и THP 1 окрашивали пре
ПЦР в реальном времени (qRT PCR).
паратом anti HLA ABC FITC («Miltenyi Biotec»,
Количественная ПЦР в реальном времени
Германия) или препаратом для изотопного
(qRT PCR). Тотальную РНК выделяли из клеток
контроля REA Control FITC («Miltenyi Biotec»,
с помощью реагента Trizol («Life Technologies»,
Германия) в течение 30 мин при 4 °C в соответ
США), следуя инструкциям производителя. Для
ствии с инструкциями производителя. Клетки
синтеза кДНК с помощью набора Verso cDNA
анализировали с помощью проточной цитомет
synthesis kit («Thermo Fisher Scientific», США)
рии, как было описано выше.
использовали 1 микрограмм РНК. qRT PCR
проводили с использованием красителя
2×
SYBR Green («Takara Bio», Япония) и ген спе
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
цифичных праймеров (табл. S1 в Приложении)
на термоциклере C1000 с оптическим модулем
Гомеостаз цинка поддерживается путем регу
CFX96 («Bio Rad», США). Все протоколы про
ляции экспрессии тканеспецифичных транс
водились дважды. Продукты амплификации
портеров цинка. Тем не менее, насколько нам
ПЦР также анализировали с помощью электро
известно, пока не было проведено детальных
фореза в агарозном геле с окрашиванием этидиум
исследований экспрессии генов в функциональ
бромидом для подтверждения чистоты ампли
но различных тканях. В настоящей работе мы
фицированного продукта ПЦР. Для нормализа
определяли уровень внутриклеточного цинка с
ции результатов, полученных на клетках RD и
помощью флуоресцентной микроскопии и
THP 1, использовали ГАФДГ и β актин соответ
спектрофлуориметрии. Профили экспрессии
ственно. Продукты ПЦР всех образцов также
MTs, ZnTs и ZIPs анализировали с помощью ко
анализировали путем определения кривой плав
личественной ПЦР в реальном времени
ления для подтверждения чистоты продукта
(qRT PCR).
амплификации. В качестве отрицательного
Уровни внутриклеточного Zn в условиях из
контроля использовали соответствующий обра
бытка или недостатка цинка. Клетки THP 1
зец мРНК без обратной транскриптазы. Отно
(рис. 1, а-в) и RD (рис. 1, г-е), культивируемые
сительный уровень экспрессии мРНК рассчи
с цинком и TPEN в течение 4 ч, демонстрирова
тывали по методу 2-ΔΔCT.
ли флуоресценцию этилового эфира Zinquin в
Окрашивание внутриклеточных цитокинов для
сравнении с соответствующими контролями,
определения TNF α. Экспрессию цитокина
что указывает на то, что интернализация Zinquin
TNF α в ответ на введение пула пептидов Tat
может быть использована для количественного
исследовали в иммунных клетках, т.е. только в
определения цинка в этих клетках. Соответ
клетках THP 1. Вкратце, совместно культиви
ственно, средний уровень цинка в обеих линиях
руемые клетки THP 1 обрабатывали Tat
клеток был значительно (p < 0,05) повышен при
(100 нг/мл), затем добавляли ингибитор транс
добавлении цинка (7,92 ± 0,10 нМ в клетках
порта белков GolgiStop™ («BD Biosciences»,
THP 1 и 13,5 ± 0,44 нМ в клетках RD) в сравне
США) и инкубировали не более 12 ч. Реакцию
нии с контролем (1,97 ± 0,103 нМ в клетках
останавливали центрифугированием клеток и
THP 1 и 5,78 ± 0,3 нМ в клетках RD). В то же
промыванием жидкостью Sheath fluid (BD FACS
время было обнаружено существенное (p < 0,05)
Flow, «BD Biosciences», США). Затем клетки ре
снижение уровня цинка в этих клетках после об
суспендировали в
250
мкл раствора цито
работки TPEN (0,94 ± 0,06 нМ в клетках THP 1
фикс/цитоперм в темноте при комнатной тем
и 3,98 ± 0,45 нМ в клетках RD) в сравнении с
пературе в течение 20 мин. Клетки промывали
контролем (табл. S2 в Приложении). Таким об
БИОХИМИЯ том 86 вып. 2 2021
204
АЛЛУРИ и др.
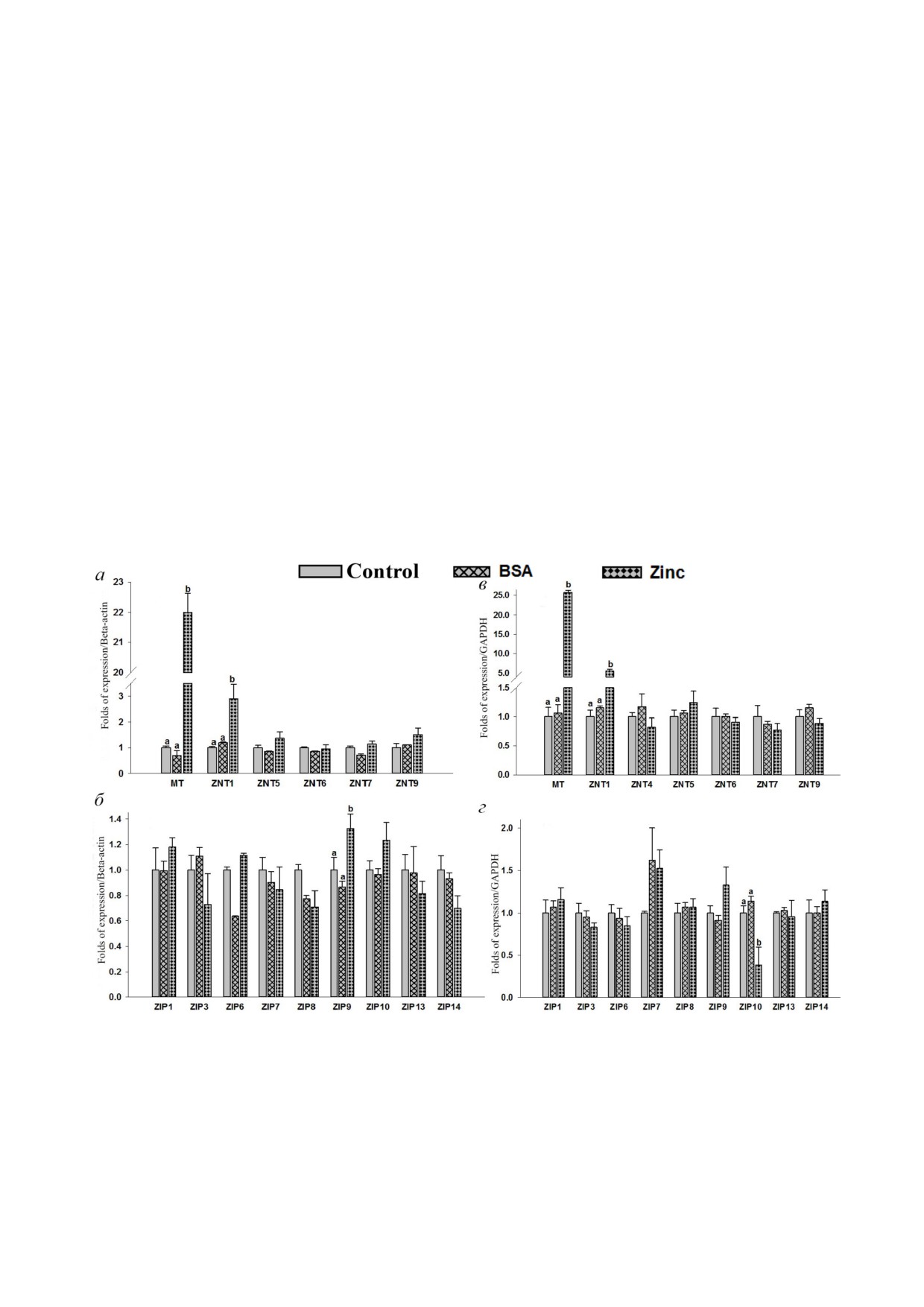
Рис. 1. Определение содержания внутриклеточного цинка в клетках RD и THP 1. Клетки обрабатывали 25 мкМ цинка
или 5 мкМ TPEN в течение 4 ч. Затем клетки промывали PBS для удаления красителя вне клеток и получали изображе
ния при λex 359 нм/λem 485 нм, фиксируя синюю флуоресценцию комплекса цинка и этилового эфира Zinquin. Клетки
THP 1: а - использованные в качестве контроля не обработанные красителем; б - обработанные 25 мкМ цинка; в - об
работанные 5 мкМ TPEN. Аналогично клетки RD: г - использованные в качестве контроля не обработанные красителем;
д - обработанные 25 мкМ цинка; е - обработанные 5 мкМ TPEN; инкубировали с 10 мкМ этилового эфира Zinquin в те
чение 30 мин. (С цветными вариантами рис. 1, 5 и 6 можно ознакомиться в электронной версии статьи на сайте: http://
sciencejournals.ru/journal/biokhsm/)
