БИОХИМИЯ, 2021, том 86, вып. 2, с. 214 - 227
УДК 615.31+616.43
ГЕПАТОПРОТЕКТОРНОЕ ДЕЙСТВИЕ НАГРУЖЕННЫХ
ПОЛИДАТИНОМ ХИТОЗАНОВЫХ НАНОЧАСТИЦ У КРЫС
С САХАРНЫМ ДИАБЕТОМ: МОДУЛЯЦИЯ МЕТАБОЛИЗМА
ГЛЮКОЗЫ, ОКИСЛИТЕЛЬНОГО СТРЕССА И БИОМАРКЕРОВ
ВОСПАЛИТЕЛЬНЫХ ПРОЦЕССОВ*
© 2021
А.М. Абд Эль#Хамид1, А.И. Юсеф2, С.М. Абд Эль#Тваб2,
А.А.Г. Эль#Шахави3, А. Абдель#Монеим2**
1 Chemistry Department, Faculty of Science, Taibah University, 30002 Al Madinah Al Munawarah, Saudi Arabia
2 Molecular Physiology Division, Faculty of Science, Beni Suef University,
62511 Beni Suef, Egypt; E mail: adel.hassan@science.bsu.edu.eg
3 Materials Science and Nanotechnology Department, Faculty of Postgraduate Studies for Advanced Sciences (PSAS),
Beni Suef University, 62511 Beni Suef, Egypt
Поступила в редакцию15.06.2020
После доработки 03.09.2020
Принята к публикации 09.09.2020
Полидатин (PD - polydatin) обладает широким набором фармакологической активности, однако его
действие при диабетическом поражении печени изучено мало. Целью настоящей работы было исследование
возможности защитного эффекта хитозановых наночастиц, нагруженных полидатином (PD$CSNPs - poly$
datin$loaded chitosan nanoparticles), или свободным полидатином при поражениях печени, ассоциированных
с диабетом. Диабет у крыс индуцировали при помощи никотинамида и стрептозоцина. Животные были раз$
делены на шесть групп: нормальный контроль, диабетический контроль, а также крысы, получавшие перо$
рально ежедневно в течение 4$х недель полидатин, хитозановые наночастицы с полидатином, ненагружен$
ные хитозановые наночастицы или метформин. Добавление в пищу полидатина и нагруженных полидати$
ном хитозановых наночастиц вызывало существенное снижение уровня глюкозы в крови и перекисного
окисления липидов в печени, снижение активности трансаминаз и ферментов метаболизма углеводов в сы$
воротке крови, в том числе сукцинатдегидрогеназы и пируваткиназы. В то же время в печени, напротив,
наблюдалось повышение уровня гликогена и глутатиона, а также активность антиоксидантных ферментов,
таких как супероксиддисмутаза, глутатионпероксидаза, каталаза и глюкозо$6$фосфатдегидрогеназа, в срав$
нении с контрольными диабетическими крысами. Кроме того, значительно снижалась экспрессия мРНК
фактора некроза опухолей α и интерлейкина 1β, в то время как уровни мРНК транспортера глюкозы 2 и глю$
кокиназы значительно повышались по сравнению с диабетическим контролем. На основании полученных
результатов мы заключили, что PD$CSNPs и PD оказывали положительное воздействие при диабетическом
поражении печени путем модуляции экспрессии транспортера глюкозы 2, влияющей на активность фер$
ментов метаболизма углеводов, а также подавляли окислительный стресс и воспаление. Было показано, что
PD$CSNPs являются более эффективным препаратом по сравнению со свободным PD, вероятно, благодаря
более высокой биодоступности и пролонгированному высвобождению PD из наночастиц.
КЛЮЧЕВЫЕ СЛОВА: диабетическое поражение печени, нагруженные полидатином хитозановые наночас$
тицы, ферменты метаболизма углеводов, транспортер глюкозы 2, окислительный стресс.
DOI: 10.31857/S0320972521020068
Принятые сокращения: АФК - активные формы кислорода (reactive oxygen species); CAT - каталаза (catalase);
DM - сахарный диабет (diabetes mellitus); G6PD - глюкозо$6$фосфатдегидрогеназа (glucose$6$phosphate dehydrogenase);
GK - глюкокиназа (glucokinase); GLUT2 - транспортер глюкозы 2 (glucose transporter 2); GPX - глутатионпероксидаза
(glutathione peroxidase); GSH - восстановленный глутатион (reduced glutathione); IL$1β - интерлейкин 1β (interleukin 1β);
LPO - перекисное окисление липидов (lipid peroxidation); MDA - малоновый диальдегид (malondialdehyde); NA - нико$
тинамид (nicotinamide); PD - полидатин (polydatin); PD$CSNPs - нагруженные полидатином хитозановые наночастицы
(polydatin$loaded chitosan nanoparticles); PK - пируваткиназа (pyruvate kinase); SDH - сукцинатдегидрогеназа (succinate
dehydrogenase); SOD - супероксиддисмутаза (superoxide dismutase); STZ - стрептозотоцин (streptozotocin); TNF$α - фак$
тор некроза опухолей α (tumor necrosis factor α), TP - общий белок (total protein).
biokhimiya, в рубрике «Papers in Press», BM20$162, 02.11.2020.
** Адресат для корреспонденции.
214
ДЕЙСТВИЕ ПОЛИДАТИНА ПРИ ПОРАЖЕНИИ ПЕЧЕНИ
215
ВВЕДЕНИЕ
В последние десятилетия большое внимание
было уделено поиску альтернативных методов
Сахарный диабет (DM - diabetes mellitus) яв$
лечения диабета и связанных с ним осложне$
ляется комплексной, распространенной и серь$
ний, разработке эффективных и безопасных ме$
езной проблемой, касающейся здоровья челове$
тодов лечения на основе растительных препара$
ка. Это заболевание характеризуется хроничес$
тов [9]. Полидатин (абб. PD от англ. polydatin),
кой гипергликемией, которая является резуль$
также известный как пицеид (pieceid; 3,4′,5$три$
татом дефицита инсулина и/или эффектов тако$
гидроксистильбен$3$β$D$глюкозид), был пер$
го дефицита [1]. Сахарный диабет ассоциирован
воначально выделен из корней и корневищ тра$
с нарушением функций и поражениями различ$
вы Polygonum cuspidatum, произрастающей в Ки$
ных органов, которые в конечном итоге приво$
тае, которая традиционно используется при ле$
дят ко многим опасным для здоровья осложне$
чении лихорадки, боли, кашля и гипертензии.
ниям и высокой заболеваемости и смертности
Также было показано, что PD может использо$
среди больных диабетом [1, 2]. В печени, кото$
ваться как мощный агент детоксикации при хо$
рая является одним из основных поражаемых
лестатическом поражении печени [10]. Антиок$
органов, сахарный диабет вызывает долговре$
сидантное действие полидатина осуществляется
менные метаболические дисфункции, приводя$
путем регулирования продукции АФК и функ$
щие к поражению тканей и способствующие
ционирования митохондрий; противовоспали$
прогрессированию различных заболеваний, та$
тельное действие PD состоит в снижении про$
ких как неалкогольная жировая болезнь печени,
дукции провоспалительных цитокинов [11].
цирроз и гепатоцеллюлярная карцинома [3].
Применение полидатина в клинической прак$
Среди больных диабетом широко распростране$
тике ограничено, несмотря на его многообеща$
но поражение печени; в частности, у 75% боль$
ющие фармакологические характеристики, вви$
ных диабетом 2$го типа диагностирована неал$
ду низкой биодоступности в связи с его хими$
когольная жировая болезнь печени [4].
ческой нестабильностью в водной среде при ще$
Неконтролируемая гипергликемия, ассоции$
лочных значениях рН, плохой растворимости в
рованная с сахарным диабетом, вызывает
воде и значительного пресистемного метаболиз$
окислительные повреждения, способствует
ма [12]. Несколько исследовательских групп ра$
инициации воспалительных процессов и приво$
ботают над решением этой проблемы путем раз$
дит к возникновению и прогрессированию за$
работки систем доставки лекарств, которые
болеваний печени [5]. Lucchesi et al. [6] показа$
могли бы защитить полидатин от деградации,
ли, что гипергликемия приводит к окислитель$
повысить растворимость загруженного вещест$
ному стрессу в клетках печени, который прояв$
ва в водных растворах, обеспечить направлен$
ляется повышением уровня активных форм
ную доставку лекарства к возможным мишеням
кислорода (АФК) и снижением экспрессии ан$
и паттерны непрерывного высвобождения [13].
тиоксидантных маркеров. Большое количество
Целью настоящего исследования было изучение
АФК может продуцироваться в ходе окисли$
вероятного положительного эффекта нагружен$
тельного фосфорилирования в митохондриях
ных полидатином хитозановых наночастиц
или другими путями, такими как аутоокисление
(PD$CSNPs) и свободного полидатина при по$
глюкозы, неферментативное гликирование, ак$
ражении печени, ассоциированном с диабетом,
тивация протеинкиназы C (абб. PKC от англ.
с акцентом на модуляцию активности фермен$
protein kinase С), а также метаболические пути
тов метаболизма углеводов и экспрессию транс$
гексозамина и сорбитола [5, 7]. Воспаление мо$
портера глюкозы GLUT2, а также окислитель$
жет также быть важным механизмом поражения
ный стресс и экспрессию провоспалительных
печени у больных диабетом. В диабетическом
маркеров.
состоянии происходит повышение экспрессии
фактора некроза опухолей α (абб. TNF$α от
англ. tumor necrosis factor α) и его рецептора
МАТЕРИАЛЫ И МЕТОДЫ
TNF$R1, что способствует повышению экспрес$
сии индуцибельной синтазы оксида азота (абб.
