БИОХИМИЯ, 2021, том 86, вып. 2, с. 236 - 247
УДК 57.021
РОЛЬ P ГЛИКОПРОТЕИНА В ОГРАНИЧЕНИИ ПРОНИЦАЕМОСТИ
КЛЕТОЧНЫХ МЕМБРАН ПРИ ОКИСЛИТЕЛЬНОМ СТРЕССЕ*
© 2021
А.В. Щулькин**, Ю.В. Абаленихина, И.В. Черных,
П.Д. Ерохина, Е.Н. Якушева
Рязанский государственный медицинский университет им. академика И.П. Павлова, 390026 Рязань, Россия;
электронная почта: alekseyshulkin@rambler.ru
Поступила в редакцию 15.07.2020
После доработки 03.09.2020
Принята к публикации 09.09.2020
P гликопротеин является одним из наиболее клинически значимых представителей ABC транспортеров в
силу участия в транспорте биобиотиков и ксенобиотиков через цитоплазматическую мембрану. Известно,
что активность и экспрессия P гликопротеина может изменяться под воздействием различных химических
веществ, факторов внешней среды, патологических процессов. В настоящем исследовании была изучена
роль P гликопротеина в ограничении проницаемости цитоплазматических мембран в условиях окислитель
ного стресса. Клетки аденокарциномы ободочной кишки человека (Caco 2), гиперэкспрессирующие дан
ный белок транспортер, культивировали с пероксидом водорода в питательной среде в концентрациях
0,1-50 мкМ в течение 72 ч. Транспорт субстрата P гликопротеина фексофенадина через мембрану клеток
оценивали в специальных трансвелл системах. Количество белка транспортера и транскрипционного фак
тора Nrf2 анализировали методом иммуноферментного анализа (ИФА). Концентрацию белковых
SH групп, продуктов перекисного окисления липидов и карбонильных производных белков определяли
спектрофотометрически. По результатам исследования пероксид водорода в концентрациях 0,1-5 мкМ не
оказывал достоверного влияния на изучаемые показатели, однако в концентрации 10 мкМ вызывал сниже
ние уровня белковых SH групп, что сопровождалось повышением количества Nrf2. Последний, в свою оче
редь, увеличивал содержание и активность белка транспортера P гликопротеина, сдерживающего расту
щую проницаемость клеточной мембраны. Увеличение концентрации прооксиданта до 50 мкМ вызывало
усиление окислительного стресса, что проявлялось снижением уровня SH групп, повышением концентра
ции продуктов перекисного окисления липидов, карбонильных производных белков и снижением содержа
ния P гликопротеина, что в итоге приводило к значительному повышению проницаемости цитоплазмати
ческой мембраны. Таким образом, транспортная и защитная роль P гликопротеина, заключающаяся в ог
раничении проницаемости клеточной мембраны, зависит от выраженности окислительного стресса и реа
лизуется только при незначительных повреждениях мембран.
КЛЮЧЕВЫЕ СЛОВА: P гликопротеин, Nrf2, окислительный стресс, проницаемость биологической мем
браны, цитотоксичность пероксида водорода, окислительное повреждение липидов и белков, восстанов
ленные тиолы.
DOI: 10.31857/S0320972521020081
ВВЕДЕНИЕ
некоторых гигантских вирусах [1]. Геном чело
века включает 49 генов ABC транспортеров,
ABC транспортеры (англ. абб. ABC от ATP
объединенных в семь подсемейств (обозначае
Binding Cassette) - это ATP зависимые белки
мых латинскими буквами A-G). Известно, что
транспортеры, обеспечивающие энергозависи
нарушение функционирования 18 транспорте
мый перенос веществ через цитоплазматичес
ров связано с развитием заболеваний (напри
кие мембраны. ABC транспортеры выявлены в
мер, нарушение ABCC7 вызывает муковисци
трех доменах (надцарствах) биологических ви
доз, ABCB11 - прогрессирующий семейный
дов - бактериях, археях и эукариотах, а также в
внутрипеченочный холестаз, и т.д.), а 20 из них
Принятые сокращения: АФК - активные формы кислорода; ВЭЖХ - высокоэффективная жидкостная хромато
графия; ИФА - иммуноферментный анализ; клетки линии Caco 2 - клетки аденокарциномы ободочной кишки челове
ка; ОП - оптическая плотность; ПОЛ - продукты перекисного окисления липидов; DTNB - 5,5′ дитиобис(2 нитробен
зойная кислота); MTT - 3 (4,5 диметилтиазол 2 ил) 2,5 дифенил 2Н тетразолиум бромид; Nrf2 - транскрипционный
фактор NF E2 related factor 2; P gp - P гликопротеин.
biokhimiya, в рубрике «Papers in Press», BM20 223, 18.01.2021.
** Адресат для корреспонденции.
236
РОЛЬ P gp В МЕМБРАНЕ ПРИ ОКИСЛИТЕЛЬНОМ СТРЕССЕ
237
вовлечены в транспорт лекарственных веществ
цию перекисного окисления липидов (ПОЛ),
[2]. ABC транспортеры имеют общую структур
что приводит к образованию пор, уменьшению
ную организацию: каждая молекула переносчи
толщины бислоя, нарушению текучести и, в ко
ка состоит из двух трансмембранных доменов,
нечном счете, повышению ее проницаемости
которые образуют транспортный канал и обес
[12].
печивают распознавание и связывание субстра
С другой стороны, гиперпродукция АФК и
тов, а также двух нуклеотид связывающих доме
развитие окислительного стресса увеличивают
нов, которые гидролизуют ATP [3, 4].
экспрессию редокс чувствительного тран
Самым изученным белком из суперсемей
скрипционного фактора Nrf2 (англ. NF E2
ства ABC транспортеров является P гликопро
related factor 2) [13], который, в свою очередь,
теин (он же - белок множественной лекар
может повысить экспрессию P gp [14], ограни
ственной устойчивости 1, P gp, ABCB1 белок,
чивающего проницаемость мембраны для
MDR1) [5, 6]. P gp представляет собой крупный
субстратов.
трансмембранный гликопротеин с молекуляр
Таким образом, при гиперпродукции АФК
ной массой 170 кДа (из них 150 кДа приходится
проницаемость цитоплазматической мембраны
на пептидную часть). P gp обладает широкой
для субстратов P gp может как повыситься (за
субстратной специфичностью. Его субстратами
счет разрушения мембраны), так и понизиться
являются преимущественно липофильные ве
(за счет повышения активности белка транс
щества с молекулярной массой 330-4000 Да, от
портера), что на данный момент не изучено и
носящиеся к следующим фармакологическим
послужило целью настоящего исследования.
группам: противоопухолевые, гипотензивные и
антигистаминные препараты, сердечные глико
зиды, антиагреганты, антикоагулянты, стероид
МАТЕРИАЛЫ И МЕТОДЫ
ные и тиреоидные гормоны, антибиотики, ин
гибиторы ВИЧ протеиназы, иммунодепрессан
Культивирование клеток линии Caco 2. Ис
ты и др. [7].
следование выполнено на линии клеток адено
Впервые P gp был выделен Juliano and Ling в
карциномы ободочной кишки человека
1976 г. из цитоплазматической мембраны клеток
(Caco 2) (ЦКП «Коллекция культур клеток поз
яичника китайского хомячка, отобранных по
воночных», Санкт Петербург, Россия). Клетки
устойчивости к колхицину и демонстрирующих
культивировали при 37 °C и 5% ном содержании
перекрестную резистентность к широкому
CO2 в инкубаторе WS 189C («World Science», Ко
спектру амфифильных веществ [8]. В дальней
рея) в среде DMEM с высоким содержанием
шем этот белок транспортер был обнаружен в
глюкозы (4 500 мг/л), также содержащей L глу
энтероцитах кишечника, гепатоцитах, эпители
тамин (4 мМ), 15% бычьей сыворотки, 100 ед./мл
альных клетках почечных канальцев, эндотели
пенициллина и 100 мкг/мл стрептомицина (все
альных клетках гистогематических барьеров.
составляющие производства «Sigma Aldrich»,
Поэтому в настоящее время принято считать,
Германия). После достижения 70-90% конфлю
что P gp играет важную роль в развитии резис
ентности клетки снимали с флакона добавлени
тентности опухолей к химиотерапии [9], а также
ем раствора трипсин ЭДТА (0,25% трипсина и
в транспорте эндогенных и экзогенных веществ
0,2% ЭДТА, «Sigma Aldrich», Германия) и высеи
[10].