разом, обе линии клеток THP 1 и RD реагируют
также значительно (p
< 0,05) повышалась
на уровень цинка, и эти результаты подтвержда
(в 1,3 раза) по сравнению с контролем при их
ют данные, полученные нами ранее на клетках
обработке экзогенным цинком (рис. 2, б).
кости [36].
Аналогично значительное (p < 0,05) повыше
Экспрессия MT, hZnTs и hZIPs в ответ на до
ние экспрессии металлотионеина (в 25 раз) и
бавление цинка. Уровни экспрессии MT, hZnTs и
ZnT1 (в 5,6 раза) в клетках RD указывает на их
hZIPs (h - human, белки клеток человека) в
положительный ответ на избыток цинка в среде
клетках THP 1 измеряли с помощью qRT PCR
(рис. 2, в). Более того, методом вестерн блот
при добавлении 25 мкМ цинка (ZnSO4). Значи
тинга было подтверждено повышение уровня
тельное (p < 0,05) повышение (в 22 раза) уровня
белка ZnT1 в этих клетках (рис. 3, б). При иссле
мРНК металлотионеина (MT) в клетках THP 1
довании различных белков ZIPs было обнаруже
свидетельствует о сильном ответе на добавление
но значительное снижение (в 0,3 раза) уровня
цинка (рис. 2, а; табл. S3 в Приложении). Ана
экспрессии белка ZIP10 в сравнении с необра
логично наблюдалось значительное (p < 0,05)
ботанным контролем (рис. 2, г).
повышение (в 2,9 раза) уровня экспрессии
Добавление цинка не оказало заметного вли
мРНК ZnT1 в клетках THP 1 (рис. 2, а). Более
яния на экспрессию других белков ZnTs
того, данное наблюдение было подтверждено
(ZnT4-ZnT7 и ZnT9) и ZIPs (ZIP1, ZIP3,
повышенной экспрессией белка ZnT1
ZIP6-ZIP9, ZIP13 и ZIP14). В клетках THP 1 и
(рис. 3, а). Экспрессия ZIP9 в клетках THP 1
RD экспрессии белков ZnT2, ZnT3, ZnT8,
БИОХИМИЯ том 86 вып. 2 2021
РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ ТРАНСПОРТЕРОВ ЦИНКА
205
ZnT10, ZIP2, ZIP4, ZIP5 и ZIP12 выявлено не
тельно повышались уровни MT и ZnT1, что
было.
подтверждало полученные ранее данные in vitro
Насколько нам известно, к настоящему мо
(клетки THP 1) и in vivo (тонкий кишечник, пе
менту не опубликовано работ о кинетике
чень и почки) [42-44]. Несмотря на функцио
экспрессии большинства транспортеров цинка
нально противоположные роли в поддержании
в мышцах человека (преобладающий резервуар
гомеостаза цинка в презентируемых тканях
Zn) в ответ на флуктуации уровня цинка и усло
(THP 1 - моноциты и клетки RD - мышцы), их
вия воспаления. Экспрессия металлотионеина
общий способ регуляции через MTF 1 опосре
MT, цинк запасающего белка, зависит от уровня
дованную активацию в присутствии или отсут
цинка и может оказывать влияние на множество
ствии цинка [40, 41] предполагает наличие об
внутриклеточных процессов [37, 38]. В клетках
щего способа оценки уровня цинка в организме
THP 1 и RD было выявлено значительное по
в целом. Поэтому мы предположили, что ZnT1
вышение уровня MT (в 22-25 раз в клетках
является одним из основных транспортеров
THP 1 и RD) в ответ на добавление цинка. Эти
цинка, регулирующих содержание цинка в мы
результаты согласуются с ранее полученными
шечных клетках и в моноцитах, что согласуется
данными in vitro исследований касательно отве
с его повсеместной экспрессией в качестве эф
та THP 1 на избыток цинка [39].
флюксного транспортера в плазматической
Металлозависимый фактор транскрипции
мембране различных типов клеток [45]. Отсут
MTF играет важную роль в активации тран
ствие реакции некоторых из белков ZnTs и ZIPs
скрипции MT и ZnT1 в ответ на металлы
на добавление Zn может быть связано с их
[40, 41]. Поэтому, как и ожидалось, в клетках
ролью в гомеостазе Zn. Например, ZnT4 и ZnT6
THP 1 и RD при достаточности цинка значи
переносят Zn из внутриклеточных компартмен
Рис. 2. Экспрессия генов транспортеров цинка в клетках THP 1 и RD. Тотальная РНК была выделена и затем синтезиро
вана кДНК. Определение относительного количества мРНК проводили методом qPCR с использованием интеркалирую
щего красителя SYBR Green. мРНК нормализовали относительно мРНК β актина и ГАФДГ. Клетки THP 1: а - уровни
мРНК MT, hZnTs; б - уровни мРНК hZIPs. Клетки RD: в - уровни мРНК MT, hZnTs; г - уровни мРНК hZIPs. Клетки
были обработаны 25 мкМ цинка в течение 4 ч. Столбцы представляют среднее значение ± SE трех повторов (n = 3) трех
независимых экспериментов. Верхние индексы означают значимые различия при p < 0,05. Достоверность различий меж
ду обработанными и контрольными клетками анализировали с помощью метода ANOVA и последующего множественно
го сравнения по критерию Даннета (Dunnett’s t test)
БИОХИМИЯ том 86 вып. 2 2021
206
АЛЛУРИ и др.