Препараты и реагенты. Полидатин, никоти$
iNOS от англ. inducible nitric oxide synthase) и
намид (NA) и стрептозотоцин (STZ) были при$
продукции в печени оксида азота [8]. Такое по$
обретены в «Sigma$Aldrich» (США). Метформин
вышение уровня провоспалительных цитокинов
(MET) был получен в «Merck KGaA» (Герма$
усиливает негативные эффекты глюкозотоксич$
ния). Хитозановые наночастицы с полидатином
ности и приводит к дисфункции митохондрий,
(PD$CSNPs) были синтезированы с помощью
окислительному стрессу и поражению печени
модифицированного метода ионного гелеобра$
[3, 8].
зования [14] и охарактеризованы по ранее опи$
БИОХИМИЯ том 86 вып. 2 2021
216
ЭЛЬ$ХАМИД и др.
санной Abdel$Moneim et al. методике [15]. Ос$
Активность аспартаттрансаминазы (AST) и
тальные реагенты и материалы были приобрете$
аланинтрансаминазы (ALT) в сыворотке крови
ны у стандартных коммерческих поставщиков.
определяли с помощью наборов «Biosystems
Лабораторные животные. Крысы$самцы по$
Company» (Испания). Колориметрическое оп$
роды Wistar albino (весом 120-140 г) были при$
ределение активности сукцинатдегидрогеназы
обретены в компании «VACSERA» (Египет).
(SDH) и пируваткиназы (PK) в сыворотке крови
Животные содержались в хорошо проветривае$
проводили с помощью наборов «BioVision»
мых клетках при нормальных атмосферных ус$
(США). Общую концентрацию белка (абб. TP от
ловиях с нормальным 12$часовым световым
англ. total protein) и концентрацию альбумина
циклом свет/темнота. Работа с эксперименталь$
определяли с помощью наборов от «Spinreact»
ными животными осуществлялась в соответ$
(Испания) и «HUMAN» (Германия) соответ$
ствии с требованиями Институционального ко$
ственно, согласно инструкции производителя.
митета по содержанию и использованию лабо$
Ткань печени гомогенизировали (10%) в
раторных животных (IACUC) Университета Бе$
нормальном физиологическом растворе. Супер$
ни$Суэфа (номер разрешения от IACUC:
натант получали в результате центрифугирова$
BSU$FS$2018$8).
ния гомогената при 3000 об/мин в течение
Моделирование диабета с использованием ни#
10 мин. Гомогенат печени использовали для оп$
котинамида и стрептозотоцина (NA/STZ) у крыс.
ределения уровня перекисного окисления ли$
Стрептозотоцин (50 мг/кг массы тела) растворя$
пидов (LPO) (анализ уровня малонового диаль$
ли в холодном цитратном буфере (pH 4,5) и не$
дегида MDA), глюкозо$6$фосфатдегидрогеназы
медленно интраперитонеально вводили раствор
(G6PD), восстановленного глутатиона (GSH), а
крысам, голодавшим в течение ночи, спустя 15
также определения активности супероксиддис$
мин после интраперитонеальной инъекции
мутазы (SOD), каталазы (CAT) и глутатионпе$
раствора никотинамида (110 мг/кг массы тела),
роксидазы (GPx) с помощью специальных на$
приготовленного в нормальном физиологичес$
боров «Biodiagnostic» (Египет) в соответствии с
ком растворе [16]. Спустя неделю после инъек$
инструкциями производителя.
ции отбирали крыс для дальнейшей работы - с
Количественная ПЦР. Препарат тотальной
уровнем глюкозы в крови натощак (абб. FBG от
РНК из ткани печени получали с помощью на$
англ. fasting blood glucose) ≥ 200 мг/дл.
бора для экстракции компании «Qiagen» (США)
Схема эксперимента. Животные были разде$
в соответствии с инструкциями производителя.
лены на шесть групп (по 6 особей в каждой груп$
Чистоту выделенной РНК (по соотношению
пе): нормальные контрольные крысы, контроль$
A260/A280) и её концентрацию определяли с
ные крысы с диабетом (D), крысы с диабетом,
помощью двухволнового спектрофотометра
получавшие полидатин (D + PD, 50 мг/кг массы
(«Beckman», США). Тотальную РНК (0,5-2 мкг)
тела), крысы с диабетом, получавшие нагружен$
использовали для получения кДНК с помощью
ные полидатином хитозановые наночастицы
набора обратной транскрипции «High$capacity
(D + PD$CSNPs, в количестве, эквивалентном
cDNA reverse transcription kit» («Fermentas»,
PD 50 мг/кг массы тела), крысы с диабетом, ко$
США) согласно протоколу производителя. Ко$
торые получали эквивалентное количество хито$
личественную ПЦР в реальном времени и ана$
зановых
наночастиц
без
полидатина
лиз продуктов реакции проводили с использова$
(D + CSNPs), и крысы с диабетом, которые по$
нием системы StepOne Real$Time PCR («Applied
лучали перорально метформин - стандартный
Biosystems», США) с соответствующим программ$
агент, вызывающий гипогликемию (D + MET,
ным обеспечением, версия 3.1. Температура от$
100 мг/кг массы тела). Все препараты вводили
жига праймеров была оптимизирована под ис$
крысам ежедневно с помощью желудочного зон$
пользованные наборы праймеров (табл. 1). Ко$
да; размер дозы пересчитывали каждую неделю в
личественное определение проводили с исполь$
соответствии с изменениями веса животного.
зованием программного обеспечения «Applied
Биохимический анализ. За день до умерщвле$
Biosystems» методом ΔΔCt. Значение RQ предс$
ния (после 4$х недель эксперимента) из лате$
тавляет собой кратность изменения по сравне$
ральной хвостовой вены голодавших в течение
нию с нормальным контролем.
ночи (8-10 ч) крыс производили забор крови.
Гистологические исследования. Для проведе$
Выделенная сыворотка крови была использова$
ния гистологического анализа брали по пять
на для спектрофотометрического определения
животных из каждой группы. Вкратце, неболь$
содержания глюкозы с использованием набора
шой кусочек печени фиксировали в 10%$ном
компании «Spinreact» (Испания). Содержание
формалине при нейтральных значениях рН в те$
гликогена в печени определяли по методу
чение 24 ч. Ткань обезвоживали растворами эта$
Seifter et al. [17].
нола с повышающимися концентрациями (70%,
БИОХИМИЯ том 86 вып. 2 2021
ДЕЙСТВИЕ ПОЛИДАТИНА ПРИ ПОРАЖЕНИИ ПЕЧЕНИ
217
Таблица 1. Пары праймеров, использованные для проведения количественной ПЦР в реальном времени
Ген
Прямой праймер (5′→3′)
Обратный праймер (5′→3′)
β$актин
TGTTTGAGACCTTCAACACC
CGCTCATTGCCGATAGTGAT
GK
CAACTGGACCAAGGGCTTCAA
TGTGGCCACCGTGTCATTC
GLUT2
CTGGAGCCCTCTTGATGGGA
CCAGTCCTGAAATTAGCCCACA
IL$1β
TGATGTCCCATTAGACAGC
GAGGTGCTGATGTACCAGTT
TNF$α
GCGACGTGGAACTGGCAGAAG
GGTACAACCCATCGGCTGGCA
Примечание. GK - глюкокиназа; GLUT2 - транспортер глюкозы 2; IL$1β - интерлейкин 1β; TNF$α - фактор некроза
опухолей α.
95% и 100%), погружали в парафин, с помощью
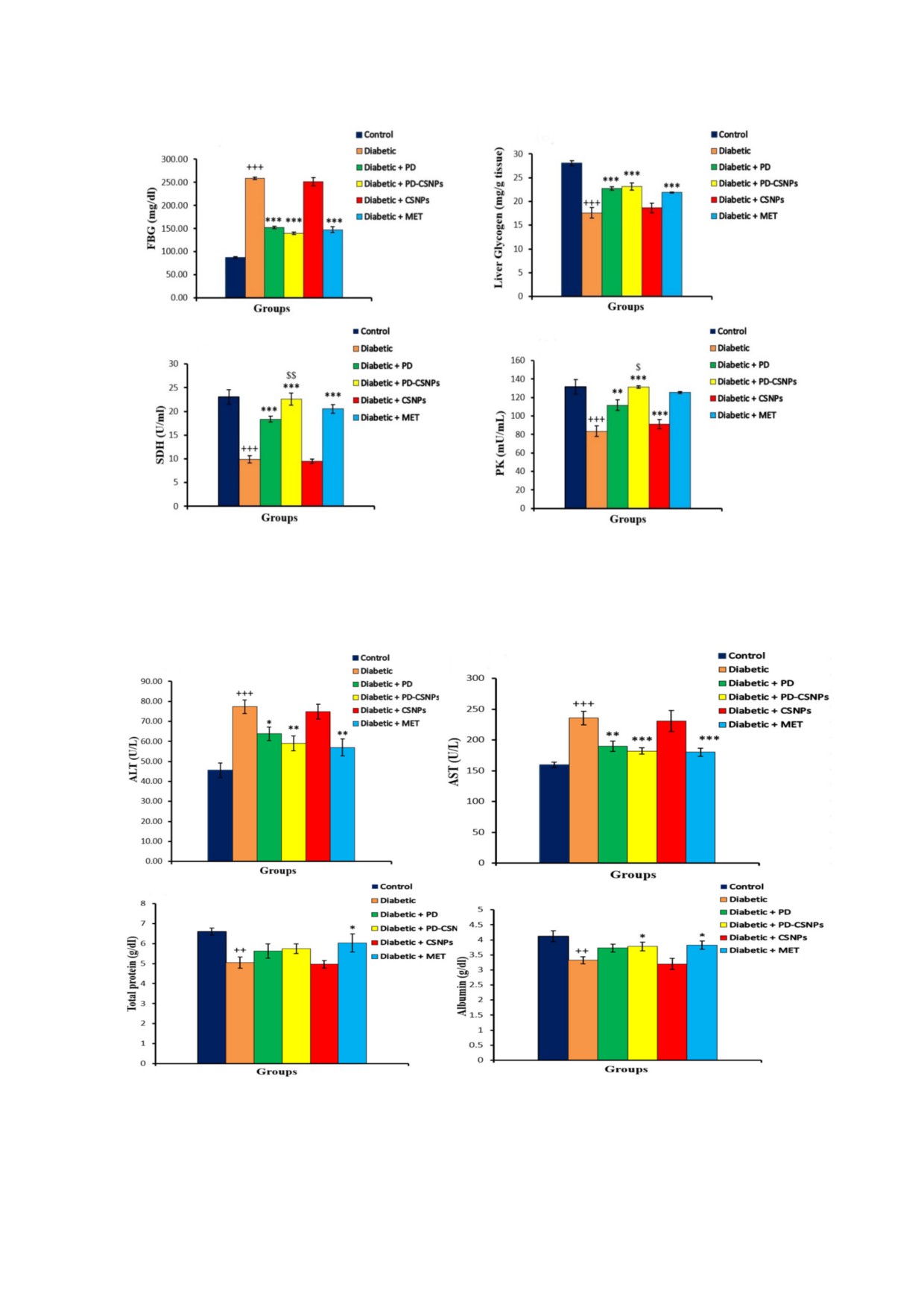
активности и AST, и ALT в сыворотке крови диа$
микротома делали срезы толщиной 4-5 мкм и
бетических крыс по сравнению с нормальными
проводили окрашивание гематоксилином и
контрольными крысами (p < 0,001). Примене$
эозином. С помощью светового микроскопа ис$
ние PD или PD$CSNPs вызывало значительное
следовали каждый препарат в пяти полях зрения
снижение активности AST и ALT у этих крыс.