вали: в
96 луночный планшет
(«Corning»,
Активность и экспрессия P gp могут изме
США) - для оценки цитотоксичности пер
няться под воздействием различных химических
оксида водорода; в
6 луночные планшеты
веществ, факторов внешней среды и патологи
(«Corning», США) - для оценки влияния перок
ческих процессов [11]. Например, рифампицин
сида водорода на количество P gp, Nrf2 и концен
и гипоксия могут активировать функциониро
трацию продуктов перекисного окисления ли
вание белка транспортера, а циклоспорин и ве
пидов, карбонильных производных белков и со
рапамил его ингибируют [7], что, в свою оче
держания белковых тиоловых SH групп; в спе
редь, влияет на проникновение субстратов белка
циальные трансвелл системы (12 мм Transwell®
транспортера внутрь клеток через цитоплазма
планшеты с поликарбонатной мембранной
тическую мембрану (повышает или снижает со
вставкой с порами 0,4 мкм, «Corning», США) -
ответственно).
для оценки влияния пероксида водорода на про
Одним из факторов, влияющих на проница
ницаемость клеточных мембран для субстрата
емость цитоплазматической мембраны, являет
P gp [15]. Клетки линии Caco 2 культивировали
ся гиперпродукция активных форм кислорода
в течение 21 сут, поскольку на данном сроке
(АФК). АФК при взаимодействии с цитоплазма
происходит их спонтанная дифференцировка в
тической мембраной клеток вызывают актива
клетки, подобные энтероцитам тонкого кишеч
БИОХИМИЯ том 86 вып. 2 2021
238
ЩУЛЬКИН и др.
ника, гиперэкспрессирующие P gp. Смену
ния). Цитоплазматическую (экстраядерную)
культуральной среды проводили раз в два дня, а
фракцию переносили в отдельные пробирки и
на последней неделе - каждый день.
использовали для определения концентрации
Окислительный стресс моделировали добав
тиоловых (SH ) групп.
лением в культуральную среду пероксида водо
Аналогичным образом лизировали клетки,
рода H2O2 («Sigma Aldrich», Германия) в конеч
которые параллельно инкубировали в 6 луноч
ных концентрациях 0,1; 0,5; 1; 5; 10 и 50 мкМ и
ных планшетах для определения концентрации
инкубацией в течение 72 ч. Смену среды прово
продуктов ПОЛ и карбонильных производных
дили каждые 24 ч. На каждый эксперимент бы
белков, из расчета на 3 × 106 клеток - 450 мкл
ло выполнено по 3 повторения.
лизирующего буфера [17].
Тест на выживаемость (цитотоксический тест).
Анализ содержания P gp и Nrf2 в клетках ли
Клетки засеивали в 96 луночный планшет из
нии Caco 2. В лизате клеток методом ИФА опре
расчета 104 клеток на каждую лунку и культиви
деляли количество P gp и Nrf2 коммерческими
ровали в течение 21 сут., затем добавляли куль
наборами (Human Permeability glycoprotein
туральную среду с H2O2. После окончания инку
ELISA Kit и Human Nuclear factor erythroid 2
бации в каждую лунку добавляли по 20 мкл
related factor 2 ELISA Kit соответственно; «Blue
0,5% ного раствора 3 (4,5 диметилтиазол 2
gene», Китай). Светопоглощение измеряли при
ил) 2,5 дифенил 2Н тетразолиум бромида (MTT)
450 нм на иммуноферментном планшетном ана
и инкубировали еще в течение 2 ч, затем раствор
лизаторе Stat Fax 2100 («Awareness Technology»,
с MTT удаляли и добавляли 200 мкл 1% ного
США). Количество белка в пробах анализирова
раствора диметилсульфоксида
(«ПанЭко»,
ли методом Бредфорда (Pierce Coomassie Plus
Россия)
[16]. Поглощение измеряли через
(Bradford) Assay Kit, «ThermoFisher», США).
10 мин при 530 нм на иммуноферментном план
Определение концентрации белковых тиоло
шетном анализаторе Stat Fax 2100 («Awareness
вых (SH ) групп. Концентрацию белковых тио
Technology», США)[16].
ловых групп определяли по разнице между уров
Выживаемость клеток Caco 2 в присутствии
нем общих и низкомолекулярных SH групп.
пероксида водорода рассчитывали по формуле:
Анализ содержания общих SH групп в лизате
(ОП опытных лунок - ОП среды)/(ОП конт
клеток проводили по методу Эллмана с 5,5′ ди
рольных лунок - ОП среды) × 100%, где ОП -
тиобис(2 нитро) бензойной кислотой (DTNB)
оптическая плотность.
в неденатурирующих условиях [18]. К 100 мкл
Приготовление клеточных лизатов для анали
образца добавляли
100
мкл
2
мМ DTNB
за. Клетки засеивали в 6 луночные планшеты из
(«Serva», Германия) в
1
М Tris HCl буфере
расчета 105 клеток на каждую лунку и культиви
(pH 8,0) и 1000 мкл дистиллированной воды.
ровали в течение 21 сут., затем добавляли куль
После 30 мин экспозиции количественно оце
туральную среду с H2O2. После окончания экс
нивали содержание 5 тио 2 нитробензойной
позиции клетки снимали с лунок раствором
кислоты при 412 нм на Stat Fax 2100 («Awareness
трипсин ЭДТА (0,25% трипсина и 0,2% ЭДТА,
Technology», США). Концентрацию SH групп
«Sigma Aldrich», Германия).
рассчитывали исходя из коэффициента экстинк
Часть клеток (2 × 106) трехкратно промывали
ции ε412 = 13,6 мМ-1см-1 [19]. Для определения
фосфатным буфером pH 7,4 («ПанЭко», Рос
содержания низкомолекулярных SH групп про
сия), лизировали трехкратным циклом замора
бу предварительно смешивали с охлажденной
живания размораживания в 200 мкл буфера при
5% ной трихлоруксусной кислотой («Химмед»,
-20 °C и использовали для проведения ИФА.
Россия), инкубировали на льду
15
мин,
Вторую часть клеток (1 × 106) промывали
затем центрифугировали при 11 000 g (CM 50,
изотоническим раствором NaCl («Медпро»,
«Eppendorf», Германия) 5 мин при 4 °C. Полу
Россия), добавляли 150 мкл лизирующего буфе
ченный супернатант нейтрализовали с по
ра (50 мМ pH 7,4 Tris HCl, 150 мМ KCl, 0,5%
мощью 1% ного NaOH («Химмед», Россия) и
тритон X 100, смесь ингибиторов протеиназ
использовали для определения низкомолеку
(4 (2 аминоэтилбензенсульфонил) флуорида
лярных SH групп по реакции с DTNB.
гидрохлорид (AEBSF)
2 мМ, апротинин
Определение концентрации продуктов пере
0,3
мкМ, бестатин
130
мкМ, ЭДТА 1мМ,
кисного окисления липидов и карбонильных про
транс эпоксисукцинил L лейциламидо(4 гуа
изводных белков. В лизате клеток с помощью
нидино)бутан (E 64) 14 мкМ, лейпептин 1 мкМ,
коммерческих наборов определяли концентра
«Sigma Aldrich», Германия)), встряхивали на
цию продуктов перекисного окисления липидов
шейкере и инкубировали на льду в течение
(ПОЛ) («Elabscience», Китай) и карбонильных
10 мин. Затем центрифугировали в течение
производных белков («Sigma Aldrich», Герма
10 мин при 5000 g (CM 50, «Eppendorf», Герма
ния).
БИОХИМИЯ том 86 вып. 2 2021
РОЛЬ P gp В МЕМБРАНЕ ПРИ ОКИСЛИТЕЛЬНОМ СТРЕССЕ
239
Метод определения продуктов ПОЛ - мало
(«Sigma Aldrich», Германия) и 1% диметилсуль
нового диальдегида и 4 гидрокси олефинов
фоксида («ПанЭко», Россия). Затем добавляли
(4 гидрокси 2 ноненаля и 4 гидрокси 2 гексе
субстрат P gp - фексофенадин («Sigma Aldrich»,
наля) - основан на их взаимодействии с 1 ме
Германия) - в апикальную камеру в конечной
тил 2 фенилиндолом с образованием стабиль
концентрации 150 мкМ [22]. Через 1, 2 и 3 ч за
ного хромофора, который имеет максимум по
бирали образцы из базолатеральной камеры
глощения при 586 нм [20].
реципиента для определения концентрации
Метод определения карбонильных произ
субстрата (a&b транспорт, обусловленный пас
водных белков основан на их взаимодействии с
сивной диффузией против функционирования
2,4 динитрофенилгидразином с образованием
P gp). В аналогичных трансвелл системах оце
2,4 динитрофенилгидразонов, которые регист
нивали транспорт фексофенадина из базолате
рировали при длине волны 375 нм. Концентра
ральной камеры в апикальную (b&a транспорт,
цию карбонильных производных белков рас
обусловленный пассивной диффузией и функ
считывали исходя из коэффициента экстинк
ционированием P gp). Для этого субстрат в той
ции ε375 = 22 мМ-1см-1 [21].