Рис. 3. Вестерн блоттинг для определения общего уровня транспортеров ZnT1 в клетках THP 1 и RD после добавления
или удаления цинка на 4 ч. а - Клетки THP 1; б - клетки RD; обрабатывали цинком или TPEN (хелатор цинка).
Control - контроль, Zinc - добавление 25 мкМ цинка, TPEN - удаление цинка путем внутриклеточного хелатирования.
ГАФДГ использовалась в качестве внутреннего контроля в случае клеток RD, а β актин -для клеток THP 1 для нормали
зации содержания белка
тов на периферию при достаточно высокой кон
казано значительное повышение экспрессии
центрации Zn [46]. Также был показан более вы
ZIP9 (в 1,81 раз), ZIP10 (в 4,7 раза) и ZIP14
раженный ответ ZIP4 на недостаток Zn в срав
(~4,9
раз) и снижение экспрессии ZIP13
нении с его достаточным уровнем [46].
(в 0,3 раза) в клетках THP 1 в условиях дефицита
Кроме того, результаты настоящей работы
цинка (рис. 4, б). В то же время недостаток Zn не
согласуются с результатами Overbeck et al., кото
оказывал заметного влияния на экспрессию
рые наблюдали низкий уровень экспрессии бел
ZnT4, ZnT7, ZnT9, а также ZIP1, ZIP3, ZIP6,
ка ZnT4 и отсутствие экспрессии ZnT2, ZnT3 и
ZIP7 и ZIP8. С другой стороны, в условиях дефи
ZnT8 в клетках THP 1, культивированных с
цита цинка в клетках THP 1 экспрессия ZnT2,
сульфатом цинка [43]. В то время как никаких
ZnT3, ZnT8, ZnT10, ZIP2, ZIP4, ZIP5 и ZIP12 не
значительных изменений уровня мРНК ZnT5 в
детектировалась. В случае ZIP2, полученные на
клетках THP 1 в присутствии цинка отмечено
ми результаты противоречат ранее полученным
не было, все же было зарегистрировано сниже
данным о важной роли этого белка в Zn дефи
ние уровня белка [43]. Известно, что MTF 1 иг
цитных клетках THP 1, что может быть связано с
рает двойственную роль, активирующую или
методологическими различиями в инкубации с
ингибирующую, в зависимости от уровня Zn.
5
мкМ TPEN в течение
18
ч или же с
Он снижает экспрессию ZIP10 в условиях доста
16 мкМ TPEN в течение 4 ч для создания условий
точности Zn и повышает её при дефиците Zn
внутриклеточной недостаточности цинка [35, 39].
[47]. Тем не менее при добавлении цинка к клет
Аналогично в ответ на дефицит цинка клет
кам экспрессия ZIP10 в клетках THP 1 сущест
ки RD отвечали значительным снижением
венно не изменялась, в то время как в клетках
экспрессии MT (в 0,5 раза) и ZnT1 (в 0,3 раза)
RD было зафиксировано ее снижение, что подт
(рис. 4, в), что было подтверждено уровнем
верждает данные работы Ryu et al. (2008) [48].
экспрессии белков (рис. 3, б). Однако в этих ус
Подавление экспрессии ZIP10 в условиях доста
ловиях экспрессия ZnT5, ZnT7, ZIP6, ZIP8,
точности Zn может быть связано с механизмом
ZIP10, ZIP13 и ZIP14 значительно повышалась
регуляции, включающим связывание MTF 1 с
в 1,9; 1,5; 1,5; 1,4; 2,5; 1,7 и 1,6 раза соответ
металл респонсивными элементами (MRE), в
ственно по сравнению с контролем (рис. 4, в, г).
ходе которого MTF 1 приостанавливает тран
В отличие от клеток THP 1, в условиях дефици
скрипцию Pol II [46].
та цинка в клетках RD повышалась экспрессия
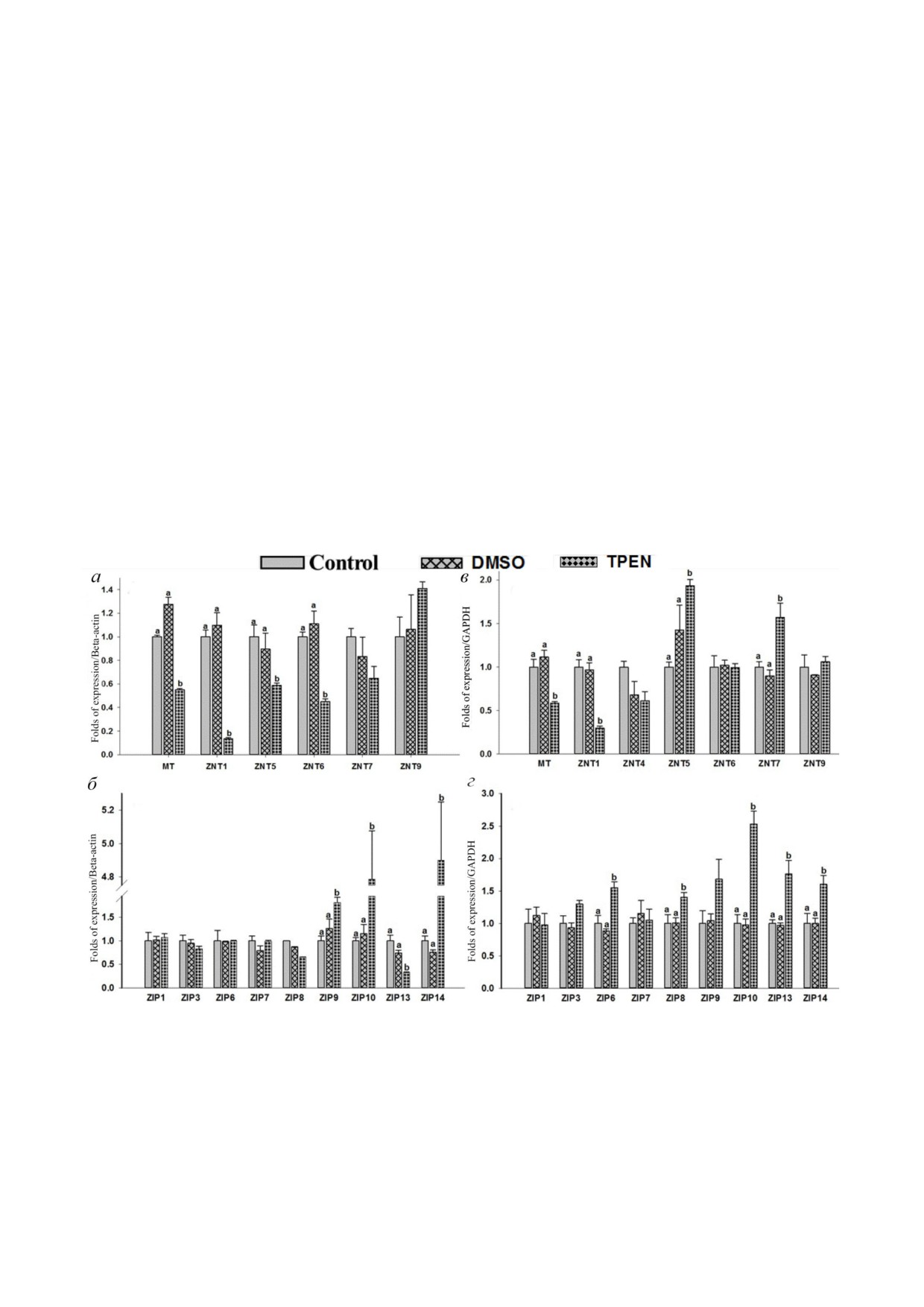
Влияние дефицита цинка на экспрессию MT,
ZnT5. Тем не менее дефицит Zn не оказывал за
ZnTs и ZIPs в клетках THP 1 и RD. Элиминация
метного влияния на паттерны экспрессии дру
внутриклеточного цинка с помощью TPEN при
гих белков ZnTs (ZnT4, ZnT6, ZnT9) и ZIPs
водила к значительному снижению экспрессии
(ZIP1, ZIP3, ZIP7). Также наблюдалось некото
MT (в 0,55 раза), ZnT1 (в 0,13 раз), ZnT5
рое повышение экспрессии ZIP9, но оно не бы
(в 0,58 раз) и ZnT6 (в 0,44 раза) в клетках THP 1
ло статистически достоверным. Экспрессия
в сравнении с контролем (рис. 4, а). Уровень син
восьми транспортеров цинка не детектирова
теза мРНК коррелировал с уровнем белка ZnT1,
лась (ZnT2, ZnT3, ZnT8, ZnT10 и ZIP2, ZIP4,
определяемым методом вестерн блоттинга
ZIP5, ZIP12). Таким образом, в клетках THP 1 и
(рис. 3, а). Данные по снижению экспрессии MT,
RD в условиях дефицита Zn транспортеры ZIPs
ZnT1, ZnT5 и ZnT6 подтверждают ранее полу