[18].
Общая концентрация белка у крыс в диабети$
Статистическая обработка результатов. Полу$
ческой группе была значительно снижена по
ченные данные анализировали с помощью
сравнению с нормальным контролем (p < 0,01),
программы SPSS, version 20 for Windows («SPSS
но была значительно выше в группе крыс, по$
Inc.», США). Для сравнения эксперименталь$
лучающих метформин, по сравнению
ных групп использовали метод однофакторного
с контрольными диабетическими крысами
дисперсионного анализа ANOVA и затем метод
(p < 0,05). Концентрация сывороточного альбу$
группирования выборок с наименее значимой
мина была также существенно снижена по
разницей для множественных сравнений. Раз$
сравнению с нормальной контрольной группой
личия считали достоверными при значении
(p < 0,01), но значительно возрастала (p < 0,05)
p < 0,05.
после применения PD$CSNPs или метформина
(рис. 2).
Влияние PD и PD#CSNPs на окислительный
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
стресс и антиоксидантные маркеры в печени. В
сравнении с нормальными контрольными кры$
Влияние PD и PD#CSNPs на метаболизм угле#
сами было обнаружено заметное повышение
водов. Одним из прямых проявлений наруше$
(p < 0,001) уровня малонового диальдегида (мар$
ния гомеостаза углеводов является изменение
кера перекисного окисления липидов) у диабе$
активности ферментов метаболизма углеводов.
тических крыс. Также наблюдалось заметное
На рис. 1 показано, что применение препаратов
снижение (p < 0,05) активности SOD, CAT, GPx,
PD и PD$CSNPs снижало уровень FBG
G6PD и уровня GSH в диабетической группе по
(p < 0,001), в то время как активность SDH и PK
сравнению с нормальной контрольной группой.
в сыворотке крови заметно повышалась по срав$
Как и предполагалось, применение PD и
нению с контрольными диабетическими крыса$
PD$CSNPs вызывало повышение уровня GSH и
ми. Применение PD и PD$CSNPs также вызы$
активности SOD, GPx, CAT и G6PD параллель$
вало повышение содержания гликогена в пече$
но со снижением продукции MDA по сравне$
ни (p < 0,001). При этом препарат PD$CSNPs
нию с диабетическими контрольными крысами.
вызывал более выраженный эффект в сравне$
Таким образом, применение PD, PD$CSNPs и
нии со свободным полидатином. Эффектив$
MET может значительно снизить окислитель$
ность действия PD и PD$CSNPs была сравнима
ный стресс и повысить экспрессию антиоксидант$
с положительным эффектом метформина в
ных маркеров в ткани печени крыс, страдающих
сравнении с диабетическими контрольными
диабетом (табл. 2).
крысами (рис. 1).
Влияние PD и PD#CSNPs на экспрессию
Влияние PD и PD#CSNPs на активность
GLUT2, IL#1β, TNF#α и глюкокиназы (GK). В пе$
трансаминаз и белковый профиль. Повышение
чени контрольных крыс с диабетом экспрессия
активности AST и ALT в сыворотке крови явля$
мРНК GLUT2 и GK была снижена в сравнении
ется стандартным биохимическим маркером
с нормальной контрольной группой (p < 0,001).
поражения печени. Мы выявили повышение
Применение PD или PD$CSNPs повышало
БИОХИМИЯ том 86 вып. 2 2021
218
ЭЛЬ$ХАМИД и др.
а
б
в
г
Рис. 1. Влияние PD и PD$CSNPs на уровень глюкозы натощак в сыворотке крови (а), гликогена в печени (б) и активность
ключевых ферментов метаболизма углеводов, SDH (в) и PK (г). Результаты представлены в виде среднего значения ± стан$
дартная ошибка. +++, p < 0,001 в сравнении с нормальной контрольной группой; **, p < 0,01; ***, p < 0,001 в сравнении с
диабетической контрольной группой; $, p < 0,05; $$, p < 0,01 в сравнении с диабетическими крысами, получавшими по$
лидатин (Diabetic + PD). (С цветными вариантами рис. 1-5 можно ознакомиться в электронной версии статьи на сайте:
а
б
в
г
Рис. 2. Влияние PD и PD$CSNPs на ферменты печени (а - ALT, б - AST) и белковый профиль у диабетических крыс (в -
общий белок, г - альбумин). Результаты представлены в виде среднего значения ± стандартная ошибка. ++, p < 0,01;
+++, p < 0,001 в сравнении с нормальной контрольной группой; *, p < 0,05; **, p < 0,01; ***, p < 0,001 в сравнении с диа$
бетической контрольной группой
БИОХИМИЯ том 86 вып. 2 2021
ДЕЙСТВИЕ ПОЛИДАТИНА ПРИ ПОРАЖЕНИИ ПЕЧЕНИ
219
Таблица 2. Влияние PD и PD$CSNPs на окислительный стресс и антиоксидантные маркеры в печени диабетических
крыс
Параметр
LPO (нмоль
SOD
CAT
GSH
GPx
G6PD
MDA/100 мг
(Ед/г ткани)
(K × 10-2)
(нмоль/100 мг
(мЕд/100 мг
(мЕд/мл)
Группа
ткани)
ткани)
ткани)
Контроль
23,33 ± 0,50
14,53 ± 0,68
97,50 ± 6,82
170,24 ± 25,78
112,03 ± 2,52
15,13 ± 1,21
Диабет
39,98 ± 2,85+++
8,24 ± 0,49+++
56,67 ± 4,75++
105,60 ± 15,88++
79,52 ± 6,04+++
6,09 ± 0,34+++
Диабет + PD
33,01 ± 0,93*
10,46 ± 0,58*
79,78 ± 2,55*
149,82 ± 9,58*
91,40 ± 2,00*
9,65 ± 0,20**
Диабет + PD$CSNPs
30,08 ± 1,16**
11,13 ± 0,43**
85,61 ± 5,47*
167,68 ± 7,85**
95,18 ± 2,80**
13,23 ± 0,73***$$
Диабет + CSNPs
38,42 ± 2,33
8,29 ± 0,53
65,30 ± 14,31
112,20 ± 14,36
83,50 ± 4,63
7,03 ± 0,28
Диабет + MET
33,16 ± 2,30*
11,02 ± 0,65**
81,86 ± 8,49*
158,63 ± 9,47*
91,53 ± 4,21*
11,49 ± 0,65***
Примечание. Результаты представлены в виде среднего значения ± стандартная ошибка. ++, p < 0,01 и +++, p < 0,001 в
сравнении с нормальной контрольной группой; *, p < 0,05 и **, p < 0,01 в сравнении с диабетической контрольной груп$
пой; $$, p < 0,01 в сравнении с группой «Диабет + PD».