же концентрации добавляли в базолатеральную
Анализ выполняли на иммуноферментном
камеру, а затем через 1, 2 и 3 ч забирали образцы
планшетном анализаторе Stat Fax
2100
из апикальной камеры для определения концен
(«Awareness Technology», США).
трации фексофенадина.
Оценка транспорта субстрата P гликопротеи
Транспорт маркерного субстрата рассчиты
на (фексофенадина) через мембрану клеток линии
вали по формуле [23]:
Caco 2. Оценку проникновения субстрата P gp
через билипидную мембрану клеток линии
Caco 2 оценивали в специальных трансвелл
,
системах (рис. 1). Трансвелл система представ
лена двумя камерами: апикальной и базолате
ральной. Дно апикальной камеры является по
где Papp - коэффициент кажущейся проницае
лупроницаемой мембраной, на которую высеи
мости (см/с), dQ/dt - изменение количества
вали клетки линии Caco 2 с плотностью 105/см2
субстрата в камере реципиенте за время инку
и культивировали в течение 21 сут.
бации (мкМ × мл/с), A - площадь полупроница
Целостность клеточного монослоя оценива
емой мембраны лунки в трансвелл системе
ли по величине трансэпителиального сопротив
(см2), C0 - начальная концентрация субстрата в
ления, которое определяли с помощью вольт
камере доноре (мкМ).
метра Millicell ERS 2 («Millipore», США). Усло
Определение концентрации фексофенадина.
вием для выполнения транспортных экспери
Концентрацию фексофенадина в транспортной
ментов было значение выше 500 мОм × см2. В
среде определяли методом высокоэффективной
лунки трансвелл системы добавляли культу
жидкостной хроматографии (ВЭЖХ) с УФ де
ральную среду с H2O2 в тестируемых концентра
тектированием при длине волны 220 нм. Иссле
циях. После окончания инкубации культураль
дование выполнялось на ВЭЖХ системе
ную среду заменяли на транспортную среду,
«Стайер» (Россия) по оригинальной методике
представляющую собой раствор Хэнкса
[24]. Полученная проба транспортной среды
(«Sigma Aldrich», Германия) с 25 мМ HEPES
(50 мкл), содержащая фексофенадин, разводи
Рис. 1. Структура трансвелл системы. Данная система представлена двумя камерами: апикальной и базолатеральной. Дно
апикальной камеры является полупроницаемой мембраной, на которую высеивали клетки линии Caco 2 с плотностью
105/см2
БИОХИМИЯ том 86 вып. 2 2021
240
ЩУЛЬКИН и др.
лась в 150 мкл подвижной фазы, и 100 мкл полу
ченного раствора вводились в хроматограф.
При анализе использовалась хроматографи
ческая колонка Synergi 4 μm Polar RP 80 Å
(250 × 4,6) («Phenomenex», США). Температура
разделения - 45 °C, скорость потока - 1 мл/мин.
Состав подвижной фазы: 128 мл ацетонитрила
(«PanReac AppliChem», Испания), 267,4 мл воды
деионизированной, 6,33 мл кислоты уксусной
ледяной («PanReac AppliChem», Испания) с до
бавлением триэтиламина («PanReac AppliChem»,
Испания) до pH 6,7. Время удерживания фексо
фенадина в данных условиях составляло
12,8 мин. Количественное определение прово
дилось методом абсолютной калибровки по
площади пиков. Аналитический диапазон мето
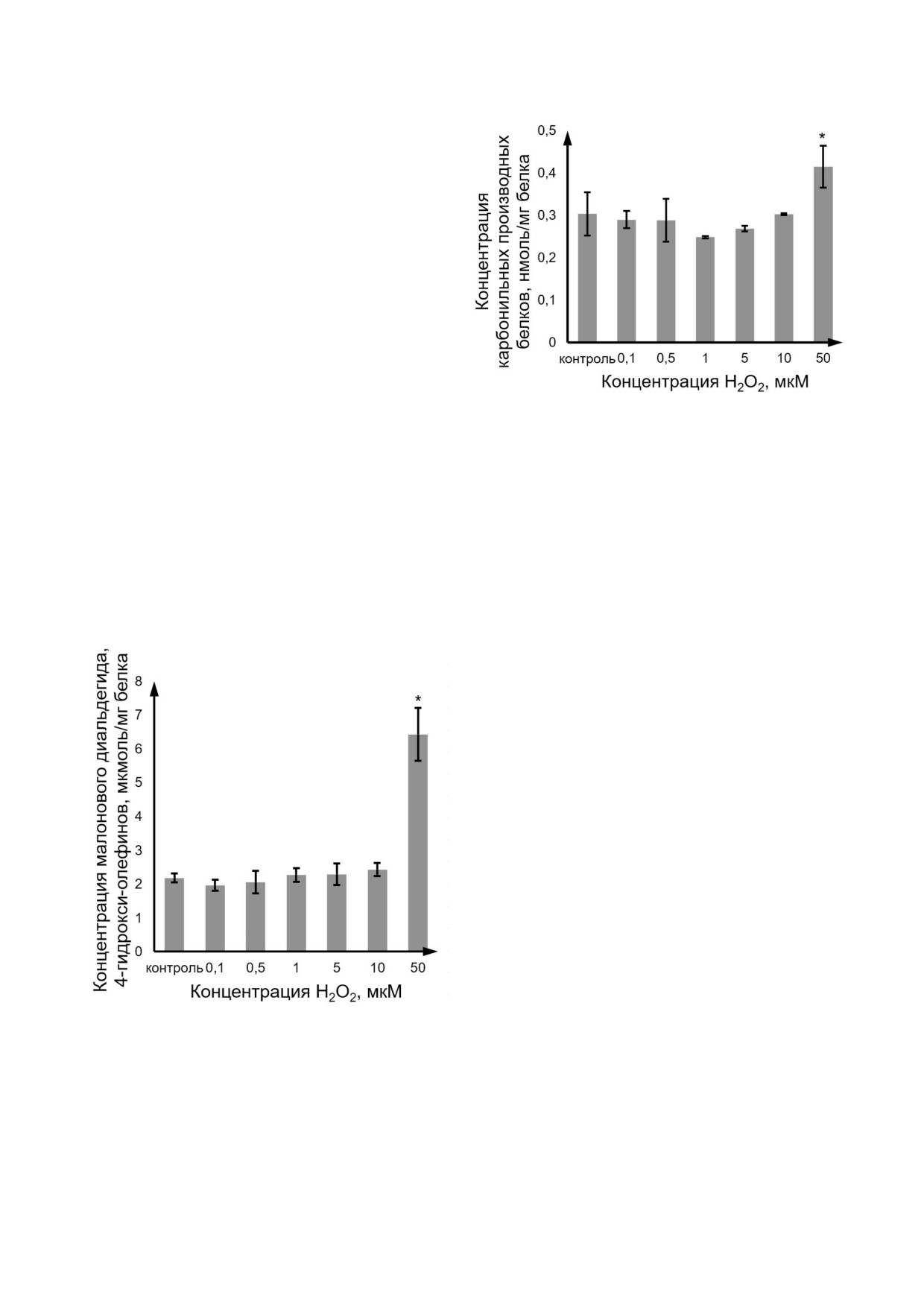
Рис. 3. Изменение концентрации карбонильных производ
дики составлял 1,2-57,4 мкМ.
ных белков при индукции окислительного стресса перок
Статистический анализ. Полученные резуль
сидом водорода в концентрациях 0,1-50 мкМ в течение
таты анализировали с помощью программ
72 ч. Количественное определение карбонилированных
белков проводили в лизате клеток Caco 2 (M ± SD, n = 3)
«StatSoft Statistica 13.0» (США, номер лицензии
фотометрическим методом с использованием коммерчес
JPZ811I521319AR25ACD W) и Microsoft Excel
кого набора «Sigma Aldrich», Германия. * - Достоверное
for Mac ver. 16.24 (ID02984 001 000001). Резуль
отличие от контроля, p < 0,05
таты представлены в виде M ± SD. Для оценки
статистической значимости различий использо
вали дисперсионный анализ (ANOVA), попар
считали различия при p < 0,05. Корреляцион
ные сравнения выполняли с помощью критерия
ный анализ проводили с помощью критерия
Ньюмена-Кейлса. Статистически значимыми
Пирсона.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Окислительное повреждение липидов и белков
клеток линии Caco 2 под действием H2O2. Окис
лительное повреждение липидов оценивали по
уровню продуктов перекисного окисления (ма
лонового диальдегида и 4 гидрокси олефинов),
белков - по концентрации белковых тиоловых
(SH ) групп и содержанию карбонильных про
изводных белков. H2O2 в концентрациях
0,1-10 мкМ достоверно не влиял на уровень
продуктов ПОЛ, а в концентрации 50 мкМ по
вышал данный показатель на 194,9% (p = 0,0002)
по сравнению с контролем (рис. 2).