экспрессируются в различной степени.
ченные результаты на клетках THP 1 в условиях
Было показано, что двойственная роль
дефицита цинка [39, 49]. В то же время было по
MTF 1 является причиной снижения экспрес
БИОХИМИЯ том 86 вып. 2 2021
РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ ТРАНСПОРТЕРОВ ЦИНКА
207
сии ZIP10 при избытке цинка и повышения его
Влияние Tat индуцированного воспаления на
экспрессии в условиях дефицита цинка [47]. На
MHC I и внутриклеточный TNF α. С помощью
ши данные по ZIP10 в клетках THP 1 и RD со
метода флуоресцентно активированной сорти
гласуются с этими наблюдениями: экспрессия
ровки клеток (FACS) в совместно культивиро
ZIP10 в клетках THP 1 и RD в условиях дефици
ванных клетках THP 1 и RD оценивался эффект
та цинка повышалась в 4,7 и 2,5 раза соответ
Tat (100 нг/мл) на MHC I. Было обнаружено,
ственно [46, 47]. Среди разнообразных эффлюкс
что уровни экспрессии MHC I в присутствии
ных и инфлюксных транспортеров цинка MT,
Tat остаются без изменений в обоих типах кле
ZnT1 и ZIP10, вероятно, регулируются сходны
ток по сравнению с соответствующими контро
ми механизмами в условиях избытка и недостат
лями (рис. 5). В то же время в клетках THP 1
ка цинка в моноцитах и мышечных клетках. При
значительно повышался уровень внутриклеточ
этом мы наблюдали противоположные паттерны
ного TNF α (~3,6 раза) в присутствии Tat
экспрессии ZIP13 и ZIP14 в клетках THP 1 и RD
(100 нг/мл) в сравнении с необработанными
в условиях дефицита цинка. С помощью нокаута
контрольными клетками (рис. 6).
генов Zip13 и Zip14 у мышей было показано, что
Для понимания эффекта воспаления на
эти транспортеры Zn регулируют Zn опосредо
транспортеры цинка мы провели совместное
ванные сигнальные пути, и поэтому аномальная
культивирование клеток THP 1 и мышечных
их экспрессия вызывала нарушения роста и го
клеток RD для стимулирования перекрестных
меостаза костей [46]. Эти данные способствуют
связей между репрезентативными тканями и об
понимаю роли MTF 1 в регуляции MT, ZnT1 и
работали их пептидами HIV 1 Tat в течение 8 ч.
ZIPs (ZIP10, ZIP13 и ZIP14) в ответ на измене
В предыдущих работах по изучению воспали
ния уровня Zn в клетках THP 1 и RD.
тельного ответа клеток THP 1 на обработку бел
Рис. 4. Экспрессия генов транспортеров цинка в ответ на обработку TPEN в клетках THP 1 и RD. Выделяли препарат то
тальной РНК и синтезировали кДНК. Определение относительного количества мРНК проводили методом qPCR с ис
пользованием интеркалирующего красителя SYBR Green. мРНК нормализовали относительно мРНК β актина и ГАФДГ.
Уровни экспрессии мРНК в клетках, обработанных 5 мкМ TPEN: а - уровни мРНК MT и hZnTs в клетках THP 1;
б - уровни мРНК hZIPs в клетках THP 1; в - уровни мРНК MT и hZnTs в клетках RD; г - уровни мРНК hZIPs в клетках
RD. Столбцы представляют среднее значение ± SE трех повторов (n = 3) трех независимых экспериментов. Верхние ин
дексы означают значимые различия при p < 0,05. Достоверность различий между обработанными и контрольными клет
ками анализировали с помощью метода ANOVA и последующего множественного сравнения по критерию Даннета
(Dunnett’s t test)
БИОХИМИЯ том 86 вып. 2 2021
208
АЛЛУРИ и др.
Рис. 5. Экспрессия MHC I в обработанных Tat клетках RD и THP 1. Совместно культивируемые мышечные клетки RD
(а) и моноциты THP 1 (б) были обработаны набором пептидов HIV 1 Tat в течение 8 ч, и экспрессия MHC I оценивалась
с помощью проточной цитометрии. Данные показаны на гистограмме: клетки, культивированные в обычной среде (конт
рольная гистограмма) с наложенной гистограммой для экспериментальных клеток, обработанных Tat (показаны различ
ными цветами)
ком Tat (100 нМ в течение 12 ч) было показано
повышение уровня TNF α [50]. Наши данные
также демонстрируют значительное повышения
уровня внутриклеточного TNF α в совместно
культивированных клетках THP 1, подвергших
ся воздействию белка Tat.
Влияние Tat на экспрессию транспортеров цин
ка в совместно культивированных клетках THP 1.