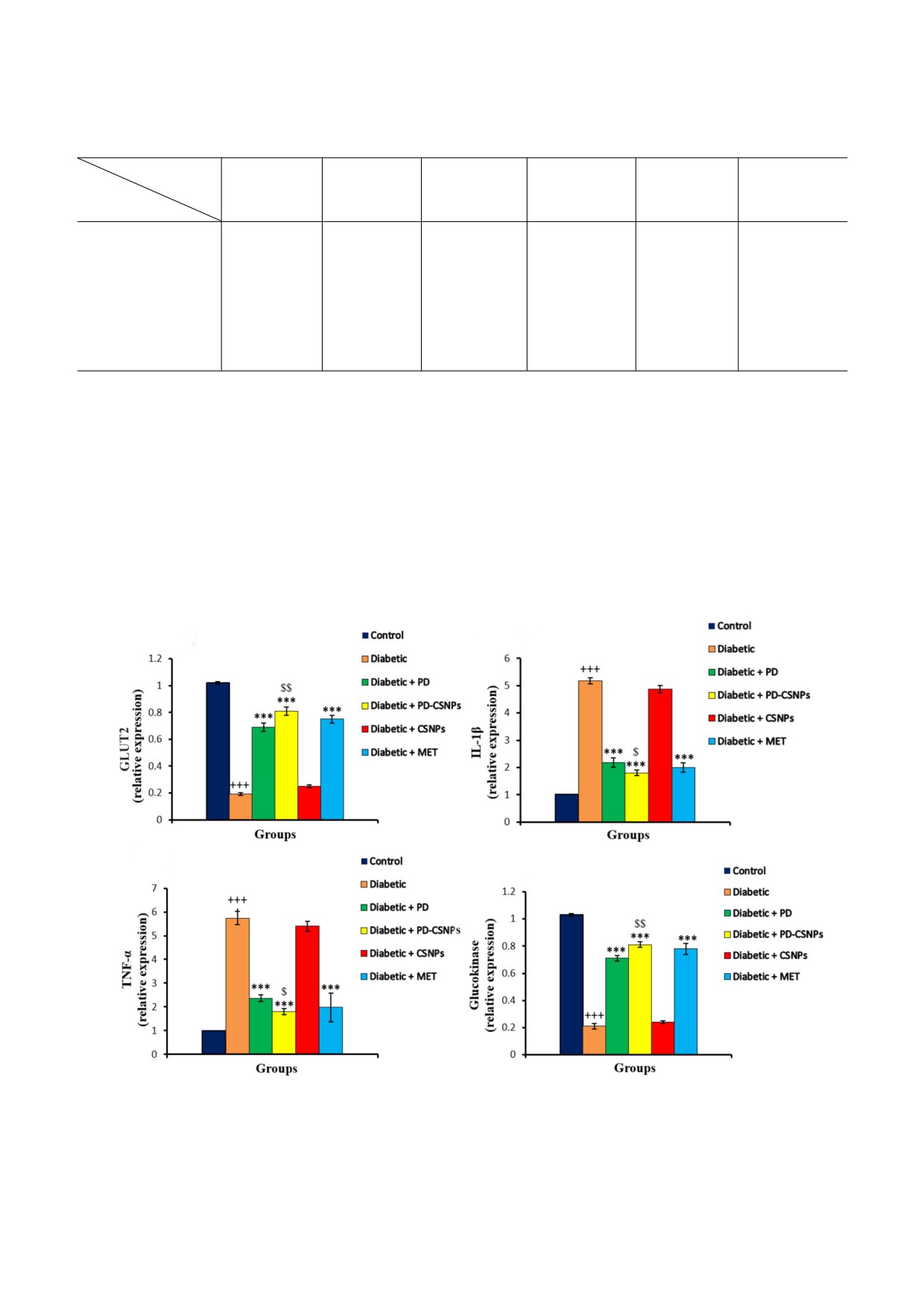
экспрессию GLUT2 и GK относительно группы
Что касается воспалительных цитокинов, то
диабетического контроля (p < 0,001). Следует
было обнаружено повышение уровня мРНК
отметить, что использование PD$CSNPs вызы$
TNF$α и IL$1β в ткани печени крыс из диабети$
вало более выраженный положительный эф$
ческой контрольной группы в сравнении с нор$
фект по сравнению с группой, получавшей сво$
мальной контрольной группой (p < 0,001). При$
бодный полидатин (p < 0,01) (рис. 3).
менение PD или PD$CSNPs вызывало сниже$
а
б
в
г
Рис. 3. Влияние PD и PD$CSNPs на экспрессию GLUT2 (а), IL$1β (б), TNF$α (в) и GK (г). Результаты представлены в ви$
де среднего значения ± стандартная ошибка. +++, p < 0,001 по сравнению с нормальной контрольной группой;
***, p < 0,001 по сравнению с диабетической группой; $, p < 0,05 и $$, p < 0,01 по сравнению с диабетической группой, по$
лучавшей полидатин (Diabetic + PD)
БИОХИМИЯ том 86 вып. 2 2021
220
ЭЛЬ$ХАМИД и др.
ние экспрессии TNF$α по сравнению с диабети$
чие отдельных гепатоцитов с пикнотическими и
ческой контрольной группой (p < 0,001). Поло$
кариолитическими ядрами (рис. 4, б). У диабе$
жительный эффект в группе PD$CSNPs был бо$
тических крыс, получавших ненагруженные
лее значительным в сравнении с группой, полу$
CSNPs, наблюдались жировые изменения, рас$
чавшей свободный PD (p < 0,05). При этом при$
ширенные гиперемированные синусоиды, вакуо$
менение и PD, и PD$CSNPs приводило к замет$
лярная дегенерация гепатоцитов и отдельные
ному улучшению по сравнению с использовани$
гепатоциты с пикнотическими и кариолитичес$
ем MET (рис. 3).
кими ядрами (рис. 4, в). В то же время примене$
Влияние PD и PD#CSNPs на гистологию пече#
ние PD$CSNPs вызывало заметные улучшения в
ни. Гистологическое исследование печени нор$
большей части печени (рис. 4, г). Использова$
мальных контрольных крыс показало типичную
ние PD$CSNPs инициировало более выражен$
структуру с центральной веной, синусоидами и
ный защитный эффект против индуцируемых
гепатоцитами (рис. 4, а). Срезы печени конт$
диабетом гистологических аномалий в печени
рольных животных с диабетом показали значи$
по сравнению с применением свободного PD
тельные поражения в виде жировых изменений,
(рис. 4, д). Как и ожидалось, MET также способ$
расширенных гиперемированных синусоидов,
ствовал улучшению состояния ткани печени
вакуолярную дегенерацию гепатоцитов и нали$
(рис. 4, е).
а
в
д
б
г
е
Рис. 4. Микрофотографии срезов печени, окрашенных гематоксилином и эозином: а - контрольные нормальные крысы,
у которых наблюдается нормальная структура с центральной веной (CV - central vein), синусоидами (стрелка) и гепатоци$
тами (H); б - контрольные животные с диабетом, у которых наблюдаются жировые изменения (F), расширенные гипере$
мированные синусоиды (стрелка), вакуолярная дегенерация гепатоцитов (V) и отдельные гепатоциты с пикнотическими
(P) и кариолитическими (K) ядрами; в - диабетические крысы, получавшие ненагруженные хитозановые наночастицы, у
которых наблюдаются жировые изменения (F), расширенные гиперемированные синусоиды (стрелка), вакуолярная деге$
нерация гепатоцитов (V) и отдельные гепатоциты с пикнотическими (P) и кариолитическими (K) ядрами; г - диабетичес$
кие крысы, которые получали нагруженные полидатином хитозановые наночастицы, у которых наблюдается заметное
улучшение в ткани печени с центральной веной (CV); д - диабетические крысы, получавшие полидатин, показана цент$
ральная вена (CV) и умеренно выраженные улучшения в большинстве гепатоцитов; е - диабетическая группа животных,
которые получали MET$HCl, зафиксировано улучшение состояния большинства гепатоцитов и центральной вены (CV).
Единица шкалы - 100 мкм
БИОХИМИЯ том 86 вып. 2 2021
ДЕЙСТВИЕ ПОЛИДАТИНА ПРИ ПОРАЖЕНИИ ПЕЧЕНИ
221
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
В случае диабета 2$го типа нечувствитель$
ность к глюкозе часто ассоциируется с наруше$
Печень является одним из главных органов,
нием функционирования транспортера глюко$
испытывающих воздействие хронических мета$
зы 2 (GLUT2) [23]. Глюкокиназа является физио$
болических нарушений, ассоциированных с диа$
логическим сенсором глюкозы в глюкозозави$
бетом, которые в конечном итоге вызывают по$
симых тканях, включая поджелудочную железу
ражение печени и прогрессирование многих за$
и печень [24]. В настоящей работе мы показали,
болеваний, таких как неалкогольная жировая
что экспрессия мРНК GLUT2 и GK в печени
болезнь печени, цирроз и гепатоцеллюлярная
крыс с диабетом была снижена. После введения
карцинома [3]. Полидатин, биологически актив$
крысам препаратов PD и PD$CSNPs происходи$
ное вещество растительного происхождения, об$
ло восстановление уровня мРНК GLUT2 и GK
ладает многими полезными с точки зрения тера$
практически до нормальных значений, более
пии свойствами, включая антиоксидантную и
выраженный эффект наблюдался в группе
противовоспалительную активность [11], что де$
PD$CSNPs. GLUT2 является основной изофор$
лает его хорошим кандидатом для смягчения по$
мой транспортера глюкозы в печени, где он
ражения печени у больных диабетом. В настоя$
участвует в транспорте глюкозы в обоих направ$
щей работе показано, что пероральное введение
лениях и, таким образом, играет ключевую роль
свободного полидатина и нагруженных полида$
в гомеостазе глюкозы [25]. Снижение уровня
тином хитозановых наночастиц в течение 4$х не$
GLUT2 приводит к падению глюкозозависимой
дель после индукции диабета приводило к смяг$
секреции инсулина из$за ограниченного коли$
чению поражения печени у диабетических крыс.
чества глюкозы, поступающего в β$клетки под$
Эффекты PD и PD$CSNPs как потенциаль$
желудочной железы [26], и может таким образом
ных гипогликемических агентов у крыс с диабе$
быть одной из причин гипергликемии. В печени
том оказались сходны с действием MET. Мет$
в ответ на высокий уровень глюкозы в плазме
формин может снижать уровень глюкозы в кро$
крови GLUT2 транспортируется из цитоплазмы
ви путем повышения чувствительности к инсу$
на плазматическую мембрану, где он служит в
лину и ускорения усвоения глюкозы [19]. Анти$
качестве основного переносчика глюкозы из
гипергликемический эффект PD и PD$CSNPs у
плазмы в гепатоциты [27]. Утрата функций глю$
диабетических крыс подтверждается снижением
кокиназы, помимо нарушения метаболизма уг$
уровня FBG в сравнении с контрольными диа$
леводов, также ассоциирована с гипергликеми$
бетическими крысами. Ранее мы показали, что
ей или гипогликемией [28], и изменения уровня
применение PD и PD$CSNPs способствовало
глюкозы в крови находятся под влиянием
нормализации уровня инсулина и заметному
экспрессии GK в печени [29]. Следовательно,
снижению уровня гликированного гемоглобина
восстановление уровня GLUT2 повысит усвое$
(HbA1c), а также параметров гомеостатической
ние глюкозы и тем самым поможет бороться с
модели оценки резистентности к инсулину (абб.
гипергликемическими состояниями. Так, у
HOMA$IR от англ. homeostatic model assessment
страдающих ожирением и диабетом крыс Цуке$
of insulin resistance) у получавших эти препараты
ра экспрессия GK в печени снижается по мере
крыс$диабетиков [15]. В сравнении с контроль$
усиления тяжести заболевания. У мышей гипер$
ными крысами, у крыс с диабетом наблюдался
экспрессия GK в печени способствовала норма$
дефицит гликогена в печени, который значи$
лизации уровня глюкозы, в то время как дефи$
тельно смягчался при применении PD и
цит GK был ассоциирован с гипергликемией
PD$CSNPs. В число факторов, приводящих к
[30].