Аналогичным образом содержание карбо
нильных производных белков в лизате клеток
Caco 2 повышалось только при концентрации
H2O2 50 мкМ - на 36,5% (p = 0,001) (рис. 3).
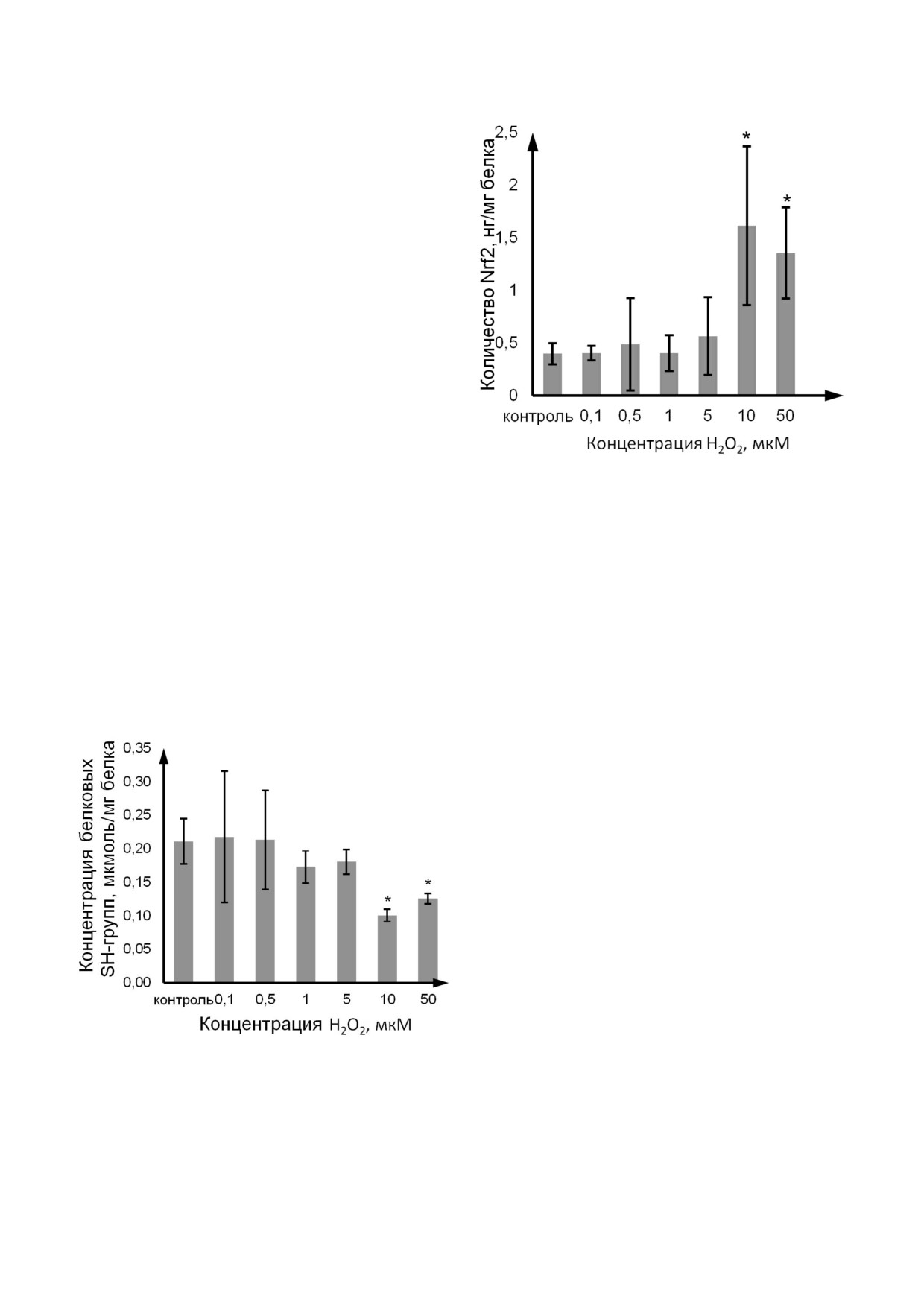
В то же время уровень белковых тиоловых
(SH ) групп при воздействии H2O2 в концентра
ции 10 мкМ уменьшался на 52,4% (p = 0,021) и
Рис. 2. Изменение концентрации продуктов перекисного
окисления липидов (малонового диальдегида и 4 гидрокси
при концентрации 50 мкМ - на 38,1% (p = 0,022)
олефинов) при индукции окислительного стресса перок
по сравнению с контролем. При концентрации
сидом водорода в концентрациях 0,1-50 мкМ в течение
пероксида водорода 0,1; 0,5; 1 и 5 мкМ уровень
72 ч. Количественное определение продуктов ПОЛ прово
восстановленных тиолов не изменялся относи
дили в лизате клеток Caco 2 (M ± SD, n = 3) фотометри
тельно значений контрольной группы (рис. 4).
ческим методом с использованием коммерческого набора
«Elabscience», Китай. * - Достоверное отличие от контро
Таким образом, установлено, что H2O2 на
ля, p < 0,05
клетках линии Caco 2 вызывает повреждение
БИОХИМИЯ том 86 вып. 2 2021
РОЛЬ P gp В МЕМБРАНЕ ПРИ ОКИСЛИТЕЛЬНОМ СТРЕССЕ
241
липидов и белков в концентрации от 50 мкМ,
при этом окисление белковых SH групп, играю
щих регуляторную роль, происходит уже при
концентрации прооксиданта 10 мкМ.
Цитотоксическое действие H2O2 на клетки ли
нии Caco 2. Цитотоксическое действие H2O2
оценивалось в ходе MTT теста. В интактной
группе клеток выживаемость клеток составила
100%, при действии пероксида водорода в кон
центрациях 0,1-10 мкМ - статистически значи
мо не отличалась от значений группы контроля
и составила 87,6 ± 9,3%. При концентрации
H2O2 50 мкМ выживаемость клеток снижалась
до 58,8 ± 11,5% (p = 0,02). В совокупности с опи
санными выше результатами эти данные свиде
тельствуют о том, что H2O2 вызывает поврежде
ние и гибель клеток только при развитии выра
женного окислительного стресса, проявляюще
гося повреждением липидов и белков.
Влияние H2O2 на количество P gp и Nrf2. Ко
Рис. 5. Изменение количества Nrf2 при индукции окисли
тельного стресса пероксидом водорода в концентрациях
личество Nrf2 в клетках линии Caco 2 повыша
0,1-50 мкМ в течение 72 ч. Количество Nrf2 определяли в
лось по сравнению с контролем под действием
лизате клеток Caco 2 (M ± SD, n = 3) с помощью ИФА.