В клетках THP 1, культивированных совместно
с клетками RD, был зарегистрирован воспали
тельный ответ на Tat в виде повышенного в
1,4 раза уровня IL 1β (рис. 7, а). В то же время
различий в уровне IL 6 в сокультивируемых
THP 1 по сравнению контрольными сокульти
вируемыми клетками не было. В работах по изу
чению влияния белка Tat на макрофаги при об
работке им в течение 4 ч было показано повыше
ние уровней IL 6 и IL 1β [51]. В настоящей рабо
те была продемонстрирована значительная
Рис. 6. Экспрессия TNF α в обработанных Tat клетках
THP 1. Совместно культивируемые клетки THP 1 обраба
экспрессия IL 1β в клетках THP 1, обработан
тывали набором пептидов Tat в концентрации 100 нг/мл в ных Tat; однако для IL 6 такого эффекта обнару
течение 8 ч. Экспрессию TNF α определяли с помощью
жено не было. Tat индуцированное воспаление
проточной цитометрии с окрашиванием внутриклеточного
цитокина TNF α. Данные показаны на гистограмме: конт
приводило к значительному (p < 0,05) снижению
рольная гистограмма показана синим цветом; клетки, об
(в 0,5 раза) уровня транспортеров цинка ZIP7 и
работанные Tat, показаны красным цветом. Сдвиг означа
ZIP14 в совместно культивированных клетках
ет повышение экспрессии TNF α. Клетки, культивиро
THP 1 по сравнению с необработанными конт
ванные в обычной среде (контрольная гистограмма синего
цвета) с наложенной гистограммой для эксперименталь
рольными клетками (рис. 7, б). Эти результаты
ных клеток, обработанных Tat (показана красной линией)
согласуются с ответом дендритных клеток (DCs)
БИОХИМИЯ том 86 вып. 2 2021
РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ ТРАНСПОРТЕРОВ ЦИНКА
209
мыши на обработку липополисахаридом (LPS),
результате транспортировки цинка в печень
при которой происходит снижение экспрессии
[54, 55]. Согласно литературным данным, сти
ZIP6 и ZIP10, в то время как экспрессия ZnT1,
муляция воспаления посредством LPS в денд
ZnT4 и ZnT6 повышается, вызывая снижение
ритных клетках приводит к снижению экспрес
содержания цинка в цитозоле [52]. В сокульти
сии ZIP6 и повышению MHC II [56]. Таким об
вированных с RD клетках THP 1 после обработ
разом, полученные нами данные об отсутствии
ки белком Tat наблюдалось повышение уровня
изменений уровня MHC I могут быть обуслов
ZIP9, однако оно не было статистически досто
лены неизменным уровнем ZIP6 при воздей
верным. При этом значительных изменений
ствии Tat. Кроме того, очевидно, что дендрит
уровней MT, ZnT1, ZnT4-ZnT7, ZnT9, ZIP1,
ные клетки с повышенной экспрессией ZIP6 не
ZIP3, ZIP6, ZIP8, ZIP10 и ZIP 13 в сокультиви
способны активировать антиген специфичные
рованных с RD клетках THP 1, обработанных
клетки CD4+ Th [52].
белком Tat, не наблюдалось.
Известно, что ZIP14 локализуется на плаз
Аналогично влияние Tat на экспрессию бел
матической мембране и импортирует цинк в ци
ков ZIPs (инфлюксные транспортеры цинка)
топлазму из внеклеточной среды. В настоящей
предполагает значительное снижение экспрес
работе было показано, что экспрессия ZIP14 в
сии ZIP7 и ZIP14 в совместно культивирован
ответ на белок HIV 1 Tat снижается. Мы пред
ных клетках THP 1. Эти данные согласуются с
полагаем, что снижение экспрессии ZIP14 мо
предыдущими исследованиями действия воспа
жет способствовать сохранению уровня сыворо
ления на клетки эпителия легких [53]. Напротив
точного цинка, что может благоприятствовать
же, инфекция и воспаление вызывают повыше
инфицированию ВИЧ. С другой стороны, не
ние экспрессии ZIP14 посредством IL 6/IL 1β,
достаточное количество цинка в клетках THP 1
приводящее к возникновению гипоцинкемии в
приводит к запуску апоптоза [57, 58].
Рис. 7. Экспрессия генов в сокультивируемых клетках THP 1 и RD. После выделения РНК определяли относительное ко
личество мРНК с использованием ПЦР в реальном времени с окрашиванием SYBR Green. Уровни мРНК нормализовали
относительно мРНК β актина и ГАФДГ. Уровни экспрессии мРНК в клетках, обработанных Tat (100 нг/мл) в течение 8 ч:
а - уровни экспрессии белков hZnTs в клетках THP 1; б - белков hZIPs в клетках THP 1; в - белков hZnTs в клетках RD;
г - белков hZIPs в клетках RD. Столбцы представляют среднее значение ± SE (n = 3). Верхние индексы обозначают ста
тистически достоверные различия при p < 0,05 с использованием парного независимого критерия
5 БИОХИМИЯ том 86 вып. 2 2021
210
АЛЛУРИ и др.
Влияние Tat на экспрессию транспортеров
точного цинка путем скоординированной регу
цинка в совместно культивированных клетках
ляции MT и транспортеров цинка и совместно
RD. В совместно культивированных с THP 1
го действия для модуляции транзиторных изме
клетках RD наблюдается значительное
нений уровня цинка. Продемонстрировано, что
(p < 0,05) повышение уровня IL 6 (в 2 раза) в
ZIP10 подвергается дифференциальному регу
сравнении с необработанными контрольными
лированию в мышцах, так как его уровень сни
клетками, что предположительно свидетель
жался при избытке цинка, но оставался неиз
ствует о провоспалительном ответе на Tat
менным в клетках THP 1 в тех же условиях.
(рис. 7, в). В то же время уровни IL 1β в сов
HIV 1 Tat опосредованное воспаление вызыва
местно культивированных клетках RD, обрабо
ло снижение экспрессии некоторых транспор
танных и не обработанных пептидами Tat, от
теров цинка, например, ZIP7 и ZIP14, особенно
носительно одинаковы. Ответ металлотионеи
в клетках THP 1, сокультивируемых с клетками
на (MT) на обработку Tat заключался в значи
RD, в то время как в мышечных клетках RD
тельном повышении (в 2,3 раза) его содержа
уровни белков ZIPs были повышены. Это может
ния в совместно культивированных клетках RD
указывать на перенаправление цинка в мышцы
по сравнению с контрольными необработан
и работать таким образом в качестве пассивного
ными клетками. Ранее было показано, что
резервуара цинка. Снижение количества моно
Tat индуцированные воспалительные ответы
цитов у больных ВИЧ/СПИД, вероятно, опос
являются тканеспецифичными, и дифферен
редованно снижением уровня внутриклеточно
циальная экспрессия была подтверждена пат
го цинка, что приводит к запуску апоптоза в мо
терном экспрессии IL 6: в мононуклеарных
ноцитах. Несмотря на то что результаты настоя
клетках периферической крови PBMCs - ми
щей работы основаны на модели клеточных ли
нимальный ответ, в THP 1 - слабый и в астро
ний, это может помочь нам понять природу раз
цитах - сильный ответ на воздействие пепти
личных ответов клеток на уровень цинка и вос
дов Tat [59].