снижению содержания гликогена в печени крыс
Гликолиз представляет собой единственный
с диабетом, входит инсулиновая недостаточ$
путь катаболизма глюкозы. Потребление глюко$
ность, которая, в свою очередь, приводит к ак$
зы с пищей является необходимым для регуля$
тивации процессов глюконеогенеза и гликоге$
ции гликолитического пути в печени [31]. Пиру$
нолиза [20]. Гликоген является основной внут$
ваткиназа (PK) является одним из скорость$
риклеточной формой запасания глюкозы в пе$
лимитирующих ферментов гликолиза, что также
чени, и его уровень является прямым свидетель$
оказывает прямое влияние на скорость и на$
ством активности инсулина, т.к. инсулин стиму$
правление всего метаболизма углеводов [32]. PK
лирует гликогенсинтазу и, соответственно, де$
катализирует реакцию синтеза пирувата и ATP
понирование гликогена [21]. Эти результаты
путем переноса фосфатной группы с фосфо$
согласуются с данными Wang et al. [22], которые
енолпирувата на ADP [33], она ускоряет процесс
наблюдали повышение содержания гликогена в
усвоения глюкозы и способствует гликолизу.
печени мышей с диабетом 2$го типа после вве$
SDH участвует в процессах как гликолиза, так и
дения им полидатина.
цикла трикарбоновых кислот (абб. TCA от англ.
БИОХИМИЯ том 86 вып. 2 2021
222
ЭЛЬ$ХАМИД и др.
tricarboxylic acid cycle) [34]. Активация действия
шего исследования, в печени крыс с диабетом
SDH в электрон$транспортной цепи и TCA мог$
происходит значительное увеличение концент$
ла бы способствовать усвоению глюкозы в пече$
рации малонового диальдегида, а также сниже$
ни [35]. В настоящей работе показано, что ак$
ние уровня GSH и снижение активности анти$
тивность PK и SDH в сыворотке крови диабети$
оксидантных ферментов, таких как SOD, GPx,
ческих крыс значительно снижена. После введе$
CAT и G6PD, в сравнении с нормальными конт$
ния крысам препаратов PD и PD$CSNPs актив$
рольными крысами. Дисфункция митохондрий,
ность этих ключевых ферментов метаболизма
вызванная гипергликемией, и стресс эндоплаз$
углеводов значительно возрастала. Повышение
матического ретикулума способствуют аккуму$
активности PK и SDH может быть вторичным
ляции АФК, которая, в свою очередь, способ$
по отношению к благоприятному влиянию
ствует процессу повреждения клеток и вносит
PD$CSNPs на уровень инсулина и приводить,
вклад в развитие диабетических осложнений,
как указано выше, к усилению гликолиза и усвое$
приводя к появлению необратимых окислитель$
ния глюкозы для генерирования энергии. Наши
ных модификаций [43].
результаты
согласуются
с
выводами
Основным внутриклеточным восстановите$
Meghana et al. [36], которые показали, что при$
лем является НАДФН. Действительно, для
менение полидатина вызывает повышение
функционирования всей антиоксидантной сис$
уровня SDH в животной модели алкогольного
темы необходим восстановитель. Глюко$
поражения печени. Предположительно полида$
зо$6$фосфатдегидрогеназа является основным
тин оказывает антидиабетические эффекты пу$
источником НАДФН, поэтому нарушение ак$
тем стимулирования усвоения внутриклеточной
тивности G6PD может приводить к изменению
глюкозы за счет повышения экспрессии
уровня НАДФН, таким образом влияя на всю
GLUT2, активации пентозофосфатного пути за
антиоксидантную систему [44]. Таким образом,
счет повышения уровня SDH, а также восста$
снижение уровня НАДФН в результате ингиби$
навливая гликолиз в печени путем регуляции
рования активности G6PD делает клетки очень
ключевых ферментов метаболизма углево$
чувствительными к повреждающему действию
дов [37].
окислителей. В соответствии с нашими резуль$
Для диагностики функционального состоя$
татами было показано, что сахарный диабет ве$
ния печени используется анализ белкового про$
дет к ингибированию активности G6PD у экс$
филя [38]. В настоящей работе показано значи$
периментальных животных с диабетом и в куль$
тельное снижение содержания сывороточного
тивированных эндотелиальных клетках [45].
альбумина и общего белка в крови животных
Кроме того, у больных диабетом наблюдается
диабетической группы. Это может быть связано
пониженная активность G6PD в печени [46].
с повышенной скоростью конверсии аминокис$
Пониженная активность G6PD приводит к сни$
лот в глюкозу, глюконеогенеза, и конверсией
жению уровня внутриклеточного НАДФН, по$
гликогенных аминокислот в CO2 и H2O [39].
вышению содержания АФК в клетках, а также
Также это может быть ассоциировано с наруше$
способствует окислительному стрессу. Более то$
ниями функций и структуры клеток печени, ко$
го, АФК могут инициировать перекисное окис$
торые характеризуются низким уровнем сыво$
ление липидов путем окисления полиненасы$
роточного альбумина и общего белка [40]. Вве$
щенных жирных кислот в мембранах гепатоци$
дение крысам с диабетом PD или PD$CSNPs
тов, что приводит к повышению проницаемости
привело к заметному повышению уровня сыво$
клеточных мембран и последующим поврежде$
роточного альбумина и общего белка, что, пред$
ниям клеток [47], что также может вызывать по$
положительно, опосредовано гипогликемичес$
вышение активности сывороточных ALT и AST.
ким эффектом полидатина. Эти результаты сог$
Как и предполагалось, применение PD или
ласуются с данными Ince et al. [41], где было по$
PD$CSNPs у крыс с диабетом в значительной
казано, что полидатин дозозависимо восстанав$
степени препятствовало окислению липидов за
ливает уровень общего белка и альбумина в кро$
счет повышения уровня активности антиокси$
ви крыс, нарушенные цисплатин$индуцирован$
дантов и антиоксидантных ферментов (GSH,
ной токсичностью.
GPx, SOD, CAT и G6PD) и снижения уровня
Как правило, дефицит инсулина и сопут$
MDA в сравнении с контрольными крысами с
ствующая гипергликемия наблюдаются в ре$
диабетом. Zhang et al. [48] показали, что полида$
зультате нарушения функционирования β$кле$
тин снижает окислительный стресс и замедляет
ток, которые считаются основными посредни$
процесс поражения печени через снижение
ками в процессе стимулирования продукции
уровня MDA и повышение активности SOD,
АФК, сопровождающем поражение печени при
GSH, GPx и CAT в печени. Повышенная актив$
сахарном диабете [42]. Согласно результатам на$
ность G6PD, наблюдаемая у крыс в группах, по$
БИОХИМИЯ том 86 вып. 2 2021
ДЕЙСТВИЕ ПОЛИДАТИНА ПРИ ПОРАЖЕНИИ ПЕЧЕНИ
223
лучающих PD и PD$CSNP, объясняет повышен$
щающей стратегией в борьбе с осложнениями
ные уровни антиоксидантных ферментов.
сахарного диабета. Насколько нам известно,
НАДФН, в основном производимый G6PD, яв$
данная работа является первым исследованием
ляется важнейшим кофактором для каталазы и
действия PD$CSNPs как гепатопротекторного и
субстратом глутатионредуктазы, поддерживая
противовоспалительного средства, направлен$
каталазу в активном состоянии и обеспечивая
ного против поражения печени при диабете. В
восстановление глутатиона с образованием
ряде работ показано, что полидатин влияет на
GSH, который является основным акцептором
окислительный стресс и таким образом - на
свободных радикалов [49]. Антиоксидантная
воспалительный ответ [59, 60]. Гепатопротек$
роль G6PD, являющейся основным источником
торное действие свободного PD и PD$CSNPs
НАДФН, недавно была хорошо описана в рабо$
проявлялось в стимулировании активности
те Leopold et al [50].
ферментов метаболизма углеводов, ингибирова$
Ozer et al. [51] показали, что активность ALT
нии окислительного стресса путем повышения
и AST может быть использована в качестве мар$
активности антиоксидантных ферментов и
керов поражения печени. Повышенная актив$
впоследствии модулировании провоспалитель$
ность трансаминаз, выявленная в настоящей ра$
ных цитокинов.
боте, является в большей степени следствием
Гистологическое исследование показало на$
гипергликемии, приводящей к диабетическому
личие множества гистопатологических измене$
поражению печени. Мы предполагаем, что пе$
ний в печени диабетических крыс, включая жи$
роральное применение PD и PD$CSNPs обеспе$
ровые изменения, расширенные гиперемиро$
чивает антиоксидантную защиту против пере$
ванные синусоиды, вакуолярную дегенерацию
кисного окисления липидов благодаря мощной
гепатоцитов, а также отдельные гепатоциты с
активности PD в качестве акцептора свободных
пикнотическими и кариолитическими ядрами.
радикалов [52]. Предположительно, данные
Эти результаты согласуются с данными более
препараты также оказывают гепатопротектор$
ранних исследований, в которых отмечались
ный эффект при ассоциированном с диабетом
отдельные гистопатологические изменения в
поражении печени, снижая активность ALT и
печени диабетических крыс [61, 62]. В соответ$
AST у экспериментальных крыс. Эти результаты
ствии с результатами биохимических исследо$
согласуются с данными Lai et al. [53], которые
ваний и анализом экспрессии генов, примене$
описали сходные эффекты полидатина.
ние PD$CSNPs и PD вызывало заметные улуч$
В ряде работ было показано, что при диабете
шения в большинстве образцов печени диабе$
наблюдается повышение продукции некоторых
тических крыс, подтверждая гепатопротектор$
провоспалительных цитокинов, таких как
ное действие этих препаратов. Следует отме$
IL$1β, IL$6 и TNF$α, которые способствуют
тить, что нагруженные полидатином хитозано$
воспалению и развитию гепатического стеатоза,
вые наночастицы оказывали более выраженное
приводящего в итоге к поражению печени [54].