H2O2 в концентрациях 10 мкМ и 50 мкМ на
* - Достоверное отличие от контроля, p < 0,05
302,5% (p = 0,02) и 237,5% (p = 0,03) соответ
ственно, а при остальных концентрациях про
оксиданта - достоверно не изменялось (рис. 5)
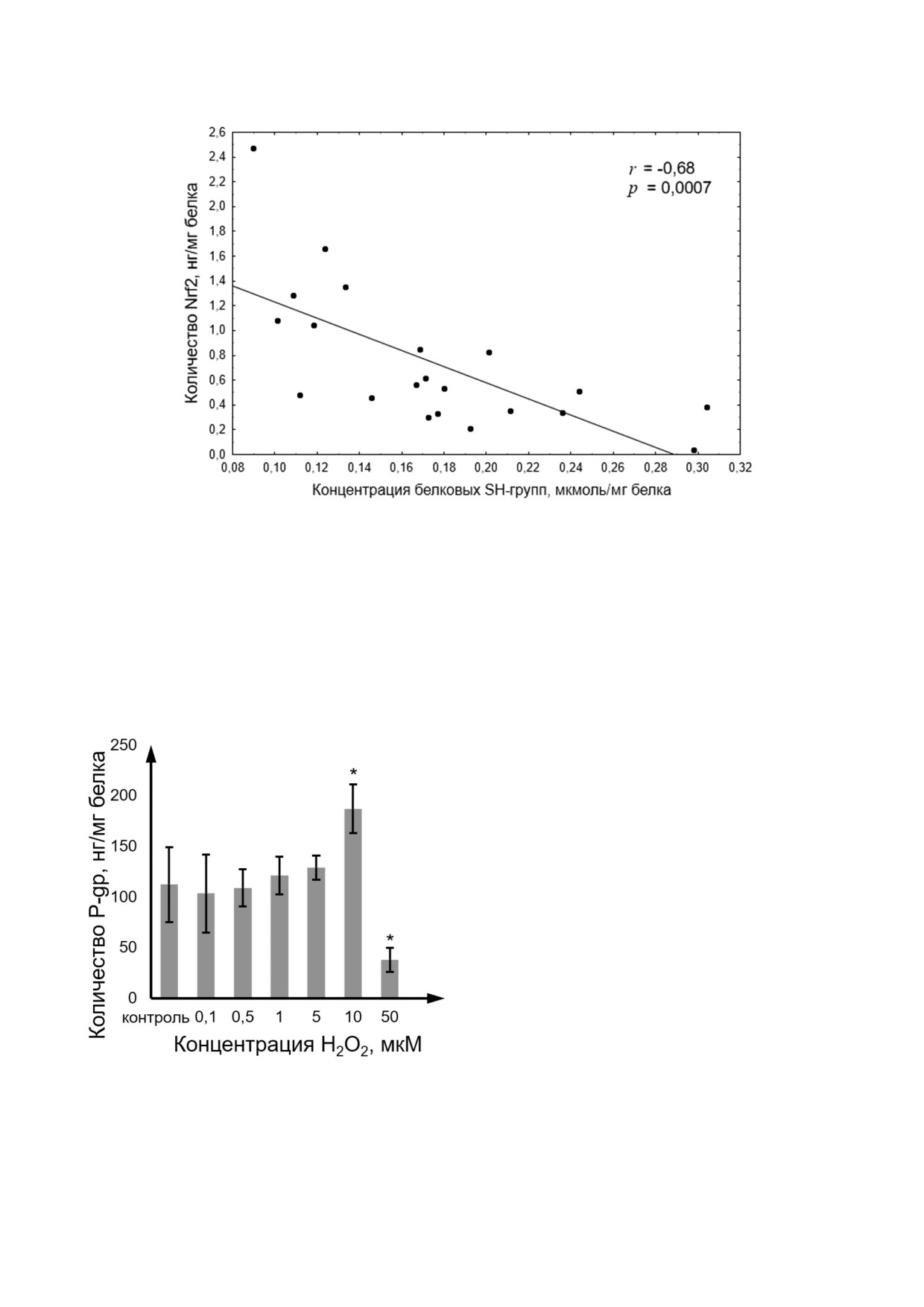
Количество P gp в клетках линии Caco 2,
Показано, что в диапазоне концентраций
оцененное методом ИФА, составило 110,8 нг/мг
H2O2 от 0,1 до 50 мкМ уровень Nrf2 был обратно
белка или 23,5 нг/106 клеток. H2O2 в концентра
пропорционален содержанию белковых тиоло
циях 0,1; 0,5; 1; 5 мкМ не влиял на количество
вых (SH ) групп (рис. 6), что согласуется с клас
белка транспортера. В концентрации 10 мкМ
сическими представлениями о механизме регу
пероксид водорода повышал содержание P gp
ляции Nrf2 [25].
на 68,9% (p = 0,01), а в концентрации 50 мкМ -
снижал его на 65,8% (p = 0,01) по сравнению с
показателями контроля (рис. 7)
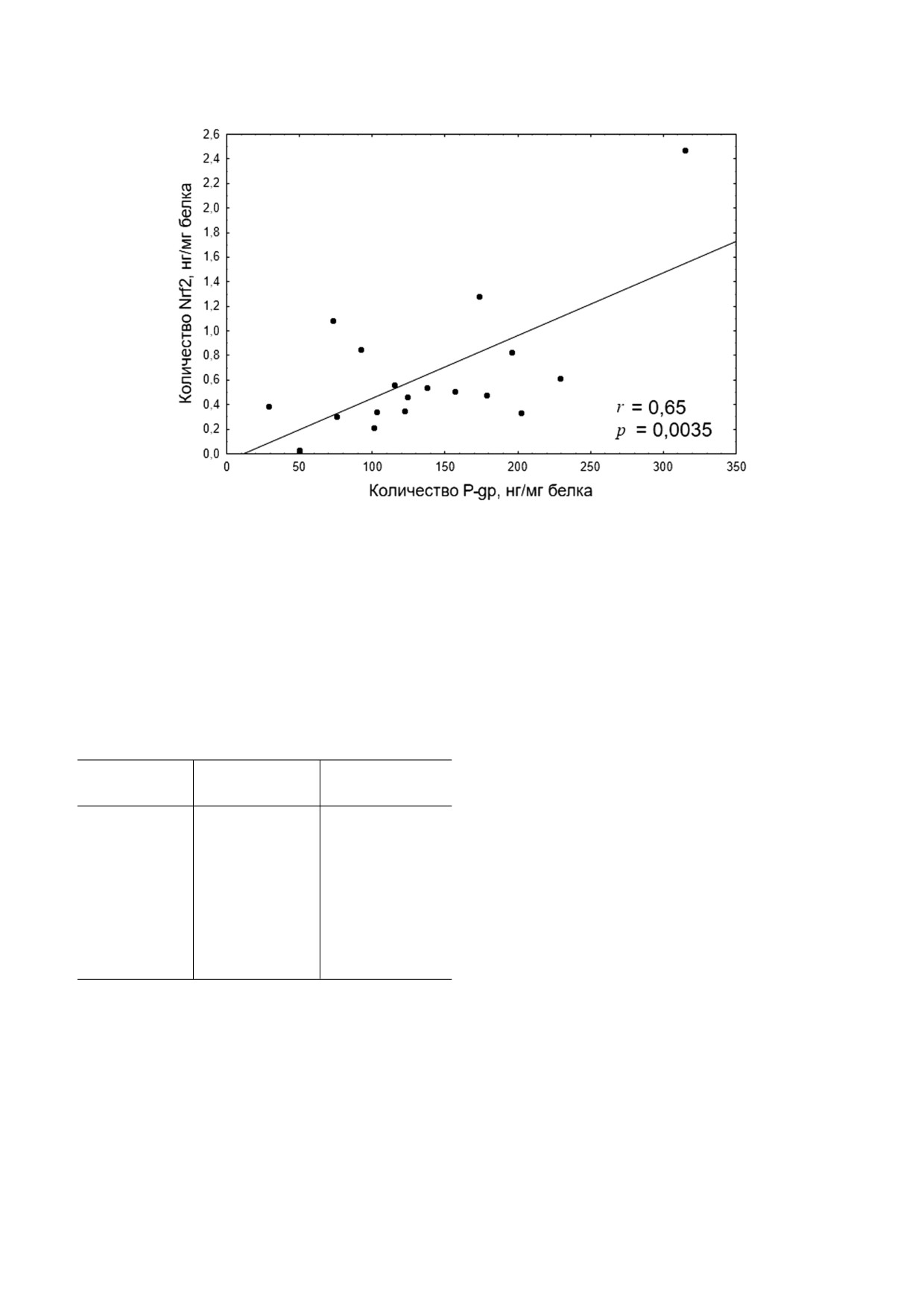
Анализ результатов выявил, что уровень P gp
в лизате клеток Caco 2 был прямо пропорцио
нален содержанию Nrf2 в экспериментах с H2O2
в диапазоне концентраций от 0,1 до 10 мкМ
(рис. 8).
Таким образом, окислительный стресс и
окисление SH групп повышает уровень тран
скрипционного фактора Nrf2, который, в свою
очередь, увеличивает количество белка транс
портера P gp. Усиление окислительного стресса
приводит к снижению количества P gp, несмот
ря на сохраняющийся повышенный уровень
Nrf2.
Влияние H2O2 на транспорт субстрата P gp
(фексофенадина) через билипидную мембрану кле
ток линии Caco 2. Коэффициент кажущейся про
ницаемости Papp b&a фексофенадина, характери
Рис. 4. Изменение концентрации белковых SH групп при
зующий его транспорт из базолатеральной каме
индукции окислительного стресса пероксидом водорода в
ры в апикальную за счет пассивной диффузии и
концентрациях 0,1-50 мкМ в течение 72 ч. Количествен
функционирования P gp, составил 3,08 × 10-6 ±
ное определение тиоловых (SH ) групп проводили фото
± 0,94 × 10-6 см/с, коэффициент кажущейся
метрическим методом по реакции с DTNB («Serva», Герма
ния) в лизате клеток Caco 2 (M ± SD, n = 3). * - Достовер
проницаемости Papp a&b, оценивающий транс
ное отличие от контроля, p < 0,05
порт из апикальной камеры в базолатеральную
7 БИОХИМИЯ том 86 вып. 2 2021
242
ЩУЛЬКИН и др.
Рис. 6. Корреляционная зависимость между концентрацией SH групп (мкмоль/мг белка) и количеством Nrf2 (нг/мг бел
ка) при индукции окислительного стресса пероксидом водорода в концентрациях 0,1-50 мкМ в течение 72 ч. Коэффици
ент корреляции r = -0,68, p = 0,0007, что свидетельствует об обратной зависимости между концентрацией восстановлен
ных тиолов и количеством транскрипционного фактора Nrf2
за счет пассивной диффузии против функции
счет пассивной диффузии против работы белка
P gp, составил 1,13 × 10-6 ± 0,6 × 10-6 см/с. Из
транспортера (p = 0,013) (таблица).