паление.
Аналогично значительно повышался уро
вень транспортеров цинка ZnT5 (в 1,6 раза),
Финансирование. Выполнение данной рабо
ZIP7 (в 1,7 раза), ZIP8 (в 2,6 раза) и ZIP9 (в
ты проходило при поддержке грантов, предо
1,6 раза) в совместно культивированных клетках
ставленных К. П. М. Наиру, и гранта, выделен
RD в ответ на обработку Tat (рис. 7, г). В то же
ного С. Гошу (5/9/1137/2014 NUT) Индий
время заметных изменений уровня других бел
ским советом по медицинским исследованиям
ков ZnTs (ZnT1, ZnT4, ZnT6, ZnT7, ZnT9) и
(Indian Council of Medical Research), Индия. Ав
ZIPs (ZIP1, ZIP3, ZIP6, ZIP10, ZIP13, ZIP14) в
торы выражают благодарность д ру Б. Динешу
обработанных Tat клетках RD после их сокуль
Кумару, научному сотруднику G & Head DTRC,
тивирования с клетками THP 1 зарегистрирова
за совместное использование оборудования и
но не было (рис. 7, в и г).
реагентов из проекта ICMR Taskforce № HIV/
Влияние белка Tat на экспрессию белков
62/47/2016 ECD II.
ZIPs, инфлюксных транспортеров цинка, сви
Конфликт интересов. Авторы заявляют об от
детельствует о значительном снижении
сутствии конфликта интересов.
экспрессии ZIP7 и ZIP14 в совместно культиви
Благодарности. Авторы выражают благодар
рованных клетках THP 1, в то время как в клет
ность CSIR - New Delhi (Индия) за финансовую
ках RD было зафиксировано повышение
поддержку, оказанную Кирану Аллури для JRF и
экспрессии ZIP7, ZIP8 и ZIP9. Возможно, мы
SRF (09/484/(0050)/2012 EMR 1).
шечные клетки дают сигнал моноцитам снижать
Соблюдение этических норм. В данной статье
экспрессию белков ZIPs, чтобы они могли за
не содержатся результаты работ с участием лю
пасти больше цинка. Однако настоящая работа
дей или лабораторных животных, выполненных
ограничена отсутствием данных по транскрип
кем либо из авторов статьи.
ционной регуляции MTF 1 и маркеров апопто
Дополнительные материалы. Приложение к
за, и потому мы не можем проверить наше пред
статье на английском языке опубликовано на
положение.
сайте журнала
«Biochemistry» (Moscow)
Похоже, что два функционально противопо
ложных типа клеток, RD (мышцы) и THP 1 (мо
ноциты), поддерживают гомеостаз внутрикле
journal/10541), том 86, вып. 2, 2021.
БИОХИМИЯ том 86 вып. 2 2021
РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ ТРАНСПОРТЕРОВ ЦИНКА
211
СПИСОК ЛИТЕРАТУРЫ
1.
Kambe, T., Tsuji, T., Hashimoto, A., and Itsumura, N.
plementation to prevent immunological failure in HIV
(2015) The physiological, biochemical, and molecular
infected adults, Clin. Infect. Dis., 50, 1653 1660.
roles of zinc transporters in zinc homeostasis and metabo
16.
Tang, A. M., Graham, N. M., Kirby, A. J., McCall, L. D.,
lism, Physiol. Rev., 95, 749 784.
Willett, W. C., and Saah, A. J. (1993) dietary micronutrient
2.
Jackson, M. (1989) Physiology of Zinc: General Aspects,
intake and risk of progression to Acquired
Zinc in Human Biology, pp. 1 14.
Immunodeficiency Syndrome (AIDS) in Human
3.
Beck, F. W., Kaplan, J., Fine, N., Handschu, W., and
Immunodeficiency Virus Type 1 (HlV 1) infected homo
Prasad, A. S. (1997) Decreased expression of CD73 (ecto 5′
sexual men, Am. J. Epidemiol., 138, 937 951.
nucleotidase) in the CD8+ subset is associated with zinc defi
17.
Lee, S. P., and Han, M. K. (1996) Zinc stimulates Mg2+
ciency in human patients, J. Lab. Clin. Med., 130, 147 156.
dependent 3′ processing activity of human immunodefi
4.
Prasad, A. S. (2008) Zinc in human health: effect of zinc
ciency virus type 1 integrase in vitro, Biochemistry, 35,
on immune cells, Mol. Med., 14, 353 357.
3837 3844.
5.
Giacconi, R., Malavolta, M., Costarelli, L., Busco, F.,
18.
Rice, W. G., Schaeffer, C. A., Harten, B., Villinger, F.,
Galeazzi, R., et al. (2012) Comparison of intracellular zinc
South, T. L., et al. (1993) Inhibition of HIV 1 infectivity by
signals in nonadherent lymphocytes from young adult and
zinc ejecting aromatic C nitroso compounds, Nature, 361,
elderly donors: role of zinc transporters (Zip family) and
473 475.
proinflammatory cytokines, J. Nutr. Biochem., 23, 1256
19.
Reid, W., Sadowska, M., Denaro, F., Rao, S., Foulke, J., et
1263.
al. (2001) An HIV 1 transgenic rat that develops HIV
6.
Mao, X., Kim, B. E., Wang, F., Eide, D. J., and Petris, M. J.
related pathology and immunologic dysfunction, Proc.
(2007) A histidine rich cluster mediates the ubiquitination
Natl. Acad. Sci. USA, 98, 9271 9276.
and degradation of the human zinc transporter, hZIP4, and
20.
Reid, W., Abdelwahab, S., Sadowska, M., Huso, D., Neal, A.,
protects against zinc cytotoxicity, J. Biol. Chem., 282,
et al. (2004) HIV 1 transgenic rats develop T cell abnor
6992 7000.
malities, Virology, 321, 111 119.
7.
Weaver, B. P., and Andrews, G. K. (2012) Regulation of
21.
Purvis, S. F., Jacobberger, J. W., Sramkoski, R. M., Patki,
zinc responsive Slc39a5 (Zip5) translation is mediated by
A. H., and Lederman, M. M. (1995) HIV type 1 Tat pro
conserved elements in the
3′ untranslated region,
tein induces apoptosis and death in Jurkat cells, AIDS Res.
Biometals, 25, 319 335.
Hum. Retroviruses, 11, 443 450.
8.
Mocchegiani, E., Giacconi, R., Cipriano, C., and
22.
Bettaccini, A. A., Baj, A., Accolla, R. S., Basolo, F., and
Malavolta, M. (2009) NK and NKT cells in aging and
Toniolo, A. Q. (2005) Proliferative activity of extracellular
longevity: role of zinc and metallothioneins, J. Clin.
HIV 1 Tat protein in human epithelial cells: expression pro
Immunol., 29, 416 425.
file of pathogenetically relevant genes, BMC Microbiol., 5, 20.
9.
Nishida, K., Hasegawa, A., Nakae, S., Oboki, K., Saito, H.,
23.