защитное действие по сравнению со свободным
В настоящей работе показано, что уровни мРНК
полидатином.
IL$1β и TNF$α в печени диабетических крыс
Метформин является одним из широко ис$
были повышены, в то время как применение PD
пользуемых лекарств при диабете и считается
и PD$CSNPs ингибировало экспрессию этих
препаратом первой линии при лечении диабета
мРНК в печени. При этом препарат PD$CSNPs
2$го типа. Клинические преимущества MET в
показал более выраженный противовоспали$
основном связаны со специфическим сниже$
тельный эффект по сравнению со свободным
нием выработки глюкозы в печени и улучшени$
PD. TNF$α является одним из основных цито$
ем периферической чувствительности к инсули$
кинов, чья экспрессия в печени значительно
ну; поэтому печень является основной ми$
увеличивается у диабетических крыс, что ведет к
шенью MET [63]. Однако лечение метформи$
воспалению и апоптозу клеток печени и ассоции$
ном часто ассоциировано с побочными эффек$
ровано с развитием окислительного стресса и
тами со стороны желудочно$кишечного тракта,
гипергликемии [55, 56]. Также описан вклад
такими как тошнота, рвота, диарея, металличес$
IL$1β при диабете, TNF$α - при алкогольном и
кий привкус и дискомфорт в брюшной полости.
вирусном гепатите, диабете и поражении пече$
MET также снижает абсорбцию витамина B12,
ни в результате ишемии/реперфузии [57, 58]. В
что приводит к развитию анемии [64, 65]. Таким
связи с существующей ассоциацией между хро$
образом, существует необходимость разработки
ническим воспалением и осложнениями диабе$
новых альтернативных препаратов растительно$
та, включая поражение печени, определение те$
го происхождения для лечения диабета и его ос$
рапевтических мишеней, способных снижать
ложнений с минимизацией побочных эффек$
провоспалительный ответ, является многообе$
тов.
БИОХИМИЯ том 86 вып. 2 2021
224
ЭЛЬ$ХАМИД и др.
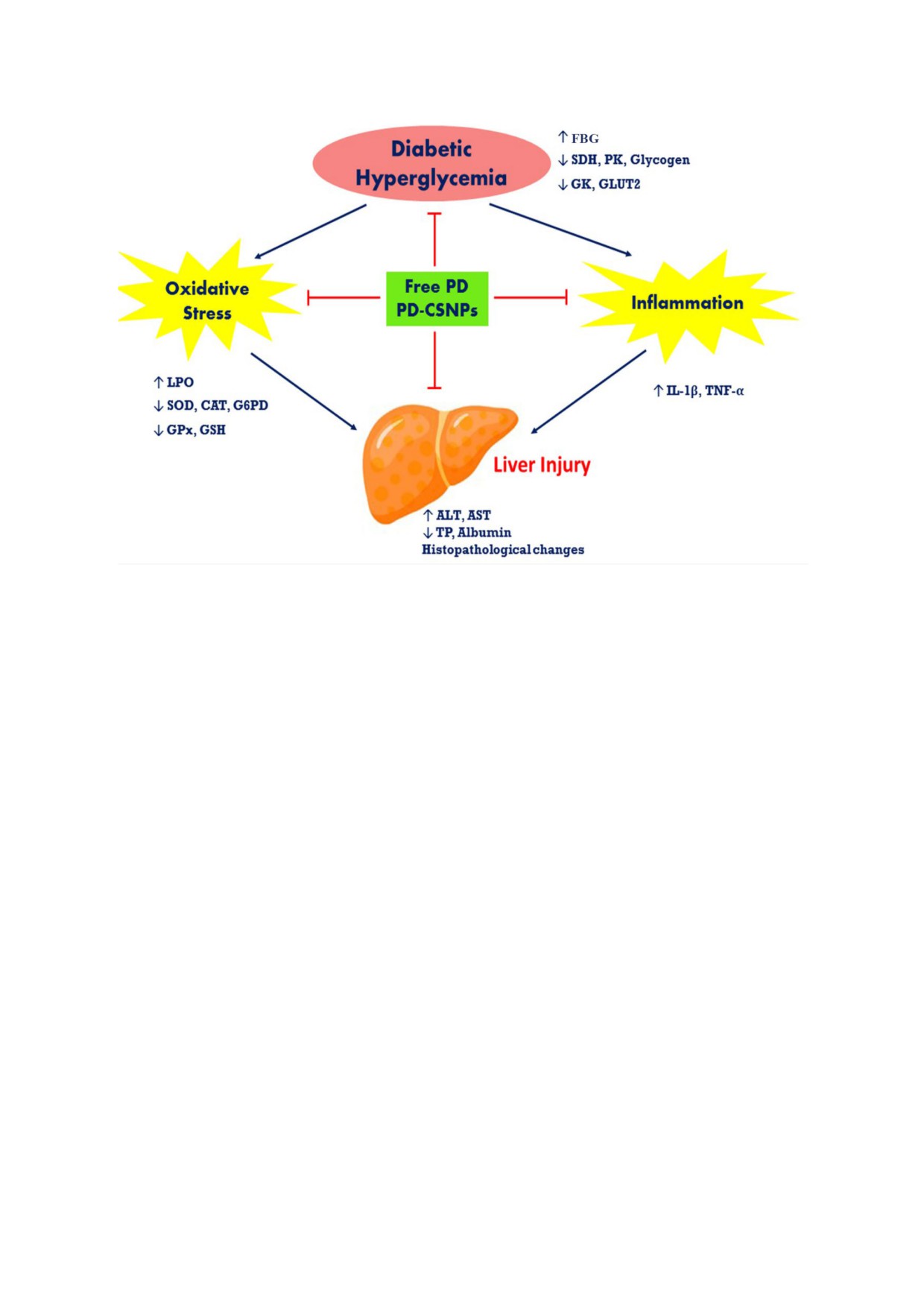
Рис. 5. Общая схема путей гепатопротекторного действия свободного PD и PD$CSNPs в печени крыс с диабетом
В связи с этим мы протестировали полида$
ный эффект против поражения печени в сравне$
тин как потенциальное средство для лечения
нии со свободным PD. Также было показано,
поражения печени, т.к. он является безопасным
что гепатопротекторное действие PD$CSNPs у
фитопрепаратом и обладает различными фарма$
крыс с диабетом может быть связано с (1) про$
кологическими активностями [10, 11]. В наших
тиводиабетическим действием путем модулиро$
предыдущих работах было продемонстрирова$
вания экспрессии GLUT2 и активности фер$
но, что хитозановые наночастицы также без$
ментов метаболизма углеводов; (2) модулирова$
опасны и могут повысить терапевтическую эф$
нием окислительного стресса благодаря антиок$
фективность полидатина, что может объяснять$
сидантным свойствам PD; и (3) противовоспа$
ся пролонгированным высвобождением, повы$
лительным действием путем снижения содержа$
шенной абсорбцией и высокой биодоступ$
ния провоспалительных цитокинов (рис. 5). На$
ностью PD в комбинации с CSNPs [15]. Наше
деемся, что результаты нашей работы также бу$
исследование подтвердило, что PD$CSNPs яв$
дут способствовать повышению интереса к изу$
ляется перспективным препаратом, имеющим
чению терапевтической роли PD$CSNPs при
ряд преимуществ перед MET, особенно в случае
других заболеваниях печени.
диабетического поражения печени, вызванного
окислительным стрессом. Препарат PD$CSNPs
Финансирование. Нет (выполнение данной
может быть адаптирован на молекулярном уров$
работы не поддерживалось каким$либо гран$
не для увеличения эффективности и минимиза$
том).
ции побочных эффектов. Тем не менее до прове$
Конфликт интересов. Авторы заявляют об от$
дения клинических исследований необходимо
сутствии конфликта интересов.
провести токсикокинетические и фармакокине$
Соблюдение этических норм. При работе с ла$
тические исследования нового препарата
бораторными животными все процедуры были
PD$CSNPs для оценки и оптимизации баланса
выполнены в соответствии с этическими стан$
между эффективностью и токсичностью разра$
дартами Институционального комитета по со$
ботанного препарата.
держанию и использованию лабораторных жи$
В настоящей работе показано, что препарат
вотных (IACUC) Университета Бени$Суэфа в
PD$CSNPs является биологически совмести$
Египте (номер разрешения IACUC: BSU$FS$
мым, а также имеет более выраженный защит$
2018$8).
БИОХИМИЯ том 86 вып. 2 2021
ДЕЙСТВИЕ ПОЛИДАТИНА ПРИ ПОРАЖЕНИИ ПЕЧЕНИ
225
СПИСОК ЛИТЕРАТУРЫ
1.
Cho, N., Shaw, J. E., Karuranga, S., Huang, Y., da Rocha
18.
Bancroft, J. D., and Gamble, M. (2002) In Theory and
Fernandes, J. D., Ohlrogge, A. W., and Malanda, B. (2018)
Practice of Histological Techniques, 5th Ed., Churchill
IDF Diabetes Atlas: global estimates of diabetes prevalence
Livingstone, pp. 172$175.
for 2017 and projections for 2045, Diabetes Res. Clin.
19.
Xu, C., Liu, W., Zhang, D., Cao, X., Shi, H., and Li, X.
Pract., 138, 271$281.
(2018) Interactions between dietary carbohydrate and met$
2.
Ahlqvist, E., Van Zuydam, N. R., Groop, L. C., and
formin: implications on energy sensing, insulin signaling
McCarthy, M. I. (2015) The genetics of diabetic complica$
pathway, glycolipid metabolism and glucose tolerance in
tions, Nat. Rev. Nephrol., 11, 277$287.
blunt snout bream Megalobrama amblycephala,
3.
Bedi, O., Aggarwal, S., Trehanpati, N., Ramakrishna, G.,
Aquaculture, 483, 183$195.
and Krishan, P. (2019) Molecular and pathological events
20.