представленных данных видно, что транспорт,
H2O2 в концентрациях 0,1-5 мкМ достовер
обусловленный функционированием P gp, поч
но не влиял на транспорт субстрата P gp фексо
ти в 3 раза выше транспорта, происходящего за
фенадина из апикальной камеры в базальную и
в обратном направлении. При этом сохранялось
превышение транспорта, осуществляемого P gp
и пассивной диффузией, над транспортом пас
сивной диффузией против работы изучаемого
белка транспортера (p < 0,05), что характеризует
стабильное состояние его функциональной ак
тивности и проницаемости мембраны.
В концентрации 10 мкМ H2O2 вызывал по
вышение коэффициента кажущейся проницае
мости Papp b&a фексофенадина по сравнению с
контролем на 98,1% (p = 0,056) и не влиял на ко
эффициент кажущейся проницаемости Papp
a&b. Данные результаты свидетельствуют о том,
что транспорт фексофенадина, обусловленный
P gp, повышается, при этом транспорт субстра
та пассивной диффузией против функциониро
вания белка транспортера достоверно не изме
няется.
При увеличении концентрации H2O2 до
50 мкМ происходило повышение коэффициен
та кажущейся проницаемости Papp b&a фексо
Рис. 7. Изменение количества P gp при индукции окисли
фенадина на 260,7% (p = 0,001) и увеличение
тельного стресса пероксидом водорода в концентрациях
коэффициента кажущейся проницаемости Papp
0,1-50 мкМ в течение 72 ч. Количество P gp определяли в
лизате клеток Caco 2 (M ± SD, n = 3) с помощью ИФА.
a&b на 1029,2% (p = 0,001) по сравнению с конт
* - Достоверное отличие от контроля, p < 0,05
ролем (таблица). При этом коэффициент кажу
БИОХИМИЯ том 86 вып. 2 2021
РОЛЬ P gp В МЕМБРАНЕ ПРИ ОКИСЛИТЕЛЬНОМ СТРЕССЕ
243
Рис. 8. Корреляционная зависимость между количеством Nrf2 (нг/мг белка) и P gp (нг/мг белка) при индукции окисли
тельного стресса пероксидом водорода в концентрациях 0,1-10 мкМ в течение 72 ч. Коэффициент корреляции r = 0,65,
p = 0,0035, что свидетельствует о прямой зависимости между количеством транскрипционного фактора Nrf2 и уровнем
P gp при условии целостности цитоплазматической мембраны
щейся проницаемости Papp b&a (обусловлен
зий против функции белка транспортера)
ный P gp и пассивной диффузий) выравнивал
(p > 0,05).
ся с коэффициентом кажущейся проницаемос
Таким образом, показано, что H2O2 в концен
ти Papp a&b (обусловленным пассивной диффу
трации 10 мкМ повышает активность P gp в
клетках линии Caco 2. При более высокой кон
центрации H2O2 происходит повреждение как
Влияние H2O2 на транспорт субстрата P gp (фексофенади
клеточной мембраны, так и белка транспортера
на) через билипидную мембрану клеток Caco 2 (M ± SD,
n = 3)
P gp, что следует из данных по количеству бел
ка транспортера в лизате клеток - это приводит
Серии
Papp b&a,
Papp a&b,
к существенному повышению проницаемости
эксперимента
×10-6 см/с
×10-6 см/с
клеточной мембраны в обоих направлениях.
Контроль
3,08 ± 0,94
1,13 ± 0,6
0,1 мкМ H2O2
3,02 ± 0,85
1,23 ± 0,59
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
0,5 мкМ H2O2
3,19 ± 0,53
1,32 ± 0,26
1 мкМ H2O2
3,73 ± 1,29
1,79 ± 0,14
P gp - ATP зависимый белок транспортер,
обеспечивающий эффлюкс экзогенных и эндо
5 мкМ H2O
2
3,37 ± 0,41
1,18 ± 0,45
генных веществ из клеток в межклеточное
10 мкМ H2O2
* 6,10 ± 0,82
2,64 ± 1,44
пространство и биологические жидкости. В опу
50 мкМ H2O2
** 11,11 ± 2,96
**12,76 ± 3,39
холевых клетках P gp способствует их защите от
цитостатиков, в кишечнике - препятствует аб
Примечание. Клетки Caco 2 после экспозиции с перокси
сорбции биобиотиков и ксенобиотиков, в эндо
дом водорода (72 ч) инкубировали в транспортной среде с телии гематоэнцефалического барьера - про
добавлением фексофенадина (150 мкМ) в апикальную или
никновению субстратов в ткань мозга, в печени
базолатеральную камеры соответствующих трансвелл сис
и почках - выводит вещества в желчь и мочу [6].
тем. Забирали образцы через 1, 2 и 3 ч из противоположной
камеры реципиента: базолатеральной (a&b транспорт) и Некоторые лекарственные вещества и условия
апикальной камеры (b&a транспорт) для определения кон
среды могут изменять скорость синтеза и актив
центрации фексофенадина. Концентрацию фексофенади
ность P gp. Например, показано, что Nrf2 может
на в транспортной среде определяли методом ВЭЖХ с
УФ детектированием. * - Отличие от контроля, р = 0,056,
стимулировать синтез белка транспортера при
** - достоверное отличие от контроля, p < 0,001.
окислительном повреждении гепатоцитов [26],
БИОХИМИЯ том 86 вып. 2 2021
7*
244
ЩУЛЬКИН и др.
нервных клеток [27], клеток гематоэнцефали
мембраны [36]. Соотношение характера струк
ческого барьера [28] и карциномы легкого A549
турного и функционального повреждения
[14], что приводит к повышению активности
мембраны при разной степени выраженности
P gp и соответственно снижает проницаемость
окислительного стресса недостаточно изучено.
мембраны для его субстратов.
Насколько нам известно, ранее исследований, в
В настоящем исследовании в опытах in vitro
которых анализировались бы количественные и
на линии клеток Caco 2 оценивалось количест
функциональные параметры мембранного
во P gp и проницаемость его субстрата фексо
транспортера P gp при развитии окислительно
фенадина через билипидную мембрану при
го стресса, не проводилось.
окислительном стрессе разной степени выра
В ходе данной работы было показано, что
женности, вызванном H2O2. Период распада
развитие окислительного стресса, вызванного
пероксида водорода составляет от миллисекунд
H2O2 в концентрации 10 мкМ, приводило к
до секунд, что позволяет ему проникать в клет
окислению белковых SH групп, что вызывало
ку сквозь мембрану и оказывать внутриклеточ
повышение количества Nrf2. Данный тран
ное воздействие. Проникновение пероксида
скрипционный фактор стимулировал увеличе
водорода сквозь клеточную мембрану может
ние содержания и активности P gp, что прояв
также осуществляться с участием аквапоринов
лялось в повышении транспорта субстрата бел
[29]. H2O2 является стабильным и малореактив
ка транспортера из базолатеральной камеры
ным агентом, вследствие чего может действо
трансвелл системы в апикальную (за счет пас
вать как вторичный внутриклеточный посред
сивной диффузии и функционирования P gp).
ник [30]. Неконтролируемое взаимодействие
Логично было ожидать, что транспорт фексофе
пероксида водорода с переходными металлами
надина из апикальной камеры в базолатераль
(Fe2+ или Cu+) по реакции Фентона приводит к
ную (за счет пассивной диффузии против рабо
образованию АФК [31]. В свою очередь, актив
ты белка транспортера) при этом снизится. Од
ные формы кислорода могут быть нейтрализо
нако в нашем исследовании данный показатель
ваны антиоксидантными системами или накап
не изменялся. Скорее всего, это связано с тем,
ливаться в клетках, вызывая окислительный
что при воздействии H2O2 в концентрации
стресс, что приводит к повреждению белков,
10 мкМ увеличивается проницаемость билипид
липидов, ДНК, мутагенезу и гибели клеток. В
ной мембраны, но повышение активности P gp
условиях окислительного стресса наиболее
нивелирует данное нарушение.
чувствительными к повреждению являются се
Увеличение концентрации H2O2 до 50 мкМ
росодержащие аминокислотные остатки (мети
усиливало окислительный стресс, вызывало на
онин и цистеин), что имеет регуляторное значе
копление малонового диальдегида и 4 гидрокcи
ние [32, 33].
олефинов (4 гидрокси 2 ноненаля и 4 гидрок
На изменение соотношения восстановлен
си 2 гексеналя), карбонильных производных
ных и окисленных SH групп в белках реагирует
белков, а также снижало количество P gp, при
Nrf2 - редокс чувствительный транскрипцион
этом уровень Nrf2 оставался повышенным, что,
ный фактор. Его экспрессия повышается при
скорее всего, связано с локализацией изучае
развитии окислительного стресса и направлена
мых белков. P gp находится преимущественно в
на защиту клетки от воздействия свободных ра
цитоплазматической мембране, подверженной
дикалов [13, 34]. В нормальных условиях дан
воздействию H2O2 в первую очередь, а Nrf2 - в
ный транскрипционный фактор находится в
цитоплазме и ядре [37], более защищенных от
комплексе с белком репрессором Keap1, их свя
воздействия прооксиданта в инкубационной
зывание регулируется рядом протеинкиназ.