Frankel, A. D., Bredt, D. S., and Pabo, C. O. (1988) Tat
et al. (2009) Zinc transporter Znt5/Slc30a5 is required for
protein from human immunodeficiency virus forms a
the mast cell mediated delayed type allergic reaction but
metal linked dimer, Science, 240, 70 73.
not the immediate type reaction, J. Exp. Med., 206, 1351
24.
Canani, R. B., Ruotolo, S., Buccigrossi, V., Passariello, A.,
1364.
Porcaro, F., Siani, M. C., and Guarino, A. (2007) Zinc
10.
Liu, M. J., Bao, S., Gálvez Peralta, M., Pyle, C. J.,
fights diarrhoea in HIV 1 infected children: in vitro evi
Rudawsky, A. C., et al. (2013) The zinc transporter
dence to link clinical data and pathophysiological mecha
SLC39A8 is a negative feedback regulator of NF κB
nism, AIDS, 21, 108 110.
through zinc mediated inhibition of IKK, Cell Rep., 3,
25.
Li, J. C., Yim, H. C., and Lau, A. S. (2010) Role of
386.
HIV 1 Tat in AIDS pathogenesis: its effects on cytokine
11.
Taniguchi, M., Fukunaka, A., Hagihara, M., Watanabe,
dysregulation and contributions to the pathogenesis of
K., Kamino, S., Kambe, T., Enomoto, S., and Hiromura,
opportunistic infection, AIDS, 24, 1609 1623.
M.
(2013) Essential role of the zinc transporter
26.
Matsui, M., Warburton, R. J., Cogswell, P. C., Baldwin, A.
ZIP9/SLC39A9 in regulating the activations of Akt and
S., Jr., and Frelinger, J. A. (1996) Effects of HIV 1 Tat on
Erk in B cell receptor signaling pathway in DT40 cells,
expression of HLA class I molecules, J. Acquir. Immune
PLoS One, 8, e58022.
Defic. Syndr. Hum. Retrovirol., 11, 233 240.
12.
Haraguchi, Y., Sakurai, H., Hussain, S., Anner, B. M., and
27.
Gherardi, R. K. (1994) Skeletal muscle involvement in
Hoshino, H. (1999) Inhibition of HIV 1 infection by zinc
HIV infected patients, Neuropathol. Appl. Neurobiol., 20,
group metal compounds, Antiviral Res., 43, 123 133.
232 237.
13.
Mocchegiani, E., and Muzzioli, M. (2000) Therapeutic
28.
Illa, I., Nath, A., and Dalakas, M. (1991) Immunocyto
application of zinc in human immunodeficiency virus
chemical and virological characteristics of HIV associated
against opportunistic infections, J. Nutr., 130, 1424S
inflammatory myopathies: similarities with seronegative
1431S.
polymyositis, Ann. Neurol., 29, 474 481.
14.
Bobat, R., Coovadia, H., Stephen, C., Naidoo, K. L.,
29.
Belec, L., Meillet, D., Hernvann, A., Gresenguet, G., and
McKerrow, N., Black, R. E., and Moss, W. J. (2005) Safety
Gherardi, R. (1994) Differential elevation of circulating
and efficacy of zinc supplementation for children with
interleukin 1 beta, tumor necrosis factor alpha, and inter
HIV 1 infection in South Africa: a randomised double
leukin 6 in AIDS associated cachectic states, Clin. Diagn.
blind placebo controlled trial, Lancet, 366, 1862 1867.
Lab. Immunol., 1, 117 120.
15.
Baum, M. K., Lai, S., Sales, S., Page, J. B., and Campa, A.
30.
Llovera, M., Garcia Martinez, C., Agell, N., Lopez
(2010) Randomized, controlled clinical trial of zinc sup
Soriano, F. J., Authier, F. J., Gherardi, R. K., and Argiles,
БИОХИМИЯ том 86 вып. 2 2021
5*
212
АЛЛУРИ и др.
J. M. (1998) Ubiquitin and proteasome gene expression is
45.
Palmiter, R. D., and Findley, S. D. (1995) Cloning and
increased in skeletal muscle of slim AIDS patients, Int. J.
functional characterization of a mammalian zinc trans
Mol. Med., 2, 69 73.
porter that confers resistance to zinc, EMBO J., 14, 639
31.
Gonzalez Cadavid, N. F., Taylor, W. E., Yarasheski, K.,
649.
Sinha Hikim, I., Ma, K., et al. (1998) Organization of the
46.
Hara, T., Takeda, T. A., Takagishi, T., Fukue, K., Kambe, T.,
human myostatin gene and expression in healthy men and
and Fukada, T. (2017) Physiological roles of zinc trans
HIV infected men with muscle wasting, Proc. Natl. Acad.
porters: molecular and genetic importance in zinc homeo
Sci. USA, 95, 14938 14943.
stasis, J. Physiol. Sci., 67, 283 301.
32.
Tibaduiza, E. C., and Bobilya, D. J. (1996) Zinc transport
47.
Lichten, L. A., Ryu, M. S., Guo, L., Embury, J., and
across an endothelium includes vesicular cotransport with
Cousins, R. J. (2011) MTF 1 mediated repression of the
albumin, J. Cell. Physiol., 167, 539 547.
zinc transporter Zip10 is alleviated by zinc restriction,
33.
Sreenivasulu, K., Raghu, P., and Nair, K. M.
(2010)
PLoS One, 6, e21526.
Polyphenol rich beverages enhance zinc uptake and metal
48.
Ryu, M. S., Lichten, L. A., Liuzzi, J. P., and Cousins, R.
lothionein expression in Caco 2 cells, J. Food Sci., 75,
J.
(2008) Zinc transporters ZnT1 (Slc30a1), Zip8
H123 H128.
(Slc39a8), and Zip10 (Slc39a10) in mouse red blood cells
34.
Coyle, P., Zalewski, P. D., Philcox, J. C., Forbes, I. J.,
are differentially regulated during erythroid development
Ward, A. D., et al. (1994) Measurement of zinc in hepato
and by dietary zinc deficiency, J. Nutr., 138, 2076 2083.
cytes by using a fluorescent probe, zinquin: relationship to
49.
Hamon, R., Homan, C. C., Tran, H. B., Mukaro, V. R.,
metallothionein and intracellular zinc, Biochem. J., 303,
Lester, S. E., et al. (2014) Zinc and zinc transporters in
781 786.
macrophages and their roles in efferocytosis in COPD,
35.
Chen, L., Frister, A., Wang, S., Ludwig, A., Behr, H., et al.
PLoS One, 9, e110056.
(2009) Interaction of vascular smooth muscle cells and
50.
Chen, P., Mayne, M., Power, C., and Nath, A. (1997) The
monocytes by soluble factors synergistically enhances
Tat protein of HIV 1 induces Tumor Necrosis Factor α
interleukin 6 and MCP 1 production, Am. J. Physiol.
production implications for HIV 1 associated neurological
Heart Circ. Physiol., 296, H987 H996.
diseases, J. Biol. Chem., 272, 22385 22388.
36.
Alluri, K., Nair, K. P., Kotturu, S. K., and Ghosh, S.
51.
Nath, A., Conant, K., Chen, P., Scott, C., and Major, E. O.