Pari, L., and Murugan, P. (2005) Effect of tetrahydrocur$
involved in the pathogenesis of diabetes$associated nonal$
cumin on blood glucose, plasma insulin and hepatic key
coholic fatty liver disease, J. Clin. Exp. Hepatol., 9, 607$
enzymes in streptozotocin induced diabetic rats, J. Basic
618.
Clin. Physiol. Pharmacol., 16, 257$274.
4.
Angulo, P. (2002) Nonalcoholic fatty liver disease, N. Engl.
21.
Vats, V., Yadav, S., and Grover, J. (2004) Ethanolic extract
J. Med., 346, 1221$1231.
of Ocimum sanctum leaves partially attenuates streptozo$
5.
Brownlee, M. (2001) Biochemistry and molecular cell
tocin$induced alterations in glycogen content and carbo$
biology of diabetic complications, Nature, 414, 813$820.
hydrate metabolism in rats, J. Ethnopharmacol., 90, 155$
6.
Lucchesi, A. N., Freitas, N. T. D., Cassettari, L. L.,
160.
Marques, S. F. G., and Spadella, C. T. (2013) Diabetes
22.
Wang, Y., Ye, J., Li, J., Chen, C., Huang, J., Liu, P., and
mellitus triggers oxidative stress in the liver of alloxan$
Huang, H. (2016) Polydatin ameliorates lipid and glucose
treated rats: a mechanism for diabetic chronic liver disease,
metabolism in type 2 diabetes mellitus by downregulating
Acta Cir. Bras., 28, 502$508.
proprotein convertase subtilisin/kexin type 9 (PCSK9),
7.
Kangralkar, V. A., Patil, S. D., and Bandivadekar, R. M.
Cardiovasc. Diabetol., 15, 19, doi: 10.1186/s12933$015$
(2010) Oxidative stress and diabetes: a review, Int. J. Appl.
0325$x.
Pharm. Sci. Res., 1, 38$45.
23.
Thorens, B., Wu, Y. J., Leahy, J. L., and Weir, G. C. (1992)
8.
Ingaramo, P. I., Francés, D. E., Ronco, M. T., and
The loss of GLUT2 expression by glucose unresponsive
Carnovale, C. E. (2013) in Hot Topics in Endocrine and
beta cells of db/db mice is reversible and is induced by the
Endocrine Related Diseases (Fedele, M., ed.), IntechOpen,
diabetic environment, J. Clin. Invest., 90, 77$80.
London, doi: 10.5772/53684.
24.
Liu, D., Regenstein, J. M., Diao, Y., Qiu, J., Zhang, H.,
9.
Coman, C., Rugina, O. D., and Socaciu, C. (2012) Plants
Li, J., Zhao, H., and Wang, Z. (2019) Antidiabetic effects
and natural compounds with antidiabetic action, Not. Bot.
of water$soluble Korean pine nut protein on type 2 diabet$
Horti. Agrobot. Cluj. Napoca., 40, 314$325.
ic mice, Biomed. Pharmacother.,
117,
108989,
10.
Fanga, J., Luoa, L., Kea, Z., Liua, C., Yina, L., et al.
doi: 10.1016/j.biopha.2019.108989.
(2019) Polydatin protects against acute cholestatic liver
25.
Sole, S. S., and Srinivasan, B. P. (2012) Aqueous extract of
injury in mice via the inhibition of oxidative stress and
tamarind seeds selectively increases glucose transporter$2,
endoplasmic reticulum stress, J. Funct. Foods, 55, 175$183.
glucose transporter$4, and islets’ intracellular calcium lev$
11.
Ji, H., Zhang, X., Du, Y., Liu, H., Li, S., and Li, L. (2012)
els and stimulates β$cell proliferation resulting in improved
Polydatin modulates inflammation by decreasing NF$κB
glucose homeostasis in rats with streptozotocin$induced
activation and oxidative stress by increasing Gli1, Ptch1,
diabetes mellitus, Nutr. Res., 32, 626$636.
SOD1 expression and ameliorates blood-brain barrier per$
26.
Weir, G. C. (1993) The relationship of diabetes, loss glu$
meability for its neuroprotective effect in pMCAO rat
cose$induced insulin secretion, and GLUT2, J. Diabetes
brain, Brain Res. Bull., 87, 50$59.
Complications, 7, 124$129.
12.
Cheng, W., Li, X., Zhang, C., Chen, W., Yuan, H., and Xu,
27.
Zhao, F. Q., and Keating, A. F. (2007) Expression and reg$
S. (2017) Preparation and in vivo$in vitro evaluation of
ulation of glucose transporter in the bovine mammary
polydatin$phospholipid complex with improved dissolu$
gland, J. Dairy Sci., 90, E76$E86.
tion and bioavailability, Int. J. Drug Dev. Res., 9, 39$43.
28.
Gloyn, A. L., Odili, S., Zelent, D., Buettger, C.,
13.
Amri, A., Chaumeil, J., Sfar, S., and Charrueau, C. (2012)
Castleden, H. A. J., et al. (2005) Insights into the structure
Administration of resveratrol: what formulation solutions
and regulation of glucokinase from a novel mutation
to bioavailability limitations? J. Control Release, 158, 182$
(V62M), which causes maturity$onset diabetes of the
193.
young, J. Biol. Chem., 280, 14105$14113.
14.
Nagpal, K., Singh, S., and Mishra, D. (2013) Optimization
29.
Zhang, Y., Wei, Z., Liu, G., Deng, K., Yang, M., et al.
of brain targeted chitosan nanoparticles of Rivastigmine for
(2019) Synergistic effects of dietary carbohydrate and tau$
improved efficacy and safety, Int. J. Biol. Macromol., 59,
rine on growth performance, digestive enzyme activities
72$83.
and glucose metabolism in juvenile turbot Scophthalmus
15.
Abdel$Moneim, A., El$Shahawy, A., Yousef, A. I., Abd El$
maximus L., Aquaculture, 499, 32$41.
Twab, S. M., Elden, Z. E., and Taha, M. (2020) Novel
30.
Postic, C., Shiota, M., Niswender, K. D., Jetton, T. L.,
polydatin$loaded chitosan nanoparticles for safe and effi$
Chen, Y., et al. (1999) Dual roles for glucokinase in glucose
cient type 2 diabetes therapy: in silico, in vitro and in vivo
homeostasis as determined by liver and pancreatic beta
approaches, Int. J. Biol. Macromol., 154, 1496$1504.
cell$specific gene knock$outs using Cre recombinase, J.
16.
Masiello, P., Broca, C., Gross R., Roye, M., Manteghetti,
Biol. Chem., 274, 305$315.
M., Hillaire$Buys, D., Novelli, M., and Ribes, G. (1998)
31.
Li, R., Liu, H., Dong, X., Chi, S., Yang, Q., Zhang, S.,
Experimental NIDDM: development of a new model in
and Tan, B. (2018) Molecular characterization and
adult rats administered streptozotocin and nicotinamide,
expression analysis of glucose transporter1 and hepatic
Diabetes, 47, 224$229.
glycolytic enzymes activities from herbivorous fish
17.
Seifter, S., Dayton, S., Novic, B., and Muntwyler, E.
Ctenopharyngodon idellus in respond to a glucose load after
(1950) The estimation of glycogen with the anthrone
the adaptation to dietary carbohydrate levels, Aquaculture,
reagent, Arch. Biochem., 25, 191$200.
792, 290$299.
6 БИОХИМИЯ том 86 вып. 2 2021
226
ЭЛЬ$ХАМИД и др.
32.
Kayne, F. J. (1973) Pyruvate kinase, in The Enzymes, 3rd
through modulation of PPARγ, proinflammatory cytokines
Edn., Vol. VIII (Boyer, P. D., ed.) Academic Press, New
and oxidative stress, Biomed. Pharmacother., 88, 52$60.
York, pp. 353$382.
48.
Zhang, H., Yu, C. H., Jiang, Y. P., Peng, C., He, K., Tang,
33.
Gupta, V., and Bamezai, R. N. (2010) Human pyruvate
J. Y., and Xin, H. L. (2012) Protective effects of polydatin
kinase M2: a multifunctional protein, Protein Sci., 19,
from Polygonum cuspidatum against carbon tetrachloride$
2031$2044.
induced liver injury in mice, PLoS One, 7, e46574,
34.
Oyedotun, K. S., and Lemire, B. D. (2004) The quaternary
doi: 10.1371/journal.pone.0046574.
structure of the saccharomyces cerevisiae succinate dehy$
49.
Kirkman, H. N., Rolfo, M., Ferraris, A. M., and Gaetani,
drogenase, J. Biol. Chem., 279, 9424$9431.
G. F. (1999) Mechanisms of protection of catalase by
35.
Choudhury, H., Pandey, M., Hua, C. K., Mun, C. S., Jing,
NADPH. Kinetics and stoichiometry, J. Biol. Chem., 274,
J. K., et al. (2017) An update on natural compounds in the
13908$13914.
remedy of diabetes mellitus: a systematic review, J. Tradit.
50.
Leopold, J. A., Zhang, Y. Y., Scribner, A. W., Stanton, R.
Complement. Med., 8, 361$376.
C., and Loscalzo, J. (2003) Glucose$6$phosphate dehy$
36.
Koneru, M., Sahu, B. D., Gudem, S., Kuncha, M.,
drogenase overexpression decreases endothelial cell oxi$
Ravuri, H. G., Kumar, J. M., Kilari, E. K., and Sistla, R.
dant stress and increases bioavailable nitric oxide,
(2017) Polydatin alleviates alcohol$induced acute liver
Arterioscler. Thromb. Vasc. Biol., 23, 411$417.
injury in mice: Relevance of matrix metalloproteinases
51.
Ozer, J., Ratner, M., Shaw, M., Bailey, W., and Schomaker,
(MMPs) and hepatic antioxidants, Phytomedicine, 27, 23$
S. (2008) The current state of serum biomarkers of hepato$
32.
toxicity, Toxicology, 245, 194$205.