среде.
Keap1, с одной стороны, способствует убикви
Снижение количества P gp и повреждение
тинированию и протеосомальной деградации
билипидной мембраны в условиях окислитель
Nrf2 (необходимым условием для этого процес
ного стресса (концентрация H2O2 50 мкМ) при
са является наличие двух остатков цистеина в
водило к повышению транспорта субстрата белка
молекуле Keap1), а с другой - предотвращает
транспортера фексофенадина через цитоплаз
его проникновение из цитоплазмы в ядро [35].
матическую мембрану как из базолатеральной
После активации комплекс Keap1-Nrf2 диссо
камеры в апикальную, так и наоборот - из апи
циирует, и Nrf2 транслоцируется в ядро, где свя
кальной в базолатеральную. Причем значение
зывается с элементами антиоксидантного ответа
коэффициента кажущейся проницаемости Papp
(англ. абб. ARE) и активирует транскрипцию за
b&a соответствовало коэффициенту кажущейся
щитных ферментов [35].
проницаемости Papp a&b. Биохимические изме
В то же время окислительный стресс приво
нения при воздействии H2O2 в концентрациях
дит к повышению проницаемости билипидной
50 мкМ сопровождались существенным по
БИОХИМИЯ том 86 вып. 2 2021
РОЛЬ P gp В МЕМБРАНЕ ПРИ ОКИСЛИТЕЛЬНОМ СТРЕССЕ
245
вышением гибели клеток по результатам
Таким образом, увеличение количества Nrf2
MTT теста.
при окислительном стрессе стимулирует повы
Полученные результаты могут иметь важное
шение содержания P gp, что сдерживает расту
практические значение. Например, считается,
щую проницаемость клеточной мембраны. Уси
что P gp в гематоэнцефалическом барьере явля
ление окислительного стресса вызывает пов
ется одним из ключевых факторов, ограничива
реждение мембран и белка транспортера P gp,
ющих проникновение лекарственных веществ
что приводит к значительному повышению про
субстратов в головной мозг, снижая тем самым
ницаемости цитоплазматической мембраны в
эффективность терапии церебральных заболе
обоих направлениях. Таким образом, транспорт
ваний (острого нарушения мозгового кровооб
ная и защитная роль P gp, заключающаяся в ог
ращения, эпилепсии), а его ингибирование рас
раничении проницаемости клеточной мембра
сматривается в качестве терапевтической мише
ны, зависит от выраженности окислительного
ни при лечении данных патологий [38, 39].
стресса и реализуется только при незначитель
В нашем исследовании показано, что разви
ных повреждениях мембран.
тие выраженного окислительного стресса (кото
рое наблюдается и при данных неврологических
Финансирование. Работа выполнена при фи
заболеваниях) приводит к повреждению били
нансовой поддержке гранта Президента Рос
пидных мембран и P gp, что сопровождается
сийской Федерации для государственной под
повышением проницаемости цитоплазматичес
держки молодых российских ученых - кандида
ких мембран. То есть в условиях патологии за
тов наук МК 1856.2020.7.
щитная роль P gp сохраняется только при ком
Конфликт интересов. Авторы заявляют об от
пенсированном окислительном стрессе, что го
сутствии конфликта интересов.
ворит в пользу того, что данный белок транс
Соблюдение этических норм. Настоящая
портер с большой вероятностью не стоит рас
статья не содержит описания каких либо иссле
сматривать в качестве перспективной мишени
дований с участием людей или животных в каче
для терапевтического воздействия.
стве объектов.
СПИСОК ЛИТЕРАТУРЫ
1.
Davidson, A. L. (2008) Structure, function, and evolution
cell mutants, Biochem. Biophis. Acta, 455, 155162,
of bacterial ATP binding cassette systems, Microbiol. Mol.
doi: 10.1016/0005 2736(76)90160 7.
Biol. Rev., 72, 317 364, doi: 10.1128/MMBR.00031 07.
9. Pokharel, D., Roseblade, A., Oenarto, V., Lu, J. F., and
2.
Vasiliou, V., Vasiliou, K., and Nebert, D. W.
(2009)
Bebawy, M. (2017) Proteins regulating the intercellular
Human ATP binding cassette (ABC) transporter family,
transfer and function of P glycoprotein in multidrug resis
Hum. Genomics, 3, 281 290, doi: 10.1186/1479 7364 3 3
tant cancer, Ecancermedicalscience, 11, e768, doi: 10.3332/
281.
ecancer.2017.768.
3.
Licht, A., and Schneider, E. (2011) ATP binding cassette
10. Borst, P., and Schinkel, A. H. (2013) P glycoprotein
systems: Structures, mechanisms, and functions, Cent. Eur.
ABCB1: a major player in drug handling by mammals, J.
J. Biol., 6, 785 801, doi: 10.2478/s11535 011 0054 4.
Clin. Invest., 23, 4131 4133, doi: 10.1172/JCI70430.
4.
Esser, L., Zhou, F., Pluchino, K. M., Shiloach, J., Ma, J.,
11. Yano, K., Tomono, T., and Ogihara, T. (2018) Advances in
et al. (2017) Structures of the multidrug transporter P gly
studies of P glycoprotein and its expression regulators,
coprotein reveal asymmetric ATP binding and the mecha
Biol. Pharm. Bull., 41, 11 19, doi: 10.1248/bpb.b17 00725.
nism of polyspecificity, J. Biol. Chem., 292, 446 461,
12. Van der Paal, J., Neyts, E. C., Verlackt, C. C. W., and
doi: 10.1074/jbc.M116.755884.
Bogaerts, A. (2016) Effect of lipid peroxidation on mem
5.
Sharom, F. J. (2011) The P glycoprotein multidrug trans
brane permeability of cancer and normal cells subjected to
porter, Essays Biochem., 50, 161178, doi: 10.1042/
oxidative stress, Chem. Sci., 7, 489 498, doi: 10.1039/
bse0500161.
C5SC02311D.
6.
Якушева Е. Н., Титов Д. С. (2018) Структура и функ
13. Raghunath, A., Sundarraj, K., Nagarajan, R., Arfuso, F.,
ционирование белка множественной лекарственной
Bian, J., Kumar, A. P., Sethi, G., and Perumala, E. (2018)
устойчивости 1, Биохимия, 8, 1148 1172, doi: 10.1134/
Antioxidant response elements: Discovery, classes, regula
S0320972518080043.
tion and potential applications, Redox. Biol., 17, 297 314,
7.
Кукес В. Г., Грачев С. В., Сычев Д. А., Раменская Г. В.
doi: 10.1016/j.redox.2018.05.002.
(2008) Метаболизм лекарственных средств. Научные
14. Wu, B., Li, H. X., Lian, J., Guo, Y. J., Tang, Y. H., Chang,
основы персонализированной медицины: руковод
Z. J., Hu, L. F., Zhao, G. J., Hong, G. L., and Lu, Z. Q.
ство для врачей?, Гэотар&Медиа, Москва, 304 с.
(2019) Nrf2 overexpression protects against paraquat
8.
Juliano, R. L., and Ling, V. (1976) A surface glycoprotein
induced A549 cell injury primarily by upregulating P glyco
modulating drug permeability in Chinese hamster ovary
protein and reducing intracellular paraquat accumulation,
БИОХИМИЯ том 86 вып. 2 2021
246
ЩУЛЬКИН и др.
Exp. Ther. Med., 17, 1240 1247, doi: 10.3892/etm.2018.
26.
Thakkar, N., Slizgi, J. R., and Brouwer, K. L. R. (2017)
7044.
Effect of liver disease on hepatic transporter expression and
15.
Якушева Е. Н., Щулькин А. В., Черных И. В., Попова
function, J. Pharm Sci., 106, 2282 2294, doi: 10.1016/
Н. М., Котлярова А. А., Слепнев А. А. (2019) Метод
j.xphs.2017.04.053.
анализа принадлежности лекарственных веществ к
27.
Grewal, G. K., Kukal, S., Kanojia, N., Saso, L., Kukreti,
субстратам и ингибиторам белка транспортера гли
Sh., and Kukreti, R. (2017) Effect of oxidative stress on ABC
копротеина Р in vitro, Обзоры по клинической фармако&
transporters: contribution to epilepsy pharmacoresistance,
логии и лекарственной терапии, 17, 71 78, doi: 10.7816/
Molecules, 22, e365, doi: 10.3390/molecules22030365.
RCF17171 78.
28.
Aryal, M., Fischer, K., Gentile, C., Gitto, S., Zhang, Y.
16.
Tolosa, L., Donato, M. T., and Gómez Lechón, M. J.