(2020) Transcriptional regulation of zinc transporters in
(1999) Transient exposure to HIV 1 Tat protein results in
human osteogenic sarcoma (Saos 2) cells to zinc supple
cytokine production in macrophages and astrocytes A hit
mentation and zinc depletion, Biol. Trace Elem. Res., 194,
and run phenomenon, J. Biol. Chem., 274, 17098 17102.
360 367.
52.
Hojyo, S., and Fukada, T. J. (2016) Roles of zinc signaling
37.
Davis, S. R., and Cousins, R. J. (2000) Metallothionein
in the immune system, Immunol. Res., 2016, 6762343,
expression in animals: a physiological perspective on func
doi: 10.1155/2016/6762343.
tion, J. Nutr., 130, 1085 1088.
53.
Lang, C. J., Murgia, C., Leong, M., Tan, L. W., Perozzi,
38.
Alluri, K., Nair, K. P., and Ghosh, S. (2019) Differential
G., et al. (2006) Anti inflammatory effects of zinc and
expression of zinc transporters in functionally contrasting
alterations in zinc transporter mRNA in mouse models of
tissues involved in zinc homeostasis, Nucleosides
allergic inflammation, Am. J. Physiol. Lung Cell. Mol.
Nucleotides Nucleic Acids, 18, 1 5.
Physiol., 292, L577 L584.
39.
Cao, J., Bobo, J. A., Liuzzi, J. P., and Cousins, R. J. (2001)
54.
Liuzzi, J. P., Lichten, L. A., Rivera, S., Blanchard, R. K.,
Effects of intracellular zinc depletion on metallothionein
Aydemir, T. B., et al. (2005) Interleukin 6 regulates the
and ZIP2 transporter expression and apoptosis, J. Leukoc.
zinc transporter Zip14 in liver and contributes to the
Biol., 70, 559 566.
hypozincemia of the acute phase response, Proc. Natl.
40.
Andrews, G. K. (2000) Regulation of metallothionein gene
Acad. Sci. USA, 102, 6843 6848.
expression by oxidative stress and metal ions, Biochem.
55.
Lichten, L. A., Liuzzi, J. P., and Cousins, R. J. (2009)
Pharmacol., 59, 95 104.
Interleukin 1β contributes via nitric oxide to the upregula
41.
Langmade, S. J., Ravindra, R., Daniels, P. J., and
tion and functional activity of the zinc transporter Zip14
Andrews, G. K. (2000) The transcription factor MTF 1
(Slc39a14) in murine hepatocytes, Am. J. Physiol.
mediates metal regulation of the mouse ZnT1 gene, J. Biol.
Gastroint. Liver Physiol., 296, G860 G867.
Chem., 275, 34803 34809.
56.
Kitamura, H., Morikawa, H., Kamon, H., Iguchi, M.,
42.
Cousins, R. J., Blanchard, R. K., Popp, M. P., Liu, L.,
Hojyo, S., et al. (2006) Toll like receptor mediated regula
Cao, J., et al. (2003) A global view of the selectivity of zinc
tion of zinc homeostasis influences dendritic cell function,
deprivation and excess on genes expressed in human
Nat. Immunol., 7, 971 977.
THP 1 mononuclear cells, Proc. Natl. Acad. Sci. USA, 100,
57.
Li, C. J., Friedman, D. J., Wang, C., Metelev, V., and
6952 6957.
Pardee, A. B. (1995) Induction of apoptosis in uninfected
43.
Overbeck, S., Uciechowski, P., Ackland, M. L., Ford, D.,
lymphocytes by HIV 1 Tat protein, Science, 268, 429 431.
and Rink, L. (2008) Intracellular zinc homeostasis in
58.
Chen, D., Wang, M., Zhou, S., and Zhou, Q. (2002) HIV
leukocyte subsets is regulated by different expression of
1 Tat targets microtubules to induce apoptosis, a process
zinc exporters ZnT 1 to ZnT 9, J. Leuk. Biol., 83, 368
promoted by the pro apoptotic Bcl 2 relative Bim, EMBO
380.
J., 21, 6801 6810.
44.
Liuzzi, J. P., Blanchard, R. K., and Cousins, R. J. (2001)
59.
Joshi, P. C., and Guidot, D. M. (2011) HIV 1 transgene
Differential regulation of zinc transporter 1, 2, and 4
expression in rats induces differential expression of tumor
mRNA expression by dietary zinc in rats, J. Nutr., 131, 46
necrosis factor alpha and zinc transporters in the liver and
52.
the lung, AIDS Res. Ther., 8, 36.
БИОХИМИЯ том 86 вып. 2 2021
РЕГУЛЯЦИЯ ТРАНСКРИПЦИИ ТРАНСПОРТЕРОВ ЦИНКА
213
LEVELS OF ZINC TRANSPORTERS mRNA DEPENDING ON ZINC STATUS
AND HIV 1 TAT INDUCED INFLAMMATION IN MUSCLE (RD)
AND MONOCYTE (THP 1) CELL LINES*
K. Alluri1, S. Reddy Yathapu2, N. Babu Kondapalli3,
R. Hemalatha3, K. M. Nair4*, and S. Ghosh1**
1 Molecular Biology Division, ICMR National Institute of Nutrition,
500007 Hyderabad, India; e mail: bihongo@yahoo.com
2 Drug Toxicology Division, ICMR National Institute of Nutrition, 500007 Hyderabad, India
3 Microbiology and Immunology Division, ICMR National Institute of Nutrition, 500007 Hyderabad, India
4 Micronutrient Division, ICMR National Institute of Nutrition,
500007 Hyderabad, India; e mail: nairthayil@gmail.com
Monocytes and muscles demonstrate functionally contrasting behavior under conditions of zinc deficiency with rela
tion to zinc storage system (muscle retain zinc in contrast to monocytes). We aimed to understand the effects of zinc
status and HIV 1 Tat mediated inflammation on expression of zinc transporters in these types of cells. Expression of
zinc transporters [ZnTs, ZIPs, and metallothionein (MT)] was quantified by qRT PCR in RD, THP 1 cells sepa
rately and in co cultured THP 1-RD cells. ZnT1 protein expression levels were confirmed by Western blot.
Significant increase of MT and ZnT1 mRNA in response to zinc supplementation and decrease during zinc deficien
cy indicates significance of the genes encoding transporters in maintaining zinc homeostasis in these tissues. In the
RD cells ZIP10 exhibited inverse relation to zinc status whereas no correlation was found in the THP 1 cells. Tat
induced inflammation resulted in the significant elevation of MT, IL6, ZIP7, ZIP8, ZIP9 transcripts in the co cul
tured RD cells, whereas THP 1 cells demonstrated increased IL 1β levels and reduced levels of ZIP7 and ZIP14.
Zinc status and HIV 1Tat induced inflammation appear to influence differential expression of MT, ZnTs, and ZIPs
in the muscle and monocyte cells.
Keywords: zinc transporters, ZIPs, ZnTs, co culture, HIV 1 Tat, THP 1, RD
БИОХИМИЯ том 86 вып. 2 2021