37.
Enes, P., Panserat, S., Kaushik, S., and Oliva$Teles, A.
52.
Huang, Q. H., Xu, L. Q., Liu, Y. H., Wu, J. Z., Wu, X., et
(2009) Nutritional regulation of hepatic glucose metabo$
al. (2017) Polydatin protects rat liver against ethanol$
lism in fish, Fish Physiol. Biochem., 35, 519$539.
induced injury: involvement of CYP2E1/ROS/Nrf2 and
38.
Soji$Omoniwa, O., Muhammad, N. O., Usman, L. A., and
TLR4/NF$κB p65 pathway, Evid. Based. Complement
Omoniwa, B. P. (2014) Effect of leaf essential oil of Citrus
Alternat. Med.,
2017,
7953850, doi:
10.1155/2017/
sinensis at different harvest time on some liver and kidney
7953850.
function indices of diabetic rats, Int. J. Biol. Vet. Agric. Food
53.
Lai Xue, L., Wub, K., Qiu, H., Huang, B., Chen, R., Xie,
Eng., 8, 484$488.
W., and Jiang, Q. (2017) Polydatin exhibits the hepatopro$
39.
Abdel$Moneim, A., Abd El$Twab, S. M., Ashour, M. B.,
tective effects through PPAR$a/$b signaling pathway in
and Yousef, A. I. (2016) Hepato$renal protective effects of
streptozocin$induced diabetic mice, J. Funct. Foods, 36,
gallic acid and p$coumaric acid in nicotinamide/strepto$
341$347.
zotocin$induced diabetic rats, Int. J. Bioassays, 5.6, 4641$
54.
Perumpail, R. B. (2015) Pathogenesis of hepatocarcino$
4649.
genesis in non$cirrhotic nonalcoholic fatty liver disease:
40.
Korish, A. A., and Arafah, M. M. (2013) Camel milk ame$
potential mechanistic pathways, World J. Gastroenterol., 7,
liorates steatohepatitis, insulin resistance and lipid peroxi$
2384$2388.
dation in experimental non$alcoholic fatty liver disease,
55.
Ingaramo, P. I., Ronco, M. T., Francés, D. E., Monti, J.
BMC Complement Altern. Med., 13, 264, doi: 10.1186/
A., Pisani, G. B., et al. (2011) Tumor necrosis factor alpha
1472$6882$13$264.
pathways develops liver apoptosis in type 1 diabetes melli$
41.
Ince, S., Arslan Acaroz, D., Neuwirth, O., Demirel, H. H.,
tus, Mol. Immunol., 48, 1397$1407.
et al. (2014) Protective effect of polydatin, a natural pre$
56.
Xiao, Y., Chen, L., Fan, Y., Yan, P., Li, S., and Zhou, X.
cursor of resveratrol, against cisplatin$induced toxicity in
(2019) The effect of boletus polysaccharides on diabetic
rats, Food Chem. Toxicol., 72, 147$153.
hepatopathy in rats, Chem. Biol. Interact., 308, 61$69.
42.
Schmatz, R., Perreira, L. B., Stefanello, N., Mazzanti, C.,
57.
Ehses, J., Lacraz, G., Giroix, M., Schmidlin, F., Coulaud,
Spanevello, R., et al. (2012) Effects of resveratrol on bio$
J., et al. (2009) IL$1 antagonism reduces hyperglycemia
markers of oxidative stress and on the activity of delta
and tissue inflammation in the type 2 diabetic GK rat,
aminolevulinic acid dehydratase in liver and kidney of
Proc. Natl. Acad. Sci. USA, 106, 13998$14003.
streptozotocin$induced diabetic rats, Biochimie, 94, 374$
58.
Ding, W., and Yin, X. (2004) Dissection of the multiple
383.
mechanisms of TNF$α$induced apoptosis in liver injury, J.
42.
Fiorentino, T. V., Prioletta, A., Zuo, P., and Folli, F. (2013)
Cell Mol. Med., 8, 445$454.
Hyperglycemia$induced oxidative stress and its role in dia$
59.
Du, Q. H., Peng, C., and Zhang, H. (2013) Polydatin: a
betes mellitus related cardiovascular diseases, Curr. Pharm.
review of pharmacology and pharmacokinetics, Pharm.
Des., 19, 5695$5703.
Biol., 51, 1347$1354.
44.
Xu, Y., Osborne, B. W., and Stanton, R. C. (2005) Diabetes
60.
Li, T., Cai, S., Zeng, Z., Zhang, J., Gao, Y., Wang, X., and
causes inhibition of glucose$6$phosphate dehydrogenase
Chen, Z. (2014) Protective effect of polydatin against burn$
via activation of PKA, which contributes to oxidative stress
induced lung injury in rats, Respir. Care, 59, 1412$1421.
in rat kidney cortex, Am. J. Physiol. Renal. Physiol., 289,
61.
Teoh, S. L., Latiff, A. A., and Das, S. (2009) A histological
F1040$F1047.
study of the structural changes in the liver of streptozotocin$
45.
Ulusu, N. N., Sahilli, M., Avci, A., Canbolat, O., Ozansoy,
induced diabetic rats treated with or without Momordica
G., et al. (2003) Pentose phosphate pathway, glutathione$
charantia (bitter gourd), Clin Ter., 160, 283$286.
dependent enzymes and antioxidant defense during oxida$
62.
Song, D., Yin, L., Wang, C., and Wen, X. (2020) Zhenqing
tive stress in diabetic rodent brain and peripheral organs:
recipe attenuates non$alcoholic fatty liver disease by regu$
effects of stobadine and vitamin E, Neurochem. Res., 28,
lating the SIK1/CRTC2 signaling in experimental diabetic
815$823.
rats, BMC Complement. Altern. Med.,
20,
27,
46.
Cedola, N., Cabarrou, A., Auciello, N., Doria, I., Ponce,
doi: 10.1186/s12906$019$2811$2.
D. L., and Baylon, N. (1974) The liver in human diabetes.
63.
Song, R. (2016) Mechanism of metformin: a tale of two
Concentration of some induced enzymes, Acta
sites, Diabetes Care, 39, 187$189.
Diabetologica, 12, 263$271.
64.
Nathan, D. M., Buse, J. B., Davidson, M. B., Ferrannini, E.,
47.
Hosni, A. A., Abdel$Moneim, A. A., Abdel$Reheim, E. S.,
Holman, R. R., Sherwin, R., Zinman, B., American
Mohamed, S. M., and Helmy, H. (2017) Cinnamaldehyde
Diabetes Association, and European Association for Study
potentially attenuates gestational hyperglycemia in rats
of Diabetes (2009) Medical management of hyperglycemia
БИОХИМИЯ том 86 вып. 2 2021
ДЕЙСТВИЕ ПОЛИДАТИНА ПРИ ПОРАЖЕНИИ ПЕЧЕНИ
227
in type 2 diabetes: a consensus algorithm for the initiation
65. Abdel$Moneim, A, Abdel$Reheim, E. S., Semmler, M.,
and adjustment of therapy - a consensus statement of the
and Addaleel, W. (2019) The impact of glycemic status and
American Diabetes Association and the European
metformin administration on red blood cell indices and
Association for the Study of Diabetes, Diabetes Care, 32,
oxidative stress in type 2 diabetic patients, Malays J. Med.
193$203.
Sci., 26, 47$60.
HEPATOPROTECTIVE EFFECTS OF POLYDATIN#LOADED CHITOSAN
NANOPARTICLES IN DIABETIC RATS: MODULATION
OF GLUCOSE METABOLISM, OXIDATIVE STRESS,
AND INFLAMMATION BIOMARKERS*
A. M. Abd El#Hameed1, A. I. Yousef2, S. M. Abd El#Twab2,
A. A. G. El#Shahawy3, and A. Abdel#Moneim2**
1 Chemistry Department, Faculty of Science, Taibah University, 30002 Al Madinah Al Munawarah, Saudi Arabia
2 Molecular Physiology Division, Faculty of Science, Beni Suef University,
62511 Beni Suef, Egypt; e mail: adel.hassan@science.bsu.edu.eg
3 Materials Science and Nanotechnology Department, Faculty of Postgraduate Studies for Advanced Sciences (PSAS),
Beni Suef University, 62511 Beni Suef, Egypt
Polydatin (PD) has a broad range of pharmacological activities; however, its effects on diabetic liver damage are poor$
ly studied. This work is aimed to explore possible protective effects of polydatin$loaded chitosan nanoparticles
(PD$CSNPs) or PD against liver damage associated with diabetes. Diabetes was induced in rats using nicoti$
namide/streptozotocin treatment. Experimental rats were then divided into six groups: normal control rats, diabetic
control rats, and rats orally treated with PD, PD$CSNPs, equivalent unloaded CSNPs, or metformin daily for 4
weeks. Treatment with PD and PD$CSNPs significantly reduced the blood glucose content, lipid peroxidation in the
liver, and activities of serum transaminases and carbohydrate metabolism enzymes (including succinate dehydroge$
nase and pyruvate kinase); by contrast, liver glycogen content, glutathione concentration, and activities of antioxidant
enzymes (superoxide dismutase, glutathione peroxidase, catalase, and glucose$6$phosphate dehydrogenase) were
markedly increased compared with control diabetic rats. Furthermore, the expression of tumor necrosis factor α and
interleukin$1β mRNAs was significantly downregulated, while the expression of glucose transporter 2 and glucoki$
nase mRNAs was strongly upregulated vs. control diabetic rats. We concluded that PD$CSNPs and PD ameliorate
diabetic liver damage by modulating glucose transporter 2 expression, affecting the activity of carbohydrate metabo$
lism enzymes, and suppressing oxidative stress and inflammation, while PD$CSNPs being more efficient than PD
alone, probably due to higher bioavailability and prolonged release.
Keywords: diabetic liver damage, polydatin$loaded chitosan nanoparticles, carbohydrate metabolism enzymes, glu$
cose transporter 2, oxidative stress
БИОХИМИЯ том 86 вып. 2 2021
6*