Z., and McDannold, N. (2017) Effects on P glycoprotein
(2015) General cytotoxicity assessment by means of the
expression after blood brain barrier disruption using
MTT assay, Methods Mol. Biol.,
1250,
333348,
focused ultrasound and microbubbles, PLoS One, 3,
doi: 10.1007/978 1 4939 2074 7_26.
e0166061, doi: 10.1371/journal.pone.0166061.
17.
Tinnikov, A. A., and Samuels, H. H. (2013) A novel cell
29.
Rodrigues, O., Reshetnyak, G., Grondin, A., Saijo ,Y.,
lysis approach reveals that caspase 2 rapidly translocates
Leonhardt, N., Maurel Ch., and Verdoucq, L. (2017)
from the nucleus to the cytoplasm in response to apoptotic
Aquaporins facilitate hydrogen peroxide entry into guard
stimuli, PLoS One, 8, e61085, doi: 10.1371/journal.pone.
cells to mediate ABA and pathogen triggered stomatal
0061085.
closure, Proc. Natl. Acad. Sci. USA, 114, 9200 9205,
18.
Boschi Muller, S., Azza, S., Sanglier Cianferani, S.,
doi: 10.1073/pnas.1704754114.
Talfournier, F., Dorsselear, A. V., and Branlant, G. (2000)
30.
Sies, H. (2017) Hydrogen peroxide as a central redox sig
A sulfenic acid enzyme intermediate is involved in the cat
naling molecule in physiological oxidative stress: oxidative
alytic mechanism of peptide methionine sulfoxide reduc
eustress, Redox Biol., 11, 613 619, doi: 10.1016/j.redox.
tase from Escherichia coli, J. Biol. Chem., 275, 35908
2016.12.035.
35913, doi: 10.1074/jbc.M006137200.
31.
Smirnoff, N., and Arnaud, D. (2019) Hydrogen peroxide
19.
Ellman, L. G. (1959) Tissue sulfhydryl groups, Arch.
metabolism and functions in plants, New Phytol., 2, 1197
Biochem. Biophys.,
82,
7077, doi:
10.1016/0003
1214, doi: 10.1111/nph.15488.
9861(59)90090 6.
32.
Vogelsang, L., and Dietz, K. J. (2020) Regulatory thiol
20.
Gérard Monnier, D., Erdelmeier, I., Régnard, K., Moze
oxidation in chloroplast metabolism, oxidative stress
Henry, N., Yadan, J. C., Chaudière, J. (1998) Reactions of
response and environmental signaling in plants, Biochem.
1 methyl 2 phenylindole with malondialdehyde and 4
J., 477, 1865 1878, doi:10.1042/BCJ20190124.
hydroxyalkenals. Analytical applications to a colorimetric
33.
Poole, L.B. (2015) The basics of thiols and cysteines in
assay of lipid peroxidation, Chem. Res. Toxicol., 11, 1176
redox biology and chemistry, Free Radic. Biol. Med., 1,
1183, doi: 10.1021/tx9701790.
148 157, doi: 10.1016/j.freeradbiomed.2014.11.013.
21.
Weber, D., Davies, M. J., and Grunea, T.
(2015)
34.
Kang, K. A., and Hyun, J. W. (2017) Oxidative stress, Nrf2,
Determination of protein carbonyls in plasma, cell
and epigenetic modification contribute to anticancer drug
extracts, tissue homogenates, isolated proteins: focus on
resistance, Toxicol. Res., 33, 1 5, doi: 10.5487/TR.2017.33.
sample preparation and derivatization conditions, Redox
1.001.
Biol., 5, 367 380, doi: 10.1016/j.redox.2015.06.005.
35.
Wen, Zh., Liu, W., Li, X., Chen, W., Liu, J., Wen, Zh., and
22.
Petri, N., Tannergren, C., Rungstad, D., and Lennernäs,
Liu, Zh. (2019) A protective role of the NRF2 Keap1
H. (2004) Transport characteristics of fexofenadine in the
pathway in maintaining intestinal barrier function, Oxid.
Caco 2 cell model, Pharmac. Res., 21, 13981404,
Med. Cell. Longev., e1759149, doi:
10.1155/2019/
doi: 10.1023/B:PHAM.0000036913.90332.b1.
1759149.
23.
Elsby, R., Surry, D. D., Smith, V. N., and Gray, A. J. (2008)
36.
Itri, R., Junqueira, H. C., Mertins, O., and Baptista, M. S.
Validation and application of Caco 2 assays for the in vitro
(2014) Membrane changes under oxidative stress: the
evaluation of development candidate drugs as substrates or
impact of oxidized lipids, Biophys. Rev., 6, 4761,
inhibitors of P glycoprotein to support regulatory submis
doi: 10.1007/s12551 013 0128 9.
sions, Xenobiotica,
38,
11401164, doi:
10.1080/
37.
Yoon, D. S., Choi, Y., and Lee, J. W. (2016) Cellular local
00498250802050880.
ization of NRF2 determines the self renewal and
24.
Ерохина П. Д., Абаленихина Ю. В., Щулькин А. В.,
osteogenic differentiation potential of human MSCs via
Черных И. В., Попова Н. М., Слепнев А. А., Якушева
the P53-SIRT1 axis, Cell Death Dis.,
7, e2093,
Е. Н. (2020) Изучение влияния прогестерона на актив
doi: 10.1038/cddis.2016.3.
ность гликопротеина Р in vitro, Российский медико&био&
38.
Fromm, M. F. (2004) Importance of P glycoprotein at
логический вестник имени академика И.П. Павлова, 28,
blood-tissue barriers, Trends Pharmacol. Sci., 25, 423 429,
135 142, doi: 10.23888/PAVLOVJ2020282135 142.
doi: 10.1016/j.tips.2004.06.002.
25.
Lennicke, C., Rahn, J., Lichtenfels, R., Wessjohann, L. A.,
39.
Wang, G. X., Wang, D. W., Liu, Y., and Ma, Y. H. (2016)
and Seliger, B. (2015) Hydrogen peroxide - production,
Intractable epilepsy and the P glycoprotein hypothesis,
fate and role in redox signaling of tumor cells, Cell Commun.
Int. J. Neurosci., 126, 385 392, doi: 10.3109/00207454.
Signal., 13, e39, doi: 10.1186/s12964 015 0118 6.
2015.1038710.
БИОХИМИЯ том 86 вып. 2 2021
РОЛЬ P gp В МЕМБРАНЕ ПРИ ОКИСЛИТЕЛЬНОМ СТРЕССЕ
247
P GLYCOPROTEIN ROLE IN LIMITATION OF PERMEABILITY
OF CELL MEMBRANES DURING OXIDATIVE STRESS*
A. V. Shchulkin**, Yu. V. Abalenikhina, I. V. Chernykh, P. D. Erokhina, and E. N. Yakusheva
Ryazan State Medical University, 390026 Ryazan, Russia; E&mail: alekseyshulkin@rambler.ru
P Glycoprotein (P gp) is one of the most clinically significant representatives of the ABC transporter superfamily due
to its participation in the transport of biotic components and xenobiotics across the plasma membrane. It is known
that various chemicals, environmental factors, and pathological processes can affect P gp activity and expression. In
this study, we investigated the role of P gp in limiting the cell membrane permeability during oxidative stress. Human
adenocarcinoma colon cells (Caco 2) overexpressing P gp were cultured for 72 h in the medium containing hydro
gen peroxide (0.1 50 μM). The transport of the P gp substrate fexofenadine was evaluated in a special Transwell sys
tem. The amounts of P gp and Nrf2 transcription factor were analyzed by the enzyme linked immunosorbent assay.
The concentration of SH groups in proteins and the contents of lipid peroxidation products and protein carbonyl
derivatives were determined spectrophotometrically. Hydrogen peroxide at a concentration of 0.1 5 μM did not sig
nificantly affect the studied parameters, while incubation with 10 μM H2O2 decreased in the level of SH groups in cell
lysates and increased in the amount of Nrf2 in the cell lysates. Nrf2, in its turn, mediated an increase in the content
and activity of the P gp transporter, thus limiting the increasing permeability of the cell membrane. Hydrogen perox
ide at a concentration of 50 μM promoted oxidative stress, which was manifested as a decrease in the content of SH
groups, increase in the concentration of lipid peroxidation products and protein carbonyl derivatives, and decrease in
the P gp level, which led to a significantly increased permeability of the plasma membrane. These results show that
the transport and protective roles of P gp, in particular, reduction of the cell membrane permeability, are affected by
the intensity of oxidative stress and can be manifested only if the extent of membrane damage is insignificant.
Keywords: P glycoprotein (P gp), Nrf2, oxidative stress, permeability of biological membranes, hydrogen peroxide
cytotoxicity, oxidative damage of lipids and proteins, reduced thiols
БИОХИМИЯ том 86 вып. 2 2021